novel role for sp1 in phorbol ester enhancement of human
TRANSCRIPT

Novel Role for Sp1 in Phorbol Ester Enhancement of HumanPlatelet Thromboxane Receptor Gene Expression*
(Received for publication, February 27, 1996)
Drew D. D’Angelo, Brian G. Oliver, Michael G. Davis, T. Scott McCluskey, andGerald W. Dorn II‡
From the University of Cincinnati and the Cincinnati Veterans Administration Medical Center,Cincinnati, Ohio 45267-0542
Expression of platelet thromboxane receptors is tran-scriptionally increased duringmegakaryocytic differen-tiation stimulated by phorbol 12-myristate 13-acetate(PMA). We previously cloned and characterized the pro-moter region of the human thromboxane receptor geneand localized PMA-responsive elements to a region be-tween 1.84 and 1.95 kilobase pairs (kb) 5* of the tran-scription initiation site (D’Angelo, D. D., Davis, M. G.,Houser, W. A., Eubank, J. J., Ritchie, M. E., and Dorn, G.W., II (1995) Circ. Res. 77, 466–474). Herein we report thelocalization of the PMA response element to a 14-nucle-otide C-rich sequence, flanked by an octanucleotide in-verted repeat, located 21.938 to 21.925 kb 5* of the tran-scription start site of this gene. We further identify thePMA-responsive enhancer factor that binds to this C-rich sequence as Sp1. Heterologous thromboxane recep-tor gene promoter/thymidilate kinase reporter con-structs transfected into K562 cells exhibited PMAresponsiveness when the C-rich element was includedwith additional 3* sequence from 21.924 to 21.84 kb.However, mutations of the C-rich element that dis-rupted a GC box located on the inverse strand elimi-nated PMA responsiveness and, in gel mobility shift as-says, eliminated binding of Sp1. PMA treatment of K562cells significantly increased, by 5-fold, Sp1 binding tothe C-rich element and increased both phosphorylatedand nonphosphorylated Sp1 protein levels by 2-fold.Furthermore, PMA treatment transiently increased Sp1mRNA levels prior to increasing thromboxane receptormRNA, suggesting that up-regulation of Sp1 contributesto up-regulation of thromboxane receptors. Finally, wehave detected an unidentified K562 nuclear protein thatbinds specifically to the sense strand of the C-rich se-quence overlapping the Sp1 binding site and that, bystabilizing a double stem-loop conformation of this DNAsegment, may also play a role in Sp1 regulation of thisgene. These studies are the first to describe regulatoryand regulated roles for Sp1 in PMA-responsive geneexpression and suggest that modulation of Sp1 levelscontrols thromboxane receptor expression duringmegakaryocytic differentiation.
Thromboxane A2 is one of the most potent platelet-aggregat-ing and vasoconstricting substances known and is crucial formaintenance of normal hemostasis (1). While considerable at-tention has been directed toward measuring increased throm-boxane production in various disease states including acutecoronary ischemic syndromes (2, 3), relatively little is knownabout how target tissue responses to thromboxane may bemodulated. Two intriguing studies have reported increasednumbers of platelet thromboxane receptors, with enhancedplatelet aggregation, in acute myocardial infarction and unsta-ble angina pectoris (4, 5). The observation that this thrombox-ane receptor abnormality was reversible after acute myocardialinfarction (4) and the discovery of an identical abnormality inpatients with unstable angina (the clinical precursor of com-pleted myocardial infarct) (5), strongly suggest that unidenti-fied factors can up-regulate platelet thromboxane receptors invivo and that the resulting platelet hypersensitivity to throm-boxane precedes, and can contribute to, thrombotic coronaryocclusion.Since platelets lack a significant capacity for protein synthe-
sis, platelet thromboxane receptor number must be determinedprior to platelet origination from platelet precursor megakaryo-cytes (6). Therefore, most investigations into the determinantsof platelet thromboxane receptor expression have been carriedout in cultured megakaryoblast-like cells. We previously re-ported that the platelet-like cultured human leukemic cell linesK562 and CHRF-288–11 exhibited increased levels of throm-boxane receptor protein and steady-state mRNA expressionafter megakaryocytic differentiation was stimulated with ei-ther thrombin or phorbol 12-myristate 13-acetate (PMA)1 (7, 8).These findings suggested that physiologic activation of proteinkinase C could enhance thromboxane receptor gene expressionin platelet progenitor cells and ultimately up-regulate plateletthromboxane receptors.In an initial attempt to delineate the mechanism for phorbol
ester-stimulated enhancement of thromboxane receptor geneexpression, we cloned and characterized the promotor region ofthe human thromboxane receptor gene and localized PMA re-sponsiveness to a region located between 1.84 and 1.95 kbupstream of the start of transcription (Ref. 8, GenBankTM
accession number U30503). This region of the thromboxanereceptor gene promotor specifically bound an unidentified K562nuclear protein, the levels of which were increased in nuclearextracts from PMA-treated cells.In the present study we have identified the PMA response
element in the human thromboxane receptor gene promotor,determined the identity of the associated DNA binding factor,
* This work was supported by National Institutes of Health GrantsHL49267 and P50 52318–01, the American Heart Association 1995Council on Circulation Boots Cardiovascular Research Prize, a Univer-sity of Cincinnati Cardiovascular Center Interdepartmental ResearchAward, and a fellowship stipend from the American Heart Association,Ohio affiliate. The costs of publication of this article were defrayed inpart by the payment of page charges. This article must therefore behereby marked “advertisement” in accordance with 18 U.S.C. Section1734 solely to indicate this fact.‡ An Established Investigator of the American Heart Association,
supported with funds contributed in part by its Ohio Affiliate. To whomcorrespondence should be addressed: University of Cincinnati and theCincinnati VA Medical Center, 231 Bethesda Ave., ML 542, Cincinnati,OH 45267-0542.
1 The abbreviations used are: PMA, phorbol 12-myristate 13-acetate;CAT, chloramphenicol acetyltransferase; kb, kilobase pair(s); bp, basepair(s); EMSA, electrophoretic mobility shift assay; AP, activatingprotein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 33, Issue of August 16, pp. 19696–19704, 1996Printed in U.S.A.
19696
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

and have defined a mechanism for PMA enhancement of tran-scription of this gene. We describe a novel action for transcrip-tion factor Sp1 acting as a phorbol ester-responsive transacti-vating factor for the thromboxane receptor gene in K562 cells.These studies represent the first detailed examinations of tran-scriptional regulatory mechanisms for any eicosanoid receptorand the first report of Sp1 involvement in protein kinase Cregulation of gene expression.
