nutrient ions of plants and the ion activation of plant enzymes.* the role of salts
TRANSCRIPT

NUTRIENT IONS OF PLANTS AND THE ION ACTIVATION OF PLANT ENZYMES.*
BY GEZA DOBY AND R. P. HIBBARD.
(From the Department of Botany, Michigan State College, East Lansing, and the Agrochemical Department of The Economical Faculty, Royal
Hungarian University, Budapest, Hungary.)
(Received for publication, January 10, 1927.)
INTRODUCTION.
The role of salts and of their ions in the metabolism of the cell still remains one of the most important problems in the biological sciences.
It was believed that enzyme activity would be modified in plants when growing in solutions lacking certain salts or ions, and to test this idea out a number of experiments was undertaken with sugar beets as the test crop. At certain arbitrary periods during the growth of the plants, leaf material was gathered and the enzymes extracted. Certain tests were then made on these enzymes to determine their activity under various conditions.
Method.
1. Sand Cultures.
The plants used in the experiments were grown in pure quartz sand cultures in enamelled earthenware crocks of 2 gallon capacity. There were seventeen of these cultures, nine being reserved to be watered with a complete nutrient solution (containing all the essential salts in proper balance) and the other eight to be watered with a solution like the above with the exception of a deficiency of potassium. In this solution another salt, sodium, was substituted for the potassium. Care had to be exercised also
* Published by the permission of the Director of the Experiment Station of the Michigan State College.
405
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

406 Plant, Nutrient Ions and Enzymes
to adjust the solution to the same osmotic concentration as that of the complete solution. The construction of the cultures is shown in Fig. 1. The gravel layer at the top of the crock, as shown in the sketch acted as a mulch to check a too rapid evapora- tion of water and to restrain the growth of alga.
The sugar beet plants’ were grown in the field from pure line seed. When about 2 months old they were carefully transplanted to pots and brought into the greenhouse. Here they were left for a while to become accustomed to the new environmental conditions before being again transplanted into the culture crocks
l%G. 1. Details of quartz sand cultures. C, crock; T, glass tube for aeration and exchange of solution; G, quartz gravel; X, quartz sand; Ch, cloth.
described above. Transplanting was done carefully and the roots freed of soil particles by a jet of water. Three plants of uniform size were put in each crock.
The complete nutrient solution developed by Stoklasa and Matousek (11) and reported as supporting luxuriant growth was slightly modified for our purpose and also found to give excellent
1 We are indebted to E. E. Down, Plant Breeder, for these excellent plants.
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

G. Doby and R. P. Hibbard
growth. A glance at Table I will show that the modification consisted in eliminating certain ions that occurred frequently and in using magnesium sulfate in the place of calcium sulfate.
The two sets of sand cultures thus prepared were saturated with their respective solutions, which in 10 minutes were partially withdrawn, by means of a suction pump, to a final water content of 16 per cent. The total weight was then recorded on the jar. At weekly intervals the solutions were renewed in the same way and then sucked out to original weight. By this means the water content from week to week throughout the growing season was approximately maintained. The cultures were set on a rotating table in the greenhouse. Additional light (five 200 watt nitrogen-filled Mazda lamps) was used during the winter months. The plants were carefully watched and kept free from insect and
TABLE I.
Salts and Volume Molecular Concentration for Three Types of Solutions Compared.
~__ ~~~~~- gm. Urn. !7m. cm. vn. Qrn. c/m. l7m.
Complete . 1.150,0.9060.246,0.100,0.010 Minus potassium.. . 10.9060.246iO. 100,O. 101 0.96 Stoklasa and Matousek .11.1509.500~ i0. 100,O. 101~0~200,0~100
fungous pests. The tests on these plants were made at two differ- ent periods during their growth, February 10 to 14 and April 7 to 13. On the first date, about 2$ months after being trans- planted, the plants were in good vigorous condition, those in the complete nutrient being from one-third to one-fourth larger than the plants in the potassium-deficient nutrient. At the beginning of the second period the plants in the potassium-deficient nutrient had gained on an average of 46 per cent on their previous meas- urements, while those in the complete nutrient had made a gain of 75 per cent. Since all other conditions for growth were the same, it is evident that potassium has much to do with the vigor and growth of plants. What then can be the influence of a potassium-deficient nutrient on enzyme activity? How would the enzymes extracted from the leaves of plants in these two types of nutrient differ in amount, character, and salt activation?
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Plant Nutrient Ions and Enzymes
2. Enzyme Preparations and Preliminary Tests.
The two enzymes for study were those chiefly responsible for carbohydrate changes, amylase and saccharase. Both of these were extracted from the leaves at the same time by the follow- ing method: 20 to 30 gm. of fresh leaf material were shredded up after the petioles had been thrown out. A definite weighed sample was then mixed with clean quartz sand and ground in a porcelain mortar until it had become a thin paste. The juice was then pressed out by hand through cheese-cloth, diluted with an equal quantity of distilled water, and centrifuged for 5 minutes at 2000 R.P.M. The clear liquid was pipetted off and used immediately. Toluene was added as a preservative where necessary. This method of making the enzyme preparation is preferable to that in which the leaf material is first dried and then extracted. The enzyme appears to be obtained in unmodified form by this method.
The presence of amylase in the sugar beet was demonstrated long ago by Gonnermann (8) and later by Brasse (2). No quanti- tative experiments were reported. Later Palladin and Popoff (9) isolated amylase from sugar beet leaves and the leaves of many other kinds of plants. These men reported in addition more amylase in young leaves than in older ones.
Saccharase was shown by Gonnermann (8) to be present in the sugar beet plant. Later Stoklasa, Jelinek and Vftex (10) found this enzyme in roots undergoing anaerobic respiration. Cohn (4) made a study of the roots and leaves separately and found it always in the leaf, but absent in the roots that were growing normally. At the same time Bodnar (1) showed that it was absent in healthy roots but present in roots suffering from a disease, commonly known as “root-rot” (Schwanzfaule)..
Several preliminary experiments, which will be briefly noted, had to be conducted in order to become acquainted with the activity of the enzyme preparation and the various experimental conditions. For these studies good vigorous plants were selected from the general supply brought into the greenhouse from the field and potted in their own soil. The enzyme preparations were so active that considerable dilution (10: 1000) was necessary at the start. No buffers were used as it was thought that natural
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

