myopia of selection: does organizational adaptation limit ... · there is reason to believe that...
TRANSCRIPT

This paper develops and tests a model of the effective-ness of selection processes in eliminating less fit organi-zations from a population when organizations are under-going adaptive change. Stable organizational traits, suchas a search strategy or routine, do not imply that an orga-nization’s performance will remain stable over time orthat cross-sectional differences in performance will per-sist. These properties create the possibility that popula-tion-level selection processes will be inefficient in thatorganizations with potentially superior long-run perfor-mance will be selected out. We theorize that organization-al-level adaptation often results in fluctuations in currentperformance across time. These fluctuations may attenu-ate the degree to which current performance differencesamong organizations are indicative of future perfor-mance. As a consequence, search strategies that generatesystematically different performance trajectories, even ifthey share a common long-run outcome, will generatediffering survival rates. These ideas are explored using aformal simulation model employing the framework of NKperformance landscapes. Our central finding is that selec-tion may be systematically prone to errors and that theseselection errors are endogenous to, and differ markedlyacross, firms’ search strategies.•The past 40 years has seen the emergence of significantinterest in examining the population dynamics of organiza-tions from an evolutionary perspective (Campbell, 1965; Han-nan and Freeman, 1977; Nelson and Winter, 1982). A defin-ing feature of the evolutionary perspective is that selectiondisproportionately removes organizations that are less fit. Yetthere is reason to believe that this selection process may notbe effective in eliminating organizations undergoing change.First, selection does not operate directly on relatively stableorganizational traits, such as strategies or routines that arefundamental to firm-level heterogeneity. Instead, selectionoperates on the manifestations of those traits in terms ofobservable performance outcomes such as productivity,patents, new products, product quality and cost, marketshare, sales, or profits. Moreover, even stable organizationaltraits, including an organization’s search strategy, may lead toperformance manifestations that are highly variable. To theextent that organizations’ stable traits lead to differences inthe nature of learning, they may give rise to significant differ-ences in the reliability of performance over time. Conse-quently, organizational populations may experience differentsurvival patterns in the selection process not merely basedon some inherent superiority of one form or another but alsoas influenced by the dynamics of change that different formsgenerate over time.
Second, though change processes and their effects on per-formance play out over time, selection processes are funda-mentally myopic in that they cannot “see” those futureeffects. Selection is the outcome of a complex web of atom-istic and, to some degree, also inherently myopic decisionsabout the distribution of resources across firms. The quintes-sential example of myopia is that of individual purchase deci-sions in product markets, where products’ characteristics,such as quality and price, drive consumers’ choice and hence
© 2007 by Johnson Graduate School,Cornell University. 0001-8392/07/5204-0586/$3.00.
•Financial support from Mack Center forTechnological Change and Innovation atthe Wharton School is gratefully acknowl-edged. We have benefited from com-ments on prior drafts by Sendil Ethiraj,Anne Marie Knott, Myles Shaver, NicolajSiggelkow, and Sidney Winter, as well asreviewers and participants at the Acade-my of Management Meeting, theBYU/Utah Winter Strategy Conference,Informs Annual Meeting, and seminarparticipants at U.C. Berkeley, the Univer-sity of Maryland, University of Michigan,University of Pennsylvania, and the Uni-versity of Wisconsin. Finally, we thankHuggy Rao and three anonymous review-ers for a helpful set of comments on aprior draft.
Myopia of Selection:Does OrganizationalAdaptation Limit theEfficacy of PopulationSelection?
Daniel LevinthalUniversity of PennsylvaniaHart E. PosenUniversity of Michigan
586/Administrative Science Quarterly, 52 (2007): 586–620

selection outcomes for the product. Even forward-lookingactors in capital markets are myopic decision makers in thatthey must construct expectations of future performance sole-ly based on such indicators of past and current performanceas sales, profits, or patent filings. One important conse-quence of the myopia of selection is that even if selection iseffective in removing inferior organizations at one point intime, it may be ineffective over time in that it may removeorganizations that, had they survived, would have gone on todo well.
The question is whether there are conditions under whichmyopic selection can identify adapting organizations at inter-mediate points in time, while they are still in the midst ofchange, that will be superior in the long run. The existing lit-erature provides some important rationales as to why selec-tion processes may fail in this respect, indicating why inferiororganizations, populations, or technologies may dominate theset of survivors (Barron, West, and Hannan, 1994; Carroll andHarrison, 1994; Barnett and Burgelman, 1996; Barnett, 1997).Three broad classes of answers have been proposed. First,selection does not act on a single dimension of performancebut, in fact, on a multitude of potential performance criteria.Thus the outcome of the selection process can be as much afunction of political and social competition as it is technicalcompetition on the merits of alternatives (Anderson andTushman, 1990). Second, organizational slack acts as a bufferto stave off failure even when organizations are weak from atechnical efficiency standpoint (Levinthal, 1991b; Barnett,Greve, and Park, 1994; Barron, West, and Hannan, 1994).Third, path dependence can arise from network externalities(David, 1985; Arthur, 1989) whereby relative market shares ofalternative forms or technology influence selection forces,and thus outcomes at a given point in time are in part a func-tion of prior choices and outcomes (Barron, West, and Han-nan, 1994; Carroll and Harrison, 1994; Barnett and Burgel-man, 1996; Barnett, 1997).
A different reason, however, is that selection may be system-atically prone to errors, which is suggested by the results ofCarroll and Harrison’s (1994) examination of the extent towhich selection processes are historically efficient. Buildingon March and Olsen (1989), Carroll and Harrison (1994: 720)framed the issue as the extent to which “organizationalarrangements observed at a particular point in time representthe unique outcome of some systematic process such ascompetition.” Focusing on differential selection among typesof organizations, they employed a computational model ofthe evolution of two populations of fully inert organizationsunder an ecological selection regime based on legitimationand competition. They concluded that deviations from histori-cal efficiency may arise from differences in organizations’order of entry into a population due to the positive feedbackeffects of legitimation. As a result, it is possible for a popula-tion of inferior organizations to come to dominate the set ofsurviving firms if they enter first.
Carroll and Harrison (1994) focused on entry order in theirmodel. Alternatively, as developed here, one can start withthe assumption that organizations are engaged in a learning
587/ASQ, December 2007
Myopia of Selection

process and thus exhibit performance manifestations thatvary over time. The unfolding of an organization’s particularhistory is then a function of both its idiosyncratic foundingconditions and the strategies that guide its learning. Strate-gies that lead to higher variance in performance over time arelikely to increase the extent to which selection is prone to errin selecting out organizations that would have gone on toachieve a high level of performance. In turn, selection errors,in which inferior firms survive selection, would decrease theaverage performance of the set of surviving firms. Whenorganizations within a population are distinguished by strate-gies that differ in reliability and performance, an importantelement of selection consists of sorting among organizationsin a single population. Under these conditions, differencesacross populations in reliability over time alone are sufficientto generate outcomes in which the population with inferiormean long-run performance, but higher reliability, dominatesthe set of surviving firms.
In the model developed in this paper, we highlight the centralrole of reliability in performance. In contrast to ecologicalmodels in which selection favors reliability (Hannan and Free-man, 1977, 1984), however, our model imposes no a priorirelationship between reliability and mortality. Rather, the roleof reliability emerges endogenously from a model in whichfirms that are subject to selection employ strategies that areinert, but these fixed search strategies guide organizationallearning.
Important stable attributes of organizations such as searchstrategies or routines may differ in their contribution not onlyto organizational performance at a given point in time butalso to the variance across time. In turn, these stable traitsdifferentially affect the extent to which selection is prone toerrors. We denote this effect on the efficacy of selection as“selectability,” which is an organization-level construct thatmeasures the information content embodied in current per-formance as a critical determinant of the efficacy of selectionin identifying organizations that will be superior in the longrun. For organizations that exhibit high levels of selectability,intermediate performance manifestations will provide superi-or information about their long-run performance, and conse-quently, selection in populations of such organizations will bemore effective over time. Thus, even when a set of searchstrategies across organizations shares common long-runproperties, the dynamics by which organizations reach thislong-run performance level have important implications forthe selectability among organizational populations and theefficacy of selection as a mechanism of population-leveladaptation.
Questions about the nature of organizational change and itsconsequences for the efficacy of selection, though relativelyunexplored, are not new. Campbell (1960, 1965), in laying thefoundation of evolutionary perspectives in the social sci-ences, suggested one important factor driving the efficacy ofselection. He argued that “if discovery or expansions ofknowledge are [to be] achieved, blind variation is requisite”;blind implies that “specific correct trials are no more likely tooccur .|.|. than specific incorrect trials” (Campbell, 1982: 86).
588/ASQ, December 2007

Nevertheless, the organizational analogue differs from its bio-logical foundations in a critical way: organizations engage inadaptive trial-and-error search strategies that are intended toenhance their survival prospects (Simon, 1955; Cyert andMarch, 1963; Nelson and Winter, 1982). Simon (1969: 95), incontrast to Campbell, argued that such “trial and error is notcompletely random or blind.” This classic debate highlights acentral but unresolved question related to the efficacy ofselection as a mechanism of population-level adaptation: Isthe efficacy of selection invariant to the characteristics of theprocesses (e.g., blind vs. non-blind variation) that guide orga-nizational change?
To gain insight into the effect of differential reliability on theefficacy of selection, we implemented a formal agent-basedmodel using an NK methodology (Kauffman, 1993; Levinthal,1997; Rivkin, 2000). This formal structure offers a number ofcritical advantages over other forms of theorizing. First, ourtheory involves the intersection of three central features oflearning models: partially stochastic outcomes, path depen-dence, and interdependence among the adapting entities.Under these conditions, verbal theorizing is inadequate forthe complexity of theorizing, and closed-form analytical mod-els are nearly impossible to implement (Lane, 1993). Second,our theory is multilevel in that it involves adaptation at boththe individual and population levels. The agent-based modelallows us both to examine individual histories and to under-stand how these histories aggregate to population-level out-comes. Finally, agent-based simulation methods allow us tointegrate existing theory into the model. For example, ratherthan treat variance across time as exogenous, our formalmodeling approach allows differences in variance to emergeas a consequence of organizational choices in a manner con-sistent with extant theory.
We started with the premise that organizations are engagedin trial-and-error adaptation intended to enhance their perfor-mance in the face of selection pressure. We make three cen-tral assumptions in constructing our model: (1) the character-istics of the adaptation process are determined byorganizations’ search strategies, which we treat as fixed overtime; (2) selection is myopic, occurring on the basis of perfor-mance at one point in the midst of this adaptation process;and (3) the efficacy of selection is assessed based on real-ized future performance. We began by examining a baselinemodel in which we assessed the efficacy of selection acrossorganizations employing a very simple, fully blind adaptationstrategy. Over the subsequent four experiments, we com-pared the baseline model to alternative implementations oftrial-and-error search strategies that varied in reliability.
SELECTION ACROSS ADAPTIVE ORGANIZATIONS
Our concern is with the efficacy of selection as a mechanismof population-level adaptation. Selection acts by removinginferior organizations from the population, but evolutionarytheory does not imply that selection processes lead to opti-mal outcomes (Winter, 1964). As Simon (1991: 166) noted,“.|.|. evolutionists sometimes talk about survival of the fittest.But in fact, natural selection only predicts that survivors will
589/ASQ, December 2007
Myopia of Selection

