neural coding: hybrid analog and digital signalling in axons
TRANSCRIPT

DispatchR585
Neural Coding: Hybrid Analog andDigital Signalling in Axons
Mammalian axons are thought to act as digital signaling devices,conveying information only by the timing and rate of all-or-none actionpotentials. Two recent studies now show that synaptic potentials canalso spread far down the axon and influence action potential-triggeredtransmitter release in a graded, ‘analog’ manner. Axons thus encodeinformation both about subthreshold and suprathreshold synapticactivity.
Beverley Clarkand Michael Hausser
Axons connect neurons. They aresignalling devices, transmittingaction potentials which triggerneurotransmitter release fromsynaptic boutons along the axon.In the classical view, synaptic inputto the soma and dendrites ofa neuron is funnelled into the axon,and at the initiation site — in theaxon initial segment or first node ofRanvier [1–3] — a decision is madeto fire an action potential oncea threshold level of depolarizationis exceeded. As the action potentialis an all-or-none signal, it can beviewed as a ‘digital’ signal, with itsinitiation thus representing ananalog-to-digital conversion. Theaction potential is then transmitteddown the axon to the synapticboutons, where it serves as thenecessary trigger for synapticrelease. Action potentialpropagation is thought to bequite reliable, making axonsrelatively uninteresting in signalprocessing terms, acting assimple point-to-point relays.Furthermore, in this view, theinformation transmitted by axonsis solely encoded by the numberand timing of action potentialsin the axon.
Two recent studies [4,5] havechallenged this view. Usingsophisticated techniques to makewhole-cell patch-clamp recordingsdirectly from the axons of neuronsin brain slices, Alle and Geiger [4]and Shu et al. [5] have shown, intwo different neuronal types, that,in addition to action potentials,axons can transmit subthresholdsynaptic potentials surprisinglyefficiently (Figure 1), and that thisspread of subthreshold potentialscan have a significant impact on
synaptic transmission triggeredby action potentials.
Alle and Geiger [4] recordeddirectly from mossy fiber boutons[6], several hundred microns fromthe soma of dentate gyrus granulecells. They found that activatingsynaptic input to the granule cellsresulted in large subthresholdexcitatory postsynaptic potentials(EPSPs) in the mossy fiberboutons, with EPSP amplitudesthat were graded with stimulationintensity. They demonstrated bylocal application of blockers ofsynaptic transmission that theseaxonal EPSPs, which they calledexcitatory presynaptic potentials,or EPreSPs, are propagatedfrom the soma and not locallygenerated in the axon. Thus, the
axon must be capable of efficienttransmission of subthresholdEPSPs down the axon. Bymeasuring the decay of EPreSPamplitude as a function of distancefrom the soma, they estimatedthat the space constant — l, thedistance at which the voltagedecays to 37% of its originalvalue — for propagation ofEPreSPs is 430 mm.
Shu et al. [5] used a novelapproach to record intracellularlyfrom axons. They noticed that thecut end of unmyelinated pyramidalcell axons in brain slices oftenforms a membrane ‘bleb’ which islarge enough to permit whole-cellpatch-clamp recording. Bysimultaneously recording from thesoma and such axon blebs, theyshowed that subthresholdmembrane potentials at the somaspread efficiently down the axon. Inparticular, spontaneous barragesof synaptic activity triggered bynetwork ‘up’ states [7] could beobserved hundreds of micronsdown the axon (Figure 1), witha space constant l of 417 mm. Thespace constant values obtainedby the two studies are surprisinglylarge given the fine calibre ofpyramidal cell and granule cell
Soma
Axon
Single EPSP Network activity
100 µm
100 ms20 ms
Current Biology
Figure 1. Synaptic potentials spread efficiently down the axon.
A schematic representation of a mammalian central neuron showing a simultaneousrecording at the soma and several hundred microns down the axon. Single EPSPs(left) and depolarizations due to synchronous network activity (right) propagate effi-ciently to the axonal recording site and thus can modulate action potential-driven neuro-transmitter release.
