naosite: nagasaki university's academic output...
TRANSCRIPT

This document is downloaded at: 2020-08-02T13:46:11Z
Title Larval Metamorphosis of the Sea Urchins, Pseudocentrotus depressus andAnthocidaris crassispina Induced by Microbial Films
Author(s) Siti Akmar Khadijah binte Ab Rahim
Citation (2004-03-31)
Issue Date 2004-03-31
URL http://hdl.handle.net/10069/20898
Right
NAOSITE: Nagasaki University's Academic Output SITE
http://naosite.lb.nagasaki-u.ac.jp

LarvalMetamorphosisoftheSeaUrchins,
鳶鰯・・8伽伽s吻・θ5s麗5and14励・・∫伽s徽5s棚nα
InducedbyMiαobialFilms
December,、2003
G㎜U紐ESCHOOLOFSCIENCE州pTECHNOLOGY
NAGASAKI UNIVERSITY
JAPAN
SITIAKMARKHADIJAH箪INTIABRAHIM

Larval Metamorphosis of the Sea Urchins,
Pseudocentrotus depressus and Anthocidaris crassispina
Induced by Microbial Films
A dissertation submitted in partial fulfillment of the requirement
for the degree of
Doctor of Philosophy in Marine Science
by
SITI A~MAR KHADIJ~AH~~ BINTl AB ~HIM
Approved by the Research Advisory Committee
Prof. Hitoshi Kitamura
Course Supervisor & Chairperson
Prof. Yuji Fujita prof. Kenji Hara
December, 2003
Graduate School of Science and Technology
Nagasaki University, Japan

ACKNOWLED GEMENTS
I would like to express my sincere thanks and gratitude for help received
during this work to . . .
' My honorable research supervisor Prof. Hitoshi Kitamura, for his
able guidance and constructive criticisms during the course of this
mvestrgation.
e Ministry of Education, Science, Sports and Culture of Japan
(MONBUSHO), for awarding the scholarship.
' Prof. ~Y~uji Fujita and Prof. Kenji Hara, for their valuable advice and
suggestions as referees for doctoral dissertation.
' Dr. Cyril Glenn Satuito, for his kind ~elp and scholastic advice in
writing up this thesis.
' Prof. Toru Takita, for his kindness to introduce me with my research
supervrsor.
e Universiti Malaysia Sarawak (UNIMAS), fbr granting me the study
leave to conduct the present research.
' My lab-mates: Mrs. Lee, Mr. Fujikawa, Mr. Jouuchi, Mr. Shiraishi,
Mr. Taniguchi, Miss Kouda, Miss Yokoyama and other students of
Laboratory of Biological Environment Science for their kind help
during my work.
e Kiwakai Ai_ 'kido Club members.
' Kokusai Kouryuu Juku (Chikyukan) members.

●●
細岨andfriend亀f・rtheirsupP・rtandunderstanding
Final1第Ials・wisht・c・nveymydeepsense・fgratitudet・my
bel・vedfamily;with・uttheirenc・uragementandblessin&this
achievementwould not have been possible.

to my parents. . .

CONTENTS
Page
English Abstract ------ __ i
Japanese Abstract ------ __ v
List of Figures ------ __ vii
Chapter - I. ueneral mtroductron ---- 1
Chapter - II. Larval metamorphosis of the sea urchins,
Pseudocentrotus depressus and Anthocidaris
crasslspma m response to microbial films formed
in the sea and diatom-based films formed in the
mass-production tanks -------------------------------_ 6
Chapter - 111. The biological factors of diatom-based film for
larval metamorphosis of the sea urchins,
Pseudocentrotus depressus and Anthocidaris
crassrspma ---- ____ 26

Chapter-IV=
Ch叩ter」V
Refbrences
Characteristic ofthe chemical cue in diatom.based
films for larval metamorphosis ofthe seaurchins,
鳶εμ40cεn∫7伽s吻rεs5μ5andオn孟hoc∫伽∫s
C7αSS嗣ρ必α 一一一一一一一一一一一一一一一一一一一一一一一一一一一一_
General discussion and conclusions 一.舳一一一一.一.一.一一
45
59
66

AB STRACT
Most of marine benthic invertebrates possess planktonic larvae,
which gain the ability to select a suitable habitat for settlement and
metamorphose into juveniles during their competent stage. The
selection of substrate for larval metamorphosis is influenced by chemical
cues from microbial ~ilms, algae (heterospecific) and conspecific adults;
and also by physical factors. In the western part of Japan, diatom-based
film grown on plastic plates is used to induce larval metamorphosis of
sea urchin during the mass-production of juveniles. However, the role
of periphytic diatoms and bacteria co-existing in the films is poorly
understood. Therefore, this study focused on the larval response of two
commercially important species of sea urchin, Pseudocentrotus
depressus and Anthocidaris crassispina to microbial films developed in
the sea, diatom culturing tanks and laboratory conditions.
In Chapter - I, the relationship between microbial film and larval
metamorphosis of marine invertebrates especially of fisheries important
species were outlined. The role of microbial film in aquaculture for
mass-production of sea urchin juveniles was focused in this study.
In Chapter - II, the induction of larval metamorphosis of two
species of sea urchin by micyobial films formed in the sea and in the tank
(5-tonne) for sea urchin juvenile mass-production was studied for several
years. Films formed in the sea showed that the metamorphosis (%) of P
i

depressus increased gradually in accordance with the immersion period
of film formed on glass slides, whereas the larval metamorphosis of A.
irassispina showed a bell shaped response curve. In the tank, although
the diatom-based films showed a low inducing activity for larval
metamorphosis of A. crassispina, the metamorphosis of P depressus
larvae increased linearly in accordance with the diatom density. These
suggest that diatom-based films could promote the larval metamorphosis
of P depressus, but are less important in A. crassispina . Based on other
reports, I concluded that the high response of the former towards
nucroDral nlm tnan tne tatter rs due to the di~tt~erence in their natural
habitats where juveniles of P depressus are found in deep rocky subtidal
areas while ofA. crassispina live in shallow intertidal coralline algae bed.
Microbial films on rock surfaces act as nietamorphosis inducer for larvae
of P depressus, while the cue from algae induces larval metamorphosis
ofA. crassispina .
Chapter - 111 investigated the roles of diatoms and bacteria which
comprised laboratory-cultured diatom-based films (several diatom
species including bacteria) in the induction of metamorphosis in sea
urchin larvae. These diatom-based films induced larval metamorphosis
in both sea urchins, but the response rate was higher in P depressus.
Diatoms collected on the glass-fiber filters induced metamorphosis in
larvae of both species, although the inducing activity was significantly
less than that of the diatom-based films. No larva metamorphosed on
ii

bacterial films that formed from the filtrates, suggesting that bacteria
alone cannot induce metamorphosis and that diatoms play a major role in
the induction. Antibiotic treatment during assay reduced the inducing
activity in diatom-based films, even though the treatment did not control
bacterial growth. None of five isolated species of periphytic diatom
induced larval metamorphosis by itself and this suggests that no certain
diatom species was directly involved as an inducer. For diatom-based
film cultured in the presence qf germanium dioxide (Ge02) or antibiotic,
both species of sea urchins showed a similar response, in which
reduction in diatom and bacteria density resulted in a decrease in the
original inducing activity. There seems to be a synergistic effect
between diatom and bacteria in inducing larval metamorphosis.
Chapter - IV attempted to elucidate the characteristic of chemical
cue in laboratory-cultured diatom-based film. Films subjected to
treatment with 45'C heat, 0.1 N HCI and glutaraldehyde (5%) were no
longer inductive for either sea urchins, while those films treated with
40'C heat or EtOH (5%- and 10%-EtOH) showed a significant reduction
in the inducing activity. However, Iectin (LCA, SBA and WGA)
treatments had no effect. It i~ suggested that metamorphosis-inducing
cue(s) of live diatom-based films was unstable and sugar-related
compounds do not play a main role in the metamorphosis.
Chapter - V concluded that (1) microbial films acted as an inducer
for larval metamorphosis for both species of sea urchin although the
iii

response was different for each species and correlated with their natural
habitat. (2) It seems that diatoms and bacteria in the diatom-based
films produced a synergistic effect for induction of metamorphosis in sea
urchin larvae. (3) The metamorphosis-inducing cue(s) in diatom-based
films was unstable and sugar-related compounds do not play a main role
in the metamorphosis. This study provides initial information on the
role of microbial films on induction of larval metamorphosis in two
species of sea urchins, P depressus and A. crassispina and may improve
the techniques of juveniles mass-production of fisheries important
species of marine invertebrates especially sea urchins and other
echinoderms. Moreover, the identification of the chemical structure of
the cues and the receptor sites in the larva are important for
understanding the mechanism and chemical ecology of metamorphosis
in sea urchin larvae.
iv

La「va1弊etam・rphgsis・ftheSeaUrchins,P躍・cεn繊s吻ε5s麗sand。4n孟hoα伽7∫sc7αs3砂∫nαInducedbyMicrobialFilms
長崎大学大学院生産科学研究科
シティ・アクマル・カディジャ・ビンティ・アブル・ラヒム
(SitiAkmar KhadijahbinteAb Rahim)
海洋無脊椎動物の多くは浮遊幼生を経て、底棲性の成体になる。幼生から成体
への変態には、種々の基質と接触することが重要であり、この際に化学的な刺激
(ケミカルシグナル)や物理的な刺激を感受し、変態に至るものと考えらる。基
質としては・海藻や微生物フィルムなどが知られており、西日本におけるウニ種
苗生産でも、波板に付着珪藻フィルムを繁殖させて幼生の変態誘導を行ってい
る・しかし・微生物フィルムによるウニ幼生の変態機構についての知見は少ない。
そこで本研究では・西日本で水産上重要なアカウニ(」附θαの。θ四伽o云α5
砲ρ兜33α3)及びムラサキウニ(。4滋hoα幽万5α25磯ρ血8)幼生を対象として、
海域、大型水槽および室内実験にて、微生物フィルムと変態との関係を様々な観
点から追究した。
第1章では、ウニ幼生の変態と微生物フィルムとの関係について研究の概要を
まとめた。
第2章では、海域およびウニ種苗生産水槽(5トン)において形成された微生
物フィルムとアカウニ、ムラサキウニ幼生の変態誘起との関係を数年間にわたり
調べた。その結果、海域フィルムでは、アカウニの場合、垂下日数に従い変態率
が上昇したが1ムラサキウニでは山形(ベルシャープ)の傾向となった。種苗生
産水槽では、付着珪藻主体のフィルムが形成されるが、アカウニでは、珪藻密度
と変態活性とには明瞭な正の相関関係が認められた。しかし、ムラサキウニでは
全体に活性が認められない傾向となった。既報との関連から、アカウニは微生物
フィルムに反応し、ムラサキウニ幼生は海藻との関係で変態するものと結論され
た。
第3章では、付着珪藻フィルム(多種の付着珪藻および付着細菌)を室内にお
いて培養し、変態活性を確認すると共に、付着珪藻と付着細菌の役割について検
討した。ガラス繊維濾紙により珪藻と細菌とを大まかに分離したところ、珪藻側
V

に活性が認められた。先の付着珪藻フィルムから5種類の付着珪藻を単離した
が、これらに変態活性は認められなかった。また、珪藻の増殖阻害剤である二酸
化ゲルマニュームを添加した場合、および抗生物質を添加して珪藻フィルムを形
成させた場合は、両種共に、抗生物質添加でより大きく活性炉低下した。以上の
ことから、珪藻と綿菌とは各々単独では誘起活性が低く、両者が混合して活性が
発現してくるものと推察した。
第4章では、室内培養の付着珪藻フィルムについて、熱(300C-450C)、EtOH
(1%、5%及び10%)、塩酸(0.1N及び1.O N)の各処理、さらにはグルタール
アルデヒド固定を行ったところ、両種共に、活性は400C、EtOH(10%)、塩
酸(0.1N)処理で著しく低下した。またレクチン(LCA,SBA,WGA)添加で
は活性に影響がなかった。これらより培養珪藻フィルムの変態誘起物質は、著し
く不安定な物質であると推定された。また珪藻や細菌などの生死が活性発現に大
きく関係していることも示唆された。
以上のように、アカウニ及びムラサキウニ幼生に対して、両種で感受性は異な
るものの、微生物フィルムが変態にとって重要であることを明らかにした。次に
微生物フィルムの主な構成生物である付着珪藻と細菌とは共存して変態に関わ
るものと考察した。またフィルムの持つ変態誘起物質は化学的に不安定な物質で一
あると推定した。これらの知見は、ウニ類を含めた水産無脊椎動物の種苗生産技
術の改良、さらには海洋生物の変態機構の更なる解明につながるものと思わ液
る。
vi

LIST OF FIGURES
Page
Figure - 2.1. Methods for the larval culture. ------------------------- 9
Figure - 2.2. Larval assay procedure. ---------------------------_____ 11
Figure - 2.3. Preparation of (a) microbial films in the sea and
(b) diatom-based films in the mass-production
tanks. ---------------------------___________________________ 13
Figure - 2 4 Larval metamorphosis of P d ' ' ' , epressus m response
to microbial films of different immersion period
formed in the sea. --- ____ 16 Figure - 2.5. Larval metamorphosis ofA, crassispma m response
to microbial films of different immersion period
formed in the sea. --------------------___________________ 17
Figure - 2 6 Larval metamorphosis of P d ' ' ' . epressus m response
to diatom-based films of di~erent diatom density
formed in the tank. -------------_________________________ 19
Figure - 2.7. Larval metamorphosis ofA. crassispma m response
to diatom-based films of different diatom density
formed in the tank. ------------__________________________ 20
Figure - 2.8. Larval metamorphosis of two species of sea urchin,
P depressus and A, crassispina in response to
diatom-based films of different diatom density
formed in the tank in May. ---------------------------- 22
vii