be fit enough, that is, fitter than their losing competitors.”While the term “fitter” implies non-random elimination ofless-fit organizations, it leaves open a wide range of possibleoutcomes. Selection may lead to less than optimal outcomesfor two reasons. First, there may be errors in selection at anypoint in time. For example, consider ten organizations, onlythree of which will survive selection. Ranking the organiza-tions in terms of performance, if selection is perfectly effec-tive, then only the three top organizations should survive. If,in contrast, selection is error prone, such that the top organi-zation fails and those ranked second through fourth survive,then selection will have increased average fitness but wouldnot have been perfectly effective. Second, less salient in thisdiscourse, but perhaps more important in an organizationalcontext, is that the entities over which selection is operatingmay themselves be changing over time. In part, this lack ofattention in the literature may stem from the importance puton stable organizational traits in evolutionary arguments(Cohen et al., 1996). Yet, as the biological analogy suggests,a fixed genetic structure need not imply a fixed phenotype.Consider the obvious case of human development from frag-ile newborn to robust toddler, sprouting adolescent, andmature and eventually elderly adult. Clearly, selection at inter-mediate points in this developmental process, even if it isoptimal in terms of removing the weakest individuals at agiven point in time, may not be optimal over time because itremoves slower developing individuals who may eventuallyexhibit superior attributes. In the context of selection acrossadaptive organizations, even optimal selection in the shortterm need not generate optimal long-run outcomes.
The fact that selection forces operate on entities that them-selves are undergoing a dynamic process of adaptation raisesan important issue about the efficacy of selection in an orga-nizational context. A key factor underlying firms’ heterogene-ity and, in turn, forming the basis for differential selection, isthe fact that organizations engage in different strategies as tohow they go about establishing an economically viable entity.It is well recognized that distinct strategies may imply differ-ential long-run performance and, as a consequence, mayhave a significant impact on organizations’ survival prospects.It is less well recognized that distinct strategies, becausethey imply the possibility that they will unfold differently overtime, may also have an impact on organizations’ survivalprospects independent of long-run performance.
In our modeling, we focus on the implications of organization-al adaptation for the efficacy with which selection processesidentify organizations that will be superior in the long run andremove those that will be inferior. The existing literature inpopulation ecology and organization theory more broadly hasfocused significant attention on a related but different issue,the implications of organizational change for the mortality riskof individual firms. In doing so, this literature has identifiedtwo broad classes of mortality risks: the first related to riskassociated with the content of change and the second relat-ed to risk in the process of change. In both cases, organiza-tional change, even if it has long-run positive performance
590/ASQ, December 2007

implications, may have short-run negative performance impli-cations that increase mortality.
Risks in the content of change reflect the problems faced byboundedly rational firms in identifying a suitable destinationstate, or desired set of outcomes. Change from the existingstate to a new state entails a significant mortality risk, as theinformation required to evaluate the performance conse-quences of such a change may not be available. Moreover,change to a very distant destination state (exploration) entailsmore risk than change to an adjacent state (exploitation orlocal search), as adjacent states are more likely to result insimilar performance. Thus change leads to instability becauseorganizations implement changes that may have detrimentalperformance. The instability associated with risk in the con-tent of change arises because of the difficulty of evaluatingalternatives without actually implementing them. At thestable extreme, “off-line” evaluation of alternative destinationstates (Gavetti and Levinthal, 2000), using cognitive represen-tations to evaluate alternatives without implementing them,reduces content risk by eliminating the worst alternativesprior to exposing them to selection forces. At the unstableextreme, content risk is greater when search is blind,because “specific correct trials are no more likely to occur.|.|. than specific incorrect trials” (Campbell, 1982: 86). Whensearch is blind, alternatives must be implemented andsubjected to selection in order to assess their value. Contentrisk thus reflects the intelligence embodied by changeefforts—the ability (or inability) to evaluate alternatives andselect only those that have positive and immediate effects onperformance.
Process risks reflect the problems that arise because of theembedded nature of organizational structures. Theory distin-guishes between changes in core and peripheral organization-al elements (Thompson, 1967; Hannan and Freeman, 1984;Levinthal, 1997). Core elements of structure are, by defini-tion, those that are highly interdependent with other organi-zational elements. In the microcomputer industry, Romanelliand Tushman (1994: 1147) argued, organizational culture,strategy, structure, power distributions, and control systemsconstitute core elements. In Silicon Valley start-ups, Baron,Burton, and Hannan (1999) studied founders’ “organizationalblueprints” as a core organizational feature that dictates theextent of formal organizational structures that serve as con-trol and coordination mechanisms. Although changes inperipheral elements are considered to be feasible, researchhas called into question the survival benefit of changes incore elements (Singh, House, and Tucker, 1986; Haveman,1992; Amburgey, Kelly, and Barnett, 1993; Barnett and Car-roll, 1995; Dobrev, Kim, and Carroll, 2003). The risk of changein core organizational features arises because changerequires significant adjustments across a wide variety ofinterdependent organizational elements, and this in turnengenders instability. Even small changes in core elementsmay set off a cascade of related changes that disrupt existingorganizational linkages, disturb communication channelsacross organizational boundaries, and disable routines (Han-nan and Freeman, 1984). The risk of mortality is further
591/ASQ, December 2007
Myopia of Selection

increased because content and process issues interact—interdependence across core elements of the organizationmakes it more difficult to evaluate the performance conse-quences of a change in content.
One strategy organizations use to manage the problem of thecomplexities of change is to engage in the change processessequentially rather than in parallel, making individual changesto organizational content in one domain, while holding all elseconstant. That said, at higher levels of interdependence,firms may need to make concurrent changes across multipleorganizational elements (Rivkin, 2000; Sorenson, 2003), andattempts to do otherwise introduce significant organizationalinstability. For example, Sorenson (2003), in a study of theeffects of the interdependence associated with the co-pro-duction of components among computer workstation manu-facturers, found that interdependence retards performanceand increases mortality in all but high-velocity environments.Ethiraj and Levinthal (2004), in a simulation study of organiza-tional adaptation, demonstrated that reciprocal interdepen-dence (Thompson, 1967) leads to significant instability in theface of local search, which can only be overcome by impos-ing a strict hierarchy.
To the extent that organizational age (and to a lesser extent,size) is a suitable proxy for routines and elaborated organiza-tional structure, empirical work has documented a correlationbetween age and mortality in the face of change (Delacroixand Swaminathan, 1991; Kelly and Amburgey, 1991;Amburgey, Kelly, and Barnett, 1993; Miller and Chen, 1994).Highlighting the role of organizational structures and routines,Dobrev, Kim, and Carroll (2003) demonstrated, in a study ofthe U.S. auto industry, that change requiring a reorientationof structures and the introduction of novel routines increasesthe risk of failure, but change building on existing structuresand routines does not. That said, results in many studieshave been mixed (Baum and Amburgey, 2001), in partbecause age (and size) is a proxy not only for structure butalso for accumulated resources. For example, research hashighlighted the strategic choices that older and larger firmsmake to buffer themselves from failure (Miner, Amburgey,and Stearns, 1990; Barron, West, and Hannan, 1994).
The organizations literature in general and the populationecology literature in particular thus suggest that change itselfexposes a firm to risk (Hannan and Freeman, 1984; Miner,Amburgey, and Stearns, 1990; Haveman, 1992; Amburgey,Kelly, and Barnett, 1993) arising from the difficulty of identify-ing a suitable destination state (content risk) and interdepen-dence among elements of the elaborated structure (processrisk). The causal chain reflects the argument that content andprocess issues give rise to reductions in the reliability of per-formance over time that, in turn, increase the risk of mortali-ty. Still unaddressed, however, is the related question ofwhat the consequences of reduced reliability would be forthe efficacy of selection as a driver of population-leveladaptation.
If selection is to be effective, it must be able to identify orga-nizations at intermediate points in time that will be superior
592/ASQ, December 2007

in the long run. This challenge reflects the difficulties in esti-mating future performance. Although selection processesmay operate through a variety of mechanisms, inevitably,selection must be based on current and past performance.Even if selection processes such as financial markets are for-ward looking, information about future performance expecta-tions can be based only on current and past data and out-comes. For instance, the early performance of a newventure-capital-backed technology firm—such as the extentto which it has achieved its early sales milestone—can beused to assess its potential. Similarly, in educational con-texts, standardized testing is used to assess students’ futureeducational performance. In both cases, the intermediatemanifestations of performance before the adaptation processis complete may provide only a noisy signal about long-termperformance. More generally, it is unclear to what extent theintermediate performance manifestations of adapting organi-zations provide a good signal about future performance.
In our modeling, we focus on how heterogeneity across orga-nizations in reliability during the adaptation process affectsthe quality of the information on which selection acts and, inturn, the efficacy of selection. We illustrate how the informa-tion content of current performance as it applies to predictingfuture performance decreases as the instability with whichadaptation proceeds increases. The structure of the modelclosely reflects its theoretical foundation in the populationecology and organizations literature. Organizations engage ina process of local search for alternatives (Cyert and March,1963; Nelson and Winter, 1982; Stuart and Podolny, 1996). Insuch a process, the set of potential destination states is limit-ed to those near to the origin state, and the correlation inperformance over time is high relative to exploratory search.But even in local search processes, organizational searchchoices affect the adaptive dynamics. Building on the prior lit-erature discussed above, the reliability of adaptation is, inpart, jointly the result of differences across firms in howsearch is pursued with regard to (1) the mode by which alter-natives are evaluated, which affects the degree of contentrisk, and (2) the imposition of organizational structure (inte-grated vs. functional), which affects the degree of processrisk. The imposition of structure may be further refined interms of how effort is allocated over time across elements ofstructure (parallel vs. sequential allocation of effort). Takentogether, we denote these choices collectively as a searchstrategy, a coherent set of rules governing learning. We havechosen to model the mode of evaluation and the nature oforganizational structures because they closely reflect the the-oretical foundations of the population ecology literature’sfocus on reliability, although other elements of search strate-gies, not modeled here, may also contribute to differentialreliability.
First, evaluation to assess the performance impact of analternative may be conducted on-line or off-line. For on-lineevaluation, an organization must implement that alternative inorder to assess its implications for performance. As such, on-line evaluation reflects the content risk associated with thedifficulties embodied in identifying a destination state for
593/ASQ, December 2007
Myopia of Selection

