neurotrophic factors and their receptors - university of liverpool
TRANSCRIPT

Neurotrophic factors and their receptors
Mariano Barbacid
Bristol-Myers Squibb Pharmaceutical Research Institute, Princeton, USA
Several new members of the nerve growth factor family of neurotrophins,
which comprises nerve growth factor itself, brain-derived neurotrophic factor
and neurotrophins-3, -4 (also known as neurotrophin5) and -6, have been
isolated in recent years. Their signaling receptors have been identified as the
Trk family of tyrosine protein kinases, thus facilitating the dissection of the
signaling pathways responsible for mediating their trophic properties. More
recently, the advent of gene targeting has made it possible to generate strains
of mice lacking neurotrophins and their receptors. Analysis of the phenotypes
of these mutant animals has provided detailed information on the role that
neurotrophins and their receptors play in the ontogeny of the mammalian
nervous system.
Current Opinion in Cell Biology 1995, 7:148-l 55
Introduction
Growth factors play a critical role in the development and maintenance of the mammalian nervous system. Some of these factors (e.g. platelet-derived growth fac- tor and fibroblast growth factor) have pleiotropic ac- tivities and are often studied in systems other than the nervous system [l-3]. Other factors have a more restricted activity on neuronal cells and are classified as ‘neurotrophic factors’. Among them, the most in- tensively studied include the ciliary neurotrophic fac- tor (CNTF) [4], the recently discovered glial-derived neurotrophic factor (GDNF) [5*] and the members of the nerve growth factor (NGF) family, now known as ‘neurotrophins’ [6,7]. Each of these neurotrophic factors exerts its biological activities through a different class of receptors. Neurotrophins signal through the Trk family of tyrosine protein kinases [8-lo]. In addition, neuro- trophins interact with a second receptor, ~75, a member of the tumor necrosis factor receptor superfamily [ll]. CNTF signals though a multimeric receptor complex which includes binding and signaling elements [12,13]. The binding subunit is specific for CNTF and does not have transmembrane or cytoplasmic domains, but instead anchors to the plama membrane by a glycosyl phosphatidylinositol linkage [14]. The signaling subunits include gp130, a transmembrane glycoprotein previously found in various cytokine receptors [15], and the p-sub- unit of the receptor for leukemia inhibitory factor [16]. The cytoplasmic domains of both of these molecules interact with members of the Jak family of cytoplas- mic tyrosine kinases, which are likely to be the primary
mediators of CNTF activity [12,13]. CNTF shares these signaling subunits with a variety of cytokine receptors, including the receptors for leukemia inhibitory factor, interleukin-6, oncostatin M and, possibly, interleukin-1 1 [12,13]. How these growth factors can elicit such differ- ent biological responses by activating the same signaling receptor subunits is a subject of intense study at this time [12,13]. Finally, the receptor for GDNF has not as yet been identified; however, the limited structural similarities between GDNF and the transforming growth factor-p superfamily [5*] raise the possibility that GDNF may signal through multimeric serine/threonine kinase receptors.
During the past year, several important developments have occured in the field of neurotrophins and their receptors. The generation of strains of mice carrying targeted mutations in the genes encoding several neu- rotrophins, as well as in those encoding the ~75 and the Trk kinase receptors, has provided a wealth of infor- mation regarding the role that these molecules play in the ontogeny of the mammalian nervous system [17]. In consideration of the space limitations, this review will fo- cus on neurotrophins and their receptors. Other exciting developments regarding CNTF and its signal transduc- tion pathways have been recently summarized in several reviews [4,12,13] and will not be discussed here. GDNF, initially thought to be specific for dopaminergic neurons [5-l, appears to have a broader specificity, as it can also support the survival of motorneurons [18]. Further stud- ies are needed to define the full range of neurotrophic activities of this novel growth factor.
148
Abbreviations
BDNF-brain-derived neurotrophic factor; CNTF-ciliary neurotrophic factor; EGFGpidermal growth factor; GDNF-glial-derived neurotrophic factor; MAP-mitogen-activated protein; NCF-nerve growth factor;
NT-neurotrophin; PLC-phospholipase C.
0 Current Biology Ltd ISSN 0955-0674

Neurotrouhic factors and their receutors Barbacid 149
The NCF family of neurotrophins
This family of growth factors includes NGF, brain-de- rived neurotrophic factor (BDNF), neurotrophin (NT)-3 and NT-4 (also known as NT-4/5 and NT-5) [2]. A molecule recently isolated from the teleost fish Xiphopho- rus, NT-6, probably represents a novel member of this f&nily [19*]. These neurotrophins bind specifically to the three known Trk tyrosine kinase receptors, T&A, TrkB and TrkC [8-lo]. Whereas NGF binds to T&A, BDNF and NT-4 interact with TrkB (Fig. 1). NT-3 appears to be more promiscuous and binds to each of the Trk receptors, but its primary biological responses are me- diated by TrkC (see below). In addition, each of these neurotrophins binds with low affinity to ~75, a receptor structurally unrelated to the Trks and whose precise role in mediating neurotrophin activity remains to be eluci- dated (see below) [ll] (Fig. 1). Whether NT-6 binds to any of these receptors is not known at the present time.
NGF recruitment ?
Qo
NGF BDNF NT-4 NT-3
TrKA TrkB
Signal transduction
TrKC
0 1995 Current Opinion in Cell Biology
Fig. 1. Functional interactions between neurotrophins and their cog- nate receptors. Each of the neurotrophins binds with similarly low affinity (nanomolar range) to ~75. The neuronal defects observed in p75 null mice, however, suggest a functional interaction of this re- ceptor with NGF (solid arrow) but not with BDNF or NT-3 (dashed arrows). The interactions of NCF with TrkA and of BNDF and NT-4 with TrkB are highly specific. NT-3 primarily activates TrkC recep- tors; however, this neurotrophin can also interact with the TrkA and TrkB receptors (see text).