EXPERIMENTAL PROCEDURES
Materials—The K562 (chronic myelogenous leukemia) cell line wasobtained from the ATCC (Rockville, MD). The Fast CAT chloramphen-icol acetyltransferase kits were from Molecular Probes (Eugene, OR).All radiochemicals were purchased from Dupont NEN. Oligonucleotideswere synthesized and purified at the University of Cincinnati CoreDNA Facility. Double-stranded oligonucleotides corresponding to tran-scription factor binding sites (AP-1, AP-2, AP-3, Sp1, GRE, NF-1,NKkB, OCT-1, and CRCB) were purchased from Stratagene, La Jolla,CA. Sp1 affinity-purified rabbit polyclonal antibody was purchasedfrom Santa Cruz Biotechnology (Santa Cruz, CA). Purified Sp1 wasfrom Promega (Madison, WI). Unless otherwise indicated, other re-agents were of the highest quality available from Sigma or Life Tech-nologies, Inc.Construction of Thromboxane Gene Promotor/pBLCAT5 Chime-
ras—A numbering system designating the start of transcription atposition 11 will be used throughout (8). Constructs (see Fig. 1, A and B)containing sequences 22127 (22.13/21.84), 22048 (22.05/21.84), and21957 (21.96/21.84) to 21843 were made using restriction enzymesBamHI, PstI, or DraIII, respectively, along with RsaI. The fragmentswere blunt-ended using T4 DNA polymerase and subcloned into thefilled-in XbaI site of pBLCAT5. A construct containing sequences22127 to 21957 (22.13/21.96) was made using restriction enzymesBamHI and DraIII followed by blunting with T4 polymerase and sub-cloning into the filled-in XbaI site of pBLCAT5. A construct spanning21944 to 21919 (21.94/21.92) was engineered using two oligonucleo-tides (sense, 59-AAGCTGCCCCCGCCCCCACCCGCTT-39; antisense,59-AAGCTGGGTGGGGGCGGGGGCAGCTT-39), which were annealedprior to subcloning into the filled-in XbaI site of pBLCAT5. A constructspanning22127 to21919 (22.13/21.92) was engineered by polymerasechain reaction with Pfu polymerase (98 °C for 1 min/50 °C for 1 min/72 °C for 2 min) (primers: sense, 59-ACGGATCCATGGGGAGCT-GGGCACCAG-39; antisense, 59-ACAAGCTTAAGCTGGGTGGGGGCG-GGG-39). The resultant polymerase chain reaction product was cut withBamHI and HindIII and subcloned into BamHI/HindIII-cut pBLCAT5.The mutant construct 21.96/21.84 M was constructed using two over-lapping oligonucleotides (sense, 59-CACCTGGTGTCCTGGAAGC-TGAAAAATAAAAAAAAAAGCTTCCTGACTTTGGCTGTGTCCAGAGCTAAGA-39; antisense, 59-GTACCCCCGTGGCTGCGGTTTCCGGC-GGGGGCCGGCAGGGAG-CGTCTATCTTAGCTCTGGACACAGCCAAAGTCAGGA-39), which were annealed followed by fill-in with T4 DNApolymerase and subcloning into the XbaI-filled-in site of pBLCAT5.During this process a fragment was also obtained that contained se-quences 21923 to 21843 (21.92/21.84), presumably due to exonucleaseactivity of T4 polymerase.Promoter Analysis—Promoter activity in genomic fragments was
characterized using chimeric thromboxane receptor gene-chloramphen-icol acetyltransferase (CAT) constructs in the heterologous thymidilatekinase promotor CAT expression vector pBLCAT5 (9). The chimeraswere transfected in duplicate into K562 cells as described previously(8). Briefly, 10 mg of DNA (total) plus 25 ml of Lipofectamine (LifeTechnologies, Inc.) were incubated at room temperature for 45 min.K562 cells grown to a density of 105 cells/ml in RPMI containing 10%fetal calf serum were washed and resuspended in serum-free RPMI ata density of 3 3 106 cells/ml. The DNA/Lipofectamine mixture wasmixed with 800 ml of cells, aliquoted in one well of a 6-well dish, andplaced in a 95% air, 5% CO2 tissue culture incubator at 37 °C. After 5hours, 4 ml of RPMI, 10% fetal calf serum was added. Unless otherwiseindicated, 100 nM PMA or vehicle (95% ethanol) was added to cells 24 hafter transfection. For basal activity studies, transfection efficiency wasmonitored by cotransfecting pCMVb (1 mg) (Clontech) in which theb-galactosidase gene is driven by the cytomegalovirus promoter. Theresultant CAT activity levels were then normalized for b-galactosidaseactivity.Forty-eight hours following transfection, cells were pelleted by cen-
trifugation and lysed in 0.25 M Tris-HCl, pH 7.4, plus three cycles offreeze-thawing followed by microcentrifugation to remove particulate
matter. The supernatant was heated to 65 °C for 7 min to destroyendogenous acetyltransferase activity and was stored at 270 °C for upto 3 days prior to assay. CAT activity was measured using the Fast CATchloramphenicol acetyltransferase assay kit and the manufacturer’srecommended protocol. Fluorescent acetylated products were resolvedby thin layer chromatography on silica gels, acetylated and nonacety-lated products were separately pooled, and fluorescence was measuredon a Photon Technologies spectrofluorometer (490 nm excitation and512 nm emission). In all studies, transfection of promoterless pBLCAT5served to measure background activity, and pcDNA3CAT (driven by thecytomegalovirus promoter) served as an index of maximal CAT expres-sion by K562 cells.Extract for b-galactosidase activity was prepared as described for
CAT assays except that the 65 °C incubation was omitted. Cell extractwas incubated with 0.26 mg of o-nitrophenyl-b-D-galactoside, 0.1 M
MgCl2, 5 M b-mercaptoethanol at 37 °C for 30 min to 1 h. The reactionwas stopped with 1 M Na2CO3, and b-galactosidase activity was quan-tified by its absorbance at 410 nm.Preparation of K562 Nuclear Extract—Nuclear extract from vehicle
or PMA-treated K562 cells was prepared as described previously (8).Briefly, K562 cells were pelleted and washed in hypotonic buffer fol-lowed by Dounce homogenization. Nuclei were pelleted at 3300 3 g,extracted with 300 mM KCl for 30 min, and dialyzed against 100 mM
KCl followed by centrifugation at 25,000 3 g. Aliquots of nuclear extractwere stored at 270 °C prior to use.DNA Footprinting—DNA footprinting was performed using the Core
Footprinting system (Promega). Briefly, promotor fragment 21.96/21.84 in the EcoRV site of pBluescript (Stratagene, La Jolla, CA) wasexcised with BamHI/XhoI. The fragment was 32P-labeled with T4polynucleotide kinase followed by EcoRI digestion to leave the 39 endlabeled. Probe (50,000 cpm) was incubated with 50 ng of purified Sp1with or without 25 ng of an oligonucleotide corresponding to a genuineSp1 binding site in 50 ml containing 25 mM Tris-HCl, pH 8.0, 50 mM
KCl, 6.75 mMMgCl2, 0.5 mM EDTA, 10% glycerol, 0.5 mM dithiothreitol,10 mM ZnCl2 for 1 h at room temp followed by 10 min on ice. DNase I(0.15 units) was added and incubated for 1 min at room temperature.The resultant products were size-separated on an 8 M urea, 6% poly-acrylamide gel and visualized by autoradiography at 270 °C with in-tensifying screen for 48 h.Gel Mobility Shift Assays—Gel mobility shift assays were performed
essentially as described previously (8). Briefly, 10,000 cpm of 32P-la-beled single- or double-stranded DNA was incubated with 1 mg of K562cell nuclear extract, with or without a 50-fold molar excess of a com-peting oligonucleotide for 1 h. DNA-protein complexes were resolved byelectrophoresis through 5% polyacrylamide and visualized by autora-diography at 270 °C with intensifying screen for 16 h. Relativeamounts of shifted probes were quantitated with an LKB Ultro XL laserdensitometer (Pharmacı̀a Biotech Inc.).Double-stranded probes were constructed by combining equimolar
amounts of sense and antisense strands and heating to 100 °C for 2 minfollowed by slowly cooling to room temperature. Prior to most studiesdouble-stranded probe was purified by electrophoresis through 15%polyacrylamide followed by elution into 10 mM Tris, 0.1 mM EDTA, pH7.4.Depletion of K562 cell nuclear extract of Sp1 was accomplished by
immunoprecipitation of Sp1 protein. 400 mg of K562 cell nuclear extractwas included with 2 mg of Sp1 polyclonal antibody for 1 h at 4 °C.Protein A cell suspension (100 ml of 10% solution) was added andincubated overnight at 4 °C. Precipitate was removed by centrifugation,and supernatant was used in subsequent electrophoretic mobility shiftassay (EMSA) studies.RNA Analysis—RNA was extracted from PMA or vehicle-treated
K562 cells using Chomczynski’s method (10). Poly(A)1 RNA was en-riched by binding to oligo(dT)-cellulose and size-fractionated by electro-phoresis in 1% agarose gels containing 3% formaldehyde before blottingonto Hybond N1 membranes by vacuum transfer. An Sp1 cDNA probe(gift of Dr. Robert Tjian, University of California at Berkeley (11)), athromboxane receptor cDNA probe, or a b-actin cDNA probe (8) was32P-labeled and hybridized to the blot overnight at 42 °C in 50% form-amide/6 3 SSPE (1 3 SSPE is 0.15 M NaCl, 0.015 M Na3 citrate, pH 7.0),0.5% SDS, 100 mg/ml denatured herring sperm DNA. Membranes werewashed at 60 °C in 0.1 3 SSC (1 3 SSC is 0.15 M NaCl, 0.01 M NaH2P04, 0.001 M EDTA, pH 7.4), 0.5% SDS for 1 h and exposed overnight onx-ray film at 270 °C with intensifying screens.Western Analysis—K562 cell nuclear extract (0.5 mg) from vehicle or
PMA-treated cells was size-separated by electrophoresis through a dis-continuous 4–10% SDS-PAGE gel followed by transfer to nitrocellulose.Sp1 was visualized using an affinity-purified rabbit polyclonal antibody
Sp1 and Thromboxane Receptor Gene 19697
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
following the manufacturer’s recommended protocol using an anti-rab-bit horseradish peroxidase-conjugated secondary antibody and a chemi-luminescence detection system (Amersham Corp.). Relative amounts ofSp1 protein in vehicle and PMA-treated K562 cells were quantitatedusing a LKB Ultro XL laser densitometer (Pharmacia).Statistical Methods—Data are reported as mean 6 S.E. unless oth-
erwise stated. If normally distributed, multiple data sets were com-pared using a one way analysis of variance. Significant differencesbetween individual means were determined using a two-tailed group ttest and the Bonferroni procedure. If data were not normally distrib-uted, multiple data sets were compared using one way analysis ofvariance on ranks followed by Dunn’s test for comparison of individualgroups. Statistical significance was assumed at p , 0.05.