Current Biology Vol 16 No 15R586
Analog Digital
Pre
Post
Hybrid
Current Biology
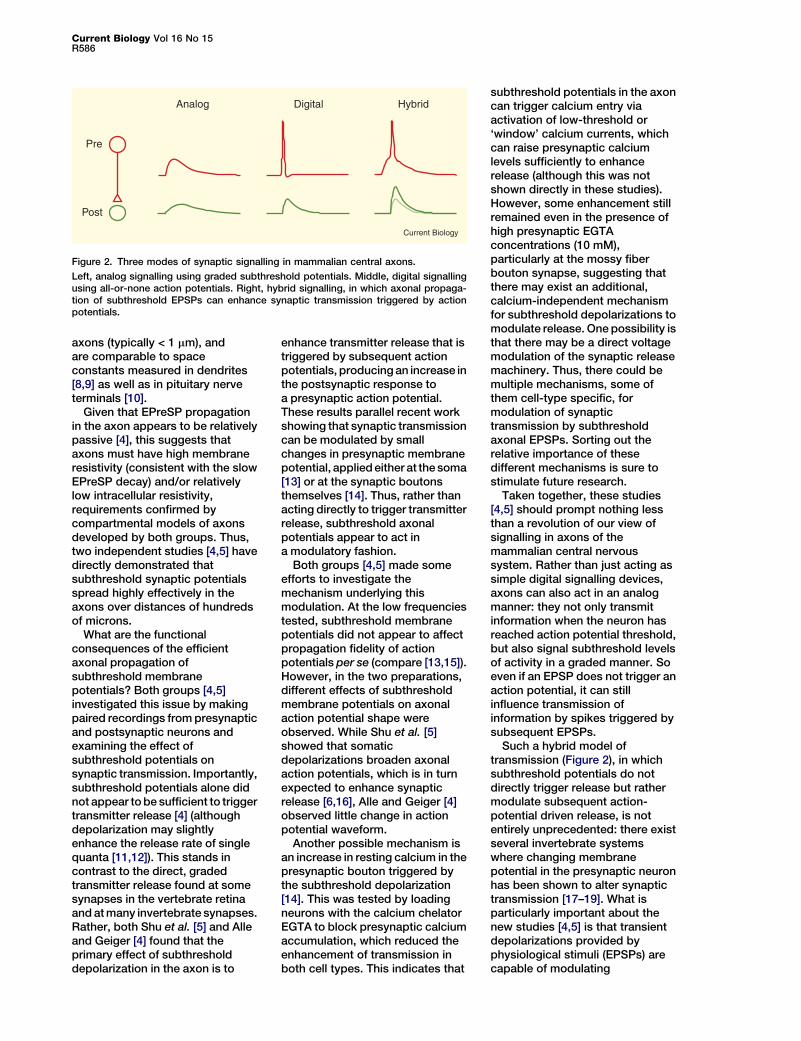
Figure 2. Three modes of synaptic signalling in mammalian central axons.
Left, analog signalling using graded subthreshold potentials. Middle, digital signallingusing all-or-none action potentials. Right, hybrid signalling, in which axonal propaga-tion of subthreshold EPSPs can enhance synaptic transmission triggered by actionpotentials.
axons (typically < 1 mm), andare comparable to spaceconstants measured in dendrites[8,9] as well as in pituitary nerveterminals [10].
Given that EPreSP propagationin the axon appears to be relativelypassive [4], this suggests thataxons must have high membraneresistivity (consistent with the slowEPreSP decay) and/or relativelylow intracellular resistivity,requirements confirmed bycompartmental models of axonsdeveloped by both groups. Thus,two independent studies [4,5] havedirectly demonstrated thatsubthreshold synaptic potentialsspread highly effectively in theaxons over distances of hundredsof microns.