G. Doby and R. P. Hibbard 409
ones in sufficient quantities were present in plant juices. Further- more, under the conditions arbitrarily set, it was necessary to study the influence of alkali salts without the disturbance of other buffers. The salts used in the study of enzyme activation were potassium nitrate, sodium nitrate, and potassium chloride. Studies were also made to determine the proper salt concentra- tions and to find out which salt and which ion exerted the greatest influence.
In testing for amylase and saccharase in young and older leaves it was found that young leaves showed more amylase than older ones. It did not appear to be a matter of the size, but just a matter of age. In regard to saccharase activity the young leaves contained more enzyme than the older ones. Great care then had to be exercised in the collection of material for comparative tests., All leaf collections were made at noon.
The method of procedure for determining the diastatic power or the sucroclastic activity of the leaf juice was that used by the senior author in his studies on the enzymes of the potato leaf (5, 6). For the amylase studies the method consisted in incubat- ing a 5 cc. sample for 24 hours at 38”C., and then determining by Wohlgemuth’s (13) calorimetric iodine method the diastatic power. In the tables this is represented as Di$, (diastatic power at 38°C. for 24 hours). The 5 cc. sample contains a definite amount of a 1 per cent starch solution, a definite concentration of enzymatic juice, and 0.1 cc. of toluene as a preservative.
The details of the method as used in the studies on saccharase are as follows: 2 cc. of enzymatic preparation are put into a total of 50 cc. of a solution which is made up of saccharase at 5 per cent, toluene (0.1 cc.) as a preservative, and the necessary amount of activating salt to reach the concentration called for in the test. This solution is then put into the incubator at 38°C. At certain time intervals, previously determined upon, 5 cc. quantities are withdrawn and treated with 1.5 cc. of lead sub- acetate (saturated solution diluted 2:5). After shaking, 0.5 cc. of a 12.8 per cent NazSOI solution is introduced and the solution shaken again. It is now centrifuged and 4 cc. of the clear liquid are mixed with 1 cc. of a 5 per cent solution of Na2C03 (made from crystals). This is again centrifuged and the clear solution is polarized in a 20 cm. tube at 20°C. The constant (Ic) of the
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Plant Nutrient Ions and Enzymes
reaction is calculated according to von Euler and Svanberg (7), R-FL
k = f + log d+L where t is time in hours from the beginning
of the reaction, R the initial rotation, L the final rotation, and d the observed rotation at t time.
TABLE II.
Amylase Activity in Plants under Two Types of Nutrient Conditions.
Type of nutrition. I
D38” 24h D Apr.
DFeb. Feb. 10. Apr. 8.