RESULTS
The Upstream Region of the Human Thromboxane ReceptorGene Promotor Contains Phorbol Ester-responsive RegulatoryElements—We have previously used human thromboxane re-ceptor gene promotor-CAT chimeras transfected into K562 cellsto localize a position- and orientation-independent phorbol es-ter-responsive element to the 110-bp fragment between 1.84and 1.95 kb upstream of the start of transcription (8). These
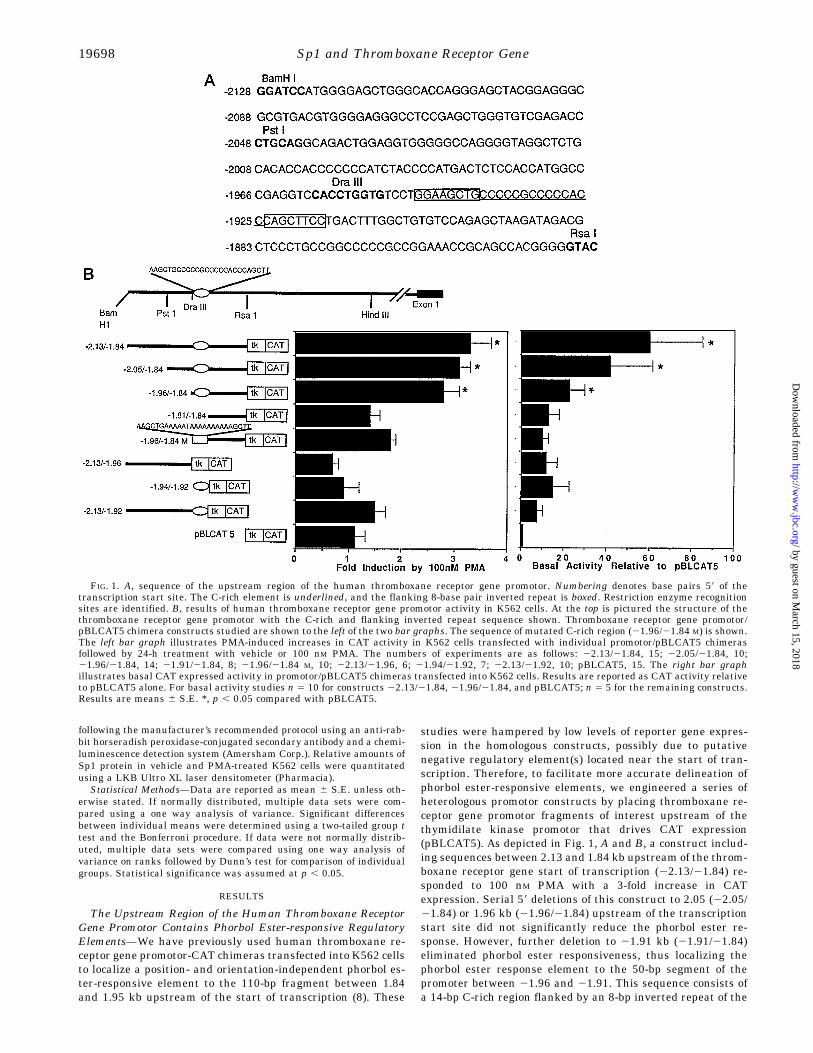
studies were hampered by low levels of reporter gene expres-sion in the homologous constructs, possibly due to putativenegative regulatory element(s) located near the start of tran-scription. Therefore, to facilitate more accurate delineation ofphorbol ester-responsive elements, we engineered a series ofheterologous promotor constructs by placing thromboxane re-ceptor gene promotor fragments of interest upstream of thethymidilate kinase promotor that drives CAT expression(pBLCAT5). As depicted in Fig. 1, A and B, a construct includ-ing sequences between 2.13 and 1.84 kb upstream of the throm-boxane receptor gene start of transcription (22.13/21.84) re-sponded to 100 nM PMA with a 3-fold increase in CATexpression. Serial 59 deletions of this construct to 2.05 (22.05/21.84) or 1.96 kb (21.96/21.84) upstream of the transcriptionstart site did not significantly reduce the phorbol ester re-sponse. However, further deletion to 21.91 kb (21.91/21.84)eliminated phorbol ester responsiveness, thus localizing thephorbol ester response element to the 50-bp segment of thepromoter between 21.96 and 21.91. This sequence consists ofa 14-bp C-rich region flanked by an 8-bp inverted repeat of the
FIG. 1. A, sequence of the upstream region of the human thromboxane receptor gene promotor. Numbering denotes base pairs 59 of thetranscription start site. The C-rich element is underlined, and the flanking 8-base pair inverted repeat is boxed. Restriction enzyme recognitionsites are identified. B, results of human thromboxane receptor gene promotor activity in K562 cells. At the top is pictured the structure of thethromboxane receptor gene promotor with the C-rich and flanking inverted repeat sequence shown. Thromboxane receptor gene promotor/pBLCAT5 chimera constructs studied are shown to the left of the two bar graphs. The sequence of mutated C-rich region (21.96/21.84 M) is shown.The left bar graph illustrates PMA-induced increases in CAT activity in K562 cells transfected with individual promotor/pBLCAT5 chimerasfollowed by 24-h treatment with vehicle or 100 nM PMA. The numbers of experiments are as follows: 22.13/21.84, 15; 22.05/21.84, 10;21.96/21.84, 14; 21.91/21.84, 8; 21.96/21.84 M, 10; 22.13/21.96, 6; 21.94/21.92, 7; 22.13/21.92, 10; pBLCAT5, 15. The right bar graphillustrates basal CAT expressed activity in promotor/pBLCAT5 chimeras transfected into K562 cells. Results are reported as CAT activity relativeto pBLCAT5 alone. For basal activity studies n 5 10 for constructs 22.13/21.84, 21.96/21.84, and pBLCAT5; n 5 5 for the remaining constructs.Results are means 6 S.E. *, p , 0.05 compared with pBLCAT5.
