observations on dicystid gregarines from marine...
TRANSCRIPT

Observations on Dicystid Gregarines fromMarine Worms.
By
D. L. Mackinnon, D.Sc,and
H. N. Ray, Ph.D.From the Zoology Department, King's College, London.
With Plates 20 and 21.
CONTENTS.PAGE
I N T R O D U C T I O N . . . . . . . . . . 439
M A T E R I A L A N D M E T H O D S . . . . . . . . 4 4 3
D I C Y S T I D G R E G A R I N E S F R O M T W O P O L Y C H A E T E W O R M S . . 4 4 4
A . P o l y r h a b d i n a s p i o n i s Ming , n . v a r . b i f u r c a t e , f r om
S c o l e l e p i s f u l i g i n o s a . . . . . . 444
B . P o l y r h a b d i n a p o l y d o r a e Caul l . a n d Mesn . ( ? ) f rom
P o l y d o r a f l a v a . . . . . . . 447
D I C Y S T I D G R E G A R I N E S F R O M T H A L A S S E M A N E P T U U I . . 450
A . H e n t s c h e l i a t h a l a a s e m a e n .gen . , n . s p . . . 4 5 1
B . L e c y t h i o n t h a l a s s e m a e n .gen . , n . s p . . . . . 4 5 3
S P O R E F O R M A T I O N I N G R E G A R I N E S F R O M T H E G U T O F T H A L A S S E M A 4 5 6
S Y S T E M A T I C P O S I T I O N O F T H E D I C Y S T I D G R E G A R I N E S . . . 4 6 0
S U M M A R Y . . . . . . . . . . 4 6 1
R E F E R E N C E S . . . . . . . . . . 4 6 3
E X P L A N A T I O N O F P L A T E S . . . . . . . . 4 6 4
I N T R O D U C T I O N .
THE ' dicystid' gregarines are a little-known group of Sporozoa,and since modern technical methods have been introduced intoprotozoology they have scarcely been studied at all. Webecame interested in some that we found abundantly in certainmarine worms at Plymouth; but our preliminary inquiry intothe work of earher observers revealed so many discrepancies,

440 D. L. MACKINNON AND H. N. BAY
and the confusion in the nomenclature appeared to be so greatthat we think it not amiss to prelude what we ourselves have tosay with a brief summary of relevant statements that appearin the papers we have consulted.
The following definition was given by Leger in 1892. 'Lesveritables dicystidees comprendront seulement deux segments:l'un intracellulaire, l'appareil de fixation: l'autre resultant duprolongement pousse en dehors de la cellule et dans lequel iln'apparait pas de septum dans la suite: c'est l'equivalent duprotomerite et du deutomerite reunis: il renferme le noyau.'
Leger took as his type a gregarine that he found in the gutof the aquatic larvae of certain dipterous insects; to this he gavethe name S c h n e i d e r i a . The somewhat elaborate epimeriteof S c h n e i d e r i a is shed when the organism is full-grown; themonocystid-like bodies then associate in pairs by their anteriorends, and in the resulting gametocysts spores are formed beforeevacuation from the host.
A year previous to the appearance of Leger's paper, Min-gazzini (1891 a) had published a note on a gregarine which hecalled U l i v i n a e l l i p t i c a , from the intestine of the poly-chaete worm C i r r a t u l u s f i l i g e r u s . He did not observeits epimerite and his drawings show the main part of the bodyonly, in which there is no distinct septum between the anteriorand posterior portions, although the endoplasm in the former isof different quality and is somewhat sharply delimited from thatwhich lies behind it. U l i v i n a e l l i p t i c a is closely relatedto, if not actually identical with, Syc i a o p i n a t a , which inhis paper of 1892 Leger also records from the gut of A u d o u i n i a( C i r r a t u l u s ) l a m a r k i i . But Leger evidently regardedS y c i a as a tricystid. He described its development prettyfully, with a young intracellular stage, and found that game-tocysts are formed within the worm's body, although he couldnot trace the formation of their contents beyond the sporoblast
1 We have frequently found the trophozoites of Syoia (Ulivina?)in A u d o u i n i a (Ci r ra tu lus ) t e n t a c u l a t u s (Mont.) at Plymouth,and we consider that it is as definitely 'dicystid' as the gregarines fromother marine worms that we shall describe in this paper.

DICYSTID GBEGARINES 441
In 1893 Leger noted two 'true dicystids' from the polychaeteworms Nereis cultr ifera and Polydora agassizi i .For these he created the genus Doliocystis , of Avhich heexpressly states that a characteristic feature is the simpleand rudimentary character of the epimerite; in Doliocystisnereidis it is 'un simple bouton', and in Doliocystispolydorae 'un tronc de cone a petite base inferieure' which' se continue directement avec l'extremite anterieure du secondsegment, allonge en forme de col'.
In 1891 Mingazzini published other papers on gregarinesbesides that in which he described U1 i v i n a; and in one ofthese (18916) he created the genus Polyrabdina , of whichhis account runs as follows: 'Specie dimorfe. Individui e formacli nematode piriforme. I primi hanno il corpo allungato fusi-forme, e la cuticula striata longitudinalmente da rialzi numerosifinissimi.' The organisms so described were found in the intes-tine of the polychaete Spio fuliginosus, and Mingazziniconsidered that in its 'first form' his Polyrabdina wasprobably identical with Gregarina spionis recorded fromthat worm by Kolliker in 1845 and 1849. He slightly elaboratedhis description in a later paper (1893), gave some very feebledrawings, and again remarked that the second form 'whichKolliker did not see has the shape of a very small nematode'.
In 1897 Mesnil and Caullery recorded from Spio mart i -nensis a gregarine which they then regarded as Gregarinaspionis Kolliker; and in 1901 the same authors referred to agregarine with intracellular epimerite living in Scolelepis(Spio) fuliginosa and 'appartenant au genre Dolio-cystis Leger'. Liihe (1904), on Mesnil and Caullery's authority,also referred to this gregarine from Scolelepis as a speciesof Doliocystis .
Mesnil (1907) returned to the question, and remarked, 'IIexiste, je crois, chez toutes les Annelides de la famille desSpionidiens, des Gregarines aplaties, a contour ovoiide, qui sontdes vraies Dicystidees. J'ai etudie en particulier une especeparasite de Scolelepis fuliginosa, a epimerite caduc,qui envoie dans la cellule epitheliale une couronne de prolonge-ments, parfois ramifiees, sans doute amibo'ides.'

