observations on egg hatching in the estuarine crab sesarma ... · completed, hatching...
TRANSCRIPT

1 Manuscript accepted 15 October 1991.20kayama University, College of Liberal Arts and Sciences, Tsushima 2-1-1, Okayama 700, Japan.
NEWLY OVIPOSITED EGGS of decapod crustaceans are attached to the abdominal appendages of the females, where they are ventilatedduring development by movements of thepleopods. When embryonic development iscompleted, hatching (egg-membrane ruptureor breakage) occurs. Hatching in marine species occurs during the larval stage, and larvalrelease is associated with vigorous abdominalfanning behavior (e.g., DeCoursey 1979).
A remarkable feature of larval release issynchrony with environmental cues such asday-night, tidal, or lunar cycles. Nocturnal release rhythms have been observed for a number of crustaceans. In the lobster, Homarus,larvae are released by the female a few minutesafter darkness on successive days (Ennis 1973,Branford 1978). Hatch time of the fiddlercrab, Vea, is closely correlated with high tideat night (DeCoursey 1979). Precisely timedrhythms in larval release implies that hatching
Pacific Science (1992), vol. 46, no. 4: 484-494© 1992 by University of Hawaii Press. All rights reserved
Observations on Egg Hatching in the Estuarine Crab Sesarma haematocheir1
MASAYUKI SAIGUSA2
ABSTRACT: A female of the terrestrial crab Sesarma haematoeheir incubates30,000-50,000 eggs on her abdomen. After 1 month of embryonic development,zoeae larvae are released into estuarine waters within 3-5 sec by means ofvigorousfanning motions of the abdomen. Hatching (breakage of the outer egg membrane) occurs on land just before larval release. The release behavior itself doesnot cause rupture of the egg case, nor has the presence of a "hatching enzyme"been obviously demonstrated. Hatching seems to be induced by mechanicalrupture of the egg case. The pressure responsible for hatching may be producedeither by the larva itself, or by osmotic swelling of thin inner membranes encasingthe larva, although neither of these hypotheses is sufficient at present to explainthe complete hatching mechanism. If hatching is explained by such mechanisms,then there remains the question ofhow hatching is synchronized among the largenumber of embryos attached to the female. Hatching of detached embryos issynchronized to some extent, but the degree of synchronization is less than thatoccurring in the larvae carried by the female. This observation suggests thatstimuli from the female are important in establishing highly synchronizedhatching. The ecological significance of the hatching system is also discussed.
time is also synchronized within a batch ofeggs attached to a female.
Little is known about the hatching mechanism in Crustacea. It may be mechanical,nonmechanical, or a combination of both.Osmotic hatching has been suggested for anumber ofcrustaceans. In this hypothesis, therigid outer egg membrane may be burst byosmotic pressure inside the thin inner membrane (Yonge 1937, Marshall and Orr 1954,Davis 1965), or by pressure caused by theswelling larva itself (Davis 1964).
The crab Sesarma haematocheir burrows onland surrounding small estuaries in Japan andalong the coastlines of Southeast Asia. Inearly summer, a female incubates 30,00050,000 eggs in her folded abdomen for ca. Imonth. She emerges at riverside or seashorewhere the zoeae are released. About the timeof high tide at night, the females immersethemselves in water and vigorously flex theirabdomens as clouds of newly hatched larvaeare swept away. This larval release behavioris completed in 3-5 sec, and all the zoeae arereleased from the pleopods. After larval release, the females return to land and incubate
484

Egg Hatching in an Estuarine Crab-SAIGUSA
the next clutch, which will appear within afew days. Each female incubates and releaseslarvae two or three times between July andSeptember.