change. Off-line evaluation allows an organization to imple-ment only those alternatives that are believed to lead to per-formance improvements. Off-line evaluation decreases thecontent risk of mortality but does not necessarily eliminate it,because the a priori screening of alternatives may well beimperfect. Although, on average, both methods of evaluationallow organizations to proceed in the direction that leads tomarginal performance gains, off-line evaluation does so withgreater stability.
Second, search strategies can differ in how organizationsstructure the problem-solving search effort. We consideredan underlying task structure in which the problem facingorganizations can be divided into two sub-problems, such asresearch and development (R&D) and marketing, or domesticand international business. Given the important role of organi-zational structure in structural inertia theory, we examinedhow different structures affect the degree of reliability overtime during adaptation. The simplest organizational structureis fully integrated. In this structure, the organization does nottreat the problem as decomposable. For example, there areno separate marketing and R&D departments. Instead,search is structured in an integrated manner across both sub-problems. In this sense, firms search across one large prob-lem landscape. Alternatively, organizations may employ afunctional structure consisting of two functional units, suchas separate R&D and marketing departments. Each functionalunit focuses on a particular sub-problem and as such, firmsengage in local search on two distinct sub-problem land-scapes.
If an organization has a functional structure, a set of searchrules governing the allocation of resources across the func-tional units must be considered. Two fundamentally differentsearch strategies are possible: parallel and sequential. In par-allel search strategies, the organization allocates its searcheffort evenly across both units. In sequential search strate-gies, it first devotes all search effort to one unit, and onlystarts searching across the other after it has made a certaindegree of progress on the first. As an example of sequentialsearch, entrepreneurial firms in the technology sectors oftenaddress the problem of marketing only after technologicalsearch has yielded a working prototype. Parallel searchenables integrated trial-and-error learning across the entiresystem, which tends to reveal problems more rapidly and, indoing so, stimulates learning. At the same time, this tight-coupling approach tends to decrease the rate of early perfor-mance improvement. This problem arises as adaptation inone domain, which improves performance locally, negativelyaffects performance in the other domains and thus leads toremedial action. In contrast, sequential search purposefullysuppresses learning in one domain in order to enhance learn-ing in another (Baldwin and Clark, 2000), producing superiorearly performance gains in one unit at the expense ofreduced stability over time (i.e., a non-monotonic learningcurve).
The challenge to the efficacy of selection across organizationsin the midst of adaptation is identifying the organizations thatwill have superior long-run performance. The selection
594/ASQ, December 2007

such that the rate of performance improvement varies con-siderably over time. To the extent that adaptation is unreli-able, selection is less effective at identifying organizationsthat will be superior in the long run. In this sense, the effica-cy of selection is endogenous to the organizational search
595/ASQ, December 2007
Myopia of Selection
process is significantly confounded by organizational hetero-geneity in search strategies that affect both long-run perfor-mance and the reliability of performance over time as theorganization moves toward the long-run performance out-come. In particular, the path of adaptation over time can becharacterized by two basic types of reliability exhibited byorganizational learning curves: short-wave reliability and long-wave reliability, as depicted by the grey learning curves infigure 1. While short-wave reliability (figure 1, A) reflectsnear-term performance cycling, long-wave reliability (figure1, B) reflects non-monotonicity in performance improvement
Figure 1. Learning curves and intertemporal correlation in performance.
A: Short-wave reliability
Selection
Selection
Per
form
ance
Per
form
ance
Time (t) T
Time (t) T
B: Long-wave reliability

strategies that drive differential adaptation trajectories. In ourcomputational model of adaptation, we examine systematicdifferences in reliability across organizations and the effect ofthose differences on the information available in the selectionprocess.
COMPUTATIONAL MODEL OF ADAPTATION IN THEPRESENCE OF INTERMEDIATE SELECTION
We developed an agent-based simulation model to examinethe efficacy of selection across heterogeneous organizationsundergoing adaptation. We built our model on two centralassumptions, discussed above: (1) that adaptation is theresult of local search among policy choices guided by searchstrategies, and (2) selection occurs on the basis of currentperformance outcomes. We employed as an analytical toolthe NK model developed by Kauffman (1993), which hasfound growing acceptance in the management literature instudies of search (Levinthal, 1997), imitation (Rivkin, 2001),organizational structure (Rivkin and Siggelkow, 2003), andorganizational architecture (Ethiraj and Levinthal, 2004). Thebasic model consists of a structure for defining a fitness land-scape (the problem space to be searched), search rules thatguide adaptation, and a selection mechanism. Each of theseelements is briefly characterized below and a detailed mathe-matical treatment is included in the Appendix.
Landscape Structure and Fitness Functions
Our primary objective was to explore how the process ofadaptation affects the efficacy of selection. We modeled atask environment consisting of two interconnected sub-prob-lem landscapes (e.g., R&D and marketing; domestic andinternational activity, etc.). The payoff to a set of choices inone problem domain, or landscape, depends on the choicesmade in the other. We assumed that the boundaries of thesetwo landscapes, along with the level of interdependenciesbetween them, were defined exogenously and thus were notsubject to managerial discretion in the short term. Our mod-eling of this task environment is related to recent efforts tostudy issues of organizational design (Rivkin and Siggelkow,2003; Siggelkow and Levinthal, 2003; Ethiraj and Levinthal,2004), in which interactions across sub-problems (land-scapes) play a central role.
A set of choices about how an organization wishes to com-pete and operate is represented as a location in policy spacecharacterized by a vector of policy choices, N1 = (a1
1, .|.|. , a1n1),
and N2 = (a21, .|.|. , a
2n2), of lengths n1 and n2, respectively,
where the superscript represents the landscape (sub-prob-lem), and the subscript represents the policy choice. We leteach policy choice correspond to a particular binary decisionvariable, where, for example, the choice of 1 might imply thechoice of using flash memory rather than a disk drive in thedesign of a new portable music player. Consider the examplein figure 2. An organization’s location on each landscape isrepresented by vectors of length n1 = n2 = 4, such that N1 =a1
1a12a
13a
14 and N2 = a2
1a22a
23a
24. Thus there are 2n1 = 2n2 = 24 pos-
sible combinations of policy choices in each space. The firm’sposition in the overall landscape can be represented by the
596/ASQ, December 2007

vector N of length n = n1 + n2, such that N = {N1, N2} = (a11,
.|.|. , a1n1, a2
1, .|.|. , a2n2).
Interactions between policy choices can occur both within alandscape and across landscapes (Simon, 1962). As an exam-ple, a within-sub-problem interaction in R&D would be theinteraction between a laptop’s weight and screen size, whilean across-sub-problem interaction might occur between R&Dand marketing in the form of an interaction between a lap-top’s weight and choice of consumer- versus business-saleschannels. Interactions within a landscape are characterized bythe vector KW
j (length kW ) of policy choices with which policya1
j (or a2j ) interacts on its own landscape (where j indexes the
policy choice). The example in figure 2 demonstrates theinteraction space in which kW = 2 (landscape 1 in the upper-left quadrant and landscape 2 in the lower right, which repre-sent two sub-problems). In the example, the payoff to policya1
1 (choice of 0 or 1) in landscape 1 is a function of the choic-es made for the policies in the vector KW
1 = (a12, a
13), this rela-
tionship between policies is characterized by an “x” in theappropriate cell. In the example in figure 2, if we let N1 =a1
1a12a
13a
14 = 1110, then KW
1 = (1,1), and the fitness contributionof policy choice a1
1 is a function of selecting a 1 for bit a11, as
well as selecting a12 = 1 and a1
3 = 1. Changing to a13 = 0 would
alter the fitness contribution made by the choice of a11 = 1,
while the policy choice setting for a14 = 1 or 0 has no effect
on the fitness contribution of a11.
Interactions across landscapes are characterized by the vec-tor KB
j (length kB) of policy choices in the other landscape that
597/ASQ, December 2007
Myopia of Selection
Figure 2. Example of a set of firm choices across two landscapes (N1 and N2) and the interactions betweenthem.*
a11 a1
2 a13 a1
4
a11 x x x o
a12 x x x o
a13 x x x o
a14 x x x o
o x x x a21
o x x x a22
o x x x a23
o x x x a24
a21 a2
2 a23 a2
4
Landscape 1 (N1)
Landscape 2 (N2)
*The x’s represent interactions across choices within a landscape (sub-problem) while the o’s representinteractions across landscapes. For example, the payoff to choice a1
1 is a function of the choices for a13, a
14, but
also a22.

affect the performance contribution of a policy choice in thefocal landscape (the “o”s in the upper right and lower left).That is, interdependencies across activities on the two land-scapes complicate the learning process as search activity inone domain both informs and affects progress in the other. Inthe example, with kB = 1, the fitness contribution of a1
1 is alsoa function of the choices made for the policies in the vectorKB
1 = (a22). In total, the payoff associated with an individual pol-
icy choice is conditional on all policy choices (within andacross landscapes) with which it interacts. In our example,the payoff associated with policy choice 1 in landscape 1, a1
1,is �1
1(a11|KW
1 , KB1) = �1
1 (a11|a1
2, a13, a2
2). The payoffs to each ofthe 21+kW+kB distinct vectors associated with a policy choiceare generated by a draw from a uniform distribution u[0,1].The fitness associated with a position on landscape 1, N1, issimply the average of the fitness contributions of each of theindividual policy choices, such that
�1j =
n1
1n1
�j=1
�1j (a
1j |KW
j , KBj ).
There is a analogous expression for the fitness associatedwith N2. The total fitness is the average fitness across
1landscapes, such that �j = 2(�1
j + �2j ).
Rules Governing Search
Search takes the form of local trial-and-error learning, whichcorresponds to “hill climbing” in a multiple-peaked perfor-mance landscape (Levinthal, 1997). Search is performed byevaluating the consequences for fitness of changing a ran-domly selected policy choice aL
j where L is landscape 1 or 2.Search strategies can differ both in how alternatives are eval-uated and in the imposition of organizational structure.
Evaluation reflects the mechanism by which the alternativesidentified in the trial-and-error search process are assessedand, in particular, the difference between blind and non-blindsearch. On-line evaluation reflects a context in which it is notpossible to determine the value of a change in a single policyelement without implementing the change (Gavetti andLevinthal, 2000). Thus “specific correct trials are no morelikely to occur .|.|. than specific incorrect trials” (Campbell,1982: 86). As a consequence, fitness at the end of the periodmay be either higher or lower than at the beginning of theperiod. If fitness is lower, then in the next period, the organi-zation returns to its earlier position and attempts to testanother policy choice. Once all single policy choice changeshave been examined and found to be inferior, search ceases.In contrast, off-line evaluation is such that, if performance isimproved by making the change, it is implemented (Gavettiand Levinthal, 2000), otherwise the potential policy change isdiscarded. As such, off-line evaluation entails search in whichincorrect trials are not subjected to population selection, andas such, off-line search is non-blind. Off-line evaluation is pos-sible when the efficacy of a choice can be determined usingsuch things as cognitive evaluation, computer models, struc-tured experiments, animal models, or prototypes. For
598/ASQ, December 2007

instance, a particular combination of chemicals may be evalu-ated in vivo in a laboratory, without having to test it directlyby selling the chemical in the product market.
Organizational structure reflects differences in the set of con-straints imposed on the search process. We implementedtwo alternative structures. The first structure is integrated. Inthis structure, an organization chooses a policy choice toevaluate from the entire set of N = {N1, N2} = (a1
1, .|.|. , a1n1, a2
1,.|.|. , a2
n2) policies, with evaluation based on the change in fit-ness across the global landscape. The second structure isfunctional, in which the organization chooses a multidivisionalstructure that matches that of the task environment. Thusthe organization is divided into two functional subunits, withsearch constrained to within-unit policy changes. Whensearch is conducted in unit 1, only policy choices N1 =a1
1a12a
13a
14 may be examined, and only the consequences for
local unit 1’s performance are evaluated. As noted earlier, thefunctional structure entails two further sets of choices. Effortcan be allocated in parallel or sequentially. In parallel alloca-tion, in each period, the organization attempts one searcheffort in each functional unit. In sequential allocation, in theearly periods (prior to period 30), the organization allocates allresources to search in the first landscape, and only in laterperiods does search begin in the second landscape.
Figure 3 highlights a typology of search strategies that ariseat the intersection of modes of evaluation and the impositionof organizational structure. We implemented four types ofsearch strategies covering four of the quadrants (and sub-quadrants) of figure 3: (Q1) Baseline: an integrated structurewith on-line evaluation—which represents the simplestsearch strategy; (Q2) Integrated: an integrated structure withoff-line evaluation; (Q3a) Parallel: a functional structure withparallel allocation of effort and off-line evaluation; and finally(Q3b) Sequential: a functional structure with sequential allo-cation of effort and off-line evaluation.
599/ASQ, December 2007
Myopia of Selection
Q1BaselineStrategy
Figure 3. Evaluation mode versus organizational structure: A matrix of alternative strategies.
Evaluation Mode of Alternatives
Blind
Q4a Q3aParallel Strategy Parallel
Sequential Tem
po
ral
Allo
cati
on
of
Sea
rch
Eff
ort
No
Str
uct
ure
(In
teg
rate
d)
Org
aniz
atio
nal
Str
uct
ure
Str
uct
ure
(Fu
nct
ion
al)
Q4b Q3bSequential Strategy
Non-Blind(Off-line)
Q2IntegratedStrategy