Figure - 3.1.
Figure - 3.2.
Figure - 3.3.
r=igure - 3.4.
Figure - 3.5.
Page
Diatom-based film cultured in the laboratory. ------ 28
Preparation of diatoms on GF/C filter and
bacterial film developed from filtrate. --------------- 30
Larval metamorphosis of P depressus in response
to diatom-based films, diatoms on filters, bacterial
films and antibiotics-treated diatom-based films
(0.5 mg ml~1 penicillin G potassium and 0.01 mg ml~1
of streptomycin sulphate) during assay. ------------- 34
Larval metamorphosis ofA. crassispina in response
to diatom-based films, diatom on filters, bacterial
films and antibiotic-treated diatom-based films
(0.5 mg ml~1 penicillin G potassium and 0.01 mg ml~1
of streptomycin sulphate). ----------------------------- 35
Larval metamorphosis of P depressus in response
to diatom-based films cultured with germanium
dioxide. Ge02 (6.0 ug ml~1) and cultured with
antibiotic mixture (3.0 mg ml~1 penicillin G
potassium and 0.06 mg ml~1 of streptomycin
sulphate). ----------------------------------------------___ 37
viil

Page
Figure - 3.6.
Figure - 3.7.
Figure - 3.8.
Figure - 4.1.
Figure - 4.2.
Figure - 4.3.
Larval metamorphosis ofA. crassispina in response
to diatom-based films cultured with germanium
dioxide, Ge02 (6.0 ug ml~1) and cultured with
antibiotic mixture (3 .O mg ml~1 penicillin G
potassium and 0.06 mg ml~1 of streptomycin
sulphate). ------------------------------___________________
Larval metamorphosis of P depressus in response
to five species of periphytic diatom isolated from
diatom-based film. ---------------------------___________
Larval metamorphosis ofA crassis ' ' . pma m response
to five species of periphytic diatom isolated from
diatom-based film. --------------------------____________
Summary of heat and chemical treatments. ---------
Larval metamorphosis of P depressus in response
to diatom-based films treated with heat (300C, 350C,
400C and 450c). ________________________________________
Larval metamorphosis ofA. crassispina in response
to diatom-based films treated with heat (300C, 350C,
400C and 450C). ----------------________________________
38
40
41
48
50
51
ix

Figure - 4.4.
Page
Larval metamorphosis of P depressus in response
to diatom-based films treated with EtOH (1%, 5 %
and 10%), HCI (0.1 N and 1.0 N) and glutaraldehyde
(5%). ---------------------------------------.-------------- 53
Figure - 4.5.
Figure - 4.6.
Figure - 4.7.
Larval metamorphosis ofA. crassispina in response
to diatom-based films treated with EtOH (1%, 5%
and 10%), HCI (0.1 N and 1.0 N) and glutaraldehyde
(5%). ------------------------------------------------------ 54
Larval metamorphosis of P depressus in response
to diatom-based films treated with lectins (LCA,
SBA and WGA). -----------------------------------_____ 55
Larval metamorphosis ofA. crassispina in response
to diatom-based films treated with lectins (LCA,
SBA and WGA). -------------------------------_________ 56
x

CHAPTER - I
GENERAL INTRODUCTION
The life cycle of most marine invertebrates includes a planktonic
larval stage prior to a benthic adult phase. A critical phase during the
larval lifespan occurs when dispersed larvae gain the ability to select a
suitable habitat for settlement and metamorphose into juveniles. This
selection determines the long-term survival and consequently; the larvae
ot' a wide range of marine invertebrates do not settle and metamorphose
unless they encounter specific conditions, ensuring for their growth and
survival (Crisp, 1974; Cameron and Schroeter, 1980; Crisp et al. , 1985;
Snelgrove et al. , 1999).
Typically, settling larvae exhibit a specific searching behavior upon
contact with a suitable cue such as microbial film, macroalgae
(heterospecific) and conspecific adults; and also physical factors for
mstance, water flow, Iight and surface texture (Pawlik, 1992). Many
invertebrate larvae also exhibit the ability to delay metamorphosis until a
suitable substrate is available (Barker, 1977; Pechenik et al., 1993;
Mercier et al., 2000).
Microbial film is typically described as a succession of changes,
beginning with the formation of a layer of organic molecule film
consisting of amino acids, glycoproteins and humic materials, and
1

advancing to the colonization by bacteria, diatoms, fungi and protozoa
(Loeb and Niehof, 1975; Mitchell and Kirchman, 1 984; Costerton et al. ,
1995; Bhosle and Wagh, 1997; Tsurumi and Fusetani, 1998). The
formation of microbial films is a prerequisite for the metamorphosis of
marine invertebrate larvae (Zobell and Allen, 1935; Crisp, 1974; Pawlik,
1992; Johnson et al., 1997; Wieczorek and Todd, 1998).
Cues from microbial films induce larval metamorphosis of
coelenterate (Leitz and Wagner, 1 993), bryozoans (Brancato and
Woollacott, 1982; Keough and Raimondi, 1995), annelid (Kirchman et al.
1982; Unabia and Hadfield, 1999; Lau and Qian, 2001), mollusks
(Kawamura and Kikuchi, 1992; Slattery, 1992; Satuito et al. , 1995,
Robert and Nicholson, 1997) and echinoderms (Cameron and
Hinegardner, 1974; Barker, 1977), but has no effect on barnacles (Maki
et al., 1988).
Anrong echinoderms, it has been demonstrated that the larvae of
starfishes C , oscinasterias calamaria (Barker, 1 977) and Asterias rubens
(Barker and Nichols, 1983), settled on many kinds of hard substrates
covered with microbial films. The recruitment of crown-of-thorns
starfish Acanthaster planci was also associated with the periphytic
bacteria of crustose coralline algae (Johnson et al. , 199la, 199lb;
Johnson and Sutton, 1994). Bacteria-induced metamorphosis of the
sand dollar, Arachnoides placenta, was also reported in beakers used for
larval culture (Chen and Run, 1989). In sea urchins, Cameron and
2

Hinegardner (1974) observed that laboratory-reared Lytechinus pictus
and Arbacia punctulata larvae underwent metamorphosis in response to
microbial film. Similarly; field experiments on larval settlement in
Strongylocentrotus franciscanus and S. purpuratus (Cameron and
Schroeter, 1980; Rowley, 1989) and S. droebachiensis (Pearce and
Scheibling, 1991) demonstrated that surfaces with their microbial film
removed did not induce metamorphosis, whereas settlement did occur on
intact films.
Diatom-based film had been used in aquaculture for producing
juveniles of sea urchin, sea cucumber and abalone to sustain the wild
stock. In Japan, economically important sea urchin juveniles
(Anthocidaris ' ' Hemicentrotus crassl spma, pulcherrimus,
Pseudocentrotus depressus, S. intermedius, S. nudus) are being
mass-produced annually (Kltamura et al. , 1993). A flat or corrugated
plastic plate ("nami-ita") covered with periphytic diatoms in 5- to
15-tonne tanks is the most widespread method used in fish farms for
inducing the settlement and metamorphosis of sea urchi (T n ani and Ito,
1979; Ito, 1984; Ito et al. , 1991; Kawahara, 1996; Kitamura et al., 2000),
sea cucumber (Ito and Kitamura, 1997) and abalone (Kawamura and
Kikuchi, 1992; Kawamura, 1 996) Iarvae.
Despite the fact that success in mass-production of sea urchin
juveniles can be attributed to the successfiil use of diatom-based films as
inducers of metamorphosis in sea urchin larvae, the role of periphytic
8

diatoms and bacteria co-existing in the films is poorly understood. We
need more information on the roles of diatoms and bacteria as inducing
factors, in order to understand the importance of microbial film.
Therefore, this study focused on the larval response of two species of sea
urchin, P depressus and A. crassispina, which are important in Japanese
aquaculture, to microbial films formed in the sea and diatom-based films
formed in the tank used for culturing the periphytic diatoms (Chapter -
II) .
It is also important to understand the roles of diatom and bacteria
in diatom-based t~ilm and~ the mechanism involve in the induction of
larval metamorphosis of these two species of sea urchins. In Chapter -
III, studies were done to elucidate the biological factors in
laboratory-cultured diatom-based films that induce metamorphosis in sea
urchin larvae. We investigated the effect of diatoms and bacteria on
larval metamorphosis using diatom-based films cultured in the presence
of germanium dioxide or antibiotic mixture. In addition, we evaluated
the metamorphosis-inducing activity of five periphytic diatom species
isolated from the films.
In Chapter - IV, attempts were made to elucidate the characteristics
of the chemical cue in laboratory-cultured diatom-based film that
induced larval metamorphosis of P depressus and A. crassispina larvae.
The metamorphosis-inducing activity of film-associated cue was
evaluated after heat, ethanol, hydrochloric acid, glutaraldehyde or lectin
4

treatments.
5

CHAPTER - II
Larval metamorphosis of the sea urchins, Pseudocentrotus depressus
and Anthocidaris crassispina in response to microbial films formed in
the sea and diatom-based fihn formed in the mass-production tanks
II - I Ihtroduction
Larval metamorphosis is a nonrandom process that involves the
surface exploration and the final choice of suitable site is guided by
specific signals associated with a marine substratum (Pawlik, 1992;
Wieczorek and Todd, 1998). It has been observed in fieiu experiments
that echinoderm larvae metamorphosed on microbial films of substratum
and crustose coralline algae (Barker and Nichols, 1983; Johnson et al. ,
199la). Surfaces with their microbial film removed did not induce
larval metamorphosis of the sea urchins Stron8ylocentrotus franciscanus,
S. purpuratus (Cameron and Schroeter, 1980; Rowley, 1989) and S.
droebachiensis (Pearce and Scheibling, 1991).
In Japan, two popular methods are used for the annual
mass-production of five economically important sea urchins juveniles
(Anthocidaris ' ' Hemicentrotus pulcherrimus , crassl spma ,
Pseudocentrotus depressus, S. intermedius, S. nudus) (Kitamura et al. ,
6

1993). That is, competent larvae are induced to metamorphose on flat
or corrugated plastic plates ("nami-ita") covered with periphytic
diatom-based films in the western part of Japan and small discoid green
alga Ulvella lens in the northern region (Takahashi et al., 2002).
The studies mentioned above indicated that microbial films,
especially diatom-based films, have an importaht role in larval
metamorphosis of sea urchins. However, the role of periphytic diatom
and bacteria co-existing in the film still remains unknown. We need
more information on the role of diatom and bacteria as inducing factors
m order to understand the importance of microbial film.
Therefore, this study focused on the larval response of two species
of sea urchin, P depressus and A. crassispina, to microbial films in the
sea and diatom-based films formed in the tanks used for culturing the
periphytic diatoms.
II - 2 Materials and methods
For larval culture of two species of sea urchin, broodstocks of
Pseudocentrotus d epressus were purchased and stocked in the Nagasaki
City Fishery Center, Nagasaki, Japan while that of Anthocidaris
crassrspma were sampled from Nomo Bay, Nagasaki, Japan. Larvae of
P depressus and A. crassispina were cultured in the laboratory to the late
7

