mushroom corals (fungiidae) in the davao gulf, philippines
TRANSCRIPT

198
Bos & Hoeksema: Fungiidae and associated fish fauna
Mushroom corals (Fungiidae) in the Davao Gulf, Philippines, with records of associated fish and other cryptofauna
Arthur R. Bos1, 2, 3* & Bert W. Hoeksema2, 4
Abstract. Mushroom corals (Fungiidae) have rarely been studied in the southeast of the Philippines and even less studies have reported fish fauna associated with fungiids. The present study provides a species overview, relative abundance, and associated fish and invertebrate fauna of the mushroom corals from the Davao Gulf (southeast Mindanao). Thirty-three mushroom coral species belonging to 13 genera were recorded with Danafungia horrida being the most common coral with a relative frequency of 20.4%. By two additional records from previous studies, the mushroom coral fauna of southeastern Mindanao consists of at least 35 species, placing it in the centre of the Coral Triangle. The reports of the poorly-known species Podabacia sinai and Sandalolitha boucheti constitute first records for the Philippines, hence range extensions for both species. Furthermore, six rare species were encountered: Cycloseris mokai, Halomitra clavator, Heliofungia fralinae, Lithophyllon spinifer, Pleuractis gravis, and Podabacia motuporensis. The associated cryptofauna consisted predominantly of fish, shrimps, and acoel flatworms. Commensal fish were mainly represented by the Gobiidae and Tripterygiidae, with the goby Eviota rubriceps dwelling on 17 mushroom coral species, being the most common associated fish species. The largemouth triplefin Ucla xenogrammus was encountered twice on individuals of Pleuractis paumotensis. Apart from the commensal shrimps Cuapetes lacertae, C. kororensis, and the acoel flatworm Waminoa sp., unidentified brittle stars (Ophiuroidea), and a crab were found in association with mushroom corals. This study resulted in new records of associated fauna for Fungiidae, whereas cryptobenthic species of various other taxa have been reported from mushroom corals studied elsewhere. Based on the diversity of Fungiidae in the central Philippines and other areas in the Coral Triangle, it may be expected that the species list of the Davao Gulf may be further expanded in the future.
Key words. Caridea, Eviota, Gobiidae, Ophiuroidea, Scleractinia, Tripterygiidae
RAFFLES BULLETIN OF ZOOLOGY 65: 198–206Date of publication: 12 June 2017http://zoobank.org/urn:lsid:zoobank.org:pub:8F75D446-3D56-4FED-A308-334BDDEDE872
© National University of SingaporeISSN 2345-7600 (electronic) | ISSN 0217-2445 (print)
1Department of Biology, The American University in Cairo, P.O. Box 74, New Cairo 11835, Egypt2Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, The Netherlands3Davao del Norte State College, New Visayas, 8105 Panabo, The Philippines; Email: [email protected] (*corresponding author)4Institute of Biology Leiden, Leiden University, P.O. Box 9505, 2300 RA Leiden, The Netherlands
INTRODUCTION
Mushroom corals (Scleractinia: Fungiidae) constitute a family of stony corals, which are commonly encountered on Indo-Pacific coral reefs (Hoeksema, 1989). This family is represented by over 50 species with the majority (ca. 80%) being free-living when fully-grown, and the others being encrusting or permanently attached (Hoeksema, 1989, 2014; Gittenberger et al., 2011; Benzoni et al., 2012). While the occurrence of attached species is restricted to solid substrates during their entire sedentary life phase (Hoeksema, 2009; Gilbert et al., 2015), corals of free-living species detach themselves from the substrate (Hoeksema & Yeemin, 2011) and become mobile, which is advantageous during competitive interactions with other sessile organisms (Chadwick & Loya, 1992; Hoeksema & de Voogd, 2012;
Hoeksema et al., 2014) and enables the corals to escape. Moreover, it allows the corals to inhabit coastal and offshore habitats as well as shallow reef flats and deep reef bases, including sandy bottoms and mesophotic reefs (Goffredo & Chadwick-Furman, 2000; Hoeksema, 2012a, 2012b; Waheed & Hoeksema, 2013, 2014; Lane & Hoeksema, 2016).
Many species of scleractinian corals, including Fungiidae, may serve as habitat for an associated fauna represented by a large variety of species (Scott, 1987; Stella et al., 2011; Hoeksema et al., 2017). They belong to the reef cryptofauna, because they are usually well hidden due to their small size, endolithic position, or the use of camouflage (Hoeksema et al., 2012). Mushroom corals offer various kinds of habitats for cryptobenthic fauna. For example, the free-living mushroom coral Heliofungia actiniformis Quoy & Gaimard, 1833 with its unique long tentacles, provides shelter to various species of invertebrates and fish (Hoeksema & Fransen, 2011; Bos, 2012; Hoeksema et al., 2012; Bos & Hoeksema, 2015). Some of the associated organisms have never been recorded from other coral species and appear to be host-specific, such as the pipefish Siokunichtys nigrolineatus Dawson, 1983 and the commensal shrimp Cuapetes kororensis (Bruce, 1969) (Hoeksema et al., 2012). Other fungiids host endolithic organisms living inside the coral skeleton, such as boring molluscs (Gittenberger & Gittenberger, 2011; Owada & Hoeksema, 2011) or fauna dwelling on the coral’s upper
Conservation & Ecology

199
RAFFLES BULLETIN OF ZOOLOGY 2017
surface, such as acoelomorph flatworms (Hoeksema & Farenzena, 2012), coral gall crabs (van der Meij & Hoeksema, 2013; van der Meij et al., 2015) and serpulid tube worms (Hoeksema & ten Hove, 2014). The underside of mushroom corals may provide shelter to various invertebrates, which in some cases feed on their hosts (Gittenberger & Hoeksema, 2013; Hoeksema et al., 2013a, 2013b; Alamaru et al., 2016). These hidden organisms may temporarily lose their shelter when their mobile host corals are accidently turned over and need to find an alternative hiding place until the host coral returns in an upright position (Gittenberger & Hoeksema, 2013; Hoeksema & Bongaerts, 2016). Most studies of fungiid-associated fauna have reported invertebrates and, with the exception of H. actiniformis, very little is known about mushroom coral species serving as habitat for cryptobenthic fish fauna.
