niacin treatment of stroke increases synaptic plasticity...
TRANSCRIPT

Niacin Treatment of Stroke Increases Synaptic Plasticityand Axon Growth in Rats
Xu Cui, PhD; Michael Chopp, PhD; Alex Zacharek, MS; Cynthia Roberts, BS; Benjamin Buller, BS;Madalina Ion, BS; Jieli Chen, MD
Background and Purpose—Niacin is the most effective medication in current clinical use for increasing high-densitylipoprotein cholesterol. We tested the hypothesis that niacin treatment of stroke promotes synaptic plasticity and axongrowth in the ischemic brain.
Methods—Male Wistar rats were subjected to 2 hours of middle cerebral artery occlusion and treated with or withoutNiaspan (a prolonged-release formulation of niacin, 40 mg/kg) daily for 14 days starting 24 hours after middle cerebralartery occlusion. The expression of synaptophysin, Nogo receptor, Bielschowsky silver, brain-derived neurotrophicfactor, and its receptor tropomyosin-related kinase B were measured by immunohistostaining and Western blot,respectively, in the ischemic brain. Complementing in vivo studies, primary cultured neurons were used to test the effectof niacin and high-density lipoprotein on neurite outgrowth and brain-derived neurotrophic factor/tropomyosin-relatedkinase B expression.
Results—Niaspan treatment of stroke significantly increased synaptophysin, Bielschowsky silver, brain-derived neurotrophicfactor/tropomyosin-related kinase B expression, and decreased Nogo receptor expression in the ischemic brain compared withmiddle cerebral artery occlusion control animals (P�0.05, n�8/group). Niacin and high-density lipoprotein treatmentsignificantly increased neurite outgrowth and brain-derived neurotrophic factor/tropomyosin-related kinase B expression inprimary cultured neurons. Tropomyosin-related kinase B inhibitor attenuated niacin-induced neurite outgrowth (P�0.05,n�6/group).
Conclusions—Niacin treatment of stroke promotes synaptic plasticity and axon growth, which is mediated, at leastpartially, by the brain-derived neurotrophic factor/tropomyosin-related kinase B pathways. (Stroke. 2010;41:00-00.)
Key Words: axon growth � HDL-cholesterol � niacin � plasticity � stroke � synaptic
Synaptic plasticity and axon growth are related to behav-ioral change and functional recovery after brain and
spinal cord injury.1 Functional alterations in motor cortexorganization are accompanied by changes in dendritic andsynaptic structure.2 Cortical stimulation-induced functionalimprovements after stroke are mediated by synaptic structuralplasticity.3 Increases in dendritic arborization and spinedensity are potential morphological strategies that enable thebrain to reorganize its neuronal circuits.4
High-density lipoprotein cholesterol (HDL-C) is critical inmaintaining the homeostasis of cell membrane cholesteroland thus plays an essential role in the regulation of synapticfunction and cell plasticity.5 The expression of brain-derivedneurotrophic factor (BDNF) and its receptor tropomyosin-related kinase B (TrkB) supports neuron survival and axongrowth after neuronal injury.6,7 Voluntary exercise leads to anendogenous upregulation of BDNF and associated proteinsinvolved in synaptic function and enhances functional recov-ery after traumatic brain injury.8 Synaptic plasticity is alsoinfluenced by mutations in BDNF.9
Niacin (nicotinic acid) is the most effective medication incurrent clinical use for increasing HDL-C.10 Our previousstudy showed that Niaspan (a prolonged-release formulationof niacin) treatment of stoke significantly increases serumHDL-C level and improves functional outcome in rats.11
However, whether niacin increases synaptic plasticity andaxon growth and whether BDNF/TrkB play a role in niacin-induced synaptic plasticity and axon growth has not beeninvestigated. In this study, we tested a novel hypothesis, thatniacin treatment of stroke promotes synaptic plasticity andaxon growth in the ischemic brain in rats. In addition, thecontributions of BDNF/TrkB to niacin-induced synaptic plas-ticity and axon growth were investigated.