442 D. L. MACKINNON AND H. N. BAY
Brasil (1908 and 1909) made a further study of Leger's type-species of D o l i o c y s t i s and of others that he had observed.In his view, these gregarines show affinities wifchLankesteriaand so lead over to the coelomic 'monocystids'. He decidedthat Op h io d ina Mingazz in i is synonymous with Dol io-c y s t i s , but that Leger's name has the priority.1 The chiefcontribution by Brasil to our knowledge of the life-history ofD o l i o c y s t i s is that, while the young parasite is intra-cellular, it later on drops out completely into the gut lumen andfixes itself to the surface of the epithelium by the epimerite,which functions as a sort of sucker. Referring to Mesnil'sgregarine from Sco le lep i s , Brasil comments, 'Sont-ce la deaD o l i o c y s t i s ? Port probablement'.
By 1914, however, Caullery and Mesnil had apparentlydecided that the gregarine from Scole lepis was not aD o l i o c y s t i s . They pointed out that Mingazzini had un-doubtedly included under his P o l y r a b d i n a (or P o l y r h a b -d ina , as Caullery and Mesnil prefer to spell the name) twodistinct organisms. Mingazzini's 'nematoid form' is a speciesof Se l en id ium, a genus founded by Giard in 1884; so thatMingazzini's 'pear-shaped form' alone should bear the name ofP o l y r h a b d i n a . Caullery and Mesnil proceeded to give someaccount of this dicystid from the gut of Sco le lep i s , and theycalled it P o l y r h a b d i n a sp ionis (Kolliker), though it isobvious, when one reads Kolliker's papers, that the organismhe observed was a 'nematoid form' and a species of Seleni-d ium. 2 (Elsewhere, one of us (H. N. R.) has given somefurther description of this Se l en id ium sp ionis (Kolliker).)Caullery and Mesnil further noted the existence of three otherspecies of P o l y r h a b d i n a — P o l y r h a b d i n a p o l y d o r a e(Leger) from P o l y d o r a c i l i a t a , P o l y r h a b d i n a b r a s i l iCaullery and Mesnil from Spio m a r t i n e n s i s , and Po ly -
1 Kamm (1922) considers that both O p h i o i d i n a and D o l i o c y s t i sare synonymous with L e c u d i n a M i n g a z z i n i , a genus founded twoyears earlier—1891. Bhatia (1930) agrees with Kamm.
2 ' Die Bewegungen dieses Schmarotzers sind longsam und ganz deutlich;sie kommen wie bei G. s i p u n c u l i durch sichtbare Contraktionen undExpansionen der Leibeshohle zu Stande' (Kolliker, 1849).

DICYSTID GREGARINES 448
rhabdina pygospionis C. and M. from Pygospioniaset icornis .
While studying the Protozoa in the gut of Scolelepisfuliginosa and allied polychaetes at Plymouth, we havefrequently come across dicystid gregarines; and since no first-hand observations on these organisms have been published sinceCaullery and Mesnil's brief description of Polyrhabdinain 1914, we think it worth while to place on record such factsas we have noted, for, as may be gathered from what has beensaid above, trustworthy accounts and illustrations are few, andwe are persuaded that some of the earlier observations wereincorrect. We have also found two dicystid gregarines in thegut of the echiurid worm, Thalassema neptuni Gartner,which seem deserving of some description.
MATERIAL AND METHODS.
All the worms we examined came from Plymouth. Westudied them there and also in the laboratory in London, wherewe were able to keep them alive and healthy for some weeksat a time. We found that only those specimens of Thalas-sema obtained by dredging were well infected; a dozen or socollected on the shore between tide-marks contained almost no
It is not easy to dissect out entire the gut of a polychaetesuch as Scolelepis; nor, when this is done, can one seethrough its relatively thick wall what parasites it contains.So we examined samples of the gut-contents, and then, if itseemed to be infected, we made smears and cut sections. Withthe echiurid it was possible to recognize the parasites easilyenough through the gut wall, so our practice here was to takeout the gut entire, stretch it on a slide, and examine it underthe slight pressure of a cover-slip.
We attach much importance to studying the parasites alive.There was usually enough gut-fluid obtainable for this purpose;when this had to be diluted, we found that the gregarines stoodthe addition of sea-water better than that of ordinary physio-logical salt-solution.
Smears and pieces of infected gut were fixed in Brasil'sNO. 295 G g

444 D. L. MACKINNON AND H. N. BAY
modification of Bouin-Duboscq's fluid1 and also in Schaudinn'sfluid. Helly's fluid was occasionally used. The stains employedwere Delafield's haematoxylin, Heidenhain's iron haematoxylinand Mann's methyl blue eosin. A double staining with Delafieldand borax-carmine was sometimes employed with effect.Material fixed in Helly's fluid was stained in Altmann's acidfuchsin and toluidin blue. Various counterstains were employedfor the haematoxylin preparations; of these, saturated solutionof chromotrop 2E (1 per cent, in absolute alcohol) was foundthe most useful. Sections were usually cut 8-10/x thick.
DlCYSTID GRBGARINES FROM TWO PoLYCHAETE WORMS.
A. P o l y r h a b d i n a sp ion i s Ming, n.var. b i f u r c a t a fromScole lep is fu l ig inosa Clpdc.
Eighty-five of the two hundred and fifty specimens ofSco le lep i s fu l ig inosa examined by us contained a cepha-line gregarine that agrees in many respects with CauUery andMesnil's description of P o l y r h a b d i n a s p i o n i s .
The organism (fig. 1, PI. 20) lives in the middle portion ofthe gut, and is almost always attached to the wall by itsepimerite. This is a very definite, transparent, knob-shapedstructure, with a circlet of fourteen to sixteen minute teethat its base, and, as its crown, two much larger, diverging, claw-like processes, which are flattened towards their tips (fig. 4,PI. 20). CauUery and Mesnil figure these processes, or 'bar-belures', as at least four in number, sometimes branched; andin one drawing, where the epimerite is seen in end-on view, asmany as nine are shown. Moreover, according to these authors,the whole epimerite is intracellular and the processes are saidto be amoeboid structures which draw nourishment from thehost-cell. We have examined many sections, but have neverfound the epimeritic claws lying within the substance of thecell; they are merely applied to the surface of the epithelium,
1 Sat. solution of picric acid in 90 per cent, alcohol . . 2 partsSat. solution of corrosive sublimate in distilled water . 3 „Formalin . . . . . . . . . 1 ,,Glacial acetic acid . . . . . . . 2 „

DICYSTID GREGARINES 445
sometimes embracing more than one cell. There is no evidenceat all that they are amoeboid or capable of absorbing food; wenever find that they are perforated at their tips, and theyseem indeed to be cuticular anchors, comparable with theepimeritic outgrowths of gregarines like N i n a ( P t e r o -c e p h a l u s ) . The whole surface of the epimerite stains blackwith iron haematoxylin and pink with Mann's methyl blueeosin.
We have found it difficult to decide whether the differencesin the structure of the dicystid gregarines observed by us andby Caullery and Mesnil from the same worm (though admittedlyfrom different localities) are such as to justify the creation ofa new species. On the whole, we think it best to regard thePlymouth type as a new variety, b i f u r c a t a , of P o l y r h a b -d i n a s p i o n i s Mingazzini.
Behind the epimerite the main mass of the body of a full-grown gregarine is a sac-like thing measuring about 180/x x 36ft;it is always circular in cross-section (fig. 3, PI. 20)—not flattened,as Mesnil seems to suggest that these dicystid gregarines ofpolychaetes are wont to be. There is a thick pellicle, which islongitudinally grooved. The pellicle is very much thinner atthe bottom of the grooves, and it seems probable that fluidnourishment from the worm's gut can be absorbed along theselines. The ridges between the grooves stain deeply with ironhaematoxylin, and each contains three to five siderophilousfibres longitudinally disposed. (Caullery and Mesnil make noreference to these characteristic striations.) The ridges convergetowards the anterior and posterior ends of the deutomerite andmay actually become concrescent. The gregarine appears quiteimmobile when alive, and there is no evidence that the fibrilson the ridges are myonemes. We do find, however, that a layerof circularly disposed fibrils lies beneath the longitudinalgrooves; and the variation in width of the grooves and theseeming breadth of the dark, fibrillar ridges between them maybe affected by the contraction of these elements.
The endoplasm in the main portion of the body appears darkand granular when alive, but clear and alveolar in fixed speci-mens, except where parasites are present. As in Syc ia
Gg2