The three-dimensional structure of NGF has been re- solved by X-ray crystallography (at 2.3A resolution; [20]) and shown to consist of two identical subunits. Based on previous sequence analysis, each subunit has 118 amino acids residues and is generated by cleav- age of a precursor polypeptide of 252 residues [1,7]. The NGF protomers are elongated and contain three twisted anti-parallel p-sheet segments linked by turns. These protomers are linked to each other by a ‘cys- teine knot’, a novel structure also found in transform- ing growth factor-b and platelet-derived growth factor B-subunit homodimers, and display a twofold symmetry with parallel long axes [20]. The structure of the other neurotrophins remains to be elucidated; however, their close similarities to each other (47 residues are identical in all four mammalian neurotrophins) suggest that they are likely to have similar tertiary structures.
Site-directed mutagenesis studies have provided impor- tant information regarding those residues critical for the
interaction of neurotrophins with the p75 and TrkA receptors. The interaction of NGF with p75 is medi- ated by a positively charged region that consists of at least three lysine residues located in two spatially ad- jacent loops (I and V) [21]. These positively charged residues are conserved in NT-3 and NT-4, but not in BDNF, which contains three consecutive positively charged residues in loop V that may compensate for the absence of the two lysines present in loop I in the other neurotrophins [22]. These differences may ex- plain the somewhat distinct dissociation kinetics shown by BDNE
The interaction of NGF with TrkA is more complex and requires cooperativity of multiple contact sites de- rived horn at least five different domains within the NGF molecule [21,22]. These sites are concentrated on one side of the NGF dimer with both protomers contributing some of the critical residues. Overall, the TrkA binding site appears as a flat, continuous surface ap- proximately parallel to the twofold axis of the molecule. Because of its twofold symmetry, the active NGF dimers have two potential T&A binding sites which may facili- tate the formation of T&A receptor homodimers, a step required for receptor activation [23].
Historically, the biological activity of the neurotrophins has been studied using explants of primary neurons or neuronal progenitors in culture, as well as certain im- mortal cell lines such as PC12 and MAH cells [6,24]. In the case of NGF, pioneering in viva studies using neutralizing antibodies (either passively administered into the placenta or generated by immunization of pregnant rats with NGF) provided the first conclusive evidence for a role of neurotrophins during the development of the mammalian nervous system [6]. These studies have been described in detailed elsewhere [6,24] and will not be discussed here. In this review, I will focus on recent results obtained with ‘knockout’ mice devoid of either NGF, BDNF or NT-3 (see below).
Trk receptors
The structural features of the Trk family of receptors have been reviewed recently [25]. The unique combina- tion of structural motifs, particularly in their extracellu- lar, ligand-binding domains, indicate that they constitute a novel receptor subfamily (Fig. 2). Their kinase catalytic domain is most closely related to a series of novel orphan receptors that include the mammalian RORl, ROR2, DDR (also known as Ptk-3, Nep or T&E), Tyro-10 and TKT as well as the Torpedo RTK tyrosine kinases [l 11.
Several Trk receptor isoforms have been identified 111,251. Two TrkA kinases differ by the presence of six amino acid residues (VSFSPV, in the single letter code for amino acids) located in the extracellular domain near the transmembrane region [26,27]. Both isoforms appear to have similar biological properties, but they have dif- ferent patterns of expression. Whereas the TrkA isoform carrying the VSFSPV sequence is primarily expressed in
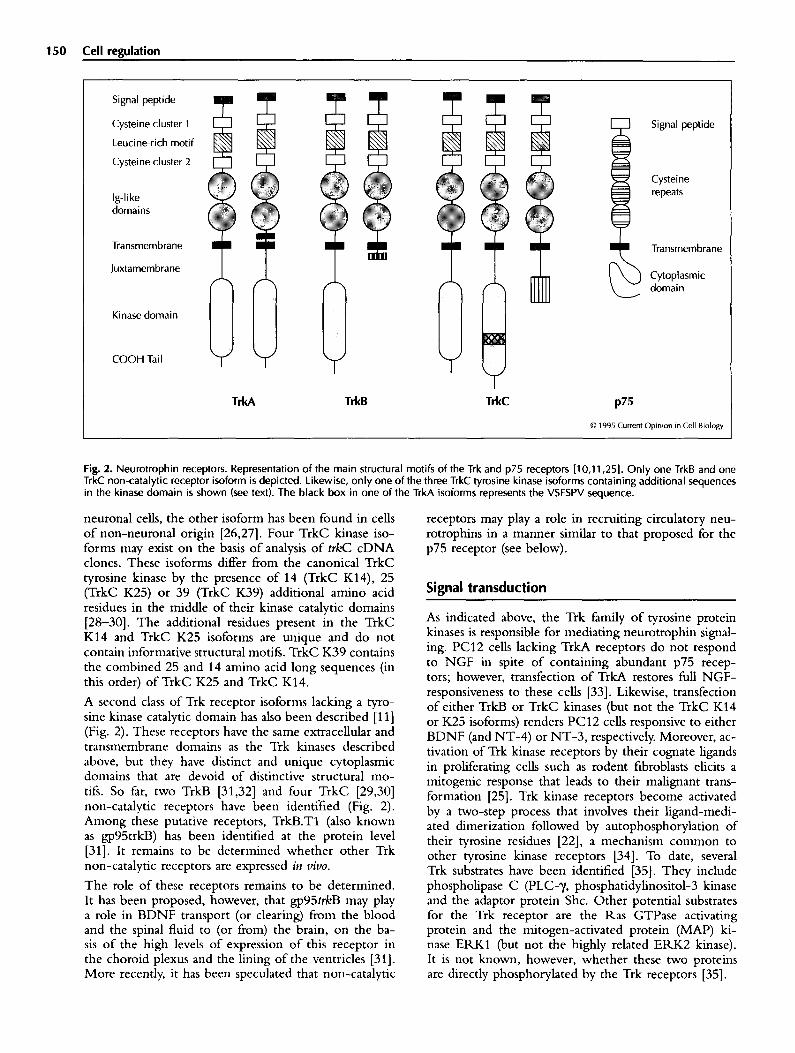
150 Cell reaulation
Signal peptide
Cysteine cluster 1
Leucine-rich motif
Cysteine cluster 2
Ig-like domains
Transmembrane
Juxtamembrane
Kinase domain
COOH Tai I
TrkA TrkB TrkC
Signal peptide
Cysteine repeats
Transmembrane
Cytoplasmic domain
P’5
0 1995 Current Opinion in Cell Biology
Fig. 2. Neurotrophin receptors. Representation of the main structural motifs of the Trk and p75 receptors [lo,1 1,251. Only one TrkB and one TrkC non-catalytic receptor isoform is depicted. Likewise, only one of the three TrkC tyrosine kinase isoforms containing additional sequences in the kinase domain is shown (see text). The black box in one of the TrkA isoforms represents the VSFSPV sequence.