eight-armed competent stage with a fully developed echinus rudiment
(Kitamura et al. , 1993) (Fig. 2.1). Since the habitat and spawning
season of these two species differed, comparison on the larval response
was evaluated during May when both larvae were available.
Spawning of the sea urchins was initiated by 0.5 M KCI solution.
Eggs were rinsed with filtered seawater (FSW, WhatmanR glass fiber
filter, GF/C; I .2 um), fertilized with a dilute suspension of sperm and
rinsed several times with FSW. After ca. 20 h, swimming, prism larvae
were transferred to two 30-liter tanks filled with 25 Iiters of FSW
(stocking density ca. I Iarva ml~1). ~1'he culture tanks were continuously
aerated (200 ml min~1) in a dark room. About half of the seawater (10
liter) was renewed every day! Polyethylene nets, with mesh openings
of 111 and 225 um, were used sequentially as the larvae grew to filter off
the larvae. Food for the larvae was added after refilling the tanks.
The diatom Chaetoceros gracilis was used as food for larvae at
concentrations of 0.5xl04 to 3.0xl04 cells ml~1 in the culture tank; the
concentration was increased as the larvae grew. C. gracilis was
propagated under unialgal conditions in 1-liter flat-bottom flasks with
mixture of 900 ml of sterilized FSW, I ml of sodium metasilicate
nonahydrate (45mg ml~1) and 0.5 ml of KW21 medium (Daiichi Seikou
Co., Kumamoto, Japan) at 25'C. The flasks were illuminated at ca.
10,000 Lux and aerated (10 Iiter min~1) continuously. The diatom
increased in number from an initial density of 5.0xl04 cells ml~1 to more
8
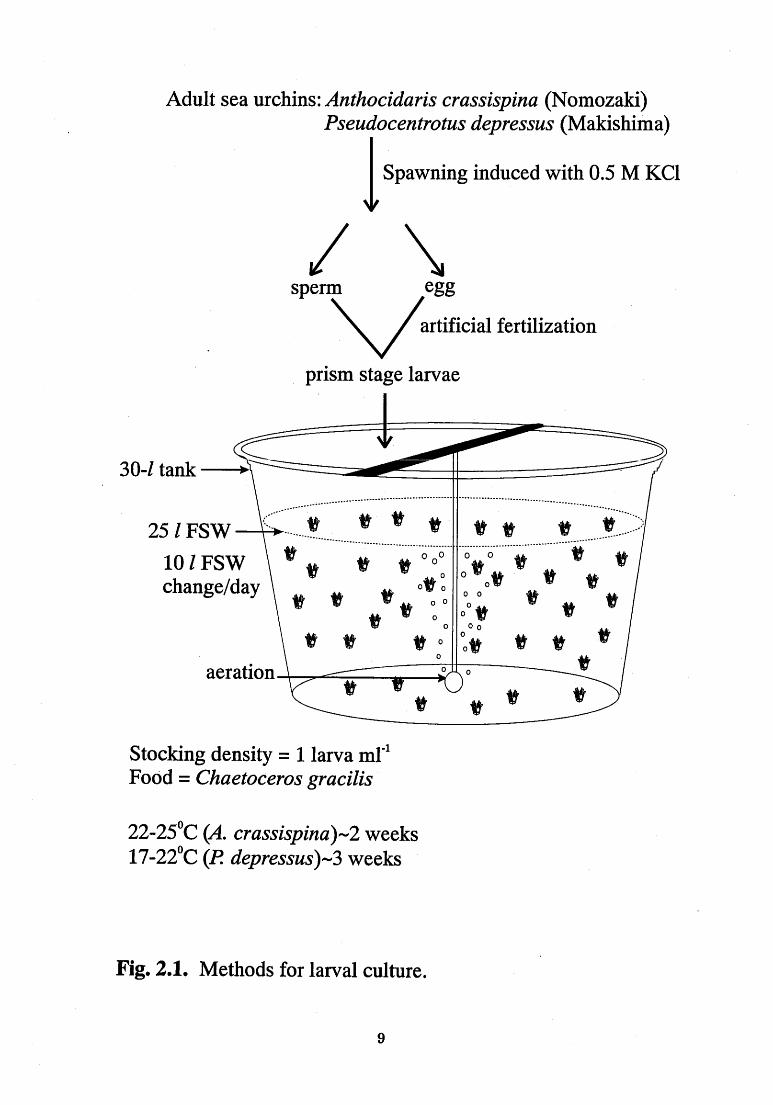
Adult sea urchins: Anthocidaris crassispina (Nomozaki)
Pseudocentrotus depressus (Makishima)
Spawning induced with 0.5 M KCl
sperm egg artificial fertilization
prism stage larvae
30-1 tank --~
25 1 FSW
10 1 FSW change/day
<:::~"~:::::: v ~ V ~
~V *" day ~ V ~~V ~"
1~ ooo c ov ~
:~~ ~ ~ vv ~
~~
~v ~ v ~ ation o
I~
Stocking density = I Iarva nil~1
Food = Chaetoceros gracilis
22_250C (A. crassispina)-2 weeks
17-220C (P depressus)-3 weeks
Fig. 2.1. Methods for larval culture.
9

than 1.0xl07 cells ml~1 within I week.
The larvae of P depressus were cultured at 17 to 22'C for ca. 3
weeks from October to March. In addition, competent larvae were also
obtained from the Nagasaki City Fishery Center, Nagasaki, Japan and the
Saga Prefectural Fish Farming Center, Chinze-cho, Saga, Japan in May.
The larval cultures of A. crassispina were done at 22 to 250C for ca. 2
weeks from May to August.
Larval assays for Pseudocentrotus depressus and Anthocidaris
crassispina were conducted 4 to 5 times from October to March and
once m May; and 6 to 7 times from May to August, from 1 997 to 2002,
respectively, In May; we had one chance to conduct the larval assay of
both species of the sea urchins simultaneously. In all assays, 200-ml
glass (PyrexR) beakers filled with 100 ml FSW were used (Fig. 2.2).
Subsequently, thirty 8-armed competent larvae were placed into each
beaker containing a piece of filmed glass slide or filmed beaker. These
beakers were kept in a dark room at 18'C and 22'C for P d . epressus and
A crasslspma respectrvely. Two types of control groups were
prepared in each assay: Iarvae exposed only to FSW, to ensure that the
larvae used were not spontaneously undergoing metamorphosis (negative
control); and larvae exposed to diatom-based films, to ensure that the
larvae used were competent to metamorphose (positive control).
Larvae were observed under a stereoscopic microscope after 24 h.
Larval response was assessed in terms of the proportion of
lO

Assay period P depressus : Oct. - Mar. &~)
A, crassispina : Aug
100 ml FSW
30 Iarvae ' 24 h (dark)
180C: P depressus
220C: A, crassispina
Microbial film (sea) Laboratory grown or diatom-based film (tank) diatom-based film
on glass slide
(38X26 mm)
Metamorphosis (%) = no,ofjuveniles X 100
30
Fig. 2.2. Larval assay procedure.
11

1~
metamorphosed individuals (%), defined as the percentage of individuals
that had undergone full metamorphosis to the juvenile form. All assays
were conducted in three to six replicates.
Microbial film samples in the sea were prepared by immersing
glass slides at Makishima Bay from a raft located near a jetty belonging
to the Nagasaki City Fishery Cepter, Nagasaki, Japan (Fig. 2.3a). Half
portion of glass slides (38x26 mm) were placed in plastic holders and
immersed vertically I .5 m below the sea surface. Immersions were
done almost throughout the year in 1996, 1997 and 1999, with each
immersion periods between I and 25 days. The means of diatom and
bacterial densities at twenty random spots on each glass slide were
recorded using a light microscope at 200x magnification and using the
phase-contrast microscope at 400x magnification, respectively. At the
same time, the genera of diatoms and other components in the film were
recorded.
Simultaneously with this experiment, similar glass slides were
immersed in 5-tonne tanks that contained plastic plates coated with
periphytic diatoms for the mass-production of sea urchin
Pseudocentrotus depressus juveniles (Fig. 2.3b). Each immersion
period was between I and 54 days. Navicula, Amphora, Achnanthes
and Nitzschia were the predominant diatom species, which proliferated
on the plates. Mass-production of sea urchin juveniles was carried out
twice a year (May and October) and the periphytic diatom plates in tanks
12

/ (a) Microbial film (sea)
- Immersion period: 1-25 days
R af t
1.5 m
III
Glass slide
(38X26 mm)
~
Plate
(b) Diatom-based film ("nami-ita" tank)
- Immersion period: 1-54 days
y
7
7
7
y
~
~
~
~
n Nami-ita'f
lll 111
Glass slide
(38X26 mm)
Plate
5-tonne tank
Fig. 23. Preparation of (a) microbial films in the sea and
(b) diatom-based films in the mass-production
tanks.
13

were maintained throughout the year.
To propagate periphytic diatoms on the plates, a flowing seawater
system with aeration was used. The light intensity inside the tanks was
controlled by covering the tanks with black mesh sheets (on a sunny day;
70-90% reduction in light intensity and on a cloudy/rainy day, 50%).
Fertilizers for diatom growth were added to the tank, and pesticides
(0.5-1.0 ppm of trichloroform, which is a safe level for sea urchin
juveniles) were used to eliminate the copepods that graze on the diatom
films. Plates coated with periphytic diatoms were washed with
high-pressure seawater (1-2 times per week) in order to select the highly
adhesive and small type of diatoms. Diatom and bacterial densities on
the glass slides were estimated following the same procedure mentioned
earlier. Microbial films and diatom-based films were then subjected to
larval assays described earlier.
All data are expressed as means (:!:SD) of three to six replicates
from three trials. Data were analyzed using the Pearson correlation test
(P<0.01) for the relationship between larval metamorphosis (%) and
diatom density in the tank film. One-way ANOVA followed by
Tukey's multiple comparison test (a=0.05) was used to compare the
metamorphosis (%) of control and other treatments D 'ff . I erences were
considered significant at P<0.001 .
14

II - 3 Results
Throughout all the experiments, the metamorphosis (%) of
competent larvae of Pseudocentrotus depressus and Anthocidaris
crassispina in clean beakers containing FSW was always O%.
The relationship between immersion period (days) and
metamorphosis (%) of two species of sea urchins on microbial films
formed in the sea is shown in Fig. 2.4 and 2.5. Metamorphosis of P
depressus increased gradually with immersion period (October to May)
(Fig. 2.4). Films immersed between 8 days and 23 days showed more
than 20% metamorphosis. On the other hand, the larval response ofA.
crassispina (Fig. 2.5) followed a bell-shaped curve from May to August.
Metamorphosis was high between 8-day and 15-day films, and the
metamorphosis-inducing activity decreased with immersion period. In
one-third of the total trials, the larvae of A. crassispina did 'not
metamorphose in response to microbial film.
Throughout the experiments in the sea, salinity and temperature at
the immersion site were 30-340/00 and 14-30'C, respectively! During
the rainy season (June to mid-July), the main components of the films
were detritus and silt. In the winter season, periphytic diatoms, such as
Navicula, Nitzschia, Amphora, Achnanthes, Licmophora and Cocconeis,
became predominant and the density reached about l05 cells cm~ .
During other seasons, aside from periphytic diatoms, other organisms
15

Oct.-Jan., May
~ ~~.'
~ ~ ~::
~ o ~ o ~:
1 OO
80
60
JA 4U
20
O
*
A A 11FO
A o. AO 6 A
*
* o A
AA
,
A
JL
e'o
A~ eO
A
A Oct
o Nov
e Dec
c Jan
* May
O 5 1 O 1 5 20 Immersion period (days)
25
Fig. 2.4. Larval metamorphosis of P depressus in response to
microbial films of different immersion period formed
in the sea. Each symbol represents the mean meta-
morphosis percentage of three replicates.
16

1 OO
80 '~, ~~OO
,~, , ea_,u:1 60
o ~l ~*
o ~ An ~! '~ o ~:
20
o
May-Aug.
vV
$e
v~.~
* $$
o $$ * *
v.
v
$$ ve v ~ e * o
'v ov 'vve *e
$e
* May
o Jun
v Jut
O 5 10 15 20 Immersion period (days)
25
Fig. 2.5. Larval metamorphosis ofA. crassispina in response to
microbial films of different immersion period formed
in the sea. Each symbol represents the mean meta-
morphosis percentage of three replicates.
17

such as protozoa, fungi, algal spores, filamentous blue-green algae and
copepods were also found on the film. In the sea films, we faced a
difficulty in counting the bacterial density accurately by direct counting
method, owing to the thickness of the film.
Half portion of glass slides (38x26 mm) were immersed in the
periphytic diatom plates culturing tanks under controlled conditions with
salinity and temperature of seawater being 30-340/00 and 10-30'C,
' respectively. Diatom density ranged from 103 to 106 cells cm~2
comprising Navicula, Nitzschia, Amphora, Tabularia and Achnanthes.
In determination of the bacterial density, we faced a difficulty owing to
the thickness of the film.
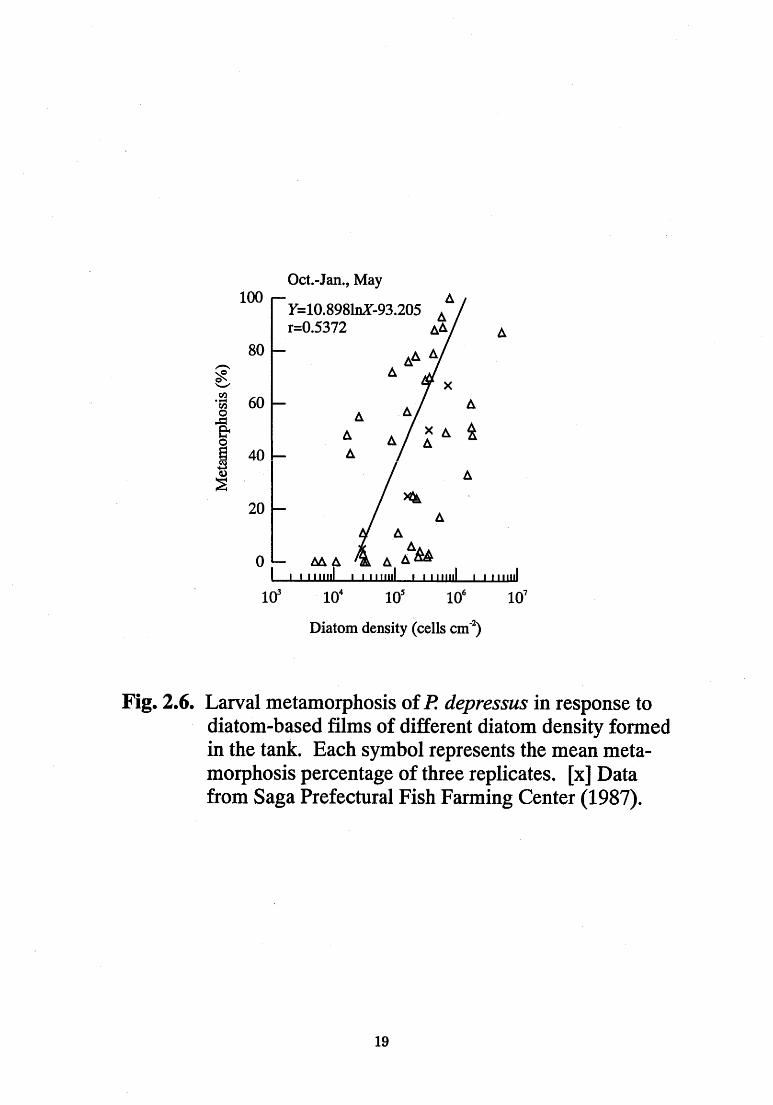
The relationship between the diatom density (cells cm~2) and
metamorphosis (%) of the diatom-based film in the tanks is shown in Fig.
2.6 and 2.7. Metamorphosis of P depressus larvae increased linearly,
with diatom density ranging from 2.0xl04 to 6.0xl05 cells cm~
(Y=10.898lnX-93.205; Pearson coefficient 0.5372, P<0.01) (Fig. 2.6).
The correlation coincided with that for the data of Saga Prefecture (1987;
Fig. 2.6). The experiment in Saga was done inside beakers by inducing
the larvae to metamorphose on corrugated plastic plates, covered with
diatom-based film, which were cut into small pieces. In contrast, the
larval metamorphosis ofA. crassispina was less than 20% in nearly 90%
of the total trials, although the diatom densities were high (Fig. 2.7).
Larvae ofA. crassispina showed lower metamorphosis on diatom-based
18
'~ ~~oe
,J enO
~::
el
O ~O
~:
100
80
60
40
20
O
Oct.-Jan., May
A Y=10.898hlX-93.205 A
r=0.5372 AA AAA
A
A ,A A A A
AAA
~!~
A AA^
A A~~
A
~
A
A
1 03 1 04 1 05 1 06
Diatom density (cells cm~2)
1 07
Fig. 2.6. Larval metamorphosis of P depressus in response to
diatom-based films of different diatom density formed
in the tank. Each symbol represents the mean meta-
morphosis percentage of three replicates. [x] Data
from Saga Prefectural Fish Farming Center (1987).
19