446 D. L. MACKINNON AND H. N. RAY
(Ulivina) the endoplasm is more transparent just behind theepimerite, and there is sometimes a fairly definite line ofdemarcation between this clearer region and the more granularendoplasm behind, though there is no actual septum. Thenucleus measures 10-5/x x7-5/x in a full-grown specimen; it liesin the middle of the body, midway between the poles of themain segment (fig. 1, PL 20); in parasitized gregarines it getscrushed to one side and stains more faintly than normal. Thenuclear sap is filled with fine, evenly disposed granules ofchromatin; usually two karyosomes are present; one is fairlylarge and stains less intensely than the other, which is alwaysa small, densely siderophilous granule. Both stain red withMann's stain.
Caullery and Mesnil maintain that, in the early part of itslife, P o l y r h a b d i n a s p i o n i s lives mainly within the host-cell. This is what one would expect to find. They give drawingsto show the entry of the anterior end of the young, undifferen-tiated organism into the cell, and its growth there. The youngeststages we have found in S c o l e l e p i s from Plymouth measureabout 7-5 ix x3/u. These are clinging to the surface of the cellsby means of the small teeth at the base of the epimerite; thecharacteristic anchors must be developed later. In otherrespects these young gregarines are just like the adults, but ona small scale. We have no evidence that the organism is evereven partially intracellular. Caullery and Mesnil's pictures ofthat stage suggest that they may have confused with P o l y -r h a b d i n a a young intracellular S e l e n i d i u m , two speciesof which are its common accompaniments in S c o l e l e p i s .One of us has described these ' schizogregarines' elsewhere(Ray, 1930 a). We are the more inclined to think that Caulleryand Mesnil were mistaken, since they undoubtedly confusedS e l e n i d i u m with intracellular stages of a coccidian-likesporozoan that is also common in the same situation (Ray,19306).
Our impression then is that the intracellular stage of P o l y -r h a b d i n a s p i o n i s , if it occurs at all, must be very brief,and that during almost the whole of its life this gregarine isextracellular, clinging to the epithelial surface by means of its

DICYSTID GREGARINES 447
epimeritic anchor. The cells to which it hangs rarely showhypertrophy.
We are unable to say anything further concerning the life-history. Where the infection is heavy, these relatively largeorganisms lie so close to one another in the narrow lumen ofthe host's gut that association might easily take place withoutany relaxation of the grip upon the epithelial cells; but we havenever seen such a thing happen, though we have searched longand carefully. Nor have we ever found spores, either withinthe worms or in the slime mixed with faecal debris that collectsround these in the aquarium.
We have often seen the curious parasites of Polyrhabdinathat Caullery and Mesnil (1897 and 1914) named Metchniko-v e 11 a; but we are as little able as were these authors to decidewhat is the true nature of these organisms. We have nothingto add to the description given by Caullery and Mesnil ofMetchnikovella, and we suppose that the species we findis Metchnikovella spionis Caullery and Mesnil, althoughtheirs was from Polyrhabdina brasi l i in Spio Marti-n en sis and the cysts in our specimens sometimes exceed thedimensions they give. On PI. 20 are three drawings (figs. 5, 6,and 7) of this parasite as we have seen it in the Polyrhab-dina from Scolelepis fuliginosa.
B. Polyrhabdina polydorae Caul, and Mesn.(?) fromPolydora flava Clpde. (= Lecudina polydoraeKamm 1922).
As already stated (p. 441), Leger (1893) described a dicystidgregarine from the intestine of the polychaete Polydoraagassizii and he named this Doliocystis polydorae .It was said to have a very simple epimerite,' un tronc de cone apetite base inferieure'. Brasil (1909) when working on other speciesof Doliocystis , referred to Doliocystis polydoraeLeger, said that he had not actually seen it, and then went onto observe that the genus Doliocystis seems to have much incommon with Lankesteria. It is rather surprising, therefore,that Brasil should consider as 'probably a Doliocyst is ' thedicystid with ramified epimerite recorded by Mesnil (1907)

448 D. L. MACKINNON AND H. N. RAY
from Scolelepis fuliginosa—a near relative, that is tosay, of the gregarine which we have just described above.
Caullery and Mesnil merely added to the confusion. Theyremarked (1914a) that the species of Polyrhabdina (Poly-rhabdina brasili) which they find in Spio mart inensishas an epimerite of fundamentally the same type as that ofPolyrhabdina spionis, and they proceed, 'Ilenestsansdoute de meme de l'epimerite du Doliocystis polydoraedecrit par Leger; nous avons pu le verifier pour une Gregarinede Poly dor a cil iata. ' And in the list of species ofPolyrhabdina at the end of their paper they insert 'P .polydorae (Leger) de Polydora ciliata' .1 Caullery andMesnil seem to agree with Brasil that Doliocystis is relatedto the monocystid gregarines; and it is not clear how they squarethis with their assertion that Doliocystis polydoraeLe"ger is a species of Polyrhabdina . Kamm (1922) appar-ently follows Caullery and Mesnil in regarding the gregarine asa Doliocystis . She refers to it as Lecudine polydorae.
Leger's accounts of what he observed should always be takenseriously. It is unlikely that so careful an observer would haveoverlooked the somewhat complicated structure that one wouldsuppose, from Caullery and Mesnil's own statement, the epimeriteof a Polyrhabdina should show.
We have not seen Polydora c i l ia ta , nor its varietya g a s s i z i i, but we have frequently found in the intestine ofPolydora flava at Plymouth a dicystid gregarine un-doubtedly related to the Polyrhabdina from Scolelepis.This may be the species observed by Caullery and Mesnil, andwe have therefore given their names as the ' authority' for Poly-rhabdina polydorae, for there is no good reason to sup-pose that Leger ever saw this organism.
We say that the dicystid from Polydora flava may beCaullery and Mesnil's species, but these authors give no realdescription of what they saw. We think it worth while to pub-lish a few notes and drawings, although unfortunately we arenot able to give any account of the life-history. We examinedmany worms, and though 75 per cent, were infected, often very
1 Polydora agassizii is a variety of Polydora ciliata.

DICYSTID GBEGARINES 449
richly, we always found the gregarines well developed butapparently not yet ripe for association.
As we have found them, the trophozoites measure 100-180/*x 30-40/x (fig. 8, PI. 20); we have never seen any young forms.
Thepellicular layer is relatively thinner than in P o l y r h a b d i n as p i o n i s, but carries comparable, though slighter, longitudinalstriations (each made up of a number of dark-staining fibrils)with thinner-walled grooves between. The nucleus measures15/A across and lies in the middle of the sac-like body; theremay be more than one karyosome, and one of these frequentlyshows a lighter core. The chromatin is less evenly disposed inthe nuclear sap than in the last species; it often forms smallclumps of irregular shape.
The epimerite has no claws at its tip; but twenty-two totwenty-four slender, dark-staining prongs arise in a circleslightly anterior to a collar of similar material at the base(fig. 9, PI. 20), and are seemingly the counterpart of the smalldenticles in that position on the epimerite of P o l y d o r as p i o n i s . By means of its epimerite the gregarine clingsclosely to an epithelial cell; but, in the adult condition anyhow,it always remains quite external thereto. The delicate prongsfrom the collar arch over and hold on to the surface of the cell,and the knob-like portion in front of them bulges into a depres-sion in it. The affected cell tends to increase considerably inthickness, but does not elongate perceptibly nor project beyondits fellows. There is no evidence that such hypertrophy asoccurs is caused by other than purely mechanical interferenceby the parasite, for the prongs of the epimerite have no apertureat their tips, nor does the knob really enter the cell anywhere.As in P o l y d o r a s p i o n i s , the epimerite seems to persistthroughout the gregarine's growth period; but as we have seenno young forms, we have no evidence as to how it develops.
A species of M e t c h n i k o v e l l a attacks this gregarine also,and, as it is not like any of the seven described by Caullery andMesnil, we propose to call it M e t c h n i k o v e l l a c a u l l e r y in.sp. Portions of the hyphal stages are shown in figs. 11 and 12(PI. 20). The tube-like 'cysts' measure 85-100/1 x-2-5-3/*(fig. 13 a, PI. 20); they scarcely taper at all towards their ends,

