nafenopin-induced hepatic microbody (peroxisome
TRANSCRIPT

NAFENOPIN-INDUCED HEPATIC MICROBODY
(PEROXISOME) PROLIFERATION AND CATALASE
SYNTHESIS IN RATS AND MICE
Absence of Sex Difference in Response
JARNARDAN K . REDDY, DANIEL L . AZARNOFF,DONALD J . SVOBODA, and JADA D . PRASAD
From the Department of Pathology and Oncology and the Clinical Pharmacology and ToxicologyCenter, University of Kansas Medical Center, Kansas City, Kansas 66103
ABSTRACTNafenopin (2-methyl-2[p-(1,2,3,4-tetrahydro-l-naphthyl)phenoxy]-propionic acid ; Su-13437), a potent hypolipidemic compound, was administered in varying concentrations inground Purina Chow to male and female rats, wild type (Csa strain) mice and acatalasemic(Csb strain) mice to determine the hepatic microbody proliferative and catalase-inducingeffects. In all groups of animals, administration of nafenopin at dietary levels of 0.125%and 0.25% produced a significant and sustained increase in the number of peroxisomes .The hepatic microbody proliferation in both male and female rats and wild type Csa strainmice treated with nafenopin was of the same magnitude and was associated with a two-foldincrease in catalase activity and in the concentration of catalase protein . The increase inmicrobody population in acatalasemic mice, although not accompanied by increase incatalase activity, was associated with a twofold increase in the amount of catalase protein .The absence of sex difference in microbody proliferative response in nafenopin-treated ratsand wild type mice is of particular significance, since ethyl-a-p-chlorophenoxyisobutyrate(CPIB)-induced microbody proliferation and increase in catalase activity occurred only inmales. Nafenopin can, therefore, be used as an inducer of microbody proliferation and ofcatalase synthesis in both sexes of rats and mice . The serum glycerol-glycerides weremarkedly lowered in all the animals given nafenopin, which paralleled the increase in livercatalase . All the above effects of nafenopin were fully reversed when the drug was with-drawn from the diet of male rats . During reversal, several microbody nucleoids were seenfree in the hyaloplasm or in the dilated endoplasmic reticulum channels resulting from arapid reduction in microbody matrix proteins after the withdrawal of nafenopin from thediet. Because of microbody proliferation and catalase induction with increasing number ofhypolipidemic compounds, additional studies are necessary to determine the interrelation-ships of microbody proliferation, catalase induction, and hypolipidemia .
INTRODUCTION
Microbodies (peroxisomes), the cytoplasmic con- liver (2), have only recently been recognized asstituents described originally in kidney (1) and ubiquitous structures in animal (3-5) and plant
344
THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974 . pages 344-358
on December 12, 2018jcb.rupress.org Downloaded from http://doi.org/10.1083/jcb.61.2.344Published Online: 1 May, 1974 | Supp Info:

cells (6) . de Duve and associates (7-8) investigatedthe biochemical composition of microbodies ofrat liver and showed that these structures possesscatalase and several oxidative enzymes. It isevident from several studies that catalase is theonly constant component of peroxisomes derivedfrom different sources, whereas the hydrogenperoxide-generating oxidative enzymes that areassociated with catalase differ in number and inspecificity (9) . On the basis of the cytochemicallocalization of catalase with the alkaline 3', 3'-diaminobenzidine medium (10, 11), microbodieshave been identified in tissues other than liverand kidney. Additional studies, however, arenecessary to characterize the composition of thesecatalase-containing structures in tissues otherthan liver and kidney either by biochemical orcytochemical procedures ; and if the structures arefound to possess at least one hydrogen peroxide-producing oxidase, they can then be unequivo-cally regarded as peroxisomes .
Although considerable progress has been madein the understanding of the functional role ofperoxisomes in plant cells, the importance of theseorganelles in cellular metabolism in animal tissuesremains to be elucidated . However, several studiessuggest that microbodies may play a significantbut as yet unexplained role in a variety of func-tions such as in gluconeogenesis, in cholesteroland steroid metabolism, in the detoxification ofhydrogen peroxide, etc . (7, 12, 13) . Studies withethyl-cY p-chlorophenoxyisobutyrate (CPIB) haveclearly established that this compound is valuablein investigating the biological behavior of micro-bodies under different experimental situationsand also suggested a possible relationship betweenmicrobodies and lipid metabolism (13-18) . CPIBis a potent hypolipidemic drug which lowersserum triglycerides and cholesterol in man (19)and in experimental animals (20) . Administrationof this drug to male rats results in a significantincrease in the number of microbody profiles inliver cells (14-16) . The CPIB-induced microbodyproliferation in male rat liver was associated witha marked increase in catalase activity resultingfrom enhanced rate of synthesis of this enzyme(21) . However, when CPIB was administered tofemale rats in doses identical to those given tomale rats for the same duration, there was noappreciable increase either in the number ofmicrobody profiles or in the liver catalase activity .Similar sex differences in microbody proliferativeand catalase response were observed in C 3H miceand in wild type (Csa strain) mice given CPIB,
whereas no sex difference in microbody prolifera-tive response was noted in acatalasemic (Csbstrain) mice (17) .
Recently we have demonstrated that adminis-tration of nafenopin (2-methyl-2[p-(l,2,3,4-tetrahydro-l-nephthyl)phenoxy]-propionic acid ;Su-13437) to male rats, by gavage at 100 mg/kgbody weight daily for 1-2 wk, results in a markedproliferation of hepatic microbody profiles withsimultaneous increase in catalase activity (22) .Nafenopin is structurally related to CPIB and onan equivalent weight basis is approximately fivetimes as potent as CPIB in producing hypo-lipidemia and hepatomegaly (23, 24) . In thepresent investigation varying concentrations ofnafenopin were administered in ground PurinaChow ad lib to both male and female rats, wildtype (Csa strain) mice and acatalasemic (Cs'strain) mice, in order to determine the dose-response relationship and to ascertain the speciesand sex differences, if any, in microbody prolifera-tive response and inducibility of liver catalase .The results indicate an absence of sex differencein microbody proliferative response in nafenopin-treated rats and wild mice, whereas CPIB-inducedmicrobody proliferation occurred only in males(16) . Accordingly, nafenopin can be used as aninducer of microbody proliferation and of catalaseenzyme synthesis in both sexes of rats and mice .These studies also emphasize the significance ofhypolipidemic drugs as effective tools for investi-gating the interrelationship of microbodies withlipid metabolism .