1 OO
80 '
~ ~~"
~ ~ 60 ~:Io
~h
~~ 40
~:
20
O
May-Aug.
Y=2.5188lnX-22.995 r=0.4636
o
oO 'O o O~-
O . er' Oo o O QOO;~~O OCO d 9
9, o
o
,
1 03 1 04 1 05 106
Diatom density (cells cm~2)
1 07
Fig. 2.7. Larval metamorphosis ofA. crassispina in response to
diatom-based films of different diatom density formed
in the tank. Each symbol represents the mean meta-
morphosis percentage of three replicates.
20

film compared to P depressus. Moreover, it was also observed that the
larvae of P depressus did not metamorphose on films with a diatom
density of <104 cells cm~ .
During the experiment with the diatom-based film, simultaneous
larval assay of these two species P depressus and A. crassispina, was
carried out in May (Fig. 2.8). As the diatom density increased from
2.0xl04 to 2.0xl05 cells cm~2, the mean metamorphosis rate for the
form~r was 39% and for the latter, 5%. Moreover, a closer correlation
between metamorphosis rate and diatom density was observed in P
depressus (Y=23.707lnX-217.14; Pearson coefficient 0.7066, P<0.01)
than in A. crassispina (Y=3.8753lnX-38.235; Pearson coefficient 0.4802,
P<0.01), which showed a clear difference between the two species in
response to the metamorphosis-inducing activity of diatom-based film.
21

100
80
~ ~~.'
~ "~ 60 ~i
~h
~ 40 ~
20
O
Pseudocentrotus depressus
Y=23.707lnX-217.14 r=0.7066
A A
A
A .A
A o A O A ..・・' ' 6
R;'~ O o
Anthocidaris crassispina
Y=3.8753lnX-38.235 r=0.4802
103 1 04 1 05 1 06
Diatom density (cells cm~2)
1 07
Fig. 2.8. Larval metamorphosis of two species of sea urchins,
P depressus and A. crassispina in response to diatom-
based films of different diatom density formed in the
tank in May. Each symbol represents the mean meta-
morphosis percentage of three replicates.
A : P depressus; O : A. crassispina
22

II - 4 Discussion
The induction of larval metamorphosis of most marine
invertebrates is influenced by chemical cues from microbial films, algae
(heterospecific) and conspecific adults; and by physical factors, such as
water flow, Iight and surface texture (Pawlik, 1992). Microbial films
induce larval metamorphosis of invertebrates such as bryozoans
(Brancato and Woollacott, 1982), hydrozoans (Leitz and Wagner, 1993),
polychaetes (Unabia and Hadfield, 1999), abalones (Kawamura and
Kikuchi, 1992), sea cucumbers (Ito and Kitamura, 1997) and sea urchins
(Ito, 1984), but has no effect on barnacles (Maki et al., 1988). Alnong
these organisms, bryozoan, hydrozoan and polychaete larvae
metamorphose on a clean surface, whereas for abalones and sea urchins,
the presence of microbial films is essential. In our study; we showed
the importance of microbial films in the larval metamorphosis of
Pseudocentrotus depressus and Anthocidaris crassispina.
In the sea, metamorphosis (%) of P depressus larvae increased
gradually with immersion period (Fig. 2.4), whereas the larval
metamorphosis of A. crassispina showed a bell-shaped response curve
(Fig. 2.5). Difference in larval response may be influenced by the film
components and growth rate of film, which reached the peak within a
shorter period during the hot season (May to August) than during the
cold season (October to May). We hypothesize that P depressus larvae
23

may also conform to the bell-shaped response curve if the immersion
period is extended beyond the 25-day period tested.
In the tank films, metamorphosis of P depressus larvae increased
linearly with diatom density ranging from 2.0xl04 to 6.0xl05 cells cm~
(Fig. 2.6), although the diatom-based films showed low
metamorphosis-inducing activity for larvae of A. crassispina (Fig. 2.7).
This pattem coincided with the data of Saga Prefecture (Ito et al. , 1991).
These results suggest that the periphytic diatoms on the tank film could
promote the larval metamorphosis of P depressus, but may be less
important in A. crassispina . Moreover, co-existing bacteria on the film
may also influence the larval response for both species of sea urchins.
There may be a difference between these two sea urchin species in
larval response towards microbial films, although the experiments were
conducted in two different seasons (Fig. 2.6, 2.7). A comparison of
results from larval assays simultaneous conducted in May (Fig. 2.8)
indicates that P depressus showed higher rate of metamorphosis than A.
crassispina in response to diatom-based films in the tank. This
difference seems to be related to their natural habitats P d . . epressus
juveniles are found in deep (8-12 m) rocky subtidal areas (Imai and Arai,
1994), whereas A. crassispina live in shallow (0.5-4.0 m) intertidal field
dominated by foliose coralline algae (Imai, 1980a, 1980b; Tsuji et al. ,
1989). Different habitats may influence larval response to certain
chemical cues in their environment.
/
24

The findings repOrted hereprovide initial information on the role of
microbial films developed in the sea and in tank$cult皿ing the periphytic
diatoms,on induction of larval metamorphosis in two species of sea
urchins,R勿r63s〃5and乱c7σss剛nα.
25

CHAPTER - 111
The biological factors of diatom-based film for larval
metamorphosis of the sea urchins, Pseudocentrotus depressus and
Anthocidaris crassispina
III - I Introduction
In mass-production of juveniles of sea urchin, sea cucumber and
abalone, diatom-based film propagated on plastic plates ("nami-ita") in
5- to 15-tonne tanks is used to induce larval metamorphosis (Ito et al. ,
1991; Kawamura and Kikuchi, 1992; Kawahara, 1996; Ito and Kitamura,
1997). Diatom-based film is mainly composed of several species of
diatoms and multispecies of bacteria, but the role of either diatom or
bacteria in the induction of larval metamorphosis remains unclear.
Chapter - 11 mentioned that the diatom-based films in the
mass-production tanks induced the larval metamorphosis of
Pseudocentrotus depressus and Anthoc ~l ' l ans crassispina even though
larval response differed between the two species. That is, diatom-based
films formed in the tanks could promote larval metamorphosis in P
depressus but were less important in A. crassispina.
In the present study; in order to clarify the role of diatoms and
26

bacteria as biological factors of metamorphosis in larvae of sea urchins,
we investigated the effect of variously treated laboratory-cultured
diatom-based films on the metamorphosis of P depressus and A.
crassispina larvae. The effect of diatoms and bacteria on larval
metamorphosis was also examined using diatom-based films cultured in
the presence of germanium dioxide or antibiotic mixture. In addition,
we evaluated the metamo h rp osis-inducing activity of five periphytic
diatom species isolated from the films.
III - 2 Materials and methods
Diatom-based f'l 1 ms were cultured in 200-ml beakers (Fig. 3.1).
The seed film was obtained from the filmed plastic plates in the 5-tonne
tanks of the Nagasaki City Fishery Center, Nagasaki, Japan, and cultured
for several days under laboratory conditions (14:10 LD cycle;
4,000-7,000 Lux; 23'C) in 50-ml flasks containing 30 ml modified
Erdschreiber culture medium (l~:tamura and Hirayama, 1984). The
film was then scraped off the inner surface of the flask, thereby
suspending it in the medium. One milliliter of this suspension,
containing 2.0-5.0xl04 diatom cells, was inoculated into each
experimental beaker containing 100 ml of the medium. The beakers
were left under the conditions mentioned above for 5 days to allow
27

辮鰯~~~肇~レ
~
~”Nami-itバco飢ed with
オ!periphyticdねt・ms 5
~
Eldschreibe1+vitamins(EV)
14:10LD cyclel4シ000-7,000Luxl230C
篇
Day O
100%EV
Day1雛圏聾斎麺醗→(顯
D&y2 Day4
50%EV FSW
Multispecies
dia重Qm and
bac⑯ria
FSW
Fig・3・LDiat・m-basedfilm?ulturedinthelab・・縫t・r弘
28

development of film. The culture medium was changed to 50%-
diluted medium after 2 days and to filtered seawater (FSW, WhatmanR
glass fiber filter, GF/C; I .2 um) after 4 days. Periphytic diatoms, viz.
Navicula, Nitzschia, Amphora, Achnanthes and Cocconeis dominated
this cultured film. Each beaker was refilled with 100 ml FSW prior to
larval assays.
In a parallel experiment, variously treated diatom-based films were
prepared. Diatom-based film was scraped off one beaker, resuspended
in 100 ml FSW, and filtered through a glass fiber filter (WhatmanR,
GF/C). The filter with diatoms (hereafter referred to as "diatoms on
filter") was placed on the bottom of a beaker, which was then filled with
FSW and subjected to larval assay! On the other hand, the filtrate was
poured into a clean beaker to allow the development of bacterial film
(hereafter referred to as "bacterial film") and subjected to larval assay
(Fig. 3.2). A bacterial film with a density of 6.0xl05 cells cm~ was
verified to have formed on the bottom of the beakers within 2 h after the
commencement of assays.
Variously treated diatom-based films were also subjected to larval
assays. The first treatment was antibiotic treatment. Diatom-based
films were treated with FSW containing an antibiotic mixture (0.5 mg
ml~1 of penicillin G potassium and 0.01 mg ml~1 of streptomycin
sulphate) and subjected to larval assay. These antibiotics were
purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan).
29

Scraped film from a beaker
Film suspension in 100 ml FSW
Diatoms on filter
GF/C filter
"~~!~=~~~~~~-~_~~~ - ~!.~~!I~~~ ~~~
.~'_ ~~"=~~_=~~~-}~~~-S~~=.~~~'~. ~~~
~. ~=~-
Fig. 3.2.
Bacterial film
Preparation of diatoms on GF/C filter and
bacterial film developed from filtrate.
30

The specified concentrations of these antibiotics had been verified to
have no toxic effect on larvae (own unpublished data).
The effect of diatoms and bacteria on larval metamorphosis was
examined by using diatom-based films with germanium dioxide (Ge02)
or antibiotic mixture added in the film culture. For this experiment,
diatom-based films were cultured in 200-ml beakers containing 100 ml
of culture medium and cultured for 5 days under the same condition as
mentioned above (Fig. 3.1). Germanium dioxide (6.0 ug ml~1) to
depress the diatom density or antibiotic mixture (3.0 mg ml~1 of
penicillin G potassium and 0.06 mg ml~1 of streptomycin sulphate) were
added during the film culture period. The culture medium was changed
to 50%-diluted medium after 2 days and to FSW after 4 days with each
reagent, respectively! Each beaker was refilled with 100 ml FSW prior
to larval assays.
Five species of periphytic diatom (Achnanthes, Amphora,
Cocconeis, Navicula and Nitzschia) were successfully isolated from the
diatom-based film and were evaluated f or metamorphosis-inducing
activity in larvae. Each species of diatom was cultured from a single
cell isolated from a diluted film suspension under the stereomicroscope
using a fine tip capillary pipette. Each type of unialgal diatom film was
cultured in beakers using the same method described above for
diatom-based films (Fig. 3.1) and was subjected to larval assay. Larval
assays were conducted as described in Chapter - 11 (Fig. 2.2).
31