It was obvious from field observations thatembryos carried by the female had not yethatched when the female appeared on theshore. That the behavior associated with release is an extremely short-lived event suggestsseveral possible mechanisms enabling larvalemergence from the egg membranes: (I) physical stimuli from the female during releasebehavior (i.e., vigorous fanning movements ofpleopods and abdomen cause breakage of theegg membrane); (2) a hatching enzyme secreted by the zoea itself or the female thatdigests the egg case; or (3) some other mechanism that controls time of hatching. To determine which of the possibilities is correct, fieldstudies were carried out to determine whenand where the larvae hatch, and the hatchingprocess was examined by microscope. Theresults raised questions about synchrony inhatching, and investigations were focused onwhether detached eggs hatch with the samesynchrony as observed when the eggs areattached to a female.
MATERJALS AND METHODS
Field Studies
This study was conducted at two sites onthe coast of the Inland Sea, Japan: the marginof the outlet of a small river at Kasaokawhere larval release activity had been observed (Saigusa 1982), and at a similar place atUshimado, Okayama Prefecture, 70 km eastof Kasaoka. A scanning electron microscopewas utilized at the latter site.
Larval release activity of S. haematocheircoincides with high tide at night (Saigusa 1982,1985). To find out when the embryos hatch,ovigerous females approaching riverside werecaptured about dusk. They were confined inempty plastic buckets and examined for hatching. Hatching was distinguished by the following features: (1) at hatching, the whole spongeor egg mass becomes somewhat sticky (that is,when touched with a finger, the hatched zoeae
485
stick to it); and (2) females with sticky eggsfroth slightly at the mouth. The sponge wasfrequently checked and the number of femaleswith hatched larvae recorded at intervals ofabout 15 min. Collecting and counting crabswas accomplished with a hand-held flashlight.
Stereomicroscope Observations
Ovigerous females were captured from thefield at Ushimado or Kasaoka and broughtto the laboratory. Small samples of eggs(embryos folded within the membranes) wereremoved from females expected to releaselarvae that night. The samples were placed ina laboratory dish containing a small quantityof diluted seawater. Salinity was not recordedas there is no clear evidence for osmoticeffects, even in distilled water. Observationson hatching were made with a stereomicroscope magnifying 20-90 times.
Scanning Electron Microscope Observations
The empty egg cases remaining immediately after larval release, and recently hatchedlarvae detached from the pleopods of femaleswith forceps, were prepared for the scanningelectron microscope (SEM) to analyze themfor evidence of a hatching enzyme, whichwould be indicated by dissolution on the eggmembrane enclosing the embryo. The sampleswere fixed for 2 hr in a solution containing2.5% glutaraldehyde in 0.1 M Na cacodylatebuffer (pH 7.4). After fixation they werewashed twice in a 1: 1 mixture of 0.1 M Nacacodylate buffer and distilled water, dehydrated through a graded series of ethanol,and dried in a Hitachi HCP- I critical pointapparatus. They were mounted on metallicstubs with double-stick tape, plated with gold,and examined.
Hatching ofDetached Embryos
Detached embryos were examined to determine whether they hatch at the same time asthose attached to the female. Crabs withembryos estimated to hatch within 2 days(based on the color of embryos) were chosenfor these experiments. Detached embryos in

486
nonaerated water, detached embryos withaeration and water turbulence, and embryosattached to the female were utilized in theexperiments. For the first condition, 3001000 embryos were removed 3-3.5 hr beforedark and placed in plastic cases (8 cm in diam.,6 cm in depth) approximately one-third filledwith dilute seawater (10%0). At 30-min intervals, the swimming zoeae were counted andremoved by pipette. A small flashlight (withtwo 1.5-V batteries) covered with red cellophane was used during the dark period.
All experiments on detached embryos wereperformed in experimental rooms in whichlight and temperature were controlled. A 15: 9(L: D) photoperiod similar to that in the fieldwas employed. Temperature was maintainedat 23 ± 1.5°C. As shown in Figure 1, somedetached embryos were suspended on a horizontal nylon thread stretched across the center of an opening of a small plastic case, andthe case was submerged; at 0.5-hr intervals thecase was transferred to another beaker thatcontained a similar quantity of diluted sea-
PACIFIC SCIENCE, Volume 46, October 1992
water. The berried crabs were separately confined in vessels containing diluted seawater(ca. I cm deep), and hatching and larvalrelease were monitored simultaneously.