Materials and MethodsAnimal Middle Cerebral Artery Occlusion andExperimental GroupsAdult male Wistar rats weighing 270 to 300 g (Jackson Laboratory,Bar Harbor, Maine) were subjected to 2 hours of right middle
Received May 4, 2010; accepted June 9, 2010.From the Department of Neurology (X.C., M.C., A.Z., C.R., B.B., M.I., J.C.), Henry Ford Hospital, Detroit, Mich; and the Department of Physics
(M.C., B.B.), Oakland University, Rochester, Mich.Correspondence to Jieli Chen, MD, Neurology Research, E&R Building, Room 3091, Henry Ford Hospital, 2799 West Grand Boulevard, Detroit, MI,
48202. E-mail [email protected]© 2010 American Heart Association, Inc.
Stroke is available at http://stroke.ahajournals.org DOI: 10.1161/STROKEAHA.110.589333
1
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from

cerebral artery occlusion (MCAO).11 Rats were gavaged starting 24hours after surgery with: (1) saline for vehicle control; or (2) 40mg/kg Niaspan (KOS Pharmaceuticals) daily for 14 days.11 Sham-operated rats underwent the same surgical procedure without sutureinsertion. These rats were euthanized 14 days after MCAO forimmunostaining (n�8/group). Additional sets of rats (n�4/group)were euthanized 5 days after MCAO, and the brain tissues wereprepared for Western blot assay.
Histological and Immunohistochemical AssessmentRats were euthanized 14 days after stroke. The brains were fixed bytranscardial perfusion with saline followed by perfusion and immer-sion in 4% paraformaldehyde before being embedded in paraffin. Astandard paraffin block was obtained from the center of the lesion(bregma �1 mm to �1 mm). A series of 6-�m thick sections werecut from the block. Every 10th coronal section for a total 5 sectionswas used for immunohistochemical staining. Antibody against Syn-aptophysin (1:1000; Chemicon), Nogo receptor (NgR; 1:50; SantaCruz), BDNF (1:300; Santa Cruz), and TrkB (1:500; Santa Cruz)immunostaining was performed. Bielschowsky silver staining wasalso used as previously described.12 Control experiments consistedof staining brain coronal tissue sections as outlined previously, butnonimmune serum was substituted for the primary antibody. Theimmunostaining analysis was performed by an investigator blindedto the experimental groups.
Immunostaining QuantificationLike our previous description,11 synaptophysin, NgR, BDNF, TrkB,and Bielschowsky silver immunohistostained sections were digitizedusing a 40� objective (Olympus BX40) using a 3-CCD color videocamera (Sony DXC-970MD) interfaced with the Micro ComputerImaging Device computer imaging analysis system (Imaging Re-search). Synaptophysin-, BDNF-, and TrkB-positive area wascounted in the ischemic boundary zone (IBZ, which adjacent to theischemic core) in each section.11 Five sections and 8 views in eachsection were counted per rat. For semiquantitative measurements ofBielschowsky silver and NgR, the positive stained areas in thebundle of striatum in the IBZ were measured. Data were analyzed ina blinded manner and presented as percentage of positive area forsynaptophysin, Bielschowsky silver, NgR, BDNF, and TrkB immu-noreactivity, respectively.
Primary Cultured Neuron and TreatmentsTo test whether niacin regulates dendrite outgrowth, primary cul-tured neuron (PCN) culture was used. Embryonic Day 17 corticalcells were isolated under the 10� microscope from embryonic brainof Wistar rats and cultured in 4-chamber slides with Neuralbasal-Amedium (GIBCO) containing 2% B27 medium supplement (GIBCO)and antibiotics for 7 days (6 chambers/group) in vitro.13 To mimicthe ischemic condition in vivo, oxygen-glucose deprivation (OGD)was induced in vitro as previously described.14 OGD was inducedwithin an anaerobic chamber. Briefly, the PCN cultures weretransferred to the anaerobic chamber (Model 1025; Forma Scientific)and were incubated in 85% N2, 10% H2, and 5% CO2 at 37°C for 1hour. The PCN cultures were removed from the anaerobic chamber,rinsed with phosphate-buffered saline, and fed with the primarymedia and the groups were divided into: (1) control; (2) high-densitylipoprotein (HDL; 80 �g/mL; Calbiochem, Cat. #437641); (3)1 mmol/L niacin; (4) 5 mmol/L niacin; (5) TrkB inhibitor (K252a,200 nmol/L; Calbiochem, Cat. #480354); (6) TrkB inhibitor�HDL;(7) TrkB inhibitor�1 mmol/L niacin; or (8) TrkB inhibi-tor�5 mmol/L niacin for 24 hours. Western blot assay and realtime-polymerase chain reaction (RT-PCR) were performed, respec-tively. The PCN cultures were also performed for neuron-specificClass III �-tubulin (TUJ1, a phenotypic marker of neural cells)immunofluorescent staining using a monoclonal anti-TUJ1 antibody(Covance; 1:1000) with Cy3 for PCN number counting and neuriteoutgrowth measurement.15
Quantification of PCN Numbers and Measurementof PCN Neurite OutgrowthTo count PCN numbers and trace the axonal arbors of fluorescentlylabeled neurons, the fluorescent photomicrographs were captured at40� magnification with a digital camera; the TUJ1-positive PCNnumbers and the length of TUJ1-positive dendrites were measuredusing Micro Computer Imaging Device analysis system.16 Theaverage number of TUJ1-positive PCNs per 40� field and theaverage length of total 20 neuronal dendrite outgrowth werepresented.