Selection
Because we were interested in the efficiency of selection,we modeled adaptation and selection (organizational exit)without considering the entry of replacement organizations.1Selection regimes vary in their intensity—weak versusstrong. Selection was implemented as the outcome of a sto-chastic process such that an organization’s probability of sur-vival is a function of its fitness relative to the fitness of thebest firm at a given point in time (Wilson and Bossert, 1971).We implemented a proportional selection model, such thatthe probability of survival at a given point in time is si,t =(�i,t/Max(�t))
Zt, where �i,t is the fitness of organization i attime t, Max(�t) is the maximum fitness achieved by any firmat time t; and Zt is the strength of selection. The proportionalselection model imposes competitive consequences on orga-nizations because the probability of failure for a given organi-zation increases with fitness of the leading organization in thepopulation at any point in time.2
SIMULATIONS
We used the fitness landscape described above as an analyti-cal tool to analyze the efficacy of selection across alternativesearch strategies. All simulations are for an n = 20 landscapeconsisting of two sub-problems where each sub-problem cor-responds to a single landscape with n1 = n2 = 10 interdepen-dent policy choices. This in turn generates approximately onemillion (220) policy choice combinations represented byunique locations on the combined landscape. The interactionstructure within each sub-landscape is specified as kW = 5,with kB = 2 interactions across landscapes. A simulation runlasts for 100 periods and includes 200 organizations seededrandomly on the landscape. In each period, an organization isallocated two search attempts. All results are averaged over100 runs, and thus we evaluated the performance of a totalof 20,000 organizations in each simulation. Our analysis pro-ceeded in four experiments. In the first experiment, weexamined the dynamics of performance improvement and, inparticular, the reliability of adaptation across alternativesearch strategies. In the second experiment, we examinedthe efficacy of continuous selection (on the basis of currentperformance) within populations of organizations that werehomogeneous in terms of search strategies. In doing so, weassessed how different strategies affected the ability ofselection to act as a mechanism of population-level adapta-tion, to determine if there are differences across strategies inselectability—the information content of current performancethat allows selection to screen out organizations in a mannercorresponding to their long-run performance. In the thirdexperiment, we examined alternative selection regimes, firstby enabling selection on the basis of past as well as currentperformance and, second, by implementing discrete selec-tion at a single point in time. In the final experiment, weallowed organizations pursuing different search strategies tocompete and examined the implications of differences acrossstrategies in selectability for the demographics of survivingfirms.
1Modeling entry entails a number of choic-es, such as whether the entry process israndom or entrants tend to be replicationsof existing organizations, which wouldaffect our assessment of the extent towhich selection improves the population’sperformance across search strategies.
2The selection model implemented here isbased on proportional performance. Sucha model is sensitive to the extent ofcross-sectional variation. We also ran themodels in this paper using a rank-orderselection criterion, and the results did notchange markedly.
600/ASQ, December 2007

Experiment 1: Reliability of Alternative Search Strategies
We began by assessing the performance trajectories of alter-native search strategies in the absence of selection. We didso by running independent simulations for each of the fourdistinct search strategies in quadrants 1 through 3 in figure 3above. Results are presented in figures 4a–4c: (a) averageperformance across time; (b) cross-sectional variation (stan-dard deviation of fitness across firms) within the populationof organizations following a given strategy; and (c) intertem-poral short wave reliability measured as Corr(Ft–1,Ft). The cen-tral performance measure of fitness in figure 4a shows thatalthough the rate of performance increase differs markedlyacross the strategies, all four strategies appear to generate acommon asymptote with an average fitness of .69. In fact,the long-run results are not quite identical, although they areindistinguishable at the three-digit level. In particular, func-tional parallel strategies, by generating perturbations in onelandscape that have an impact on search in the other land-scape, periodically enable firms to escape a poor basin ofattraction for a basin with a superior local peak. But these dif-ferences in long-run performance across strategies are onlynoticeable at higher levels of cross-landscape interdepen-dence than was implemented in this paper and have no bear-ing on our results.
All strategies use the same amount of effort in the searchprocess. Thus differences in the rate of progress can onlycome from differences in search-strategy performance char-acteristics driven by the mode of evaluation and the imposi-tion of organizational structure. The central qualitative obser-vation is that the baseline and sequential strategies not onlyexhibit slower rates of progress but also entail far greatercross-sectional variability, as shown in figure 4b. For thebaseline strategy, these observed results can be attributed tothe on-line evaluation, which results in some fraction of alter-natives generating performance decrements. These errorslead to a significant reduction in short-wave reliability thatboth decreases the rate of performance improvement andincreases cross-sectional variation, particularly between peri-ods 15 and 25, as shown in figure 4c.
Less obvious is the sharp contrast in behavior among thethree search strategies that entail off-line evaluation of alter-natives (Quadrants 2 and 3 in figure 3). Integrated search,which differs from the baseline strategy because it imple-ments only performance-enhancing alternatives, proceedswith greater reliability and, as a consequence, more rapidlythan baseline strategy search. The two functional searchstrategies, parallel and sequential, match the underlying taskstructure by imposing organizational structure and decompos-ing the problem across two functional units. For example,while marketing and R&D functions may be interconnected interms of solutions, firms typically decompose the problemacross two functional units. Functional search strategiesignore the between-unit interdependence in activities andoutcomes because doing so reduces coordination costs.
Despite this commonality, the dynamics of performance ofparallel and sequential search strategies are markedly differ-
601/ASQ, December 2007
Myopia of Selection

602/ASQ, December 2007
Figure 4a. Average performance of search strategies across time.Fi
tnes
s
0.75
0.70
0.65
0.60
0.55
0.5000 06 12 18 24 30 36 42 48 54 60 66 72 78 84 90 96
Period
Figure 4b. Cross-sectional variation (S.D. across organizations).
Sta
nd
ard
Dev
iati
on
in F
itn
ess
0.07
0.065
0.06
0.055
0.05
0.045
0.04
0.035
0.0300 07 14 21 28 35 42 49 56 63 70 77 84 91 98
Period
Co
rrel
atio
n (
F t–1, F
t) in
Fit
nes
s
Figure 4c. Short-wave reliability: Correlation in performance across time.
00 05 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95
Period
1.0
0.9
0.8
0.7
0.6
0.5
0.4
0.3

ent. The parallel-strategy firms exhibit performance thatclosely matches that of the integrated-strategy firms. In con-trast, sequential-strategy firms, by focusing on only one land-scape in the early periods, limit the potential for performanceimprovement. In particular, the potential improvement is onaverage half that of the potential improvement that a parallel-strategy firm can expect. Sequential-strategy firms, however,exhibit even higher levels of short-wave reliability in earlyperiods by allocating all their search effort to one sub-prob-lem landscape. This higher early reliability leads to rapid earlyperformance gains that are quickly offset by the limited roomfor improvement in the landscape in which search efforts areinitially focused. In addition, this decoupling of search intro-duces a second issue: the adaptation trajectory of sequential-strategy firms is highly non-monotonic. After period 30, whensequential organizations commence search in the secondlandscape, fitness improves dramatically, cross-sectional vari-ation decreases, and short-wave reliability returns to pre-period 30 levels.
All search strategies examined in the first experiment led toadaptation that converged to a common long-run perfor-mance. Nevertheless, differences across search strategies inthe mode of evaluation of alternatives and the implementa-tion of organizational structure gave rise to significant differ-ences in the level of short-wave reliability. This property, inturn, drove both the rate of performance improvement andthe level of cross-sectional variation.
Experiment 2: Implications of Reliability for the Efficacy ofSelection
In the second experiment, we examined the consequencesof differences in reliability for the efficacy of selection as amechanism of population-level adaptation, to assess theextent to which selection increases average population-levelperformance across the different search strategies by elimi-nating inferior firms. Our proposition is that selection will bemore effective at identifying superior firms when those firmspursue search strategies that exhibit higher levels of reliabili-ty in the adaptation process.
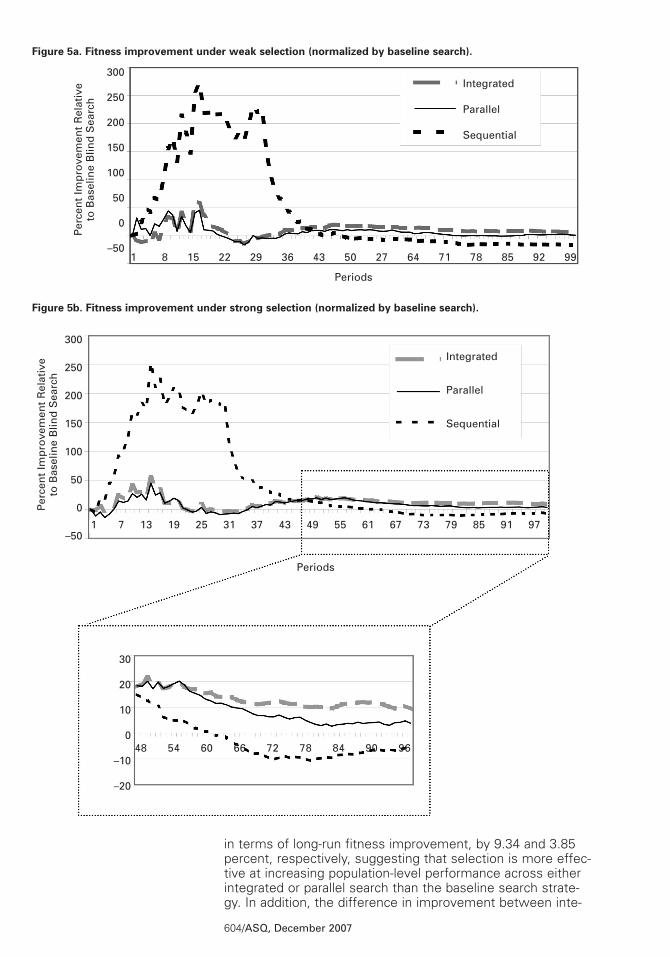
We ran this experiment under two different selectionregimes: (1) weak selection, which removes approximately50 percent of the population over the 100-period experiment,and (2) strong selection, which eliminates approximately 95percent of firms over 100 periods. The performance variableof interest was the average improvement in population-levelfitness generated by selection, as it removes organizationsdeemed to be inferior, normalized by the baseline adaptationresults. The results are presented in figures 5a (weak selec-tion) and 5b (strong selection). The figures show that strongselection over a population of integrated-search firms increas-es fitness 9.34 percent more than does such selection acrossa population of firms pursuing the baseline adaptation model.Because the results are consistent across both regimes(although less intense in the weak selection regime), wefocus our discussion on the strong selection regime.
Under the strong selection regime, both integrated and paral-lel search strategies outperform the baseline search strategy
603/ASQ, December 2007
Myopia of Selection
in terms of long-run fitness improvement, by 9.34 and 3.85percent, respectively, suggesting that selection is more effec-tive at increasing population-level performance across eitherintegrated or parallel search than the baseline search strate-gy. In addition, the difference in improvement between inte-
604/ASQ, December 2007
Figure 5a. Fitness improvement under weak selection (normalized by baseline search).
Integrated
Parallel
Sequential
1 08 15 22 29 36 43 50 27 64 71 78 85 92 99
Periods
300
250
200
150
100
50
0
–50
Per
cen
t Im
pro
vem
ent
Rel
ativ
eto
Bas
elin
e B
lind
Sea
rch
Integrated
Parallel
Sequential
Figure 5b. Fitness improvement under strong selection (normalized by baseline search).
300
250
200
150
100
50
0
–50
30
20
10
0
–10
–20
48 54 60 66 72 78 84 90 96
1 07 13 19 25 31 37 43 49 55 61 67 73 79 85 91 97
Periods
Per
cen
t Im
pro
vem
ent
Rel
ativ
eto
Bas
elin
e B
lind
Sea
rch

