obtención de carotenoides con la microalga scenedesmus
TRANSCRIPT

INFORME TÉCNICO DE LA OPCIÓN CURRICULAR EN LA MODALIDAD DE: PROYECTO DE INVESTIGACIÓN
INSTITUTO POLITÉCNICO NACIONAL UNIDAD PROFESIONAL INTERDISCIPLINARIA DE BIOTECNOLOGÍA
TÍTULO DEL TRABAJO:
Obtención de carotenoides con la microalga Scenedesmus incrassatulus en un biorreactor tipo
airlift
QUE PARA OBTENER EL TÍTULO DE INGENIERA BIOTECNÓLOGA
PRESENTA: XIMENA LEMUS VILLAFUERTE
México, D. F. Junio 2008
ASESORES Asesor externo Dra. Rosa Olivia Cañizares Villanueva Asesor interno Dr. Luis Fernández Linares
México, D.F. Junio 2009

RESUMEN
Los carotenoides son metabolitos primarios producidos por las microalgas para que
cumplan su función como pigmentos accesorios de la fotosíntesis y como moléculas
fotoprotectoras. Se ha reportado la producción de carotenoides secundarios en microalgas
en respuesta a algún tipo de estrés. En el presente trabajo se aplicaron dos condiciones de
estrés, nutricional y lumínico, por separado y en conjunto, para inducir la carotenogénesis
en la microalga Scenedesmus incrassatulus CLHE-Si01. Se llevaron a cabo cultivos con
fotoperiodo y con luz continua en fotobiorreactores de 2L, en un medio mineral, con un
flujo de aireación de 0.5 vvm y una iluminación de 400m-2 s-1. Una vez que los cultivos
hubieron alcanzado la máxima concentración volumétrica de carotenoides totales, se aplicó
el estrés. En caso de estrés nutricional se realizó un lavado celular y se sustituyó el medio
mineral por agua, mientras que en el caso de estrés lumínico se agregaron 350m-2 s-1 de luz.
Los resultados obtenidos indican que el estrés lumínico y nutricional estimularon la
carotenogénesis y que además, la luz continua es una forma adicional de estresar a la
célula. El crecimiento e inducción con fotoperiodo permitió obtener una mayor cantidad de
biomasa y de carotenoides, mientras que el crecimiento con luz continua aumentó el
porcentaje de carotenoides secundarios y la degradación de éstos. El aumento en los
carotenoides secundarios estuvo representado por el aumento de la concentración de
astaxantina+violaxantina. El fotobiorreactor que presentó el máximo contenido específico
de cetocarotenoides fue el sometido a estrés nutricional con fotoperiodo (1.2%).

AGRADECIMIENTOSAGRADECIMIENTOSAGRADECIMIENTOSAGRADECIMIENTOS
A mi papá, por siempre alentarme a cumplir mis metas.
A mi mamá por darme siempre su cariño y apoyo. A Fabi, por ser una verdadera hermana.
A la Dra. Rosa Olivia Cañizares, por su asesoría, por permitirme realizar el proyecto en su Laboratorio y también aprender en él.
Al Dr. Hugo Perales, por su asesoría y por enseñarme el amor a la investigación.
A Alfredo, por ser un compañero, un asesor y un amigo. A mis amigos de la UPIBI, del taller de teatro de la UPIBI y de Gato Encerrado, por ayudarme a reír cuando más lo necesitaba.
“One song glory, beyond the cheap colored lights… Time flies” [RENT]
“No hay árbol que el viento no haya sacudido.”
Proverbio hindú

ÍNDICEÍNDICEÍNDICEÍNDICE
Página
1. INTRODUCCIÓN
1.1 Los carotenoides…………………………………………………………..….
1.1.1 Descripción..……………………………………………………………..….
1.1.2 Clasificación……………………………………………………………..….
1.1.3 Características químicas……………………………………………………
1.1.5 Importancia en la salud humana …………………………………………....
1.1.6 Importancia industrial………………………………………………………
1.2 Los carotenoides y las microalgas…………………………………………
1.2.1 Descripción general…………………………………………… …………..
1.2.2 Función fisiológica…………………………………………………………...
1.2.3 Scenedesmus…………………………………………………………………………......
1.2.4 Biosíntesis ………………………………………………………………….........
1.2.5 Factores que afectan la biosíntesis……………………………………….………
1.3 Producción industrial: síntesis química vs. Biosíntesis…………………
1.4 Fotobiorreactores…………………………………………………………………
2. ANTECEDENTES……………………………………………………………….
3. JUSTIFICACIÓN E HIPÓTESIS……………………………………………...
4. OBJETIVOS……………………...…………………………………………………
5. MATERIALES Y MÉTODOS………………………………………………
5.1 Scenedesmus incrassatulus. …….……………………………………...…
5.2 Medio de Cultivo mineral……..………………………………………………...
5.3 Biorreactor airlift……………………………………………………………….
5.4 Protocolo experimental………….....…………………………………………….
1
1
2
3
3
7
9
9
10
12
15
18
20
21
21
23
25
26
25
25
25
27

6. RESULTADOS Y ANÁLISIS DE RESULTADOS
6.1 Crecimiento celular con fotoperiodo (FP) y luz continua…………………………
6.2 Fotobiorreactores con luz continua………………………………………………..
6.2.1 Cuantificación de clorofilas y carotenoides totales……………………………..
6.2.3 Contenido específico de carotenoides totales…………………………………...
6.2.4 Análisis por HPLC-DAD……………………………………………………….
6.2.5 Cuantificación de cetocarotenoides……………………………………………..
6.3 Fotobiorreactores con fotoperiodo………………………………………………..
6.3.1 Cuantificación de clorofilas y carotenoides totales……………………………..
6.3.3 Contenido específico de carotenoides totales…………………………………...
6.3.4 Análisis por HPLC-DAD……………………………………………………….
6.3.5 Cuantificación de cetocarotenoides……………………………………………..
6.4 LC y FP después de la aplicación de estrés……………………………………….
7. CONCLUSIONES………………………………………………………………….
8. RECOMENDACIONES PARA TRABAJOS FUTUROS…………………..……..
9. REFERENCIAS…………………..………………………………………………...
9. ANEXOS………………………..……………………………………………….….
Anexo I. Composición de algunas soluciones nutricionales para el cultivo en
laboratorio del género Scenedesmus …………………………………………………
Anexo II. Método del lavado celular …………………………………………………
Anexo III. Metodología para la cuantificación de pigmentos con metanol caliente…..
Anexo IV. Metodología para la cuantificación de la proporción de xantofilas
por Cromatografía de Alta Resolución (HPLC)……………………………………..
Anexo V. Metodología para la cuantificación de cetocarotenoides con
Dimetilsulfóxido (DMSO) ………………………………………………………….
Anexo VI. Fotografía de los fotobiorreactores………………………………………..
Anexo VII. Cromatogramas de los fotobiorreactores………………………………..
31
31
33
33
36
37
38
40
40
42
43
44
45
48
49
50
54
54
57
58
59
61
62
64

Índice de tablas y figurasÍndice de tablas y figurasÍndice de tablas y figurasÍndice de tablas y figuras
Tabla 1. Actividades biológicas de los carotenoides en el ser humano 5
Tabla 2. Carotenoides de mayor importancia industrial 8
Tabla 3. Características generales de las microalgas 9
Tabla 4. Características generales del subgénero Scenedesmus 14
Tabla 5. Contenido específico máximo de carotenoides totales en los reactores con luz
continua 37
Tabla 6. Contenido específico máximo de carotenoides totales en los reactores con
fotoperiodo 43
Figura 1. Ejemplo de carotenoides: el β-caroteno 1
Figura 2. Carotenos y xantofilas presentes en distintos organismos 2
Figura 3. Estructura del retinol 4
Figura 4. Carotenoides presentes en microalgas 10
Figura 5. Esquema del transporte de electrones y el ciclo de las xantofilas 12
Figura 6. Especie del subtipo Desmodesmus 14
Figura 7. Especie del subtipo Scenedesmus 14
Figura 8. Ruta del mevalonato a prenil fosfatos 15
Figura 9. Ruta de 1-deoxixilulosa-5-fosfato a prenil pirofosfatos 16
Figura 10. Síntesis de geranilgeranil pirofosfato (GGPP) 16
Figura 11. Biosíntesis de carotenoides en microalgas 17
Figura 12. Biorreactor airlift 26
Figura 13. Diagrama de bloques del desarrollo general de cada experimento 27
Figura 14. Cinética de crecimiento por PS y recuento celular directo en
fotobiorreactores con FP y LC 31
Figura 15. Cuantificación de pigmentos con FP y LC durante crecimiento 32
Figura 16. Cinética de crecimiento en experimentos con luz continua 33
Figura 17. Recuento celular en fotobiorreactores con luz continua 34
Figura 18. Cuantificación de clorofilas totales para fotobiorreactores con luz continua 35

Figura 19. Cuantificación de carotenoides totales para fotobiorreactores con
luz continua 36
Figura 20. Porcentajes relativos de carotenoides identificados
por HPLC-DAD (LC) 37
Figura 21. Cuantificación de cetocarotenoides en fotobiorreactores con luz continua 39
Figura 22. Cinética de crecimiento de fotobiorreactores con fotoperiodo 40
Figura 23. Recuento celular directo en fotobiorreactores con fotoperiodo 41
Figura 24. Cuantificación de clorofilas totales para fotobiorreactores con fotoperiodo 41
Figura 25. Cuantificación de carotenoides totales para fotobiorreactores
con fotoperiodo 42
Figura 26. Porcentajes relativos de carotenoides identificados
por HPLC-DAD (FP)
Figura 27. Cuantificación de cetocarotenoides en experimentos con fotoperiodo 44
Figura 28. Disminución del porcentaje de cetocarotenoides en el fotobiorreactor con FP
después de la aplicación de estrés nutricional y lumínico 45

1
1 INTRODUCCIÓN
1.1 Los carotenoides
1.1.1 Descripción
Los carotenoides son pigmentos terpenoides liposolubles, compuestos por 8 unidades de de
isopreno. Cuentan con una estructura básica lineal con dobles enlaces conjugados que
representa el grupo cromóforo, el cual absorbe la energía luminosa en la región visible del
espectro y le confiere a la molécula su coloración distintiva. La longitud de onda máxima a
la que absorbe cada carotenoide depende, entre otros factores, del número de dobles enlaces
conjugados y de la existencia de anillos cerrados en la molécula (Zechmeister, 1944, 1962),
de tal manera que se requieren 7 dobles enlaces conjugados para que un carotenoide
muestre un color perceptible (Meléndez-Martínez et al., 2007). Los carotenoides absorben
luz en la región azul y verde del espectro, y reflejan las longitudes de onda de las regiones
del amarillo, el naranja y el rojo.
Todos los carotenoides se derivan de la estructura C40H56, la cual al sufrir reacciones de:
hidrogenación, dehidrogenación, ciclización y oxidación, da lugar a diferentes compuestos
de este tipo. Como ejemplo se puede mencionar al β caroteno, (Fig. 1) el cual cuenta con
dos anillos en los extremos de su molécula.
Figura 1. Ejemplo de carotenoides: el β-caroteno (Eonseon, 2003)
Se han aislado alrededor de 600 carotenoides a partir de fuentes naturales. Los carotenoides
son sintetizados por una gran variedad de organismos, incluyendo las plantas, los hongos,
las bacterias y las algas (Fig. 2). Se estima que en la naturaleza se producen anualmente
más de 100.000.000 de toneladas de carotenoides (Meléndez-Martínez et al., 2004). La
mayor parte de esta cantidad se encuentra en forma de fucoxantina (en diversas algas) y en
los tres principales carotenoides de las hojas verdes: luteína, violaxantina y neoxantina.

2
1.1.2 Clasificación
De acuerdo a su composición química, los carotenoides se dividen en dos tipos:
Carotenos o compuestos hidrocarbonados
Xantofilas u oxicarotenos, que presentan oxígeno generalmente en los anillos
terminales de su estructura
Otro criterio de clasificación implica la forma en la que los carotenoides son sintetizados en
los organismos, y de acuerdo con esto, los carotenoides primarios son aquellos sintetizados
bajo condiciones normales y que favorecen el crecimiento, mientras que los carotenoides
secundarios se producen bajo condiciones especiales de estrés, principalmente estrés
nutricional, tal como la deficiencia de nitrógeno (Britton, 1988).
Figura 2. Carotenos y xantofilas presentes en distintos organismos (Umeno, 2005)

3
1.1.3 Características químicas
Debido a su naturaleza, los carotenoides son insolubles en agua y solubles en disolventes
no polares. Su grado de solubilidad dependerá de los grupos funcionales sustituyentes de la
molécula. Los carotenos son muy solubles en éter de petróleo y hexano, mientras que las
xantofilas se disuelven mejor en metanol o etanol. La solubilidad de los carotenoides es una
característica muy importante, puesto que influye directamente en la extracción y
purificación en los procesos industriales.
Otras propiedades de los carotenoides son: sensibilidad a la luz, el oxígeno, compuestos
ácidos y a temperaturas elevadas. Su mayor degradación se produce por reacciones de
oxidación. Dichas propiedades se reflejan en otras características como la atenuación del
nivel energético o “quenching” de los singuletes de oxígeno y el bloqueo de las reacciones
mediadas por radicales libres, también llamado “scavenging” o “trapping”.
Los carotenoides, en general, son más estables en sistemas con alto grado de insaturación
como los fotosistemas, ya que este tipo de sistemas acepta más fácilmente oxígeno y
radicales libres antes que el carotenoide. Por el contrario, en sistemas con lípidos saturados,
los carotenoides presentan mayor inestabilidad. En consecuencia estos compuestos pueden
actuar como prooxidantes o antioxidantes dependiendo del sistema donde se encuentren.
(Olmedilla et al., 2001)
Por lo anterior, se recomienda trabajar con los carotenoides en una atmósfera inerte
(nitrógeno), a baja temperatura (20ºC), en oscuridad o luz difusa, en condiciones libres de
ácido y con solventes libres de peróxido. (Pfander, 1992).
1.1.4 Importancia en la salud humana
Algunos carotenoides, además de otorgar un color atractivo a los distintos organismos
fotosintéticos, tienen la propiedad de presentar actividad como provitamina A, lo que
incrementa su importancia a nivel fisiológico y nutricional.

