na+ requirement for growth, photosynthesis, and ...jb.asm.org/content/159/1/100.full.pdf · the...
TRANSCRIPT

Vol. 159, No. 1
Na+ Requirement for Growth, Photosynthesis, and pH Regulation inthe Alkalotolerant Cyanobacterium Synechococcus leopoliensis
ANTHONY G. MILLER, DAVID H. TURPIN, AND DAVID T. CANVIN*Department of Biology, Queen's University, Kingston, Ontario, Canada, K7L 3N6
Received 22 November 1983/Accepted 15 April 1984
We have found that Na+ is required for the alkalotolerance of the cyanobacterium Synechococcusleopoliensis. Cell division did not occur at any pH in the absence of Na+, but cells inoculated into Na+-freegrowth medium at pH 6.8 did continue metabolic activity, and over a period of 48 h, the cells became twicetheir normal size. Many of these cells remained viable for at least 59 h and formed colonies on Na+-containingmedium. Cells grown in the presence of Na+ and inoculated into Na+-free growth medium at pH 9.6 rapidlylost viability. An Na+ concentration of ca. 0.5 milliequivalents -Iiter-' was required for sustained growthabove pH 9.0. The Na+ requirement could be only partially met by Li' and not at all by K+ or Rb+. Cellsincubated in darkness in growth medium at pH 6.8 had an intracellular pH near neutrality in the presence orabsence of Na+. When the external pH was shifted to 9.6, only cells in the presence of Na+ were able tomaintain an intracellular pH near 7.0. The membrane potential, however, remained high (-120 mV) in theabsence or presence of Na+ unless collapsed by the addition of gramicidin. Thus, the inability to maintain aneutral intracellular pH at pH 9.6 in the absence of Na+ was not due to a generalized disruption of membraneintegrity. Even cells containing Na+ still required added Na+ to restore photosynthetic rates to normal after thecells had been washed in Na+-free buffer at pH 9.6. This requirement was only partially met by Li' and wasnot met at all by K+, Rb+, Cs+, Mg2+, or Ca2+. The restoration of photosynthesis by added Na+ occurredwithin 30 s and suggests a role for extracellular Na+. Part of our results can be explained in terms of theoperation of an Na+/H+ antiporter activity in the plasma membrane, but some results would seem to requireother mechanisms for Na+ action.
Most organisms live within the limits of pH 4 to 9 and havegrowth optima near neutrality (19). Organisms that havegrowth optima above pH 9 have been termed alkalophilic,and those that have growth optima below pH 9, but cansurvive extended exposure above it, have been termedalkalotolerant (19). The cyanobacteria as a group are charac-teristically alkalotolerant or alkalophilic (19, 30). In fact, themedia originally designed for their growth were effectivelyunbuffered (30), but growth still continued even when the pHrose above 10.5 as a result of CO2 or HC03- uptake duringphotosynthesis (7). This ability to grow at alkaline pHimplies the presence of mechanisms for the maintenance ofan intracellular pH (pHj) more acidic than that of theexternal milieu.
Several alkalophilic Bacillus species maintain their cyto-plasm more acidic than the external medium by the exchangeof intracellular Na+ for external H+ (3, 15, 17, 21, 25). Thisexchange is mediated by membrane-associated Na+/H+antiporter activity, which, at alkaline pH, appears to operatein an electrogenic fashion and is driven by the membranepotential (21). In at least one species, Bacillus firmus, thereis the expected, obligate requirement of Na+ for growth (17),and mutants of Bacillus alcalophilus that lack Na+/H+antiporter activity are unable to grow above pH 9 (18).Operation of the Na+/H+ antiporter results in a substantialinwardly directed gradient for Na+ ions (18), and use is madeof this for Na+ solute symport (2, 3, 8, 9) into the cells andeven for flagellar rotation coupled to Na+ reentry into thecells (11). Thus, a substantial Na+ circulation across theplasma membrane of alkalophilic bacteria seems to be cen-tral to their ability to grow and survive above pH 9.
* Corresponding author.
The inability of various cyanobacteria of the genus Syne-chococcus to grow at acid pH has been previously studied(4, 13), but we are not aware of any studies on the mecha-nisms allowing cyanobacteria to grow at alkaline pH. Sincethe work of Kratz and Myers (16), it has been well knownthat a considerable concentration of Na+ is required forgrowth of some cyanobacteria. In light of the work withalkalophilic Bacillus species and the known Na' require-ment for Synechococcus leopoliensis (16), the importance ofNa+ to the ability of this species to grow above pH 9 wasinvestigated. The results presented in this paper are consis-tent with the involvement of Na+/H+ antiporter activity inthe alkalotolerance of S. leopoliensis, but additional effectsof Na+ were also noted.