neuronal cells, the other isoform has been found in cells of non-neuronal origin [26,27]. Four TrkC kinase iso- forms may exist on the basis of analysis of t&C cDNA clones. These isoforms differ from the canonical TrkC tyrosine kinase by the presence of 14 (TrkC K14), 25 (TrkC K25) or 39 (TrkC K39) additional amino acid residues in the middle of their kinase catalytic domains [28-301. The additional residues present in the TrkC K14 and TrkC K25 isoforms are unique and do not contain informative structural motifs. TrkC K39 contains the combined 25 and 14 amino acid long sequences (in this order) of TrkC K25 and TrkC K14.
A second class of Trk receptor isoforms lacking a tyro- sine kinase catalytic domain has also been described [l l] (Fig. 2). These receptors have the same extracellular and transmembrane domains as the Trk kinases described above, but they have distinct and unique cytoplasmic domains that are devoid of distinctive structural mo- tifs. So far, two T&B [31,32] and four TrkC [29,30] non-catalytic receptors have been identified (Fig. 2). Among these putative receptors, TrkB.Tl (also known as gp95trkB) has been identified at the protein level [31]. It remains to be determined whether other Trk non-catalytic receptors are expressed in t&o.
The role of these receptors remains to be determined. It has been proposed, however, that gp95trkB may play a role in BDNF transport (or clearing) from the blood and the spinal fluid to (or from) the brain, on the ba- sis of the high levels of expression of this receptor in the choroid plexus and the lining of the ventricles [31]. More recently, it has been speculated that non-catalytic
receptors may play a role in recruiting circulatory neu- rotrophins in a manner similar to that proposed for the p75 receptor (see below).
Signal transduction
As indicated above, the Trk family of tyrosine protein kinases is responsible for mediating neurotrophin signal- ing. PC12 cells lacking TrkA receptors do not respond to NGF in spite of containing abundant p75 recep- tors; however, transfection of TrkA restores full NGF- responsiveness to these cells [33]. Likewise, transfection of either Trkl3 or TrkC kinases (but not the TrkC K14 or K25 isoforms) renders PC12 cells responsive to either BDNF (and NT-4) or NT-3, respectively. Moreover, ac- tivation of Trk kinase receptors by their cognate ligands in proliferating cells such as rodent frbroblasts elicits a mitogenic response that leads to their malignant trans- formation [25]. Trk kinase receptors become activated by a two-step process that involves their ligand-medi- ated dimerization followed by autophosphorylation of their tyrosine residues [22], a mechanism common to other tyrosine kinase receptors [34]. To date, several Trk substrates have been identified [35]. They include phospholipase C (PLC-y, phosphatidylinositol-3 kinase and the adaptor protein She. Other potential substrates for the Trk receptor are the Ras GTPase activating protein and the mitogen-activated protein (MAP) ki- nase ERKl (but not the highly related ERK2 kinase). It is not known, however, whether these two proteins are directly phosphorylated by the Trk receptors [35].

Neurotrophic factors and their receptors Barbacid 151
PLC-‘y binds to a conserved tyrosine residue located in the short carboxy-terminal tail characteristic of the Trk receptor family. Phosphatidylinositol-3 kinase inter- acts with a neighboring tyrosine residue located at the carboxyl terminus of the kinase domain. The phys- iological significance of these interactions remains ob- scure, however, as Trk receptors carrying Tyr+Phe mu- tations in these residues retain their ability to transform NIH3T3 cells and to cause PC12 cells to differentiate [36*,37*]. The She binding site has been mapped to a conserved tyrosine residue located in the juxtamembrane domain [37*]. Mutation of this residue decreases signifi- cantly the mitogenic (NIH3T3 cells) and differentiative (PC12 cells) activity of the Trk receptors. Complete ablation of Trk signaling, however, requires the addi- tional mutation of the tyrosine residue responsible for PLC-y binding [36*,37*]. The TrkC kinase isoforms, TrkC K14 and TrkC K25, have normal autocatalytic kinase activity but they do not phosphorylate PLC-), or phosphatidylinositol-3 kinase [28]. These receptors mediate NT-3 dependent signal transduction, as deter- mined by the induction of c-Fos expression and DNA synthesis, but they cannot elicit full mitogenic or PC12 differentiation responses. These results, taken together, indicate that neurotrophin activity may require activa- tion of multiple signaling pathways.
One of these pathways includes the well characterized signaling elements of the Ras/Raf/MAP kinase path- way [38]. Addition of NGF to wild-type, but not to Trk-deficient, PC12 cells results in the rapid activation of Ras as well as of the downstream Raf and MAP kinases [35]. Moreover, expression of oncogenic Ras and Raf proteins in PC12 cells results in their neuronal differ- entiation in the absence of NGE A second pathway utilized by Trk receptors may involve neuron-specific signaling elements such as the recently identified SNT protein [39]. This 90 kDa protein binds to ~13, a sub- unit of the cell cycle regulatory complex that includes the cdc2 kinase and cyclin. SNT is rapidly phospho- rylated on tyrosine residues upon treatment of PC12 with NGF, but not upon treatment with mitogenic fac- tors such as epidermal growth factor (EGF).
Neurotrophins and their Trk receptors: in vivo
function
The generation of mice carrying germ line mutations in the genes encoding NGF [40”], BDNF [410*,42**] and NT-3 [43**,44**] by homologous recombination in embryonic stem (ES) cells has opened up a unique op- portunity to study the role of these neurotrophins in the development and maintenance of the mammalian ner- vous system. Similar studies involving knockout mice carrying mutations in the catalytic domains of each of the Trk kinase receptors have also provide critical in- formation regarding the role of these receptors in viva [45**-47**]. Moreover, the striking similarities between the phenotypes of mice defective for each Trk recep- tor and its cognate neurotrophin (Table 1) represent the
most compelling evidence to date supporting the con- cept that the Trk receptors mediate most, if not all, of the biological activities of the neurotrophins [17].