Sp1 and Thromboxane Receptor Gene19698
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
sequence 59-GGAAGCTG-39 (Fig. 1A). Mutation of the C-richregion (C changed to A and G changed to T) in the context of the21.96/21.84 construct (21.96/21.84 M) eliminated phorbol es-ter responsiveness, thus demonstrating the critical importanceof the C-rich element in PMA enhancement of thromboxanereceptor gene activity. A construct containing only the C-richregion plus 6 bp of the flanking inverted repeat (21.94/21.92)exhibited no phorbol ester responsiveness (although it en-
hanced basal activity of the thymidilate kinase promoter; Fig.1B). In contrast to 21.96/21.84, which included the C-richelement plus 60 bp of downstream sequence and was sufficientfor phorbol ester responsiveness, inclusion of the C-rich ele-ment plus 90 bp of upstream sequence (22.13/21.92) did notconfer phorbol ester responsiveness. Not surprisingly, the up-stream sequence alone (22.13/21.96) also had no phorbol ester
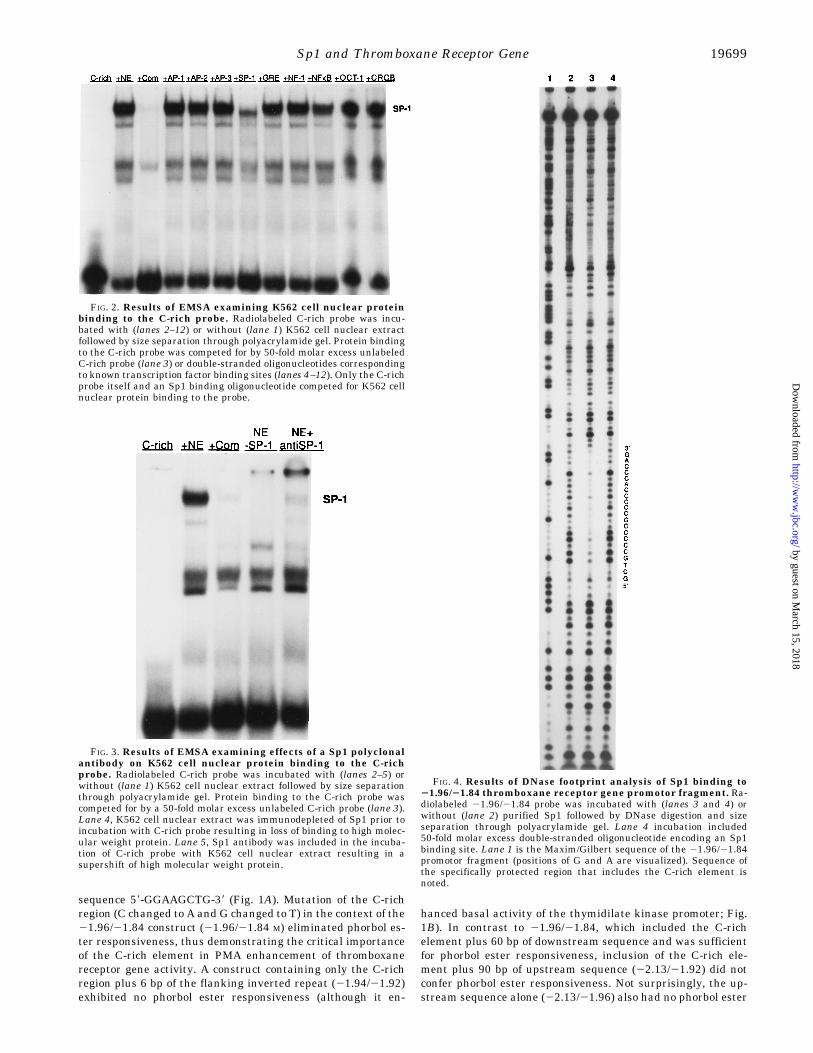
FIG. 2. Results of EMSA examining K562 cell nuclear proteinbinding to the C-rich probe. Radiolabeled C-rich probe was incu-bated with (lanes 2–12) or without (lane 1) K562 cell nuclear extractfollowed by size separation through polyacrylamide gel. Protein bindingto the C-rich probe was competed for by 50-fold molar excess unlabeledC-rich probe (lane 3) or double-stranded oligonucleotides correspondingto known transcription factor binding sites (lanes 4–12). Only the C-richprobe itself and an Sp1 binding oligonucleotide competed for K562 cellnuclear protein binding to the probe.
FIG. 3. Results of EMSA examining effects of a Sp1 polyclonalantibody on K562 cell nuclear protein binding to the C-richprobe. Radiolabeled C-rich probe was incubated with (lanes 2–5) orwithout (lane 1) K562 cell nuclear extract followed by size separationthrough polyacrylamide gel. Protein binding to the C-rich probe wascompeted for by a 50-fold molar excess unlabeled C-rich probe (lane 3).Lane 4, K562 cell nuclear extract was immunodepleted of Sp1 prior toincubation with C-rich probe resulting in loss of binding to high molec-ular weight protein. Lane 5, Sp1 antibody was included in the incuba-tion of C-rich probe with K562 cell nuclear extract resulting in asupershift of high molecular weight protein.
FIG. 4. Results of DNase footprint analysis of Sp1 binding to21.96/21.84 thromboxane receptor gene promotor fragment. Ra-diolabeled 21.96/21.84 probe was incubated with (lanes 3 and 4) orwithout (lane 2) purified Sp1 followed by DNase digestion and sizeseparation through polyacrylamide gel. Lane 4 incubation included50-fold molar excess double-stranded oligonucleotide encoding an Sp1binding site. Lane 1 is the Maxim/Gilbert sequence of the 21.96/21.84promotor fragment (positions of G and A are visualized). Sequence ofthe specifically protected region that includes the C-rich element isnoted.
Sp1 and Thromboxane Receptor Gene 19699
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
responsiveness. Taken together, these results indicated thatphorbol ester up-regulation of the thromboxane receptor generequired the C-rich element located between 21.938 and21.925 kb upstream of the start of transcription. Furthermore,this element alone was not sufficient to confer phorbol esterresponsiveness but appeared to work in concert with otherelements located between 1.92 and 1.84 kb upstream of thetranscription start site.Identification of a K562 Nuclear DNA Binding Protein That
Binds to the C-rich Phorbol Ester-responsive Element of theHuman Thromboxane Receptor Gene Promotor as Sp1—Ourfunctional studies indicated that a C-rich element between21.938 and 21.925 was required for phorbol ester responsive-ness of the thromboxane receptor gene. Computer analysis (12)suggested a potential AP-2 transcription factor binding site onthe forward strand of this sequence, while the backward strandcontained a GC box. In order to identify which transcription
factor(s) from K562 cells bound to the C-rich element we useda double-stranded oligonucleotide corresponding to the C-richsequence plus 6 bp of the flanking inverted repeat (C-richprobe) in EMSAs. C-rich probe bound to four major proteinsfrom K562 nuclear extract. Competition for C-rich probe bind-ing to the lowest and two higher molecular weight proteins waseliminated when a 50-fold molar excess of unlabeled probe wasadded to the reaction, while binding to an intermediate molec-ular weight protein was less efficiently displaced, suggestingthat the intermediate protein bound in a less specific manner(Fig. 2). As a first approach to identifying the shifted K562nuclear proteins, oligonucleotides corresponding to a number ofknown transcription factor binding sites were used as compet-itors. These studies revealed that only the Sp1 binding oligo-nucleotide was an effective competitor for C-rich probe binding,and the competition appeared to be most efficient for binding ofthe two higher molecular weight proteins (Fig. 2).