MATERIALS AND METHODSOrganism and growth conditions. S. leopoliensis UTEX
625, which is also known as Synechococcus sp. strain PCC6301 and Anacystis nidulans Tx2O (27), was obtained as anaxenic culture from the culture collection at the Universityof Texas, Austin. It was maintained under low illumination(15 microeinsteins m2
2 s-') on slants prepared with Allenmedium (1).Normal Allen medium contains 17.6 milliequivalents
(mEq) of Na+ - liter-' as NaNO3, 0.20 mEq of Na' * liter-'as Na2SiO3 * 9H20, 0.19 mEq of Na+ - liter-1 as Na2CO3,and trace amounts as sodium molybdate (1). We routinelyomit the sodium silicate for growth of unicellular cyanobac-teria in liquid cultures. The NaNO3 was replaced with 8.8mM KNO3, and the Na2CO3 was replaced with 0.19 mMK2CO3. The sodium molybdate in the trace element solutionwas replaced with molybdic acid titrated to pH 7.0 withKOH and was kept in a stock solution separate from the restof the trace elements to prevent precipitation. The K2HPO4
100
JOURNAL OF BACTERIOLOGY, JUlY 1984, p. 100-1060021-9193/84/070100-07$02.00/0Copyright © 1984, American Society for Microbiology
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
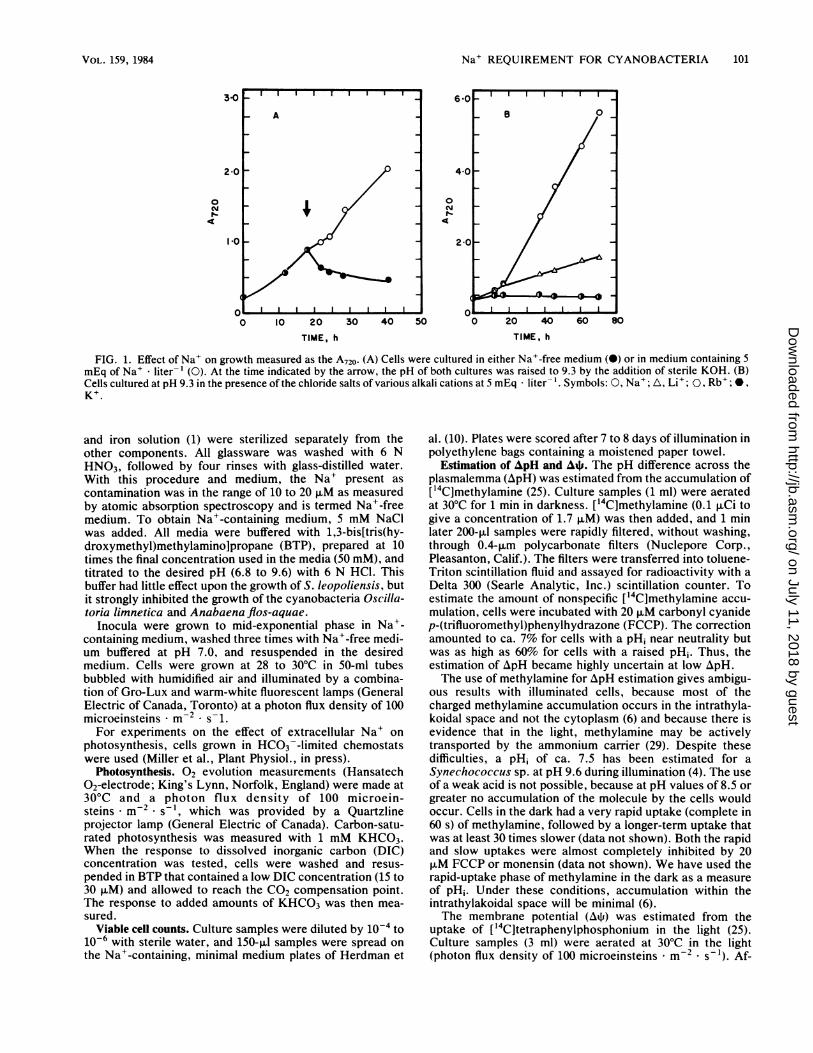
Na+ REQUIREMENT FOR CYANOBACTERIA 101
3-0
2-0
0
4C
I I II ] 20, ~
0 10 20 30 40 50 0 20 40 60 80
TIME, h TIME, h
FIG. 1. Effect of Na+ on growth measured as the A720. (A) Cells were cultured in either Na+-free medium (0) or in medium containing 5mEq of Na+ * liter-' (0). At the time indicated by the arrow, the pH of both cultures was raised to 9.3 by the addition of sterile KOH. (B)Cells cultured at pH 9.3 in the presence of the chloride salts of various alkali cations at 5 mEq * liter-'. Symbols: 0, Na+; A, Li+; 0, Rb+; 0,K+.
and iron solution (1) were sterilized separately from theother components. All glassware was washed with 6 NHNO3, followed by four rinses with glass-distilled water.With this procedure and medium, the Na+ present as
contamination was in the range of 10 to 20 puM as measuredby atomic absorption spectroscopy and is termed Na+-freemedium. To obtain Na+-containing medium, 5 mM NaClwas added. All media were buffered with 1,3-bis[tris(hy-droxymethyl)methylamino]propane (BTP), prepared at 10times the final concentration used in the media (50 mM), andtitrated to the desired pH (6.8 to 9.6) with 6 N HCI. Thisbuffer had little effect upon the growth of S. leopoliensis, butit strongly inhibited the growth of the cyanobacteria Oscilla-toria limnetica and Anabaena flos-aquae.
Inocula were grown to mid-exponential phase in Na+-containing medium, washed three times with Na+-free medi-um buffered at pH 7.0, and resuspended in the desiredmedium. Cells were grown at 28 to 30°C in 50-ml tubesbubbled with humidified air and illuminated by a combina-tion of Gro-Lux and warm-white fluorescent lamps (GeneralElectric of Canada, Toronto) at a photon flux density of 100microeinsteins -m-2 * s-1.For experiments on the effect of extracellular Na+ on
photosynthesis, cells grown in HCO3 -limited chemostatswere used (Miller et al., Plant Physiol., in press).
Photosynthesis. 02 evolution measurements (Hansatech02-electrode; King's Lynn, Norfolk, England) were made at30°C and a photon flux density of 100 microein-steins* m2 51 which was provided by a Quartzlineprojector lamp (General Electric of Canada). Carbon-satu-rated photosynthesis was measured with 1 mM KHCO3.When the response to dissolved inorganic carbon (DIC)concentration was tested, cells were washed and resus-pended in BTP that contained a low DIC concentration (15 to30 ,uM) and allowed to reach the CO2 compensation point.The response to added amounts of KHCO3 was then mea-sured.