MATERIALS AND METHODS
Animals and Treatment
The inbred rats used in these experiments were ofthe F-344 strain (Simonson Laboratories Inc ., Gilroy,Calif.) weighing between 125 and 175 g, which weremaintained in individual cages . The wild type (Csastrain) mice and the acatalasemic (Csb strain) miceused in these experiments were 2-6 mo old andweighed between 20 and 30 g . Nafenopin (CIBAPharmaceuticals, Summit, N . J .) was added to theground Purina Chow in concentrations of 0.0125%,0.05%, 0.125%, and 0.25% and was thoroughlymixed . Preliminary studies in rats indicated thatnafenopin at 0 .125% and 0.25% dietary levels wasvery effective in inducing microbody proliferation .Accordingly, studies in wild type and acatalasemicmice were conducted with these two dose levels . Atthe end of 1, 2, 3, 4, 6, and 8 wk, liver tissue for mor-phological studies was obtained from two to threeanimals from each of the treated groups after laparot-
REDDY ET AL . Nafenopin-Induced Microbody Proliferation
345

omy or sacrifice . The reversal of nafenopin-inducedmicrobody proliferation was investigated in malerats. After receiving 0.25% nafenopin for 6 wk, ani-mals were killed on days 1, 3, 5, 7, 10, and 14 afterwithdrawal of the drug for ultrastructural studies andcatalase assay.
Morphology
For light microscope examination, samples of liverwere fixed in neutral buffered formaldehyde andembedded in paraffin. 3-5 µm sections were stainedwith hematoxylin and eosin . For electron microscopestudies, tissues were fixed for 1-2 h at 4 °C in 2%osmium tetroxide buffered with S-collidine to pH 7 .4,and processed as described previously (15) . For cyto-chemical localization of catalase, selected samples ofliver from treated and untreated animals were fixedin 2 .5% glutaraldehyde buffered with 0 .1 M sodiumcacodylate, pH 7 .4, for 4 h at 4 °C. After fixation, thetissues were rinsed overnight in 0.1 M cacodylatebuffer containing 0 .2 M sucrose. Then the tissues werefinely chopped with a Sorvall "chopper" (IvanSorvall, Inc ., Newtown, Conn .) and incubated at37 °C for 30-45 min in the 3',3'-diaminobenzidineoxidation medium of Novikoff and Goldfischer (10)and processed for electron microscope examination .
Assay of Liver Catalase Activity
After determining that nafenopin at 0.125% and0.25% caused maximal proliferation of microbodies,in both male and female animals, we limited the bio-chemical studies of catalase to these two dietary levels .The animals were killed by light ether anaesthesiaand the livers were removed and weighed . 5% liverhomogenates were prepared to which sodium deoxy-cholate (DOC) was added to give a final DOC con-centration of 0 .5% (25) . This mixture was incubatedat room temperature for 30 min and then centrifugedin a Spinco model L ultracentrifuge (Beckman Instru-ments, Inc., Spinco Div ., Palo Alto, Calif.) using a50 rotor at 35,000 rpm for 85 min . After centrifuga-tion the lipid-containing layer at the top was carefullyremoved and the supernate was assayed for catalaseactivity at 25 °C by the spectrophotometric methoddescribed by Lück (26) . Total proteins were deter-mined by the method of Lowry et al . (27) .
346
THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974
strain) and acatalasemic (Csb strain) mice, immuno-precipitation of protein reactive with antibody specificfor catalase was undertaken. The anticatalase serumwas obtained by injecting rabbits with catalase puri-fied from the livers of Cs, strain mice. By the Ouch-terlony double diffusion method, this antiserum gavesingle precipitin lines with wild and acatalasemic liverextracts .
The content of liver catalase protein in control andnafenopin-treated wild type male and female micewas determined by the immunotitration methodreferred to above (25) . Quantitation of liver catalaseprotein in acatalasemic mice by the immunotitrationmethod could not be performed, because of low levelsof catalase activity (17, 28) and also because of rapidinactivation of catalase when incubated at 37 °C(29) . In order to compare the amount of catalaseprotein in livers of acatalasemic and wild type mice,equal amounts of liver extracts from normal andnafenopin-treated animals were incubated withquantities of antiserum that were found to neutralizethe catalase protein in normal and nafenopin-treatedwild type mice . The immunoprecipitates were cen-trifuged and washed twice with cold 0 .15 M sodiumchloride . The washed precipitates were dissolved in0.1 M sodium hydroxide and the absorbance at 280nm was measured .
Serum Cholesterol and Triglycerides
All animals were lightly anesthetized with ether .Blood was collected from the inferior vena cava fromnonfasted animals between 9 and 10 a .m. Serum totalcholesterol and/or glyceride glycerol were determinedafter quantitative separation on silicic acid from phos-pholipids by the micromethod described by Azarnoff(30) .
RESULTS
Studies in Rats
MICROBODY PROLIFERATION :
With0.0125% of nafenopin for 2 wk, the liver weightswere only slightly increased . Nafenopin given at0.125% and 0 .25% produced a marked increasein liver weights in both male and female rats(Table I). Light microscope examination of theselivers revealed greatly enlarged hepatocytes con-taining abundant acidophilic, finely granularcytoplasm . During the initial states of nafenopintreatment, numerous mitoses of liver cells wereencountered . Autoradiographic studies, after['H]thymidine pulse, during the first week ofnafenopin administration showed numerous la-beled hepatocyte nuclei in male and female rats .'
Quantitation of Liver Catalase Protein
IN RATS : Content of catalase protein in normaland nafenopin-treated male and female rats wasdetermined by immunotitration method described byGanschow and Schimke (25) .IN WILD AND ACATALASEMIC MICE : In or-
der to determine the content of liver catalase proteinamong control and nafenopin-treated wild type (Cs, 1 Reddy, J ., and Prasad, J . Unpublished observations .

TABLE IEffect of Feeding F-344 Rats with Nafenopin on Liver Weight, Liver Catalase Activity, Serum
Cholesterol, and Serum Glyceride-Glycerol Concentrations
(0 .25%)
* Nafenopin was administered in ground Purina Chow for 4 wk .$ Mean f standard error .