4
La vitamina A resulta esencial para la visión nocturna, y necesaria para mantener sanos la
piel y los tejidos superficiales. Puede suministrarse como tal, llamándose entonces retinol
(Fig. 3), como algunos análogos menos activos o como sus precursores, los carotenoides.
Figura 3. Estructura del retinol (Olmedilla et al., 2001)
No todos los carotenoides son precursores de la vitamina A, por lo que se encuentran
carotenoides tanto provitamínicos como no provitamínicos. El número de carotenoides
precursores de vitamina A oscila entre 50 y 60, destacando los carotenos (α, β - y γ-
caroteno) y algunas xantofilas (β-criptoxantina). La capacidad de los carotenos para actuar
como provitamina A depende de su conversión a retinol por los animales, así como de la
presencia de β-ionona. Los carotenos que contienen como mínimo un anillo de β-ionona
pueden convertirse en retinol. Desde este punto de vista, el carotenoide más importante es
el β-caroteno, que contiene dos anillos de β-ionona.
Actualmente, el término provitamina A se usa para todos los carotenoides que presentan
cualitativamente actividad de β-caroteno. En cuanto a éste, su actividad dependerá a su vez
de la cantidad en la que esté presente en los alimentos. La FAO/WHO (1991) establece que
la cantidad de β-caroteno equivalente a 1 µg de retinol es de 4 mg, 6 mg o 10 mg,
dependiendo de la cantidad de β-caroteno presente en la comida.
Además de su importante papel en la visión, la vitamina A es reconocida como un factor de
gran importancia en la salud infantil y la supervivencia (WHO, 1998). (Tabla 2)

5
Tabla 1. Actividades biológicas de los carotenoides en el ser humano
(Olmedilla et al., 2001)
La actividad antioxidante que tienen los pigmentos relacionados con el presente trabajo
depende de una serie de factores, como su estructura química (tamaño, número de
sustituyentes, configuración cis o trans, etc.), su concentración, la presión parcial de
oxígeno o su interacción con otros antioxidantes, como las vitaminas C y E. El mecanismo
de la actividad antioxidante del β-caroteno, está relacionado con su carácter hidrofóbico y
con su capacidad para "retirar" el oxígeno singulete y desactivar radicales libres. También
se ha demostrado que otros carotenoides, como la astaxantina, luteína, zeaxantina,
cantaxantina y el licopeno funcionan como antioxidantes. Es importante mencionar que se
ha demostrado la actividad antioxidante de estos compuestos pero también se han obtenido
resultados que demuestran lo contrario. Así, por ejemplo, algunos ensayos indican que la
actividad antioxidante de la astaxantina es superior a la de otros carotenoides, mientras que
en otros estudios se llega a la conclusión inversa (Meléndez-Martínez et al., 2004).
Se ha evaluado el papel protector de diversos antioxidantes como el β-caroteno, α-tocoferol
y el ácido ascórbico para las células humanas frente a la radiación ultravioleta, llegándose a
la conclusión de que el primero es el más eficiente, probablemente debido a su localización
en la membrana celular. La luteína y la zeaxantina (referidos como pigmento macular —
Acciones (respuestas, benéficas o adversas; fisiológicas o farmacológicas ante la
administración de estos compuestos; no se considera esencial)
-Antioxidantes -Inmunopotenciadores -Inhibición de mutagénesis y transformación -Inhibición de lesiones premalignas -Protección frente a la fotosensibilización Asociaciones (correlaciones entre los carotenoides y algún aspecto o finalidad
fisiológica o médica que puede o no mostrar una relación causal)
-Cataratas -Degeneración macular -Diversos tipos de cáncer -Enfermedades cardiovasculares

6
PM—), pueden prevenir el daño oxidativo inducido por la luz en la retina, al atenuar la luz
azul que entra en ella, y por tanto proteger frente al deterioro asociado a la edad, causante
de la degeneración macular senil.
Hay estudios que relacionan la aparición de algunos tipos de cáncer con la carencia de
ciertos carotenoides en la dieta, por lo que estos compuestos son considerados
anticancerígenos. Diversas investigaciones epidemiológicas han demostrado que el riesgo
de padecer cáncer es inversamente proporcional al consumo de vegetales y frutas ricos en
carotenoides. No obstante que muchos de estos estudios se han centrado en el β-caroteno,
existen otros carotenoides eficaces en la prevención de esta enfermedad, como son la β-
criptoxantina, zeaxantina, astaxantina e incluso el carotenoide no coloreado fitoeno. En un
estudio reciente, se propuso una relación inversa entre el consumo de alimentos ricos en
luteína (como espinaca o lechuga) y el cáncer de colon, tanto en hombres como en mujeres.
De igual forma, se ha demostrado que los carotenoides típicos del pimiento rojo (Capsicum
annuum L.) como la capsantina y sus ésteres, y la capsorrubina, entre otros, son agentes
antitumorales efectivos. La protección de los pigmentos carotenoides frente al cáncer y a
otras enfermedades crónicas, podría atribuirse además de a sus propiedades antioxidantes, a
efectos como la inhibición de la proliferación celular, a la mejora de la diferenciación
celular, a la estimulación de la comunicación intercelular y a la filtración de la luz azul,
entre otros.
Desde hace tiempo se ha postulado que los carotenoides actúan como potenciadores
positivos de la respuesta inmune, y se ha encontrado que elevadas dosis de β -caroteno
aumentan la relación entre los linfocitos CD4 y CD8, que suele ser muy baja en pacientes
seropositivos.
Se ha estimado que el consumo promedio de vitamina A oscila entre los 744 y 811
equivalentes de retinol por día en los hombres y los 530 y 716 equivalentes de retinol por
día en las mujeres. Tomando como base los equivalentes de retinol, se estima que
aproximadamente un 26% y un 34% de la vitamina A consumida por hombres y mujeres,
respectivamente, es proporcionada por los carotenoides provitamínicos.

7
1.1.5 Importancia industrial
La importancia de los carotenoides en la salud humana, repercute en la generación
industrial de productos como vitaminas y complementos. Sin embargo, los carotenoides
tienen otras aplicaciones en este ámbito (Tabla 3), siendo una de las más importantes su
función como pigmento en la industria alimentaria, puesto que se considera que hay
seguridad en su consumo.
En la avicultura, la luteína resulta un complemento de gran importancia, pues la coloración
de la carne de aves es uno de los parámetros de calidad y también un atractivo. El consumo
mundial de luteína es de aproximadamente 350 000 kg anuales. México consume alrededor
de 180 000 kg, Estados Unidos 70 000 kg y Europa alrededor de 100 000 kg anuales (Agro
2000, 2001), de tal forma que la industria avícola mexicana consume más del 50 por ciento
de los pigmentos naturales a base de luteína.
En la acuacultura, la astaxantina principalmente, y la cantaxantina en segundo lugar, son
utilizadas como complementos alimenticios para animales como salmones y crustáceos, los
cuales no son capaces de sintetizar estos pigmentos y los absorben normalmente de su
medio marino, donde son sintetizados por el fitoplancton. Cuando estos organismos son
criados por el hombre en estanques aislados, es necesario suministrarles dicho pigmento en
la dieta. La ventaja de producir astaxantina a partir de microalgas o de cualquier otro
organismo vivo, es que no es necesaria su purificación posterior, ya que es asimilable tal
cual por el pez o crustáceo en cultivo. La ausencia de astaxantina como suplemento, daría
como resultado carne pálida y poco atractiva a la vista. El precio elevado de los pigmentos
hace que sea uno de los insumos que más incide en los costos de producción de la industria
salmonera, representando entre un 18% a 22% (Pokniak y Bravo, 2000).
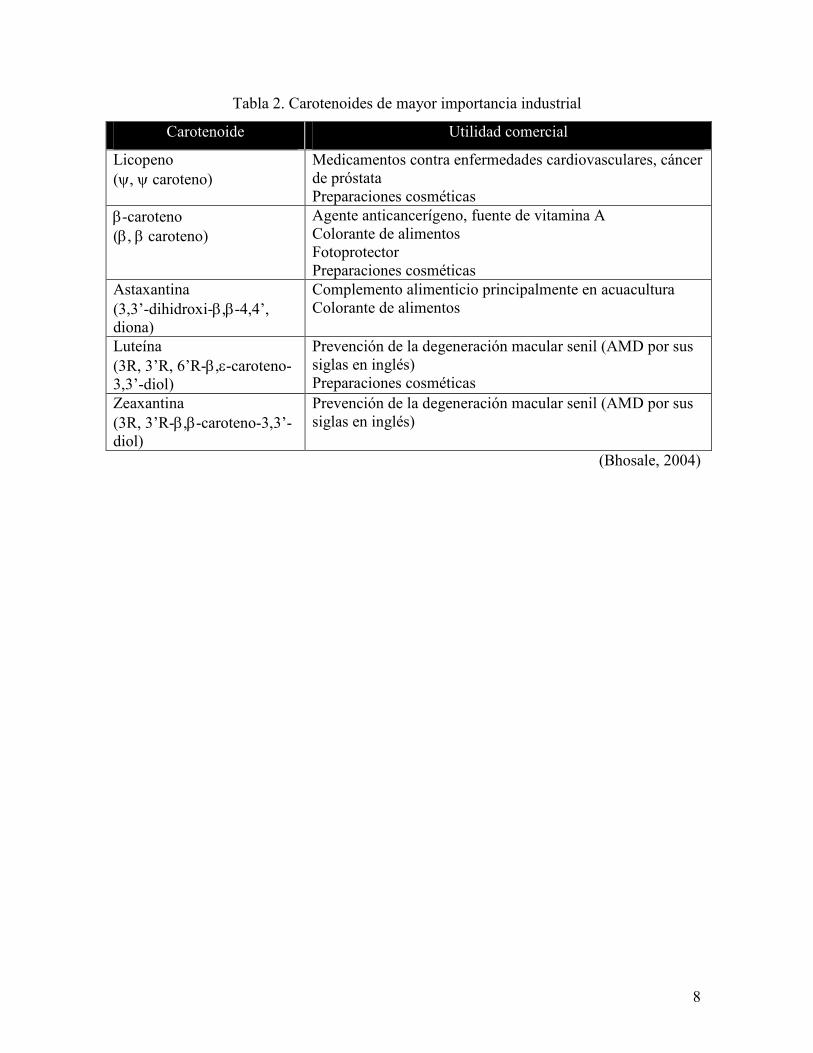
8
Tabla 2. Carotenoides de mayor importancia industrial
Carotenoide Utilidad comercial
Licopeno (ψ, ψ caroteno)
Medicamentos contra enfermedades cardiovasculares, cáncer de próstata Preparaciones cosméticas
β-caroteno (β, β caroteno)
Agente anticancerígeno, fuente de vitamina A Colorante de alimentos Fotoprotector Preparaciones cosméticas
Astaxantina (3,3’-dihidroxi-β,β-4,4’, diona)
Complemento alimenticio principalmente en acuacultura Colorante de alimentos
Luteína (3R, 3’R, 6’R-β,ε-caroteno-3,3’-diol)
Prevención de la degeneración macular senil (AMD por sus siglas en inglés) Preparaciones cosméticas
Zeaxantina (3R, 3’R-β,β-caroteno-3,3’-diol)
Prevención de la degeneración macular senil (AMD por sus siglas en inglés)
(Bhosale, 2004)

9
1.2 Las microalgas y los carotenoides
1.2.1 Descripción general
Las microalgas son organismos eucariotes microscópicos que contienen clorofila y que
llevan a cabo fotosíntesis oxigénica. En la tabla 4 se presentan sus características generales.
Tabla 3. Características generales de las microalgas
Característica Descripción
Tipo de organización celular
Unicelular, colonial o filamentosa
Motilidad Pueden tener o no motilidad, durante toda su existencia o solamente en alguna etapa de su crecimiento
Reproducción Sexual o asexual Nutrición Auxótrofos, mixótrofos, heterótrofos facultativos u obligados Dependencia de la luz
Existe una gran cantidad de especies fotótrofas obligadas, es decir, que no pueden crecer en ausencia de luz
Pigmentación Distintos colores. A pesar de que todas las microalgas tienen clorofila se encuentran algas de color rojo o marrón, atribuyéndose este color a la presencia de pigmentos que enmascaran el color verde de la clorofila
Ambiente Generalmente ambientes acuáticos como los lagos, los estanques y el mar, sin embargo, también se les puede encontrar viviendo sobre todo tipo de tierra. La mayoría vive libremente, mientras que algunas viven en relación simbiótica con otros organismos
Las algas y especialmente las microalgas sintetizan una gran variedad de carotenoides
(Fig. 4), incluyendo aquellos sintetizados por las plantas, tales como las xantofilas
violaxantina, neoxantina y luteína. También sintetizan exclusivamente carotenoides como
diatoxantina, diadinoxantina y fucoxantina.

10
Figura 4. Carotenoides presentes en microalgas (Eonseon, 2003)
1.2.2 Función fisiológica
Los carotenoides tienen dos principales funciones en las microalgas: la primera es como
pigmento accesorio en la fotosíntesis y la segunda es la fotoprotección.
Como pigmentos accesorios, es importante recalcar que si bien las clorofilas son los
pigmentos que reciben la mayor cantidad de energía, la luz que incide en el
microorganismo está compuesta por varias longitudes de onda, y la presencia de pigmentos

11
con distintas propiedades de absorción, como los carotenoides, asegura que un mayor
porcentaje de los fotones incidentes estimule la fotosíntesis.
Por otro lado, una de las funciones más importantes de los pigmentos carotenoides, es
extraer el exceso de energía de las moléculas excitadas de clorofila y disiparla como calor.
De no ser absorbido este exceso de energía, la clorofila basal se activaría y transferiría esta
energía al oxígeno. Entonces se produciría una forma ultrarreactiva de oxígeno sencillo
(1O*) ó singulete, la cual puede destruir moléculas biológicas y causar la muerte celular.
Los carotenoides disipan el radical peróxido y también la clorofila excitada adquiriendo el
estado triplete, que a su vez se disipa desprendiendo calor al medio.
El ciclo de las xantofilas: fotoprotección
En casi todos los eucariotes fotosintéticos, la mayoría de las xantofilas están ligadas junto
con las clorofilas a los Complejos de Captura de Luz (LHC por sus siglas en inglés), los
cuales absorben y transfieren energía de excitación a los centros de reacción. Las xantofilas
no participan como parte funcional del complejo, sino como elemento estructural, teniendo
una labor importante en la fotoprotección (Fig. 5).
En la fotosíntesis el transporte de electrones por los portadores de la membrana tilacoidal
como el NADP, implica el bombeo de protones desde el estroma cloroplástico al lumen
tilacoidal. La acumulación de los protones en el lumen tilacoidal, tiene como consecuencia
una baja en el pH, causando un disparo en la activación de la enzima violaxantina de-
epoxidasa. Esta enzima, localizada en la cara interna de los sacos tilacoidales, transforma
el diepóxido violaxantina (V) en zeaxantina (Z) deepoxidada a través del intermediario
monoepoxidado anteraxantina (A). La zeaxantina es capaz de recibir la energía
directamente de la clorofila excitada, disipándola posteriormente en forma de calor. El
proceso se invierte cuando la luz desaparece, disminuye su intensidad progresivamente o
cuando los protones pasan a través de la ATPasa. La conversión de zeaxantina a
violaxantina es realizada por la enzima zeaxantina epoxidasa, resultando en una
acumulación de zeaxantina.

12
Figura 5. Esquema del transporte de electrones y el ciclo de las xantofilas (Manrique,
2003)
Es importante mencionar que el ciclo de las xantofilas es un proceso estimulado no sólo por
la existencia y/o intensidad de la luz, sino también a otros factores ambientales como la
temperatura extrema y la deficiencia de nutrientes (Demmig-Adams et al. 1996).
1.2.3 Género Scenedesmus
La microalga utilizada en el presente trabajo fue Scenedesmus incrassatulus, perteneciente
al subgénero Scenedesmus que a su vez es parte del género Chlorophyta.
Las Chlorophyta o algas verdes, son abundantes en la naturaleza; incluyen organismos
macro y microscópicos de agua dulce. Poseen clorofila a y b, y distintos carotenoides.
Tienen almidón como producto de reserva, el cual, a diferencia de otras algas, se forma en
el cloroplasto. Su pared celular es celulósica aunque algunas especies carecen de ésta. Los
cloroplastos pueden contener pirenoides.
La explotación comercial de las microalgas verdes comprende principalmente los géneros
Chlorella, Dunaliella y Haematococcus. Dunaliella spp., habitante de ambientes
hipersalinos, ha sido estudiada extensivamente debido a su producción de β -caroteno.