Viable cell counts. Culture samples were diluted by 10' to10-6 with sterile water, and 150-pul samples were spread onthe Na+-containing, minimal medium plates of Herdman et
al. (10). Plates were scored after 7 to 8 days of illumination inpolyethylene bags containing a moistened paper towel.
Estimation of ApH and A+. The pH difference across theplasmalemma (ApH) was estimated from the accumulation of["4C]methylamine (25). Culture samples (1 ml) were aeratedat 30°C for 1 min in darkness. [14C]methylamine (0.1 ,Ci togive a concentration of 1.7 ,uM) was then added, and 1 minlater 200-,ul samples were rapidly filtered, without washing,through 0.4-,um polycarbonate filters (Nuclepore Corp.,Pleasanton, Calif.). The filters were transferred into toluene-Triton scintillation fluid and assayed for radioactivity with aDelta 300 (Searle Analytic, Inc.) scintillation counter. Toestimate the amount of nonspecific [14C]methylamine accu-mulation, cells were incubated with 20 puM carbonyl cyanidep-(trifluoromethyl)phenylhydrazone (FCCP). The correctionamounted to ca. 7% for cells with a pH, near neutrality butwas as high as 60% for cells with a raised pH1. Thus, theestimation of ApH became highly uncertain at low ApH.The use of methylamine for ApH estimation gives ambigu-
ous results with illuminated cells, because most of thecharged methylamine accumulation occurs in the intrathyla-koidal space and not the cytoplasm (6) and because there isevidence that in the light, methylamine may be activelytransported by the ammonium carrier (29). Despite thesedifficulties, a pHi of ca. 7.5 has been estimated for aSynechococcus sp. at pH 9.6 during illumination (4). The useof a weak acid is not possible, because at pH values of 8.5 orgreater no accumulation of the molecule by the cells wouldoccur. Cells in the dark had a very rapid uptake (complete in60 s) of methylamine, followed by a longer-term uptake thatwas at least 30 times slower (data not shown). Both the rapidand slow uptakes were almost completely inhibited by 20puM FCCP or monensin (data not shown). We have used therapid-uptake phase of methylamine in the dark as a measureof pHi. Under these conditions, accumulation within theintrathylakoidal space will be minimal (6).The membrane potential (A/v) was estimated from the
uptake of [14C]tetraphenylphosphonium in the light (25).Culture samples (3 ml) were aerated at 30°C in the light(photon flux density of 100 microeinsteins * m-2 s - 1). Af-
VOL. 159, 1984
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
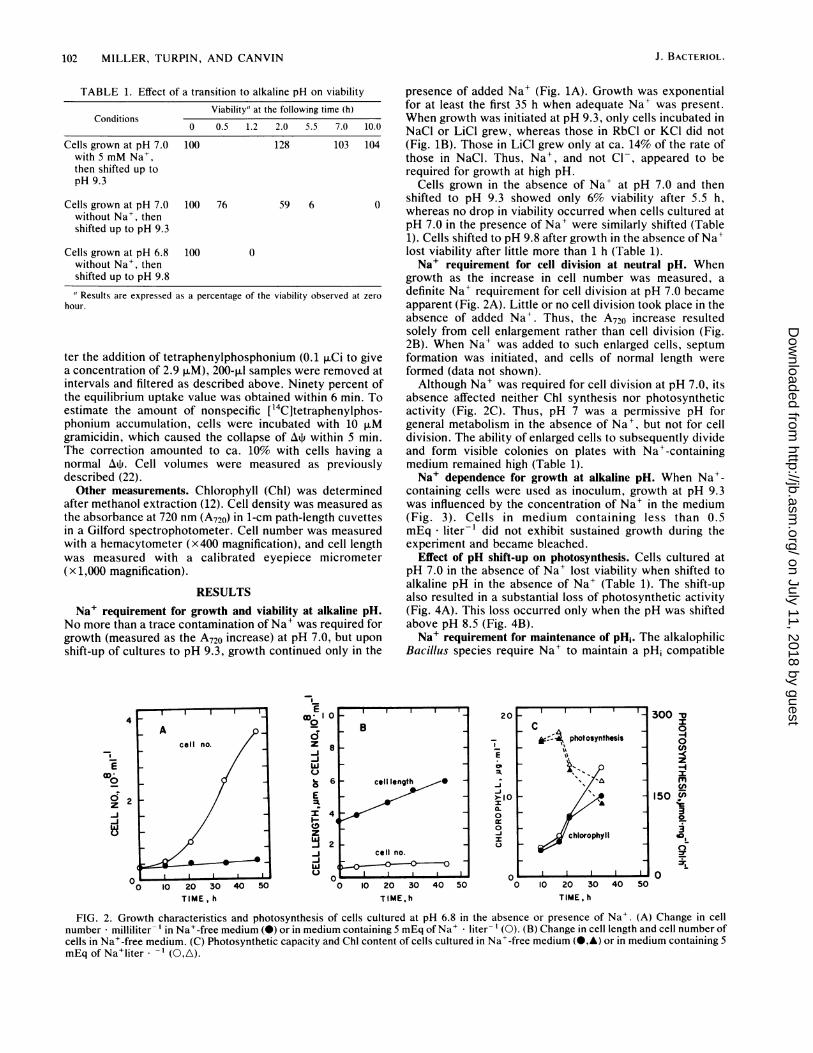
102 MILLER, TURPIN, AND CANVIN
TABLE 1. Effect of a transition to alkaline pH on viabilityViability" at the following time (h)
Conditions0 0.5 1.2 2.0 5.5 7.0 10.0
Cells grown at pH 7.0 100 128 103 104with 5 mM Na+,then shifted up topH 9.3
Cells grown at pH 7.0 100 76 59 6 0without Na+, thenshifted up to pH 9.3
Cells grown at pH 6.8 100 0without Na+, thenshifted up to pH 9.8
" Results are expressed as a percentage of the viability observed at zerohour.