Increase in the number of microbody profileswas evident in both sexes of rats given variousdietary concentrations of nafenopin . I-µm sectionsof Epon-embedded material clearly suggestedthat the increase in microbodies is generalized inthe cytoplasm of liver cells throughout the entireliver lobule. With 0.0125% nafenopin, the micro-body increase was slight when compared to thedegree of proliferation encountered with higherdose levels. The microbody profiles were mostnumerous in liver cells of male and female rats(Fig. I) given 0 .125% nafenopin. With 0.257,nafenopin, there was no additional increase innumber of microbody profiles . The microbodyproliferation in both sexes of rats treated with0.125 ,70 and 0.26 ,70 nafenopin was found to be ofthe same magnitude . Prolonged treatment withnafenopin resulted in the persistence of an in-creased number of peroxisomal profiles in maleand female rats . Conspicuous variations in sizeand shape of microbody profiles were apparent .Nucleoids were observed in many microbody pro-files of different sizes and configurations . Ab-normalities in size and shape of peroxisomal pro-files were striking in animals given 0.0125%nafenopin. Several of these microbody profilescontained striations in the matrix (Fig . 2), similarto those seen in CPIB-treated animals (14). Anumber of microbody profiles in liver cells ofanimals on prolonged nafenopin treatment pos-sessed a somewhat flocculent matrix . However,
all these organelles stained positively for catalasewhen incubated in the alkaline 3',3'-diamino-benzidine medium . This reaction was inhibitedwhen tissues were incubated in the presence ofaminotriazole .
In addition to showing microbody proliferation,the liver cells of nafenopin-treated animals re-vealed marked proliferation of smooth endoplas-mic reticulum. Isolated channels of rough endo-plasmic reticulum were also frequently encoun-tered. Mitochondria appeared normal in size,shape, and number. They did not show any in-clusions or other obvious structural abnormality(Fig . 1) . There was no increase in the number oflysosomes or lipid droplets in nafenopin-treatedlivers. Many hepatocyte nucleoli were hyper-trophied .MICROBODY REVERSAL : Rats treated with
nafenopin for 6 wk showed increase in the numberof microbody profiles to the same extent as evi-denced at 4 and 8 wk . Gradual depletion in thenumber of microbody profiles was noted duringthe first 7 days after withdrawal of nafenopin . By14 days after the drug was discontinued, the num-ber of microbody profiles returned to normal . Thereduction in microbody profiles was imperceptiblein that there was no appreciable evidence of cellu-lar autophagy. Detailed examination of liversduring the first week of nafenopin withdrawalrevealed certain interesting morphological fea-tures (Figs . 3-7). Marked decrease in electron
REDDY ET AL. Nafenopin-Induced Microbody Proliferation
347
Group*No . ofanimals Liver weight $
Liver catalaseactivity $
Serumcholesterol I
Serum glyceride-glycerol$
g/100 g body weight U/mg protein mg/100 ml mg/100 mlMale rats
Controls 6 3 .75 f 0.42 38.0 f 3 .9 82 f 5 10.6 f 2 .8Nafenopin 5 8 .09 f 0 .52 74.9 f 5 .1 86 f 9 2 .9 f 0 .6
(0 .125%)Nafenopin 6 8 .34 f 0.48 86.1 f 4.3 79 f 8 1 .6 f 0.2
(0 .25%)Female rats
Controls 6 3 .21 f 0.36 31 .2 f 3 .7 76 f 4 9.7 f 2 .4Nafenopin 4 7 .95 f 0.18 71 .4 f 4.2 66 f 4 2.5 f 0.3
(0 .125%)Nafenopin 5 8.10 f 0.04 82 .7 f 1 .25 74 f 6 2 .3 f 0 .5

FiauRE 1 Female rat liver . Nafenopin 0.125% in the diet for 4 wk. The microbody proliferation infemale rat liver cells was comparable to that observed in male rats . Significant variation in size andshape of microbody profiles (mb) is evident. Nucleoids (n) . X 14,500 .
3 48

opacity of the matrix was noted in some micro-body profiles. Several dilated endoplasmic reticu-lum channels contained crystalline microbodynucleoids or cores (Figs. 3 and 4) without thebackground matrix proteins, suggesting a gradualdepletion of matrix proteins presumably due tocessation of synthesis after nafenopin withdrawal .Extrusion of nucleoids into hyaloplasm was alsoseen. At 7 days after withdrawal of nafenopin,occasional examples of nucleoids lying free in thehyaloplasm were also encountered (Figs. 6 and 7) .LIVER CATALASE : The results in Table I
indicate that the activity of catalase in the liverin both male and female rats increased about two-fold on administration of 0 .125% or 0.25% nafeno-pin. When liver catalase activities were determinedin male rats treated with 0.25% nafenopin for2-14 days, it was found that the activity of cata-lase increases progressively, reaching the steadystate level in 8-12 days . Nafenopin appeared aseffective as CPIB in increasing the liver catalaseactivity . It is of interest to note that the inducedsteady state of catalase activity with these twodrugs is nearly the same, i.e. approximately twicethat of the normal steady state .
The results of immunotitration of liver catalasein male and female rats treated with 0 .25%nafenopin for 4 wk are shown in Fig . 8. Theamount of anticatalase serum needed to precipi-tate completely the catalase activity from 1 ml of5% liver extract of either male or female rattreated with nafenopin was two times greaterthan that required for extracts from livers of un-treated controls . These results indicate that theamount of catalase protein in liver extracts ofmale and female rats given nafenopin was thesame and that nafenopin causes a twofold increasein the amount of liver catalase protein when com-pared to controls .
By 10 days after withdrawal of nafenopin fromthe diet, the catalase activity in liver reverted tonormal values which paralleled the reduction inthe number of hepatic microbody profiles .SERUM CHOLESTEROL AND SERUM GLYC-
ERIDE LEVELS : Administration of nafenopinat 0.125% and 0 .25% to both male and femalerats produced no significant cholesterol-loweringeffect at 4 wk . In direct contrast, however, a pro-found reduction in serum glyceride levels was ob-tained in both males and females (Table I) .Within 7-10 days after withdrawal of the drugthe serum glyceride levels returned to normal,
which paralleled the reduction in catalase activityand the number of microbody profiles .
Studies in Mice
MICROBODY PROLIFERATION : Nafenopinwas given to both sexes of wild type (Csa strain)mice and acatalasemic (Csb strain) mice at dietarylevels of 0.125 0/0 and 0.25%. The gain in bodyweight was not affected at 0.125 % dose level,whereas at 0 .25% level there was some reductionin weight gain. However, at both these levels,nafenopin produced a marked increase in theweight of the liver in both males and females .In wild type mice fed 0.125% nafenopin for 4-8wk the liver weight was approximately 25% ofthe body weight (Table II) . Similar increases inliver weight were also seen in both sexes of acatala-semic mice treated with nafenopin . Examinationof these livers with the light microscope showedlarge polygonal liver cells with abundant cyto-plasm. Several mitotic figures were seen in thelivers of animals killed during the first 10 days ofnafenopin administration, suggesting that nafeno-pin-induced increases in liver weight are due inpart to cell multiplication and cellular hyper-trophy .