Since coral-associated organisms appear to be selectively linked to one or more hosts, areas with high densities of potential host corals may also have an abundant and species-rich associated fauna. The Coral Triangle region in the central Indo-Pacific harbours the highest diversity of reef coral species in the world (Hoeksema, 2007). Recently, this was confirmed for mushroom corals (Waheed & Hoeksema, 2013, 2014; Hoeksema & Lane, 2014; Waheed et al., 2015b; Lane & Hoeksema, 2016). As a consequence, the diversity of coral-associated fauna may also be high and studies on symbiotic relationships should ideally be conducted within the Coral Triangle. The Philippines is part of the Coral Triangle and knowledge of its mushroom coral fauna is limited to a few locations, which may no longer be complete and accurate (e.g., Hoeksema, 1989; Veron & Hodgson, 1989; Veron & Fenner, 2000; Licuanan, 2009). Hence, information on species ranges, coral diversity patterns, and associated fauna from the Philippines is still poor, which can be overcome by adding species records from areas that so far have remained understudied.
The Davao Gulf, situated in the southern Philippines, provides an excellent setting for the study on associated fauna of mushroom corals. Previous studies on Philippine corals have neither included the Davao Gulf nor southeast Mindanao (Nemenzo, 1986; Veron & Hodgson, 1989; Lucuanan & Capili, 2004; Huang et al., 2015). Therefore, knowledge about coral reefs and the marine fauna of the Davao Gulf is limited. Recent studies have shown that these coral reefs vary in their reef fauna compositions compared to other locations in the Philippines (Bos & Smits, 2013; Bos & Gumanao, 2013; Bos, 2014). Furthermore, these reefs support ecological processes, such as corallimorpharian polyps feeding on the crown-of-thorns sea star Acanthaster planci (Bos et al., 2008, 2011; Bos, 2011), that have so far not been observed elsewhere. The Davao Gulf borders the area where the westward North Equatorial Current divides itself into the southwest-ward Mindanao Current and the Kuroshio Current, which continues its way northward to Taiwan and Japan along the east coast of the Philippines archipelago (Hoeksema, 2007: Fig. 11). This is of biogeographical importance because larval dispersal through these currents is shaping species richness patterns. Although southeast Mindanao is part of the Coral Triangle,
located near its northeastern boundary (see Hoeksema, 2007; Veron et al., 2015), there are no records supporting this position. Because previous studies on mushroom coral fauna in the Coral Triangle are available for comparison and considering the strategic marine biogeographical position of the area, the present study aims at describing and quantifying the mushroom corals in the Davao Gulf in the southern Philippines. Because fungiids constitute microhabitats for associated organisms adding to local marine biodiversity, the present study further aims at identifying fish and other fauna living on these mushroom corals.
METHODS
Mushroom corals were studied in the Davao Gulf, Philippines between 11 March and 2 May 2016. A total of 19 surveys were conducted along the western coasts of the islands of Samal and Talikud using SCUBA (see Bos et al., 2013 for details about the location of the study area) and 11 additional surveys were conducted in shallow patch reefs while snorkelling. The vast majority of surveys was conducted during the day (between 0900 and 1700 hours), whereas four surveys were conducted after sunset. During each survey, mushroom corals were searched for using the roving diving technique, which is the most effective method for detecting mushroom coral species (Hoeksema & Koh, 2009). Each observed mushroom coral was carefully approached avoiding disturbance of associated fauna. Close-up photographs were taken of the oral side, and in some cases also of the underside of the corals for documentation and identification purposes. Relative frequency (the percentage of individuals per mushroom coral species related to all individuals encountered during the study) was used to provide a quantitative measure of mushroom coral density.
Organisms were considered associated fauna when touching tissue and/or the skeleton of mushroom corals. Associated fish species were identified using Froese & Pauly (2016) and associated invertebrates were identified using Bruce (1992) and Gosliner et al. (1996). The World Register of Marine Species (WoRMS Editorial Board, 2016) was used for updating nomenclature of species names. Total Length (TL) of fish was estimated to the nearest 5 mm. Numbers of organisms using mushroom corals as habitat were counted per mushroom coral species and per individual coral and the nature of the association was recorded. The water depth of the surveys ranged from 0 to 31 m. Water temperature was measured at 1°C accuracy and ranged from 27 to 30°C during the study period.
RESULTS
Fungiidae. A total of 515 mushroom coral individuals, belonging to 13 genera and representing 33 species, were studied in the coral reefs in the Davao Gulf (Table 1). Danafungia horrida was the most common species with a relative frequency of 20.4%. Lithophyllon repanda, Pleuractis granulosa, Fungia fungites, P. paumotensis, and P. moluccensis, in order of decreasing relative frequency (>5%), were also commonly encountered (Table 1).