ter the addition of tetraphenylphosphonium (0.1 ,uCi to givea concentration of 2.9 ,uM), 200-,ul samples were removed atintervals and filtered as described above. Ninety percent ofthe equilibrium uptake value was obtained within 6 min. Toestimate the amount of nonspecific [14C]tetraphenylphos-phonium accumulation, cells were incubated with 10 ,uMgramicidin, which caused the collapse of Aip within 5 min.The correction amounted to ca. 10% with cells having anormal Ai,. Cell volumes were measured as previouslydescribed (22).Other measurements. Chlorophyll (Chl) was determined
after methanol extraction (12). Cell density was measured asthe absorbance at 720 nm (A720) in 1-cm path-length cuvettesin a Gilford spectrophotometer. Cell number was measuredwith a hemacytometer (x400 magnification), and cell lengthwas measured with a calibrated eyepiece micrometer(x 1,000 magnification).
RESULTSNa+ requirement for growth and viability at alkaline pH.
No more than a trace contamination of Na+ was required forgrowth (measured as the A720 increase) at pH 7.0, but uponshift-up of cultures to pH 9.3, growth continued only in the
presence of added Na+ (Fig. 1A). Growth was exponentialfor at least the first 35 h when adequate Na+ was present.When growth was initiated at pH 9.3, only cells incubated inNaCl or LiCl grew, whereas those in RbCl or KCI did not(Fig. iB). Those in LiCl grew only at ca. 14% of the rate ofthose in NaCl. Thus, Na+, and not Cl-, appeared to berequired for growth at high pH.
Cells grown in the absence of Na+ at pH 7.0 and thenshifted to pH 9.3 showed only 6% viability after 5.5 h,whereas no drop in viability occurred when cells cultured atpH 7.0 in the presence of Na+ were similarly shifted (Table1). Cells shifted to pH 9.8 after growth in the absence of Na+lost viability after little more than 1 h (Table 1).Na+ requirement for cell division at neutral pH. When
growth as the increase in cell number was measured, adefinite Na+ requirement for cell division at pH 7.0 becameapparent (Fig. 2A). Little or no cell division took place in theabsence of added Na+. Thus, the A720 increase resultedsolely from cell enlargement rather than cell division (Fig.2B). When Na+ was added to such enlarged cells, septumformation was initiated, and cells of normal length wereformed (data not shown).Although Na+ was required for cell division at pH 7.0, its
absence affected neither Chl synthesis nor photosyntheticactivity (Fig. 2C). Thus, pH 7 was a permissive pH forgeneral metabolism in the absence of Na', but not for celldivision. The ability of enlarged cells to subsequently divideand form visible colonies on plates with Na+-containingmedium remained high (Table 1).Na+ dependence for growth at alkaline pH. When Na+-
containing cells were used as inoculum, growth at pH 9.3was influenced by the concentration of Na+ in the medium(Fig. 3). Cells in medium containing less than 0.5mEq * liter-' did not exhibit sustained growth during theexperiment and became bleached.
Effect of pH shift-up on photosynthesis. Cells cultured atpH 7.0 in the absence of Na+ lost viability when shifted toalkaline pH in the absence of Na+ (Table 1). The shift-upalso resulted in a substantial loss of photosynthetic activity(Fig. 4A). This loss occurred only when the pH was shiftedabove pH 8.5 (Fig. 4B).Na+ requirement for maintenance of pH;. The alkalophilic
Bacillus species require Na+ to maintain a pH, compatible
10 20 30 40 50
TIME, h
I
D6I 0
0z 8-J-Jw_'-60E
164I-
z.J2-j-Jw-)
v0 10 20 30 40 50
TIME,h
20
IO
E
0
-i-J
100
0-J
0
0 10 20 30
TIME, h
FIG. 2. Growth characteristics and photosynthesis of cells cultured at pH 6.8 in the absence or presence of Na+. (A) Change in cellnumber milliliter-' in Na+-free medium (0) or in medium containing 5 mEq of Na+ liter- l (0). (B) Change in cell length and cell number ofcells in Na+-free medium. (C) Photosynthetic capacity and Chl content of cells cultured in Na+-free medium (@4,) or in medium containing 5mEq of Na+liter * -l (0,A).
4
IE
0602z
0i
B
cell length g
cell no.