Throughout this study, enumeration of diatoms and bacterial
densities were counted from filmed glass slides (38x26 mm) prepared in
a similar manner as filmed beakers. Mean diatom and bacterial
densities were taken from twenty random spots on each glass slide and
were counted using a light microscope at 200x magnification and a
phase contrast microscope at 400x magnification, respectively.
All data are expressed as means (:!:SD). Differences between
control and treatments were analyzed using one-way ANOVA followed
by Tukey's multiple comparison test. Results were assessed significant
at P<0.001 .
III - 3 Results
Throughout all the experiments, the metamorphosis (%) of
competent larvae of Pseudocentrotus depressus and Anthocidaris
crassispina in clean beakers containing only FSW (negative control) was
always O%. By contrast, the diatom-based films (positive control) that
consisted of multispecies periphytic diatoms and bacteria, showed larval
metamorphosis, ranging from 87% to 95% in P depressus and from 5 1 %
to 79% inA. crassispina.
The metamorphosis-inducing activities of the diatoms on filters,
the bacterial films and the diatom-based films treated with antibiotics
32

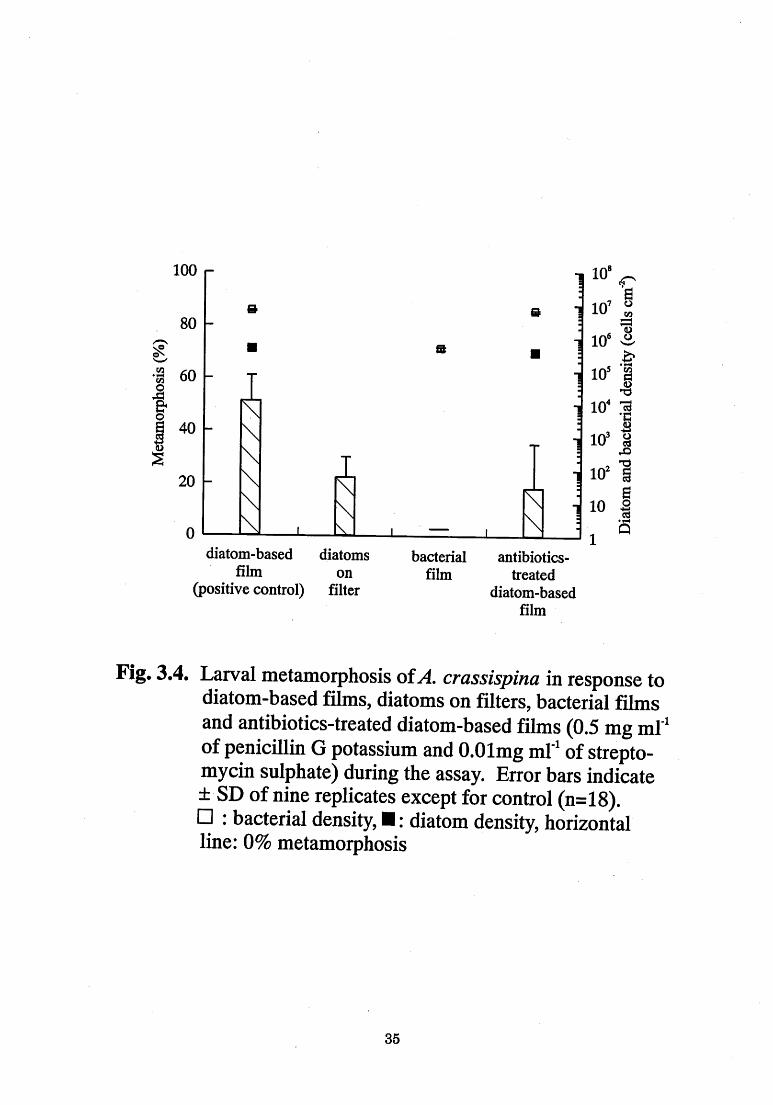
during the assay for both sea urchins are shown in Fig. 3.3 and 3 .4. In
P depressus (Fig. 3.3), the diatom-based films (positive co~trol) induced
95 % metamorphosis, while the diatoms on filters induced 67% Iarval
metamorphosis. However, no metamorphosis occurred on bacterial
films. Anthocidaris crassispina (Fig. 3.4) showed a response similar to
that of P depressus. Diatom-based films (positive control) induced
5 1 % metamorphosis, while the diatoms on filters induced 22% Iarval
metamorphosis. Bacterial films also did not induce larvae of A.
crassispina to metamorphose. In both species of, sea urchin, the
diatoms on filters showed lower activity than the diatom-based films
(p<d.OO1).
In assays with P depressus, the bacterial density of film treated
with antibiotics was 6.9xl06 cells cm~ while that of non-treated film
(positive control) was 7.1xl06 cells cm~2 24 h after the start of assay.
Thus, treatment with antibiotics did not reduce bacterial density; but it
did drastically reduced the inducing activity of diatom-based films (Fig.
3.3, 3.4). Assays employing larvae ofA. crassispina showed the same
trend. The rate of metamorphosis decreased to 3% and 18% in P
depressus and A. crassipina, respectively.
33

'~ ~~ee
¥J
O ~l ~l
O ~ Q) ~:
100
80
60
40
20
O
diatom-based f i Im
(positive control)
e
a
diatoms
on f ilter
bacterial antibiotics-f ilm treated
diatom-based f ilm
108 ~_
70S 10 ca
=t o 106 o ~ ~ 10s ~s~va
(D ~1
104 ~i::
o 103 *oe8
~ 11 102 ~
~ 10 _oee
,*
1 O
Fig. 3.3. Larval metamorphosis of P depressus in response to
diatom-based films, diatoms on filters, bacterial films
and antibiotics-treated diatom-based films (0.5 mg ml~1
of penicillin G potassium and 0.0lmg ml~1 of strepto-
mycin sulphate) during the assay. Error bars indicate
:b SD of nine replicates except for control (n=18).
[1 : bacterial density; I : diatom density; horizontal
line: O% metamorphosis
34
~ ~~.'
~ ~:lOa:I
~ O ~O
:~
IOO
80
60
40
20
O
e
l a:
s
I
diatom-based diatoms f ilm on
(positive control) filter
bacterial antibiotics-
film treated diatom-based
f ilm
108 ~,~
'~
107 o gD
~ -o 106 o ,~ ~?
105 *F~uD
o ~:'
104 ~~:.::
o ~' In3 o lu cs
~ ~! 102 ~
~ 10 o *, Gs
,* 1 C~
Fig. 3.4. Larval metamorphosis ofA. crassispina in response to diatom-based films, diatoms on filters, bacterial films
and antibiotics-treated diatom-based films (0.5 mg ml~1
of penicillin G potassium and 0.0lmg ml~1 of strepto-
mycin sulphate) during the assay. Error bars indicate
:t SD of nine replicates except for control (n= 1 8).
[] : bacterial density, I : diatom density, horizontal
line: O% metamorphosis
35

In diatom-based films cultured without addition of germanium
dioxide (Ge02), nor of antibiotic mixture (positive control), mean
metamorphosis rate of P depressus and A. crassispina were 87% and
79%, respectively (Fig. 3.5, 3.6). For films cultured with Ge02,
metamorphosis of P depressus was reduced significantly to 35%
(P<0.001) when diatom density was reduced by two orders of magnitude
(1.0xl03 cells cm~ , compared to the positive control film, 4.6xl05 cells
cm~2) (Fig. 3.5). Bacterial density on the Ge02 film was similar to that
on the positive control film. In contrast, metamorphosis-inducing
activity was reduced drastically to 9% (P<0.001) in film cultured with
antibiotic mixture, and bacterial density was reduced by one order of
magnitude (5.3xl05 cells cm~ , compared to the positive control film,
5.6xl06 cells cm~2). Diatom density on antibiotic films was similar to
that on the positive control film.
A. crassispina (Fig. 3.6) also showed similar response to that of P
depressus. The metamorphosis of film cultured with Ge02 Was reduced
significantly to 52% (P<0.001) when diatom density was reduced by two
orders of magnitude (2.4xl03 cells cm~ , compared to the positive control
film, 3.2xl05 cells cm~2). Larval metamorphosis was reduced to 16%
(P<0.001) in film cultured with antibiotic mixture, when the bacterial
density was reduced by an order of magnitude (4.8xl05 cells cm~ ,
compared to the positive control film, 4.2xl06 cells cm~2).
36

1 OO
80
~ ~~.'
~ '"* 60 ~i
~ ~o 40
~:
20
O
diatom-based f ilm
Gpositive control)
A
e
I ~
B
c
+ G, e02 + antibiotic
mixture
108 ~-
70~ 10 GQ
~ ~ 106 ~
~ .* 10s ~ o 1:'
104 ~!:'
o 103 ~o,s
~ ~' 102 ~
~ 10 o ~ es
.* 1 C:i
Fig. 3.5. Larval metamorphosis of P depressus in response to
diatom-based film cultured with germanium dioxide,
Ge02 (6.0 u g ml~1) and cultured with antibiotic mixture
(3 .O mg ml~1 of penicillin G potassium and 0.06 mg ml~1
of streptomycin sulphate). Error bars indicate i SD of
eighteen replicates. Values not sharing common letters
are significantly different (P<0.001). [] : bacterial
density; I : diatom density
37

100
80
~ .¥'
~ '~ 60 ~!
~ ~o 40
:~
20
O
diatom-based f ilm
(positive control)
A a:
~ B
c
+ Ge02 + antibiotic mixture
108 ~_ '~
107 ~
~ ~ o 106 ~'o
~ 10s ~ o ~:'
104 ~i::
_o 103 cso
~ ~' 102 ~
~ 10 -eeo
,* 1 C:I
Fig. 3.6. Larval metamorphosis ofA. crassispina in response to
diatom-based film cultured with germanium dioxide
Ge02 (6.0 u g ml~1) and cultured with antibiotic mixture
(3.0 mg ml~1 of penicillin G potassium and 0.06 mg ml~1
of streptomycin sulphate). Error bars indicate :1: SD of
eighteen replicates. Values not sharing common letters
are significantly different (P<0.001). [1 : bacterial
density; I : diatom density
38

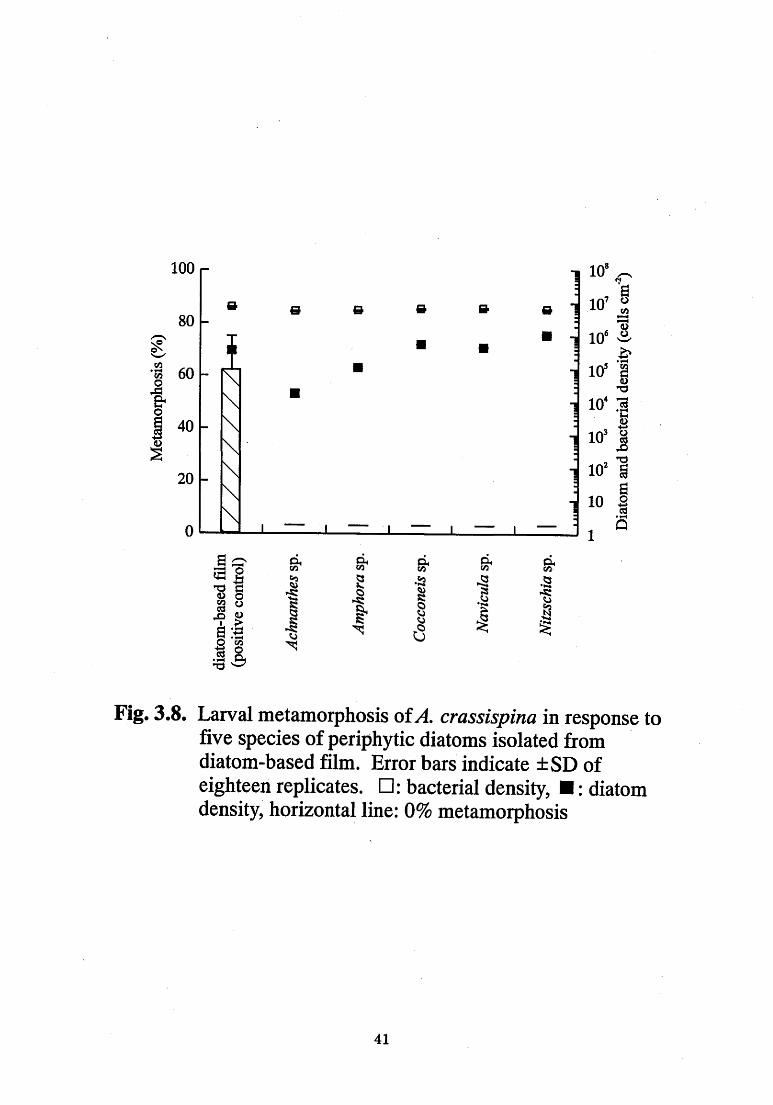
None of the five species of diatom isolated from the diatom-based
film(、4chnαn孟hε&。4醒ρho鵜Cocconε鶴ハ砂v∫cμ伽and荒砿sch∫α)induced
larval metamorphosis in either gfthe two species ofsea urchin(Fig.3.7,
3.8);nevertheless,the metamorphosis rate of R喫ρrε3sμ3and1生
c■αssゆ功α exposed to multisp停cies diato血一based films (positive
“ontrol)were89%and62%,respective1》乙
39