Mice defective for either NGF [40**] or TrkA receptors [46**] display severe sensory defects characterized by a complete loss of nociceptive activity (Table 1). These mice fail to react to deep pinpricks in their whisker pads and rear paws [40”,46**]. In addition, they ex- hibit deficiencies in thermoception as they can stay on top of a 60°C hot plate for at least 10s. Neuro- anatomical examination of the TrkA- and NGF-null mice revealed extensive neuronal cell loss in trigemi- nal, dorsal root and sympathetic ganglia [40**,46**]. In the dorsal root ganglia, the vast majority of the missing neurons correspond to those of small size, a population known to be NGF-dependent. The sympathetic ganglia are severely shrunken and contain only a few neurons [40”,46”].
A similar correlation can be found between the phe- notypes of mice targeted in the genes encoding NT-3 [43**,44**] and TrkC [47**] (Table 1). These mice also display severe sensory defects, but of a nature distinct h-om those observed in NGF- and T&A-defective mice. Mice lacking NT-3 or its cognate TrkC kinase receptors have normal nociception, but they are defective in pro- prioception, the sensory activity responsible for localiz- ing the limbs in space [43**,44**,47**]. As a consequence, these targeted mice display abnormal movements of an athetotic nature that result in highly abnormal limb pos- tures. This sensory defect is due to the complete absence of Ia muscle afferents, the projections derived from large proprioceptive dorsal root ganglion neurons that connect primary endings of muscle spindles in the periphery to motor pools in the ventral region of the spinal cord [43**,44**,47**]. TrkC and NT-3 mutant mice have limited life spans most probably because of additional neuronal defects; however, a few TrkC (but not NT-3) mutant mice have survived for over six months. The ap- parently more severe phenotype of the NT-3 targeted mice might simply be attributed to differences in strain or in the animal colony However, the TrkC defective mice may express non-catalytic TrkC receptor isoforms, as the gene targeting took place within their t&C tyro- sine kinase sequences. Whether mutant mice carrying a completely inactivated t&C gene display the shorter life span of the NT-3 knockouts remains to be investigated.
Comparison of the phenotypes of mutant mice deficient in BDNF [41**,42**] and in its TrkB kinase receptor [45**] has been less straightforward (Table 1). TrkB mutants display a severe phenotype that results in the death of most animals in the first postnatal week due to their inability to feed [45**]. BDNF-defective mice also have short life spans; however, many animals survive at least two weeks. These mice display defective movement coordination and balance, with head bobbing and spin- ning followed by long periods of inactivity. This defect is likely to be due to atrophy and loss of vestibular gan- glion neurons, which results in defective innervation of the inner ear [41**,42**]. No such defects were observed
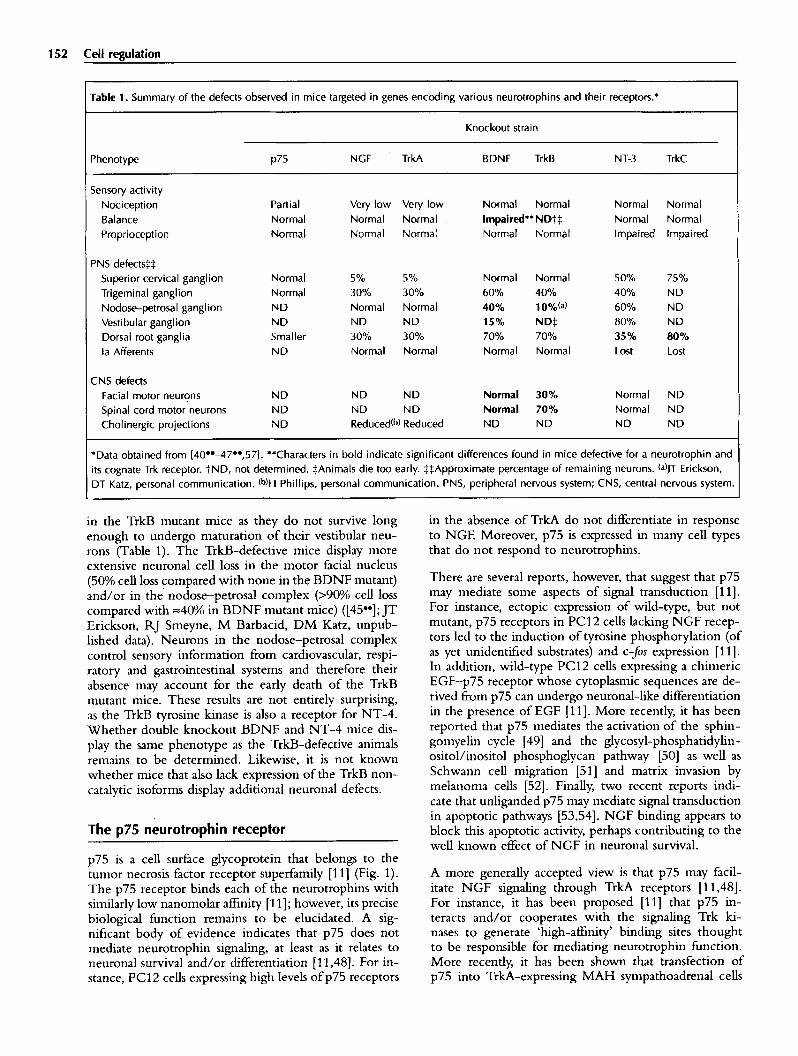
152 Cell regulation
Table 1. Summary of the defects observed in mice targeted in genes encoding various neurotrophins and their receptors.*
Knockout strain
Phenotype p75 NGF TrkA BDNF TrkB NT-3 TrkC
Sensory activity Nociception Partial Very low Very low Normal Normal Normal Normal Balance Normal Normal Necmal . Impaired** ND-~:]: Normal Normal Proprioception Normal Normal Normal Normal Normal Impaired Impaired
PNS defects~::[: Superior cervical ganglion Normal 5% 5% Normal Normal 50% 75% Trigeminal ganglion Normal 30% 30% 60% 40% 40% ND Nodose-petrosal ganglion ND Normal Normal 40% 10% (a) 60% ND Vestibular ganglion ND ND ND 15% ND:]: 80% ND Dorsal root ganglia Smaller 30% 30% 70% 70% 35% 80% la Afferents ND Normal Normal Normal Normal Lost Lost
CNS defects Facial motor neurons ND ND ND Normal 30% Normal ND Spinal cord motor neurons ND ND ND Normal 70% Normal ND Cholinergic projections ND Reduced (b) Reduced ND ND ND ND
*Data obtained from [40°'~7°',57]. **Characters in bold indicate significant differences found in mice defective for a neurotrophin and its cognate Trk receptor. LND, not determined. :[:Animals die too early. :[::[:Approximate percentage of remaining neurons. (a)JT Erickson, DT Katz, personal communication. (b)H Phillips, personal communication. PNS, peripheral nervous system; CNS, central nervous system.