13
El subgénero Scenedesmus se caracteriza por formar cenobios o conformaciones de 2, 4, 8
ó hasta 16 células. Incluye más de 200 especies, las cuales comparten las características
señaladas en la tabla 4. El subgénero se subdivide a su vez en:
Figura 7. División de Scenedesmus
Las especies del subtipo Scenedesmus (Fig. 7) tienen paredes celulares en arreglo de
multicapas. La capa interna contiene celulosa embebida en una matriz de hemicelulosa,
mientras que la capa externa (con tres subcapas) consiste en el politerpeno esporopolenina.
Por otra parte, el subtipo Desmodesmus (Fig. 6) se caracteriza por una pared celular más
complicada, con una capa adicional de esporopolenina que rodea a todo el cenobio.
Scenedesmus
Desmodesmus
Scenedesmus
Acutodesmus
Scenedesmus

14
Tabla 4. Características generales del subgénero Scenedesmus
(Kessler, 1982)
Figura 6. Especie del subtipo Desmodesmus
Figura 7. Especie del subtipo Scenedesmus
- Presencia de hidrogenasa, mostrando que al ser iluminadas, son capaces de liberar H2
a bajas presiones parciales de O2 y de H2, y de reducir CO2 con H2, H2S o algún otro
donador orgánico de hidrógeno
- Formación de carotenoides secundarios, tales como astaxantina y cantaxantina, cuando
crecen en deficiencia de nitrógeno, fósforo o de fierro
- Liberación de proteasas extracelulares, revelado por la licuefacción de gelatina
- Liberación de amilasas extracelulares, indicado por la hidrólisis de almidón
- Como organismos de agua dulce, muestran una baja tolerancia a la sal
- El cambio de oscuridad a luz provoca la sincronización celular (Simmer y Sodomkova,
1967)
- Baja tolerancia a los ácidos
- La propagación de Scenedesmus suele ser por autocenobios, aunque también se ha
observado la producción de gametos biflagelados en S. obliquus. (Trainor y Burg, 1965)

15
1.2.4 Biosíntesis de carotenoides
Originalmente, se creyó que la ruta de biosíntesis de carotenoides de las microalgas era la
del acetato/mevalonato, sin embargo, últimamente se ha establecido la ruta del
piruvato/gliceraldehido 3-fosfato para algas verdes (Schwender et al., 1996). En la figura
2, se muestran las dos rutas:
Ruta del mevalonato a prenil fosfatos
El mevalonato se forma a a partir del acetil-CoA y conduce a la formación de isopentenil
pirofosfato (IPP), que es la unidad formadora de terpenoides.
Figura 8. Ruta del mevalonato a prenil fosfatos (Sandmann, 2000)
Ruta de 1-deoxixilulosa-5-fosfatro a prenil pirofosfatos
También llamada ruta de 2-C-metil-D-eritritol-4-fosfato (Fig. 9) por ser éste el primer
producto que se convierte a prenil pirofosfato. No todos los pasos de esta ruta han sido
identificados.
Partiendo de piruvato y 3-P gliceraldehido se forma una unidad de dos carbonos, y
subsecuentes reacciones conducen al 2-C-metil-D-eritritol 2,4-ciclodifosfato. Se sospecha
que la síntesis de este producto es el punto en el que se bifurca la ruta, ya sea para formar
IPP o dimetilalil pirofosfato (DMAPP).
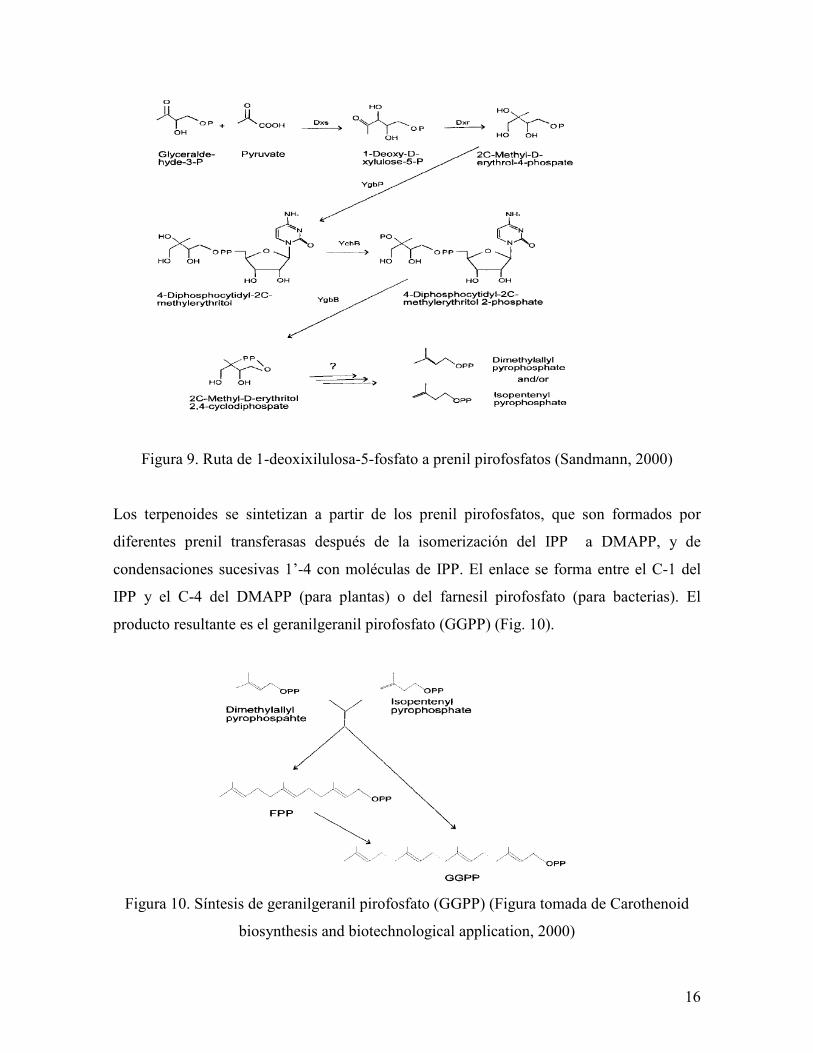
16
Figura 9. Ruta de 1-deoxixilulosa-5-fosfato a prenil pirofosfatos (Sandmann, 2000)
Los terpenoides se sintetizan a partir de los prenil pirofosfatos, que son formados por
diferentes prenil transferasas después de la isomerización del IPP a DMAPP, y de
condensaciones sucesivas 1’-4 con moléculas de IPP. El enlace se forma entre el C-1 del
IPP y el C-4 del DMAPP (para plantas) o del farnesil pirofosfato (para bacterias). El
producto resultante es el geranilgeranil pirofosfato (GGPP) (Fig. 10).
Figura 10. Síntesis de geranilgeranil pirofosfato (GGPP) (Figura tomada de Carothenoid
biosynthesis and biotechnological application, 2000)

17
Para la formación de un caroteno de 40 carbonos se parte del fitoeno. La condensación de
dos moléculas de GGPP con un ciclopropilcarbonil entre el carbono 1 del primer GGPP y
los carbonos 2 y 3 del segundo GGPP dan lugar al prefitoeno pirofosfato. Al eliminarse el
pirofosfato, el 15-cis fitoeno restante es convertido a licopeno.
El licopeno sufre distintas reacciones de desaturación, hidroxilación, epoxidación,
glicosilación, o-metilación, transferencia de grupos acilo y fenilo, ciclidación para formar
los distintos carotenoides existentes en la naturaleza. La figura 5 muestra algunas de estas
reacciones y sus productos.
Las enzimas implicadas en la biosíntesis de carotenoides están asociadas a membranas o se
encuentran integradas en ellas.
La biosíntesis de carotenoides en microalgas se lleva a cabo como se describió
anteriormente, hasta llegar al licopeno. Posteriormente, las células siguen la ruta descrita en
la figura 11.
Figura 11. Biosíntesis de carotenoides en microalgas (Jin et al., 2003)

18
1.2.5 Factores que afectan la biosíntesis de carotenoides en microalgas
La eficiencia de la biosíntesis de carotenoides implica principalmente dos cuestiones: las
condiciones de cultivo, y el nivel y actividad de las enzimas biosintéticas de carotenoides.
En ausencia de herramientas de biología molecular que mejoren la actividad de las enzimas
implicadas, las condiciones de cultivo son la variable a manipular en la mayoría de los
casos.
A continuación se enumeran los factores más importantes de los cultivos de microalgas que
tienen como objetivo la sobreproducción de carotenoides:
*Luz
En las microalgas, la producción y la acumulación de carotenoides se ve afectada
positivamente por la irradiación con luz blanca, sin embargo, la intensidad, el horario y el
tiempo en que ésta es administrada, varía con el microorganismo. La teoría de la
fotoinducción abarca dos aspectos importantes. El primero es que el mejoramiento de la
producción volumétrica (mg ml-1), está asociado al mejoramiento del crecimiento. El
segundo aspecto es que la acumulación celular (mg g-1) de los carotenoides está asociada al
aumento de la actividad de las enzimas implicadas en su ruta biosintética. Por ejemplo, la
microalga Dunaliella sp., hiperproductora de β-caroteno, para crecer y sintetizar
carotenoides, requiere de alta intensidad luminosa, de un estrés causado por sales y de
limitación de nutrientes.
*Temperatura
La temperatura es uno de los principales parámetros a controlar en un cultivo. De hecho, en
los cultivos de Dunaliella y Haematococcus, es el factor a controlar más importante. En un
experimento realizado por Orset y Young (1999), con Dunaliella, los niveles de caroteno
(excepto para el α-caroteno), disminuyeron cuando la temperatura varió de 34 a 17 ºC. Sin
embargo, cuando la radiación luminosa fue de 1,000 µmol m2 s-1 a 19ºC, los niveles de β-
caroteno se incrementaron.

19
*Iones metálicos
El papel biológico de algunos iones metálicos en los microorganismos se desconoce, sin
embargo se han hecho estudios al respecto en el caso de la producción de carotenoides. En
los experimentos de Kobayashi et al. (1992), la producción de astaxantina por H. pluvialis
mejoró cuando el medio se enriqueció con una sal ferrosa. Tjahjono et al. (1994) dedujeron
que esto se debía a la generación del radical hidroxi, el cual estimula la síntesis de
carotenoides por la reacción de Fenton (H2O2 + Fe 2+ → Fe 2+ +HO− +HO*).
*Deficiencia de nitrógeno
El nitrógeno es un constituyente esencial de las proteínas estructurales y funcionales de las
células algales, representando del 7-10% del peso celular seco. En general, las microalgas
tienen una capacidad limitada para producir materiales de reserva de nitrógeno, con
excepción de la cianoficina y ficocianica en cianobacterias, las cuales en condiciones
limitadas de nitrógeno, degradan los ficobilisomas. Entonces, el flujo de carbono es
conducido a la síntesis de lípidos o carbohidratos, en vez de a síntesis de proteínas.
Numerosos estudios indican que la biosíntesis y la acumulación de lípidos en forma de
triacilglicerol se ve beneficiada por la limitación de nitrógeno en el medio. (Thompson,
1996).
La acumulación de carotenoides secundarios, es una característica importante de muchas
algas cuando son cultivadas con deficiencia de nitrógeno; esto es comúnmente acompañado
por una disminución en el contenido celular de clorofila. Ben-Amotz et al. (1982)
demostraron que la producción de β-caroteno se ve aumentada en células de Dunaliella
crecidas con deficiencia de nitrógeno en el medio. De igual forma, Borowitzka et al. (1991)
demostraron que una baja concentración de nitrógeno, es uno de los factores más
importantes para estimular la síntesis y acumulación de astaxantina, Zhekisheva et al.
(2002) reportaron que bajo condiciones limitantes de nitrógeno, Haematococcus pluvialis
produjo 5 picogramos de ácidos grasos por cada picogramo de astaxantina, sugiriendo que
estos dos procesos podrían estar relacionados, de tal manera que los glóbulos de aceite
mantuvieran el alto contenido de los ésteres de astaxantina.

20
1.4 Producción industrial de carotenoides: síntesis química vs. biosíntesis
La producción de carotenoides se realiza por síntesis química o por biosíntesis.
Entre las ventajas de la síntesis química, destaca la pureza de los compuestos obtenidos, su
consistencia, y bajo costo de producción. Por otro lado, también presenta ciertas
desventajas: la complejidad de la síntesis de algunos carotenoides. Si bien se pueden
utilizar procesos similares para producir distintos carotenoides, normalmente la síntesis de
un nuevo carotenoide requiere el desarrollo de toda una nueva ruta química. Otro gran
inconveniente que presenta la síntesis química de carotenoides, es la producción de mezclas
de estereoisómeros, algunos de los cuales no se encuentran en la naturaleza y no son tan
activos como los presentes en ella, por lo que quedan inhabilitados para el consumo por sus
efectos secundarios indeseables.
Con relación a la biosíntesis de carotenoides, ésta presenta varias ventajas. Considerando
que existen más de 600 carotenoides en la naturaleza, se puede encontrar una gran
capacidad biosintética en ella, sin tener que desarrollar procesos complejos de síntesis
química. A diferencia de ésta, al saber la ruta de biosíntesis de un carotenoide se puede
tener conocimiento de la biosíntesis de varios carotenoides. De igual manera, la producción
de estos pigmentos por medio de organismos vivos, evita la formación de estereoisómeros
perjudiciales para la salud. Finalmente, la capacidad biosintética de organismos
sobreproductores puede ser mejorada gracias a la tecnología del DNA recombinante y otras
herramientas de biología molecular.
Sin embargo, resulta prudente también mencionar las desventajas de la biosíntesis de
carotenoides. Los sistemas biológicos suelen producir una mezcla de carotenoides, lo que
hace indispensable aplicar un proceso de purificación y extracción, el cual aumenta los
costos de producción. Otro inconveniente es que estos pigmentos son componentes
intracelulares, y que por lo tanto no son secretados al medio en un proceso de fermentación,
volviendo imprescindible su extracción a partir de la célula y en consecuencia, aumentan
los costos de producción.

21
1.5 Fotobiorreactores
En general, los fotobiorreactores son reactores especiales que, a diferencia de los reactores
comunes, reciben luz de manera que se satisfagan las necesidades de un microorganismo
fotosintético. Asimismo, dependiendo del microorganismo, estos pueden ser sistemas
abiertos o cerrados. Los sistemas abiertos se usan para cepas que crecen a gran velocidad o
a condiciones extremas de pH y salinidad. Para asegurar que se les suministre suficiente
luz, son puestos al aire libre, de manera que reciban la luz solar.
Por otro lado, los sistemas cerrados pueden incluir reactores de columna burbujeada, tipo
airlift, es decir, reactores que garanticen una alta transferencia de masa y bajos tiempos de
circulación, además de una aireación eficiente y bajo esfuerzo de corte.
2. ANTECEDENTES
La producción de carotenoides por microalgas del género Scenedesmus no ha sido
ampliamente estudiada. Si bien es cierto que miembros de este género han sido muy
utilizados como organismos modelo en el campo de la limnología, tecnología y manejo del
agua, alimentación de zooplankton y efecto de sustancias químicas en su crecimiento
(Wiltshire, 2000), la producción de carotenoides como tal, a la fecha ha sido el objetivo de
pocos experimentos con este género. El ejemplo más reciente y representativo de dichos
experimentos es el artículo publicado en la revista Process Biochemistry, “Acumulación y
metabolismo de astaxantina en Scenedesmus obliquus (Chlorophyceae)” de Qin et al.,
2008, en el que se describe el cultivo de S. obliquus en un sistema en dos etapas. En la
primera etapa, la microalga se cultivó bajo condiciones de inducción (incubación a 30ºC e
iluminación de 180 µmol m2 s-1) por 48 horas. Se analizó la composición de los
carotenoides obtenidos y se identificaron 7 cetocarotenoides. En la segunda etapa, realizada
bajo condiciones normales (25ºC e iluminación de 80 µmol m2 s-1) por 72 horas, la
concentración de carotenoides disminuyó del 59.84% al 6.57%. Los resultados sugieren que
la biosíntesis de astaxantina y los otros carotenoides puede ser controlada con las
condiciones de cultivo.