F, ~~ee
~'
O ~ el O ~ Q) ~:
1 OO
80
60
40
20
o
e
I
e
l
e
l
e
l
e
l
~ l~ ~ q:1,,:!o
1$~l Q)O ~o ~o l> ~~~ ~!oca
~~
e~ C':I
~*~
~:
~ ~~!)
,~:
v:~
L~
~ E~~
~:
pl Ga EQ .~ ~) ~: q) ~) ~) Cb ~J
~l co
_~~$
~: ~b .~ ~~~
Ga~
.~ ~~)
:~!._.~e
~
108
~~ '~
107 o ~,D
~ ~ In6 o u~ ~ 105 '~
o =1 104 ~j
.F: 103 *o'L)c:l
~ 102 ~' ~ ~
10 _o~s
.~ 1 O
Fig. 3.7. Larval metalnorphosis of P depressus in response to
five species of periphytic diatoms isolated from
diatom-based film. Error bars indicate :1: SD of .
eighteen replicates. [] : bacterial density, I : diatom
density; horizontal line: O% metamorphosis
,
40
~ ~~.'
~ ~
~::
~ o ~O
~:
100
80
60
40
20
o
e s
l
5
l
e
l
e
l
e l
~(:r ~::O -S! ~'$::
OO ~O ~O l .> ~:i-~,**
Oc'D ~'eS O '~ie'
eo~
~:~
~;
~: ~~:cL)
~~
~$::*
L~$
~ ~~~
~:
olE'e
ee .~ qD ~: cb ~) c!)
s ~)
O1 C':I
'~~~$
~: ~) .~ ~~~
pl ca-
.~ ~~ ~)
.1~.__N~':
~
108 ~_
70~ 10 c'b
~ -68 10 -~ .* 10s ~ o 11
104 ~!_oE1
103 ~cso
JD It
102 ~
~ 10 -o~5
.~ 1 C:I
Fig. 3.8. Larval metamorphosis ofA. crassispina in response to
five species of periphytic diatoms isolated from
diatom-based film. Error bars indicate :!: SD of
eighteen replicates. [1 : bacterial density~ I : diatom
density, horizontal line: O% metamorphosis
41

-~
III - 4 Discussion
Diatom-based films have been used to induce larval metamorphosis
during mass-production of sea urchins juveniles (Ito et al. , 1991;
Kawahara, 1996). In the present paper, the results of a preliminary
study to elucidate the roles of diatoms and bacteria on larval
metamorphosis of Pseudocentrotus depressus and Anthocidaris
crassispina have been reported.
Attempts were made to evaluate separately the
metamorphosis-inducing activity of diatoms and bacteria from
laboratory-cultured diatom-based film. Diatoms on the filter induced
metamorphosis in larvae of both species, even though the activity was
significantly lower than that of the control (Fig. 3 .3, 3.4). By contrast,
bacterial films did not induce metamorphosis in larvae of either species.
These results suggest that bacteria alone cannot induce larval
metamorphosis and that diatoms play a major role in the induction of
larval metamorphosis in these sea urchins. However, it should be noted
that the metamorphosis-inducing activity of the diatoms on filter showed
a significant decrease compared to the film; therefore, it can be assumed
that a metamorphosis-enhancing factor in diatom-based film passed
through the filter. It is also possible to hypothesize that the mixture of
diatoms and bacteria provide a synergistic effect for the
metamorphosis-inducing activity of diatom-based film. A different
42

explanation could be a difference in the configuration of the diatom films
cultured on beakers and those collected on filters. This reduction in
metamorphosis-inducing potency requires further investigation.
In the present investigation, treatment with antibiotics (penicillin
and streptomycin) drastically reduced the metamorphosis-inducing
activity of diatom-based films, even though bacterial growth was not
controlled. The concentrations of antibiotics used in this experiment
should had no adverse effect on the sea urchin larvae (own unpublished
data); I suggest that the antibiotic mixture may have affected the
condition of the diatom-based films.
The larvae of P depressus 'and A. crassispina showed similar
response towards the film cultured with germanium dioxide and
antibiotic mixture (Fig. 3.5, 3.6). For both species, reduction in
bacterial density had a greater effect on the metamorphosis-inducing
activity of films than did reduction in diatom density; with a drastic
decrease of 80% to 90% in the former, compared to 35% to 60% in the
latter. Based on this results, it is possible that the
metamorphosis-inducing activity of diatom-based film for larval
metamorphosis of these sea urchins may be due to the synergistic effect
of diatoms and bacteria as mentioned earlier (Fig. 3.3, 3.4).
None of the five isolated species of periphytic diatom induced
metamorphosis in larvae of P depressus and A. crassispina (Fig. 3.7,
3 .8). This suggests that no single diatom species is directly involved as
43

an inducers of metamorphosis in sea urchin larvae. The elucidation of
the role of diatoms in combination with other factors requires further
investigation.
Although diatom-based films cultured in the laboratory induced
larval metamorphosis in both P depressus and A. crassispina, the
response differed between the two species; higher metamorphosis (%)
was observed in P depressus than in A. crassispina . This observation
is consistent with the results shown in Chapter - II; that is, diatom-based
films formed in the sea urchin juvenile mass-production tanks promote
larval metamorphosis in P depressus but are less important in A.
crassl spma .
44

CHAPTER - IV
Characteristic of the chemical cue in diatom-based films for larval
metamorphosis of the sea urchins, Pseudocentrotus depressus and
Anthocldans crassispina
IV - I Introduction
As mentioned in the previous chapters, competent larvae of sea
urchins Pseudocentrotus depressus and Anthocidaris crassispina must be
able to detect the metamorphosis-inducing cue released by suitable
inducers such as microbial films in the sea, diatom-based films formed in
the sea urchin juveniles mass-production tanks and laboratory-grown
f ilms .
A number of researchers have studied the chemical cues for larval
metamorphosis of sea urchins derived from algae, although real chemical
cues have been rarely identified. Free fatty acids, such as
eicosapentaenoic acid and arachidonic acid extracted from coralline red
alga Corallina pilullfera, were reported to induce larval metamorphosis
of P depressus and A. crassispina (Kitamura et al., 1992; 1993).
Glycoglycerolipids from green alga Ulvella lens induced larval
metamorphosis of Stron8ylocentrotus intermedius (Takahashi et al.,
45

2002). A water-soluble complex of the sugar floridoside and isethionic
acid isolated from red alga Delisea pulchra triggered metamorphosis in
Holopneustes purpurascens larvae (Williamson et al. , 2000).
Dibromomethane that was detected in the seawater, where U. Iens or
coralline red algae were previously cultured, induced larval
metamorphosis of S. nudus (Taniguchi et al. , 1994). In larvae of P
depressus and A. crassispina, high concentration of synthetic
dibromomethane induced metamorphosis at 3,800 ppm and 2,200 ,ppm,
respectively (Koh et al., 1996).
An amino acid, L-glutamine also induced larval metamorphosis in
several sea urchins, Hemicentrotus pulcherrimus (Yazaki, 1995), P
depressus (Yazaki and Harashima, 1994) and S. intermedius (Naidenko,
1996). Naidenko (1996) also reported that the epiphytic calcareous
alga Melobesia sp., colonizing the older sea grass Zostera marina,
induced metamorphosis of S. intermedius larvae and this alga contained
L-glutamine. Larvae of S. droebachiensis metamorphosed in response
to GABA-mimetic molecules contained in coralline red alga
Lithothamnion glaciale (Pearce and Scheibling, 1990).
In the present study, we attempted to elucidate the characteristics of
the chemical cue in the laboratory-cultured diatom-based films that
induced larval metamorphosis of P depressus and A. crassispina larvae.
The metamorphic activity of potential cues in film was evaluated after
heat, ethanol, hydrochloric acid, glutaraldehyde or lectin treatments.
46

IV - 2 Materials and methods
The nature of the chemical cues in laboratory-cultured
diatom-based films for larval metamorphosis was investigated by heating,
ethanol (EtOH), hydrochloric acid (HCl), glutaraldehyde and lectin
treatments, and subjected to larval assays (Fig. 4.1). Heat and lectins
treatments were carried out to determine whether the cue is heat-stable
and is a sugar-related compound. HCI and glutaraldehyde treatments
were done to check the induction activity of non-living films.
Diatom-based films were heated (30'C, 35'C, 40'C or 45'C) in a
water incubator for 30 min. Diatom-based films were also treated for
30 min with EtOH (1%, 5% or 10%) or HCI (0.1 N or 1.0 N). In the
glutaraldehyde (5%) treatment, diatom-based films were treated for 24 h,
then rinsed with FSW 3 times per day for 3 consecutive days. The
treatments of diatom-based films were also performed with the lectins
LCA (Lentil Agglutinin), SBA (Soybean Agglutinin), or WGA (Wheat
Germ Agglutinin) at 100 ug ml~1 concentration for 2 h. The lectins
were purchased from Wako Pure Chemical Industries, Ltd. (Osaka,
Japan). Prior to larval assays, all filmed beakers were rinsed with FSW
for 3 times and refilled with 100 ml FSW. Larval assays were
conducted as described in Chapter - 11 (Fig. 2.2).
Throughout this study; Iaboratory-cultured diatom-based film was
prepared as mentioned earlier in Chapter - 111 (Fig. 3.1). Diatom and
47

Treatment
Hea‡(30min)
300C,350C,400C,450C
EtOH(30min)
1%,5%,10%
HCl(30min)
0.1N,1.ON
Glutaraldehyde,5%(24h)
.恥ctins・100國m1(2h)
LCA,SBA,WGA
rinsewith
FSW
Larval
assay
Fig.4.1。Summary ofheat and chemical treatments.

bacterial densities were enumerated as described in Chapter - 111.
Statistical analyses were done as mentioned in Chapter - 111.
IV - 3 Results
Throughout all the experiments, the rate of metamorphosis in
competent larvae of Pseudocentrotus depressus and Anthocidaris
crassispina in negative controls was always O%. By contrast, the
laboratory-cultured diatom-based films (positive control) that consisted
of multispecies periphytic diatoms and bacteria, induced larval
metamorphosis greater than 90% in P depressus and from 61% to 85%
in A. crassispina . The diatom and bacterial density was determined
after heat, ethanol (EtOH), hydrochloric acid (HCl), glutaraldehyde or
lectin treatments mentioned below, and no significant difference was
observed compared to the untreated film.
Larval metamorphosis in both species of sea urchins exposed to
diatom-based films treated with heat is shown in Fig. 4.2 and 4.3. For P
depressus (Fig. 4.2), diatom-based films (positive control) showed 93 %
metamorphosis and the metamorphosis-inducing activity decreased
drastically to O% at 40'C and 45'C (P<0.001). ForA. crassispina (Fig.
4.3), the larval response was similar to P depressus. Diatom-based
films (positive control) showed 61% metamorphosis and the
49

'~ ~:~ee
'~'
~:O
~ O ~ o :~
1 OO
80
60
40
20
O
diatom-based film
(positive control)
~
E: i l~lli '!'
.
~=
al l
,i
~~i
ll
i ll l lr '
'l
~' ji~ii,
.I ~ta
~l
'l:
I ~ 'ii
~l
':~: ~"' il 'i~ii
iiil
ii ~
l --
I~ i:1 :
300C 35"c 400C 450c
108 ~,~
'~
107 o c'a
-~ o 106 o ,J ~ ." 105 ~ o =1
104 .ce~~
~a)
In3 o lu ~~ ~'
102 ~
~ o 10 ~~ Gs .* C:I
1
Fig. 4.2. Larval metamorphosis of P depressus in response to diatom-based film treated with heat (300C, 350C, 400C
and 450C). Error bars indicate :!: SD of eighteen
replicates. [] : bacterial density; I : diatom density;
horizontal line: O% metamorphosis
50

l~* ~~OO
~J . Ga_,
O ~ 81 O ~Q)
~:
100
80
60
40
20
o
~l
jii~i
~ l!i
~pl
jl
j~~i*
lli~i*,~
~lpi~
' ~~
j~i~~
j~i~i~
~~~,':1!
~i
l~
~jr;'~i
.~i
p*
~ .~
s
l
di~i#
jr'
:E
l
diatom-based 300C film
(positive control)
35'c 40'c 450C
108 ~,_.
'~
107 o c':I
~ ~ o 106 ~~
~ 5 '~uD 10 ,sl *Q) ~)
104 .~ ,~ ~o
103 ~c~o
=1 102 ~
~ 10 o ~' ee
.* O 1
Fig. 4.3. Larval metamorphosis ofA. crassispina in response to
diatom-based film treated with heat (300C, 350C, 400C
and 450C). Error bars indicate :1: SD of eighteen
replicates. [] : bacterial density; I : diatom density;
horizontal line: O% metamorphosis
51

metamorphosis-inducing activity decreased drastically to 10% and O% at
40'C and 45'C, respectively (P<0.001).
The metamorphosis-inducing activity of diatom-based films treated
with EtOH, HCI or glutaraldehyde is shown in Fig. 4.4 and 4.5. For P
depressus (Fig. 4.4), metamorphosis was also reduced significantly to
50% in 5 %-EtOH and to 7% in 10%-EtOH-treated diatom-based films
(P<0.001). By contrast, Iarvae did not metamorphose in response to
HCI (0.1 N or I .O N) or 5% glutaraldehyde treated diatom-based films
(P<0.001). For A. crassispina (Fig. 4.5), the larval response was
similar to P depressus except in diatom-based films subjected to
5 %-EtOH treatment. In A. crassispina, metamorphosis decreased
significantly to 22% only in 10%-EtOH-treated diatom-based films
(P<0.001), but not in 5%-EtOH. Diatom-based films treated with HCl
or glutaraldehyde did not induce metamorphosis (P<0.001).
On the other hand, Iectin (LCA, SBA or WGA) treatments did not
affect the metamorphosis-inducing activity of diatom-based films either
in P depressus (Fig. 4.6) or A. crassispina (Fig. 4.7).
52

'~ ~~eo
'v'
uDO
~:1
B~
O ~ 'L)
~:
1 OO
80
60
40
20
o
~ dll~:'
d ~ ~i:
p lpll~I~
~ B S g!: 9: l
~~~!f*
~ d~::
i!r
~l~!' E
!!1li~' !!!~
illli*' '
illii~:~" l~~~
~l~llii ~i!l ~
'd:**!!*
l~"' .*.
~~ii~' jpii~i
.'~~d '~k ~・*
diatom- 1% 5% 10% 0.1 N based
EtOH
108 -~~
70 10 va --o 60 10 -~~
10s c'aoF~
~' 104 ~I,~
o ~ 103 ~c:'o
~1 102 ~
~ o 10 -~3 .* C:I
1
1.0 N 5% glutar-
aldehyde
(positive control)
Fig. 4.4. Larval metamorphosis of P depressus in response to
diatom-based film treated with EtOH (1%, 5% and
10%). HCI (0.1 N and 1.0 N) and glutaraldehyde (5%).
Error bars indicate :1: SD of eighteen replicates.
[] :, bacterial density; I : diatom density; horizontal
line: O% metamorphosis
53