450 D. L. MACKINNON AND H. N. RAY
and these contain very little of the dark-staining pluggingmaterial that is so obvious in M e t c h n i k o v e l l a s p i o n i s(fig. 13 b, PI. 20). The ' spores' measure about 3/a x -75 /u (fig. 13 c,PI. 20): there are 20 to 30 within each cyst. In some of theinfected gregarines in smears, we find that the epimerite hasbeen torn away, and the cysts of the M e t c h n i k o v e l l a areseen escaping, as they might from a bottle of which the stopperhad been removed. It seems improbable that this is theirnormal mode of emergence. Perhaps the host gregarines dis-integrate and deposit their contents in the gut; we havenever found completely free cysts, and we have often seen thespores escaping from the cysts into the impoverished endoplasmof a heavily infected gregarine.
DICYSTID GREGARINES FROM T H A L A S S E M A N E P T U N IGARTNER
The gregarines we have described above are from the gutof polychaete worms; and this seems to be the habitat of mostof the species known.
The following notes refer to two dicystids that we find veryfrequently in the intestine of an echiurid worm at Plymouth,T h a l a s s e m a n e p t u n i Gartner, where they occur in com-pany with the holotrichous ciliate P t y s s o s t o m a t h a l a s -s e m a e Hentschel, and a peculiar acephaline gregarine whichwe describe elsewhere.1
We are reluctant to give new generic names to these organisms,for it is obvious that, until much more is known about the life-cycle of the dicystid gregarines, it is futile to express an opinionas to their relationships or to decide which of their morphologicalcharacters is to be considered as of 'generic value'. With thetricystid gregarines, the structure of the spore seems to be thebest criterion of relationship; and, next to that, the form of theepimerite. For reasons that will appear later, we can say nothingdefinite about the spores of the gregarines from T h a l a s s e m a .
1 In 1881 Lankester described a 'gregarine' from T h a l a s s e m an e p t u n i , but this was found in the egg-sacs of a female, where it hadinvaded the developing eggs. We have seen this organism, and haverecently stated (1929) that it is a coccidian.

DICYSTID GREGARINES 451
Their epimerites are complicated structures which seem toanchor them to the gut-wall throughout their growth astrophozoites; this rules out our placing them in the genusDoliocystis . We have seen Anchorina sag i t t a t aLeuck. from the gut of the polychaete Capitella cap i t a t a ,a dicystid which, in spite of its peculiar form, has points ofsimilarity to Doliocystis; it has almost nothing incommon with the gregarines from Thalassema. We havecarefully examined Sycia (Ulivina?) from Audouinia,and we regard it as something quite distinct from our parasites.Their epimeritic structure is unlike that of Polyrhabdina ,described above. Among well-authenticated genera thereremains only Schneideria; but this dicystid is parasitic inaquatic insect larvae, and its spores develop while the gameto-cysts are still within the body of the host. Whereas, so far aswe can judge, the associated gametocytes of the gregarines fromThalassema do not develop further until they have reachedthe water outside.
We therefore feel obliged to create two new genera, H e n t -sche 1 ia and Lecythion, to include the dicystid gregarinesfrom the echiurid.
A. Hentschelia thalassemae n.gen., n.sp.The trophozoites are most abundant in the middle portion
of the gut, i.e. the part of the intestine that is accompanied bythe collateral tube; in the narrower, thick-walled region theymay be so abundant as almost to block the lumen. Associatingforms occur further back and are found in the rectum chiefly,whence they are evacuated with the faecal balls.
We have found a few young stages. These lie within theepithelial cells (fig. 14, PI. 20). The smallest we have notedmeasure about 15-22 ju x23/x. Already the inner end of theparasite is partially differentiated as the epimerite; the nucleusis relatively enormous. The host-cell is considerably enlargedand has paler contents than have its neighbours, and even atthis early stage it tends to get dragged up from the rest of theepithelium, to which it remains attached by rooting processesof cytoplasm. As in cephaline gregarines generally, it would

452 D. L. MACKINNON AND H. N. BAY
appear that, as the organism increases in length, its posteriorsegment breaks through into the gut and hangs free there,while continuing to grow to full size; the epimerite, however,always remains lodged within the host-cell, and so firmly that,when one makes a smear, the two usually come away together.
Including its epimerite, the full-grown trophozoite (fig. 15,PI. 20), measures 75-98/x in length; the greatest width of suchforms is 30-45/z. The posterior end is often pointed like thefree end of a lemon. But the deutomeritic segment varies verymuch in relative length and breadth from specimen to specimen;and this suggests that there may be a layer of circularly dis-posed myonemes below the pellicle though we have never observedany contraction, and the gregarines remain quite inert whendetached from their hold. The pellicle is very thin comparedwith that of P o l y r h a b d i n a j i t s surface bears fine, close-set,inconspicuous striations; just behind the epimerite, linesrunning transversely to these may sometimes be seen.
The nucleus lies about the centre of the body; its diameter is22-5 ̂ in a large specimen. It is crammed with minute chromatingranules, arranged in strings or scattered more irregularly;often the granules form three or four small clumps. There isusually one fairly large karyosome, which is often pale, butsometimes stains more darkly; in addition to this there may bea smaller karyosome.
The anterior end of the main segment narrows to a smallneck, whence arises the epimerite. The surface of this transparentstructure stains rather deeply with iron haematoxylin. As wehave said, it is always deeply embedded in a cell, a conditionof things very unlike what we have noted in the previousspecies. As the gametocytic stage approaches, the epimerite iseasily lost, and isolated epimerites lying within shrivelledepithelial cells may often be seen in sections of the gut. Theepimerite (fig. 16, PI. 20) measures about one-sixth of the totallength of the full-grown gregarine; it is a broadly spreadingumbrella-like structure, of which the margin is divided intofour or five lobes, each fluted on the anterior surface andelaborately frilled around its edge. The fluting of the lobesis continued on to the rounded apex of the umbrella as a series

DICYSTID GREGARINES 453
of converging striations. It is sometimes possible to see thata slight pellicular ridge stands up from the posterior segmentaround the base of the neck of the epimerite. Just anterior tothe epimerite in the fixed and stained host cell, there usuallylies a vacuole crossed by a few protoplasmic strands.
Individuals about to associate shorten and thicken. As wehave said, they generally lose their epimerites before associating,but occasionally they may associate precociously (fig. 22, PI. 21).Association is lateral, and the pair are usually disposed head-to-tail. Very commonly they are of different sizes. Whenattachment has been effected by circular areas on theapposed sides, the gametocytes become almost spherical, sothat their combined bodies have the outline of a figure 8(fig. 23, PI. 21). At this stage their pellicular striations fade,and it becomes impossible to distinguish the associated pairsof H e n t s c h e l i a from those of its fellow gregarines, exceptby the nuclear structure. In this condition they are passed outinto the sea-water with the faeces of the worm.
B. L e c y t h i o n t h a l a s s e m a e n.gen., n.sp.
Attached to epithelial cells in the same part, of the gut withH e n t s c h e l i a we find, though less frequently, anotherdicystid. This organism, when full-grown, has the form of anelongated soda-water bottle, of which the epimerite representsthe neck and cork1 (fig. 17a, PI. 20).
Adult trophozoites measure about 135^. x52/i; the epimeritecontributes about one-fifth of the length, or sometimes less.The posterior segment is often pointed posteriorly, so that itsoutline is that of a lemon. The epimerite (fig. 21, PI. 20) isa flexible, thin-walled structure; in half-grown specimens it islonger in proportion to the posterior segment than it is in theolder trophozoites; its greatest width is about 11 /*. After leavingthe main segment it swells out slightly, then narrows again,and finally expands at its tip into a slight cone, the cork of thebottle, around the base of which are some fourteen or fifteen
1 It has considerable resemblance to the tricystid gregarine Phialoid.esLeger.