22
En el Laboratorio de Biotecnología de microalgas del Departamento de Biotecnología y
Bioingeniería del CINVESTAV previo al presente estudio, se realizó un proyecto
relacionado con los carotenoides producidos en especies del género Scenedesmus:
“Carotenogénesis en Scenedesmus acutus” (Castelblanco, 2007), mientras que en la
Facultad de Estudios Superiores de Iztacala se llevó a cabo el proyecto: “Caracterización
fisiológica de Scenedesmus incrassatulus durante la síntesis de pigmentos” (Vega, 2006).
En el trabajo “Carotenogénesis en Scenedesmus acutus” se realizó un seguimiento de la
producción de carotenoides y en su correlación con el consumo de nutrientes y
metabolismo. Se cultivó la microalga en un fotobiorreactor tipo columna burbujeada con un
volumen de trabajo de 2 L, a 25 ºC, 200µmoles m-2 s-1. Se observó que S. acutus creció en
forma exponencial durante los primeros siete días, con una µmax= 0.49 d-1 y alcanzando una
concentración máxima de biomasa de 2.0 g L-1 a los 39 días de cultivo. Al día 11 de
cultivo el consumo de nitratos y fosfatos fue de 97% y 98%, respectivamente. A los 7 días
hubo una disminución del contenido de clorofilas totales y de carotenoides totales, los
cuales fueron identificados por HPLC-DAD como: β-caroteno, luteína, violaxantina y
neoxantina. La luteína fue el carotenoide presente en mayor concentración, con un valor
que disminuyó de 160.8 µg g-1 PS a 40.9µg g-1 PS en el día 29.
En “Caracterización fisiológica de Scenedesmus incrassatulus durante la síntesis de
pigmentos” se describieron los procesos metabólicos de Scenedesmus incrassatulus durante
la síntesis de carotenoides. S. incrassatulus fue cultivado en medio mineral (Perales, 2004)
en frascos de 1 L por 30 días, a 25 ºC, una densidad de flujo fotónico de 250 µmoles m-2 s-1
con periodo de luz/oscuridad de 14/10 hr y un flujo de aire de 0.5 vvm, partiendo de un
inóculo de 1.3* 106 cel mL-1 provenientes de un cultivo sincronizado. En el trabajo se
menciona que la disminución de la actividad metabólica de S. incrassatulus a un nivel basal
de mantenimiento se encuentra estrechamente relacionada con la limitación de nutrientes
como fósforo y nitrógeno en el medio de cultivo, la alta densidad celular, el aumento de la
concentración de carotenoides totales y la disminución en el contenido de clorofilas
almacenadas en el espacio citoplasmático; la carotenogénesis está relacionada con la
inhibición del metabolismo respiratorio y fotosintético. Asimismo, se determinó que en las

23
condiciones experimentales utilizadas, la microalga en cuestión sintetizó isozeaxantina,
zeaxantina, astaxantina y cantaxantina como pigmentos secundarios. Se observó que
durante la carotenogénesis, S. incrassatulus experimentó cambios morforlógicos en su
ultraestructura como aumento en el tamaño celular, acumulación de materiales
electrodensos en la pared celular y aparición y desaparición de estructuras celulares.
El reactor utilizado en el presente trabajo fue caracterizado por Vega-Estrada et al.
(2005).con un cultivo de Haematococcus pluvialis. El biorreactor cuenta con un kLa entre
10 y 32 h-1.
3. JUSTIFICACIÓN
Los carotenoides se destacan por cumplir numerosas funciones en el ser humano. La
principal de ellas, su actividad provitamínica A, junto con su desempeño como
antioxidantes y su consecuente prevención del cáncer, como inmunopotenciadores en caso
de enfermedades como el VIH y como compuestos que ayudan a prevenir la degeneración
macular senil, hace de estos pigmentos unos de los compuestos más importantes para
mejorar la salud.
Asimismo, la importancia comercial de los carotenoides en áreas como la acuacultura y la
avicultura, la industria de los cosméticos, etc., hace necesaria su obtención en forma
natural, cuestión que orienta a la biosíntesis, pues la síntesis química en varias ocasiones
está acompañada de estereoisómeros no naturales que perjudican la salud. La biosíntesis
comercial de carotenoides está dominada por especies como Dunaliella salina (β-caroteno),
Haematococcus pluvialis (astaxantina), Phaffia rhodozyma (astaxantina) y Tagetes erecta
(luteína).
En los casos anteriores, cada organismo presenta cualidades y desventajas. Por ejemplo, en
el caso de H. pluvialis, su alta productividad (4% del peso celular seco) resulta atractiva, sin
embargo, la extracción de este pigmento resulta complicada, debido a su acumulación en
quistes. Este problema no se presenta en la levadura P. rhodozyma, sin embargo, su baja

24
productividad (0.3% por peso seco) provoca que el proceso sea incosteable para muchos
productores.
Por otra parte, la luteína es únicamente extraída con fines comerciales de la flor mexicana
Cempasúchil o flor de muerto, pero al igual que en H. pluvialis, la dificultad de su
extracción vuelve la producción de luteína un proceso costoso.
Los usos de los carotenoides son tan diversos que se espera que su mercado global sea de
919 mdd para el año 2015. En el año 2007, el mercado de los carotenoides fue de 766 mdd,
considerándosele un mercado con crecimiento anual del 2.3% para los siguientes 8 años.
El β-caroteno es el pigmento que ocupa el mayor porcentaje del mercado, alcanzando
ventas de 247 mdd por su consumo como complemento alimenticio. La astaxantina ocupa
el segundo lugar en el mercado con 219 mdd en el mismo año, por sus usos en la
acuacultura.
Por las consideraciones anteriormente expuestas, es imprescindible buscar fuentes naturales
de carotenoides con alta productividad y facilidad de obtención, que satisfagan las
necesidades humanas, tanto de salud como comerciales. En el presente trabajo se investigó
la producción de carotenoides por la microalga Scenedesmus incrassatulus, pues no se ha
caracterizado su producción, aunque se sabe de su capacidad para producir carotenoides
bajo condiciones de estrés, lo cual la convierte en una productora potencial de dichos
pigmentos.
HIPÓTESIS
La aplicación de estrés nutricional y lumínico inducirá la carotenogénesis en la microalga
Scenedesmus incrassatulus.

25
4. OBJETIVOS
Objetivo general
Caracterizar la producción de carotenoides por la microalga Scenedesmus incrassatulus en
cultivo por lote utilizando un reactor tipo airlift
Objetivos específicos
• Inducir la producción de carotenoides en la microalga Scenedesmus incrassatulus
por eliminación de nutrientes del medio de cultivo y aplicación de estrés lumínico
en fotobiorreactores con fotoperiodo y con luz continua.
• Extraer, identificar y cuantificar los carotenoides presentes en la microalga
Scenedesmus incrassatulus después de inducir la carotenogénesis

26
5. MATERIALES Y MÉTODOS
5.1 Scenedesmus incrassatulus
La cepa que se usó fue S. incrassatulus CLHE-Si01, donada por el Laboratorio de
Hidrobiología experimental de la Escuela Nacional de Ciencias Biológicas, del Instituto
Politécnico Nacional.
5.2 Medio de cultivo mineral libre de EDTA (Anexo I)
El medio de cultivo empleado en el presente trabajo ha sido usado en trabajos previos para
el crecimiento de diversas cepas del género Scenedesmus (Perales, 2007). El hecho de que
este medio sea libre de EDTA evita la quelación de iones importantes como el Fe, el cual
no puede ser asimilado con la misma facilidad cuando está quelado que cuando se
encuentra libre en el medio de cultivo (Soeger y Hegewald, 1988).
5.3 Biorreactor airlift
Se utilizó un biorreactor tipo airlift, con una capacidad nominal de 2 litros, caracterizado
anteriormente por Vega-Estrada et al. (2005). El diagrama del reactor se encuentra en la
figura 12:
A Tapón de hule: filtro de aire (1), salida de aire (2), electrodo de pH o sensor de O2 (3),
alimentación de medio (4), tubo para muestreo (5), aguja (6), lámparas (7)
B Cilindro
C Placa
Figura 12. Biorreactor airlift (Vega-Estrada et al., 2005). Las dimensiones se muestran en cm

27
5.4 Protocolo experimental
Se observó la inducción de la carotenogénesis bajo 2 diferentes periodos de luz, fotoperiodo
12/12 hr y luz continua, aplicando estrés lumínico o nutricional o la combinación de ambos
de tal forma que la investigación comprendió los siguientes experimentos:
Con luz continua:
-Aplicación de estrés nutricional
-Aplicación de estrés nutricional y lumínico
-Sin aplicación de estrés
Con fotoperiodo:
-Aplicación de estrés nutricional
Aplicación de estrés lumínico
-Aplicación de estrés nutricional y lumínico
-Sin aplicación de estrés
5.4. 1 Metodología
Cada experimento comprendió tres fases: establecimiento del cultivo de Scenedesmus
incrassatulus, cinéticas de crecimiento de la microalga y fase de inducción de la
carotenogénesis o aplicación de estrés.
2. Inoculación a partir de matraz semilla
1. Sincronización del cultivo
3. Crecimiento
Determinaciones
analíticas
4. Aplicación de estrés
(Perales et al., 2004)
Determinaciones
analíticas
Figura 13. Diagrama de bloques del desarrollo general de cada experimento

28
5.4.1.1 Sincronización del cultivo
En esta fase se llevó a cabo la preparación del cultivo semilla. Para ello se realizaron 3
resiembras en medio mineral en botellas de 500mL, una resiembra cada cuatro días, para
asegurar que las células se dividieran al mismo tiempo, eso es de manera sincrónica.
5.4.1.2 Inoculación a partir de matraz semilla
El biorreactor de 2 L fue inoculado al 10%(v/v), equivalente a 200 mL del matraz semilla.
5.4.1.3 Crecimiento
Durante el crecimiento se mantuvieron las siguientes condiciones de cultivo:
• Aireación: 0.5 vvm
• Iluminación: 400 µmol m-2 s-1
• Temperatura: 22 ºC
• Volumen de trabajo: 2 L
• Tipo de medio: mineral (Anexo I)
5.4.1.4 Aplicación de estrés
El tipo de estrés aplicado fue nutricional o lumínico o la combinación de ambos. La
aplicación de estrés se hizo cuando el cultivo alcanzó su mayor crecimiento, lo cual fue
establecido por medio de las determinaciones analíticas efectuadas a lo largo de este
periodo.
Para la aplicación de estrés nutricional se realizó un lavado de células con el fin de eliminar
los nutrientes del medio mineral (Anexo II), mientras que para la aplicación de estrés
lumínico se agregaron 350 µmol m-2 s-1 de luz con una lámpara Tecnolite ML-150
teniendo cuidado en mantenerla a una distancia de aproximadamente 20 cm de manera que
la elevación de temperatura no fuera excesiva y causara la lisis celular.
Durante la aplicación de estrés el cultivo se mantuvo bajo las mismas condiciones a las que
estaba durante el crecimiento.

29
5.4.1.5 Determinaciones analíticas
5.4.1.5.1 Peso seco (PS)
Se filtraron 10 mL de cultivo a través de una membrana Millipore de 0.5 µm previamente
puesta a peso constante. La membrana conteniendo la biomasa se colocó en un horno a
70ºC de manera que alcanzara el peso constante. Después de 24 horas la membrana se pesó
y la diferencia de este peso con el de la membrana antes de filtrar el cultivo determinó el
peso seco.
5.4.1.5.2 Recuento celular directo
El conteo celular se realizó en una cámara de Neubauer, donde se contaron las células en
todos los cuadrantes para obtener un valor promedio que fue multiplicado por un factor de
104 para obtener el número de células mL-1.
5.4.1.5.3 Clorofilas (a y b) y carotenoides totales (Anexo III).
Se realizó una extracción con metanol caliente a 2mL del cultivo. Se determinó la
absorbancia del extracto metanólico a 666, 653 y 470 nm. Se calculó la concentración de
clorofilas a y b y carotenoides totales en base a las ecuaciones propuestas por Wellburn
(1994).
5.4.1.5.4 Perfil de carotenoides por HPLC – DAD (Anexo IV)
Al final de los experimentos, se tomaron alícuotas de 25 mL de cultivo y se extrajeron las
xantofilas totales con HEAT (Hexano: Etanol: Acetona: Tolueno; 10:6:7:7). El extracto fue
leído en un Cromatógrafo de Líquidos de Alta Resolución (HPLC) marca Hewlett Packard
series 1100.
Los picos de los cromatogramas fueron identificados en base a los tiempos de retención, a
las máximas longitudes de onda características de cada pigmento y al espectro de absorción
desarrollado por cada pico.

30
5.4.1.5.5 Concentración final de cetocarotenoides (Anexo V)
El contenido específico de cetocarotenoides (gramos de cetocarotenoides/gramos de peso
seco) fue determinado a partir de alícuotas de 4mL del cultivo tomadas al final de los
experimentos, siendo a su vez determinado el peso seco.
Es importante mencionar que la técnica empleada en esta determinación (Boussiba, 1992)
fue diseñada para Haematococcus pluvialis, por lo que se considera que cuando las células
son verdes el valor obtenido en la cuantificación se refiere a los cetocarotenoides, pero
cuando las células de H. pluvialis forman quistes y se tornan rojas, el valor obtenido por la
técnica se puede considerar exclusivo del cetocarotenoide astaxantina.