grated and parallel search is large, given that the results fromexperiment 1 showed only relatively small differences inshort-wave reliability. This highlights an as yet unexploredcost of decomposing problems into constituent parts. Evensmall differences in reliability that arise through the imposi-tion of organizational structure can alter the ability of selec-tion to facilitate effective population-level adaptation.
The results for the sequential search strategy are more com-plex. Selection-driven performance improvements are anorder of magnitude larger for sequential search in the earlyperiods, reaching a peak of 250 percent of the rate exhibitedby the baseline search strategy, but this dissipates rapidlywith the activation of search efforts related to the secondsub-problem (landscape 2). In the long run, selection acrossfirms pursuing a sequential search strategy is less effectivethan equivalent selection across the baseline blind-searchmodel by 6.7 percent.
To assess the significance of these differences in long-runperformance contingent on surviving selection, we conduct-ed unpaired t-tests of the mean performance across eachstrategy at period 100. The results of this test for the strongselection regime are presented in table 1. Each data point inthe table contains the difference in fitness at period 100 asthe column less the row. Thus after selection, the average fit-ness of a population of firms pursuing the integrated searchstrategy is .0047 greater than that of a population of firmspursuing the baseline strategy, a difference that is highly sig-nificant at the .001 level. There are two central results fromthis table. First, selection is significantly less effective acrossfirms pursuing the baseline blind search strategy than eitherthe integrated or the parallel strategies. Second, selection issignificantly less effective across the sequential strategy thanall the other search strategies examined.
To examine why selection is more or less effective acrosssome search strategies, we conducted two further sets ofanalyses. First, we calculated the selection-error rateobserved at period 100 to determine how many of the firms(out of 200) that survived selection were actually in the top100 firms in period 100. The results of this analysis, whichare presented in figure 6, are read as follows: in the weakselection regime with firms pursuing the baseline search
605/ASQ, December 2007
Myopia of Selection
Table 1
Difference in Mean Fitness at Period 100 under a Strong Selection
Regime*
Strategy Baseline Integrated Parallel
Integrated .0047•••• .— .—(.0013) .— .—
Parallel .0027• –.0020 .—(.0015) (.0015) .—
Sequential –.0035••• –.0081•••• –.0061••••(.0015) (.0015) (.0017)
• p < .10; •• p < .05; ••• p < .01; •••• p ≤ .001.* Standard errors are in parentheses. The difference in fitness is the averagerow fitness less the average column fitness, with a positive difference indi-cating greater row fitness.

strategy, of the 102 firms that survive selection, 41 of thesurviving firms were not in the top 102 of the 200 firms, thusgenerating a 39-percent error rate. In general, the results ofthe error-rate analysis are consistent with the performance-improvement examination. A shift from the baseline searchstrategy to integrated search leads to a decrease in the errorrate from 71 percent to 65 percent, with the error rateincreasing to 66 percent for parallel search, consistent withthe long-term performance differences exhibited in figure 5b.The error rate across sequential search was the highestobserved, at 77 percent.
Thus the efficacy of selection as a mechanism of population-level adaptation, highlighted by differences in population fit-ness under selection, is driven by the extent to which theselection process is error prone, removing organizations inearlier periods that would have gone on to do well if they hadsurvived. In particular, for all search strategies exceptsequential search, there appears to be a strong relationshipbetween the extent of short-wave reliability and selection-error rates. As short-wave reliability increases, error ratesappear to decrease, and selection becomes more effective.
Second, we examined the nature of information available forselection at a given point in time, focusing on how hetero-geneity across organizations in the reliability of adaptationaffects the quality of the information on which selection acts.The challenge over time for selection is in estimating futureperformance. To what extent do the intermediate perfor-mance manifestations of adapting organizations provide agood signal about future performance? If differences acrosssearch strategies affect the degree of reliability of adaptation,then this may distort the information provided by contempo-raneous performance, rendering it less informative aboutlong-run performance and in turn limiting the effectiveness ofintermediate selection.
To examine this issue, we measured long-wave reliability bycalculating the extent to which fitness at a given point in timeis correlated with long-run fitness, Corr(Ft,F100), which pro-vides a direct measure of the informativeness of intermedi-ate performance levels. For example, if the correlationbetween fitness at period 30 and 100 is 100 percent, then fit-ness at period 30 is perfectly informative about long-run fit-ness. If, in contrast, the correlation is only 5 percent, thenintermediate fitness provides little useful information aboutlong-run fitness. The results of this analysis are shown in fig-ure 7, which plots the correlation in fitness between period tand period 100 across each of the search strategies. Forexample, in period 15, the correlation between contempora-neous and long-term fitness for the integrated strategy isapproximately 52 percent—that is, fitness at period 15explains 52 percent of the variance in long-term performance.In contrast, for the baseline blind-search strategy model, thecorrelation is only 36 percent. For the sequential strategy, thecorrelation drops further to 27 percent.
For the baseline, parallel, and sequential strategies, the short-wave and long-wave reliability results are broadly similar interms of rank ordering. In contrast, for the sequential strate-
606/ASQ, December 2007

gy, while the short-wave reliability is superior to that of thebaseline strategy, the long-wave reliability is greatly inferior.This reversal stems from the temporal decomposition of thesearch effort in the sequential strategy. Because the twosub-problems (landscapes) are interdependent, the onset ofsearch in the second landscape in period 30 makes obsoletemuch of the firm-level learning in the first landscape. As aconsequence, the selective removal of inferior sequential-strategy firms prior to period 30 is much more likely to be inerror. The early performance improvement generated byselection across sequential-strategy firms, shown in figure 5babove, leads to rapid improvement in the population’s perfor-mance and is consistent with the very high level of short-wave reliability that this strategy demonstrates at the outsetof the simulation. But this advantage is quickly overwhelmedby the cost associated with a significant decrease in long-wave reliability. In the long run, this reliability deficit leads toselection errors that decrease the ability of selection to act asa mechanism of population-level adaptation.
607/ASQ, December 2007
Myopia of Selection
Figure 6. Selection error rates across search strategies at period 100.
Baseline Integrated Parallel Sequential
Weak Selection Strong Selection
Selection Regime
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%
Err
or
Rat
e (%
)
Figure 7. Long-wave reliability: The information content in contemporaneous performance.
1.0
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0.00 06 12 18 24 30 36 42 48 54 60 66 72 78 84 90 96
Period
Co
rrel
atio
n (
F t,F10
0) in
Fit
nes
s
Baseline
Integrated
Parallel
Sequential

Finally, we examined the extent to which our results weresensitive to differences across strategies in cross-sectionalvariation. We implemented a rank-order selection regime thatwas insensitive to cross-sectional variation. Our results (avail-able from the authors) were robust to this alternative specifi-cation. Differences in cross-sectional variation do affect theefficacy of selection, but when there are also substantial dif-ferences in reliability across strategies, differences in cross-sectional variation will tend to be swamped by the contribu-tion of these differences in reliability over time.
Experiment 3: Efficacy of Alternative Selection Regimes
The previous experiment demonstrated that the efficacy ofselection varied significantly across search strategies as afunction of the reliability of adaptive efforts. In that experi-ment, selection was continuous, occurring in each period,and was implemented on the basis of contemporaneous per-formance alone. In the third experiment, we explored theefficacy of two alternative selection regimes that, althougharbitrary in their construction, reflect important dimensions ofselection regimes observed in practice: (1) continuous selec-tion with the use of historical information and (2) discreteselection at period 15.
We implemented selection on the basis of historical, ratherthan simply current performance. We used twenty periods ofhistorical data and ran the simulation models from experi-ment 2 to ascertain if more information enhances the effica-cy of selection. As an indication of the efficacy of selection,we considered the error-rate results, which are reported infigure 8. The height of the bars represents the percentage oforganizations that survive selection erroneously. The num-bers above the bars represent the comparison with theresults from experiment 2 graphed in figures 5a and 5b.Thus, for the baseline blind-search strategy, the use ofhistorical data under a strong selection regime led to an error
608/ASQ, December 2007
Figure 8. Selection error rate with performance history.*
Baseline Integrated Parallel Sequential
Weak Selection Strong Selection
Selection Regime
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%
* Numbers above the columns represent an increase or decrease in the error rate from the continuousselection mode results graphed in figure 6.
Err
or
Rat
e (%
)
–2.35% 7.99% 4.05%
6.35%
–1.22% 6.68% 8.53%
4.44%

rate of 71 percent, which is a reduction of 1.22 percent rela-tive to the error rate observed in experiment 2 (e.g., currentperformance only). In contrast, for the integrated, parallel,and sequential strategies, the error rate increased by 6.68,8.53, and 4.44 percent, respectively.
The results are informative in two respects. First, for themore reliable strategies (integrated, parallel), historical dataare less useful predictors of future success than is currentperformance, because performance at any point in time forthese monotonic strategies provides information on thelower bound of the local peak, and therefore historical perfor-mance data from points lower down the peak is less informa-tive about future performance than is current performance.Thus the counterintuitive result is that including historicaldata diminishes the efficacy of selection by increasing errorrates. Second, for the less reliable strategies, which exhibitnon-monotonic performance gains, the results diverge. Thebaseline strategy suffers from a deficit of short-wave reliabili-ty. Because adaptation is blind, in any given period, the orga-nization may implement a trial that reduces performance.Here, the use of historical data, which in effect averagesbackward, captures prior-period results that were at timessuperior (higher up the local peak) to the current period, thusproviding better information on which selection could act andresulting in a lower error rate. In contrast, sequential searchsuffers from a deficit in long-wave reliability. In this case, theuse of historical data implies that after period 30 (the onsetof landscape 2 search), pre-period 30 data are still consid-ered, and as a consequence, the error rate increases.
Discrete selection represents a regime with intermittentrather than continuous selection and is in this sense similarto that of a venture capital regime. We examined the out-come of a discrete selection event at period 15. Error-rateresults on the efficacy of selection are reported in figure 9.
609/ASQ, December 2007
Myopia of Selection
Figure 9. Discrete selection error rate with performance history.*
Weak Selection Strong Selection
Selection Regime
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%
Baseline Integrated Parallel Sequential
33.1%
16.7% 17.0%
30.0%
15.1%
12.2% 6.7%
15.1%
Err
or
Rat
e (%
)
* Numbers above the columns represent an increase or decrease in the error rate from the continuousselection mode results graphed in figure 6.