The microbody proliferation was most pro-nounced in the livers of both sexes of wild typeand acatalasemic mice given 0 .125% nafenopin(Fig. 9) . Further increase in the number of micro-body profiles was not seen with the higher doselevel. In previous studies, we did not observe anyincrease in the number of microbody profiles inthe female wild type (Csa strain) mice treatedwith CPIB, although CPIB produced a marked in-crease in hepatic microbody population in bothsexes of acatalasemic (Csb strain) mice (17) . Theabsence of sex difference in microbody prolifera-tion in response to nafenopin, in rats and mice,is of particular interest .The microbody profiles were numerous and
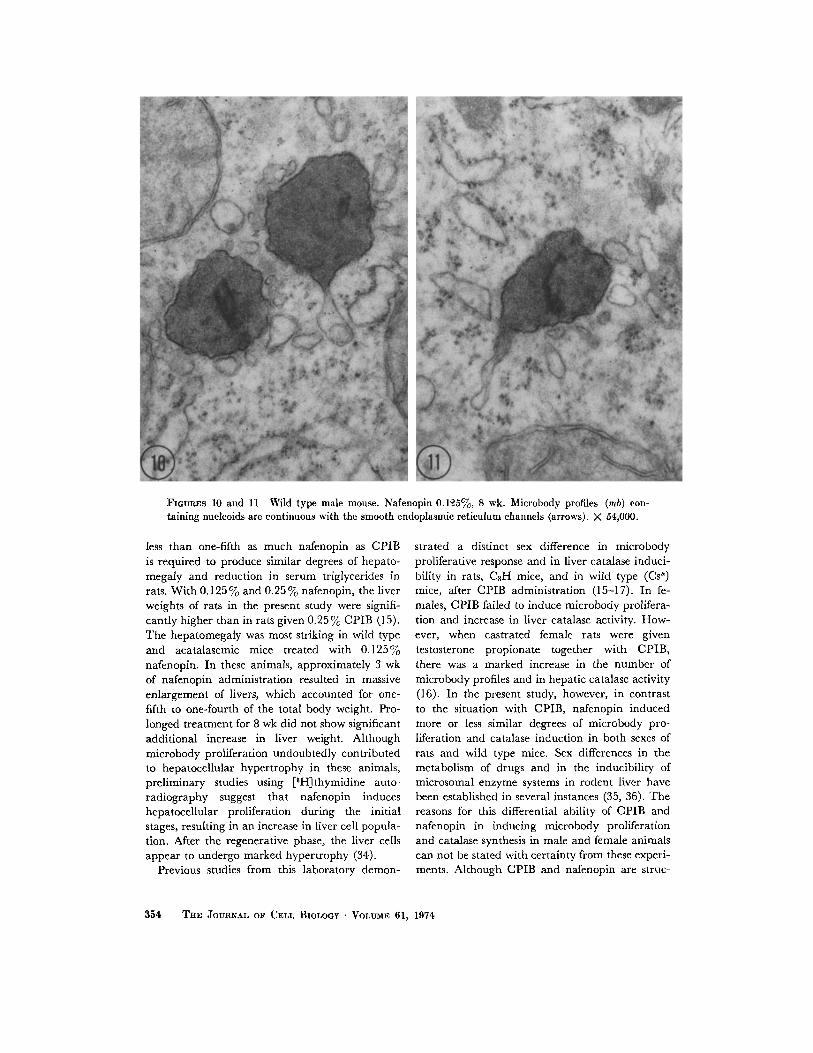
showed a great deal of variation in size andshape. Continuities between adjacent microbodyprofiles and between microbody profiles andsmooth endoplasmic reticulum channels werenoted frequently (Figs . 10 and 11) . Structuralchanges in mitochondria were not seen, andbecause of marked increase in microbody profiles,the mitochondria appeared sparse. Proliferationof smooth endoplasmic reticulum was observedin the liver cells of all animals treated with nafeno-pin .
REDDY ET AL . Nafenopin-Induced Microbody Proliferation
349

350
THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974

LIVER CATALASE : The results of the deter-mination of catalase activity in livers of acatala-semic and wild type mice are shown in Table II .After nafenopin treatment, the catalase activityin male and female wild type mice (Csa strain) wassignificantly increased . The amount of catalaseprotein in liver of untreated and treated Csa micewas determined by the immunotitration method .
a06
0.1
0.14
a1e
0.22
0.26
0.3ml OF ANTICATALASE SERUM ADDED TO
I m l OF LIVER EXTRACT
FIGURE 8 Quantitation of liver catalase by immuno-precipitation in male and female rats treated with0.25% nafenopin for 4 wk . Increasing quantities ofanticatalase serum were added to 1-ml samples ofdeoxycholate-treated liver extracts from normalmales (A-A), normal females (o-o), nafenopin-treated males (A-A) and nafenopin-treated females(0--0) . Dashed lines are extrapolations of the linearportions of the titration curves to zero catalase ac-tivity .
0.02
For complete precipitation of the catalase activityfrom I ml of 2 .5% liver extract of Cs' mice treatedwith nafenopin for 8 wk, approximately 0 .64 mlof anticatalase serum was required, whereas only0.31 ml antiserum was required to precipitate thecatalase activity from I ml of 2 .5% liver extractsfrom untreated wild mice . From these findings itcan be reasonably assumed that a twofold increasein concentration of catalase protein occurs inlivers of Csa mice treated with nafenopin .
Acatalasemic (Csb) mice treated with nafenopinshowed only a minimal increase in liver catalaseactivity (Table II), although the microbodypopulation in liver cells was comparable to thatencountered in male and female wild type (Csa)mice. Studies of Aebi et al . (29) demonstratedthat acatalasemic mutant mice have an unstablecatalase that is unusually heat labile . The catalasein the present studies was assayed on liver extractstreated earlier with DOC and kept at room tem-perature for 30 min (see Materials and Methods) .Accordingly, the low levels of catalase in nafeno-pin-treated Csb mice may be, in part, due to in-activation at room temperature.