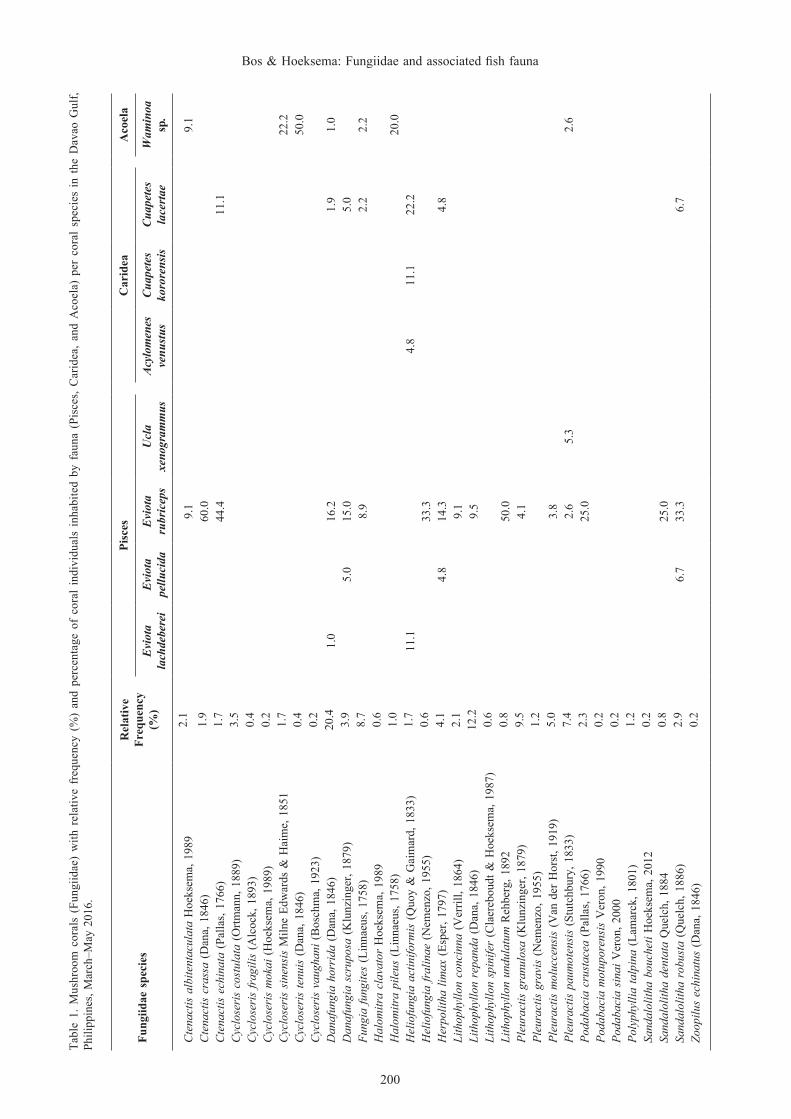
200
Bos & Hoeksema: Fungiidae and associated fish faunaTa
ble
1. M
ushr
oom
cor
als
(Fun
giid
ae)
with
rel
ativ
e fr
eque
ncy
(%)
and
perc
enta
ge o
f co
ral
indi
vidu
als
inha
bite
d by
fau
na (
Pisc
es, C
arid
ea, a
nd A
coel
a) p
er c
oral
spe
cies
in
the
Dav
ao G
ulf,
Phili
ppin
es, M
arch
–May
201
6.
Fung
iidae
spe
cies
Rel
ativ
e Fr
eque
ncy
(%)
Pisc
esC
arid
eaA
coel
a
Evi
ota
lach
debe
rei
Evi
ota
pellu
cida
Evi
ota
rubr
icep
sU
cla
xeno
gram
mus
Acyl
omen
es
venu
stus
Cua
pete
s ko
rore
nsis
Cua
pete
s la
cert
aeW
amin
oa
sp.
Cte
nact
is a
lbite
ntac
ulat
a H
oeks
ema,
198
92.
19.
19.
1C
tena
ctis
cra
ssa
(Dan
a, 1
846)
1.9
60.0
Cte
nact
is e
chin
ata
(Pal
las,
1766
)1.
744
.411
.1C
yclo
seri
s co
stul
ata
(Ortm
ann,
188
9)3.
5C
yclo
seri
s fr
agili
s (A
lcoc
k, 1
893)
0.4
Cyc
lose
ris
mok
ai (H
oeks
ema,
198
9)0.
2C
yclo
seri
s si
nens
is M
ilne
Edw
ards
& H
aim
e, 1
851
1.7
22.2
Cyc
lose
ris
tenu
is (D
ana,
184
6)0.
450
.0C
yclo
seri
s va
ugha
ni (B
osch
ma,
192
3)0.
2D
anaf
ungi
a ho
rrid
a (D
ana,
184
6)20
.41.
016
.21.
91.
0D
anaf
ungi
a sc
rupo
sa (K
lunz
inge
r, 18
79)
3.9
5.0
15.0
5.0
Fung
ia fu
ngite
s (L
inna
eus,
1758
)8.
78.
92.
22.
2H
alom
itra
clav
ator
Hoe
ksem
a, 1
989
0.6
Hal
omitr
a pi
leus
(Lin
naeu
s, 17
58)
1.0
20.0
Hel
iofu
ngia
act
inifo
rmis
(Quo
y &
Gai
mar
d, 1
833)
1.7
11.1
4.8
11.1
22.2
Hel
iofu
ngia
fral
inae
(Nem
enzo
, 195
5)0.
633
.3H
erpo
litha
lim
ax (E
sper
, 179
7)4.
14.
814
.34.
8Li
thop
hyllo
n co
ncin
na (V
erril
l, 18
64)
2.1
9.1
Lith
ophy
llon
repa
nda
(Dan
a, 1
846)
12.2
9.5
Lith
ophy
llon
spin
ifer
(Cla
ereb
oudt
& H
oeks
ema,
198
7)0.
6Li
thop
hyllo
n un
dula
tum
Reh
berg
, 189
20.
850
.0Pl
eura
ctis
gra
nulo
sa (K
lunz
inge
r, 18
79)
9.5
4.1
Pleu
ract
is g
ravi
s (N
emen
zo, 1
955)
1.2
Pleu
ract
is m
oluc
cens
is (V
an d
er H
orst
, 191
9)5.
03.
8Pl
eura
ctis
pau
mot
ensi
s (S
tutc
hbur
y, 1
833)
7.4
2.6
5.3
2.6
Poda
baci
a cr
usta
cea
(Pal
las,
1766
)2.
325
.0Po
daba
cia
mot
upor
ensi
s V
eron
, 199
00.
2Po
daba
cia
sina
i Ver
on, 2
000
0.2
Poly
phyl
lia ta
lpin
a (L
amar
ck, 1
801)
1.2
Sand
alol
itha
bouc
heti
Hoe
ksem
a, 2
012
0.2
Sand
alol
itha
dent
ata
Que
lch,
188
40.
825
.0Sa
ndal
olith
a ro
bust
a (Q
uelc
h, 1
886)
2.9
6.7
33.3
6.7
Zoop
ilus
echi
natu
s (D
ana,
184
6)0.