Western BlotRats were euthanized 5 days after MCAO and brain tissues wereextracted from the IBZ tissue. Equal amounts of cell lysate weresubjected to Western blot analysis, as previously described.17 PCNcells were harvested after 24 hours of treatment for Western blot.Total protein was isolated from treated cells with TRIzol (Invitrogen)following a standard protocol. Heat the protein samples in 1%sodium dodecyl sulfate for approximately 20 minutes at 60°C torecover the protein activity. Specific proteins were visualized usinga SuperSignal West Pico chemiluminescence kit (Pierce). Thefollowing primary antibodies were used: anti-BDNF (1:1000; SantaCruz), anti-TrkB (1:1000; Santa Cruz), and anti-�-actin (1:2000;Sigma).
Real-Time Polymerase Chain ReactionPCN cultures were harvested after 24 hours of treatment and totalRNA was isolated with TRIzol (Invitrogen). Quantitative PCR wasperformed using the SYBR Green RT-PCR method on an ABI 7000PCR instrument (Applied Biosystems), as previously described.11
The following primers for RT-PCR were designed using PrimerExpress software (ABI). BDNF forward: TAC TTC GGT TGC ATGAAG GCG; reverse: GTC AGA CCT CTC GAA CCT GCC. TrkBforward: TCA TCA AGT CAG AGG TGA CAG G; reverse: ACTGGG TAC ACT CCT TCT CTC G. glyceraldehyde-3-phosphatedehydrogenase: forward: AGA ACA TCA TCC CTG CAT CC;reverse: CAC ATT GGG GGT AGG AAC AC. Each sample wastested in triplicate, and samples were obtained from 6 independentexperiments that were used for analysis of relative gene expressiondata using the 2���CT method.
Statistical AnalysisTwo-way analysis of variance was performed on data of thepercentage of positive area for synaptophysin, NgR, Bielschowskysilver, BDNF, and TrkB in the ischemic brain. If an overall treatmentgroup effect was detected at P�0.05, Tukey test after post hoc testwas used for multiple comparison. Independent-sample t test wasused for testing the expression of BDNF and TrkB by Western blotassay in the brain tissues between 2 groups. One-way analysis ofvariance and least significant difference analysis after post hoc testwas performed to assess data of BDNF and TrkB expression byWestern blot and RT-PCR assay in the ischemic brain and PCNcultures and the neurite outgrowth in vitro. All data are presented asmean�SE.