To determine the quantity of catalase proteinin acatalasemic mice, 0 .31 ml and 0.64 ml ofanticatalase serum were added, respectively, to 1ml samples of 2 .5% liver extracts from controland nafenopin-treated animals . The amount ofanticatalase serum used was the same that wasfound to precipitate all the catalase activity inwild type mice by the immunotitration method(see above) . The amount of protein precipitatedfrom nafenopin-treated, acatalasemic mouseliver extracts was approximately twice that pre-cipitated from control mouse liver extracts . Fur-thermore, the amounts of protein precipitated
FIGURE 2 Male rat treated with 0.0125% nafenopin for 2 wk. Several microbody profiles containedstriations in the matrix (arrows) . Nucleoid (n) . X 65,000 .
FIGURES 3-7 Figures 3-7 are from the livers of male rats killed between 2 and 10 days after withdrawalof nafenopin after they have been on the drug for 6 wk .
FIGURES 3-5 Microbody nucleoids (n) are seen within the irregular endoplasmic reticulum channelswithin 5-7 days after discontinuation of nafenopin . Electron-opaque microbody proteins are not seenin these membranes . In Fig . 3, the dumb-bell-shaped channel (arrow) contains a residue of electron-opaque material similar to that present in the channel containing nucleoid, possibly suggesting that thematrix proteins have dried up after withdrawal of nafenopin . Figs . 4 and 5 . X 71,000 . Fig. 3. X 49,000 .
FIGURES 6 and 7 Nucleoids (n) are seen in the hyaloplasm . By 5-7 days after removal of nafenopinfrom the diet, some microbody nucleoids were seen lying free in cytoplasm after dissociating from micro-body profiles. X 52,000 .
REDDY ET AL . Nafenopin-Induced Microbody Proliferation 351

352
TABLE II
Effect of Feeding Wild Type (Cs" Strain) Mice and Acatalasemic (Csb Strain) Mice withNafenopin on Liver Weight, Liver Catalase Activity, and Serum
Glyceride-Glycerol Levels .
* Nafenopin (0 .125%) was given in ground Purina Chow for 8 wk .
from the liver extracts of acatalasemic and wildtype mice were also comparable . From theseresults it can be assumed that nafenopin causesapproximately a twofold increase in the quantityof liver catalase protein in acatalasemic mice .SERUM GLYCERIDE LEVELS :
The resultslisted in Table II indicate that serum glyceride-glycerol levels are considerably lower in theacatalasemic mice than in the wild type mice .These findings in untreated controls are in agree-ment with the observations of Goldfischer et al .(31) . After treatment with 0 .1250/, nafenopin for8 wk, the serum glyceride levels in both sexes ofacatalasemic and wild type mice were markedlylowered (Table II) . Serum cholesterol levels ofthese mice were not determined in this study .
DISCUSSION
Unlike other cellular constituents, microbodies(peroxisomes) in hepatic parenchymal cells areinfrequently altered in number or morphology byexperimental manipulations . However, markedlyreduced activities of catalase and/or other perox-isomal enzymes in the liver have been observed incertain conditions, particularly in tumor-bearinganimals (32) . Quantitative studies to date indicatethat CPIB and acetylsalicylic acid are the onlyknown compounds that are capable of inducingsignificant increase in the number of microbodyprofiles in male rat liver cells and that the micro-body proliferative response induced by CPIB ismore pronounced in comparison to that inducedby acetylsalicylic acid (14, 15, 33) . Concomitant
THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974
with the increase in microbody profiles, the livercatalase activity is increased twofold after CPIBtreatment due to enhanced synthesis of thisenzyme (21) . Additional studies by Svoboda,Azarnoff, and Reddy (16) demonstrated that inrats, CPIB-induced microbody proliferation andrise in catalase levels are dependent, to a signifi-cant degree, upon male sex hormones.
In the present study, microbody proliferationin both male and female rats treated with higherdoses of nafenopin (0 .125% and 0.25%) was verypronounced . Marked increase in the number ofmicrobody profiles was also observed in both sexesof wild type and acatalasemic mice given nafeno-pin. The microbody proliferation induced with0.125% nafenopin in these animals was of thesame degree as that induced by 0.25% CPIB(14, 15, 17) . In addition, these studies have alsoshown a twofold increase in the hepatic catalaseactivity and in the amount of catalase proteinin both sexes of rats and wild type mice treatedwith nafenopin . Although the increase in catalaseactivity was not significant in acatalasemic micegiven nafenopin, the amount of liver catalaseprotein in these animals was approximately equalto that found in nafenopin-treated wild type mice .
Nafenopin, like CPIB, is a phenolic ether whichproduces in rats a significant reduction in serumtriglycerides, but does not significantly reduce theserum cholesterol (24) . Serum glycerol-glycerideswere significantly lowered in rats and mice givennafenopin in the present investigation . The studiesof Best and Duncan (24) indicate that somewhat
Group*No. ofanimals Liver weight
Liver catalaseactivity
Serum glyceride-glycerol
g1100 g body weight U/mg protein mg/100 ml
Wild type (Cs) miceMales control 6 5 .3 f 0 .30 38 .8 f 3 .1 11 .6 t 1 .8Males nafenopin 4 24 .6 t 2 .80 76 .3 t 5 .6 4 .2 t 0.4Females control 5 4 .9 t 0 .26 37 .2 t 4 .3 10.5 t 1 .3Females nafenopin 4 26 .5 f 1 .70 71
f 5 .4 4.1 t 0.6Acatalasemic (C S b) MiceMales control 6 5 .6 t 0 .25 10 .5 ± 3 .6 5 .7 t 1 .7Males nafenopin 4 18 .7 t 1 .90 15.2 t 2 .7 1 .9 t 0 .6Females control 4 5 .0 f 0 .40 11 .3 t 3 .2 5 .3 f 2 .2Females nafenopin 4 19 .8 f 2 .75 13 .4 f 2 .8 2 .1 f 0 .8

FIGURE 9 Female wild type mouse (Cs' strain) . Nafenopin 0.125% for 4 wk . Marked increase in numberof microbody profiles (mb) is seen . The microbody proliferation in male Cs' mice treated with nafenopinwas of similar magnitude . X 19,000 .