Complete.. . . . . . . . . . . 1s 5 0.28 Minus potnssium.. . . 33 25 0.75
TABLE III.
Sa.lt Activation of Amylase Extracted from Leaves of Plants Growing in Cultures Containing a Complete Nutrient as Compared with Similar
Samples of Amylase from Leaves of Plants Growing in C&w-es Otherwise Complete with the Exception of Potassium.
Concentration of salt.
Check ............................ NKNO~. ...........................
“ NaNOz ...........................
“ KCI. ...........................
Check ............................. 0.01 NKNOI .......................
0.01 “N&NO 3 ...................... 0.01 “KC1 .........................
D
1st period, Feb. 10, 1925.
Complete. Minus potassium.
per cent per cent 100 100 139 300 139 300 278 600
100 100 129 200 11s 200 294 400
2nd period. Apr. 8, 1925.
100 100 294 588 147 588 789 1176
100 100 100 400 loo 400 100 1600
3. Expetiments on Enzyme Preparations Obtained from Plants Growing under Two Sets of Nutrient Conditions.
With these preliminary experiments out of the way, it was next in order to extract the juice from the leaves of plants growing under conditions of complete nutrition and al&o under conditions of a nutrient lacking potassium, and make certain comparative
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from
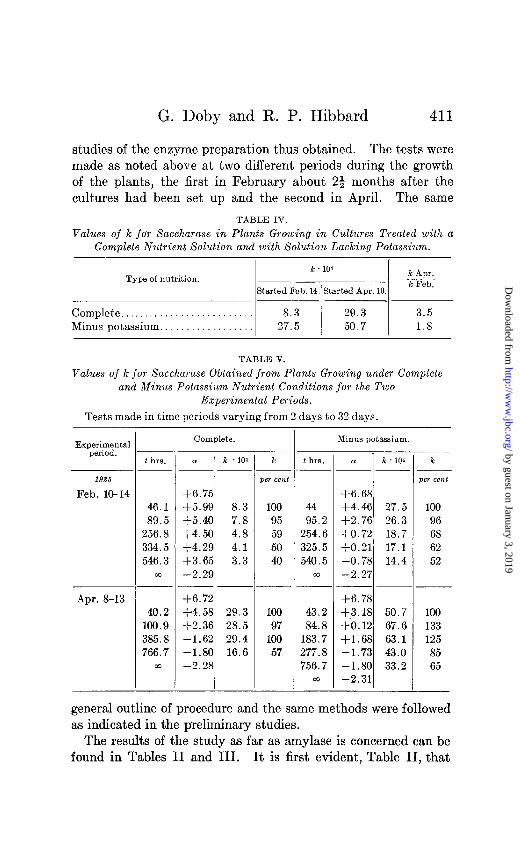
G. Doby and R. P. Hibbard 411
studies of the enzyme preparation thus obtained. The tests were made as noted above at two different periods during the growth of the plants, the first in February about 23 months after the cultures had been set up and the second in April. The same
TABLE IV.
Values of k jar Saccharase in Plants Growing in Cultures Treated with a Complete Nutrient $olution and with Solution Lacking Potassium.
Type of nutrition. k . 104
k Apr.
Started Feb. ld.lStarted Apr. 10. k Feb.
Complete. Minus potassium.
TABLE V.
Values of k for Saccharase Obtained from Plants Growing under Complete and Minus Potassium Nutrient Conditio,ns for the T.UO
Experimental Periods.
Tests made in time periods varying from 2 days to 32 days.
19.%%
Feb. 10-14
Apr. S-13
t hrs.
46.1 89.5
256.8 334.5 546.3
00
40.2 100.9 385.8 766.7
m
Compl&.
o( k 104
+6.75 +5.99 8.3 +5.40 7.8 $4.50 4.8 f4.29 4.1 +3.65 3.3 -2.29 --
+6.72 f4.58 29.3 +2.36 28.5 -1.62 29.4 -1.80 16.6 -2.28
k
wr cent
100 95 59 50 40
loo
97
100 57
Minus potassium.
t hrs.
44 95.2
254.6 325.5 540.5
m
43.2 84.8
183.7 277.8 756.7
co
+6.68 +4.46 27.5 +2.76 26.3 $0.72 18.7 $0.21 17.1 -0.78 14.4 -2.27
+6.78 +%18 50.7 f0.12 67.6 +‘1.68 63.1 -1.73 43.0 -1.80 33.2 -2.31
T k
per cent
100 96 68 62 52
loo
133 125 85 65
general outline of procedure and the same methods were followed as indicated in the peliminary studies.
The results of the study as far as amylase is concerned can be found in Tables II and III. It is first evident, Table II, that
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