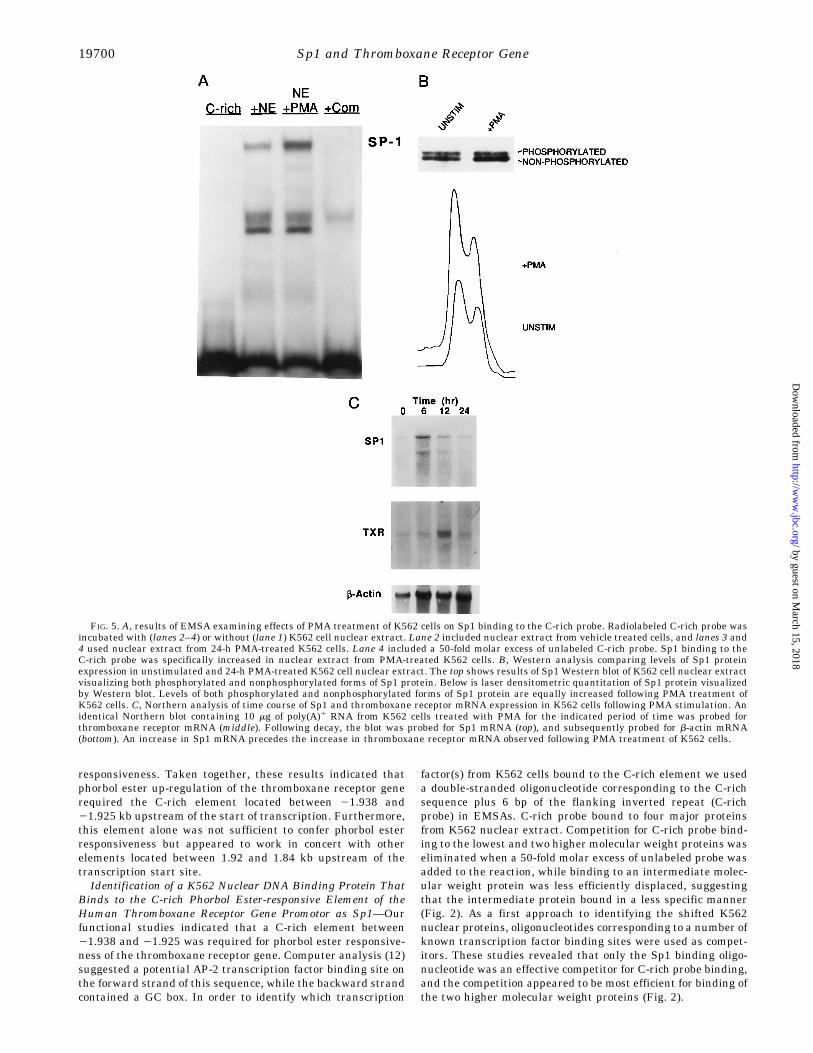
FIG. 5. A, results of EMSA examining effects of PMA treatment of K562 cells on Sp1 binding to the C-rich probe. Radiolabeled C-rich probe wasincubated with (lanes 2–4) or without (lane 1) K562 cell nuclear extract. Lane 2 included nuclear extract from vehicle treated cells, and lanes 3 and4 used nuclear extract from 24-h PMA-treated K562 cells. Lane 4 included a 50-fold molar excess of unlabeled C-rich probe. Sp1 binding to theC-rich probe was specifically increased in nuclear extract from PMA-treated K562 cells. B, Western analysis comparing levels of Sp1 proteinexpression in unstimulated and 24-h PMA-treated K562 cell nuclear extract. The top shows results of Sp1 Western blot of K562 cell nuclear extractvisualizing both phosphorylated and nonphosphorylated forms of Sp1 protein. Below is laser densitometric quantitation of Sp1 protein visualizedby Western blot. Levels of both phosphorylated and nonphosphorylated forms of Sp1 protein are equally increased following PMA treatment ofK562 cells. C, Northern analysis of time course of Sp1 and thromboxane receptor mRNA expression in K562 cells following PMA stimulation. Anidentical Northern blot containing 10 mg of poly(A)1 RNA from K562 cells treated with PMA for the indicated period of time was probed forthromboxane receptor mRNA (middle). Following decay, the blot was probed for Sp1 mRNA (top), and subsequently probed for b-actin mRNA(bottom). An increase in Sp1 mRNA precedes the increase in thromboxane receptor mRNA observed following PMA treatment of K562 cells.
Sp1 and Thromboxane Receptor Gene19700
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Additional experiments were performed to determine if thehigh molecular weight K562 nuclear protein that bound toC-rich probe was Sp1. First, when EMSA was performed withK562 nuclear extract immunodepleted of Sp1, the two highermolecular weight DNA binding proteins were no longer ob-served (Fig. 3, lane 4). Second, when the anti-Sp1 antibody wasincluded in the EMSA reaction, the two higher molecularweight bands “supershifted,” indicating that the antibody rec-ognized those DNA-protein complexes (Fig. 3, lane 5). Finally,when authentic Sp1 was used in EMSA studies with the C-richprobe, it shifted it in a manner identical to the higher molecu-lar weight K562 nuclear protein (not shown). These studiesconfirmed that Sp1 from K562 cell nuclear extract bound to theC-rich element and was responsible for the high molecularweight bands observed on EMSA. To determine the exact site ofSp1 binding we performed DNase footprinting using promotorfragment 21.96/21.84 and purified Sp1 protein (Fig. 4). Sp1protected the region from 21.942 to 21.922 kb from DNase
digestion. This region corresponds to the sequence 59-GCTGC-CCCCGCCCCCACCCAG-39, which includes the complete C-rich element (underlined). Taken together with our functionalstudies of phorbol ester response (see above), the DNA bindingstudies suggested that a critical feature required for PMAenhancement of thromboxane receptor gene expression wasSp1 binding to the C-rich element.Sp1 Is Up-regulated by Phorbol Ester in K562 Cells—Sp1
had not previously been implicated as mediating phorbol esterenhancement of gene expression. Therefore, we used EMSA todetermine if Sp1 binding to the thromboxane receptor genepromoter C-rich region differed quantitatively in nuclear ex-tracts from K562 cells treated with vehicle compared with 100nM PMA. PMA treatment increased Sp1 binding to the C-richprobe 5.4 6 1.2-fold over vehicle (p 5 0.02, n 5 4) (Fig. 5A). Incontrast, binding to the intermediate nuclear weight proteindid not change, thus demonstrating equal loading of nuclearextract. In addition, Sp1 protein levels, assessed by Westernblot analysis, increased 2.4 6 0.1-fold (p 5 0.002, n 5 3) inK562 cells stimulated with PMA (Fig. 5B). Interestingly, therewas an equivalent increase in both the phosphorylated andnonphosphorylated forms of Sp1 after PMA treatment. Thus,increased Sp1 binding to the thromboxane receptor gene pro-motor appeared to be due to an increase in Sp1 protein levels,not phosphorylation of existing protein. Finally, Northern anal-ysis showed that Sp1 mRNA levels increased following PMAtreatment of K562 cells and that the increase in Sp1 mRNApreceded the phorbol ester-stimulated increase in thromboxanereceptor mRNA (Fig. 5C). In aggregate, these results supportthe notion that phorbol ester treatment of K562 cells increasesthromboxane receptor gene transcription in part by increasingSp1 expression and DNA binding.Specific Nucleotide Alterations in the Phorbol Ester-respon-
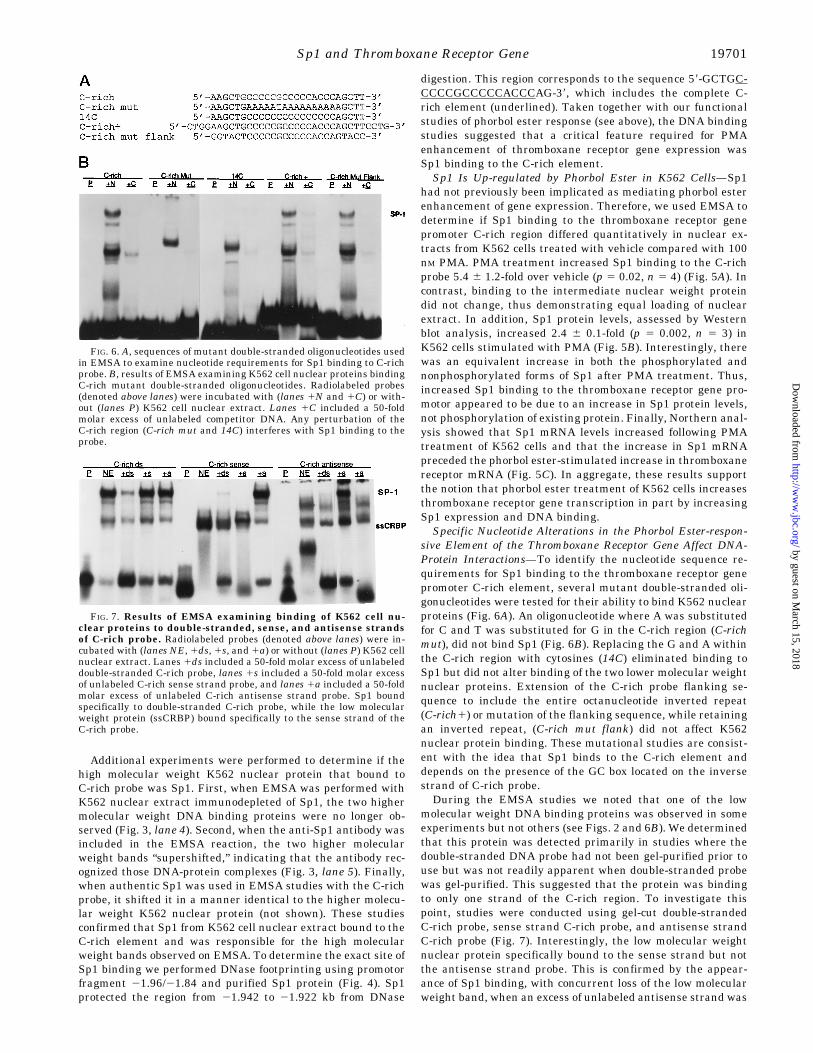