353
less than one-fifth as much nafenopin as CPIBis required to produce similar degrees of hepato-megaly and reduction in serum triglycerides inrats . With 0 .125 % and 0.25 % nafenopin, the liverweights of rats in the present study were signifi-cantly higher than in rats given 0 .25% CPIB (15) .The hepatomegaly was most striking in wild typeand acatalasemic mice treated with 0.125%nafenopin. In these animals, approximately 3 wkof nafenopin administration resulted in massiveenlargement of livers, which accounted for one-fifth to one-fourth of the total body weight. Pro-longed treatment for 8 wk did not show significantadditional increase in liver weight . Althoughmicrobody proliferation undoubtedly contributedto hepatocellular hypertrophy in these animals,preliminary studies using [ 5H]thymidine auto-radiography suggest that nafenopin induceshepatocellular proliferation during the initialstages, resulting in an increase in liver cell popula-tion, After the regenerative phase, the liver cellsappear to undergo marked hypertrophy (34) .
Previous studies from this laboratory demon-
35 4
FIGURES 10 and 11 Wild type male mouse. Nafenopin 0 .125%, 8 wk . Microbody profiles (mb) con-taining nucleoids are continuous with the smooth endoplasmic reticulum channels (arrows) . X 54,000 .
THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974
strated a distinct sex difference in microbodyproliferative response and in liver catalase induci-bility in rats, C3H mice, and in wild type (Cs")mice, after CPIB administration (15-17) . In fe-males, CPIB failed to induce microbody prolifera-tion and increase in liver catalase activity. How-ever, when castrated female rats were giventestosterone propionate together with CPIB,there was a marked increase in the number ofmicrobody profiles and in hepatic catalase activity(16) . In the present study, however, in contrastto the situation with CPIB, nafenopin inducedmore or less similar degrees of microbody pro-liferation and catalase induction in both sexes ofrats and wild type mice. Sex differences in themetabolism of drugs and in the inducibility ofmicrosomal enzyme systems in rodent liver havebeen established in several instances (35, 36) . Thereasons for this differential ability of CPIB andnafenopin in inducing microbody proliferationand catalase synthesis in male and female animalscan not be stated with certainty from these experi-ments. Although CPIB and nafenopin are struc-

turally similar hypolipidemic phenolic ethers,there may exist differences in their metabolism .It is possible that the microbody proliferativeeffect of CPIB is dependent upon excessive ana-bolic activity, whereas nafenopin may be lessdependent, or may possess androgenic activity.The absence of sex difference in response to nafeno-pin-induced microbody proliferation and catalasesynthesis may be due to the fact that this com-pound is about five times more potent than clofi-brate (CPIB) in producing hypotriglyceridemia(23, 24). Further clarification would probably beachieved by evaluation of the dose in relation toreduction of plasma cholesterol and triglyceridelevels . It is not known if CPIB, in concentrationsgreater than 0 .25% in diet, would cause micro-body proliferation in female rats and mice . Thismay not occur in view of the finding that theeffect on lipids actually decreases when CPIB isgiven in doses greater than 0 .25%. Despite theuncertainty in understanding the basic differ-ence(s) in the action of these drugs in inducingmicrobody proliferation in one or both sexes ofrats and mice, the present study clearly identifiesnafenopin as an effective inducer of microbodyproliferation and of catalase synthesis in bothsexes.
In contrast to a sex difference in response toCPIB in rats and mice cited above, no suchdifference was observed in acatalasemic "mu-tant" mice (17) . Both male and female acatala-semic mice given CPIB showed profound increasein number of hepatic microbody profiles . Thismay be in part due to the genetic makeup of thismutant mouse strain, because, within a givenspecies, genetic variation may be important indetermining the occurrence or magnitude of en-zyme induction (35) . Microbody proliferativeresponse in both sexes of acatalasemic mice afternafenopin was identical to that obtained afterthe administration of CPIB .
The increase in catalase activity and in theamount of catalase protein in liver of rats treatedeither with nafenopin or with CPIB was twicethat of controls . It is of interest to note that theinduced steady state of liver catalase in rats isnever more than twofold . Furthermore, even inanimals given diets containing both nafenopinand CPIB, the concentration of liver catalase didnot exceed this level.' The amount of liver catalaseprotein in these animals, therefore, does not
2 Reddy, J. Unpublished observations .
correspond with the severalfold increase in thenumber of microbody profiles. Although, cyto-chemically, catalase was demonstrable in all themicrobody profiles, the possibility that thesestructures proliferating under the influence ofnafenopin or CPIB (37) possess a low concentra-tion of catalase cannot be excluded . This is notsurprising in view of our earlier observation thatCPIB induces hepatic microbody proliferation inanimals treated with allylisopropylacetamide, adrug which completely inhibits the catalasesynthesis (18) .
The relationship between the increase in thenumber of hepatic microbody profiles and decreasein serum cholesterol and/or triglycerides resultingfrom the administration of CPIB and nafenopin,is not clear. Despite numerous investigations, themechanism by which CPIB elicits the hypolipi-demic and microbody proliferative responsescannot be stated with certainty. Since nafenopinis a relatively new compound in the expandingfield of hypolipidemic agents, studies dealing withthe mechanism of its action are lacking, becauseit was withdrawn from clinical studies after itwas found to produce liver tumors in mice .' Re-duction in serum glycerides and induction ofhepatic microbody proliferation and of catalaseappear to be the common properties of these twodrugs. Because of the significant increase in micro-body population and in catalase activity resultingfrom CPIB, Svoboda and Azarnoff (14) invokeda possible relationship between CPIB-inducedmicrobody proliferation and hypolipidemia . Asimilar relationship between microbodies andlipid metabolism was suggested earlier by Novi-koff and Shin (12) on the basis of the observationof Hochstein and Ernster (38) that the rat livermicrosomes catalyze lipid peroxidation that canlead to the production of hydrogen peroxide .Administration of bovine hepatic catalase has
been shown to induce a hypocholesterolemicresponse in humans and rabbits (39, 40) . Morerecently, Goldfischer et al . (31) showed that inacatalasemic mouse mutants the serum cholesteroland triglycerides are significantly lower, suggest-ing that in vivo peroxidase is responsible for hypo-lipidemia. A definite relationship between in vivoperoxidase and hypolipidemia in acatalasemicmice appears doubtful because of the recent
'Wagner, W. E. CIBA Pharmaceuticals, personalcommunication .
REDDY ET AL . Nafenopin-Induced Microbody Proliferation 355

demonstration of an abnormality of hepatic lipo-genesis in this mutant strain (41) .