$12 Plant Nutrient Ions and Enzymes
amylase from leaves of plants growing in a complete nutrient solution is less in amount than amylase extracted from leaves of plants growing in a nutrient lacking potassium. This is true for both growth periods. In the earlier period the difference was approximately twice as much, while in the second period it was 5 times as much. Again it is evident that enzymatic activity ,decreases with age, the decrease being more marked in plants growing under complete nutrient conditions. In Table III the D (diastatic power) values in percentage are recorded and show that activation occurs with all salts used and to a much greater extent
check . . * * *cd .
I 0 100 200 300 400 500
t (hours)
___ Complete. ------ Minus K. FIG. 2. First experimental period, absolute values of K.
in the material derived from plants growing in a potassium-de- ficient nutrient. The figures are more striking in the second period when the concentration of salts is 0.01 N.
The results of the study on saccharase are to be found in Tables IV and V. In the first of these two tables the k values for the enzyme are lower when obtained from plants growing under complete nutrient conditions than when taken from plants grow- ing in potassium-deficient cultures. The difference is 2 or 3 times as much. It is also clear that the k values increased in the second period. With material from plants in the complete nu-
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

G. Doby and R. P. Hibbard 413
Gent the increase was nearly 4 times, while in the other case it was nearly twice that of the first period.
In Table V a further difference is noted and that is in the be- havior of k, or, for example, the course of the reaction. In the first period (February) k (percentage) decreased in both types of nutrition, while in the second period it was constant for 16 days
60 -
50 -
40.
5 r;:
30 -
20 -
10 -
,
0 100 200 400 600
t (hours) ~ Complete. ------ Minus K.
FIG. 3. Second experimental period, absolute values of K.
for the complete nutrient and increased to the 4th day in the minus potassium nutrient. This is shown more clearly in Figs. 2 and 3 when the curves marked “check” are observed. In these graphs the absolute values of k are plotted, however.
The influence of the different salts on the absolute values of k for the different types of nutrient conditions is also shown in
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

414 Plant Nutrient Ions and Enzymes
the above graphs, the use of which eliminates the necessity of publishing the many tables prepared. Nitrates, it is observed, with but one exception were activating while the chloride was inhibiting. The exception was for the salt NaNOX and for the material obtained from plants in the potassium-deficient nutrient and for the second period. In Fig. 3, therefore, the curve for NaN03 is left out. It was also demonstrated in these experi- ments that there was a difference, through the use of salts, in the degree of enzyme activity between the two periods. There was twice as much in the earlier period as in the second.
Finally, as can be learned from Figs. 2 and 3, the k curve rises for falls much more slowly when the enzyme preparation is obtained from plants in complete nutrient condition than in the enzyme preparation from plants in potassium-deficient cultures. This is true in both growth periods, but is especially marked in the second period.
In the first period, nitrates showed considerable activating power when the enzyme preparation was obtained from plants in the potassium-deficient nutrient. In other words the curves rise to a high maximum. In the second period the same was true, but to a greater degree.
DISCUSSION OF RESULTS.
These experiments with amylase and saccharase go to show that ions may have a specific action and also an antagonistic one.
The type of nutrition and the physiological balance in a nutrient solution markedly influence the activation of enzymes by salts. Amylase preparations, for example, from leaves of plants in a complete nutrient, are not nearly as strongly activated as are the amylase preparations from leaves of plants growing in a potassium- deficient nutrient. In the latter case an unbalanced nutrient has induced an unbalanced salt or ion condition or perhaps a deficiency of a particular salt or ion in the plant and so affects the enzyme. When certain even weak concentrations, then, of these deficient salts or ions are used as activating agents the enzyme shows great activity. In the above experiments, since potassium was lacking, the plant was unable to obtain the chloride ion in sufficient quantity so that potassium chloride exerted a great influence as an activating agent.
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