ResultsNiaspan Treatment of Stroke Increases SynapticPlasticity and Axon Growth in the Ischemic BrainTo test whether Niaspan treatment of stroke induces synapticplasticity and axon growth, synaptophysin, Bielschowskysilver, and NgR immunostaining were performed. Synapto-physin is a marker for presynaptic plasticity and synaptogen-esis.18 Bielschowsky silver is a marker for axons.19,20 NgR, aneurite outgrowth inhibitor, regulates axonal growth as wellas axon regeneration after injury.12,21 Figure 1A–L shows thatthe expression of Bielschowsky sliver (Figure 1A–D) andsynaptophysin (Figure 1I–L) significantly increased in the
2 Stroke September 2010
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from
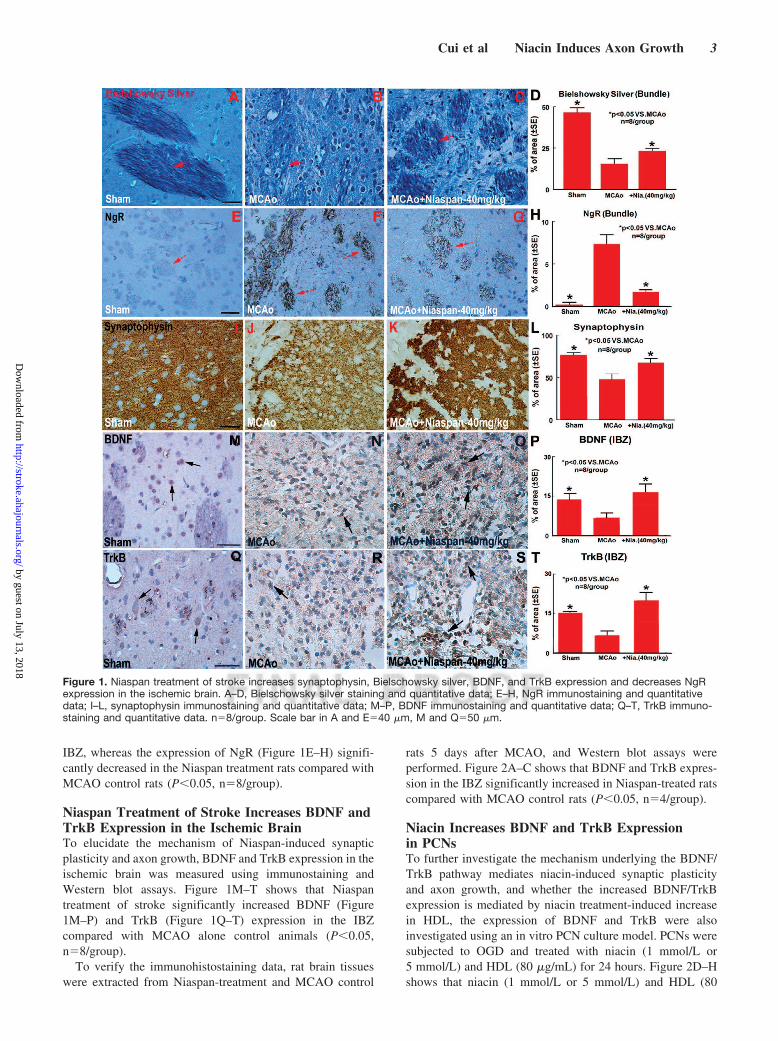
IBZ, whereas the expression of NgR (Figure 1E–H) signifi-cantly decreased in the Niaspan treatment rats compared withMCAO control rats (P�0.05, n�8/group).
Niaspan Treatment of Stroke Increases BDNF andTrkB Expression in the Ischemic BrainTo elucidate the mechanism of Niaspan-induced synapticplasticity and axon growth, BDNF and TrkB expression in theischemic brain was measured using immunostaining andWestern blot assays. Figure 1M–T shows that Niaspantreatment of stroke significantly increased BDNF (Figure1M–P) and TrkB (Figure 1Q–T) expression in the IBZcompared with MCAO alone control animals (P�0.05,n�8/group).
To verify the immunohistostaining data, rat brain tissueswere extracted from Niaspan-treatment and MCAO control
rats 5 days after MCAO, and Western blot assays wereperformed. Figure 2A–C shows that BDNF and TrkB expres-sion in the IBZ significantly increased in Niaspan-treated ratscompared with MCAO control rats (P�0.05, n�4/group).
Niacin Increases BDNF and TrkB Expressionin PCNsTo further investigate the mechanism underlying the BDNF/TrkB pathway mediates niacin-induced synaptic plasticityand axon growth, and whether the increased BDNF/TrkBexpression is mediated by niacin treatment-induced increasein HDL, the expression of BDNF and TrkB were alsoinvestigated using an in vitro PCN culture model. PCNs weresubjected to OGD and treated with niacin (1 mmol/L or5 mmol/L) and HDL (80 �g/mL) for 24 hours. Figure 2D–Hshows that niacin (1 mmol/L or 5 mmol/L) and HDL (80
Figure 1. Niaspan treatment of stroke increases synaptophysin, Bielschowsky silver, BDNF, and TrkB expression and decreases NgRexpression in the ischemic brain. A–D, Bielschowsky silver staining and quantitative data; E–H, NgR immunostaining and quantitativedata; I–L, synaptophysin immunostaining and quantitative data; M–P, BDNF immunostaining and quantitative data; Q–T, TrkB immuno-staining and quantitative data. n�8/group. Scale bar in A and E�40 �m, M and Q�50 �m.