31
6. RESULTADOS Y DISCUSIÓN
6.1 Crecimiento celular con fotoperiodo (FP) y luz continua (LC)
Los cultivos con fotoperiodo tuvieron una velocidad de crecimiento de de 0.308 ± 0.065 d-1
antes de ser estresados, la cual resultó superior a la velocidad de los crecidos con LC, donde
ésta fue de 0.168 ± 0.056 d-1. Como se observa en la figura 14 los valores de peso seco (PS)
en los biorreactores con FP y LC fueron similares hasta el día 8, después del cual los
experimentos con LC presentaron una desaceleración en el crecimiento, evento que ocurre
en los fotobiorreactores con FP hasta el día 14. En el recuento celular, los fotobiorreactores
con LC tuvieron un número máximo (14.65*106 células mL-1) el día 6 y los
fotobiorreactores con FP (22.05*106 células mL-1) hasta el día 11, coincidiendo en ambos
casos con las concentraciones máximas en cuantificación de clorofilas totales. El aumento
de PS posterior al día en el que se alcanza el mayor contenido de células puede deberse al
aumento del tamaño celular.
Figura 14. Cinética de crecimiento por PS y recuento celular directo en fotobiorreactores
con FP y LC.
Es importante destacar es que los experimentos realizados con FP presentaron un mayor
número de células que los cultivados con LC. Esto se debe a que la división celular ocurre
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 2 4 6 8 10 12 14 16t(d)
Peso seco (g/L)
0
5
10
15
20
25
No. de células* 106
Cinética de crecimiento en base a PS ( FP) Cinética de crecimiento en base a PS ( LC)
Recuento celular (FP) Recuento celular (LC)
32
principalmente al inicio del ciclo de oscuridad, situación que en los cultivos con LC no se
presenta y que por lo tanto no les permitió dividirse con la misma oportunidad que las
células mantenidas con FP (Kaftan, et al, 1999).
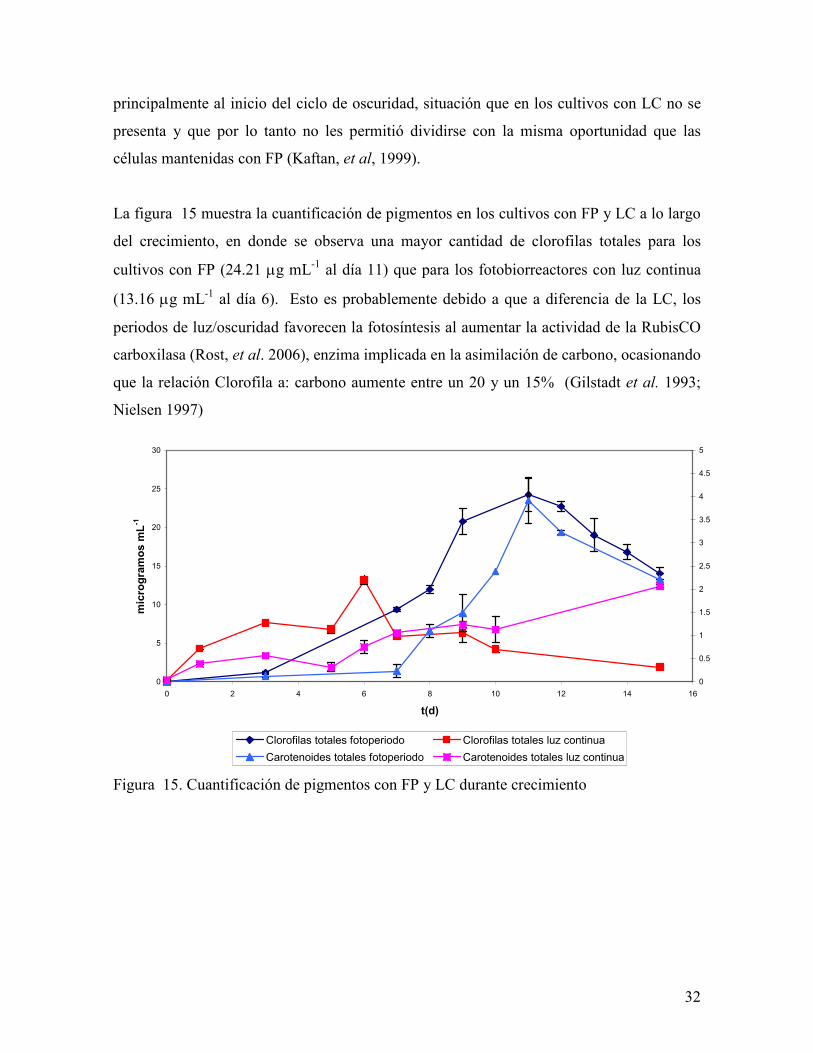
La figura 15 muestra la cuantificación de pigmentos en los cultivos con FP y LC a lo largo
del crecimiento, en donde se observa una mayor cantidad de clorofilas totales para los
cultivos con FP (24.21 µg mL-1 al día 11) que para los fotobiorreactores con luz continua
(13.16 µg mL-1 al día 6). Esto es probablemente debido a que a diferencia de la LC, los
periodos de luz/oscuridad favorecen la fotosíntesis al aumentar la actividad de la RubisCO
carboxilasa (Rost, et al. 2006), enzima implicada en la asimilación de carbono, ocasionando
que la relación Clorofila a: carbono aumente entre un 20 y un 15% (Gilstadt et al. 1993;
Nielsen 1997)
0
5
10
15
20
25
30
0 2 4 6 8 10 12 14 16
t(d)
microgramos mL-1
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Clorofilas totales fotoperiodo Clorofilas totales luz continua
Carotenoides totales fotoperiodo Carotenoides totales luz continua
Figura 15. Cuantificación de pigmentos con FP y LC durante crecimiento
33
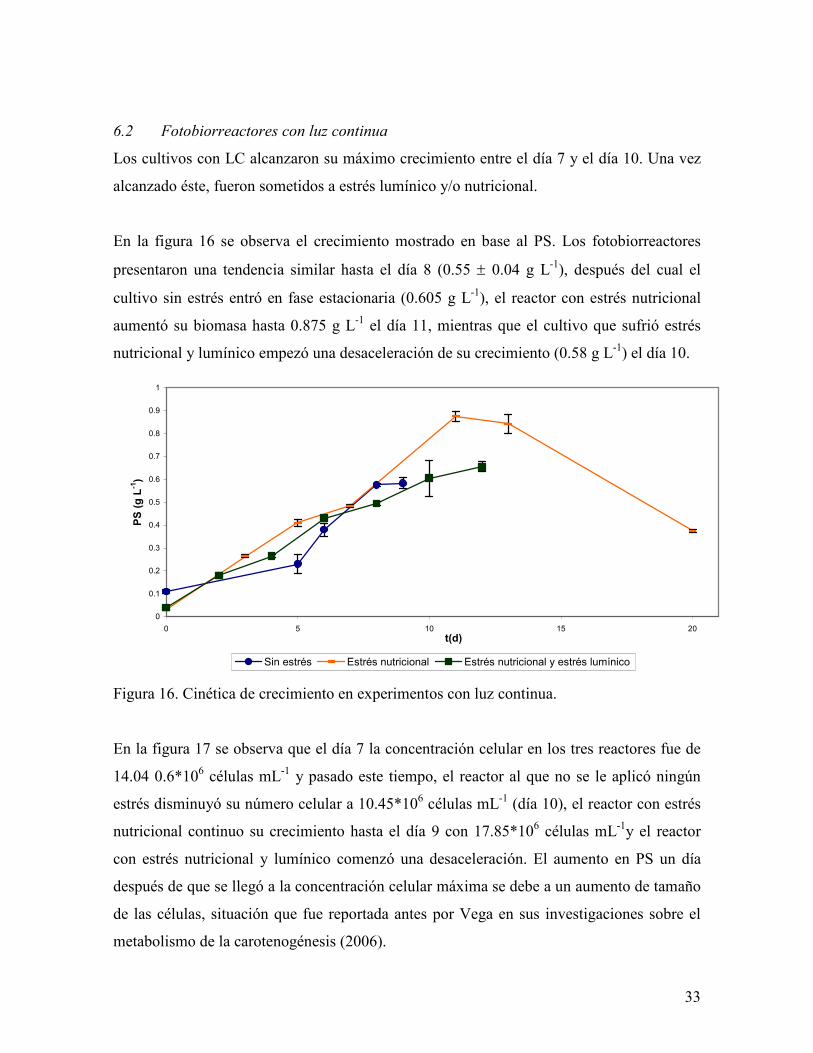
6.2 Fotobiorreactores con luz continua
Los cultivos con LC alcanzaron su máximo crecimiento entre el día 7 y el día 10. Una vez
alcanzado éste, fueron sometidos a estrés lumínico y/o nutricional.
En la figura 16 se observa el crecimiento mostrado en base al PS. Los fotobiorreactores
presentaron una tendencia similar hasta el día 8 (0.55 ± 0.04 g L-1), después del cual el
cultivo sin estrés entró en fase estacionaria (0.605 g L-1), el reactor con estrés nutricional
aumentó su biomasa hasta 0.875 g L-1 el día 11, mientras que el cultivo que sufrió estrés
nutricional y lumínico empezó una desaceleración de su crecimiento (0.58 g L-1) el día 10.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 5 10 15 20
t(d)
PS (g L-1)
Sin estrés Estrés nutricional Estrés nutricional y estrés lumínico
Figura 16. Cinética de crecimiento en experimentos con luz continua.
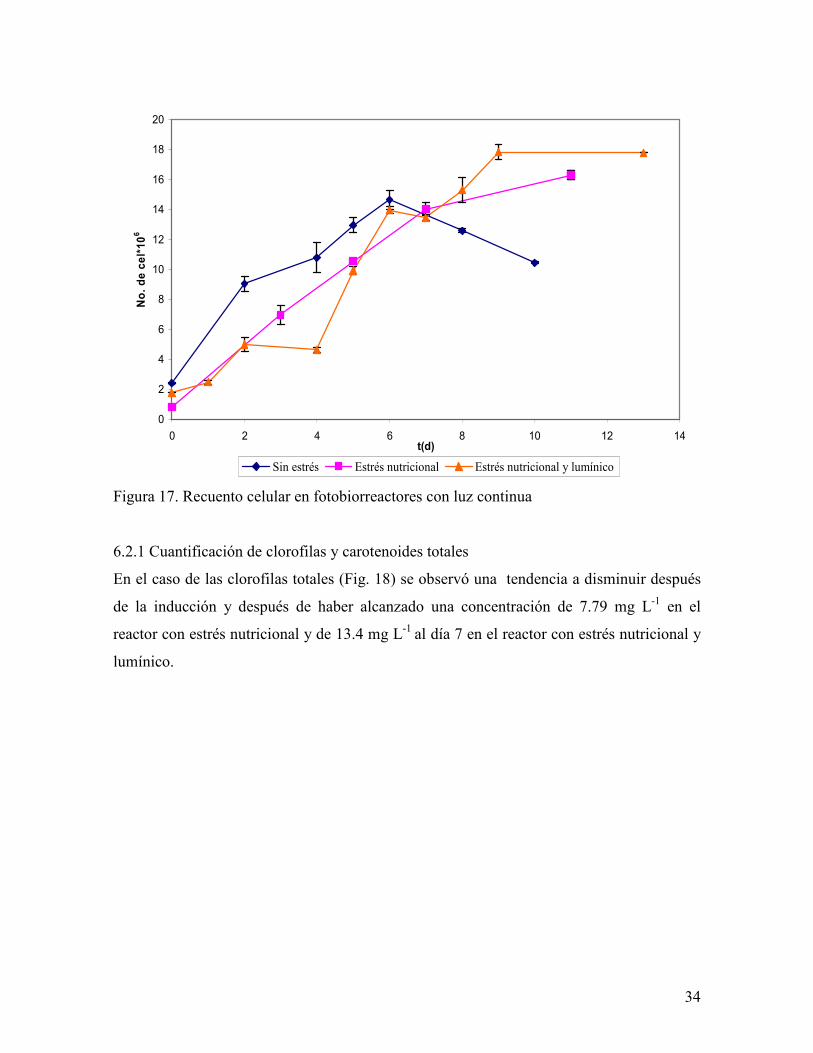
En la figura 17 se observa que el día 7 la concentración celular en los tres reactores fue de
14.04 0.6*106 células mL-1 y pasado este tiempo, el reactor al que no se le aplicó ningún
estrés disminuyó su número celular a 10.45*106 células mL-1 (día 10), el reactor con estrés
nutricional continuo su crecimiento hasta el día 9 con 17.85*106 células mL-1y el reactor
con estrés nutricional y lumínico comenzó una desaceleración. El aumento en PS un día
después de que se llegó a la concentración celular máxima se debe a un aumento de tamaño
de las células, situación que fue reportada antes por Vega en sus investigaciones sobre el
metabolismo de la carotenogénesis (2006).
34
0
2
4
6
8
10
12
14
16
18
20
0 2 4 6 8 10 12 14t(d)
No. de cel*106
Sin estrés Estrés nutricional Estrés nutricional y lumínico
Figura 17. Recuento celular en fotobiorreactores con luz continua
6.2.1 Cuantificación de clorofilas y carotenoides totales
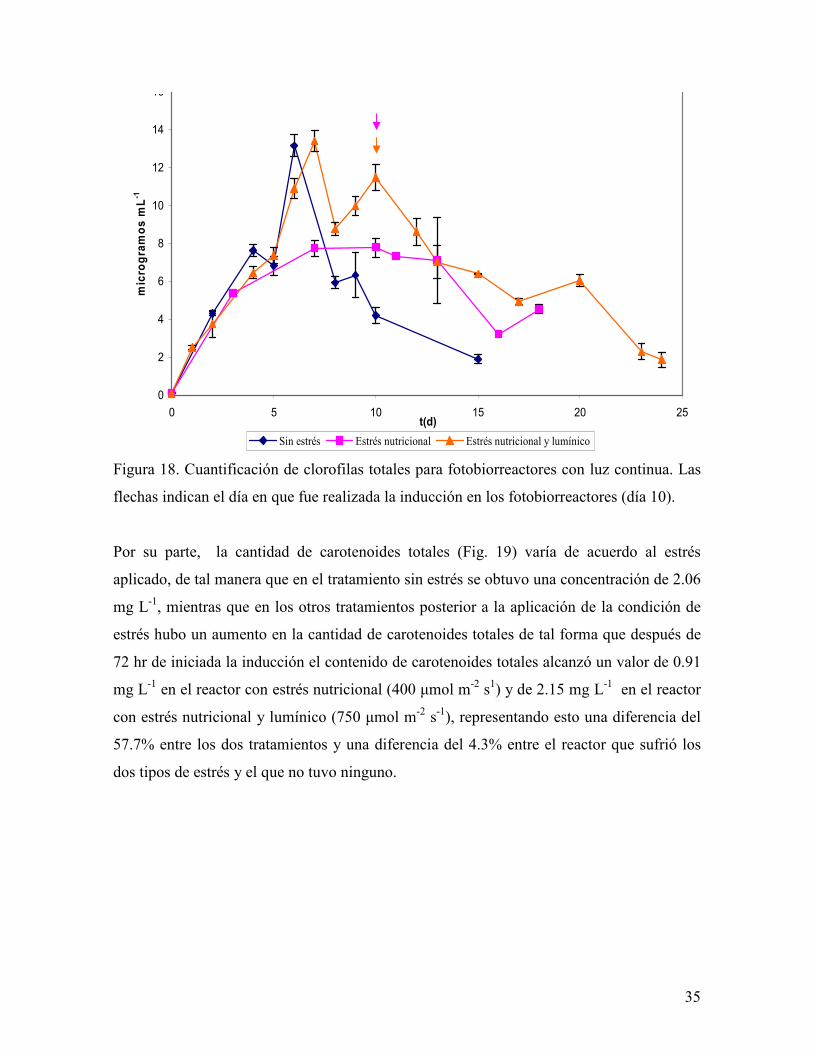
En el caso de las clorofilas totales (Fig. 18) se observó una tendencia a disminuir después
de la inducción y después de haber alcanzado una concentración de 7.79 mg L-1 en el
reactor con estrés nutricional y de 13.4 mg L-1 al día 7 en el reactor con estrés nutricional y
lumínico.
35
0
2
4
6
8
10
12
14
16
0 5 10 15 20 25t(d)
microgramos mL-1
Sin estrés Estrés nutricional Estrés nutricional y lumínico
Figura 18. Cuantificación de clorofilas totales para fotobiorreactores con luz continua. Las
flechas indican el día en que fue realizada la inducción en los fotobiorreactores (día 10).
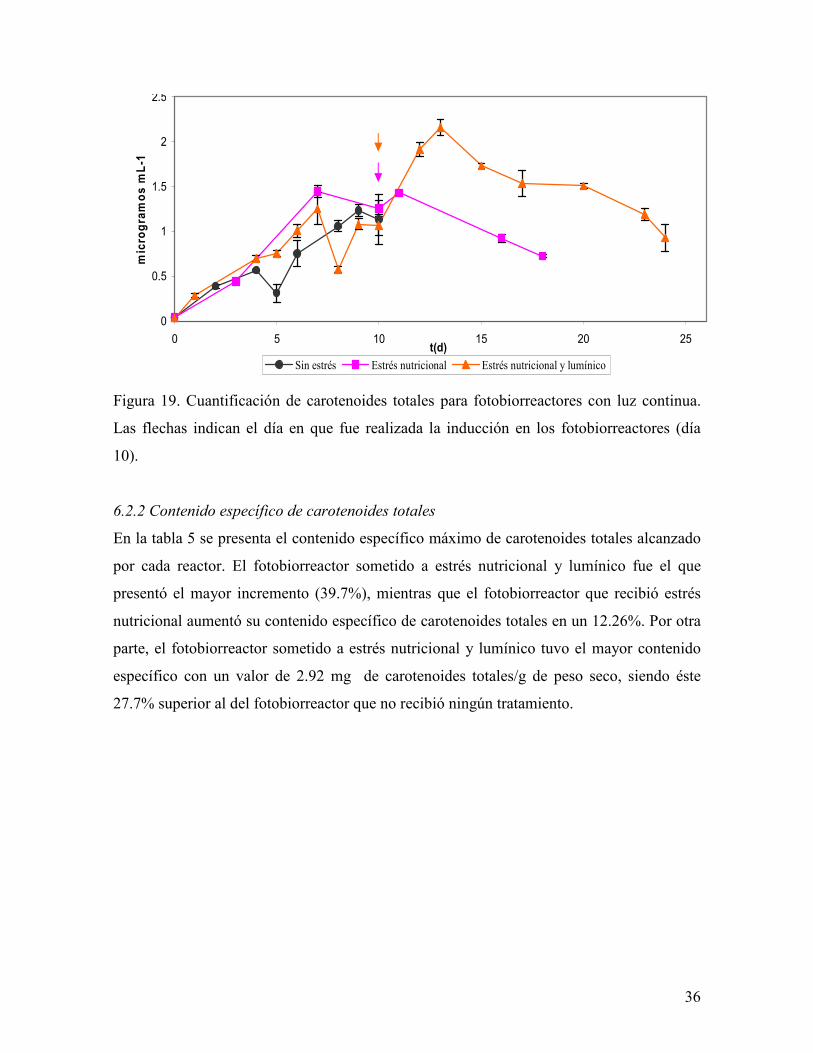
Por su parte, la cantidad de carotenoides totales (Fig. 19) varía de acuerdo al estrés
aplicado, de tal manera que en el tratamiento sin estrés se obtuvo una concentración de 2.06
mg L-1, mientras que en los otros tratamientos posterior a la aplicación de la condición de
estrés hubo un aumento en la cantidad de carotenoides totales de tal forma que después de
72 hr de iniciada la inducción el contenido de carotenoides totales alcanzó un valor de 0.91
mg L-1 en el reactor con estrés nutricional (400 µmol m-2 s1) y de 2.15 mg L-1 en el reactor
con estrés nutricional y lumínico (750 µmol m-2 s-1), representando esto una diferencia del
57.7% entre los dos tratamientos y una diferencia del 4.3% entre el reactor que sufrió los
dos tipos de estrés y el que no tuvo ninguno.
36
0
0.5
1
1.5
2
2.5
0 5 10 15 20 25t(d)
microgramos mL-1
Sin estrés Estrés nutricional Estrés nutricional y lumínico
Figura 19. Cuantificación de carotenoides totales para fotobiorreactores con luz continua.
Las flechas indican el día en que fue realizada la inducción en los fotobiorreactores (día
10).
6.2.2 Contenido específico de carotenoides totales
En la tabla 5 se presenta el contenido específico máximo de carotenoides totales alcanzado
por cada reactor. El fotobiorreactor sometido a estrés nutricional y lumínico fue el que
presentó el mayor incremento (39.7%), mientras que el fotobiorreactor que recibió estrés
nutricional aumentó su contenido específico de carotenoides totales en un 12.26%. Por otra
parte, el fotobiorreactor sometido a estrés nutricional y lumínico tuvo el mayor contenido
específico con un valor de 2.92 mg de carotenoides totales/g de peso seco, siendo éste
27.7% superior al del fotobiorreactor que no recibió ningún tratamiento.
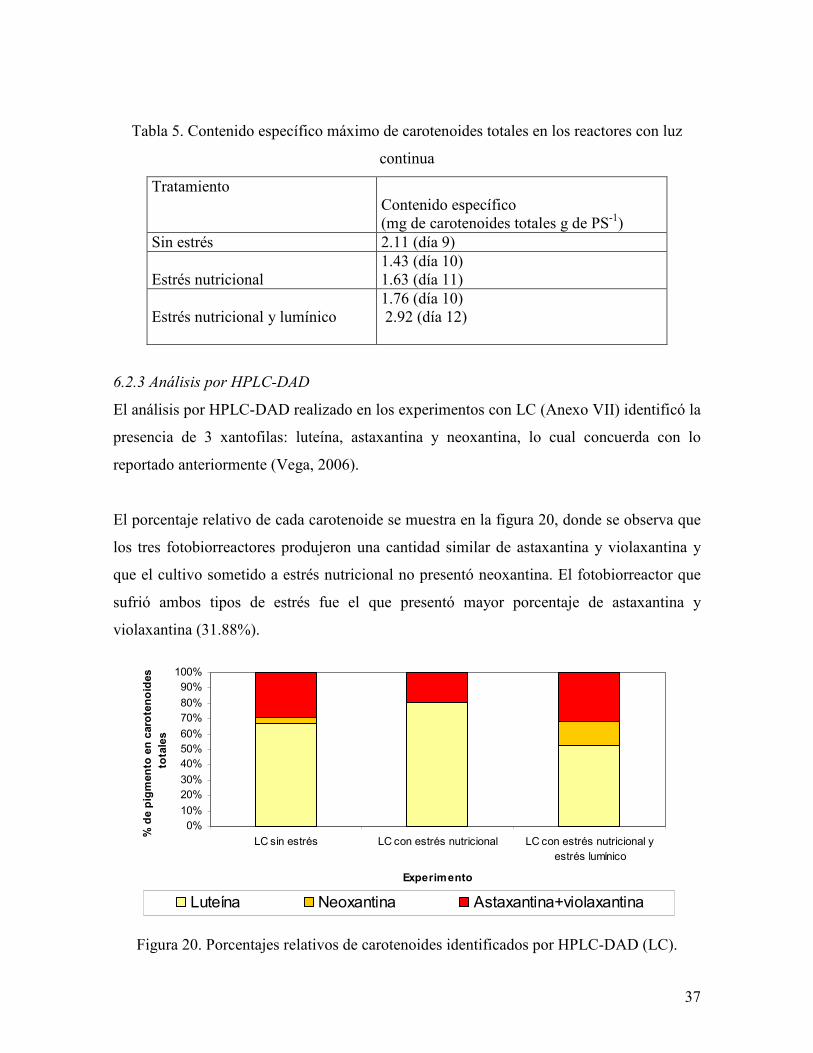
37
Tabla 5. Contenido específico máximo de carotenoides totales en los reactores con luz
continua
Tratamiento
Contenido específico (mg de carotenoides totales g de PS-1)
Sin estrés 2.11 (día 9)
Estrés nutricional 1.43 (día 10) 1.63 (día 11)
Estrés nutricional y lumínico
1.76 (día 10) 2.92 (día 12)
6.2.3 Análisis por HPLC-DAD
El análisis por HPLC-DAD realizado en los experimentos con LC (Anexo VII) identificó la
presencia de 3 xantofilas: luteína, astaxantina y neoxantina, lo cual concuerda con lo
reportado anteriormente (Vega, 2006).
El porcentaje relativo de cada carotenoide se muestra en la figura 20, donde se observa que
los tres fotobiorreactores produjeron una cantidad similar de astaxantina y violaxantina y
que el cultivo sometido a estrés nutricional no presentó neoxantina. El fotobiorreactor que
sufrió ambos tipos de estrés fue el que presentó mayor porcentaje de astaxantina y
violaxantina (31.88%).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
LC sin estrés LC con estrés nutricional LC con estrés nutricional y
estrés lumínico
Experimento
% de pigmento en carotenoides
totales
Luteína Neoxantina Astaxantina+violaxantina
Figura 20. Porcentajes relativos de carotenoides identificados por HPLC-DAD (LC).