~ ~~'e
,J
~ ~:I
~ O ~ o :~
100
80
60
40
20
O
o o
~ rr
llllk!i'
'~l jii~
* e~-*
~! ..~
piF'
5
ptli
lliii~
~l~i~
'I:1 ~
i~:'
~
~L
l
~ ~.
j~i
~!-~
,~ ~
5:
l
~
l
~:
l
diatom-
based film
(positive control)
1%
108 -'~
107 ~
-~ o 106 ~
~ .* 105 ~ o ~:l
104 ~i::
o * 103 ~cso
~1 102 ~
~ 10 o ~ ee
.-~l 1
5% 10% 0.1 N 1.0 N 5% glutar-
EtOH HCI aldehyde
Fig. 4.5. Larval metamorphosis ofA, crassispina in response to
diatom-based film treated with EtOH (1%, 5% and
10%), HCI (0.1 N and 1.0 N) and glutaraldehyde (5%).
Error bars indicate -+ SD of eighteen replicates.
[] : bacterial density; I : diatom density; horizontal
line: O% metamorphosis
54

l~ ~~ee
,~
O ~:l
~1
O ~ O ~:
100
80
60
40
20
O
~ ~~:ll! Il!
pt~iPl ~i~i~
~l 'i
~" ~i
pi~
l#~~*
ji~
pt!~ pii~~~ ~
~i!c
~iii:~gi ~li~~,j 'i
lll~i~,
~1 j~~ ~~
jpi '
diatom-based f ilm
LCA SBA WGA
108 ~,_,
'~
107 ~
~ ~ o 60 10 -~ 105 ~ o '1'
104 ~i.*
h o 1. 103 oc:'
~) ~1
102 ~ ~
10 o ~ 's
.-C:I 1
(positive control) '
Fig. 4.6. Larval metamorphosis of P depressus in response to
diatom-based films treated with lectins (LCA, SBA
and WGA). Error bars indicate :b SD of nine replicates.
[] : bacterial density, I : diatom density
55

'~. ~~ee
~' aaO
~:i
el
O ~ Q) ~:
1 OO
80
60
40
20
O
diatom-based f ilm
(positive control)__
~ l!Ii
llil~:
i
ppi~ li l~~
jllil
j ~~! ~ ~1lpi=
l
'. jil
i~:
~i~: l~~ jipii l.r
t
i
piii~ pti
d' '
jlll~1i
~
LCA S BA WGA
1 08
~'~ '~
107 ~
~ 106 ,JoQ)
~ ~ 10s '~
o 11 104 ~!,~
_o 103 cso
~ lc;
102 ~
S 10 o ~ Ge
.-1 C:I
Fig. 4.7. Larval metamorphosis ofA. crassispina in response to
diatom-based films treated with lectins (LCA, SBA
and WGA). Error bars indicate :t SD of nine replicates.
[] : bacterial density, I : diatom density
56

IV - 4 Discussion
Diatom-based films have been used to induce larval metamorphosis
during the mass-production of sea urchins juveniles (Ito et al. , 1991;
Kawahara, 1996). In this section, preliminary study was done to
investigate the nature of the chemical cue in diatom-based films on larval
metamorphosis of Pseudocentrotus depressus and Anthocidaris
crassl spma .
The larval response of P depressus and A. crassispina towards
laboratory-grown diatom-based films treated with heat (Fig. 4.2, 4.3),
ethanol (EtOH), hydrochloric acid (HCl) and glutaraldehyde (Fig. 4.4,
4.5) clearly shows that the cue is susceptible to degradation.
Glutaraldehyde, HCI and heat (above 40'C) treatments all completely
destroyed the. metamorphosis-inducing activity of diatom-based films.
This suggests that the live films possess an unstable cue that was easily
inactivated by heat. By contrast, Iectin treatment did not affect the
metamorphosis-inducing activity of diatom-based films (Fig. 4.6, 4.7),
which suggests that sugar-related compound do not play a main role in
the metamorphosis.
Metamorphosis-inducing compounds previously reported for sea
urchins are sugar floridoside and isethionic acid (Williamson et al. ,
2000), eicosapentaenoic acid (Kitamura et al. , 1993) and
glycoglycerolipids (Takahashi et al., 2002), which are relatively stable at
57

400C. Thus,the cues in diatom-based film may be difOerent from
pleviously reported ones.Dibromomethane(Taniguchiαα乙,1994)is
a possible candidate as an hlducing cue,but a high concentration(2,200
to3,800ppm)is needed to induce larval metamorphosis of these two
species of sea urchin(Kohα‘zJ.,1996).
58

CIIAPTER - V
GENERAL DISCUSSION AND CONCLUSIONS
In Japan, diatom-based film had been used in aquaculture to
mass-produce juveniles of sea urchin (Kawahara, 1996), sea cucumber
(Ito and Kitamura, 1997) and abalone (Kawamura and Kikuchi, 1992;
Kawamura, 1996) to sustain the wild stock. Mass-production of
economically important species of sea urchin juveniles (Anthocidaris
crassispina , Hemicentrotus pulcherrimus, Pseudocentrotus depressus, S.
intermedius, S. nudus) is being done annually (Kitamura et al. , 1993) by
inducing metamorphosis of competent larvae using plastic plates
("nami-ita") covered with periphytic diatoms. These studies indicated
the importance of diatom-based films in larval metamorphosis of sea
urchin, though the role of periphytic diatom and bacteria co-existing in
the film remains unknown.
In the present study; the metamorphosis-inducing activity of
microbial films formed in the sea and diatom-based films formed in
periphytic diatoms culture tanks on the larval metamorphosis of two
species of sea urchins P depressus and A. crassispina were investigated
(Chapter - II). Attempts were also made to separately evaluate the
metamorphosis-inducing activity of diatoms and bacteria from the
laboratory-cultured diatom-based films (Chapter - 111). Moreover, a
59

preliminary study was done to understand the nature of the chemical cue
in diatom-based films on larval metamorphosis of these two sea urchins
(Chapter - IV).
As has been detailed in Chapter - II, microbial films in the sea
showed that metamorphosis of P depressus larvae increased gradually
with immersion period, whereas the larval metamorphosis of A.
crassispina showed a bell-shaped response curve. Difference in larval
response may be influenced by the film components and the growth rate
of film, which reached the peak within a shorter period during spring and
summer (A. crassispina) than during the autumn and winter season (P
depressus). I hypothesize that P depressus larvae may also conform to
the bell-shaped response curve if the immersion period is extended
beyond the 25-day period tested.
In the tank films, metamorphosis of P depressus larvae increased
linearly with diatom density ranging from 2.0xl04 to 6.0xl05 cells cm~ ,
although the diatom-based films showed low metamorphosis-inducing
activity for larvae of A. crassispina. This pattern for P depressus
coincided with the data of Ito et al. (1991). These results also suggest
that the periphytic diatoms in the tank film could promote the larval
metamorphosis of P depressus, but may be less important in A.
crassispina (Chapter - II).
A comparison of results of larval assays done simultaneously in
May indicates that P depressus showed higher metamorphosis than A.
60

crassispina in response to diatom-based films in the tank. This
difference may be related to their natural habitats. P depressus
juveniles are found in deep (8-12 m) rocky subtidal areas (Imai and Arai,
1994), whereas A. crassispina live in shallow (0.5-4.0 m) intertidal field
dominated with foliose coralline algae (Imai, 1980a; 1980b; Tsuji et al. ,
1989). In other words, the microbial films on rock surfaces act as
metamorphosis inducer for larvae of P depressus, while the cue from
algae induces larval metamorphosis ofA. crassispina (Chapter - II).
In the Chapter - 111, diatom-based films grown under laboratory
conditions also induced larval metamorphosis of both sea urchins.
Furthermore, diatoms on filters induced metamorphosis in larvae of both
species, even though activity significantly decreased as compared to the
control group. By contrast, the bacterial films (filtrate) did not induce
metamorphosis in larvae of both species, suggesting that bacteria alone
cannot induce larval metamorphosis and that diatoms play a major role
in the induction of larval metamorphosis in sea urchins. However, it
should be noted that metamorphosis-inducing activity of the diatoms on
filters significantly decreased. It can be assumed that either a
metamorphosis-enhancing factor in diatom-based film passed through
the filter or a difference in surface configuration of diatom films between
those grown on beakers and those collected on filter may have affected
results.
It was also found that the antibiotic mixture (penicillin and
61

streptomycin) during larval assay destroyed or erased the
metamorphosis-inducing activity in diatom-based films, even though
bacterial density was not significantly decreased. Hence, the result
suggests that antibiotic mixture could have affected the quality of
diatom-based film community (Chapter - 111).
The larvae of both sea urchins showed similar response towards the
diatom-based film cultured with germanium dioxide and antibiotic
mixture. Reduction in bacterial density had a greater effect on the
metamorphosis-inducing activity of films than did reduction in diatotn
density, with a drastic decrease of 80% to 90% in the former as
compared to 35% to 60% in the latter. All five isolated species of
periphytic diatoms did not induce metamorphosis of both sea urchins
larvae. This suggests that no single diatom species is directly involved
as an inducer of metamorphosis in sea urchin larvae (Chapter - 111).
As has been noted above, diatom-based films cultured in the
laboratory induced larval metamorphosis in both P depressus and A.
crassispina. However, response of larvae to the diatom-based films
differed between the two species in that higher metamorphosis was
observed in P depressus than A. crassispina . This observation suggests
that diatom-based films formed in the sea urchin juvenile
mass-production tanks could promote larval metamorphosis in P
depressus but were less important in A. crassispina . Based on my study,
it is possible to hypothesize that the mixture of diatoms and bacteria
62

provide a synergistic effect for metamorphosis-inducing activity of
diatom-based film (Chapter - 111).
Laboratory-cultured diatom-based films induced larval
metamorphosis and a preliminary study was done to investigate the
nature of the metamorphosis-inducing chemical cue in diatom-based
films for P depressus and A. crassispina (Chapter - IV). The cue was
found to degrade when treated with heat (40'C), 5%- and 10%-ethanol,
0.1 N hydrochloric acid and 5 % glutaraldehyde. These results suggest
that the films possess an unstable chemical cue that was very degradable
in heat and chemically unstable. By contrast, Iectin treatment did not
affect the metamorphosis-inducing activity of diatom-based films,
suggesting that the cue is not a sugar-related compound. Previously
reported metamorphosis-inducing compounds for sea urchins were a
complex of the sugar floridoside and isethionic acid (Williamson et al. ,
2000), eicosapentaenoic acid (Kitamura et al. , 1993), and
glycoglycerolipids (Takahashi et al., 2002); all of them are relatively
stable at 40'C. Treatment of diatom-based films by ethanol (5% and
10%) may have affected the activity of diatoms and bacteria, and thus
resulting in the decrease in its metamorphosis-inducing activity.
Dibromomethane (Taniguchi et al., 1994) may be a possible candidate as
inducing cue, but high concentration was needed to induce larval
metamorphosis of these two species of sea urchins (Koh et al., 1996).
Thus, the cues in diatom-based film may be different from previously
63

reported ones.
Based on the overall findings in the present study, the microbial
films formed in the sea triggered laryal metamorphosis of P depressus
and A. crassispina although the pattern of larval response differed. In
diatom-based film formed in tanks, metamorphosis of P depressus
larvae increased linearly with diatom density; but showed low inducing
activity for larval metamorphosis of A. crassispina . The difference in
larval response of these species may be related to their natural habitat.
It is also possible to hypothesize that the mixture of diatoms and bacteria
provides a synergistic effect for the metamorphosis-inducing activity of
diatom-based film since bacteria alone or isolated periphytic diatoms
cannot induce larval metamorphosis. It is suggested that live films
possess an unstable cue(s) that was easily inactivated by heat and
sugar-related compounds do not play a main role in the metamorphosis.
The roles of periphytic diatoms and co-existing bacteria in
diatom-based film in the induction of larval metamorphosis require
further investigation. Moreover, the identification of the chemical
structure of the cues and the receptor sites in the larva are important for
understanding the mechanism and chemical ecology of metamorphosis
in sea urchin larvae. The findings reported here provide initial
information on the role of microbial films formed in the sea,
diatom-based films in periphytic diatom culturing tanks and in the
laboratory, on induction of larval metamorphosis in two species of sea
64