4 photosynthesis
- 'I
chlorophyll
1I
300 -VI0
0
z-.4IC
=10
40 50
J. BACTERIOL.
I
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
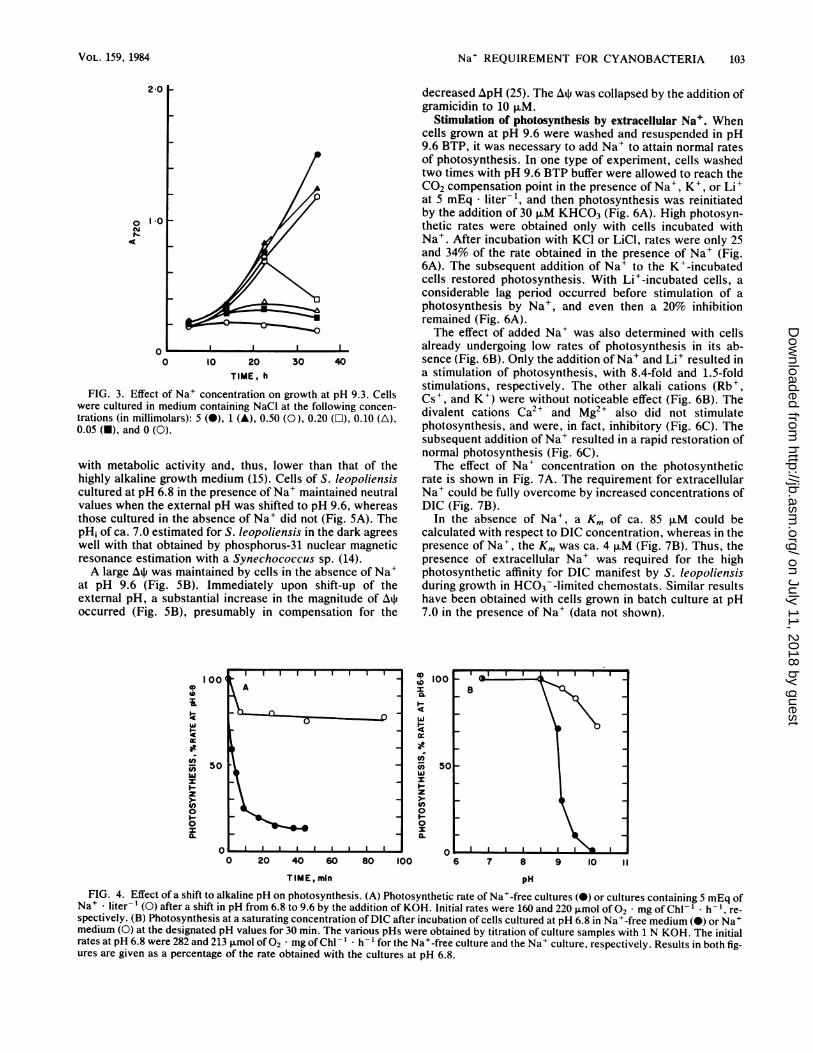
Na+ REQUIREMENT FOR CYANOBACTERIA 103
2-0
0
4
I-*oF
0 '0
I
10 20TIME, h
30 40
FIG. 3. Effect of Na+ concentration on growth at pH 9.3. Cellswere cultured in medium containing NaCl at the following concen-trations (in millimolars): 5 (0), 1 (A), 0.50 (0), 0.20 (O), 0.10 (A),0.05 (O), and 0 (0).
with metabolic activity and, thus, lower than that of thehighly alkaline growth medium (15). Cells of S. leopoliensiscultured at pH 6.8 in the presence of Na+ maintained neutralvalues when the external pH was shifted to pH 9.6, whereasthose cultured in the absence of Na+ did not (Fig. SA). ThepH1 of ca. 7.0 estimated for S. leopoliensis in the dark agreeswell with that obtained by phosphorus-31 nuclear magneticresonance estimation with a Synechococcus sp. (14).A large Ai was maintained by cells in the absence of Na+
at pH 9.6 (Fig. 5B). Immediately upon shift-up of theexternal pH, a substantial increase in the magnitude of A*occurred (Fig. SB), presumably in compensation for the
decreased ApH (25). The Ap was collapsed by the addition ofgramicidin to 10 ,M.
Stimulation of photosynthesis by extracellular Na+. Whencells grown at pH 9.6 were washed and resuspended in pH9.6 BTP, it was necessary to add Na+ to attain normal ratesof photosynthesis. In one type of experiment, cells washedtwo times with pH 9.6 BTP buffer were allowed to reach theCO2 compensation point in the presence of Na+, K+, or Li'at 5 mEq * liter-', and then photosynthesis was reinitiatedby the addition of 30 FM KHCO3 (Fig. 6A). High photosyn-thetic rates were obtained only with cells incubated withNa+. After incubation with KCI or LiCl, rates were only 25and 34% of the rate obtained in the presence of Na+ (Fig.6A). The subsequent addition of Na+ to the K+-incubatedcells restored photosynthesis. With Li+-incubated cells, aconsiderable lag period occurred before stimulation of aphotosynthesis by Na+, and even then a 20% inhibitionremained (Fig. 6A).The effect of added Na+ was also determined with cells
already undergoing low rates of photosynthesis in its ab-sence (Fig. 6B). Only the addition of Na+ and Li+ resulted ina stimulation of photosynthesis, with 8.4-fold and 1.5-foldstimulations, respectively. The other alkali cations (Rb+,Cs+, and K+) were without noticeable effect (Fig. 6B). Thedivalent cations Ca2+ and Mg2+ also did not stimulatephotosynthesis, and were, in fact, inhibitory (Fig. 6C). Thesubsequent addition of Na+ resulted in a rapid restoration ofnormal photosynthesis (Fig. 6C).The effect of Na+ concentration on the photosynthetic
rate is shown in Fig. 7A. The requirement for extracellularNa+ could be fully overcome by increased concentrations ofDIC (Fig. 7B).
In the absence of Na+, a Km of ca. 85 ,uM could becalculated with respect to DIC concentration, whereas in thepresence of Na+, the Km was ca. 4 ,uM (Fig. 7B). Thus, thepresence of extracellular Na+ was required for the highphotosynthetic affinity for DIC manifest by S. leopoliensisduring growth in HCO3 -limited chemostats. Similar resultshave been obtained with cells grown in batch culture at pH7.0 in the presence of Na+ (data not shown).