38
La luteína ha sido reportada anteriormente como el principal carotenoide primario
producido por especies de Scenedesmus (Bishop, 1996; Orosa et al., 2001; Qin et al.,
2008). En el caso de los tres cultivos realizados con LC la luteína fue el carotenoide
predominante. El mayor porcentaje de luteína fue presentado por el fotobiorreactor que
recibió estrés nutricional (80.33%), mientras que los otros dos reactores manifestaron un
porcentaje menor, con 67.15% el fotobiorreactor que no fue sometido a ningún estrés y con
52.28% el fotobiorreactor sometido a estrés nutricional y lumínico. La disminución que
presenta el pigmento en el cultivo que sufrió ambos tipos de estrés probablemente se deba
no sólo a que la luteína represente un metabolito primario en esta especie, sino también a
que el pigmento fue degradado por el efecto de la luz. Esto sugiere que la actividad
antioxidante de la luteína es más limitada que la de otros carotenoides, de hecho presenta
una constante de rompimiento de oxígeno singulete (kq= 0.8*1010 L mol-1 s-1) menor que
otras xantofilas como la astaxantina. (kq= 2.4*1010 L mol-1 s-1) (Di Mascio et al. 1989; Di
Mascio et al. 1990; Di Mascio et al. 1991).
Por otro lado el contenido similar de astaxantina en los tres cultivos sugiere que este
carotenoide es producido cuando el cultivo es crecido a LUZ CONTINUA y su proporción
se mantiene constante aún cuando después sea sometido a condiciones de estrés nutricional
y lumínico.
6.2.4 Cuantificación de cetocarotenoides
El análisis por HPLC-DAD revela la presencia de astaxantina, de manera que los
porcentajes presentados de cetocarotenoides se identifican principalmente como dicho
pigmento (Fig. 21).
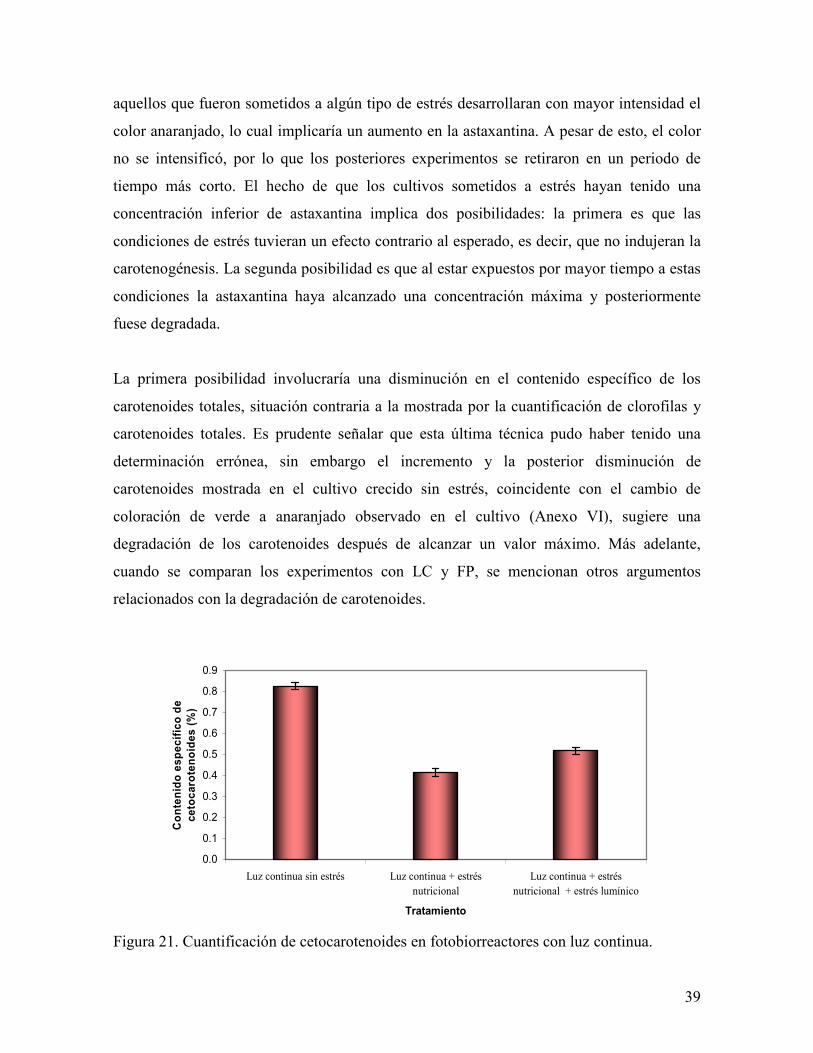
El fotobiorreactor donde se logró el mayor porcentaje de astaxantina (0.83 %) fue el que no
recibió ningún estrés, mientras que los otros dos reactores presentaron un porcentaje similar
entre ellos (0.47± 0.07%). Se debe tomar en cuenta que los reactores se retiraron en
diferentes días (a los 40 días LC con estrés nutricional, a los 25 días LC con estrés
nutricional y lumínico y a los 15 días LC sin estrés), ya que en un principio se esperaba que
39
aquellos que fueron sometidos a algún tipo de estrés desarrollaran con mayor intensidad el
color anaranjado, lo cual implicaría un aumento en la astaxantina. A pesar de esto, el color
no se intensificó, por lo que los posteriores experimentos se retiraron en un periodo de
tiempo más corto. El hecho de que los cultivos sometidos a estrés hayan tenido una
concentración inferior de astaxantina implica dos posibilidades: la primera es que las
condiciones de estrés tuvieran un efecto contrario al esperado, es decir, que no indujeran la
carotenogénesis. La segunda posibilidad es que al estar expuestos por mayor tiempo a estas
condiciones la astaxantina haya alcanzado una concentración máxima y posteriormente
fuese degradada.
La primera posibilidad involucraría una disminución en el contenido específico de los
carotenoides totales, situación contraria a la mostrada por la cuantificación de clorofilas y
carotenoides totales. Es prudente señalar que esta última técnica pudo haber tenido una
determinación errónea, sin embargo el incremento y la posterior disminución de
carotenoides mostrada en el cultivo crecido sin estrés, coincidente con el cambio de
coloración de verde a anaranjado observado en el cultivo (Anexo VI), sugiere una
degradación de los carotenoides después de alcanzar un valor máximo. Más adelante,
cuando se comparan los experimentos con LC y FP, se mencionan otros argumentos
relacionados con la degradación de carotenoides.
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Luz continua sin estrés Luz continua + estrésnutricional
Luz continua + estrésnutricional + estrés lumínico
Tratamiento
Contenido específico de
cetocarotenoides (%)
Figura 21. Cuantificación de cetocarotenoides en fotobiorreactores con luz continua.
40
6.3 Fotoperiodo (12/12)
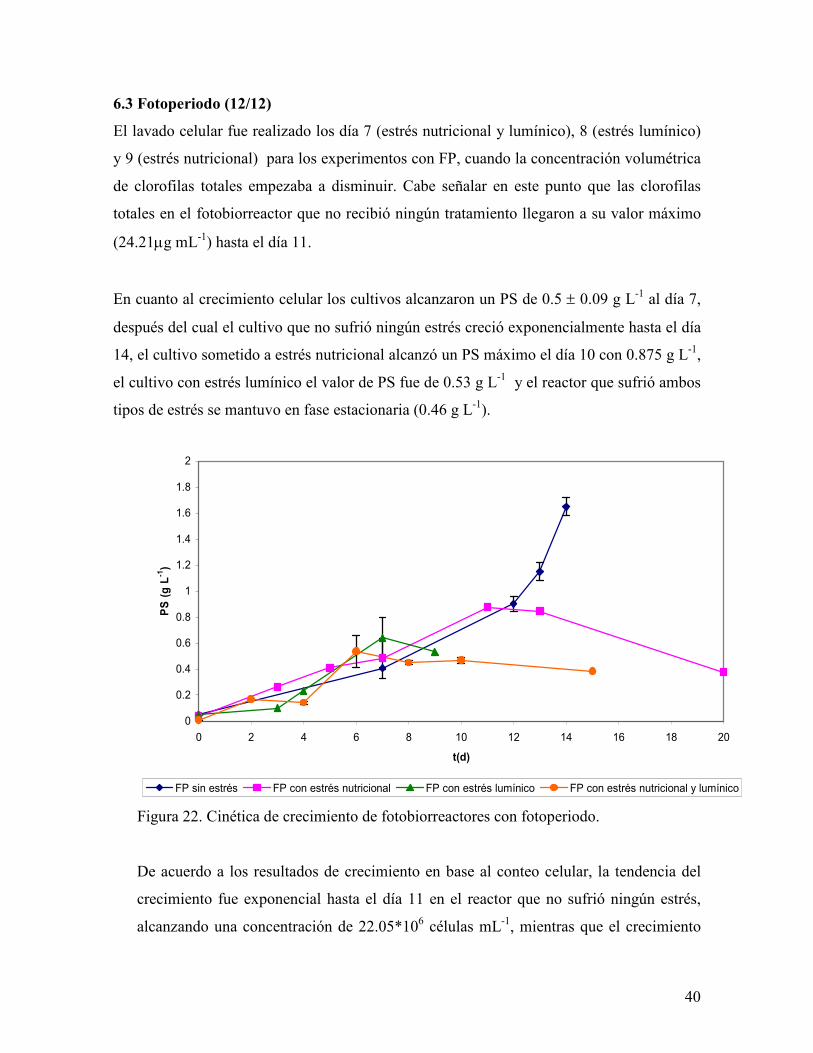
El lavado celular fue realizado los día 7 (estrés nutricional y lumínico), 8 (estrés lumínico)
y 9 (estrés nutricional) para los experimentos con FP, cuando la concentración volumétrica
de clorofilas totales empezaba a disminuir. Cabe señalar en este punto que las clorofilas
totales en el fotobiorreactor que no recibió ningún tratamiento llegaron a su valor máximo
(24.21µg mL-1) hasta el día 11.
En cuanto al crecimiento celular los cultivos alcanzaron un PS de 0.5 ± 0.09 g L-1 al día 7,
después del cual el cultivo que no sufrió ningún estrés creció exponencialmente hasta el día
14, el cultivo sometido a estrés nutricional alcanzó un PS máximo el día 10 con 0.875 g L-1,
el cultivo con estrés lumínico el valor de PS fue de 0.53 g L-1 y el reactor que sufrió ambos
tipos de estrés se mantuvo en fase estacionaria (0.46 g L-1).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
0 2 4 6 8 10 12 14 16 18 20
t(d)
PS (g L-1)
FP sin estrés FP con estrés nutricional FP con estrés lumínico FP con estrés nutricional y lumínico
Figura 22. Cinética de crecimiento de fotobiorreactores con fotoperiodo.
De acuerdo a los resultados de crecimiento en base al conteo celular, la tendencia del
crecimiento fue exponencial hasta el día 11 en el reactor que no sufrió ningún estrés,
alcanzando una concentración de 22.05*106 células mL-1, mientras que el crecimiento
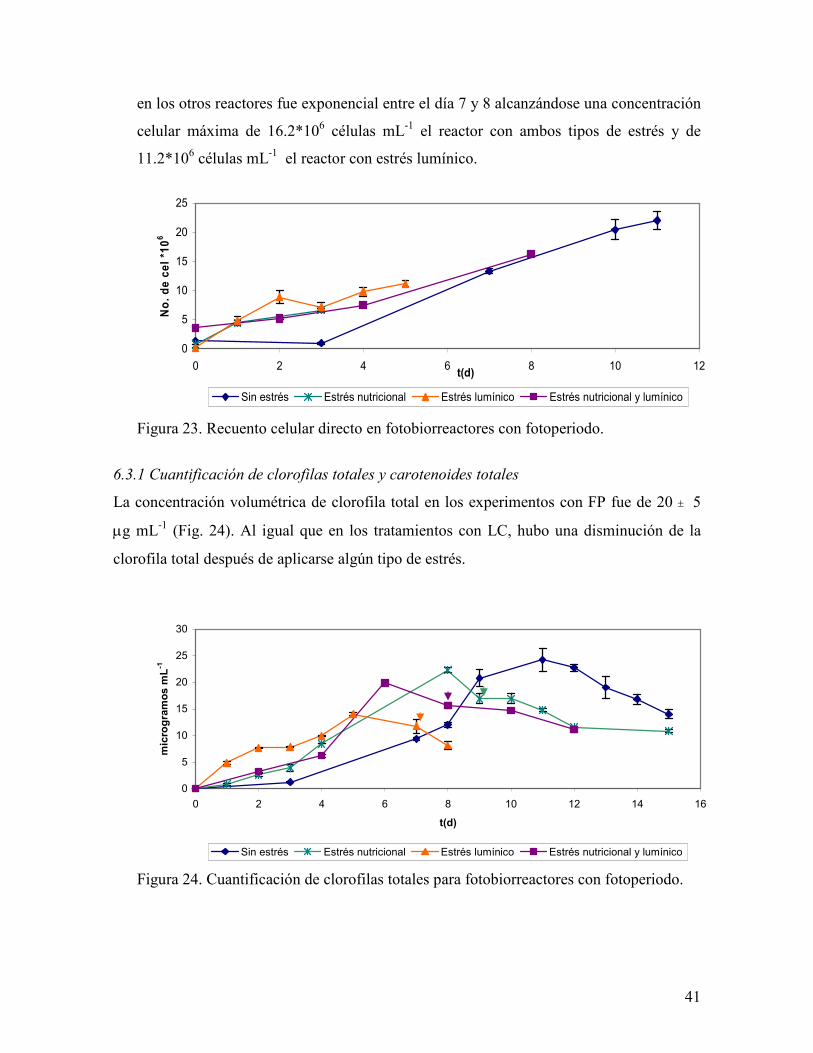
41
en los otros reactores fue exponencial entre el día 7 y 8 alcanzándose una concentración
celular máxima de 16.2*106 células mL-1 el reactor con ambos tipos de estrés y de
11.2*106 células mL-1 el reactor con estrés lumínico.
0
5
10
15
20
25
0 2 4 6 8 10 12t(d)
No. de cel *106
Sin estrés Estrés nutricional Estrés lumínico Estrés nutricional y lumínico
Figura 23. Recuento celular directo en fotobiorreactores con fotoperiodo.
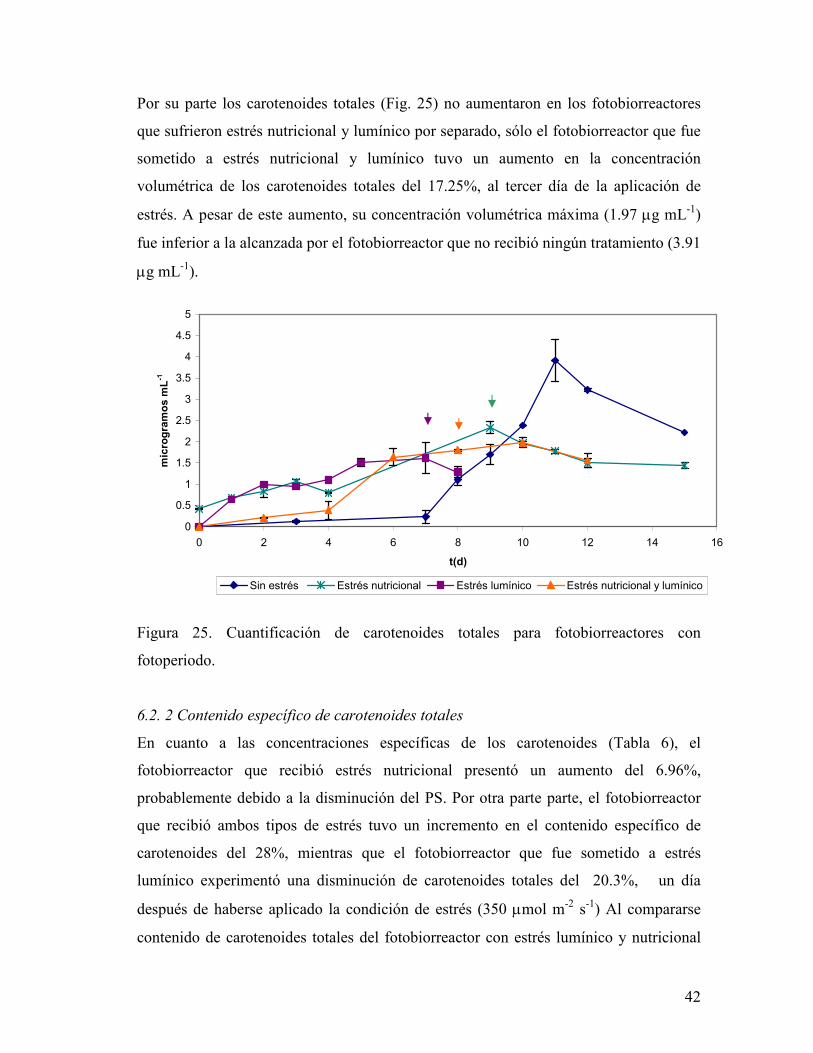
6.3.1 Cuantificación de clorofilas totales y carotenoides totales
La concentración volumétrica de clorofila total en los experimentos con FP fue de 20 ± 5
µg mL-1 (Fig. 24). Al igual que en los tratamientos con LC, hubo una disminución de la
clorofila total después de aplicarse algún tipo de estrés.
0
5
10
15
20
25
30
0 2 4 6 8 10 12 14 16
t(d)
microgramos mL-1
Sin estrés Estrés nutricional Estrés lumínico Estrés nutricional y lumínico
Figura 24. Cuantificación de clorofilas totales para fotobiorreactores con fotoperiodo.
42
Por su parte los carotenoides totales (Fig. 25) no aumentaron en los fotobiorreactores
que sufrieron estrés nutricional y lumínico por separado, sólo el fotobiorreactor que fue
sometido a estrés nutricional y lumínico tuvo un aumento en la concentración
volumétrica de los carotenoides totales del 17.25%, al tercer día de la aplicación de
estrés. A pesar de este aumento, su concentración volumétrica máxima (1.97 µg mL-1)
fue inferior a la alcanzada por el fotobiorreactor que no recibió ningún tratamiento (3.91
µg mL-1).
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
0 2 4 6 8 10 12 14 16
t(d)
microgramos mL-1
Sin estrés Estrés nutricional Estrés lumínico Estrés nutricional y lumínico
Figura 25. Cuantificación de carotenoides totales para fotobiorreactores con
fotoperiodo.
6.2. 2 Contenido específico de carotenoides totales
En cuanto a las concentraciones específicas de los carotenoides (Tabla 6), el
fotobiorreactor que recibió estrés nutricional presentó un aumento del 6.96%,
probablemente debido a la disminución del PS. Por otra parte parte, el fotobiorreactor
que recibió ambos tipos de estrés tuvo un incremento en el contenido específico de
carotenoides del 28%, mientras que el fotobiorreactor que fue sometido a estrés
lumínico experimentó una disminución de carotenoides totales del 20.3%, un día
después de haberse aplicado la condición de estrés (350 µmol m-2 s-1) Al compararse
contenido de carotenoides totales del fotobiorreactor con estrés lumínico y nutricional
43
con el que no fue sometido a ningún estrés, se observó un incremento del 15.56% en
dicho contenido en el primero de ellos.
Tabla 6. Contenido específico máximo de carotenoides totales en los reactores con
fotoperiodo
Tratamiento
Contenido específico
(mg carotenoides totales g de PS-1)
Sin estrés 3.58 (día 12) Estrés nutricional 3.36(día 10) Estrés nutricional y lumínico 4.24 (día 10) Estrés lumínico 1.52 (día 7)
6.3.3 Análisis por HPLC-DAD
El análisis de hizo con muestras tomadas al final de los experimentos. No se pudieron
identificar carotenoides en los experimentos que fueron sometidos a estrés lumínico y estrés
nutricional y lumínico (Anexo VI).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Fotoperiodo sin estrés Fotoperiodo con estrés nutricional
Experimento
% de pigmento en carotenoides
totales
Luteína Neoxantina Astaxantina+violaxantina
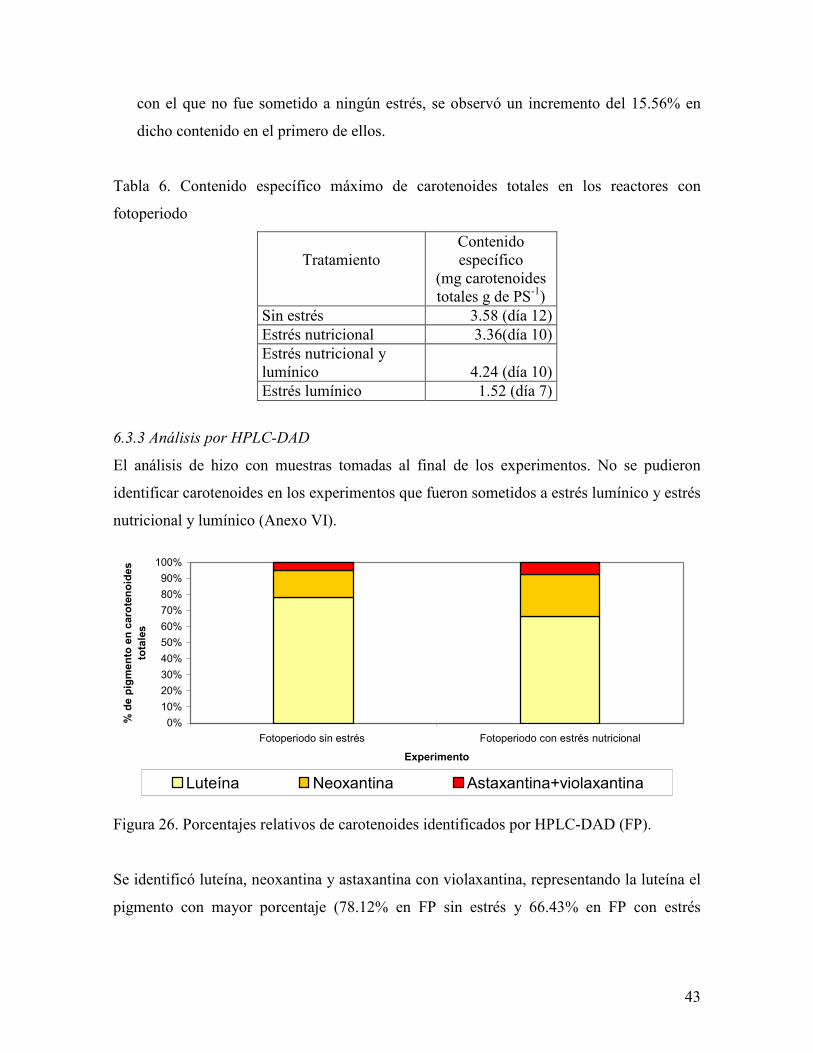
Figura 26. Porcentajes relativos de carotenoides identificados por HPLC-DAD (FP).
Se identificó luteína, neoxantina y astaxantina con violaxantina, representando la luteína el
pigmento con mayor porcentaje (78.12% en FP sin estrés y 66.43% en FP con estrés
44
nutricional). El experimento que sufrió estrés nutricional presentó un porcentaje de
astaxantina y violaxantina 2.74% más que el experimento que no sufrió ningún estrés.
6.3.4 Cuantificación de cetocarotenoides
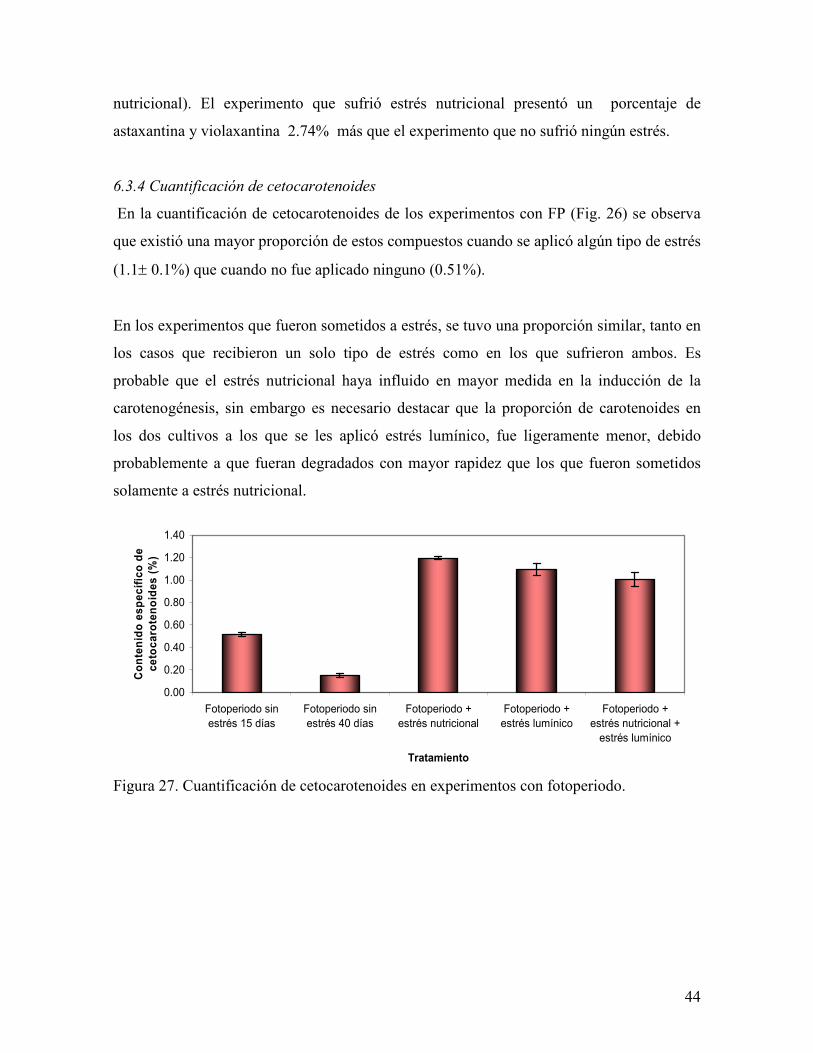
En la cuantificación de cetocarotenoides de los experimentos con FP (Fig. 26) se observa
que existió una mayor proporción de estos compuestos cuando se aplicó algún tipo de estrés
(1.1± 0.1%) que cuando no fue aplicado ninguno (0.51%).
En los experimentos que fueron sometidos a estrés, se tuvo una proporción similar, tanto en
los casos que recibieron un solo tipo de estrés como en los que sufrieron ambos. Es
probable que el estrés nutricional haya influido en mayor medida en la inducción de la
carotenogénesis, sin embargo es necesario destacar que la proporción de carotenoides en
los dos cultivos a los que se les aplicó estrés lumínico, fue ligeramente menor, debido
probablemente a que fueran degradados con mayor rapidez que los que fueron sometidos
solamente a estrés nutricional.
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
Fotoperiodo sin
estrés 15 días
Fotoperiodo sin
estrés 40 días
Fotoperiodo +
estrés nutricional
Fotoperiodo +
estrés lumínico
Fotoperiodo +
estrés nutricional +
estrés lumínico
Tratamiento
Contenido específico de
cetocarotenoides (%)
Figura 27. Cuantificación de cetocarotenoides en experimentos con fotoperiodo.