皿chins,R塵ρr8ssμs and/i1.c7α55麹ρ∫nα. These in£ormation may
improve the techniques of juveniles mass-production of fisheries
important species of ma血e invertebrates especially sea皿chins and
other echinoderms.
65

REFERENCES
Barker MF (1977) Observation on the settlement of the branchiolaria
larvae of Stichaster australis (verrill) and Coscinasterias calamaria
(Gray) (Echinodermata: Asteroidea) in the laboratory and on the
shore. J Exp Mar Biol Ecol 30: 95-l08
Barker MF, Nichols D (1983) Reproduction, recruitment and juvenile
ecology of the starfish, Asterias rubens and Marthasterias glacialis.
J Mar Biol Assoc UK 63: 745-765
Bhosle NB, Wagh AB (1997) Alnino acids in biofilm material on
aluminium panels immersed in marine waters. Biofouling 11(2):
149-166
Brancato MS, Woollacott RM (1982) Effects of microbial films on
settlement of bryozoan larvae (Bugula simplex, B. stolonifera and B.
turrita). Mar Biol 71: 51-56
Cameron RA, Hinegardner RT (1974) Initiation of metamorphosis in
laboratory cultured sea urchins. Biol Bull 146: 335-342
Cameron RA, Schroeter SC (1980) Sea urchin recruitment: effect of
substrate selection on juvenile distribution. Mar Ecol Prog Ser 2:
243 -247
Chen CP, Run JQ (1989) Larval growth and bacteria-induced
metamorphosis of Arachnoides placenta (L.) (Echinodermata:
Echinoidea). In: Ryland JS, Tyler PA (eds) Reproduction, genetics
66

and distributions of marine organisms. Olsen & Olsen,
Fredensborg, Denmark, pp. 55-59
Costerton JW, Lawandowski Z, Caldwell DE, Korber DR, Lappin-Scott
HM (1995) Microbial films. Annu Rev Microbiol 49: 711-745
Crisp DJ (1974) Factors influencing settlement of marine invertebrate
larvae. In: Grant PT, Mackie AM (eds) Chemoreception in marine
organisms. Academic Press, London, pp 177-265
Crisp DJ, Walker G, Young GA, Yule AB (1985) Adhesion and substrate
choice in mussels and barnacles. J Colloid and Interf Sci 104(1):
40-50
Imai T (1980a) On the sea urchins off Miura city. The study of
distribution, environment, growth and gonad in Jogashima. Bull
Kanagawa Pref Exp Stn I : 35-79 (in Japanese)
Imai T (1980b) On the sea urchins off Miura city! The study of
distribution, environment, growth and gonad from Hatsuse to
Kamimiyata. Bull Kanagawa Pref Exp Stn 2: 27-36 (in Japanese)
Imai T, Arai S (1994) Local pecurialities as living for the red sea urchin,
Pseudocentrotus depressus of bishamon waters off southern Miura
Peninsula, Kanagawa Prefecture, Japan. Suisanzoshoku 42:
307-3 13 (in Japanese with English abstract)
Ito S, Kitamura H (1997) Induction of larval metamorphosis in the sea
cucumber, Stichopus japonicus by periphytic diatoms .
Hydrobiologia 358: 281-284
67

Ito S, Kobayakawa J, Tani Y, Nakamura N (1991) The combined effect
of Hizikia fuslformis with attaching diatoms for larval settlement
and metamorphosis of Hemicentrotus pulcherrimus and
Pseudocentrotus depressus. Saibaigiken 19: 61-66 (in Japanese)
Ito Y (1984) The effect of benthic diatom on the acceleration of
metamorphosis of sea urchin larva. Mar Fouling 5: 15-18 (in
Japanese)
Johnsons CR, Sutton DC (1994) Bacteria on the surface of crustose
coralline algae induce the metamorphosis of crown-of-thorns
starfish. Mar Biol 120: 305-310
Johnsons CR, Sutton DC, Olson RR, Giddins R (199la) Settlement of
crown-of-thorns starfish: role of bacteria on surfaces of coralline
algae and a hypothesis for deepwater recruitment. Mar Ecol Prog
Ser 71: 143-162
Johnsons CR, Muir DG, Reysenbach AL (199lb) Characteristic bacteria
associated with surfaces of coralline red algae: a hypothesis for
bacterial induction of marine invertebrate larvae. Mar Ecol Prog
Ser 74: 281-294
Johnsons CR, Lewis TE, Nichols DS, Degnan BM (1997) Bacterial
induction of settlement and metamorphosis in marine invertebrates.
Proc 8th Int Coral Reef Sym 2: 1219-1224
Kawahara I (1996) Seed production of sea urchins (Pseudocentrotus
depressus and Hemicentrotus pulcherrimus). In: Saga Prefectural
68

Fish Farming Center (eds) Manual of Saga Prefectural Fish Farming
Center for seed production. Saga, pp 45-68 (in Japanese)
Kawamura T (1996) The role of benthic diatoms in the early life stages
of the Japanese abalone (Haliotis discus hannai). In: Watanabe Y,
Yamashita Y, Oozeki Y (eds) Survival strategies in early life stages
of marine resources. Balkema ~~, Rotterdam, pp 355-367
Kawamura T, Kikuchi S (1992) Effect of benthic diatoms on settlement
and metamorphosis of abalone larvae. Suisan-Zoshoku 40:
403-409 (in Japanese)
Keough MJ, Raimondi PT (1995) Responses of settling invertebrate
larvae to bioorganic films: effects of different types of films. J
Exp Mar Biol Ecol 185: 235-253
Kirchman D, Graham S, Reish D, Mitchell R (1982) Bacteria induce
settlement and metamorphosis of Janua (Dexiospira) brasiliensis
Grube (Polychaeta: Spirobidae). J Exp Mar Biol Ecol 56: 153-163
I~:tamura H, Hirayama K (1984) Suitable food plankton for growth of a
bryozoan Bugula neritina under laboratory conditions. Bull Jpn
Soc Sci Fish 50(6): 973-977
Kitamura H, Ito S, Tominaga S (2000) Relationship between the density
of benthic diatoms and the amount of chlorophyll-a on plastic plates
in sea urchin & sea cucumber tanks. Sessile Organisms 16(2):
21-25 (in Japanese)
Kitamura H, Kitahara S, Hirayama K (1992) Lipophilic inducers
69

extracted from extracted from Corallina pilullfera for larval
settlement and metamorphosis of two sea urchins (Pseudocentrotus
depressus and Anthocidaris crassispina). Nippon Suisan Gakh 58:
75-78
Kitamura H, Kltahara S, Koh HB (1993) T~le induction of larval
settlement and metamorphbsis of two sea urchins, Pseudocentrotus
depressus and Anthocidaris crassispina, by free fatty acids extracted
from the coralline red alga Corallina pilullfera . Mar Biol 1 15:
387-392
Koh HB, Tsuchida T, Kltamura H, Hirayama K (1996) Effect of
dibromomethane on the larval metamorphosis of the sea urchins,
Anthocidaris crassispina and Pseudocentrotus depressus. Sessile
Organisms 13(1): 7-9 (in Japanese)
Lau SCK, Qian PY (2001) Larval settlement in the serpulid polychaete
Hydroides elegans in response to bacterial films: an investigation of
the nature of putative larval settlement cue. Mar Biol 138:
321-328
Leitz T, Wagner T (1993) The marine bacterium Alteromonas espejiana
induces metamorphosis of the hydroid Hydractinia echinata . Mar
Biol 115: 173-178
Loeb GI, Neihof RA (1975) Marine conditioning film. Adv Chem Ser
145: 319-335
Maki JS, Rittschof D, Costlow JD, Mitchell R (1988) Inhibition of
70

attachment of larval barnacles, Balanus amphitrite, by bacterial
surface films. Mar Biol 97: 199-206
Mercier A, Battaglene SC, Hamel J-F (2000) Settlement preferences and
early migration of the tropical sea cucumber Holothuria scabra . J
Exp Mar Biol Ecol 249: 89-110
h4itchell R, Klrchman D (1984) The microbial ecology of marine
surfaces. In: Costlow JD, Tipper RC (eds) Marine
biodeterioration: an interdisciplinary study. Naval Institutes P, ress,
Annapolis, MD, pp. 49-56
Naidenko TK (1996) Induction of metamorphosis of two species of sea
urchin from Sea of Japan. Mar Biol 126: 685-692
Pawlik JR (1992) Chemical ecology of the settlement of benthic marine
invertebrates. Oceanogr Mar Biol Annu Rev 30: 273-335
Pearce CM, Scheibling RE (1990) Induction of metamorphosis of larvae
of the green sea urchin, Strongylocentrotus droebachiensis, by
coralline red algae. Biol Bull 179: 304-311
Pearce CM, Scheibling RE (1991) Effect of macroalgae, microbial films,
and conspecifics on the ifiduction of metamorphosis of the green sea
urchin Strongylocentrotus droebachiensis (Mtiller). J Exp Mar
Biol Ecol 147: 147-162
Pechenik JA, Rittschoff D, Schmidt AR (1993) Influence of delayed
metamorphosis on survival and growth of juvenile bamacles
Balanus amphitrite. Mar Biol 115: 287-294
71

Rahim SAKA, Li J-Y, Kitamura H (2004) Larval metamorphosis of the
sea urchins, Pseudocentrotus depressus and Anthocidaris
crassispina in response to microbial films. Mar Biol 144: 71-78
Rahim SAKA, Li J-Y, Satuito CG, Kitamura H (2004) The role of
diatom-based film as inducer of metamorphosis in larvae of two
species of sea urchins, Pseudocentrotus depressus and Anthocidaris
crassispina. Sessile Organisms 21(1): (in press)
Robert RD, Nicholson CM (1997) Variable response from abalone larvae
(Haliotis iris, H. virginea) to a range of settlement cues. Moll Res
18: 131-141
Rowley RJ (1989) Settlement and recruitment of sea urchins
(Strongylocentrotus spp.) in a sea urchin barren ground and kelp
bed: are population regulated by settlement or post-settlement
processes? Mar Biol 100: 485-494
Satuito CG, Natoyama K, Yamazaki M, Fusetani N (1995) Induction of
attachment and metamorphosis of laboratory cultured mussel
Mytilus edulis galloprovincialis larvae by microbial film. Fish Sci
61(2): 223-227
Slattery M (1992) Larval settlement and juvenile survival in the red
abalone (Haliotis rufescens): an examination of inductive cues and
substrate selection. Aquaculture 102: 143-153
Snelgrove PVR, Grassle JP, Grassle JF, Petrecca RF. Ma H (1999) Notes:
In situ habitat selection by settling larvae of marine soft-sediment
72

invertebrates. Limnol Oceanogr 44(5): 1341-1347
Takahashi Y, Itoh K, Ishii M, Suzuki M, Itabashi Y (2002) Induction of
larval settlement and metamorphosis of the sea urchin
Stron8ylocentrotus intermedius by glycoglycerolipids from the
green alga Ulvella lens. Mar Biol 140: 763-771
Tani Y, Ito Y (1979) Effects of benthic diatoms on settlement and
metamorphosis of the sea urchin, Pseudocentrotus depressus.
Suisanzoshoku 27: 148-150 (in Japanese with English abstract)
Taniguchi K, Kurata K, Maruzoi T, Suzuki M (1994) Dibromomethane, a
chemical inducer of larval settlement and metamorphosis of the sea
urchin Strongylocentrotus nudus. Fish Sci 60: 795-796
Tsuji S, Yoshiya M, Tanaka M, Kuwahara A, Uchino K (1989) Seasonal
changes in distribution and ripeness of gonad of a sea urchin
Strongylocentrotus nudus in the western part of Wakasa Bay! Bull
Kyoto Ocean Fish Sci 12: 15-21 (in Japanese)
Tsurumi K, Fusetani N (1998) Effects of early fouling communities
formed in the field on settlement and metamorphosis of cyprids of
the barnacle, Balanus amphitrite Darwin. Biofouling 12: 119-131
Unabia CRC, Hadfield MG (1999) Role of bacteria in larval settlement
and metamorphosis of the polychaete ~:ydroides elegans. Mar
Biol 133: 55-64
Wieczorek SK, Todd CD (1998) Inhibition and facilitation of settlement
of epifaunal marine invertebrate larvae by microbial biofilm cues.
73

Biofouling 12: 81-118
Williamson JE, Nys RD, Kumar N, Carson DG, Steinberg PD (2000)
Induction of metamorphosis in sea urchin Holopneustes
purpurascens by a metabolites complex from the algal host Delisea
pulchra. Biol Bull 198: 332-345
Yazaki I (1995) Quantitative analysis of metamorphosis induced by
L-glutamine in embryos of the sea urchin, Hemicentrotus
pulcherrimus. Zool Sci 12: 105-112
Yazaki I, Harashima H (1994) Induction of metamorphosis in the sea
urchin, Pseudocentrotus depressus, using L-glutamine. Zool Sci
11: 253-260
Zobell CE, Allen EC (1935) The significance of marine bacteria in the
fouling of submerged surfaces. J Bacteriol 29: 239-251
74