The main result is that discrete selection exhibits a higherselection-error rate than does continuous selection. Underthe weak selection regime, the error rate increase isbetween 16.7 and 33.1 percent. Under the strong selectionregime, the increase in error rates ranges from 6.7 to 15.1percent. Two observations are of note. First, error rates growfaster under the weak selection regime. Second, the imple-mentation of discrete selection, rather than continuous selec-tion, comes at a cost that differs across search strategies. Inparticular, although the basic pattern of the efficacy of selec-tion across alternative search strategies remains constant,the error rate increases faster across the less reliable base-line and sequential strategies than it does across the morereliable integrated and parallel strategies. The largest increasein error rate occurs in selection over the sequential strategy.The efficacy of the results from discrete selection, however,will be partly a function of the time at which selection isimplemented. Because our interest was in modeling a selec-tion regime similar to that of venture capital, we examinedearly selection, but as the time of discrete selection moveslater, the efficacy of selection will improve. At the extreme,with selection at period 100, selection will simply be propor-tionate to long-run performance.
The results of experiment 3 suggest that the general rankingof alternative strategies in terms of selectability remains rela-tively unchanged under alternative selection regimes. As onewould expect, a decrease in information on which selectionacts (discrete selection) reduces the efficacy of selection,though this reduction in efficacy is less severe in a strongselection regime. Counterintuitively, the results suggest thatin most cases, the use of more information (historical selec-tion) also serves to reduce the efficacy of selection.
Experiment 4: Competitive Implications of Differences inSelectability
The previous three experiments demonstrated that the effica-cy of selection varies significantly across search strategies asa function of the reliability of adaptation. In experiment 4, weexamined the consequences of differences across strategiesin selectability when two organizational forms (i.e., sets oforganizations pursuing different search strategies) are com-peting for dominance in a population. Each simulationinvolved 100 runs of the model with 200 firms split betweenthe two competing search strategy types.
Two central measures that are of interest are the relative pro-portion of survivors of each type of organization and the prob-ability that a given type of organization will be the winningorganization. We ran six separate simulations (three eachunder the weak and strong selection regimes) in which popu-lations of two types of firms compete: (a) baseline vs. inte-grated, (b) baseline vs. parallel, and (c) baseline vs. sequen-tial. In the absence of selection, all four strategies (Quadrants1 through 3 in figure 3) generate identical long-run perfor-mance. Contingent on surviving selection, however, this equi-finality disappears. Figure 10 illustrates the population demo-graphics results, the proportion of survivors in period 100 thatare non-baseline-strategy firms. Figure 11 illustrates the win-
610/ASQ, December 2007

ner of the competition, graphing the probability that a non-baseline-strategy firm has (or is tied for) the highest perfor-mance at period 100. On the basis of both performance met-rics, the integrated and parallel strategies generate superioroutcomes in competition with the baseline strategy. Forexample, under strong selection, the integrated-strategyfirms come to account for 63 percent of the population ofsurviving firms and an integrated-strategy firm exhibits thehighest performance amongst the survivors nearly 80 percentof the time. In contrast, the sequential-strategy firms lose tobaseline-strategy firms on both performance metrics. Theresults suggest that differences in the efficacy of selection(driven by differential reliability) across strategies have directimplications for the outcome of competition between alterna-tive organizational forms. Because selection is more errorprone within the populations of less reliable search strate-gies, many of the latently superior members of such sub-pop-
611/ASQ, December 2007
Myopia of Selection
Figure 10. Demographics of competitive outcomes (versus baseline search strategy).
Figure 11. Winner of competitive outcomes (versus baseline search strategy).
Integrated Parallel Sequential
Weak Selection Strong Selection
Selection Regime
70%
60%
50%
40%
30%
20%
10%
0%
Sh
are
of
Su
rviv
ing
Po
pu
lati
on
Integrated Parallel Sequential
Weak Selection Strong Selection
Selection Regime
1
0.8
0.6
0.4
0.2
0
–0.2
–0.4
–0.6
Win
nin
g S
trat
egy
Pro
bab
ility

ulations are mistakenly eliminated, significantly reducing thelikelihood that one such organization will be the winner. As aconsequence, organizations pursuing search strategies thatare more reliable, exhibiting superior selectability, come todominate the population of surviving organizations. In thissense, reliability and survival are correlated even when selec-tion occurs on the basis of performance. This final experi-ment highlights differential selectability, driven by differencesin the reliability of adaptive efforts, as an important factor indetermining the outcome of competition in evolutionary sys-tems in which selection occurs across organizations that arethemselves undergoing adaptation.
DISCUSSION AND CONCLUSION
Evolutionary perspectives on firms and industries havebecome quite prominent in the strategy and organizations lit-eratures (Nelson and Winter, 1982; Hannan and Freeman,1984; Baum and Singh, 1994; Burgelman, 1996). Central tothe evolutionary argument is the presence of some processof competitive exclusion and differential selection. Further-more, an important strand of the strategy and organizationsliterature has taken a firm-centered view of evolutionarydynamics in analyzing the path-dependent processes of firm-level adaptation and learning (cf. Helfat, 2003). Althoughthere has been some recognition of the need to consider theinterrelationship between the processes of adaptation andselection (Levinthal, 1991a), and some important steps havebeen taken in that direction (March and Shapira, 1992; Bar-nett, Greve, and Park, 1994; Barnett and Hansen, 1996; Den-rell and March, 2001; Greve, 2002), the micro dynamics ofthe two processes have only begun to be explored. Ourobjective in this paper was to use a simulation to explore theefficacy of selection as a mechanism of population-leveladaptation when individual organizations themselves areadapting. Our central finding from the simulations is that theefficacy with which selection discriminates among adaptiveorganizations, and as a consequence, the ability of selectionto drive population-level adaptation, is a function of theprocess of organizational change itself. Search strategies thatlead to learning outcomes that exhibit high reliability are moreselectable in that selection is less prone to error when actingacross firms that are pursuing highly reliable strategies.
We adopted the assumption of inertia with respect to organi-zations’ “genetic structure,” but this premise does notnegate the possibility, or even the likelihood, of profoundchanges in the observed properties of an organization, what abiologist would refer to as its phenotype, and, as a conse-quence, the performance outcomes it experiences. In ouranalysis, the stable fundamental organizational property wasthe firm’s search strategy, both how alterative initiatives areevaluated (on-line vs. off-line) and the structural arrange-ments of the organization (integrated vs. functional). Thesearch strategy in turn determines the broad characteristicsof the dynamics of organizational adaptation.
In examining selection, efficacy can be considered from twovery different perspectives. First, from a simple cross-sec-tional perspective, to what degree are poorly performing
612/ASQ, December 2007

firms selected out of the population at any given point intime? Second, from a longitudinal perspective, are those enti-ties that are on a superior latent adaptive trajectory more like-ly to survive? The longitudinal problem exists because organi-zations are adapting, and selection is inherently myopic. Eventhe self-consciously forward-looking selection efforts ofinvestment analysts and venture capitalists are necessarilyconstructed on the basis of current and historical data. Weheld constant cross-sectional efficacy and focused our atten-tion on the longitudinal problem, which only exists whenthere is adaptation: Under what conditions can myopic selec-tion demonstrate the possibility of prospective intelligence,identifying adapting organizations at intermediate points intime, in the midst of change, that will be superior in the longrun?
Results of our model point to the fact that the efficacy ofselection is endogenous to the dynamics of organizationaladaptation. Important stable attributes of organizations suchas strategies or routines may differ not only in their contribu-tion to organizational fitness or performance at a given pointin time but may also have differential impacts on the efficacyof selection over time. We denoted this effect on the efficacyof selection as selectability, an organization-level constructthat measures the information content embodied in currentperformance as a determinant of the efficacy of selection inidentifying organizations that will be superior in the long run.For organizations that exhibit high levels of selectability,intermediate performance provides superior informationabout long-run performance, and consequently, selection ismore effective longitudinally within populations of suchorganizations.
An important driver of selectability is the extent to whichadaptation proceeds more or less reliably over time. Our find-ing on reliability is closely related to the ideas developed inthe population ecology literature, in particular, the early argu-ment that selection occurs on the basis of reliability (Hannanand Freeman, 1984). Although later work has relaxed thisargument (cf. Hannan, 2005: 53), our results suggest thatselection on performance alone is sufficient to generate acorrelation between reliability and survival: the role of reliabili-ty emerges endogenously from a model in which inert strate-gies guide organizational learning in a competitive ecology.The correlation between reliability and survival occursbecause for patterns of adaptation that are highly reliableover time, an organization’s performance at any point in timeprovides better information for selection to act upon in filter-ing out inferior organizations (i.e., it is more selectable). Forexample, the integrated search strategy exhibits higher selec-tability than does the baseline blind strategy. As a conse-quence, selection across integrated-strategy firms is moreeffective at removing poor-quality organizations. Conditionalon survival, firms using integrated search on average outper-form those engaged in blind search, even though uncondi-tional on survival, they exhibit identical long-run performance.That said, though our model highlights the process by whichselection favors reliability, it was only tested in a stable envi-ronment. Left unexplored in our paper are the implications of
613/ASQ, December 2007
Myopia of Selection