What are the functionalconsequences of the efficientaxonal propagation ofsubthreshold membranepotentials? Both groups [4,5]investigated this issue by makingpaired recordings from presynapticand postsynaptic neurons andexamining the effect ofsubthreshold potentials onsynaptic transmission. Importantly,subthreshold potentials alone didnot appear to be sufficient to triggertransmitter release [4] (althoughdepolarization may slightlyenhance the release rate of singlequanta [11,12]). This stands incontrast to the direct, gradedtransmitter release found at somesynapses in the vertebrate retinaand at many invertebrate synapses.Rather, both Shu et al. [5] and Alleand Geiger [4] found that theprimary effect of subthresholddepolarization in the axon is to
enhance transmitter release that istriggered by subsequent actionpotentials, producing an increase inthe postsynaptic response toa presynaptic action potential.These results parallel recent workshowing that synaptic transmissioncan be modulated by smallchanges in presynaptic membranepotential, applied either at the soma[13] or at the synaptic boutonsthemselves [14]. Thus, rather thanacting directly to trigger transmitterrelease, subthreshold axonalpotentials appear to act ina modulatory fashion.
Both groups [4,5] made someefforts to investigate themechanism underlying thismodulation. At the low frequenciestested, subthreshold membranepotentials did not appear to affectpropagation fidelity of actionpotentials per se (compare [13,15]).However, in the two preparations,different effects of subthresholdmembrane potentials on axonalaction potential shape wereobserved. While Shu et al. [5]showed that somaticdepolarizations broaden axonalaction potentials, which is in turnexpected to enhance synapticrelease [6,16], Alle and Geiger [4]observed little change in actionpotential waveform.
Another possible mechanism isan increase in resting calcium in thepresynaptic bouton triggered bythe subthreshold depolarization[14]. This was tested by loadingneurons with the calcium chelatorEGTA to block presynaptic calciumaccumulation, which reduced theenhancement of transmission inboth cell types. This indicates that
subthreshold potentials in the axoncan trigger calcium entry viaactivation of low-threshold or‘window’ calcium currents, whichcan raise presynaptic calciumlevels sufficiently to enhancerelease (although this was notshown directly in these studies).However, some enhancement stillremained even in the presence ofhigh presynaptic EGTAconcentrations (10 mM),particularly at the mossy fiberbouton synapse, suggesting thatthere may exist an additional,calcium-independent mechanismfor subthreshold depolarizations tomodulate release. One possibility isthat there may be a direct voltagemodulation of the synaptic releasemachinery. Thus, there could bemultiple mechanisms, some ofthem cell-type specific, formodulation of synaptictransmission by subthresholdaxonal EPSPs. Sorting out therelative importance of thesedifferent mechanisms is sure tostimulate future research.
Taken together, these studies[4,5] should prompt nothing lessthan a revolution of our view ofsignalling in axons of themammalian central nervoussystem. Rather than just acting assimple digital signalling devices,axons can also act in an analogmanner: they not only transmitinformation when the neuron hasreached action potential threshold,but also signal subthreshold levelsof activity in a graded manner. Soeven if an EPSP does not trigger anaction potential, it can stillinfluence transmission ofinformation by spikes triggered bysubsequent EPSPs.
Such a hybrid model oftransmission (Figure 2), in whichsubthreshold potentials do notdirectly trigger release but rathermodulate subsequent action-potential driven release, is notentirely unprecedented: there existseveral invertebrate systemswhere changing membranepotential in the presynaptic neuronhas been shown to alter synaptictransmission [17–19]. What isparticularly important about thenew studies [4,5] is that transientdepolarizations provided byphysiological stimuli (EPSPs) arecapable of modulating

DispatchR587
transmission. This suggests thatthe timing relationship of EPSPsand succeeding action potentialswill be critical [4], favouringpotentiation of transmission inresponse to facilitating synapticinputs. It also providesa mechanism for short-termmemory of recent activity, wheresynaptic release depends on therecent history of synaptic input tothe neuron.