In view of the reports dealing with the effect ofcatalase on cholesterol, it appeared reasonable toassume that the hypolipidemic effect of CPIB wasthe result of endogenous elevation of liver catalaseproduced by microbody proliferation (14) . How-ever, accumulated evidence indicates that thehypolipidemic and microbody proliferative effectsof CPIB are possibly independent (15, 16, 18,and 42). In the present study, the demonstrationof microbody proliferation and catalase inductionafter administration of nafenopin, a potent hypo-lipidemic drug, again raises the question of possiblerelationship between microbody proliferation andhypolipidemia . In this regard, it is also pertinentto state that methyl clofenapate (43), a hypo-Epidemic analogue of CPIB, also causes a verysignificant hepatic microbody proliferation . Onan equivalent weight basis, methyl clofenapateappears to be 5-10 times as potent as nafenopinor CPIB in inducing microbody proliferation .These drugs promise to be of considerable valuefor studying the functional significance of micro-body proliferation in the production of hypo-chlesterolemia and hypotriglyceridemia. Further-more, acetylsalicylic acid which induces micro-body proliferation is also known to reduce serumcholesterol levels (42) . It is, therefore, necessaryto investigate the structural analogues of CPIB inan attempt to determine the interrelationships ofchemical configuration of these drugs, microbodyproliferation, catalase induction, and hypolipi-demia .
Although the role of drug-induced hepaticmicrobody proliferation is not understood, thewidespread occurrence of microbodies in severalcell types of vertebrates has given rise to addi-tional speculation regarding their functionalsignificance and has been the subject of discussionin a recent symposium . 4 In view of their wide-spread distribution among tissues and species,the possibility that microbodies may performdifferent functions in different cell types cannotbe excluded, although their role in lipid metabo-lism and in detoxification of H202 may be con-sidered universal (13) .
The observations on microbody profiles during
4 The symposium on "peroxisomes" organized byThe Histochemical Society on April 14, 1973, at its24th annual meeting held in Atlantic City, N . J .
356 THE JOURNAL OF CELL BIOLOGY • VOLUME 61, 1974
recovery after withdrawal of nafenopin weresimilar to those seen in rats after CPIB (44) . DI.-
solution of the microbody matrix, appearance ofnucleoid cores in dilated endoplasmic reticulummembranes or occasionally in the hyaloplasm,are the significant findings. Incorporation ofmicrobody profiles into lysosomes or into auto-phagocytic vacuoles was not detected. It appearsthat cessation of enhanced synthesis of peroxisomalproteins after discontinuation of nafenopin isresponsible for the reduction in the peroxisomalprotein pool. The reduction in number of micro-body profiles appears to be due to progressivelydiminishing quantities of peroxisomal matrixproteins in endoplasmic reticulum channels .
Nafenopin was supplied by Dr. William E . Wagner,CIBA Pharmaceuticals, Summit, N . J . Technicalassistance was given by Ms . Lynne Schmutz, Ms .Faye Brady, Ms. Dianne Knox, Ms . Mary AnnWickham, and Mr . Charles Sittler . Mrs. Ella M .Olson provided the editorial assistance.
This work was supported by United States PublicHealth Service grants GM-15956 and CA-5680 .Received for publication 16 August 1973, and in revisedform 3 December 1973.
REFERENCES
I . RHODIN, J. 1954. Correlation of ultrastructuralorganization and function is normal and experi-
mentally changed proximal convoluted tubulecells of the mouse kidney . Doctorate Thesis .Karolinska Institutet, Stockholm .
2 . ROUILLER, C., and W . BERNHARD . 1956 . "Micro-bodies" and the problem of mitochondrialregeneration in liver cells . J. Biophys. Biochem .Cytol . 2 (Suppl .) :355 .
3. HRUBAN, Z ., and M. RECHCIGL, JR . 1969 . Micro-bodies and related particles. Morphology, bio-chemistry and physiology . Int . Rev . Cytol.1(Suppl) :20.
4. HRUBAN, Z ., E . VIGIL, A. SLESERS, and E. HoF-
KINS. 1972 . Microbodies . Constituent organellesof animal cells. Lab. Invest. 27 :184 .
5. NoviKOFF, A . B ., P. M. NOVIKOFF, C . DAMS,and N. QUINTANA. 1973. Studies on micro-peroxisomes V . Are microperoxisomes ubiqui-tous in mammalian cells? J. Histochem . Cyto-chem . 21:737.
6. TOLBERT, N. 1971. Microbodies-peroxisomesand glyoxysomes . Annu . Rev. Plant Physiol.22:45 .
7 . DE DuvE, C ., and P. BAUDHUIN . 1966 . Peroxi-

somes (microbodies and related particles) .Physiol. Rev. 46 :323 .
LEIGHTON, F ., B. POOLE, P. LAZAROW, and C .DE DuvE. 1969 . The synthesis and turnover ofrat liver peroxisomes . I. Fractionation ofperoxisome proteins . J. Cell Biol. 41 :521 .
DE DuvE, C. 1969 . Evolution of thyperoxisome .Ann. N. Y. Acad. Sci. 168 :369 .
NOVIKOFF, A. B., and S . GOLDFISCHER . 1968 .Visualization of microbodies for light and elec-tron microscopy . J. Histochem. Cytochem . 16 :507 .
FAHIMI, H . D . 1969 . Cytochemical localization ofperoxidatic activity of catalase in rat hepaticmicrobodies . J. Cell Biol . 43 :275 .
NOVIKOFF, A . B., and W. Y. SHIN . 1964. Theendoplasmic reticulum in the Golgi zone andits relations to microbodies, Golgi apparatus,and autophagic vacuoles in rat liver cells . J.Microsc . (Paris) . 3 :187 .
Microbodies in experimentally altered cells .II . The relationship of microbody proliferationto endocrine glands . J. Cell Biol . 40 :734 .
REDDY, J ., S . BUNYARATVEJ, and D. SVOBODA .1969 . Microbodies in experimentally alteredcells . IV . Acatalasemic (CSb) mice treated withCPIB . J. Cell Biol . 42:587 .
REDDY, J ., M. CHIGA, S . BUNYARATVEJ, and D .SVOBODA . 1970 . Microbodies in experimentallyaltered cells. VII . CPIB-induced hepaticmicrobody proliferation in the absence ofsignificant catalase synthesis . J. Cell Biol. 44 :226-234.
19 . OLIVER, M. 1963 . Further observations on theeffects of atromid and of ethyl-chlorophenoxy-isobutyrate on serum lipid levels . J. Atheroscler .Res. 3:427 .