2

201
RAFFLES BULLETIN OF ZOOLOGY 2017
Several of the mushroom corals encountered in the Davao Gulf were uncommon species for the region or constituted range confirmations or range extensions (Fig. 1). The records of the little known species Podabacia sinai and Sandalolitha boucheti constitute range extensions and first records for the Philippines (Fig. 1A, 1B). The individual of Podabacia sinai was encountered in a relatively protected inshore reef at a depth of 14 m. Podabacia sinai is normally an attached species, but the single individual found in Davao Gulf had regenerated from a detached fragment. As a consequence, it remained detached and grew relatively large with a diameter of about 40 cm (Fig. 1A). Therefore, its appearance was similar to Zoopilus echinatus, but it was distinguished by its much finer septal ornamentation (see Hoeksema & Lane, 2014) as recognised from a close-up photograph. Corals of Podabacia sinai appear to be thin and fragile and therefore probably break and regenerate easily, as observed in P. dentata (Hoeksema & Dai, 1991). The single individual of Sandalolitha boucheti was found at a depth of 10 m in an offshore reef that is highly exposed to tidal currents.
The uncommon mushroom coral species Halomitra clavator, Heliofungia fralinae, and Lithophyllon spinifer are conspicuous species and can be easily identified (Fig. 1C–E), but were rarely encountered, each with a relative frequency of 0.6% (Table 1). Similarly, the uncommon species Cycloseris mokai, Pleuractis gravis and Podabacia motuporensis (Fig. 1F–H), easily confused with some of their respective congeners, had low relative frequencies (Table 1).
Associated fauna. The fauna associated with mushroom corals in the Davao Gulf were mainly represented by Pisces, Caridea, and Acoela (Table 1). Twenty-two coral species were used as habitat, but associated fauna was completely absent from 11 coral species. Pisces and Caridea were absent from all representatives of the fungiid genera Cycloseris, Halomitra, Polyphyllia, and Zoopilus, and from the coral species Lithopyllon spinifer, Pleuractis gravis, Podabacia motuporensis, and Sandalolitha boucheti.
Pisces were represented by the Gobiidae and Tripterygiidae with the goby Eviota rubriceps (Fig. 1D) being the most frequently associated fish, dwelling on 17 mushroom coral species (Table 1). This goby was regularly found in pairs on the mushroom corals. Apart from these associated E. rubriceps individuals or pairs resting on the corals, small groups of up to seven individuals were observed in close proximity of the corals at some occasions. Its congeners E. lachdeberei and E. pellucida were associated with two and three mushroom coral species respectively. The triplefin Ucla xenogrammus was encountered twice on individuals of Pleuractis paumotensis (Fig. 2A). Two individuals (TL 50–70 mm) of the cardinal fish Cheilodipterus quinquelineatus Cuvier, 1828 were observed swimming about the extended tentacles of Heliofungia actiniformis polyps (Fig. 2B).
The commensal shrimps Cuapetes lacertae (Fig. 2C) and C. kororensis were found on seven and one mushroom coral species, respectively (Table 1). During a night survey, nine individuals of C. lacertae were found on one coral
of Herpolitha limax (Table 1). The shrimp Ancylomenes venustus was only hosted by Heliofungia actiniformis.
The acoelomorph flatworm Waminoa sp. was found living on seven mushroom coral species (Table 1) and was always present in high numbers, ranging from 15 to several hundreds.
A relatively large brittle star (Ophiothrix sp.) rested one of its arms on a polyp of D. scruposa (Fig. 2C). Furthermore, four individuals of an unidentified genus of brittle star (Ophiuroidea) were found on three polyps of Danafungia horrida (Fig. 2D) and one individual of the same brittle star on a coral of D. scruposa. All of these relatively small brittle stars had their central disk at the underside of the coral while several arms were clung around the coral’s edge, seemingly hiding in the grooves between the septa of the corals. Apart from the above mentioned associated fauna, an unidentified crab was found living underneath a coral of Herpolitha limax, and two polychaetes had tubes grown into the skeletons of D. horrida and Lithophyllon concinna.
DISCUSSION
The total number of 33 Fungiidae species is high for the Davao Gulf, especially considering that this report is based on photographic records (Table 1). In an earlier study (Hoeksema, 1989), four mushroom coral species were recorded from southeast Mindanao, and two of these were not encountered in the present survey: Cycloseris cyclolites Lamarck, 1816 and C. somervillei Gardiner, 1909. They may not have been encountered during the present survey, because they generally occur in relatively deep water, especially the latter species (Hoeksema, 2012a). This implies that the total record of Fungiidae for southeastern Mindanao may be at least 35 species. The same number of mushroom corals was recorded off Kota Kinabalu, northwestern Sabah (Waheed & Hoeksema, 2014), whereas it is slightly lower than the total of Fungiidae species reported from other locations within the Coral Triangle: 40 from the central Philippines (Hoeksema & Putra, 2000) and west Papua (Hoeksema, 2008), 36 from Ternate in eastern Indonesia (Gittenberger et al., 2015), 42 from north Sulawesi (Hoeksema, unpublished data), 44 from eastern Sabah (Waheed & Hoeksema, 2013), and 39 from both the northernmost tip of Borneo (Waheed et al., 2015b), and Brunei (Lane & Hoeksema, 2016).
Two mushroom corals encountered during the present study, Podabacia sinai and Sandalolitha boucheti, were originally described from the Red Sea and Vanuatu, respectively (Veron, 2000; Hoeksema, 2012c) and had never been found in the Philippines and thus constitute new records. Both species are known from eastern Sabah (Waheed & Hoeksema, 2013), Layang-Layang atoll in the Spratly Islands (Waheed et al., 2015a), and northern Sulawesi (Hoeksema, unpublished data). Additionally, P. sinai was found in northwestern Sabah (Waheed & Hoeksema, 2014) and in west Papua, Indonesia (Hoeksema, 2008). Hence, the nearest previous records of P. sinai and S. boucheti are from northern Borneo and eastern Indonesia, respectively. The Davao Gulf is approximately at the same latitude as Layang-Layang (7°N), which so far

202
Bos & Hoeksema: Fungiidae and associated fish fauna
Fig. 1. Uncommon mushroom corals (Fungiidae) from the Davao Gulf, Philippines. A, Podabacia sinai; B, Sandalolitha boucheti; C, Halomitra clavator; D, Heliofungia fralinae with associated goby, Eviota rubriceps (arrow); E, Lithophyllon spinifer; F, Cycloseris mokai; G, Pleuractis gravis; H, Podabacia motuporensis.