45
6.4 LC y FP después de la aplicación de estrés
El contenido específico de carotenoides en los experimentos con LC fue inferior a los
obtenidos en los reactores con FP. Aún el que fue sometido a estrés lumínico y nutricional
tuvo menor contenido específico que el obtenido por FP sin estrés.
El tratamiento que presentó mayor contenido específico de carotenoides totales fue el de FP
con estrés nutricional y lumínico con un valor de 4.24mg carotenoides totales g de PS-1.
En cuanto a los porcentajes de los carotenoides presentes, se observa que S. incrassatulus
sintetiza principalmente luteína, representando en todos los cultivos más del 50% de las
xantofilas totales. En el caso de los tratamientos con LC sin estrés y LC con estrés
nutricional y lumínico, el porcentaje de luteína disminuyó debido al aumento de la mezcla
astaxantina+violaxantina. El aumento de astaxantina como carotenoide secundario se ha
reportado anteriormente en otras especies de Scenedesmus. (Hanagata et al. 1999, Orosa et
al. 2001, Qin et al. 2008).
En comparación con los experimentos con FP, los tratamientos con LC tuvieron un mayor
porcentaje de astaxantina + luteína, mas si se observa la altura de los picos de astaxantina +
violaxantina en los cromatogramas, el pico más alto pertenece a FP más estrés nutricional
con 153.79 unidades de área (AU), es decir, que si bien tuvo un menor porcentaje de
astaxantina que los experimentos con LC, presentó una mayor cantidad de estos 2
pigmentos. Esto concordaría con lo obtenido por la técnica de cuantificación de
cetocarotenoides, donde el experimento con FP y estrés nutricional mostró el mayor
porcentaje de cetocarotenoides (1.2%) de todos los experimentos realizados.
La extracción de cetocarotenoides mostró un valor más alto en fotobiorreactores cuanto
presentaban un color verde que los que viraban a un color amarillo o anaranjado, como es
el caso de los cultivos realizados con FP (Anexo VI). Como se mencionó anteriormente, en
un principio se consideró la posibilidad de una cuantificación deficiente, sin embargo al
medir el contenido específico de cetocarotenoides en el fotobiorreactor sometido a estrés
nutricional y lumínico (FP) en los días posteriores a la aplicación de estrés se observó que
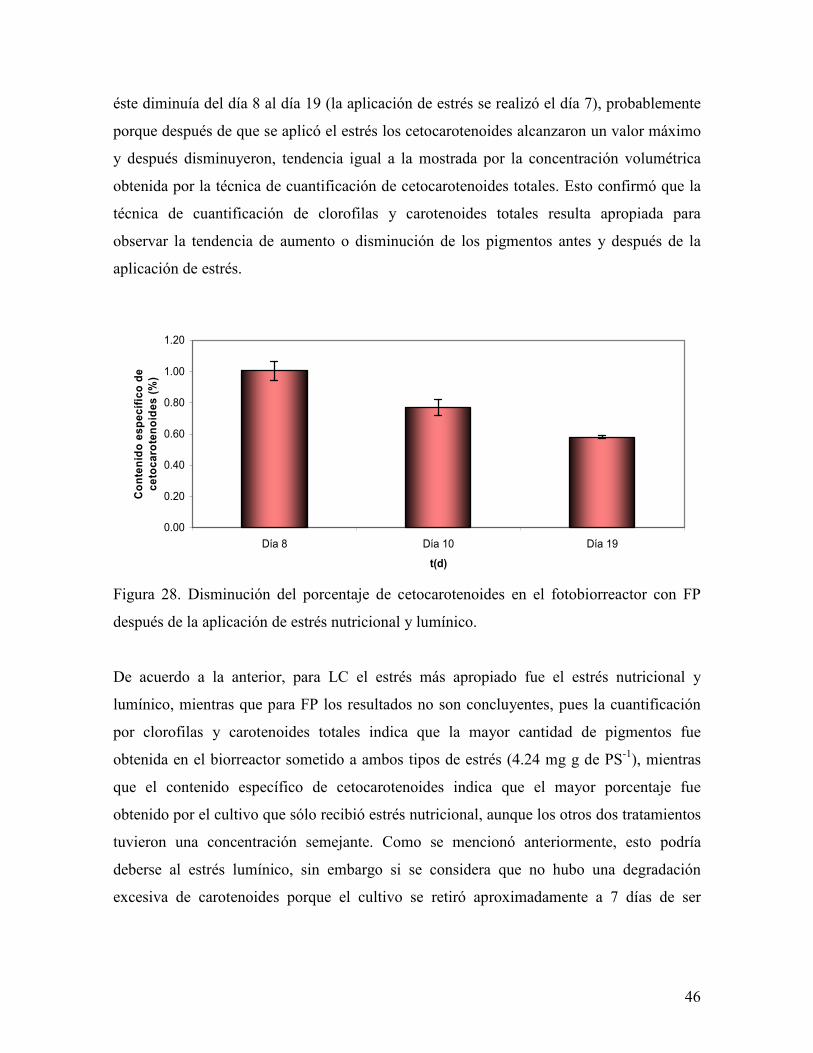
46
éste diminuía del día 8 al día 19 (la aplicación de estrés se realizó el día 7), probablemente
porque después de que se aplicó el estrés los cetocarotenoides alcanzaron un valor máximo
y después disminuyeron, tendencia igual a la mostrada por la concentración volumétrica
obtenida por la técnica de cuantificación de cetocarotenoides totales. Esto confirmó que la
técnica de cuantificación de clorofilas y carotenoides totales resulta apropiada para
observar la tendencia de aumento o disminución de los pigmentos antes y después de la
aplicación de estrés.
0.00
0.20
0.40
0.60
0.80
1.00
1.20
Día 8 Día 10 Día 19
t(d)
Contenido específico de
cetocarotenoides (%)
Figura 28. Disminución del porcentaje de cetocarotenoides en el fotobiorreactor con FP
después de la aplicación de estrés nutricional y lumínico.
De acuerdo a la anterior, para LC el estrés más apropiado fue el estrés nutricional y
lumínico, mientras que para FP los resultados no son concluyentes, pues la cuantificación
por clorofilas y carotenoides totales indica que la mayor cantidad de pigmentos fue
obtenida en el biorreactor sometido a ambos tipos de estrés (4.24 mg g de PS-1), mientras
que el contenido específico de cetocarotenoides indica que el mayor porcentaje fue
obtenido por el cultivo que sólo recibió estrés nutricional, aunque los otros dos tratamientos
tuvieron una concentración semejante. Como se mencionó anteriormente, esto podría
deberse al estrés lumínico, sin embargo si se considera que no hubo una degradación
excesiva de carotenoides porque el cultivo se retiró aproximadamente a 7 días de ser