0 20 40 60 80
TIME, min
OD 100
10 6
4
w
r.-
~i 50w
I.-
(a0
I~-0x
0*
100 6 7 8 9 10 11
pHFIG. 4. Effect of a shift to alkaline pH on photosynthesis. (A) Photosynthetic rate of Na+-free cultures (0) or cultures containing 5 mEq of
Na+ * liter-' (0) after a shift in pH from 6.8 to 9.6 by the addition of KOH. Initial rates were 160 and 220 pLmol of 02 * mg of Chl- ' h-l, re-
spectively. (B) Photosynthesis at a saturating concentration of DIC after incubation of cells cultured at pH 6.8 in Na+-free medium (0) or Na+medium (0) at the designated pH values for 30 min. The various pHs were obtained by titration of culture samples with 1 N KOH. The initialrates at pH 6.8 were 282 and 213 jLmol of02 * mg of Chl-' * h 1 for the Na+-free culture and the Na+ culture, respectively. Results in both fig-ures are given as a percentage of the rate obtained with the cultures at pH 6.8.
I I I I I I I
O -
_ |~~
l l l
I 0000
!iI
4
I4a2U;
50
zI-(00
0
VOL. 159, 1984
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

104 MILLER, TURPIN, AND CANVIN
DISCUSSION
The absence of Na+ had pleiotropic effects upon thegrowth and metabolism of the cyanobacterium S. leopolien-sis. At pH 6.8 in the absence of Na+, cells were able tophotosynthesize normally (Fig. 2C) but were unable todivide (Fig. 2A), and biomass increase was by cell enlarge-ment (Fig. 2B). Recently, Roussard-Jacquemin (28) hasshown that a strain of S. leopoliensis different from strainUTEX 625 also requires Na+ for cell division at pH 7.2 andthat cell enlargement occurred in the absence of Na+. Heperformed no studies above pH 7.2. We have found that Na+was required for viability (Table 1) and growth (Fig. 1) atalkaline pH.Na+ was also required for the maintenance of pH, at
alkaline pH (Fig. 5). In the heterotrophic bacteria, intracellu-lar Na+ is required for maintenance of pH, values more
acidic than the strongly alkaline medium, because it is Na+that is translocated outwards in exchange for the protonsneeded to acidify the cytoplasm (25). Paschinger (26) haspresented evidence for an Na+/H+ antiporter in S. leopolien-sis. He found that cells poisoned with dicyclohexylcarbodii-mide no longer actively pumped out Na+, but instead tookup Na+ in exchange for intracellular H+ (26). He interpretedthis exchange as the reversal in function of a transporternormally involved in exchanging intracellular Na+ for exter-nal H+. The requirement for intracellular Na+ for pHregulation in S. leopoliensis (Fig. SA) is consistent with theinvolvement of an Na+/H+ antiporter. The membrane poten-tial remained high at alkaline pH in the absence of Na+ (Fig.SB). This suggests that the absence of Na+ at alkaline pH didnot cause a general disruption of membrane integrity. Theinability of the cells to maintain a neutral pH, was thus due toa more specific lesion in the pH regulatory mechanism.One route of reentry for Na+ into cells of alkalophilic
bacteria (9) or Synechococcus sp. (20) is via Na+/solutesymport systems, such as the one for aminoisobutyric acid.
4
-J
w
(.1
41
~--
z
TIME, min
E
I.-
zw
0
La
TI ME, min
FIG. 5. Effect of a shift to alkaline pH on pH, and Ai/. Cells were
cultured at pH 7.0 in Na+-free medium (0) or in medium containing5 mEq of Na+ * liter-' (0), and the pH was then raised to 9.6 bytitration with 1 N KOH. (A) Effect on pHi in the dark. (B) Effect on
Aq of cells cultured in Na+-free medium. The pH was raised at thetime indicated by the first arrow, and the medium was made 10 p.Mwith respect to gramicidin at the second arrow. The cell volumeswere 285 and 95 p.l * mg of Chl-' for Na+-free cells and Na+-cultured cells, respectively.
BI min
16 nmol 02
66N K 32 Li+ s Rb+22t4 4 4
K64 164
2032
IT 4 No+
No 542.0-5Me
Ca++
FIG. 6. Effect of Na+ upon photosynthesis at alkaline pH. Allcells were grown in an HC03--limited chemostat at a growth rate of1.08 day-' (ca. 53% L,Umax). (A) Cells were allowed to reach the CO-compensation point in BTP buffer (pH 9.6) containing Na+, K+, orLi+ at 5 mEq *liter-'. Photosynthesis was then reinitiated by theaddition of 30 ptM DIC as KHCO3. (B) After the addition of 30 ,uMDIC as KHCO3, the effect on photosynthesis of Na+, K+, Cs+, orRb+ at 5 mEq * liter- l was tested. (C) As for B, but Ca2+ or Mg2+ atthe stated concentrations were used. The rates by the traces aregiven as micromoles of 02 * milligram of Chl-V hour-'.
Movement of Na+ back into the cells is down the electro-chemical gradient for Na+ (5, 26). Such symport systemswould be energetically more favorable at strongly alkalinepH than the more usual H+ symport systems operational atlower pH (25). In such Na+/solute symport systems, thepresence of Na+ seems only to reduce the Km for thecotransported solute and has no effect on Vmax (9). That is,the requirement of Na+ for solute influx can be overcome if