in the TrkB mutant mice as they do not survive long enough to undergo maturation of their vestibular neu- rons (Table 1). The TrkB-defective mice display more extensive neuronal cell loss in the motor facial nucleus (50% cen loss compared with none in the BDNF mutant) and/or in the nodose-petrosal complex (>90% cell loss compared with ~40°A in BDNF mutant mice) ([45°°]; JT Erickson, RJ Smeyne, M Barbacid, DM Katz, unpub- lished data). Neurons in the nodose-petrosal complex control sensory information from cardiovascular, respi- ratory and gastrointestinal systems and therefore their absence may account for the early death of the TrkB mutant mice. These results are not entirely surprising, as the TrkB tyrosine kinase is also a receptor for NT-4. Whether double knockout BDNF and NT-4 mice dis- play the same phenotype as the TrkB-defective animals remains to be determined. Likewise, it is not known whether mice that also lack expression of the TrkB non- catalytic isoforms display additional neuronal defects.
The p75 neurotrophin receptor
p75 is a cell surface glycoprotein that belongs to the tumor necrosis factor receptor superfam~y [11] (Fig. 1). The p75 receptor binds each of the neurotrophins with similarly low nanomolar affinity [11]; however, its precise biological function remains to be elucidated. A sig- nificant body of evidence indicates that p75 does not mediate neurotrophin signaling, at least as it relates to neuronal survival and/or differentiation [11,48]. For in- stance, PC12 cells expressing high levels of p75 receptors
in the absence of TrkA do not differentiate in response to NGE Moreover, p75 is expressed in many cell types that do not respond to neurotrophins.
There are several reports, however, that suggest that p75 may mediate some aspects of signal transduction [11]. For instance, ectopic expression of wild-type, but not mutant, p75 receptors in PC12 cells lacking NGF recep- tors led to the induction oftyrosine phosphorylation (of as yet unidentified substrates) and c-fos expression [11]. In addition, wild-type PC12 cells expressing a chimeric EGF-p75 receptor whose cytoplasmic sequences are de- rived from p75 can undergo neuronal-like differentiation in the presence of EGF [11]. More recently, it has been reported that p75 mediates the activation of the sphin- gomyelin cycle [49] and the glycosyl-phosphatidylin- ositol/inositol phosphoglycan pathway [50] as well as Schwann cell migration [51] and matrix invasion by melanoma cells [52]. Finally, two recent reports indi- cate that unliganded p75 may mediate signal transduction in apoptotic pathways [53,54]. NGF binding appears to block this apoptotic activity, perhaps contributing to the well known effect of NGF in neuronal survival.
A more generally accepted view is that p75 may facil- itate NGF signaling through TrkA receptors [11,48]. For instance, it has been proposed [11] that p75 in- teracts and/or cooperates with the signaling Trk ki- nases to generate 'high-affinity' binding sites thought to be responsible for mediating neurotrophin function. More recently, it has been shown that transfection of p75 into TrkA-expressing MAH sympathoadrenal cells

Neurotrophic factors and their receutors Barbacid 153
enhances their response to NGF [55]. Likewise, block- ing p75 binding sites in PC12 cells with either a p75 monoclonal antibody or with BDNF decreases their re- sponse to NGF [56].
Role of p75 receptors in vivo
The most relevant information regarding the function of the p75 receptor in neurotrophin signaling has been provided by analysis of mutant mice carrying a targeted p75 gene [57]. These mice display sensory and sympa- thetic defects, thus demonstrating that the p75 recep- tor is indeed required for proper neuronal development (Table 1). Interestingly, these mice do not display the defects characteristic of mice lacking BDNF (or TrkB receptors) and NT-3 (or TrkC receptors). These obser- vations indicate that ~75, despite serving as a receptor for BDNF and NT-3 in vitro, may not be involved in mediating their biological activities in viva (Fig. 1). Al- ternatively, the absence of p75 might be compensated for by other molecules such as the TrkI3 and TrkC non- catalytic receptors.
The observed defects in the p75-null mice appear to be limited to NGF-dependent neurons, mainly sensory (nociceptive) and sympathetic neurons [58*]. Moreover, these defects are much more limited than those observed in either NGF or TrkA mutant mice (Table 1). For in- stance, NGF- and T&A-null mice have lost most (>95%) of their sympathetic neurons, whereas, in contrast, the p75 null mice have normal sympathetic ganglia as well as normal innervation patterns to all their target tissues with the exception of the pineal and the sweat glands [57,58’].
These observations raise the possibility that TrkA sig- naling may require p75 receptors in only a subset of NGF-dependent neurons. A more plausible interpreta- tion of these results, however, is that p75 plays a role in recruiting circulating NGF molecules rather than in NGF signaling [48]. In support of this hypothesis, p75-defective sensory and sympathetic neurons survive well in culture in the presence of NGF [59*,60], but they need four times more NGF to achieve the same response as wild-type neurons [59*,60]. Considering the rapid dissociation kinetics of NGF binding to p75 [6], it is possible that expression of p75 receptors either in neurons or in adjacent glial cells may increase the lo- cal concentration of diffusible NGE If so, the lack of p75 receptors will only have phenotypic consequences in those neurons for which availability of NGF is limiting. Crossing p75-null mice with transgenic strains overex- pressing NGF should provide the experimental tools to test this hypothesis.