454 D. L. MACKINNON AND H. N. BAY
petal-shaped lobes, which, tilted on edge, grip the epithelialcells and serve as an anchor for the gregarine. The surface ofthe epimerite is longitudinally ridged; these striations thickenand become much more obvious at the proximal end, wherethey curve down on to the deutomerite, following there the lineof a right-handed spiral. The ' cork' bears a number of minutedenticles; and similar, but larger, denticles project from theobliquely running lines on the pellicle of the deutomerite; thetips of these deutomeritic teeth are directed backwards (fig. 17b,PI. 20). The cytoplasm within the epimerite is hyaline, but inthe terminal cone region are a few minute granules, and a groupof larger, highly refringent granules, two to eight in number, liesnear its posterior end. These granules stain deeply with ironhaematoxylin.
When the gut is teased for the making of smears, the tro-phozoites tend to come away entire, leaving behind them thecells to which they have been hanging. This is in marked con-trast to what happens with H e n t s c h e l i a . The explanationis that the epimerite of L e c y t h i o n merely clamps its gre-garine to the cell-surface, as do the claws on the epimerite ofP o l y r h a b d i n a ; in the adult condition, anyhow, it is neverintracellular. The host-cell is scarcely, if at all, hypertrophied.The point of attachment of the epimerite is generally at thebottom of a fold in the intestinal wall, and the epimerite issupported on two sides at least by the surrounding epithelium,only the deutomerite hanging out quite freely into the lumen.When alive, and while still attached to the gut-wall, thetrophozoites show certain 'metabolic' movements which oddlyresemble those of T h a l a s s e m a itself. A constriction appearsbehind the epimerite (as it does just behind the proboscis ofthe echiurid) and gradually travels backwards to the posteriorend; as it fades away, another wave of contraction begins atthe anterior end; the direction of these peristaltic waves is neverreversed. When detached from its hold, the gregarine is alsocapable of movement from place to place. It glides slowlyalong, usually hind-end foremost. This movement is notaccompanied by any contractions of the body-wall that we candetect, and it is difficult to see how the backwardly directed

DICYSTID GREGARINES 455
denticles on the pellicle can assist progression by gripping theslimy substratum.
Though we have examined many sections of the gut ofinfected T h a l a s s e m a , w e have never found in these anyevidence that L e c y t h i o n has an intracellular stage. In onesmear, however, from a heavily infected worm, we saw what isshown in fig. 20 (PI. 20). Here in a piece of detached epitheliumtwo young gregarines are embedded, and they appear to belying in vacuoles within the tissue, though whether they arebetween cells or actually intracellular it is impossible to saywith certainty. The epimerite, still very short, seems to betelescoped into the main segment; within it the characteristicsiderophilous granules show clearly, and the lobes around the'cork' are just appearing. These young parasites measure12-27 /A x 9-21 (i. All later stages that we have seen are definitelyextracellular and lie in the gut lumen. The epimerite seems togrow very quickly at first, and in a specimen 100/* long measuresabout one-quarter of the total length. The little teeth on thepellicle are relatively much larger at this stage, and we concludethat they do not increase in size as the deutomerite grows.
When full-grown, the gregarines lose their hold on theepithelium and associate in pairs. Usually the members of apair are of approximately the same size. They lie side by sideand head to tail (fig. 31, PI. 21). The epimerite persists for atime; it gradually curves over on to the free side of themain segment (fig. 32, PI. 21), and is apparently absorbed; wehave never found that it is cast off entire as in the last species.The gametocytes become firmly attached to one another bycircular areas in the middle of their apposed sides; they thenshorten and thicken, so that the pair has the outline of afigure 8. For a time it is possible to detect the characteristicdenticles on the pellicle; but when these disappear, we cannotdistinguish the association stage of this gregarine from that ofits fellow dicystid, except by the nuclear structure in stainedpreparations; for a time after association the gametocyticnuclei of L e c y t h i o n stain much more faintly and evenlythan those of H e n t s c h e l i a and still show the two karyo-somes, one much smaller and more siderophilous than the other.

456 D. L. MACKINNON AND H. N. RAY
In this condition the paired gametocytes are voided with thefaecal balls, and their further development must take place inthe sea.
SPORE FORMATION IN GREGARINES FROM THE GUT OP
T H A L A S S E M A .
We hoped to be able to trace the spore formation of ourgregarines. But though we have found gametocysts showingnuclear division, though we have seen gametes and watchedthem copulate, and though we have found ripe sporocysts andcounted the sporozoites they contain, we are unable to statewith certainty to which species of gregarine these various stagesbelong.
When they leave the body of the host, the paired gameto-cytes of H e n t s c h e l i a and of L e c y t h i o n are, as we havesaid, indistinguishable from one another except when stained,and then only if the nuclei are unchanged. And the problemis complicated by the fact that a peculiar acephaline gregarinewhich may also be present in the gut of T h a l a s s e m a hasdimensions comparable with the gametocytes of the twodicystids there. We have only twice seen this third gregarineassociating, and then in sections of a worm that contained nogregarines but the acephaline, whereas the dicystids seem toassociate very readily. Accordingly, we might infer that mostof the gametocysts we find mixed up with the excrement belongto one or other of the dicystids; but we are bound to admitthat this is mere conjecture: the acephaline may have secretbouts of association that we have not detected. For it is notpossible to observe through the thick body-wall of a living wormwhat are its intestinal parasites; and as we have had to relyfor our study of spore formation on what we found casually inthe deposited faecal balls, we may have been dealing with anassemblage of three types of gametocyst belonging to as manydifferent species. Investigation is further rendered difficult bythe fact that, as the specimens of parasitized T h a l a s s e m an e p t u n i almost always come from relatively deep water, theirgregarines must be accustomed to forming their spores in condi-tions that we cannot reproduce satisfactorily in the laboratory.