Cui et al Niacin Induces Axon Growth 3
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from
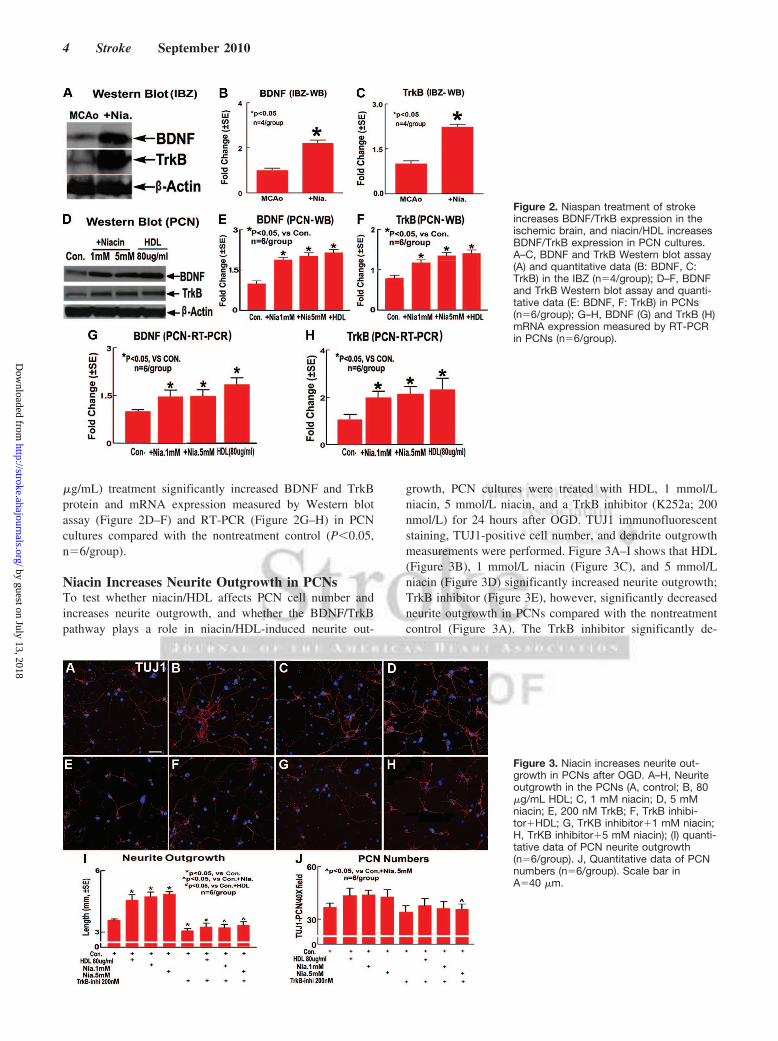
�g/mL) treatment significantly increased BDNF and TrkBprotein and mRNA expression measured by Western blotassay (Figure 2D–F) and RT-PCR (Figure 2G–H) in PCNcultures compared with the nontreatment control (P�0.05,n�6/group).
Niacin Increases Neurite Outgrowth in PCNsTo test whether niacin/HDL affects PCN cell number andincreases neurite outgrowth, and whether the BDNF/TrkBpathway plays a role in niacin/HDL-induced neurite out-
growth, PCN cultures were treated with HDL, 1 mmol/Lniacin, 5 mmol/L niacin, and a TrkB inhibitor (K252a; 200nmol/L) for 24 hours after OGD. TUJ1 immunofluorescentstaining, TUJ1-positive cell number, and dendrite outgrowthmeasurements were performed. Figure 3A–I shows that HDL(Figure 3B), 1 mmol/L niacin (Figure 3C), and 5 mmol/Lniacin (Figure 3D) significantly increased neurite outgrowth;TrkB inhibitor (Figure 3E), however, significantly decreasedneurite outgrowth in PCNs compared with the nontreatmentcontrol (Figure 3A). The TrkB inhibitor significantly de-
Figure 2. Niaspan treatment of strokeincreases BDNF/TrkB expression in theischemic brain, and niacin/HDL increasesBDNF/TrkB expression in PCN cultures.A–C, BDNF and TrkB Western blot assay(A) and quantitative data (B: BDNF, C:TrkB) in the IBZ (n�4/group); D–F, BDNFand TrkB Western blot assay and quanti-tative data (E: BDNF, F: TrkB) in PCNs(n�6/group); G–H, BDNF (G) and TrkB (H)mRNA expression measured by RT-PCRin PCNs (n�6/group).
Figure 3. Niacin increases neurite out-growth in PCNs after OGD. A–H, Neuriteoutgrowth in the PCNs (A, control; B, 80�g/mL HDL; C, 1 mM niacin; D, 5 mMniacin; E, 200 nM TrkB; F, TrkB inhibi-tor�HDL; G, TrKB inhibitor�1 mM niacin;H, TrKB inhibitor�5 mM niacin); (I) quanti-tative data of PCN neurite outgrowth(n�6/group). J, Quantitative data of PCNnumbers (n�6/group). Scale bar inA�40 �m.