Conclusions
Most of the information we had to date regarding the role of neurotrophic factors has been derived corn in vitro studies. The advent of gene targeting has provided new
means to study their function in viva. Characterization of strains of transgenic mice lacking neurotrophic factors and their receptors is making it possible to define pre- cisely the physiological role of each of these molecules, at least during development. Some of the results obtained (i.e. the observation of sympathetic and sensory defects in NGF- and T&A-null mice) were partly expected on the basis of earlier studies using immunological ap- proaches to neutralize NGF activity. Others, such as the exquisite specificity of the NT-3-TrkC pathway in pro- prioception or the limited defects displayed by the p75- null mice, were unexpected. Another important lesson learned from these gene-targeted mice is the differen- tial role that neurotrophins play in the peripheral versus the central nervous systems. In the peripheral nervous system, ablation of neurotrophin or Trk receptor genes results in massive neuronal cell death, whereas the central nervous system neurons of these mutant mice appear to be, for the most part, unaffected in spite of widespread neurotrophin and receptor expression. These observa- tions illustrate the complex mechanisms involved in the development and survival of the nervous system and pre- dict the existence of additional neurotrophic factors that await discovery.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as: . of special interest . . of outstanding interest
1. Westermark B, Heldin CH: Platelet-derived growth factor. Structure, function and implications in normal and malignant cell growth. Acta Oncol 1993, 32:101-l 05.
2. Eckenstein FP: Fibroblast growth factors in the nervous system. Neurobiology 1994, 25:1467-l 480.
3. Patterson PH: The emerging neuropoietic cytokine family: first CDF/LIF, CNTF and L-6; next ONC, MCF, CCSF? Curr Opin Neurobiol 1992, 2:94-97.
4. Sendtner M, Carroll P, Holtmann 6, Hughes RA, Thoenen H: Ciliary neurotrophic factor. / Neurobiol 1994, 25:1436-l 453.
5. Lin LF, Doherty DH, Lile JD, Bektesh S, Collins F: CDNF: a glial . cell line-derived neurotroohic factor for midbrain dooaminer-
gic neurons. Science 1993, 260:1130-l 132. ’ This paper describes the isolation and cloning of a new neurotrophic factor, glial-derived neurotrophic factor. Although initially thought to be specific for dopaminergic neurons, recent studies [l 81 indicate that it may have a broad spectrum of action.
6.
7.
8.
9.
10.
11.
12.
Levi-Montalcini R: The nerve growth factor 35 years later. Science 1987, 237:1154-1162.
Barde Y-A: Neurotrophic factors: an evolutionary perspective. / Neurobiol 1994, 25:1329-l 333.
Meakin SO, Shooter EM: The nerve growth factor family of receptors. Trends Neurosci 1992, 15:323-331.
Chao MV: Neurotrophin receptors: a window into neuronal differentiation. Neuron 1992, 9:583-593.
Barbacid M: The Trk family of neurotrophin receptors. ) Neu- robiol 1994, 25:1386-l 403.
Chao MV: The p75 neurotrophin receptor. / Neurobiol 1994, 25:1373-l 385.
Stahl N, Yancopoulos CD: The alphas, betas, and kinases of cytokine receptor complexes. Cell 1993, 74:587-590.

154 Cell reeulation
13.
14.
15.
16.
17.
18.
19. .
Davis S, Yancopoulos CD: The mofecufat biifogy of the CNTF forms of rat TrfK with different functiunaf capabilities. Neuron receptor. Curr Opin Ccjl &of 1993, 5:281-285. 1993, 10:963-974.
Davis S, Aldrich TH, Valenzuela DM, Wong VV, Furth ME, Squint0 SP, Yancopoulos CD: The recqtor for ciliiry neuro- trof&ic factor. hence 1991, 253:59-63.
31. Klein R, Conway D, Parada LF, Barbacid M: The trkB tyrosine protein fdnase gene codes for a second neurogenic receptor Ff& facks the catalytic fcinase don&n. Ce# 1990, 61:647-656.
Kishimoto T, Akira S, Taga T: lnlerleukin-6 and its receptor: a paradigm for cytokiies, Scieoce 1992, 258~593-597.
32.
Gearing DP, Thut CJ, VandeBos T, Gimpef SD, Delaney PB, King J, Price V, Cosman D, Beckmann MP: Leukemia in- hibitory factor receptor is structurally refated to the IL-6 signal &ansducer, gp130. EMBO f 1991, ?0:2839-2848.
Middlemas DS, Lindberg RA, Hunter T: trkB, a neural recep tar protein-tyrosine fdise: evidence for a full-fengtf~ and two trunca&d receptors. Mof Cell &of 1991, It:7 43-l 53.
33.
Snider WD: Functions of the nwrotropbins during nervous system ~vel~~~ what the k&outs are teaching us. Cefi 1994, 77627638.
Loeb DM, Maragos J, Martin-Zanca D, Chao MV, Parada LF, Greene LA: The trk proto-oncogene rescues NGF responsive- ness in mutant NGF-nonresponsive PC12 cuff fhtes. Ceff 1991, 66:961-966.
34. Schlessinger J, Utirich A: Growth factor signaling by receptor tyrosine kinases. Neuron 1992, 9~383-391.
Henderson CE, Phillips HS, Pollock RA, Davies AM, Lemeulie C, Armanini M, Simpson LC, Moffet B, Vandien RA, Koiiatsos VE, Rosenthal A: GDNF: a potent survival factor for motoneu- rons present in peripheral nerve and muscle. Science 1994, 266: 10&Z-1 064.
35. Kaplan DR, Stephens RM: Neurotrophin signal transduction by the Trk receptor. j ~~r~i~i 1994, 25:1404-1417.
36. l
Gotz R, Koster R, Winkler C, Raulf F, Lottspeich F, Schartl M, Thoenen H: Neurotrof&in-6 is a new member of the nerve grow& factor family. &&We $994, 372:26L%9.