203
RAFFLES BULLETIN OF ZOOLOGY 2017
has been the northernmost record for both species. The new records in the Davao Gulf, located further north-east than all other localities, therefore constitute range extensions. Both species were encountered only once during the present study (Table 1), and due to their rareness, information about their preferred habitats is limited. The present observations suggest however, that S. boucheti prefers more exposed reefs than P. sinai. Since both species are known from a few localities, their discovery in the Davao Gulf means that this area harbors rare coral species and that it is important for the reef coral fauna of the Philippines.
Three other mushroom coral species are so far only known from a few localities: Halomitra clavator, Heliofungia fralinae, and Lithophyllon spinifer (Table 1). Each of these have previously been reported from the Philippines (e.g., Huang et al., 2015) and were encountered three times during the present survey. Halomitra clavator is a very fragile species, which is usually encountered on lower reef slopes (> 10 m depth) and in clear water with little sediment accumulation (Hoeksema, 1993, 2008, 2012a; Hoeksema & Gittenberger, 2010; Waheed & Hoeksema, 2013; Gittenberger et al., 2015). In the Davao Gulf however, turbidity is
relatively high and visibility is usually no more than 10 m (A. R. Bos, pers. obs.). Heliofungia fralinae is usually also found on lower reef slopes (Hoeksema, 2012a, 2012b) and due to its well-developed capacity of asexual reproduction through budding, it is able to form large aggregations (Hoeksema, 2004). However, such an aggregation was not observed during the present study. Lithophyllon spinifer is most commonly found on soft sediment of relatively deep reef bases (Hoeksema, 1993, 2012a; Waheed & Hoeksema, 2013; Gittenberger et al., 2015; Lane & Hoeksema, 2016), which was confirmed by our observations. Also Cycloseris mokai, Pleuractis gravis, and Podabacia motuporensis are poorly known reef-slope species, predominantly found in the Philippines, Indonesia and Malaysia (Hoeksema & Putra, 2000; Hoeksema, 2008; Waheed & Hoeksema, 2013, 2014; Gittenberger et al., 2015; Huang et al., 2015).
Densities of mushroom corals vary with location and depth. E.g., in Singapore densities of more than 6 m-2 have been reported (Hoeksema & Koh, 2009) and in Indonesia densities reached values as high as 10 m-2 (Hoeksema, 2012a). In aggregations, mushroom corals may occur in extremely high densities with coral cover reaching 100% (Hoeksema &
Fig. 2. A, Individual of the largemouth triplefin, Ucla xenogrammus, on a turned-over mushroom coral, Pleuractis paumotensis; B, Juvenile of a five-lined cardinalfish, Cheilodipterus quinquelineatus, hovering between the tentacles of a Heliofungia actiniformis; C, An arm of a brittle star, Ophiothrix sp., and a shrimp, Cuapetes lacertae (arrow), on a polyp of a Danafungia scruposa; D, Two arms of another species of brittle star resting in grooves (arrow) between septa of a Danafungia horrida.

204
Bos & Hoeksema: Fungiidae and associated fish fauna
Gittenberger, 2010). Densities of mushroom corals have not been studied in the Davao Gulf, but coral cover of fungiids reached up to 2.4% of the sediment in some reefs surrounding Samal Island (Gumanao, 2009). Although mushroom coral densities may be lower in the Davao Gulf, this did not result in a significantly lower diversity of mushroom coral species.
Associations between fishes and corals have been reported from Indo-Pacific and Caribbean coral reefs with representatives of the Gobiidae (suborder Gobiodei) being the most commonly encountered (Schiemer et al., 2009; Sih & Chouw, 2009; Tornabene et al., 2013; Goatley et al., 2016), whereas also cases are known that involve fish families belonging to the suborder Blennioidei (Butter et al., 1980; Böhm & Hoeksema, 2017). Although associations between mushroom corals and shrimps have been well documented (Hoeksema et al., 2012), the association between fish and mushroom corals has long been considered to be only existent in the white pipefish, Siokunichthys nigrolineatus being a strictly host-specific associate of Heliofungia actiniformis (Phillips & Pullin, 1987; Hoeksema et al., 2012). However, 15 species of the Apogonidae, Gobiidae, Labridae and Pomacentridae were recently identified to find refuge among the tentacles of H. actiniformis (Bos, 2012; Bos & Hoeksema, 2015). In previous studies associated fish fauna in fungiids had been given little attention (Hoeksema et al., 2012). During the present study, three gobies (Eviota spp.) and one representative of the Tripterygiidae (Ucla xenogrammus) frequently settled on mushroom corals of various species (Table 1). Eviota rubriceps appears to be a habitat generalist, because it was found on 17 fungiids (Table 1). Although it was not found in Heliofungia actiniformis, a fungiid with long tentacles that is known to provide micro-habitats for small assemblages of co-habiting fish and invertebrates (e.g., Hoeksema & Fransen, 2011) during the present study, earlier records from this coral (Bos, 2012; Bos & Hoeksema, 2015) confirm its generalist behavior. Until today, apart from the well-known association between S. nigrolineatus and H. actiniformis, which surprisingly has never been reported from the Davao Gulf, associations between fishes and mushroom corals have only been studied in the Davao Gulf (Bos, 2012; Bos & Hoeksema, 2015). It is remarkable that these unique associations have neither been reported from other coral reefs in the Coral Triangle, nor from the wider Indo-Pacific region.
Bos & Hoeksema (2015) firstly reported the association between a brittle star (Ophiuroidea) and H. actiniformis. During the present study, five individual brittle stars, representing two species, were found living entirely or partly on or underneath mushroom corals with their arms exposed to the oral side of the corals (Fig. 2). The nature of this relation between brittle stars and corals is presently unknown and deserves further investigation.