4 Stroke September 2010
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from

creased HDL- (Figure 3F) and niacin- (Figure 3G–H) inducedneurite outgrowth in PCNs compared with the niacin-treatedalone group (P�0.05, n�6/group). Niacin and HDL do notaffect PCN number; TrkB inhibitor decreases PCN numbersin niacin-treated PCN compared with niacin alone-treatedPCN (P�0.05, n�6/group). These data indicate that niacintreatment significantly increased neurite outgrowth in PCNs,and niacin-induced neurite outgrowth is, at least partially,mediated by HDL-induced upregulation in BDNF/TrkB.
DiscussionFunctional recovery after acute central nervous system injuryin humans such as stroke is exceptionally limited, leaving theaffected individual with lifelong neurological deficits. Thislack of functional recovery such as motor recovery can, atleast in part, be attributed to the restriction of axon growthand synaptic plasticity.22,23
Cellular cholesterol modulates axon and dendrite out-growth and neuronal polarization under culture condi-tions.24,25 Astrocytes are a major source of HDL-C synthesisin the central nervous system.26 Growing evidence strength-ens the link between brain HDL-C metabolism and factorsinvolved in synaptic plasticity. For example, the scavengerreceptor, Class B, Type I, binds HDL and mediates theselective transfer of cholesteryl esters and �-tocopherol fromcirculating HDL to cells. Aged scavenger receptor, Class B,Type I knockout mice show deficient synaptic plasticity(long-term potentiation) in the CA1 region of the hippocam-pus. Very aged scavenger receptor, Class B, Type I knockoutmice also display selective impairments in recognition mem-ory and spatial memory.5 The cholesterol transporter ATP-binding cassette transporter A1 (ABCA1) also plays a criticalrole in brain cholesterol metabolism. Mice that specificallylacked ABCA1 in the central nervous system exhibit reducedplasma HDL-C levels and changes in synaptic ultrastructure,including reduced synapse and synaptic vesicle numbers.Disturbances in cholesterol transport in the central nervoussystem are associated with structural and functional deficitsin neurons.27 Thus, agents that increase HDL level mayincrease synaptic plasticity and axon growth after stroke.Niacin is the most potent HDL-C-increasing drug used in theclinic. Our previous study showed that Niaspan treatment ofstroke significantly increases serum HDL-C level and pro-motes functional outcome in rats.11 In this study, we find thatNiaspan treatment of stroke significantly decreases NgRexpression, upregulates synaptophysin, and increases axongrowth in the ischemic brain in rats. In addition, HDL andniacin treatment significantly increase neurite outgrowth afterOGD in PCN cultures. Therefore, Niaspan promotes func-tional outcome by increasing HDL-C, which acts in concert topromote synaptic plasticity and axon growth after stroke.
Synaptic plasticity and axon growth induced by niacintreatment of stroke may be mediated by HDL-induced up-regulation in the BDNF/TrkB axis. In the mature nervoussystem, BDNF/TrkB is crucial for regulating neuronal migra-tion, morphological and biochemical differentiation, andcontrolling synaptic function and synaptic plasticity, at thesame time as continuing to modulate neuronal survival.28,29
After somatosensory cortex injury, NgR is downregulated
specifically in cortical areas deprived of sensory input and inadjacent cortex, whereas BDNF is upregulated.30,31 NgRmRNA is downregulated in the dentate gyrus after delivery ofBDNF into the rat hippocampus formation in rats subjected tokainic acid.32 In the present study, niacin treatment of strokesignificantly increases BDNF/TrkB expression both in theischemic brain and in PCN cultures. HDL also significantlyincreases the expression in BDNF/TrkB and neurite out-growth in PCN cultures. In addition, a TrkB inhibitor signif-icantly decreases HDL- and niacin-induced neurite out-growth, which indicates that the BDNF/TrkB axis maymediate, at least in part, niacin/HDL-induced synaptic plas-ticity and axon growth.
SummaryWe demonstrated that niacin treatment of stroke promotessynaptic plasticity and axon growth in rats. The BDNF/TrkBpathways appear to contribute to niacin/HDL-induced synapticplasticity and axon growth after stroke.
AcknowledgmentsWe thank Qinge Lu and Sutapa Santra for technical assistance.