J. BACTERIOL.
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
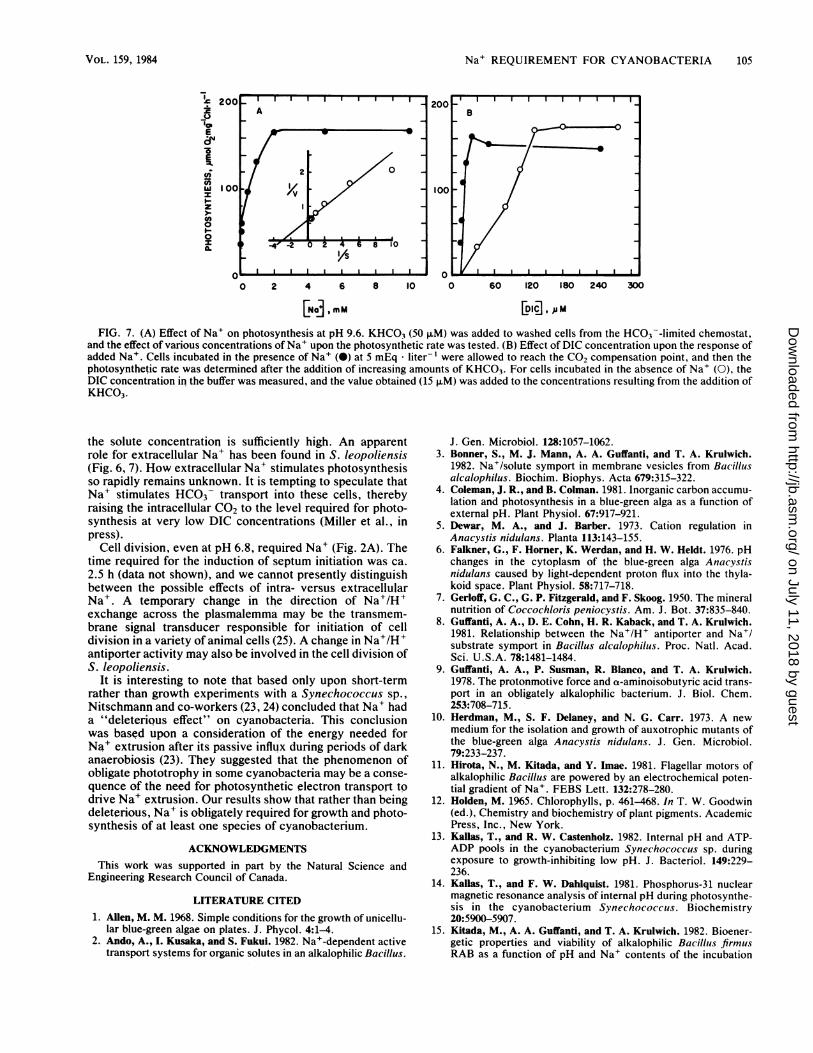
Na+ REQUIREMENT FOR CYANOBACTERIA 105
P
EU;
(nw
z
001--0Ia.
Na3, mm
0 60 120 180 240 300
LDIC,I M
FIG. 7. (A) Effect of Na+ on photosynthesis at pH 9.6. KHCO3 (50 ,uM) was added to washed cells from the HCO3--limited chemostat,and the effect of various concentrations of Na+ upon the photosynthetic rate was tested. (B) Effect of DIC concentration upon the response ofadded Na+. Cells incubated in the presence of Na+ (0) at 5 mEq lliter-' were allowed to reach the CO2 compensation point, and then thephotosynthetic rate was determined after the addition of increasing amounts of KHCO3. For cells incubated in the absence of Na+ (0), theDIC concentration in the buffer was measured, and the value obtained (15 FiM) was added to the concentrations resulting from the addition ofKHCO3.
the solute concentration is sufficiently high. An apparentrole for extracellular Na+ has been found in S. leopoliensis(Fig. 6, 7). flow extracellular Na+ stimulates photosynthesisso rapidly remains unknown. It is tempting to speculate thatNa+ stimulates HCO3 transport into these cells, therebyraising the intracellular CO2 to the level required for photo-synthesis at very low DIC concentrations (Miller et al., inpress).
Cell division, even at pH 6.8, required Na+ (Fig. 2A). Thetime required for the induction of septum initiation was ca.2.5 h (data not shown), and we cannot presently distinguishbetween the possible effects of intra- versus extracellularNa+. A temporary change in the direction of Na+/H+exchange across the plasmalemma may be the transmem-brane signal transducer responsible for initiation of celldivision in a variety of animal cells (25). A change in Na+/H+antiporter activity may also be involved in the cell division ofS. leopoliensis.
It is interesting to note that based only upon short-termrather than growth experiments with a Synechococcus sp.,Nitschmann and co-workers (23, 24) concluded that Na+ hada "deleteriQus effect" on cyanobacteria. This conclusionwas based upon a consideration of the energy needed forNa+ extrusion after its passive influx during periods of darkanaerobiosis (23). They suggested that the phenomenon ofobligate phototrophy in some cyanobacteria may be a conse-quence of the need for photosynthetic electron transport todrive Na+ extrusion. Our results show that rather than beingdeleterious, Na+ is obligately required for growth and photo-synthesis of at least one species of cyanobacterium.
ACKNOWLEDGMENTSThis work was supported in part by the Natural Science and
Engineering Research Council of Canada.
LITERATURE CITED1. Alien, M. M. 1968. Simple conditions for the growth of unicellu-
lar blue-green algae on plates. J. Phycol. 4:1-4.2. Ando, A., I. Kusaka, and S. Fukui. 1982. Na'-dependent active
transport systems for organic solutes in an alkalophilic Bacillus.
J. Gen. Microbiol. 128:1057-1062.3. Bonner, S., M. J. Mann, A. A. Guffanti, and T. A. Krulwich.
1982. Na+/solute symport in membrane vesicles from Baccillusalcalophilus. Biochim. Biophys. Acta 679:315-322.
4. Coleman, J. R., and B. Colman. 1981. Inorganic carbon accumu-lation and photosynthesis in a blue-green alga as a function ofexternal pH. Plant Physiol. 67:917-921.