Acoelomorph flatworms, in particular Acoela belonging to the genus Waminoa, have been recorded for a few areas and some coral species in the Indo-Pacific, including mushroom corals (Haapkylä et al., 2009; Barneah et al., 2012; Hoeksema & Farenzena, 2012; Cooper et al., 2015;
Ponti et al., 2016). The present record of seven host species is high considering that these only cover fungiid corals. Since several undescribed acoelomorphs have been observed in North Sulawesi, where a high concentration of species was observed on scleractinians, corallimorpharians and alcyonaceans (Farenzena & Hoeksema, unpubl. obs.), they may contribute more to marine biodiversity than previously suggested.
The nature of the relation between mushroom corals and their associated fauna is usually not clear. It is likely that various cryptobenthic species are commensals just looking for shelter underneath free-living corals or about their tentacles (Hoeksema et al., 2012). Some species of fish, shrimps and ophiuroids may indeed look for a hiding place, but may additionally receive protection from the stinging cells of the coral (e.g., Bos & Hoeksema, 2015). Moreover, we postulate that associated fauna may consume mucus from a host’s ectodermis or take food particles from a feeding host. Associated organisms may therefore be restricted to only one or few host species. Aquarium observations and experiments may help retrieving more detailed information about the role of associated fauna in relation to their coral habitats.
ACKNOWLEDGEMENTS
We thank J. Bayogan and G. Gumanao (Davao del Norte State College) for providing logistic support during the fieldwork and C. H. J. M. Fransen (Naturalis Biodiversity Center) who confirmed the identification of the commensal shrimps.
LITERATURE CITED
Alamaru A, Brokovich E & Loya Y (2016) Four new species and three new records of benthic ctenophores (Family: Coeloplanidae) from the Red Sea. Marine Biodiversity, 46: 261–279.
Barneah O, Ben-Dov E, Benayahu Y, Brickner I & Kushmaro A (2012) Molecular diversity and specificity of acoel worms associated with corals in the Gulf of Eilat (Red Sea). Aquatic Biology, 14: 277–281.
Benzoni F, Arrigoni R, Stefani F, Reijnen BT, Montano S & Hoeksema BW (2012) Phylogenetic position and taxonomy of Cycloseris explanulata and C. wellsi (Scleractinia: Fungiidae): Lost mushroom corals find their way home. Contributions to Zoology, 81: 125–146.
Böhm T & Hoeksema BW (2017) Habitat selection of the coral-dwelling spinyhead blenny, Acanthemblemaria spinosa, at Curaçao, Dutch Caribbean. Marine Biodiversity, 47: 17–25.
Bos AR (2011) Clownfishes Amphiprion clarkii and A. sandaracinos (Pomacentridae) coexist in the sea anemone Stichodactyla mertensii. Coral Reefs, 30: 369.
Bos AR (2012) Fishes (Gobiidae and Labridae) associated with the mushroom coral Heliofungia actiniformis (Scleractinia: Fungiidae) in the Philippines. Coral Reefs, 31: 133.
Bos AR (2014) Upeneus nigromarginatus, a new species of goatfish (Perciformes: Mullidae) from the Philippines. Raffles Bulletin of Zoology, 62: 745–753.
Bos AR & Gumanao GS (2013) Seven new records of fishes (Teleostei: Perciformes) from coral reefs and pelagic habitats in southern Mindanao, the Philippines. Marine Biodiversity Records, 6: e95.

205
RAFFLES BULLETIN OF ZOOLOGY 2017
Bos AR & Hoeksema BW (2015) Cryptobenthic fishes and co-inhabiting shrimps associated with the mushroom coral Heliofungia actiniformis (Fungiidae) in the Davao Gulf, Philippines. Environmental Biology of Fishes, 98: 1479–1489.
Bos AR & Smits HM (2013) First record of the dottyback Manonichthys alleni (Teleostei: Perciformes: Pseudochromidae) from the Philippines. Marine Biodiversity Records, 6: e61.
Bos AR, Gumanao GS & Salac FN (2008) A newly discovered predator of the crown-of-thorns starfish. Coral Reefs, 27: 581.
Bos AR, Mueller B & Gumanao GS (2011) Feeding biology and symbiotic relationships of the corallimorpharian Paracorynactis hoplites (Anthozoa: Hexacorallia). Raffles Bulletin of Zoology, 59: 245–250.
Bos AR, Gumanao GS, Mueller B & Saceda-Cardoza MM (2013) Management of crown-of-thorns sea star (Acanthaster planci L.) outbreaks: Removal success depends on reef topography and timing within the reproduction cycle. Ocean & Coastal Management, 71: 116–122.
Bruce AJ (1992) Two new species of Periclimenes (Crustacea: Decapoda: Palaemonidae) from Lizard Island, Queensland, with notes of related taxa. Records of the Australian Museum, 44: 45–84.
Butter ME, Wapstra M & van Dijk E (1980) Meandrina meandrites and Emblemariopsis diaphana, first record of an association between a stony coral and a fish, similar to anemone/fish relationships. Bijdragen tot de Dierkunde, 50: 87–95.
Chadwick-Furman NE & Loya Y (1992) Migration, habitat use, and competition among mobile corals (Scleractinia: Fungiidae) in the Gulf of Eilat, Red Sea. Marine Biology, 114: 617–623.
Cooper C, Clode PL, Thomson DP & Stat M (2015) A flatworm from the genus Waminoa (Acoela: Convolutidae) associated with bleached corals in Western Australia. Zoological Science, 32: 465–473.
Froese R & Pauly D (2016) FishBase. World Wide Web electronic publication. www.fishbase.org. (Last accessed 4 May 2017).
Gilbert A, Heintz T, Hoeksema BW, Benzoni F, Fernandez JM, Fauvelot C & Andréfouët S (2015) Endangered New Caledonian endemic mushroom coral Cantharellus noumeae in turbid, metal-rich, natural and artificial environments. Marine Pollution Bulletin, 100: 359–369.
Gittenberger A & Gittenberger E (2011) Cryptic, adaptive radiation of endoparasitic snails: Sibling species of Leptoconchus (Gastropoda: Coralliophilidae) in corals. Organisms, Diversity and Evolution, 11: 21–41.
Gittenberger A & Hoeksema BW (2013) Habitat preferences of coral-associated wentletrap snails (Gastropoda: Epitoniidae). Contributions to Zoology, 82: 1–25.