our model for the consequences of reliability in performanceunder conditions of environmental turbulence (Dobrev, Kim,and Carroll, 2003).
Taking the myopia of selection into account has importantimplications for our understanding of the interrelationshipbetween the processes of adaptation and selection, in partic-ular, for understanding the relative merits of exploration ver-sus exploitation strategies. March’s (1991) well-known resultis that exploratory strategies (with higher variance) tend tofavor long-run performance, while exploitive strategies favornear-term performance. But strategies that are effective inthe long run must survive a series of short-run selectiondemands (Levinthal and March, 1993). The myopia of selec-tion suggests that prospects for the long-run performanceadvantage of exploration must be offset by the higher perfor-mance variance over time that tends to obscure estimates ofan organization’s underlying quality. This difficulty of estimat-ing performance in turn increases the risk that latently superi-or firms will fail and inferior firms will survive. It would evensuggest that firms with poor endowments would choose topursue an exploration strategy, not simply because it mayenhance their long-run performance prospects but alsobecause higher variance masks the performance deficitbetween them and their rivals.
These results provide an alternative explanation for theobservation that inferior organizations often come to domi-nate the set of surviving firms. Scholars have argued that thismight occur because performance is not the sole determi-nant of the outcome of selection (Anderson and Tushman,1990). Our study suggests an additional explanation: thatselection may be systematically prone to error. In contexts inwhich selection is acting across organizations engaged inadaptation, the reliability with which adaptation occurs is cen-trally important. Strategies that are inferior may dominate ifthey exhibit more reliability in their adaptive efforts.
In examining errors in the selection process, we uncoveredan important relationship between the intensity of selectionand its error rate. Researchers have long argued that highermunificence (a less intense selection regime) gives organiza-tions access to external resources (Finkelstein and Hambrick,1990), which in turn increase their decision latitude andenhance their performance (Baum and Wally, 2003). We iden-tified an alternative mechanism by which munificence mayenhance performance, in this case, population performance.In particular, selection that is less intense (more munificent)is subject to fewer errors in selectively removing inferiorfirms. If the distinctive properties of organizations, such astacit and organizationally embedded knowledge, are lost withthe dissolution of a firm, then less intense selection, with itslower error rate may enhance social welfare.
An additional counterintuitive result is that using more infor-mation in the selection process—using historical data in addi-tion to current data—does not necessarily improve the effica-cy of selection. In fact, using more data improved theefficacy of selection only when search efforts suffered froma lack of short-wave reliability because of a blind search strat-
614/ASQ, December 2007

egy. The efficacy of selection across strategies that exhibitedrelatively high short-wave reliability (e.g., integrated and paral-lel) was strongly diminished by the use of historical data. Forsearch strategies that result in infrequent large shocks, and aconsequent lack of long-wave reliability, that devalue olderlearning (e.g., sequential strategy), the use of historical dataalso diminishes the efficacy of selection.
Many important empirical contexts have selection that is dis-crete in time, rather than continuous. Perhaps the dominantexample is that of venture capital. Our results suggest thatdiscrete selection at early points in time is significantly lesseffective than is continuous selection. This result does notrule out the viability of discrete selection regimes. When car-rying out selection is costly, a trade-off between frequencyand efficacy is to be expected. For example, in the venturecapital context, the frequency of selection is in part a func-tion of the cost of engaging in a round of selection. At thesame time, our results demonstrate that in the transitionfrom continuous to discrete selection, error rates grow fasterunder a weak selection regime than a strong selectionregime. Our model thus suggests that one would expect toobserve significantly more intense selection in a discreteselection regime than in a continuous selection regime andthus a much higher failure rate. This result is independent ofthe arguments for high failure rates that are commonly putforward about technological or market uncertainty and oppor-tunity costs for venture capitalists.
Finally, our examination of the efficacy of selection alsoenhances our understanding of the eternal question, what isa good strategy? Our results suggest an interesting answer.All else held constant, in competitive contexts in which sur-vival is conditional on the performance of other firms, higher-variance strategies (March, 1991) decouple mean perfor-mance from survival and in doing so, alter the probability ofsurvival for individual firms. At the population level, however,the survival of firms with inferior mean performance reflectsa long-run selection error. This result is particularly importantwhen increasing reliability is costly and thus reliability mustbe traded off against long-run performance. For example, amodicum of exploration may enhance long-run performance,relative to the local search we modeled, but it does so at thecost of decreased reliability. In such cases, counterintuitively,an individual firm’s attempt to enhance its performance maydiminish the efficacy of selection as a mechanism of popula-tion-level adaptation and lead to a set of surviving firms thatexhibit inferior performance. The high variance strategy maybe superior at the individual firm level but inferior at the pop-ulation level.
A logical implication of the claim that selection makes sys-tematic errors in identifying organizations that would go on todo well is that, from the perspective of individual organiza-tions, pursuing a strategy that increases selectability neednot be advantageous. For example, organizations with poorinitial strategic positions that engender limited long-runprospects may be best served by pursuing an adaptationstrategy that purposefully obfuscates their performance out-look, reducing their selectability. Such a strategy may
615/ASQ, December 2007
Myopia of Selection

Amburgey, T. L., D. Kelly, andW. P. Barnett1993 “Resetting the clock: The
dynamics of organizationalchange and failure.” Adminis-trative Science Quarterly, 38:51–73.
Anderson, P., and M. L. Tushman1990 “Technological discontinuities
and dominant designs: Acyclical model of technologi-cal change.” AdministrativeScience Quarterly, 35:604–633.
Arthur, W. B.1989 “Competing technologies,
increasing returns, and lock-inby historical events.” Eco-nomic Journal, 99: 116–131.
Baldwin, C. Y., and K. B. Clark2000 Design Rules: The Power of
Modularity. Cambridge, MA:MIT Press.
Barnett, W. P.1997 “The dynamics of competitive
intensity.” Administrative Sci-ence Quarterly, 42: 128–160.
Barnett, W. P., and R. A.Burgelman1996 “Evolutionary perspectives on
strategy.” Strategic Manage-ment Journal, 17: 5–19.
Barnett, W. P., and G. R. Carroll1995 “Modeling internal organiza-
tional change.” AnnualReview of Sociology, 21:217–236.
Barnett, W. P., H. R. Greve, andD. Y. Park1994 “An evolutionary model of
organizational performance.”Strategic Management Jour-nal, 15: 11–28.
Barnett, W. P., and M. T. Hansen1996 “The Red Queen in organiza-
tional evolution.” StrategicManagement Journal, 17:139–157.
Baron, J. N., M. D. Burton, andM. T. Hannan1999 “Engineering bureaucracy:
The genesis of formal poli-cies, positions, and structuresin high-technology firms.”Journal of Law, Economicsand Organization, 15: 1–41.
Barron, D. N., E. West, and M. T.Hannan1994 “A time to grow and a time
to die: Growth and mortalityof credit unions in New YorkCity, 1914–1990.” AmericanJournal of Sociology, 100:381–421.
Baum, J. A. C., and T. Amburgey2001 “Organizational ecology.” In
J. A. C. Baum (ed.), BlackwellCompanion to Organizations:304–326. Malden, MA: Black-well.
Baum, J. A. C., and J. V. Singh1994 Evolutionary Dynamics of
Organizations. New York:Oxford University Press.
Baum, J. R., and S. Wally2003 “Strategic decision speed and
firm performance.” StrategicManagement Journal, 24:1107–1129.
Burgelman, R. A.1996 “A process model of strategic
business exit: Implications foran evolutionary perspectiveon strategy.” Strategic Man-agement Journal, 17:193–214.
Campbell, D. T.1960 “Blind variation and selective
retention in creative thoughtas in other knowledgeprocesses.” PsychologicalReview, 67: 380–400.
1965 “Variation and selective reten-tion in socio-cultural evolu-tion.” In H. R. Barringer, G. I.Blankstein, and R. W. Mack(eds.), Social Change inDeveloping Areas: A Reinter-pretation of Evolutionary The-ory: 19–48. Cambridge, MA:Schenkman.
1982 “Evolutionary epistemology.”In H. C. Plotkin (ed.), Learn-ing, Development, and Cul-ture: Essays in EvolutionaryEpistemology: 73–107. Chich-ester, UK; New York: Wiley.
Carroll, G. R., and J. R. Harrison1994 “On the historical efficiency
of competition between orga-nizational populations.” Amer-ican Journal of Sociology,100: 720–749.
Cohen, M. D., R. Burkhart, G.Dosi, M. Egidi, L. Marengo, M.Warglien, and S. Winter1996 “Routines and other recurring
action patterns of organiza-tions: Contemporary researchissues.” Industrial and Corpo-rate Change, 5: 653–698.
Cyert, R. M., and J. G. March1963 A Behavioral Theory of the
Firm. Englewood Cliffs, NJ:Prentice-Hall.
David, P. A.1985 “Clio and the economics of
QWERTY.” American Eco-nomic Review, 75: 332–337.
616/ASQ, December 2007
enhance their own survival prospects, even though it reducesthe efficacy with which selection is able to enhance popula-tion-level performance. In this sense, systematic errors in theselection process, which are socially deleterious, may wellrepresent strategic opportunities for organizations.
Organizations engage in learning processes to enhance theirperformance prospects. Implicit in this organizational objec-tive is that such action enhances their survival prospects. Yetlearning is a long and drawn out process—end states oflearning may take months or years to achieve. Under theinfluence of the myopia of selection, the dynamics of organi-zational adaptation are a critical determinant of both thelikelihood of survival for any particular organization and, morebroadly, the efficacy of selection as a mechanism ofpopulation-level adaptation.
REFERENCES