sive Element of the Thromboxane Receptor Gene Affect DNA-Protein Interactions—To identify the nucleotide sequence re-quirements for Sp1 binding to the thromboxane receptor genepromoter C-rich element, several mutant double-stranded oli-gonucleotides were tested for their ability to bind K562 nuclearproteins (Fig. 6A). An oligonucleotide where A was substitutedfor C and T was substituted for G in the C-rich region (C-richmut), did not bind Sp1 (Fig. 6B). Replacing the G and A withinthe C-rich region with cytosines (14C) eliminated binding toSp1 but did not alter binding of the two lower molecular weightnuclear proteins. Extension of the C-rich probe flanking se-quence to include the entire octanucleotide inverted repeat(C-rich1) or mutation of the flanking sequence, while retainingan inverted repeat, (C-rich mut flank) did not affect K562nuclear protein binding. These mutational studies are consist-ent with the idea that Sp1 binds to the C-rich element anddepends on the presence of the GC box located on the inversestrand of C-rich probe.During the EMSA studies we noted that one of the low
molecular weight DNA binding proteins was observed in someexperiments but not others (see Figs. 2 and 6B). We determinedthat this protein was detected primarily in studies where thedouble-stranded DNA probe had not been gel-purified prior touse but was not readily apparent when double-stranded probewas gel-purified. This suggested that the protein was bindingto only one strand of the C-rich region. To investigate thispoint, studies were conducted using gel-cut double-strandedC-rich probe, sense strand C-rich probe, and antisense strandC-rich probe (Fig. 7). Interestingly, the low molecular weightnuclear protein specifically bound to the sense strand but notthe antisense strand probe. This is confirmed by the appear-ance of Sp1 binding, with concurrent loss of the low molecularweight band, when an excess of unlabeled antisense strand was
FIG. 6. A, sequences of mutant double-stranded oligonucleotides usedin EMSA to examine nucleotide requirements for Sp1 binding to C-richprobe. B, results of EMSA examining K562 cell nuclear proteins bindingC-rich mutant double-stranded oligonucleotides. Radiolabeled probes(denoted above lanes) were incubated with (lanes 1N and 1C) or with-out (lanes P) K562 cell nuclear extract. Lanes 1C included a 50-foldmolar excess of unlabeled competitor DNA. Any perturbation of theC-rich region (C-rich mut and 14C) interferes with Sp1 binding to theprobe.
FIG. 7. Results of EMSA examining binding of K562 cell nu-clear proteins to double-stranded, sense, and antisense strandsof C-rich probe. Radiolabeled probes (denoted above lanes) were in-cubated with (lanes NE, 1ds, 1s, and 1a) or without (lanes P) K562 cellnuclear extract. Lanes 1ds included a 50-fold molar excess of unlabeleddouble-stranded C-rich probe, lanes 1s included a 50-fold molar excessof unlabeled C-rich sense strand probe, and lanes 1a included a 50-foldmolar excess of unlabeled C-rich antisense strand probe. Sp1 boundspecifically to double-stranded C-rich probe, while the low molecularweight protein (ssCRBP) bound specifically to the sense strand of theC-rich probe.
Sp1 and Thromboxane Receptor Gene 19701
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from
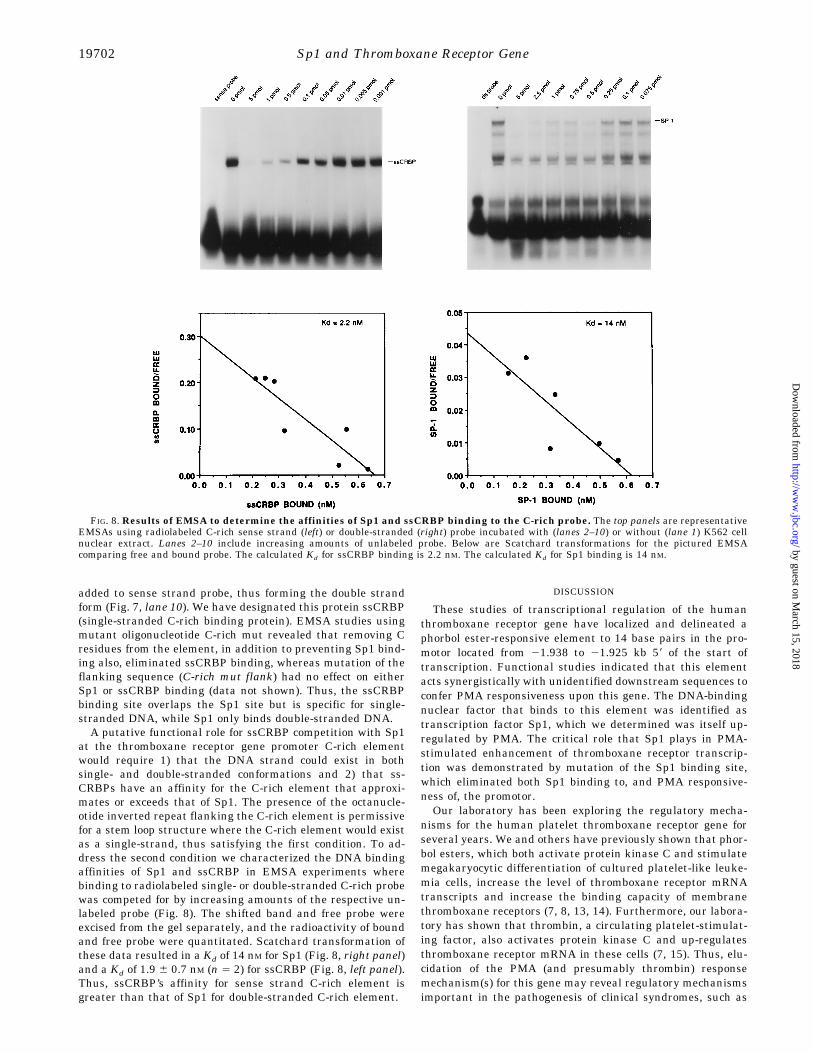
added to sense strand probe, thus forming the double strandform (Fig. 7, lane 10). We have designated this protein ssCRBP(single-stranded C-rich binding protein). EMSA studies usingmutant oligonucleotide C-rich mut revealed that removing Cresidues from the element, in addition to preventing Sp1 bind-ing also, eliminated ssCRBP binding, whereas mutation of theflanking sequence (C-rich mut flank) had no effect on eitherSp1 or ssCRBP binding (data not shown). Thus, the ssCRBPbinding site overlaps the Sp1 site but is specific for single-stranded DNA, while Sp1 only binds double-stranded DNA.A putative functional role for ssCRBP competition with Sp1
at the thromboxane receptor gene promoter C-rich elementwould require 1) that the DNA strand could exist in bothsingle- and double-stranded conformations and 2) that ss-CRBPs have an affinity for the C-rich element that approxi-mates or exceeds that of Sp1. The presence of the octanucle-otide inverted repeat flanking the C-rich element is permissivefor a stem loop structure where the C-rich element would existas a single-strand, thus satisfying the first condition. To ad-dress the second condition we characterized the DNA bindingaffinities of Sp1 and ssCRBP in EMSA experiments wherebinding to radiolabeled single- or double-stranded C-rich probewas competed for by increasing amounts of the respective un-labeled probe (Fig. 8). The shifted band and free probe wereexcised from the gel separately, and the radioactivity of boundand free probe were quantitated. Scatchard transformation ofthese data resulted in a Kd of 14 nM for Sp1 (Fig. 8, right panel)and a Kd of 1.9 6 0.7 nM (n 5 2) for ssCRBP (Fig. 8, left panel).Thus, ssCRBP’s affinity for sense strand C-rich element isgreater than that of Sp1 for double-stranded C-rich element.