Gittenberger A, Reijnen BT & Hoeksema BW (2011) A molecularly based phylogeny reconstruction of mushroom corals (Scleractinia: Fungiidae) with taxonomic consequences and evolutionary implications for life history traits. Contributions to Zoology, 80: 107–132.
Gittenberger A, Draisma S, Arbi U, Langenberg V, Erftemeijer P, Tuti Y & Hoeksema BW (2015) Coral reef organisms as bioregion indicators off Halmahera, Moluccas, Indonesia. Aquatic Conservation: Marine and Freshwater Ecosystems, 25: 743–755.
Goatley CHR, González-Cabello A & Bellwood DR (2016) Reef-scale partitioning of cryptobenthic fish assemblages across the Great Barrier Reef, Australia. Marine Ecology Progress Series, 544: 271–280.
Goffredo S & Chadwick-Furman NE (2000) Abundance and distribution of mushroom corals (Scleractinia: Fungiidae) on a coral reef at Eilat, northern Red Sea. Bulletin of Marine Science, 66: 241–254.
Gosliner TM, Behrens DW & Williams GC (1996) Coral Reef Animals of the Indo-Pacific. Sea Challengers, Monterey, 288 pp.
Gumanao GS (2009) Impacts of Marine Protected Areas in the island garden city of Samal, Philippines. Unpublished PhD Thesis, Davao del Norte State College, Panabo City, Philippines, 185 pp.
Haapkylä J, Seymour AS, Barneah O, Brickner I, Hennige S, Suggett D & Smith D (2009) Association of Waminoa sp. (Acoela) with corals in the Wakatobi Marine Park, South-East Sulawesi. Indonesia. Marine Biology, 156: 1021–1027.
Hoeksema BW (1989) Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: Fungiidae). Zoologische Verhandelingen Leiden, 254: 1–295.
Hoeksema BW (1993) Mushroom corals (Scleractinia: Fungiidae) of Madang Lagoon, northern Papua New Guinea: An annotated checklist with the description of Cantharellus jebbi spec. nov. Zoologische Mededelingen, 67: 1–19.
Hoeksema BW (2004) Impact of budding on free-living corals at East Kalimantan, Indonesia. Coral Reefs, 23: 492.
Hoeksema BW (2007) Delineation of the Indo-Malayan Centre of Maximum Marine Biodiversity: The Coral Triangle. In: Renema W (ed.) Biogeography, Time and Place: Distributions, Barriers and Islands. Springer, Dordrecht. Pp. 117–178.
Hoeksema BW (2008) Stony corals (Fungiidae). In: Hoeksema BW & van der Meij SET (eds.) Cryptic marine biota of the Raja Ampat island group. Naturalis, Leiden. Pp. 8–12.
Hoeksema BW (2009) Attached mushroom corals (Scleractinia: Fungiidae) in sediment-stressed reef conditions at Singapore, including a new species and a new record. Raffles Bulletin of Zoology, Supplement 22: 81–90.
Hoeksema BW (2012a) Distribution patterns of mushroom corals (Scleractinia: Fungiidae) across the Spermonde Shelf, South Sulawesi. Raffles Bulletin of Zoology, 60: 183–212.
Hoeksema BW (2012b) Evolutionary trends in onshore-offshore distribution patterns of mushroom coral species (Scleractinia: Fungiidae). Contributions to Zoology, 81: 199–221.
Hoeksema BW (2012c) Mushroom corals (Scleractinia: Fungiidae) of Espiritu Santo (Vanuatu, West Pacific) with the description of a new species. Zoosystema, 34: 429–443.
Hoeksema BW (2014) The “Fungia patella group” (Scleractina, Fungiidae) revisited with a description of the mini mushroom coral Cycloseris boschmai sp. n. ZooKeys, 371: 57–84.
Hoeksema BW & Bongaerts P (2016) Mobility and self-righting by a free-living mushroom coral through pulsed inflation. Marine Biodiversity, 46: 521–524.
Hoeksema BW & Dai CF (1991) Scleractinia of Taiwan. II. Family Fungiidae (with the description of a new species). Bulletin Zoological Institute Academia Sinica, 30: 201–226.
Hoeksema BW & de Voogd NJ (2012) On the run: Free-living mushroom corals avoiding interaction with sponges. Coral Reefs, 31: 455–459.
Hoeksema BW & Farenzena ZT (2012) Tissue loss in corals infested by acoelomorph flatworms (Waminoa sp.). Coral Reefs, 31: 869.
Hoeksema BW & Fransen CHJM (2011) Space partitioning by symbiotic shrimp species cohabitating in the mushroom coral Heliofungia actiniformis at Semporna, eastern Sabah. Coral Reefs, 30: 519.
Hoeksema BW & Gittenberger A (2010) High densities of mushroom coral fragments at West Halmahera, Indonesia. Coral Reefs, 29: 691.
Hoeksema BW & Koh EGL (2009) Depauperation of the mushroom coral fauna (Fungiidae) of Singapore (1860s–2006) in changing reef conditions. Raffles Bulletin of Zoology, Supplement 22: 91–101.
Hoeksema BW & Lane DJW (2014) The mushroom coral fauna (Scleractinia: Fungiidae) of Brunei Darussalam (South China Sea) and its relation to the Coral Triangle. Raffles Bulletin of Zoology, 62: 566–580.

206
Bos & Hoeksema: Fungiidae and associated fish fauna
Hoeksema BW & Putra KS (2000) The reef coral fauna of Bali in the centre of marine diversity. Proceedings of the 9th International Coral Reef Symposium, Bali, 1: 173–178.
Hoeksema BW & ten Hove HA (2014) First record of a Christmas tree worm in a mushroom coral (Loyalty Islands, Southwest Pacific). Coral Reefs, 33: 717.
Hoeksema BW & Yeemin T (2011) Late detachment conceals serial budding by the free-living coral Fungia fungites in the Inner Gulf of Thailand. Coral Reefs, 30: 975.
Hoeksema BW, van der Meij SET & Fransen CHJM (2012) The mushroom coral as a habitat. Journal of the Marine Biological Association of the United Kingdom, 92: 647–663.