This new view has severaladditional interesting functionalimplications. EPSPs attenuate asthey spread down the axon. Shuet al. [5] performed an anatomicalanalysis showing that more thanone hundred synapses made bythe axon of cortical pyramidalneurons are within a spaceconstant of the soma and thus willbe affected by subthreshold axonalmodulation. However, themagnitude of any effect ofsubthreshold depolarization onsynaptic transmission will dependon the distance of the individualsynaptic contacts from the soma:proximal synaptic contacts will bestrongly influenced, and distal onesless so. It is therefore likely thatthere exists mixed analog/digitalsignalling at proximal contacts,and purely digital signalling atdistal contacts. This may reflectdifferent functional roles ofproximal and distal connections.Proximal connections should beinfluenced more by the localnetwork context that generated thedepolarization, for exampleoscillations or other forms ofsynchrony, such that the axonaldepolarization represents a form ofpositive feedback for coordinatinglocal networks. On the otherhand, long-range connectionsexhibit entirely digital signalling,since local feedback is notnecessary.
There are many questions leftopen by these two remarkablycomplementary papers [4,5].Firstly, how general are thesefindings? Both studies recordedfrom the large, main trunk ofunmyelinated axons exhibitingrelatively small amounts ofbranching. In axons exhibitingmore branching (such as manyinterneuron axons), or in thinneraxons (particularly axon collaterals)attenuation of subthreshold
potentials should be much steeper,and so will permit less hybridsignalling. It also remains to bedetermined how well subthresholdpotentials can spread inmyelinated axons.
Secondly, can inhibitorypostsynaptic potentials (IPSPs)spread down the axon with similarefficacy to EPSPs, and if so, howdo IPSPs influence synapticrelease? Thirdly, how do axonalvoltage-gated channels influencepropagation? Alle and Geiger [4]showed that application of thesodium channel blockertetrodotoxin had only a relativelysmall effect on the EPreSPamplitude; however, thedependence on EPreSP amplitudewas not examined, and otherchannels may also contribute(particularly Ih cation channelsand also calcium channels, asexpected from the fact thatsubthreshold calcium entryappears to influence actionpotential-dependent release).
Fourthly, as action potentialpropagation itself can bemodulated by membrane potential[13,15], it is conceivable thatsynaptic potentials spreading inthe axon can either promote orinhibit propagation in differentregions of the axon. And lastly, ifaxo-axonic synapses are able totrigger presynaptic potentials, thepresent studies suggest thatthese potentials should be able tospread to other synapses toinfluence synaptic release, or evenretrogradely to influence actionpotential propagation or initiation.Such two-way traffic ofsubthreshold potentials in theaxon may have unexpectedconsequences for informationprocessing.
The new studies [4,5] shouldprovide a major stimulus to thegrowing recent change inperspective on axons as signallingdevices [20]. Rather than acting assimple, reliable transmission linesfor action potentials, axons havenow been shown to exhibit a richrepertoire of behaviour, in whichboth action potentials and synapticpotentials can be transmitted tosynapses, and can interact tomodulate synaptic release. Thissuggests that the action potentialinitiation site is no longer the final
site of synaptic integration, butthat action potentials and synapticpotentials continue to interactalong the length of the axon. Thenext step will be to understandthe computational significance ofthese interactions and how theycan be exploited to enhance thenormal functioning of themammalian central nervoussystem. Now that the axons ofmammalian neurons are finallybecoming accessible to directinvestigation using both imagingand electrophysiologicaltechniques, we can expect manymore breakthroughs from thesetiny structures.
References1. Coombs, J.S., Curtis, D.R., and
Eccles, J.C. (1957). The interpretation ofspike potentials of motoneurones.J. Physiol. 139, 198–231.
2. Clark, B.A., Monsivais, P., Branco, T.,London, M., and Hausser, M. (2005). Thesite of action potential initiation incerebellar Purkinje neurons. Nat.Neurosci. 8, 137–139.
3. Palmer, L.M., and Stuart, G.J. (2006).Site of action potential initiation in layer5 pyramidal neurons. J. Neurosci. 26,1854–1863.