DISCUSSION
These studies of transcriptional regulation of the humanthromboxane receptor gene have localized and delineated aphorbol ester-responsive element to 14 base pairs in the pro-motor located from 21.938 to 21.925 kb 59 of the start oftranscription. Functional studies indicated that this elementacts synergistically with unidentified downstream sequences toconfer PMA responsiveness upon this gene. The DNA-bindingnuclear factor that binds to this element was identified astranscription factor Sp1, which we determined was itself up-regulated by PMA. The critical role that Sp1 plays in PMA-stimulated enhancement of thromboxane receptor transcrip-tion was demonstrated by mutation of the Sp1 binding site,which eliminated both Sp1 binding to, and PMA responsive-ness of, the promotor.Our laboratory has been exploring the regulatory mecha-
nisms for the human platelet thromboxane receptor gene forseveral years. We and others have previously shown that phor-bol esters, which both activate protein kinase C and stimulatemegakaryocytic differentiation of cultured platelet-like leuke-mia cells, increase the level of thromboxane receptor mRNAtranscripts and increase the binding capacity of membranethromboxane receptors (7, 8, 13, 14). Furthermore, our labora-tory has shown that thrombin, a circulating platelet-stimulat-ing factor, also activates protein kinase C and up-regulatesthromboxane receptor mRNA in these cells (7, 15). Thus, elu-cidation of the PMA (and presumably thrombin) responsemechanism(s) for this gene may reveal regulatory mechanismsimportant in the pathogenesis of clinical syndromes, such as
FIG. 8. Results of EMSA to determine the affinities of Sp1 and ssCRBP binding to the C-rich probe. The top panels are representativeEMSAs using radiolabeled C-rich sense strand (left) or double-stranded (right) probe incubated with (lanes 2–10) or without (lane 1) K562 cellnuclear extract. Lanes 2–10 include increasing amounts of unlabeled probe. Below are Scatchard transformations for the pictured EMSAcomparing free and bound probe. The calculated Kd for ssCRBP binding is 2.2 nM. The calculated Kd for Sp1 binding is 14 nM.
Sp1 and Thromboxane Receptor Gene19702
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

acute myocardial infarction or unstable angina pectoris, inwhich both thrombin and the number of platelet thromboxanereceptors are increased (4, 16, 17).Recently, we reported the initial characterization of the hu-
man (K562 cell) thromboxane receptor gene promoter (8). Al-though our 2-kb promoter constructs were phorbol ester-re-sponsive, we could not identify a classic Fos-Jun (AP-1) bindingelement within that region of the promoter. However, severalpotential AP-2 binding sites were identified, which we postu-lated could transduce the phorbol-ester response of this gene.Our initial experiments with the heterologous thromboxanereceptor gene promoter/thymidilate kinase constructs de-scribed herein seemed to indicate that one of the putative AP-2sites, located between 21.94 and 21.92, was the PMA responseelement. However, subsequent studies indicated that AP-2 wasnot the responsible transcription factor. First, we could notdetect AP-2 in immunoblots of K562 nuclear extract.2 Second,performance of EMSA using this element and an excess ofunlabeled AP-2-binding probe failed to compete for K562 nu-clear protein binding (see Fig. 2). Finally, anti-AP-2 antibodydid not supershift any of the bands noted on EMSA.2 In con-trast, Sp1 was identified as the transcription factor that boundto the PMA response element in the thromboxane receptor genepromoter, and interestingly, the opposite strand of the putativeAP-2 site contained an Sp1-binding GC box.
Sp1 is a widely expressed RNA polymerase II transcriptionfactor, originally isolated from HeLa cells (18). The tradition-ally accepted role for Sp1 is that it binds to one or more GCboxes (consensus 59-GGGCGG-39) located near a transcriptionstart site and, in concert with other members of the RNApolymerase complex, initiates transcription (19–21). Althoughmany genes regulated by Sp1 contain multiple GC boxes, asingle Sp1 binding site is generally sufficient for transcrip-tional activation (11). While the TATA-less thromboxane recep-tor gene promotor studied herein contains GC boxes located inthe proximity of its transcription start site (on forward orreverse strand at positions 2176 to 2171, 102–107, and 272–277 (8)), our studies determined that an Sp1 binding site lo-cated approximately 1.9 kb 59 of the start of transcriptionfunctioned as an enhancer of promotor activity after PMAstimulation. This unexpected finding implies that Sp1 has verydifferent regulatory roles when it binds to upstream promoterregions compared with binding at or near the transcriptionstart site. This notion is supported by a previous report that aCAT reporter construct containing an Sp1 binding GC box 1.7kb from the transcription start site can act as a transcriptionalactivator in Drosophila SL2 cells, but only under conditionswhere Sp1 protein expression is increased (22). In short, undersome circumstances Sp1 can act as a transcriptional enhancerindependent of its actions in transcriptional initiation.Our studies indicated that Sp1 binding alone is not sufficient
for PMA enhancement of human thromboxane receptor geneexpression but that additional downstream elements locatedbetween 21.91 and 21.84 are also required. It has previouslybeen reported that Sp1 interacts with other transcription fac-tors, including GATA-1, YY-1 and NF-kB (23–25). In the case ofGATA-1, this interaction synergistically enhances transcrip-tion of globin genes during erythroid cell differentiation andrequires increased levels of Sp1 expression (25). In vitro studiesof Sp1-regulated promoter activity have also indicated a re-quirement for coactivators that are thought to tether Sp1 to thebasal transcriptional machinery (26). In a similar manner, Sp1apparently requires an as yet unidentified co-factor to functionas a PMA-responsive transcriptional enhancer of the throm-boxane receptor gene.Our finding of a single-strand-specific factor (ssCRBP) whose
DNA binding site overlaps that of Sp1 raises an interestingpossibility regarding the mechanism of PMA responsiveness inthe thromboxane receptor gene. The region flanking the Sp1/ssCRBP binding site contains an octanucleotide inverted re-peat sequence that would allow the formation of a DNA stem-loop structure such that the Sp1/ssCRBP binding site exists asa single strand permissive for ssCRBP binding, but not Sp1binding, and therefore resulting in no transcriptional enhance-ment (Fig. 9). Single- and doubled-stranded DNA configura-tions may exist in a dynamic equilibrium, with the predomi-nant form determined by the relative levels of Sp1 andssCRBP. In this scenario, increased Sp1 expression followingPMA treatment would result in a shift of this equilibriumtoward the Sp1 binding linear form of the DNA with resultanttranscriptional enhancement. Interestingly, we found thatPMA treatment did not alter ssCRBP binding to the C-richelement. Thus, regulation of Sp1 levels appears to be the majorfactor influencing the equilibrium between Sp1 and ssCRBP. Asimilar model has been proposed for the rat preprotachyki-nin-A promotor where a region functionally shown to enhancetranscription can form a stem-loop structure (27). This regioncan bind Sp1 and AP-2 when double-stranded, but the singlesense strand binds an unidentified single-strand-specific pro-tein. An element that regulates myoblast and fibroblast vascu-lar smooth muscle a-actin gene expression has also been iden-
2 D. D. D’Angelo, B. G. Oliver, M. G. Davis, T. S. McCluskey, and G.W. Dorn II, unpublished observations.
FIG. 9. Stem-loop model of competitive Sp1 and ssCRBP bind-ing to the C-rich sequence. In the stem-loop configuration (top) theC-rich element exists as a single strand permissive to ssCRBP binding,resulting in transcriptional repression. An increase in Sp1 proteinexpression, as observed following PMA treatment, would result in ashift in the equilibrium toward increased Sp1 binding and formation ofthe linear form of DNA (bottom) with subsequent transcriptionalenhancement.