Hoeksema BW, Scott C & True JD (2013a) Dietary shift in corallivorous Drupella snails following a major bleaching event at Koh Tao, Gulf of Thailand. Coral Reefs, 32: 423–428.
Hoeksema BW, Waheed Z & Alamaru A (2013b) Out of sight: Aggregations of epizoic comb jellies underneath mushroom corals. Coral Reefs, 32: 1065.
Hoeksema BW, Dekker F & de Voogd NJ (2014) Free-living mushroom corals strike back by overtopping a coral-killing sponge. Marine Biodiversity, 44: 3–4.
Hoeksema BW, van Beusekom M, ten Hove HA, Ivanenko VN, van der Meij SET & van Moorsel GWNM (2017) Helioseris cucullata as a host coral at St. Eustatius, Dutch Caribbean. Marine Biodiversity, 47: 71–78.
Huang D, Licuanan WY, Hoeksema BW, Chen CA, Ang PO, Huang H, Lane DJW, Vo ST, Waheed Z, Affendi YA, Yeemin T & Chou LM (2015) Extraordinary diversity of reef corals in the South China Sea. Marine Biodiversity, 45: 157–168.
Lane DJW & Hoeksema BW (2016) Mesophotic mushroom coral records at Brunei Darussalam support westward extension of the Coral Triangle to the South China Sea waters of Northwest Borneo. Raffles Bulletin of Zoology, 64: 204–212.
Licuanan WY (2009) Guide to the common corals of the Bolinao-Anda reef complex, northwestern Philippines. University of the Philippines Marine Science Institute, Quezon City, 178 pp.
Licuanan WY & Capili EB (2004) New records of stony corals from the Philippines previously known from peripheral areas of the Indo-Pacific. Raffles Bulletin of Zoology, 52: 285–288.
Nemenzo F (1986) Guide to the Philippine Flora and Fauna: Corals, Volume 5. Natural Resources Management Center & University of the Philippines, Manila, 273 pp.
Owada M & Hoeksema BW (2011) Molecular phylogeny and shell microstructure of Fungiacava eilatensis Goreau et al., 1968, boring into mushroom corals (Scleractinia: Fungiidae), in relation to other mussels (Bivalvia: Mytilidae). Contributions to Zoology, 80: 169–178.
Phillips DJH & Pullin RSV (1987) Siokunichthys nigrolineatus (Syngnathidae) found on Fungia sp. Copeia, 2: 509–511.
Ponti M, Fratangeli F, Dondi N, Reinach MS, Serra C & Sweet MJ (2016) Baseline reef health surveys at Bangka Island (North Sulawesi, Indonesia) reveal new threats. PeerJ, 4: e2614.
Schiemer L, Niedermüller S & Herler J (2009) The influence of colony size and coral health on the occupation of coral-associated gobies (Pisces: Gobiidae). Coral Reefs, 28: 137–142.
Scott PJB (1987) Associations between corals and macro-infaunal invertebrates in Jamaica, with a list of Caribbean and Atlantic coral associates. Bulletin of Marine Science, 40: 271–286.
Sih J & Chouw J (2009) Fish and whips: Use of gorgonians as a habitat by the large whipcoral goby, Bryaninops amplus (Larson). Raffles Bulletin of Zoology, Supplement 22: 145–157.
Stella JS, Pratchett MS, Hutchings PA & Jones GP (2011) Coral-associated invertebrates: Diversity, ecology importance and vulnerability to disturbance. Oceanography and Marine Biology: An Annual Review, 49: 43–104.
Tornabene L, Ahmadia GN, Berumen ML, Smith DJ, Jompa J & Pezold F (2013) Evolution of microhabitat association and morphology in a diverse group of cryptobenthic coral reef fishes (Teleostei: Gobiidae: Eviota). Molecular Phylogenetics and Evolution, 66: 391–400.
van der Meij SET & Hoeksema BW (2013) Distribution of gall crabs inhabiting mushroom corals on Semporna reefs, Malaysia. Marine Biodiversity, 43: 53–59.
van der Meij SET, Fransen CHJM, Pasman LR & Hoeksema BW (2015) Phylogenetic ecology of gall crabs (Cryptochiridae) as associates of mushroom corals (Fungiidae). Ecology and Evolution, 5: 5770–5780.
Veron JEN (2000) Corals of the World, Volume 2. Australian Institute of Marine Science, Townsville, 429 pp.
Veron JEN & Fenner D (2000) Corals (zooxanthellate Scleractinia) of the Calamianes Islands, Palawan Province, Philippines. RAP Bulletin of Biological Assessment, 17: 24–26, 66–80.
Veron JEN & Hodgson G (1989) Annotated checklist of the hermatypic corals of the Philippines. Pacific Science, 43: 234–287.
Veron JEN, Stafford-Smith M, DeVantier L & Turak E (2015) Overview of distribution patterns of zooxanthellate Scleractinia. Frontiers in Marine Science, 1: 81.
Waheed Z & Hoeksema BW (2013) A tale of two winds: Species richness patterns of reef corals around the Semporna peninsula, Malaysia. Marine Biodiversity, 43: 37–51.
Waheed Z & Hoeksema BW (2014) Diversity patterns of scleractinian corals at Kota Kinabalu, Malaysia, in relation to exposure and depth. Raffles Bulletin of Zoology, 62: 66–82.
Waheed Z, Benzoni F, van der Meij SET, Terraneo TI & Hoeksema BW (2015a) Scleractinian corals (Fungiidae, Agariciidae and Euphylliidae) of Pulau Layang-Layang, Spratly Islands, with a note on Pavona maldivensis (Gardiner, 1905). ZooKeys, 517: 1–37.
Waheed Z, van Mil HGJ, Hussein MAS, Jumin R, Ahad BG & Hoeksema BW (2015b) Coral reefs at the northernmost tip of Borneo: An assessment of scleractinian species richness patterns and benthic reef assemblages. PLoS ONE, 10(12): e0146006.
WoRMS Editorial Board (2016) World Register of Marine Species. Available from http://www.marinespecies.org at VLIZ, doi:10.14284/170. (Accessed 23 July 2016).