RESULTS
Synchronous Hatching and Larval Release
The purpose of the field studies was to determine when and where the embryos hatched.When females were collected at riverside aboutdusk, hatching had not yet occurred. Hatching from free animals began just after sunsetand reached a peak when the tide was highest(Figure 2, upper panel). Hatching was estimated to occur within a few minutes, andperhaps in less than I min for most of them,although it was impossible to determine theexact time required for completion of hatching. As soon as hatching was completed, thefemale produced a small amount of froth atthe mouth. Subsequently, when the female
2cm
~air
.....'. '. '. '. ,.". ,
FIGURE I. A device to provide both aeration and turbulence to eggs detached from the female. E, an egg clusterattached to a portion of the pleopods. AS, air stone. When air is forced into the water, the eggs were floated abovethe nylon thread, shown on the right.

Egg Hatching in an Estuarine Crab-SAIGUSA 487
Microscope Observations
Eggs attached to a female are at first darkbrown because of the accumulation of yolk.Four to 5 days before hatching the amount of
FIGURE 2. Distribution of hatch time recorded in thefield at Kasaoka. Collection of females was made at1830-1900 hours. Vertical straight lines (HW) indicatethe time of night high water, SS the time of sunset. Thevertical axis of the upper panel shows relative number offemales (LR) releasing larvae at night (see Saigusa 1982for larval release activity by the female; although time oflarval release by the population was examined in the field,time ofday of release in each female was roughly recordedin 1984).
yolk rapidly diminishes and the sponge turnsbrownish green. Egg size at this stage averaged 0.34 x 0.32 mm (in contrast to the estimates of young eggs at 0.2 x 0.28 mm). Wellformed compound eyes, chromatophores, andglobules of orange yolk are clearly visiblethrough the transparent egg capsule, as areoccasional movements of the body and heartbeat. Swelling, if any, was slight as hatchingapproached.
Before rupture of the egg membrane, theabdomen of the embryo curled ventrally, andthere was no fluid-filled space between thebody and the egg case. Hatching commencedwith the sudden rupture of the egg capsule. Nomotion of the body was observed at that time.In Figure 3a, the egg capsule of larva D hasalready ruptured, and the larva remains within the broken membrane. Larva C also seemsto have sloughed off the egg case, but othershave not yet hatched. Hatching of larva B isvisible in Figure 3b, and that of larva A inFigure 3c. There is a slight but obvious difference in relative positions of both eyes (bigblack spots) in larva A before and afterhatching caused by a rotation of the larvaassociated with breakage of the egg membrane. This motion stopped when the end ofthe telson was obstructed by the broken membrane. Hatched larvae remain in the foldedabdomen of the female until she shakes themoff into the water.
When the egg capsule ruptures, the second,thin membrane enclosing the larva is sometimes seen to remain intact (Figure 4a). Observations in nonaerated water show that larvaeemerging from the egg case remain motionless(e.g., larva D in Figure 3a-d) except for strongreverse peristalsis in the rectum, which appeared to be associated with an influx ofwater. As reverse peristalsis proceeds, thelarval body swells and the dorsal spine extends. When the thoracic appendages andtelson are fully developed, the larva begins toswim (Figure 4b).
Observations by SEM also indicated thatthe empty egg case consists of two membranes, an outer thick membrane serving asthe protective shell of the egg (OM in Figure5a and 5c) and an inner, thin membrane liningthe egg case (LM in Figure 5c). In addition, a
23
Aug.6.1984
19 21Time of day
II
I
I HWI
55...:
30
2010OL--f---L'P~L--,---,---r-----,
L.QI.0E::J
Z
17
'05
was splashed with water, she immediatelyvibrated her body with rapid pumping movements of the abdomen and shook all the larvaeinto the water. Ovigerous females collected atother times and placed in a similar vessel oftenexhibited the same kind of behavior, but nohatched larvae appeared at that time. Theseobservations clearly indicate that hatching isnot caused by the vigorous pumping movements of the female crab, but is induced as aresult of another synchronized process.