DICYSTID GREGARINES 457
Indeed, for a long time we could not trace further developmentin any of the ejected pairs; their nuclei sometimes elongatedand became longitudinally folded on the surface; but after thatthey lay unchanged in the sea-water for some days and thendisintegrated. But we continued our search, and we have nowa number of isolated facts to record, though whether theseshould be pieced together to make one developmental story isquite uncertain. It seems important, we notice, to examine thefaecal material as soon as possible after the worms have cometo the laboratory; gametocysts found then seem to be morehealthy and show greater power of further development whenput in a moist chamber for observation than those collectedlater on. Moreover, the echiurids rather quickly becomedefaunated and the chances of obtaining numerous associatingpairs becomes less and less after a few days.
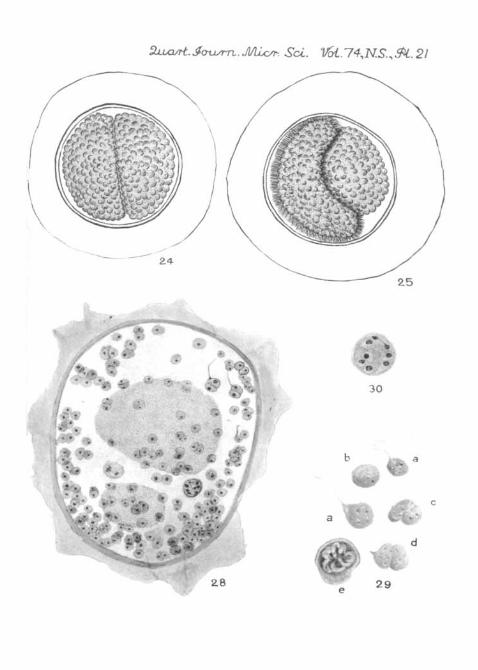
After they have been in the water for some hours, the pairedgametocytes, forming together a figure of 8, become moreclosely adpressed along the line of contact, and, where theytouch one another, their surfaces tend to become interfolded.The 'waist' of the figure 8 thus disappears and the combinedbodies have an elliptical or even a circular outline. A membraneappears surrounding this, and beyond the membrane is secreteda relatively thick, transparent envelope (fig. 24, PL 21), whichseems to be gelatinous and sticky, for bacteria and other debristend to adhere to its surface. A comparable envelope is formedround the gametocytes of N i n a ( P t e r o c e p h a l u s ) (seeLeger and Duboscq (1909)). On fixation this outer envelopeshrinks very much (figs. 26 and 28, PL 21).
Sections of fixed and stained cysts in a late figure 8 stageshow that the nuclei of the gametocytes become broad spindles,with the chromatin arranged in strings of granules stretchingfrom pole to pole; the nuclear membrane is much folded and weget the impression that fluid has passed from the nucleus intothe cytoplasm. We have not seen the completion of the firstnuclear division. The next stage we find shows the surface ofthe gametocytes becoming hummocky, and it is possible todetect, even in the living condition, that a fairly large number ofnuclei are disposed near the periphery. Pig. 26 (PL 21) is drawn

458 D. L. MACKINNON AND H. N. RAY
from a whole mount of such a cyst; it can be seen that some ofthe nuclei are dividing mitotically. Fig. 27, a, b, c, d (PI. 21)shows some of these mitotic figures, drawn from sections ofan older gametocyst; in a number of these spindles it appearsthat there are only three chromosomes.
The gametes are very slow to develop; but, as we have said,the laboratory conditions must be far from what is normal.The gametes appear as clear beads on the surface (fig. 24, PL 21),each containing a few refringent granules; in the centre of thegroup there is a large, dense, residual mass. Each gametemeasures 4-5-6/u, in diameter; those from one gametocyte areslightly larger than those from the other. In one specimen keptunder observation in the moist chamber, spherical gameteswere noticed at 11.30 a.m., and at 4 p.m. that day they were inpairs; the copulae, however, developed no further. In anothercyst in which the gametes had just formed, we noticed peculiarmovements; and while we were observing it the enveloperuptured and the gametes escaped. We then saw that the twokinds differed from one another not only in size, but in structureand behaviour. The smaller ones, measuring about 4-5/x indiameter, remained inert; whereas the larger ones, about 5—6/u. ingreatest diameter (fig. 29 a, PL 21), had become pear-shapedand moved about by means of a fairly stout flagellum arisingfrom the more pointed end. This flagellum was twice, or eventhree times, the length of the body, and, as the gamete moved,the flagellum was carried in advance. The body itself alsoshowed 'metabolic' movements, and the whole thing curiouslyresembled the flagellated stage of certain mycetozoa. Theflagellum was sometimes absorbed, and then the gamete con-tinued to move actively from side to side the clearer, pointedprominence whence the flagellum had vanished. We saw someof the flagellated gametes attach themselves to the smaller,inert gametes and gradually fuse with these (fig. 29 c, PL 21);the copula continued to move for some time by means of thegradually shortening flagellum that projected from it.
The sporocysts found in another cyst were approximatelyspherical and measured 9-10 n in diameter. The envelope wasquite well developed, but without any trace of polar thickenings.

DICYSTID GREGARINES 459
There was a small residual group of granules in the interior,and we could see the sporozoites around it, but they lay sotwisted within the sporocyst that we could not count them withcertainty. We treated some of these ripe sporocysts withWeigert's iodine solution, and one of them burst, the containedsporozoites spreading out then in such a way that it was easyto see that there were eight (PI. 29 e).
We have had so small a material to deal with that we cangive no detailed information about the gametic structure. Westained smears of the copulating gametes described above, andwe cut sections of two other cysts that contained gametes; butthe results were not very helpful. We expected to find that themale, flagellated gametes would conform to the plan of structurerevealed by Leger (1904) and by Leger and Duboscq (1909) insome tricystid gregarines in which there is pronounced aniso-gamy; and it may be that, with better stained preparations,we should get results more closely comparable with those of theFrench protozoologists. Admittedly, the copulating gametes weobserved were in conditions somewhat liable to produce abnor-mality of form, for sea-water must have got mixed with thecontents of the ruptured gametocyst, and we cannot believethat rupture of the cyst before sporocyst formation is at allusual. But they moved about actively and found their partnersas though they were quite healthy. Accordingly we werepuzzled to notice that the male gamete moves flagellum-endfirst, whereas it would appear that in Leger and Duboscq'sgregarines, the flagellum is p o s t e r i o r . We found a gametocystwith developing gametes, in which one of the gametocytes wascovered with fine, hair-like, cytoplasmic processes (fig. 25,PI. 21). These were immobile and looked like the outgrowthson the surface of certain neosporidians. If they were, as weshould be inclined to suppose, the sprouting rudiments of theflagella, then the flagellar end of the male gametes here iscertainly the reverse of what it is in S t y l o r h y n c h u s andother anisogamous gregarines that have been observed; for inthese the flagellum develops from the drawn-out region ofattachment of the male gamete to the residual mass behind it.
NO. 295 H h

460 D. H. MACKINNON AND H. N. RAY
THE SYSTEMATIC POSITION OF THE DICYSTID GREGARINES.
At one time there was a tendency among systematists totreat the dicystid gregarines as a group very distinct fromother cephalines. Brasil (1909) may have been right in supposingthat Dol iocys t i s is more nearly allied to the acephalinesthan it is to true septate forms, and Karrmi (1922), followedby Bhatia (1930), definitely placed Lecudina (Dolio-cystis) among the Acepha l ina . On the other hand, theepimerites of Schne ide r i a , Sycia , P o l y r h a b d i n a ,H e n t s c h e l i a , and Lecy th ion are as complex as those ofany tricystid.
Leger (1892) with his usual good sense remarked, 'II resultede ces observations que les differentes formes de dicystideesvraies se relient aux polycystidees par des transitions insensibles,et que la presence d'un septum ne peut constituer un caracteremorphologique serieux, susceptible d'etre invoque comme based'une classification des Gregarines.' We think that there isjustification for Leger's opinion and that it needs fresh emphasis.
Kamm (1922), as we have said, did not regard Lecud ina(Doliocystis) as a true dicystid. She created a new poly-cystid family, Polyrhabdinidae, to include P o l y r h a b d i n a ,Sycia , and Ul iv ina . The definition runs as follows:'Polycystid (septate) gregarines inhabiting the digestive tractof polychaetes. Epimerites varied,'and she adds the somewhatinconsequent remark that they ' stand near the border-line withthe Acepha l ina because of their presence only in poly-chaetes'. Schne ide r i a , being a parasite in insects, finds noplace here. The new genera we have described might fall withinthe definition of Polyrhabdinidae given by Kamm, since theechiurid worms are generally regarded as allied to the poly-chaetes. But our observations show that H e n t s c h e l i a , atany rate, differs very much in its behaviour from P o l y r h a b -dina and from Sycia .
It is abundantly clear that the dicystid gregarines are aheterogeneous group. Some occur in insects, others in polychaeteworms, and we now record two from an echiurid. Some, suchas Dol iocys t i s (Lecudina?) and H e n t s c h e l i a , are