4. Alle, H., and Geiger, J.R. (2006).Combined analog and action potentialcoding in hippocampal mossy fibers.Science 311, 1290–1293.
5. Shu, Y., Hasenstaub, A., Duque, A., Yu, Y.,and McCormick, D.A. (2006). Modulationof intracortical synaptic potentials bypresynaptic somatic membrane potential.Nature 441, 761–765.
6. Geiger, J.R., and Jonas, P. (2000).Dynamic control of presynaptic Ca(2+)inflow by fast-inactivating K(+) channels inhippocampal mossy fiber boutons.Neuron 28, 927–939.
7. Steriade, M., Timofeev, I., and Grenier, F.(2001). Natural waking and sleep states:a view from inside neocortical neurons.J. Neurophysiol. 85, 1969–1985.
8. Larkman, A.U., Major, G., Stratford, K.J.,and Jack, J.J. (1992). Dendriticmorphology of pyramidal neurones of thevisual cortex of the rat. IV: Electricalgeometry. J. Comp. Neurol. 323, 137–152.
9. Ulrich, D., and Stricker, C. (2000).Dendrosomatic voltage and chargetransfer in rat neocortical pyramidal cellsin vitro. J. Neurophysiol. 84, 1445–1452.
10. Jackson, M.B. (1993). Passive currentflow and morphology in the terminalarborizations of the posterior pituitary.J. Neurophysiol. 69, 692–702.
11. Glitsch, M., and Marty, A. (1999).Presynaptic effects of NMDA incerebellar Purkinje cells and interneurons.J. Neurosci. 19, 511–519.
12. Katz, B., and Miledi, R. (1965). Propagationof electric activity in motor nerve terminals.Proc. R. Soc. Lond. B 161, 453–482.
13. Debanne, D., Guerineau, N.C.,Gahwiler, B.H., and Thompson, S.M.(1997). Action-potential propagationgated by an axonal I(A)-like K+conductance in hippocampus. Nature389, 286–289.
14. Awatramani, G.B., Price, G.D., andTrussell, L.O. (2005). Modulation oftransmitter release by presynaptic restingpotential and background calcium levels.Neuron 48, 109–121.

Current Biology Vol 16 No 15R588
15. Monsivais, P., Clark, B.A., Roth, A., andHausser, M. (2005). Determinants ofaction potential propagation in cerebellarPurkinje cell axons. J. Neurosci. 25,464–472.
16. Borst, J.G., and Sakmann, B. (1999).Effect of changes in action potentialshape on calcium currents and transmitterrelease in a calyx-type synapse ofthe rat auditory brainstem. Philos.Trans. R. Soc. Lond. B 354,347–355.
17. Nicholls, J., and Wallace, B.G. (1978).Modulation of transmission at an
IKKe Signaling: No
IkB kinases (IKKs) are key componeninnate immunity and inflammation. Simplicate IKKs in Drosophila in seemnon-apoptotic caspase activation an
Andreas Bergmann
NF-kB transcription factors arekept inactive by cytoplasmicsequestration through complexformation with inhibitory IkBproteins [1]. NF-kB stimulation – forexample initiated by Toll-likereceptors in innate immunity orin response to proinflammatorycytokines such as TNFa – requiresdissociation of the NF-kB/IkBcomplex. IkB kinases (IKKs)were initially identified asa high-molecular weight complexcapable of site-specificphosphorylation of IkB-a [2]. Thisphosphorylation triggersubiquitin-mediated degradation ofIkB-a, and the release of NF-kBtranscription factors, whichtranslocate into the nucleus [2].Subsequent analysis identified twocatalytic subunits (IKKa and IKKb)and a structural component of thiscomplex (IKKg/NEMO). While theIKKa/b/g complex is required forNF-kB activation in response tomost NF-kB inducers, the role oftwo related kinases known asIKKe/IKKi and TBK1/NAK/T2K isless clear.