Hatching from confined females occurredsomewhat later than in free animals: larvalrelease of free animals occurred within 2 hrafter sunset, but hatch peaks of the confinedfemales were delayed 1-1.5 hr (Figure 2, lowerpanel). Time required for hatching in confinedfemales was also longer than in free animals;in some animals it took 10-20 min until theentire sponge became sticky.

300f
FIGURE 3. Hatching of detached embryos in S. haematocheir. Note the progress of embryos A and B. Hatching ofthe sponge carried by the mother crab occurred at 2315 hours on 26 July 1985. Time in minutes is shown at lower left ineach figure (a-d). See text for further details. Time of detachment was about 2200 hours on 26 July.

Egg Hatching in an Estuarine Crab-SAIGUSA 489
FIGURE 4. Hatching of S. haematocheir eggs: (a) increase in volume following rupture of the outer membrane(H" Hz, and H 3)· Arrow shows a delicate, thin membrane enclosing the larva after bursting of the egg case. Otherembryos have not yet hatched. (b) zoea larva (Z) that has begun to swim after throwing off the thin membranes.

490 PACIFIC SCIENCE, Volume 46, October 1992
FIGURE 5. Micrographs obtained by SEM: (a) egg cases from which hatching occurred and zoeae escaped. OM,outer membrane. 1M, the third membrane. (b) magnification of the third membrane (1M). (c) an empty egg capsuleshowing the thin lining membrane (LM). (d) a prezoea larva after the breakage of the outer membrane. E, a compoundeye. DP, dorsal portion of the larva. OE, another egg for which hatching has not yet occurred. Scales expressed ineach figure are all 100 /lm.
third membrane with an irregular structure(1M) protruded from the empty egg case(Figure 5a-c). This is not a so-called embryonic cuticle cast off by the larva, but is a thirdmembrane investing the embryo. This structure can also be recognized with a stereomicroscope, and its presence and sticky natureare suggested by behavior of the larvae afterhatching in that the larvae are released onlyafter vigorous movement during which theabdomen is repeatedly straightened.
No evidence was obtained from the SEMviews that the egg capsule was dissolved;rather, the outer membrane was clearlycracked on breakage (Figure 5a and c; Masamichi Yamamoto and Toshiki Makioka, pers.
comm.). The larva bulges from the brokenegg case (Figure 5d) showing that half of themembrane formed a cuplike case over thetelson-which also indicates a rupture of theouter membrane. In Figure 5d, in which thecephalic portion of the larval body is directlyexposed to the outside, the third membranemay already have been shed (or might havebeen removed during fixation or other experimental manipulation).
Hatching ofDetached Embryos
A hatching profile of detached embryosmaintained in nonaerated water is shown inFigure 6a. Hatching started shortly before

Egg Hatching in an Estuarine Crab-SAIGUSA 491
larval release began and continued all night.Hatching peaks occurred 30 min after midnight in two experiments. Hatching synchronyof detached embyros was considerably improved by exposing them to strong aeration
100 a
'":ll1;1
o~ 100.0E:>z
300 b
200
'"c'EE.~
o~ 3
'".0E:>z
18 24Time of day
• +t.336
N=395
N=290
6
N=910
N=778
and turbulence (Figure 6b). For the embryosseparated on 5 September (upper panel), mostlarvae (81 %) hatched during the 2-hr intervalbetween midnight and 0200 hours. Embryosdetached on the day of larval release (lowerpanel) showed somewhat better synchrony inhatching: 95% of hatching was concentratedin 2 hr from 2300 to 0 I00 hours. Furthermore, Figure 6b shows a difference in timingbetween the two experiments: the peak ofhatching in the embryos detached on theday of larval release occurred about 1.5 hrearlier than those separated I day before.Additional experiments confirmed the initialobservations.