DICYSTID GBBGABINBS 461
intracellular in the early stages of their life within the gut, andtheir epimeritic segment always remains intracellular; theindications are that in others, such as Polyrhabdina andLecythion, the intracellular stage, if it occurs at all, istransitory, and the epimerite of the adult is certainly extra-cellular. Doliocystis tends to lose its epimerite early;Lecythion retains the epimerite even at association. Dolio-cyst is , Sycia, and Schneideria apparently form theirspores while still in the host's body; whereas the dicystids fromThalassema, and probably also the species of Polyrhab-dina in Scolelepis and Po 1 ydora, require the stimulusof sea-water before they can proceed far with sporogony.
Parallel differences exist between tricystid gregarines. Insome tricystids the epimerite lies within the host-cell; in others,such as Nina and its allies, the epimerite is external to thecell. Genera differ from one another in the degree of develop-ment and in the persistence of the epimerite. The majorityform their spores within the host's body; but in some, such asPhia lo ides , the cysts do not mature until they are in thewater in which the infected animal lives.
Until protozoologists will be at the trouble to examinefor themselves the gregarines so inadequately described byMingazzini and his contemporaries, they cannot be in aposition to deal with the question of classification. Theconfusion that has arisen over the nomenclature, for instance,is mainly due to later authors having quoted uncritically state-ments made at a time when gregarines were scarcely known.Only a thorough knowledge of the life-histories will enable usto judge of the affinities of the dicystid gregarines with oneanother and with other forms. The fact that, among dicystids,Sycia and P o l y r h a b d i n a are infected with species ofMe t schn ikove l l a whereas H e n t s c h e l i a and Lecy-th ion are never parasitized in this way, may have its signifi-cance ; but in determining possible relationships of such gre-garines it is obvious that knowledge of the spore structure isthe matter of prime importance.
H h 2

462 D. L. MACKINNON AND H. N. RAY
SUMMARY.
1. Two species of the dicystid gregarine P o l y r h a b d i n aMingazzini have been studied in polychaete worms fromPlymouth.
(a) P o l y r h a b d i n a s p i o n i s is not the organism describedby Kolliker as G r e g a r i n a s p i o n i s , and should benamed P o l y r h a b d i n a s p i o n i s Mingazzini.
(b) We do not consider that the epimerite of P o l y r h a b -d i n a s p i o n i s is intracellular, nor do we believe thatit is an amoeboid and food-absorbing structure. Thespecimens we have found in S c o l e l e p i s f u l i g i n o s adiffer in other details from those described by Caulleryand Mesnil in the same worm from another locality, andmay constitute a new variety.
(c) The dicystid gregarine from the intestine of P o l y d o r af l ava is described by us. This may be the organismcalled by Caullery and Mesnil P o l y r h a b d i n ap o l y d o r a e , but we do not agree that this name issynonymous with D o l i o c y s t i s p o l y d o r a e Leger.
2. We describe two new dicystid gregarines from the intes-tines of the echiurid worm T h a l a s s e m a n e p t u n i Gartnerat Plymouth. These we have named H e n t s c h e l i a t h a l a s -sema e n.gen., n.sp., and L e c y t h i o n t h a l a s s e m a en.gen., n.sp.
3. Association was frequently observed in the gregarinesfrom T h a l a s s e m a . The gametocytes do not normallydevelop further until they have been passed out with theexcrement. In sea-water a cyst is formed and spores developed.Anisogamy, with flagellated male gametes, has been observed.
4. The dicystid gregarines are a heterogeneous group. Thosewith simple, transitory epimerite may be related to the acepha-lines; but the species dealt with in this paper are almost as com-plicated in structure as tricystids, and seem to differ from theseonly in the absence of a septum dividing a protomeritic froma deutomeritic segment.

DICYSTID GREGARINES 463
We wish to acknowledge grants-in-aid received by one of us(H. N. R.) from the High Commissioner for India and fromthe Layton Research Fund of King's College.
REFERENCES.
Bhatia, B. L. (1930).—"Synopsis of the genera and classification ofhaplocyte gregarines", 'Parasitology', vol. xxii.
Brasil, L. (1908).—"Le genre Doliocystis Leger", 'C. R. Acad. Sc. Paris',vol. cxlvi.
(1909).—"Documents sur quelques sporozoaires d'annelides", 'Arch.f. Protistenk.', vol. 16.
Caullery, M., and Mesnil, F. (1897).—"Sur un type nouveau (Metchniko-vella n.g.) d'organismes parasites des gregarines", 'C. R. Soc. Biol.Paris', Ser. 4, vol. 4.
(1901).—"Le parasitisme intracellulaire et la multiplication asexueedes gregarines", ibid., vol. liii.
(1914a).—"Sur 1'existence de gregarines dicystidees chez les anne-lidea polychetes", ibid., vol. lxxvii.
(19146).—"Sur les Metchnikovellidae et autres protistes parasitesdes gregarines d'annelides", ibid., vol. lxxvii.
Cecconi, J. (1905).—"Sur l'Anchorina sagittata Leuck., parasite de laCapitella capitata O. Fabr", 'Arch. f. Protistenk.', vol. 6.
Kamm, M. E. (1922).—"Studies on Gregarines", 'Illinois Biol. Monogr.',vol. vii.
Kolliker, A. (1845).—"Die Lehre von der thierischen Zelle", 'Zeitschr.f. wiss. Bot.', vol. i (ii).
(1849).—"Beitrage zur Kenntnis niederer Thiere", 'Zeitschr. f. wiss.Zool.', vol. i.
Lankester, E. R. (1881).—"On Thalassema neptuni Gaertner", 'Zool.Anz.', Jahrg. 4.
Leger, L. (1892).—"Recherches sur les gregarines", 'Tablettes zoologiques',vol. iii.
(1893).—"L'evolution des gregarines intestinales des vers marins",'C. R. Acad. Sc. Paris', vol. cxvi.
(1904).—"La reproduction sexuee chez les Stylorhynchus", 'Arch.f. Protistenk.', vol. 4.
Leger, L., and Duboscq, O. (1909).—"Etudes sur la sexualite chez lesgregarines", ibid., vol. 17.
Luhe, M. (1904).—"Bau und Entwickelung der Gregarinen", ibid., vol. 4.Mackinnon, D. L., and Ray, H. N. (1929).—"Lankester's 'gregarine' from
the eggs of Thalassema neptuni", 'Nature', vol. 124.Mesnil, F. (1907).—"Analyse de Brasil (1907)", 'Bull. Inst. Past.', vol. 5.Mesnil, F., and Caullery, M. (1897).—"Sur trois sporozoaires parasites de
la Capitella capitata", 'C. R. Soc. Biol. Paris', vol. iv.