20 . BEST, M., and C. DUNCAN . 1964. Hypolipemiaand hepatomegaly from ethyl chlorophenoxy-isobutyrate (CPIB) in the rat . J. Lab . Clin .Med. 64:634 .
21 . REDDY, J ., M. CHIGA, and D. SVOBODA. 1971 .Stimulation of liver catalase synthesis in rats byethyl p-chlorophenoxyisobutyrate . Biochem . Bio-phys . Res . Commun . 43 :318 .
22 . REDDY, J ., D . SVOBODA, and D. AZARNOFF. 1973 .
8 .
9 .
10 .
11 .
12 .
17 .
18.
Microbody proliferation in liver induced bynafenopin, a new hypolipidemic drug : com-parison with CPIB . Biochem. Biophys. Res .Comm. 52 :537-543 .
23 . HESS, R., and W. L. BENCZE . 1968 . Hypolipi-demic properties of a new tetralin derivative(CIBA 13,437-SU) . Experientia (Basel) . 24 :418 .
24 . BEST, M. M., and C . H. DUNCAN . 1970. Lipideffects of a phenolic ether (SU-13437) in therat: Comparison with CPIB . Atherosclerosis.12:185 .
25 . GANSCHOW, R . E., and R . T. SCHIMKE . 1969. In-dependent genetic control of the catalytic ac-tivity and the rate of degradation of catalase inmice . J. Biol. Chem . 244 :4649.
26 . LUCK, H . 1965 . Catalase, Methods of EnzymaticAnalysis. H. H . U. Bergmeyer, editor . Aca-demic Press, Inc ., New York . 885.
27 . LOWRY, O. H ., N . J . ROSEBROUGH, A. L. FARR,
30 . AZARNOFF, D. L . 1962 . Micromethod for thedetermination of serum lipids . J. Lab . Clin .Med. 60 :631 .
31 . GOLDFISCHER, S., P . S. ROHEIM, D . EDELSTEIN,and E. ESSNER . 1971 . Hypolipidemia in amutant strain of acatalasemic mice . Science(Wash. D . C.) . 173 :65 .
32 . KAMPSCHMIDT, R. F . 1965 . Mechanism of livercatalase depression in tumor-bearing animals :a review . Cancer Res . 25 :34 .
33 . HRUBAN, Z., H. SWIFT, and A. SLESERS . 1966 .Ultrastructural alterations of hepatic micro-bodies . Lab . Invest . 15:1884 .
34. BECKETT, R. B., R . WEISS, R. E. STITZEL, andR . J . CENEDELLA . 1972 . Studies on the hepato-megaly caused by the hypolipidemic drugsnafenopin and clofibrate . Toxicol. Appl . Pharma-col . 24 :42 .
35 . CONNEY, A. H . 1967 . Pharmacological implica-tions of microsomal enzyme induction . Pharma-col . Rev . 19 :317 .
36. KINOSHITA, F . K ., and K . P . DUBOIS . 1970 . Induc-tion of hepatic microsomal enzymes by Herban,Diuron and other substituted herbicides .Toxicol. Appl . Pharmacol. 17:406 .
37 . REDDY, J ., S . BUNYARATVEJ, and D . SVOBODA .
REDDY ET AL. Nafenopin-Induced Microbody Proliferation
357
13 . REDDY, J . K . 1973 . Possible properties of micro-bodies (peroxisomes) : Microbody proliferationand hypolipidemic drugs. J. Histochem . Cyto-
and R . J . RANDALL . 1951 . Protein measure-ment with the Folin phenol reagent. J. Biol .Chem . 193 :265 .
chem . 21 :967 . 28 . FEINSTEIN, R., J . BRAUN, and J . HOWARD . 1967 .14. SVOBODA, D . J ., and D. L. AZARNOFF . 1966 . Acatalasemic and hypocatalasemic mouse
mutants. II. Mutational variations in bloodand solid tissue catalases . Arch . Biochem . Biophys .
Response of hepatic microbodies to a hy-polipidemic agent, ethyl chlorophenoxyiso-butyrate (CPIB) . J. Cell Biol. 30 :442 . 120 :165 .
15. SVOBODA, D., H. GRADY, and D . AZARNOFF .1967 . Microbodies in experimentally alteredcells . J. Cell Biol. 35 :127 .
29 . AEBI, H., H. SUTER, and R. N . FEINSTEIN . 1968 .Activity and stability of catalase in blood andtissues of normal and acatalasemic mice . Bio-
16. SVOBODA, D., D. AZARNOFF, and J . REDDY . 1969 . chem . Genet . 2 :245 .

1969. Microbodies in experimentally alteredcells . V . Histochemical and cytochemicalstudies on the livers of rats and acatalasemicmice treated with CPIB. Am. J. Pathol . 56 :351 .
38. HOCHSTEIN, P., and L. ERNSTER . 1963 . ADP-activated lipid peroxidation coupled to theTPNH oxidase system of microsomes . Biochem .Biophys. Res. Commun . 12 :388 .
39. BARCELO, P., P . PUIG-MUSET, L . SANS SOLA, J .LAPORTE, and F. G . VALDECASAS . 1960 . Pre-mières études pharmacologiques sur l'hepato-catalase et son emploi thérapeutique . Eus.Symp. Med. Enzymol. Proc . Mil . 342 .
40. CARAVACA, J., E. G. DIMOND, S. C . SOMMERS,and R. WENS. 1967 . Prevention of inducedatherosclerosis by peroxidase . Science (Wash .D. C.) . 155:1284 .
3 58
THE JOURNAL OF CELL BIOLOGY . VOLUME 61, 1974
41 . CUADRADO, R. R., and L. A. BRICKER . 1973. Anabnormality of hepatic lipogenesis in a mutantstrain of acatalasemic mice. Biochim . Biophys.Acta 306:168 .
42 . AZARNOFF, D., and D. SvoBoDA . 1969. Micro-bodies in experimentally altered c ells. VI .Thyroxine displacement from plasma proteinsand clofibrate effect . Arch. Int. Pharmacodyn.Ther . 181 :386 .
43 . REDDY, J. K. 1974 . Hepatic microbody prolifera-
tion and catalase synthesis induced by methylclofenapate, a hypolipidemic analogue of
CPIB . Am. J. Pathol. In press .44 . SVOBODA, D., andJ . REDDY . 1972 . Microbodies in
experimentally altered cells . IX . The fate of
microbodies . Am. J. Pathol. 67 :541 .