Sp1 and Thromboxane Receptor Gene 19703
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

tified in which double-stranded DNA binds transcriptionenhancer factor-1, while both the sense and antisense strandsalone can bind other unidentified nuclear proteins (28).The current paper and a number of other recent reports have
modified our ideas regarding the role of Sp1 in gene expression,and it appears that an Sp1 mechanism of transcriptional en-hancement, such as that demonstrated for the human throm-boxane receptor gene, may be widespread. The regulatory ef-fects of Sp1 for globin gene expression during erythroid celldifferentiation were noted above (25). A similar requirementfor Sp1 has been reported for induction of the skeletal a-actinpromoter during cardiac myocyte hypertrophy (29). A regula-tory role for Sp1 has now been reported for epidermal growthfactor stimulation of the human gastrin promoter (30), cAMP-induced transcription of the bovine cyp11A gene (31) and glu-cose activation of the acetyl-CoA carboxylase promoter (32). Wehave performed preliminary studies that indicate that glu-cocorticoids, like PMA, increase K562 thromboxane receptorgene expression while increasing Sp1 mRNA expression, Sp1protein levels, and Sp1 binding to the C-rich element.3 Thethromboxane receptor gene promotor contains no glucocorticoidresponse element (8), further suggesting that Sp1 expressioncan itself be regulated by glucocorticoids as well as phorbolesters and that Sp1 modulation is a regulatory determinant ofthe expression of this and other genes.In conclusion, far from being a ubiquitously expressed
“housekeeping” transcription factor, Sp1 is a dynamically reg-ulated enhancer that appears to play a critical regulatory rolefor numerous genes and may be a final common pathway forthe regulation of gene expression by a variety of independentstimuli. Further characterizing the roles of Sp1 and ssCRBP inthromboxane receptor gene expression and determining theidentities of necessary co-factors is the object of ongoing inves-tigation in our laboratory.
REFERENCES
1. Hamberg, M., Svensson, J., and Samuelsson, B. (1975) Proc. Natl. Acad. Sci.U. S. A. 72, 2994–2998
2. Hirsh, P. D., Hillis, L. D., Campbell, W. B., Firth, B. G., and Willerson, J. T.(1977) N. Engl. J. Med. 304, 685–691
3. Fitzgerald, D. J., Roy, L., Catella, F., and FitzGerald, G. A. (1986) N. Engl.J. Med. 315, 983–990
4. Dorn, G. W., II, Liel, N., Trask, J. L., Mais, D. E., Assey, M. E., and Halushka,P. V. (1990) Circulation 81, 212–218
5. Modesti, P. A., Colella, A., Cecioni, I., Costoli, A., Biagini, D., Migliorini, A.,and Neri Serneri, G. G. (1995) Am. Heart. J. 129, 873–879
6. Dorn, G. W., II (1992) Am. J. Physiol. 262, C991–C9997. Dorn, G. W., II, Davis, M. G., and D’Angelo, D. D. (1994) Am. J. Physiol. 266,
C1231–C12398. D’Angelo, D. D., Davis, M. G., Houser, W. A., Eubank, J. J., Ritchie, M. E., and
Dorn, G. W., II (1995) Circ. Res. 77, 466–4749. Boshart, M., Kluppel, M., Schmidt, A., Schutz, G., and Luckow, B. (1992) Gene.
110, 129–13010. Chomczynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156–15911. Kadonaga, J. T., Carner, K. R., Masiarz, F. R., and Tjian, R. (1987) Cell. 51,
1079–109012. Hitachi Software Engineering (1993) HIBIO MacDNASIS Pro-DNA and Pro-
tein Sequence Analysis System, Version 3.0, Hitachi Software Engineering13. Kinsella, B. T., O’Mahony, D. J., and FitzGerald, G. A. (1994) J. Biol. Chem.
269, 29914–2991914. Nakajima, M., Yamamoto, M., Ushikubi, F., Okuma, M., Fujiware, M., and
Narumiya, S. (1989) Biochem. Biophys. Res. Commun. 158, 958–96515. Dorn, G. W., II, and Davis, M. G. (1992) Am. J. Physiol. 263, C864–C87216. Szczeklik, A., Dropinski, J., Radwan, J., and Krzanowski, M. (1992) Arterio-
scler. Thromb. 12, 548–55317. Granger, C. B., Miller, J. M., Bovill, E. G., Gruber, A., Tracy, R. P., Krucoff, M.
W., Green, C., Berrios, E., Harrington, R. A., Ohman, E. M., and Califf, R.M. (1995) Circulation 91, 1929–1935
18. Dynan, W. S., and Tjian, R. (1983) Cell 32, 669–68019. Dynan, W. S., and Tjian, R. (1983) Cell 35, 79–8720. Gidoni, D., Kadonaga, J. T., Barrera-Saldana, H., Takahashi, K., Chambon, P.,
and Tjian, R. (1985) Science 230, 511–51721. Kadonaga, J. T., Jones, K. A., and Tjian, R. (1986) Trends Biochem. 11, 20–2322. Courey, A. J., Holtzman, D. A., Jackson, S. P., and Tjian, R. (1989) Cell. 59,
827–83623. Perkins, N. D., Agranoff, A. B., Pascal, E., and Nabel, G. J. (1994) Mol. Cell.
Biol. 14, 6570–658324. Lee, J. S., Galvin, K. M., and Shi, Y. (1993) Proc. Natl. Acad. Sci. U. S. A. 90,
6145–614925. Merika, M., and Orkin, S. H. (1995) Mol. Cell. Biol. 15, 2437–244726. Pugh, B. F., and Tjian, R. (1990) Cell. 61, 1187–119727. Quinn, J. P., McAllister, J., and Mendelson, S. (1995) Biochim. Biophys. Acta
1263, 25–3428. Sun, S., Stoflet, E. S., Cogan, J. G., Strauch, A. R., and Getz, M. J. (1995)Mol.
Cell. Biol. 15, 2429–243629. Karns, L. R., Kariya, K., and Simpson, P. C. (1995) J. Biol. Chem. 270,
410–41730. Merchant, J. L., Shiotani, A., Mortensen, E. R., Shumaker, D. K., and Abrac-
zinskas, D. R. (1995) J. Biol. Chem. 270, 6314–631931. Venepally, P., and Waterman, M. R. (1995) J. Biol. Chem. 270, 25402–2541032. Daniel, S., and Kim, K-H. (1996) J. Biol. Chem. 271, 1385–13923 D. D. D’Angelo and G. W. Dorn II, unpublished results.
Sp1 and Thromboxane Receptor Gene19704
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Dorn IIDrew D. D'Angelo, Brian G. Oliver, Michael G. Davis, T. Scott McCluskey and Gerald W.
Receptor Gene ExpressionNovel Role for Sp1 in Phorbol Ester Enhancement of Human Platelet Thromboxane
doi: 10.1074/jbc.271.33.196961996, 271:19696-19704.J. Biol. Chem.
http://www.jbc.org/content/271/33/19696Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/271/33/19696.full.html#ref-list-1
This article cites 31 references, 15 of which can be accessed free at
by guest on March 15, 2018
http://ww
w.jbc.org/
Dow
nloaded from