5. Dewar, M. A., and J. Barber. 1973. Cation regulation inAnacystis nidulans. Planta 113:143-155.
6. Falkner, G., F. Horner, K. Werdan, and H. W. Heldt. 1976. pHchanges in the cytoplasm of the blue-green alga Anacystisnidulans caused by light-dependent proton flux into the thyla-koid space. Plant Physiol. 58:717-718.
7. Gerloff, G. C., G. P. Fitzgerald, and F. Skoog. 1950. The mineralnutrition of Coccochloris peniocystis. Am. J. Bot. 37:835-840.
8. Guffanti, A. A., D. E. Cohn, H. R. Kaback, and T. A. Krulwich.1981. Relationship between the Na+/H+ antiporter and Na+/substrate symport in Bacillus alcalophilus. Proc. Natl. Acad.Sci. U.S.A. 78:1481-1484.
9. Guffanti, A. A., P. Susman, R. Blanco, and T. A. Krulwich.1978. The protonmotive force and a-aminoisobutyric acid trans-port in an obligately alkalophilic bacterium. J. Biol. Chem.253:708-715.
10. Herdman, M., S. F. Delaney, and N. G. Carr. 1973. A newmedium for the isolation and growth of auxotrophic mutants ofthe blue-green alga Anacystis nidulans. J. Gen. Microbiol.79:233-237.
11. Hirota, N., M. Kitada, and Y. Imae. 1981. Flagellar motors ofalkalophilic Bacillus are powered by an electrochemical poten-tial gradient of Na+. FEBS Lett. 132:278-280.
12. Holden, M. 1965. Chlorophylls, p. 461-468. In T. W. Goodwin(ed.), Chemistry and biochemistry of plant pigments. AcademicPress, Inc., New York.
13. Kallas, T., and R. W. Castenholz. 1982. Internal pH and ATP-ADP pools in the cyanobacterium Synechococcus sp. duringexposure to growth-inhibiting low pH. J. Bacteriol. 149:229-236.
14. Kallas, T., and F. W. Dahlquist. 1981. Phosphorus-31 nuclearmagnetic resonance analysis of internal pH during photosynthe-sis in the cyanobacterium Synechococcus. Biochemistry20:5900-5907.
15. Kitada, M., A. A. Guffanti, and T. A. Krulwich. 1982. Bioener-getic properties and viability of alkalophilic Bacillus firmusRAB as a function of pH and Na+ contents of the incubation
VOL. 159, 1984
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

106 MILLER, TURPIN, AND CANVIN
medium. J. Bacteriol. 152:1096-1104.16. Kratz, W. A., and J. Myers. 1955. Nutrition and growth of
several blue-green algae. Am. J. Bot. 42:282-287.17. Krulwich, T. A., A. A. Guffanti, R. F. Bornstein, and J.
Hoffstein. 1982. A sodium requirement for growth, solute trans-port, and pH homeostasis in Bacillus firmus RAB. J. Biol.Chem. 257:1885-1889.
18. Krulwich, T. A., K. G. Mandel, R. F. Borstein, and A. A.Guffanti. 1979. A non-alkalophilic mutant of Bacillus alcalo-philus. Biochem. Biophys. Res. Commun. 91:58-62.
19. Langworthy, T. A. 1978. Microbial life in extreme pH values, p.
279-315. In D. J. Kushner (ed.). Microbial life in extremeenvironments. Academic Press, Inc., London.
20. Lee-Kaden, J., and W. Simonis. 1982. Amino acid uptake andenergy coupling dependent on photosynthesis in Anacystisnidulans. J. Bacteriol. 151:229-236.
21. Mandel, K. G., A. A. Guffanti, and T. A. Krulwich. 1980.Monovalent cation/proton antiporters in membrane vesiclesfrom Bacillus alcalophilus. J. Biol. Chem. 255:7391-7396.
22. Miller, A. G., and B. Colman. 1980. Active transport andaccumulation of bicarbonate by a unicellular cyanobacterium. J.Bacteriol. 143:1253-1259.
23. Nitschmann, W. H., and G. A. Peschek. 1982. Oxidative phos-
phorylation in intact cyanobacteria. FEBS Lett. 139:77-80.24. Nitschmann, W. H., G. Schmetterer, R. Muchl, and G. A.
Peschek. 1982. Active sodium extrusion reduces net efficienciesof oxidative phosphorylation in the strictly photoautotrophiccyanobacterium Anacystis nidulans. Biochim. Biophys. Acta682:293-296.
25. Padan, E., D. Zilberstein, and S. Schuldiner. 1981. pH homeo-stasis in bacteria. Biochim. Biophys. Acta 650:151-166.
26. Paschinger, H. 1977. DCCD induced sodium uptake by Anacys-tis nidulans. Arch. Microbiol. 113:285-291.
27. Rippka, R., J. Deruelles, J. B. Waterbury, M. Herd-man, and R. Y. Stanier. 1979. Generic assignments, strainhistories and properties of pure cultures of cyanobacteria. J.Gen. Microbiol. 111:1-61.
28. Roussard-Jacquemin, M. 1982. Sur quelques aspects du r6leessential du sodium chez Anacystis nidulans. Z. Pflanzenphy-siol. 108:305-316.
29. Turpin, D. H., S. A. Edie, and D. T. Canvin. In vivo nitrogenaseregulation by ammonium and methylamine and the effect ofMSX on ammonium transport in Anabaena flos-aquae. PlantPhysiol. 74:701-704.
30. Wolk, C. P. 1973. Physiology and cytological chemistry of blue-green algae. Bacteriol. Rev. 37:32-101.
J. BACTERIOL.
on July 11, 2018 by guesthttp://jb.asm
.org/D
ownloaded from