G. Doby and R. P. Hibbard 415
These experiments show another thing of great importance. These enzymes occur in greater quantities in material from plants in potassium-deficient nutrient than in material from plants in complete nutrient. Brown and Morris (3) report the same result with leaves kept in darkness when compared with those in light. They conclude that this is a symptom of starva- tion. Our experiments show, however, (Table II) that with amylase the decrease in the amount as the plants age is less rapid in the potassium-deficient nutrient than in the complete one. In the case of saccharase (Table IV) the increase in amount as the plants age is also less rapid in the potassium-deficient nu- trient . Sjoberg (12) noticed the decrease in the amount of amylase with age in certain tree leaves, but concluded that it was not due to external conditions. The experiments reported here show at least for sugar beets that external conditions are important.
SUMMARY.
This study of the role of electrolytes in regulating amylase and saccharase activity in the sugar beet under different nutrient conditions leads to the following conclusions.
1. Young leaves contain more of the enzyme than the older ones.
2. The quantity of the enzyme is controlled by the type of nutrition, being greater in plants growing in a potassium-deficient solution than in a complete nutrient solution.
3. Amylase is strongly activated by the chloride ion and weakly activated by the potassium, nitrate, and fluoride ions. Saccharase is activated by the nitrate ion and somewhat by the potassium ion, but paralyzed by the chloride ion. These two enzymes in turn seem to be differently activated depending on the type of nutrition.
4. These actions are characteristically modified by the condi- tion of potassium deficiency. Contrary to expectation the strongest activation is not with potassium. Potassium only slightly activates the enzymes which are merely activated by specific ions. This phenomenon is explained by supposing a differential absorption of nutrient ions from the solution and a specific action of the ions on the different enzymes.
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

416 Plant Nutrient Ions and Enzymes
5. In all enzymatic studies, the type or kind of nutrition is as important as a determination of pH values, temperature, etc.
BIBLIOGRAPHY.
1. BodnBr, J., Biochemische Untersuchung der Rtibenschwanzfaule der Zuckerriibe, Biochem. Z., 1915, lxix, 245-256.
2. Brasse, L., L’amylase dans les feuilles, Compt. rend. Acad., 1884, xcix, 878-879.
3. Brown, H. T., and Morris, G. H., A contribution to the chemistry and physiology of foliage leaves. I. The starches and diastase of leaves, J. Chem. Sot., 1893, lxiii, 604677.
4. Cohn, H., Sur la distribution de I’invertine dans les tissues de la bet- terave aux differentes epoques de la vegetation, Compt. rend. Acad., 1915, clx, 177-179.
5. Doby, G., Biochemische Untersuchungen tiber die Blattrollkrankheiten der Kartoffel. I. Die Oxydase der ruhenden Knollen, 2. PJlanzenkr., 1911, xxi, 10-17; II. Die Oxydase der ruhenden und angetriebenen Knollen, 1911, xxi, 321-336; III. Chemische Beschaffenheit kranker und gesunder Pflanzenteile, 1912, xxii, 204211.
6. Doby, G., Uber Pflanzenenzyme. II. Die Amylase der Kartoffel- knolle, Biochem. Z., 1914, lxvii, 166-181.
7. von Euler, H., and Svanberg, O., Uber die Charakterisierung von Amy- laselosungen. (Vorliiufige Mitteilung), Z. physiol. Chem., 1921, cxii, 193-230.
8. Gonnermann, M., Die Entstehung des Zuckers in der Riibe, 2. Ver. deutsch. Zucker-Ind., 1898, xlviii, 667-689.
9. Palladin, W., and Popoff, H., Uber die Entstehung der Amylase und Maltase in den Pflanzen, Biochem. Z., 1922, cxxviii, 487-494.
10. Stoklasa, J., Jelinek, J., and Vitex, E., Der anaerobe Stoffwechsel der hoheren Pflanzen und seine Beziehung zur alkoholischen Glrung, Beitr. them. Physiol. u. Path., 1903, iii, 460-509.
11. Stoklasa, J., and Matousek, A., Beitrage zur Kenntnis der Ernlhrung der Zuckerrtibe, Jena, 1916.
12. Sjoberg, K., Beitrage BUS Kenntnis der Amylase in Pflanzen. I. Uber die Bildung und das Verhalten der Amylase in lebenden Pflanzen, Biochem. Z., 1922, cxxxiii, 218-293.
13. Wohlgemuth, J., Uber eine neue Methode zur quantitativen Bestim- mung des diastatischen Eerments, Biochem. Z., 1908, ix, l-9.
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from

Geza Doby and R. P. HibbardENZYMES
ION ACTIVATION OF PLANT NUTRIENT IONS OF PLANTS AND THE
1927, 73:405-416.J. Biol. Chem.
http://www.jbc.org/content/73/2/405.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
ml#ref-list-1
http://www.jbc.org/content/73/2/405.citation.full.htaccessed free atThis article cites 0 references, 0 of which can be
by guest on January 3, 2019http://w
ww
.jbc.org/D
ownloaded from