Sources of FundingThis work was supported by National Institute on Aging RO1AG031811 (J.C.), National Institute of Neurological Disorders andStroke PO1 NS23393 (M.C.) and 1R41NS064708 (J.C.), and Amer-ican Heart Association grant 09GRNT2300151 (J.C.).
DisclosuresNone.
References1. Kolb B. Synaptic plasticity and the organization of behaviour after early
and late brain injury. Can J Exp Psychol. 1999;53:62–76.2. Nudo RJ, Plautz EJ, Frost SB. Role of adaptive plasticity in recovery of
function after damage to motor cortex. Muscle Nerve. 2001;24:1000–1019.
3. Adkins DL, Hsu JE, Jones TA. Motor cortical stimulation promotessynaptic plasticity and behavioral improvements following sensorimotorcortex lesions. Exp Neurol. 2008;212:14–28.
4. Keyvani K, Schallert T. Plasticity-associated molecular and structuralevents in the injured brain. J Neuropathol Exp Neurol. 2002;61:831–840.
5. Chang EH, Rigotti A, Huerta PT. Age-related influence of the HDLreceptor SR-BI on synaptic plasticity and cognition. Neurobiol Aging.2009;30:407–419.
6. Li F, Li L, Song XY, Zhong JH, Luo XG, Xian CJ, Zhou XF. Precon-ditioning selective ventral root injury promotes plasticity of ascendingsensory neurons in the injured spinal cord of adult rats—possible roles ofbrain-derived neurotrophic factor, TrkB and p75 neurotrophin receptor.Eur J Neurosci. 2009;30:1280–1296.
7. Gordon T. The role of neurotrophic factors in nerve regeneration. Neu-rosurg Focus. 2009;26:E3.
8. Griesbach GS, Hovda DA, Molteni R, Wu A, Gomez-Pinilla F. Voluntaryexercise following traumatic brain injury: brain-derived neurotrophicfactor upregulation and recovery of function. Neuroscience. 2004;125:129–139.
9. Johnston MV. Plasticity in the developing brain: implications for reha-bilitation. Dev Disabil Res Rev. 2009;15:94–101.
10. Elam MB, Hunninghake DB, Davis KB, Garg R, Johnson C, Egan D,Kostis JB, Sheps DS, Brinton EA. Effect of niacin on lipid and lipoproteinlevels and glycemic control in patients with diabetes and peripheralarterial disease: the ADMIT study: a randomized trial. Arterial DiseaseMultiple Intervention Trial. JAMA. 2000;284:1263–1270.
11. Chen J, Cui X, Zacharek A, Jiang H, Roberts C, Zhang C, Lu M, KapkeA, Feldkamp CS, Chopp M. Niaspan increases angiogenesis andimproves functional recovery after stroke. Ann Neurol. 2007;62:49–58.
Cui et al Niacin Induces Axon Growth 5
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from

12. Chen J, Zacharek A, Cui X, Shehadah A, Jiang H, Roberts C, Lu M,Chopp M. Treatment of stroke with a synthetic liver X receptor agonist,TO901317, promotes synaptic plasticity and axonal regeneration in mice.J Cereb Blood Flow Metab. 2010;30:102–109.
13. Hao Y, Creson T, Zhang L, Li P, Du F, Yuan P, Gould TD, Manji HK,Chen G. Mood stabilizer valproate promotes ERK pathway-dependentcortical neuronal growth and neurogenesis. J Neurosci. 2004;24:6590–6599.
14. Gao Q, Li Y, Chopp M. Bone marrow stromal cells increase astrocytesurvival via upregulation of phosphoinositide 3-kinase/threonine proteinkinase and mitogen-activated protein kinase kinase/extracellular signal-regulated kinase pathways and stimulate astrocyte trophic factor geneexpression after anaerobic insult. Neuroscience. 2005;136:123–134.
15. Halterman MW, Giuliano R, Dejesus C, Schor NF. In-tube transfectionimproves the efficiency of gene transfer in primary neuronal cultures.J Neurosci Methods. 2009;177:348–354.
16. Markus A, Zhong J, Snider WD. Raf and Akt mediate distinct aspects ofsensory axon growth. Neuron. 2002;35:65–76.
17. Chen J, Zhang ZG, Li Y, Wang Y, Wang L, Jiang H, Zhang C, Lu M,Katakowski M, Feldkamp CS, Chopp M. Statins induce angiogenesis,neurogenesis, and synaptogenesis after stroke. Ann Neurol. 2003;53:743–751.