Stephens RM, Loeb DM, Copeland TD, Pawson T, Greene LA, Kaplan DR: Trk recep&rs use redundant signaf trans. duction pathways involving WC and PLC-yl to mediate NCF responses. Neuron 1994, 12:691-705.
See I37.1 annotation.
37. Obermeier A, Bradshaw RA, Seedotf K, Choidas A, Schlessinger . J, Ullrich A: Neuronaf differentiation signals are cnntrolfed by
nerve growth factor receptor/Tilt binding sites for WC and P&y* EMBO f 1994, f 3:t 58%1590.
Describes the isolation of a novel neurotroohin. neurotroohin (NT)-6. Ex- tensive efforts using approaches based on the polymeras;! chain reaction to isolate new neurotrophins, after the rapid discovery of NT-3 and NT-4, had failed, thus sugg&ing that there might not be additional members of the neurotrophin family. it remains to be determined, however, whether NT-6 homologues exist in mammals.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
McDonald NQ, Lapatto R, Murray-Uust f, Gunning J, ylodawer A, Blundell TL: New protein fold revealed by a 2.3A resofu- tion crystal structure of nerve growth factor. Nature 1991, 354:411-414.
ibanez CF, Ebendal T, Barbany G, Murray-Rust 1, Blundell Tl, Persson Ii: Disruption of * low affinity ~eptor~~i~ site in NGF allows neuronal survival and differentiation by binding to the trk gene product. Cell “1992, 69:329-342.
lbanez CF, flag LL, Murray-Rust J, Persson i-l: An extended surface of binding to Trk tyrosine kinase receptors in NCF and BDNF allows the engineering of a multifunctional pan- neurotiophin. EMBO I 1993, 12:2281-2293.
Jing S, Tapley P, Barbacid M: Nerve growth factor mediates signal transduction through trk homodimer receptors. Neuron 1992, 9:1067-1079.
Chao MV: Growth factor signaling: where is the specificity? Celi 1992, 68:995-Y%‘.
Barbacid M: The Trk family of ~eurotrop~in receptors: mofec- ufar characterization and oncogenic activation in human tu- mors. in Mo~~u~ar genetics of nervous system tumors. Edited by Levine AJ, Schmidek HH. New York: Wiley and Sons, Inc; 1993:123-l 35.
Barker PA, Lomen-i-ioerth C, Gensch EM, Meakin SO, Glass DJ, Shooter EM: Tissue-specific alternative splicing gener- ates two isoforms of the trkA receptor. 1 Biol Chem 1993, 268:1515&15157.
Horigome K, Pryor JC, Bullock ED, Johnson EM Jr: Media- tor release from mast cells by nerve growth factor. Neuro- trophm specifiiify and receptor mediation. f Biol Chem 1993, 26~1488~-14887.
Lamb&e F, Tapley P, Barbacid M: t&C encodes multiple neurotrophin-3 receptors with distinct biological properties and substrate speeifidties. EMBO / 1993, 12:3083-3094.
Tsoulfas P, Soppet 0, Escandon E, Tessarollo L, Mendoza- Ramirez IL. Rosenthal A. Nikolics K. Parada LF: The rat trkC locus en&es multiple burogenic receptors that exhibit dif- &e&al response to n~~rophi~~ in PC12 cells. Neuron 1993, l&975-990.
Valenruela DM, Maisonpierre PC, Glass DJ, Rojas E, Nunez L, Kong Y, Gies DR, St&t TN, lp NY, Yancopoulos GD: AltemaGe
In this paper and 136.1, site-directed mutagenesis was used to define the role of individual tyrosine residues in the cytoplasmic domain of the TrkA receptor. These tyrosine residues serve as specific anchors for various TrkA substrates. Therefore, the biological activity of these mutant recep tors provides useful information regarding the relative contributions of these substrates {and their respective signaling pathways) to signailing by nerve growth factor.
38. Egan SE, Weinberg RAz The pathway to signal a&iivement. Nature 1993, 365~781-783.
39. Rabin SJ, Cleghon V, Kaplan DR: SNT, a differentiation-specific taq+ of neurotrophic factor-fnduced tyrosine fdnase activity in neurons and PC1 2 cells. Mel Ceff EGoi 1993, 13~2203-22 13.
40. l *
Crowley C, Spencer SD, Nishimura MC, Chen KS, Pitts-Meek S, Armanini MP, Ling LH, McMahon SB, Shelton DL, Levinson AD, Phillips HS: Mice lacking nerve growth factor dispfay perinataf loss of sensory and sympathetic neurons yet develop basal forebrain chofinergic neurons, Cell 1994, 76:1001-l 011.
Disruption of the nerve growth factor JNGFJ gene by homologous re- combination in embryonic stem cells results in the generation of mice that do not respond to noxious mechanical stimuli. These mice dis- play severe loss of sensory and sympathetic neurons, confirming the critical dependence of these neurons for NCF previously shown in in vitro studies.
41. Jones KR, Farinas I, Backus C, Reichardt LF: Targeted disrup . . tion of the BDNF gene perturbs brain and sensory neuron
development but not motor neuron development. Cetl 1994, 76:989-999.
See [42**1 annotation.
42. l *
Ernfors P, Lee KF, Jaenisch R: Mice lacking brain-derived neu- ro&opftic factor develop with sensory de&&s. Naiure 1994, 368: 147-l 50.
This study and [41**] independently report the generation of mice lack- ing a functional brain-derived neurotrophic factor gene. Most of these mice die after birth, although some survive for 2-4 weeks, These mice display severe deficiencies in coordination and balance, a phenotype that may result from their significantly reduced levels of vestibular sensory neurons. These mice also display neuronal deficiencies in other cranial and spinal sensory ganglia, but not in motor neurons.
43. Farinas I, Jones KR, Backus C, Wang XV, Reichardt LF: l . Severe sensory and sympathetic deficits in mice facking
neurotrophin-3. Nature 1994, 369:658-661. See [44**1 annotation.
44. . .