In Drosophila, two independentimmune signaling pathwayscontrol the activity of distinctNF-kB-like proteins [3]. Whilethe Toll/anti-fungal pathwayrequires Dorsal and Dif, theIMD/anti-bacterial pathway leadsto activation of Relish [3]. Dorsaland Dif are rendered cytoplasmic
inhibitory synapse in the central nervoussystem of the leech. J. Physiol. 281,157–170.
18. Shimahara, T., and Tauc, L. (1975).Multiple interneuronal afferents to thegiant cells in Aplysia. J. Physiol. 247,299–319.
19. Shapiro, E., Castellucci, V.F., andKandel, E.R. (1980). Presynapticmembrane potential affects transmitterrelease in an identified neuron in Aplysiaby modulating the Ca2+ and K+ currents.Proc. Natl. Acad. Sci. USA 77,629–633.
t Just NF-kB
ts of NF-kB signaling pathways inurprisingly, three recent reportsingly unrelated functions, includingd cytoskeleton organization.
in complex with the only IkB-likeprotein, termed Cactus.Phosphorylation of Cactus isrequired for its degradation [4–6],but the responsible kinase hasnot been identified. TheDrosophila genome encodes twoIKK genes. DmIKKb (or DLAK) ismost similar to human IKKb, andis involved in Relish activation[7]. That leaves the secondDrosophila IKK, DmIKKe (alsoknown as Ik2), as a candidate forthe Cactus kinase. However,recent reports [8,9], includingone in this issue of CurrentBiology [10], rule out a function ofDmIKKe as Cactus kinase.Instead, DmIKKe modulatescaspases for a non-apoptoticfunction and controls bothactin and microtubulecytoskeletons.
DmIKKe as a Negative Regulatorof Diap1 Protein StabilityAs in vertebrates, apoptosis inDrosophila is triggered byactivation of caspases, a highlyspecialized class of cell deathproteases. In surviving cells,caspases are kept inactive throughcomplex formation with inhibitorof apoptosis proteins (IAPs),most notably Drosophila IAP1(Diap1) [11]. In response tocell death-inducing signals,pro-apoptotic proteins such asReaper stimulate the ubiquitylationand degradation of Diap1,releasing caspases from IAP
20. Debanne, D. (2004). Informationprocessing in the axon. Nat. Rev.Neurosci. 5, 304–316.
Wolfson Institute for BiomedicalResearch and Department ofPhysiology, University College London,Gower Street, London WC1E 6BT, UK.E-mail: [email protected]
DOI: 10.1016/j.cub.2006.07.007
inhibition and triggering apoptosis[11]. Interestingly, the recent paperby Kuranaga et al. [9] identifiesmutations in DmIKKe
as dominant suppressors ofReaper-induced cell death [9].Subsequent analysis showed thatloss of DmIKKe increases thestability of the Diap1 protein,providing an explanation forthe observed suppression ofReaper-induced apoptosis [9].These observations suggest thatwild-type DmIKKe destabilizesDiap1, leading to Caspaseactivation, a conclusion whichwas confirmed in cell cultureexperiments and in transgenicflies.
Destabilization of Diap1 appearsto be the result ofphosphorylation by DmIKKe.Interestingly, human TBK1/NAK/T2K was able to promotephosphorylation and degradationof human XIAP [9], suggestingconservation ofIKKe-mediated control of IAPstability. DmIKKe-mediated Diap1degradation is independent ofReaper. Overexpression ofDmIKKe in cell death deficient(i.e. reaper mutant) backgroundstill induced Diap1 instability andapoptosis [9]. This is a strikingfinding, as it suggests that controlof Diap1 stability and thus caspaseactivation in Drosophila occursthrough distinct pathways,including the classical apoptoticpathway and as well as by IKKe
signaling.
DmIKKe Controls Diap1 in aNon-Apoptotic SettingDespite the fact thatoverexpression of DmIKKe inducesa strong apoptotic phenotype,developmental cell death appears