Yet another feature of the results shown inFigure 6a and 6b is a difference in the percentage of the hatching larvae in nonaerated andaerated water. Almost all embryos detachedon the day of larval release and maintained innonaerated water hatched that night. In contrast, only some of the embryos separated Iday before larval release and maintained inthe same conditions hatched (Table 1, uppermost column). The bodies of those embryosthat did not hatch were cloudy and apparentlymotionless. Embryos maintained in aeratedwater did not show these features (Table 1,lower columns). The implication of theseobservations is that the eggs may have diedbecause of an oxygen deficiency.
DISCUSSION
Observations by stereomicroscope indicatethat the egg cases of Sesarma consist of twomembranes, a thick outer membrane and aninner thin membrane, as in other decapods(e.g., Yonge 1937). In addition to these mem-
100
18 24Time of day
6
FIGURE 6. Distribution of hatching of eggs separatedfrom a female. (a) hatching monitored in still water. (b)hatching under strong aeration. Eggs were all detachedfrom one female. Time of separation was 1600 hours on5 September 1984 (upper panel) and 1530 hours on 6September 1984 (lower panel) in both illustrations. Timeof day of larval release by the female (vertical straightline) was at 2140 hours on 6 September (a portion ofhatched larvae began to swim in the vessel 20-30 minbefore the release of larvae, as shown by the dotted area).N, total number of hatched larvae in each experiment.
492 PACIFIC SCIENCE, Volume 46, October 1992
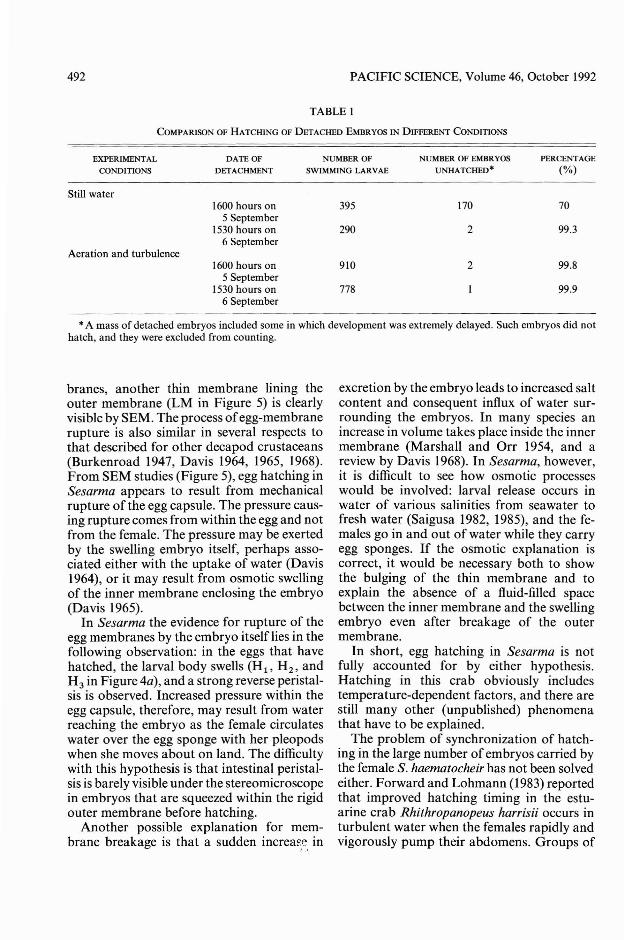
TABLE I
COMPARISON OF HATCHING OF DETACHED EMBRYOS IN DIFFERENT CONDITIONS
EXPERIMENTAL
CONDITIONS
Still water
Aeration and turbulence
DATE OF NUMBER OF NUMBER OF EMBRYOS PERCENTAGE
DETACHMENT SWIMMING LARVAE UNHATCHED· (%)
1600 hours on 395 170 705 September
1530 hours on 290 2 99.36 September
1600 hours on 910 2 99.85 September
1530 hours on 778 99.96 September
*A mass of detached embryos included some in which development was extremely delayed. Such embryos did nothatch, and they were excluded from counting.
branes, another thin membrane lining theouter membrane (LM in Figure 5) is clearlyvisible by SEM. The process ofegg-membranerupture is also similar in several respects tothat described for other decapod crustaceans(Burkenroad 1947, Davis 1964, 1965, 1968).From SEM studies (Figure 5), egg hatching inSesarma appears to result from mechanicalrupture of the egg capsule. The pressure causing rupture comes from within the egg and notfrom the female. The pressure may be exertedby the swelling embryo itself, perhaps associated either with the uptake of water (Davis1964), or it may result from osmotic swellingof the inner membrane enclosing the embryo(Davis 1965).