18. Ujike H, Takaki M, Kodama M, Kuroda S. Gene expression related tosynaptogenesis, neuritogenesis, and map kinase in behavioral sensiti-zation to psychostimulants. Ann N Y Acad Sci. 2002;965:55–67.
19. Karnezis T, Mandemakers W, McQualter JL, Zheng B, Ho PP, JordanKA, Murray BM, Barres B, Tessier-Lavigne M, Bernard CC. The neuriteoutgrowth inhibitor Nogo A is involved in autoimmune-mediated demy-elination. Nat Neurosci. 2004;7:736–744.
20. Pluchino S, Quattrini A, Brambilla E, Gritti A, Salani G, Dina G, Galli R,Del Carro U, Amadio S, Bergami A, Furlan R, Comi G, Vescovi AL,Martino G. Injection of adult neurospheres induces recovery in a chronicmodel of multiple sclerosis. Nature. 2003;422:688–694.
21. Peng X, Zhou Z, Hu J, Fink DJ, Mata M Soluble Nogo receptor down-regulates expression of neuronal Nogo-A to enhance axonal regeneration.J Biol Chem. 2009.
22. Walmsley AR, Mir AK. Targeting the Nogo-A signalling pathway topromote recovery following acute CNS injury. Curr Pharm Des. 2007;13:2470–2484.
23. Ramic M, Emerick AJ, Bollnow MR, O’Brien TE, Tsai SY, Kartje GL.Axonal plasticity is associated with motor recovery followingamphetamine treatment combined with rehabilitation after brain injury inthe adult rat. Brain Res. 2006;1111:176–186.
24. Ko M, Zou K, Minagawa H, Yu W, Gong JS, Yanagisawa K, MichikawaM. Cholesterol-mediated neurite outgrowth is differently regulatedbetween cortical and hippocampal neurons. J Biol Chem. 2005;280:42759–42765.
25. Goodrum JF, Brown JC, Fowler KA, Bouldin TW. Axonal regeneration,but not myelination, is partially dependent on local cholesterol reuti-lization in regenerating nerve. J Neuropathol Exp Neurol. 2000;59:1002–1010.
26. Mulder M. Sterols in the central nervous system. Curr Opin Clin NutrMetab Care. 2009;12:152–158.
27. Karasinska JM, Rinninger F, Lutjohann D, Ruddle P, Franciosi S, KruitJK, Singaraja RR, Hirsch-Reinshagen V, Fan J, Brunham LR, Bissada N,Ramakrishnan R, Wellington CL, Parks JS, Hayden MR. Specific loss ofbrain ABCA1 increases brain cholesterol uptake and influences neuronalstructure and function. J Neurosci. 2009;29:3579–3589.
28. Bibel M, Barde YA. Neurotrophins: key regulators of cell fate and cellshape in the vertebrate nervous system. Genes Dev. 2000;14:2919–2937.
29. Huang EJ, Reichardt LF. Neurotrophins: roles in neuronal developmentand function. Annu Rev Neurosci. 2001;24:677–736.
30. Endo T, Spenger C, Tominaga T, Brene S, Olson L. Cortical sensory maprearrangement after spinal cord injury: fMRI responses linked to Nogosignalling. Brain. 2007;130:2951–2961.
31. Josephson A, Trifunovski A, Scheele C, Widenfalk J, Wahlestedt C,Brene S, Olson L, Spenger C. Activity-induced and developmental down-regulation of the Nogo receptor. Cell Tissue Res. 2003;311:333–342.
32. Trifunovski A, Josephson A, Ringman A, Brene S, Spenger C, Olson L.Neuronal activity-induced regulation of Lingo-1. Neuroreport. 2004;15:2397–2400.
6 Stroke September 2010
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from

Jieli ChenXu Cui, Michael Chopp, Alex Zacharek, Cynthia Roberts, Benjamin Buller, Madalina Ion and
Niacin Treatment of Stroke Increases Synaptic Plasticity and Axon Growth in Rats
Print ISSN: 0039-2499. Online ISSN: 1524-4628 Copyright © 2010 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Stroke published online July 29, 2010;Stroke.
http://stroke.ahajournals.org/content/early/2010/07/29/STROKEAHA.110.589333.citationWorld Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://stroke.ahajournals.org//subscriptions/
is online at: Stroke Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer process is available in the
Request Permissions in the middle column of the Web page under Services. Further information about thisOnce the online version of the published article for which permission is being requested is located, click
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office.Strokein Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 13, 2018http://stroke.ahajournals.org/
Dow
nloaded from