464 D. L. MACKINNON AND H. N. BAY
Mingazzini, P. (1891a).—"Sulla distribuzione dello gregarine policistidee",'Rend. di. Accad. Line.', vol. vii.
(18916).—"Gregarine monooistidee nuove o poco conosoiute",ibid., vol. vii.
(1893).—"Contribute alia conoscenza degli Sporozoi", 'Ric. Labor.di. Anat. norm. d. Univ. Roma', vol. iii.
Ray, H. N. (1930a).—"Studies on some Sporozoa in polychaete worms.I. Gregarines of the genus Selenidium", 'Parasitology', vol. xxii.
(19306).—"Studies on some Sporozoa in polychaete worms. II.Dorisiella scolelepidis n.gen., n.sp.", ibid., vol. xxii.
EXPLANATION OF PLATES 20 AND 21.
All figures drawn either from living specimens or fromsmears and sections fixed in Brasil's modification of Bouin-Duboscq's fluid and stained with Heidenhain's iron haema-toxylin. A camera lucida was employed. The microscopewas a large Leitz model, fitted with 1/6" and 1/12" (oilimmersion) objectives and oculars 4, 8, and 12 (these last twocompensating oculars of Zeiss).
PLATE 20.
Po ly rhabd ina spionis n.var. b i furca ta .Fig. 1.—Trophozoite. Drawn from a smear. X 350.Pig. 2.—Young trophozoite. Drawn from a section, x 350.Kg. 3.—Transverse section of a trophozoite. X 1,000.Fig. 4.—Epimerite of the gregarine attached to the epithelium (cilia
omitted). The nearer wall has been shaved away in the section. X 1,400.Fig. 5.—A more slender trophozoite parasitized by Metchnikovel la
spionis . The parasite is in the hyphal stage. X 350.Fig. 6.—A trophozoite crammed with the cysts of Metchnikovel la .
The spores are shown in two cysts only. X 350.Fig. 7.—Cyst of Metchnikovel la spionis from Scolelepis
ful iginosa. X 1,400.
Po ly rhabd ina polydorae (?).Fig. 8.—Large trophozoite attached to the epithelium (cilia of the cells
omitted). Drawn from a section. X 350.Fig. 9.—Epimerite, showing detail of structure. X 1,400.Fig. 10.—Transverse section of a trophozoite filled with the cysts of
Metchnikovel la cau l le ry i . Note the degenerate nucleus of thehost. X 350.
Fig. 11.—Portion of the endoplasm of a parasitized trophozoite showing

DICYSTID OUtEGARINES 465
the hyphal stage of M e t c h n i k o v e l l a c a u l l e r y i . From a section.X 350.
Fig. 12.—Later stage in the hyphal development of the parasite; thehypha is becoming segmented into bead-like spores. From a section.X 1,400.
Fig. 13.—Cyst and spores of M e t c h n i k o v e l l a c a u l l e r y i : (a) theentire cyst, X 667; (6) one end of the cyst, x 2,000; (c) three spores,X 1,300.
H e n t s c h e l i a t h a l a s s e m a e n.gen., n. sp.Fig. 14.—Young intracellular trophozoite. From a section. Note the
hypertrophy of the host-cell and its projection into the lumen of the gut.X 1,000.
Fig. 15.—Mature trophozoite. From a smear. Note the intracellularepimerite. X 350.
Fig. 16.—End-on view of the epimerite. X 933-
L e c y t h i o n t h a l a s s e m a e n.gen., n.sp.Fig. 17 a.—Mature trophozoite, attached to the epithelium. From a
section. X 400.Fig. 17 6.—A small portion of the pellicle of the same, showing the
characteristic spines. X 1,400.Fig. 18.—Transverse section of a trophozoite in the region of the nucleus.
X 600.Fig. 19.—Young trophozoite, with epimerite still very short. From
a smear. X 550.Fig. 20.—Two young intracellular ( ?) gregarines. From a smear. X 667.Fig. 21.—Epimerite of a full-grown gregarine, greatly enlarged to show
detail of structure. Note the method of attachment to the epithelial cell(cilia omitted). X 1,400.
PLATE 21.
Fig. 22.—Hentschelia t h a l a s s e m a e . Associating gregarines.Note that one still retains its intracellular epimerite. The nuclei areelongating. From a smear (striations omitted). X 500.
Fig. 23.—Hentschelia t h a l a s s e m a e . Association; figure 8 stage.From a smear. X 500.
Fig. 24. Living gametocyst of (?) some hours after evacuation from theworm. Note the thick ectocyst. Gametes are developing on the surface ofthe gametocytes, and those from one are rather larger than those from theother. X 500.
Fig. 25.—Another living gametocyst, in which one of the partners iscovered with fine, hair-like processes. X 500.
Fig. 26.—Fixed and stained gametocyst, showing nuclei in process ofdivision. From a smear of the evacuated faecal balls of T h a l a s s o m a .X 500.
Fig. 27, a, h, c, and d. Four stages in the division of nuclei in an older

466 D. L. MACKINNON AND H. N. KAY
gametocyst. From a section. In two of the figures it can plainly be seenthat the number of chromosomes at this stage is three. X 2,100.
Fig. 28.—Section of a gametocyst in which gamete-formation is finished.The gametes are fusing in pairs, and some of these show a flagellum. Afew sporoblasts are shown, and one finished spore. In the centre of thecyst lies the residual body. (Only a few of the gametes that appeared inthe section have been drawn.) X 700.
Kg. 29.—Living gametes: (a) flagellated male gametes; (b) passivefemale gamete; (c) and (d) copulae; (e) ruptured sporocyst showing theeight sporozoites. X 1,800.
Fig. 30.—Sporoblast (fixed and stained) containing eight nuclei.X 1,600.
Fig. 31.—Lecythion t h a l a s s e m a e . Association. From a smear.(Pellicular striations omitted.) X 500.
Fig. 32.—A slightly later stage, in which the epimeritea have folded overand are being absorbed. From a smear. (Striationa omitted.) X 600.

14-
15
dot.

/
^ I3c
13b
10
13a
r •
18
19
\la I7b 20
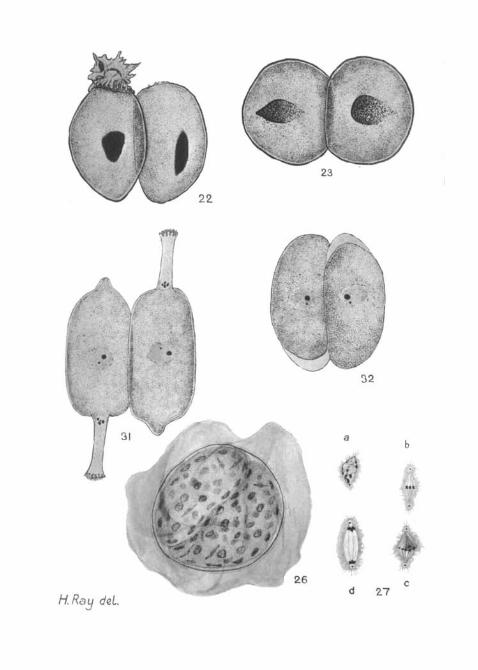
TV31
H. Ray deL
f26
32
md 27
. Jvusrn. Muyr. Sol. Vol.l^N.S.^i. 21
m30
28