In Sesarma the evidence for rupture of theegg membranes by the embryo itselflies in thefollowing observation: in the eggs that havehatched, the larval body swells (Hi' H 2 , andH 3 in Figure 4a), and a strong reverse peristalsis is observed. Increased pressure within theegg capsule, therefore, may result from waterreaching the embryo as the female circulateswater over the egg sponge with her pleopodswhen she moves about on land. The difficultywith this hypothesis is that intestinal peristalsis is barely visible under the stereomicroscopein embryos that are squeezed within the rigidouter membrane before hatching.
Another possible explanation for membrane breakage is that a sudden increas? in
excretion by the embryo leads to increased saltcontent and consequent influx of water surrounding the embryos. In many species anincrease in volume takes place inside the innermembrane (Marshall and Orr 1954, and areview by Davis 1968). In Sesarma, however,it is difficult to see how osmotic processeswould be involved: larval release occurs inwater of various salinities from seawater tofresh water (Saigusa 1982, 1985), and the females go in and out of water while they carryegg sponges. If the osmotic explanation iscorrect, it would be necessary both to showthe bulging of the thin membrane and toexplain the absence of a fluid-filled spacebetween the inner membrane and the swellingembryo even after breakage of the outermembrane.
In short, egg hatching in Sesarma is notfully accounted for by either hypothesis.Hatching in this crab obviously includestemperature-dependent factors, and there arestill many other (unpublished) phenomenathat have to be explained.
The problem of synchronization of hatching in the large number of embryos carried bythe female S. haematocheir has not been solvedeither. Forward and Lohmann (1983) reportedthat improved hatching timing in the estuarine crab Rhithropanopeus harrisii occurs inturbulent water when the females rapidly andvigorously pump their abdomens. Groups of

Egg Hatching in an Estuarine Crab-SAIGUSA
larvae are released with each pump, and thoseauthors suggested that pumping behavior itself assists in breaking the egg membrane,which enhances hatching synchrony. InSesarma, however, no particular movementsof the female's abdomen are observed uponegg hatching.
What is evident is that hatching synchronyis enhanced by aeration and water turbulence(Figure 6), there is less hatching synchronyin detached eggs than in the eggs attachedto the female (although hatching synchronyimproves somewhat when eggs are detachedon the day of release), and a hatch peak ofconfined crabs is delayed (Figures 2 and 6). Asindicated from Figures 3 and 4, some of thelarvae with outer membrane breakage haddifficulty in leaving the egg capsule. So de....tached embryos exposed to vigorously aeratedwater may have been readily released from theegg capsules, which would cause a bettersynchrony of hatching. Figure 6 also indicatesthat hatching synchrony of the detached eggswas decreased and their hatch time delayed.These features can be explained in terms oftheabsence of some cue from the female. And thedelay of hatching in confined crabs (Figure 2)may imply a delay of this cue.
Since egg hatching time is surprisingly shortfor the eggs attached to the female, it mightbe possible that a substance similar to hatching enzyme secreted by either the embryo orthe female, and which softens the egg membrane, resulting in simultaneous hatching,is present. In addition, it appears that inSesarma embryos a hatching process differentfrom embryonic development is involved, andboth mechanisms for the commencement ofthis process and for synchronization of hatching must be considered in accounting for thebreakage of the egg case.
The adaptive significance of the hatchingsystem is evident for estuarine terrestrial crabssuch as Sesarma. Because larval release occursat the water's edge, synchronization of hatchtime ensures that all the larvae are released atthe same time. Hatching just before liberationalso ensures that the larvae will not sufferoxygen deficiency and other adverse effectsthat might occur if they are held by the femalefor a longer time.