Ernfors P, Lee KF, Kucera J, Jaenisch R: lack of weurotrophiw-3 leads to deficiencies in the peripheral nervous system and loss of lib mioceptive afi+ren&. C&i 1994, 7R503-512.

Mice lacking neurotrophin-3 die within a few weeks of birth and dis- play abnormal limb movements. This defect is likely to be due to the lack of spinal proprioceptive afferents and muscle spindles. These mice display additional defects in other sensory (trigeminal and cochlear) and sympathetic ganglia, but not in enteric or motor neurons. In addition, the central nervous system of these mice appears to be normal, at least anatomically.
45. Klein R, Smeyne RJ, Wurst W, Long LK, Auerbach BA, Ioyner *• AL, Barbacid M: Targeted disruption of the trkB neurotrophin
receptor gene results in nervous system lesions and neonatal death. Cell 1993, 75:113-122.
Partial disruption of the trkB gene led to the generation of mice lack- ing TrkB tyrosine kinase receptors but not their non-catalytic isoform gp95trkB. Most of these mice die during the first postnatal week due to lack of feeding activity. They display neuronal defects in the periph- eral (trigeminal and dorsal root ganglia) and central (facial motor nucleus and spinal cord) nervous system. However, most structures of the central nervous system known to express high levels of trkB transcripts (cortex, hippocampus, etc.) appear to be normal, at least anatomically.
46. Smeyne RJ, Klein R, Schnapp A, Long LK, Bryant S, Lewin A, "• Lira SA, Barbacid M: Severe sensory and sympathetic neoro-
pathies in mice carrying a disrupted Trk/NGF receptor gene. Nature 1994, 368:246-249.
Mice lacking TrkA receptors display the same sensory and sympathetic defects as those carrying a disrupted nerve growth factor (NGF) gene, thus demonstrating that TrkA mediates NGF activity in vivo. TrkA-de- fective mice also display deficiencies in the cholinergic projections that connect basal forebrain neurons (known to be NGF-dependent) with the hippocampus and cortex.
47. Klein R, Silos-Santiago I, Smeyne RJ, Lira SA, Brambilla R, • • Bryant S, Zhang L, Snider WD, Barbacid M: Disruption of
the neurotruphln-3 receptor gene trkC eliminates ia muscle afferents and results in abnormal movements. Nature 1994, 368:249-251.
Specific disruption of those sequences encoding the catalytic domain of the TrkC tyrosine kinase receptor results in mice with severe propriocep- live sensory defects, most likely caused by the total absence of la muscle afferent projections to spinal motor neurons. These mice also have fewer large myelinated axons in the dorsal root and posterior columns of the spinal cord. Unlike mice lacking neurotrophin-3, some of the TrkC-de- fective mice survive for long periods of time (>6months).
48. Barbacid M: Nerve growth factor: a tale of two receptors. Oncogene 1993, 8:2033-2042.
49. Dobrowsky RT, Werner MH, Castellino AM, Chao MV, Hannun YA: Activation of the sphingomyelln cycle through the low- affinity neurotrophin receptor. Science 1994, 265:1596-1599.
50. Represa J, Avila MA, Miner C, Giraldez F, Romero G, Clemente R, Mato JM, Varela-Nieto I: Glycosyl-phosphatidyli- nositol/inositol phosphoglycan: a signaling system for the low-
Neuro t roph ic factors and thei r receptors Barbacid 155
52.
affinity nerve growth factor receptor. Proc Nat/Acad Sci USA 1991, 88:8016-8019.
51. Anion ES, Weskamp G, Reichardt LF, Matthew WD: Nerve growth factor and ils low-affinity receptor promote Schwann cell migration. Proc Nat/Acad Sci USA ]994, 91:2795-2799.
Herrmann JL, Menter DG, Hamada J, Marchetti D, Nakajima M, Nicolson GL: Mediation of NGF-stimulated extracelluiar matrix invasion by the human melanoma Iow-affinlty p75 neu- rotrophln receptor: melanoma p75 functions independently of trkA. Mol Biol Cell 1993, 4:]205-]216.
53. Rabizadeh S, Oh l, Zhong LT, Yang J, Bitler CM, Butcher LL, Bredesen DE: Induction of apoptosis by the low-affinity NGF receptor. Science 1993, 261:345-348.
54. Barrett GL, Bartlett PF: The p75 nerve growth factor recep- tor mediates survival or death depending on the stage of sensory neuron development. Proc Nat/ Acad Sci USA 1994, 91:6501-6505.
55. Verdi JM, Birren SJ, Ibanez CF, Persson H, Kaplan DR, Benedetti M, Chao MV, Anderson DJ: p75LNGFR regulates Irk sig- nal transduction and NGF-induced neuronal differentiation in MAH cells. Neuron 1994, 12:733-745.
56. Barker PA, Shooter EM: Disruption of NGF binding to the low affinity neurotrophln receptor p75LNTR reduces NGF binding to TrkA on PC12 cells. Neuron ]994, 13:203-215.
57. Lee KF, Li E, Huber LI, Landis SC, Sharpe AH, Chao MV, Jaenisch R: Targeted mutation of the gene encoding the low affinity NGF receptor p75 leads to deficits in the pe- ripheral sensory nervous system. Cell 1992, 69:737-749.
58. Lee KF, Bachman K, Landis S, Jaenisch R: Dependence on p75 • for innervation of some sympathetic targets. Science 1994,
263:1447-] 449. See [59 •] annotation.
59. Davies AM, Lee KF, laenisch R: p75-deficlent trigemlnal sen- . sory neurons have an altered response to NGF but not to
other neurotrophins. Neuron 1993, 11:565-574. This study and [58 •] extend the initial characterization of the p75-null mice [57] and provide support for the concept that p75 receptors might only be indispensible for those neurons (and innervations) that have ac- cess to limiting amounts of nerve growth factor.
60. Lee KF, Davies AM, laenisch R: p75-deflcient embryonic dorsal root sensory and neonatal sympathetic neurons dis- play a decreased sensitivity to NGF. Development 1994, 120:1027-1033.
M Barbacid, Department of Molecular Biology, Bristol-Myers Squibb Pharmaceutical Research Institute, PO Box 4000, Prince- ton, NJ 08543-4000, USA.