Delacroix, J., and A.Swaminathan1991 “Cosmetic, speculative, and
adaptive organizationalchange in the wine industry:A longitudinal study.” Admin-istrative Science Quarterly,36: 631–661.
Denrell, J., and J. G. March2001 “Adaptation as information
restriction: The hot stoveeffect.” Organization Science,12: 523–538.
Dobrev, S. D., T.-Y. Kim, and G. R.Carroll2003 “Shifting gears, shifting nich-
es: Organizational inertia andchange in the evolution of theU.S. automobile industry,1885–1981.” OrganizationScience, 14: 264–282.
Ethiraj, S. K., and D. Levinthal2004 “Bounded rationality and the
search for organizationalarchitecture: An evolutionaryperspective on the design oforganizations and their evolv-ability.” Administrative Sci-ence Quarterly, 49: 404–437.
Finkelstein, S., and D. C.Hambrick1990 “Top-management-team
tenure and organizational out-comes: The moderating roleof managerial discretion.”Administrative Science Quar-terly, 35: 484–503.
Gavetti, G., and D. A. Levinthal2000 “Looking forward and looking
backward: Cognitive andexperiential search.” Adminis-trative Science Quarterly, 45:113–137.
Greve, H. R.2002 “Sticky aspirations: Organiza-
tional time perspective andcompetitiveness.” Organiza-tion Science, 13: 1–17.
Hannan, M. T.2005 “Ecologies of organizations:
Diversity and identity.” Jour-nal of Economic Perspectives,19: 51–70.
Hannan, M. T., and J. Freeman1977 “The population ecology of
organizations.” AmericanJournal of Sociology, 82:929–964.
1984 “Structural inertia and organi-zational change.” AmericanSociological Review, 49:149–164.
Haveman, H. A.1992 “Between a rock and a hard
place: Organizational changeand performance under condi-tions of fundamental environ-mental transformation.”Administrative Science Quar-terly, 37: 48–75.
Helfat, C. E. (ed.)2003 The SMS Blackwell Handbook
of Organizational Capabilities:Emergence, Development,and Change. Malden, MA:Blackwell.
Kauffman, S. A.1993 The Origins of Order: Self-
Organization and Selection inEvolution. New York: OxfordUniversity Press.
Kelly, D., and T. L. Amburgey1991 “Organizational inertia and
momentum: A dynamicmodel of strategic change.”Academy of ManagementJournal, 34: 591–612.
Lane, D. A.1993 “Artificial worlds and eco-
nomics, part I.” Journal ofEvolutionary Economics, 3:89–107.
Levinthal, D. A.1991a“Organizational adaptation
and environmental selection:Interrelated processes ofchange.” Organization Sci-ence, 2: 140–145.
1991b“Random walks and organiza-tional mortality.” Administra-tive Science Quarterly, 36:397–420.
1997 “Adaptation on rugged land-scapes.” Management Sci-ence, 43: 934–950.
Levinthal, D. A., and J. G. March1993 “The myopia of learning.”
Strategic Management Jour-nal, 14: 95–112.
March, J. G.1991 “Exploration and exploitation
in organizational learning.”Organization Science, 2:71–87.
March, J. G., and J. P. Olsen1989 Rediscovering Institutions:
The Organizational Basis ofPolitics. New York: FreePress.
March, J. G., and Z. Shapira1992 “Variable risk preferences and
the focus of attention.” Psy-chological Review, 99:172–183.
Miller, D., and M. J. Chen1994 “Sources and consequences
of competitive inertia: A studyof the U.S. airline industry.”Administrative Science Quar-terly, 39: 1–23.
Miner, A. S., T. L. Amburgey, andT. M. Stearns1990 “Interorganizational linkages
and population dynamics:Buffering and transformation-al shields.” AdministrativeScience Quarterly, 35:689–713.
Nelson, R. R., and S. G. Winter1982 An Evolutionary Theory of
Economic Change. Cam-bridge, MA: Belknap Press ofHarvard University Press.
Rivkin, J. W.2000 “Imitation of complex strate-
gies.” Management Science,46: 824–844.
2001 “Reproducing knowledge:Replication without imitationat moderate complexity.”Organization Science, 12:274–293.
Rivkin, J. W., and N. Siggelkow2003 “Balancing search and stabili-
ty: Interdependencies amongelements of organizationaldesign.” Management Sci-ence, 49: 290–311.
Romanelli, E., and M. L. Tushman1994 “Organizational transforma-
tion as punctuated equilibri-um: An empirical test.” Acad-emy of Management Journal,37: 1141–1166.
Siggelkow, N., and D. A. Levinthal2003 “Temporarily divide to con-
quer: Centralized, decentral-ized, and reintegrated organi-zational approaches toexploration and adaptation.”Organization Science, 14:650–669.
Simon, H. A.1955 “A behavioral model of ratio-
nal choice.” Quarterly Journalof Economics, 69: 99–118.
1962 “The architecture of complex-ity.” Proceedings of theAmerican Philosophical Soci-ety, 106: 467–482.
1969 The Sciences of the Artificial.Cambridge, MA: MIT Press.
1991 Models of My Life. New York:Basic Books.
Singh, J. V., R. J. House, and D. J.Tucker1986 “Organizational change and
organizational mortality.”Administrative Science Quar-terly, 31: 587–611.
617/ASQ, December 2007
Myopia of Selection

APPENDIX: Construction of the NK Simulation
Task Environment
Firms operate in a task environment consisting of two nearly decomposablesub-problem spaces (landscapes). A location in landscape 1 is representedby the vector N1 (length n1) of policy choice attributes, and similarly, a loca-tion in landscape 2 is represented by the vector N2 (length n2). Taken togeth-er, a location in the combined landscape can be described by the row vectorN (length n = n1 + n2) of firm policy choice attributes such that:
N = {N1, N2} = (a11, a
12, .|.|. , a
1n1, a2
1,a22, .|.|. , a
2n2). (1)
For notational convenience, we have omitted the subscript denoting thefirm, i, from the discussion that follows. We assume that each policy choiceis binary (1 or 0) and therefore there are 2n possible combinations of organi-zational choices.
The topography of the landscape is a function of the number of interactionsbetween policy choices, which influence the degree to which the payoff of afocal policy choice depends on the value of other policy choices. Interactionscan occur both within a landscape and between landscapes (the upper-rightand lower-left quadrants in figure 2).
Consider a single policy choice j on landscape L, aLj. For notational conve-
nience, we let L = 1 or 2 where ¬L = 2 if L = 1 and ¬L = 1 if L = 2. Let KWj
represent the vector of policy choices of length kW (aLj � KW
j ) within land-scape L that affects the payoff to policy aL
j. The kW policies with which aLj
interact are specified as the kW adjacent elements such that:
KWj ={(aL
j+1, .|.|. , aLj+kW) if kW > 0
0 if kW = 0.(2)
In addition, let KBj represent the vector of policy choices of length kB on the
other landscape ¬L that affect the payoff to policy choice aLj. Specifically, we
randomly fill the vector KBj such that on average there are kB policies with
which aL.j interact. Thus, this notation implies symmetry in values above and
below diagonal. Though we implemented and ran a model with symmetry,and the results are identical, the models employed in the paper are not sym-metrical.
KBj = {(a¬L
–1, .|.|. , a¬L
–kB) if kB > 0
0 if kB = 0.(3)
We defined the fitness contribution of an individual policy choice j on land-scape L, aL
j, as �Lj, such that:
�Lj = �L
j (a Lj |aL
– ∈KWj , a¬L
– ∈KBj ) = �L
j (aLj | KW
j , KBj ). (4)
618/ASQ, December 2007
Sorenson, O.2003 “Interdependence and adapt-
ability: Organizational learningand the long-term effect ofintegration.” ManagementScience, 49: 446–463.
Stuart, T. E., and J. M. Podolny1996 “Local search and the evolu-
tion of technological capabili-ties.” Strategic ManagementJournal, 17: 21–38.
Thompson, J. D.1967 Organizations in Action: Social
Science Bases of Administra-tive Theory. New York:McGraw-Hill.
Wilson, E. O., and W. H. Bossert1971 A Primer of Population Biolo-
gy. Sunderland, MA: SinauerAssociates.
Winter, S. G.1964 “Economic natural selection
and the theory of the firm.”Yale Economic Essays, 4:225–272.

The fitness contribution of each policy choice, �Lj , can take 21+kW+kB
uniquevalues that are assigned as independent and identically distributed drawsfrom a uniform distribution u[0,1].
When k = kW = kB = 0, the payoff to a policy choice depends on that choicealone. As k increases, the payoff to a policy choice depends not only on thatchoice but also on the value of k other policies. In general, low levels ofinteractions result in a smooth landscape (at the extreme, when kW + kB = 0,the landscape has only a single peak), and high levels of interactions result ina very rugged landscape with many local peaks, reflecting a highly complextask environment. Asymmetry in the levels of interactions within andbetween the two nearly decomposable problem spaces, where kW >> kB,produces a landscape that can be described as nearly decomposable. In theextreme, when kB = 0, the landscape is fully decomposable.
The fitness of a firm with the policy choice (location) vector NL on landscapeL is defined as the average of the fitness contribution of each of its NL policychoices such that:
1 1�L =
nL
nL
�j=1
�Lj (aL
j |aL– ∈KW
j , a¬L
– ∈KBj ) =
nL
nL
�j=1
�Lj (a
Lj |KW
j , KBj ). (5)
A firm’s fitness on the combined landscape is the average of its fitness oneach individual landscape such that:
1� =
2 (�1 + �2). (6)
Search
Adaptation is modeled as a local search process in which a firm selects aninnovation from an alternative set consisting of all policy-choice sets thatvary from the current set by a single policy attribute. Thus at any location onan individual sub-problem landscape (L = 1 or 2), there are nL alternativesfrom which to choose.
Firms are allocated two search efforts per period and as such, each period issubdivided into two sub-periods t = ta and t = tb. Search varies across organi-zational structures and methods of evaluation which can be divided into twomain forms based on organizational structure: (a) functional and (b) non-func-tional. The parallel search strategy, which employs a functional organizationalstructure with off-line evaluation, proceeds as follows. In the first sub-periodof a given period t such that t = ta, the firm selects at random one of its N1
tlandscape 1 policy choices to evaluate for possible change. Consider the pol-icy choice a1
j,t and its inverse a^1j,t (recall that policy choices are binary such
that a flip entails switching a 1 to a 0 and vice versa). Without moving to thenew location in landscape 1, the firm compares its expected fitness after theflip to its fitness prior to the flip and moves to the new location (accepts theflip) only if the move results in an improvement in fitness. The evaluation offitness is local, such that the firm considers only landscape 1 performance inassessing the outcome of a search attempt. After concluding search in land-scape 1 (L = 1), the firm then conducts search in sub-period t = tb in land-scape 2 (L = 2). More formally, if:
1nL
n L
�m=1
�Lm,t(a
Lm,t; a
L¬(m,j),t, aLj|KW
m,tKBm,t) >
1nL
nL
�m=1
�Lm,t(a
Lm,t; a
L¬(m,j) ,t , a
Lj,t|KW
m,t, KBm,t)
thenNL
t = (aL¬j,t
, a^Lj,t) otherwise,
(7){NLt = (aL
¬j,t, aL
j,t).
The second functional structure gives rise to the sequential search strategy,which is identical to the parallel strategy with the exception of the allocationof effort across the two sub-problem landscapes. Prior to period 30, sequen-
619/ASQ, December 2007
Myopia of Selection

tial-strategy firms employ both search efforts in landscape 1. Post period 30,they follow the parallel strategy outlined above, allocating one effort to eachsub-problem landscape.
Integrated search strategies come in two forms, baseline blind (on-line evalu-ation) and integrated (off-line evaluation). For these strategies, a policychoice is chosen from the global landscape Nt = {N1
t,N2t } = (a1
1, a12, .|.|. , a
1n1a2
1,a2
2, .|.|. , a2n2). Two search efforts are made, one each in sub-periods t = ta and
t = tb. The performance consequences of each search effort is evaluatedanalogously to equation 7, but taking into account the global (rather thansub-problem) landscape. On-line evaluation is done by moving immediatelyto the new location regardless of the performance consequences. If the flipcauses a performance decrease, then at the start of the next period, the firmreturns to its earlier location. Off-line evaluation is conducted (as earlier) witha move to the new location only if the move improves performance.
Selection
We examined the outcome of the developmental process under selectionregimes that vary in timing, intensity, and duration. The probability of firm isurviving a selection event in period t is defined as:
�i,tZt
.si,t = ( Max(�t)) (8)
where Max(�t) is the maximum fitness at time t by all firms in the popula-tion. Zt defines the strength of selection. Because time is discrete, we treatZt as a vector of real numbers [0, ∞] that define the strength of selection ineach time period from t = 0 to t = tmax such that Zt = (zt, .|.|. , ztMax
). A random number is drawn from a uniform distribution u[0,1]. If si,t is greater than orequal to the realization of the draw, then the firm survives. When Zt = 0, theprobability of survival for firm i in period t is si,t = 1 (100%). As Zt increases,the probability of survival conditional on the firm’s fitness declines.
Simulation Model
The simulation models 200 firms as they search under risk of selection for100 periods (tmax = 100). The simulation proceeds as follows. At time t = 0,each firm is randomly allocated an initial location in both landscape 1, N1
i,0,and landscape 2, N2
i,0. Each period t is divided into two sub-periods (parts aand b), and firms are allocated two search efforts per period. At the end ofeach full period, after all firms have completed their search effort, firms aresubject to selection. Results are averaged over 100 runs of the simulation.
620/ASQ, December 2007