493
ACKNOWLEDGMENTS
The SEM micrographs were made at theUshimado Marine Biological Laboratory,Okayama University. I am indebted to Professors Masao Yoshida (deceased) and Masamichi Yamamoto at the laboratory, who gaveme facilities for the work. Dr. TaichirowGotow, Mie University, helped me in preparing samples for SEM and with photography.
LITERATURE CITED
BRANFORD, J. R. 1978. The influence of daylength, temperature and season on thehatching rhythm of Homarus gammarus.J. Mar. BioI. Assoc. u.K. 58: 639-658.
BURKENROAD, M. D. 1947. Reproductive activities of decapod Crustacea. Am. Nat.81 :392-398.
DAVIS, C. C. 1964. A study of the hatchingprocess in aquatic invertebrates. XIII. Eventsofec1osion in the American lobster, Homarusamericanus Milne-Edwards (Astacura, Homaridae). Am. MidI. Nat. 72:203-210.
---. 1965. A study of the hatching processin aquatic invertebrates: XX. The blue crab,Callinectes sapidus Rathbun. ChesapeakeSci. 6:201-208.
---. 1968. Mechanisms of hatching inaquatic invertebrate eggs. Oceanogr. Mar.BioI. Annu. Rev. 6:325-376.
DECOURSEY, P. J. 1979. Egg-hatching rhythmsin three species of fiddler crabs. Pages399-406 in E. Naylor and R. G. Hartnoll,eds. Cyclic phenomena in marine plantsand animals. Pergamon Press, Oxford.
ENNIS, G. P. 1973. Endogenous rhythmicityassociated with larval hatching in the lobster Homarus gammarus. J. Mar. BioI.Assoc. U.K. 53:531-538.
FORWARD, R. B., Jr., and K. J. LOHMANN.1983. Control of egg hatching in the crabRhithropanopeus harrisii (Gould). BioI. Bull.(Woods Hole) 165: 154-166.
MARSHALL, S. M., and A. P. ORR. 1954.Hatching in Calanusfinmarchicus and someother copepods. J. Mar. BioI. Assoc. U.K.33: 393-40 I.
SAIGUSA, M. 1982. Larval release rhythm

494
coinciding with solar day and tidal cycles inthe terrestrial crab Sesarma. BioI. Bull.(Woods Hole) 162: 371-386.
---. 1985. Tidal timing of larval releaseactivity in non tidal environment. lpn. l.Ecol. 35:243-251.
Words of the LagoonFishing and Marine LYrein the Palau District of MicronesiaR. E. JOHANNES"Records a measure of what fishermen knewabout the natural world before either anthropology or marine biology existed and leaves us inawe of all that has been forgotten."
-Oceans"A valuable book for marine biologists, anthropologists, and local people. It grew out of a deepconviction that a valuable source of knowledgewas both unnoticed and unexploited and thatthe time to tap it was overdue. Johannes hasmade his point well."-Pacific Studies$15.00 paper, illustrated
PACIFIC SCIENCE, Volume 46, October 1992
YONGE, C. M. 1937. The nature and significance of the membranes surrounding thedeveloping eggs of Homarus vulgaris andother Decapoda. Proc. Zool. Soc. Lond.ser. A 107: 499-517 (plus 1 plate).
A Natural History ofCaliforniaALLAN A. SCHOENHERRIn this comprehensive and abundantly illustrated book, Schoenherr describes a state with agreater range of landforms, a greater variety ofhabitats, and more kinds of plants and animalsthan any area of equivalent size in all of NorthAmerica. A region ofunparalleled natural beautyand richness, it is also the st;\te with the greatestnumber of endangered species.Schoenherr fosters a keen appreciation forCalifornia's natural diversity and unique beauty,much of which is threatened.California Natural History Guides$38.00 cloth, illustrated
At bookstores or order toll-free 1-800-822-6657.
University of California Press