47
sometido a estrés a diferencia de los fotobiorreactores con LC que fueron sometidos a las
condiciones de estrés por más tiempo. (15 a 30 días después de la aplicación de estrés).
Por otra parte, si bien los porcentajes de cetocarotenoides (astaxantina) obtenidos en los
experimentos con LC y FP demuestran una carotenogénesis inmediata después de la
aplicación de estrés, no son comparables con los porcentajes obtenidos por la microalga
superproductora Haematococcus pluvialis, pues ésta presenta un 4% de astaxantina por PS
mientras que la presente cepa en su máximo valor mostró 1.2% (FP con estrés nutricional).
Finalmente, cabe señalar que aún cuando en los experimentos realizados con LC se obtuvo
una menor cantidad de biomasa y de pigmentos, se debe considerar que la degradación de
clorofilas fue más rápida y por lo tanto los pigmentos fueron mostrados con mayor rapidez.
De esta manera, el proceso de extracción de carotenoides se facilita, pues la presencia de
clorofilas es escasa y no resulta necesaria su separación de los carotenoides, además de que
no es requerida una posterior eliminación del solvente. Esto representaría una disminución
en los costos de producción, además de que es posible que se pudiera suministrar
directamente a peces y aves de cría.

48
7. CONCLUSIONES
Los resultados obtenidos indican que el estrés lumínico y nutricional estimularon la
carotenogénesis y que además, la luz continua es una forma adicional de estresar a la
célula.
El crecimiento e inducción con fotoperiodo permitió obtener una mayor cantidad de
biomasa y de carotenoides, mientras que el crecimiento con luz continua aumentó el
porcentaje de carotenoides secundarios y la degradación de éstos.
El aumento en los carotenoides secundarios estuvo representado por el aumento de la
concentración de astaxantina+violaxantina.
El fotobiorreactor que presentó el máximo contenido específico de cetocarotenoides fue el
sometido a estrés nutricional con fotoperiodo (1.2%).

49
8. RECOMENDACIONES PARA TRABAJOS FUTUROS
Realizar cultivos de Scenedesmus incrassatulus crecidos con fotoperiodo y posteriormente
aplicar estrés nutricional y lumínico con luz continua.
Realizar ensayos con las técnicas de cuantificación de clorofilas y carotenoides totales,
contenido específico de cetocarotenoides y HPLC-DAD empleando soluciones estándares
de pigmentos de manera que se pueda determinar la exactitud y la sensibilidad de las
técnicas.
Determinar el tiempo exacto al cual los carotenoides totales y contenido específico de
cetocarotenoides alcanza su punto máximo y luego empieza a disminuir realizando
mediciones espaciadas por periodos más cortos en los primeros días después de la
aplicación de estrés en cultivos de S. incrassatulus.
Emplear el cultivo obtenido después de que alcanzó la máxima concentración de
carotenoides totales y el contenido máximo de cetocarotenoides, para alimentar animales de
acuacultura y avicultura, de tal manera que se estudie el efecto de los carotenoides de S.
incrassatulus en la coloración y el crecimiento de estos animales.

50
9. REFERENCIAS
Ausich, R.L., (1997) Commercial opportunities for carotenoid production by
biotechnology. Pure Appl. Chem., 69(10): 2169-2173.
Barbo Hernández, L., (2003) Técnica analítica para la cuantificación de la proporción de
xantofilas por Cromatografía de Líquidos de Alta Resolución (HPLC). Laboratorio de
Biogeoquímica. Facultad de Estudios Superiores Iztacala. Universidad Nacional Autónoma
de México
Olmedilla, B.A., Granado L.F., Blanco N.I. (2001) Carotenoides y salud humana. “Serie
Informes”. Fundación Española de la Nutrición. Madrid. 57 pp.
Liaaen-Jensen, S., (1979) Carotenoids- A chemosystematic approach. Pure Appl. Chem. 51:
661-675.
Bhosale, P. (2004) Environmental and cultural stimulants in the production of carotenoids
from microorganisms. Appl. Microbiol. Biotechnol. 63: 351–361.
Bishop, N., (2006) The β,ε-carotenoid, lutein, is specifically required for the formation of
the oligomeric forms of the light harvesting complex in the green alga, Scenedesmus
obliquus. Journal of Photochemistry and Photobiology B: Biology. 36: 279-283
Boussiba S. , Fan L., Vonshak A. (1992) Enhancement and determination of astaxanthin
accumulation in green alga Haematococcus pluvialis. Method. Enzymol. 213: 386-391.
Breithaupt, D.E. (2007) Modern application of xanthophylls in animal feeding- a review.
Trends Food Sci. Technol. 18: 501-506.
Britton G (1988) Biosynthesis of carotenoids. en Plant Pigments. London: Academic Press
Ltd. 133–182.

51
Cardozo, H.M., Guraratini, K.T., Barros M.P., et al. (2007) Metabolites from algae with
economical impact. Comparative Biochem. Physiol., 146: 60–78.
Eonseon, J.P., Kum L.J., Hong, M.H., Sang, Ch.M. (2003) Xanthophylls in Microalgae:
From Biosynthesis to Biotechnological Mass Production and Application. J. Microbiol.
Biotechnol. 13(2): 165–174.
Gildstat, M., Johnson, G., Sakshaud, E., (1993) Photosynthetic parameters, pigment
composition and respiration rates of the marine diatom Skeletonema costatum grown in
continuous light and a 12:12 h light-dark cycle. J. Plankton Res. 15: 939–951.
Kaftan, D., Meszaros, T., Whitmarsh, J., Nedbal, L., (1999) Characterization of
Photosystem II Activity and Heterogeneity during the Cell Cycle of the Green Alga
Scenedesmus quadricauda, Plant Physiol. 120: 433–441
Luteína, pigmento garantía de calidad para pollos (2001) Agro 2000. No. 31.
Margalith, P., (1999) Production of ketocarotenoids by microalgae. Appl Microbiol
Biotechnol. 51: 431-438
Meléndez-Martínez, A.J., Vicario, I.M., Heredia, F.J., (2004) Importancia nutricional de los
pigmentos carotenoides. Archivos latinoamericanos de la nutrición. 54(2): 149-154.
Mcghie T., Gary A. D., Color in fruit of the genus Actinidia: Carotenoid and chlorophyll
compositions(2002) . 50(1): 117-121
Nielsen, M. V. (1997) Growth, dark respiration and photosynthetic parameters of the
coccolithophorid Emiliania huxleyi (Prymnesiophyceae) acclimated to different daylength–
irradiance combinations. J. Phycol. 33: 818–822

52
Perales-Vela, H.V., González-Moreno, S., Montes-Horcasitas, C., Cañizares-Villanueva,
R.O. (2007) Growth, photosynthetic and respiratory responses to sub-lethal copper
concentrations in Scenedesmus incrassatulus (Chlorophyceae) Chemosphere. 67: 2274–
2281.
Pfander, H. (1992) Carotenoids: an overview. Meth.Enzymol. Academic Press. 213: 3-13.
Pokniak, J.R.; Bravo, G.I.. (2000) El color es obligatorio en los salmónidos. Tecno Vet:
Año 6(2).
Rost, B., Riebesell, U., Sultëmeyer, D., (2006) Carbon acquisition of marine
phytoplankton: Effect of photoperiod length. Limnol. Oceanogr. 51(1): 12–20.
Sandmann, G. (2000) Carotenoid Biosynthesis and Biotechnological Application, Archiv.
Biochem.Biophys. 385: 4–12.
Tomaselli, L. (2004) The Microalgal Cell. En: Amos Richmond (ed) Handbook of
Microalgal Culture. Blackwell science Ltd. pp. 3-17.
Schwender, J.S., Lichtenthaler, M., Hartmut K. y Rohmer, M. (1996) Biosynthesis of
isoprenoids (carotenoids, sterols, prenyl side-chains of chlorophylls and plastoquinone) via
a novel pyruvate/glyceraldehyde 3-phosphate non-mevalonate pathway in the green alga
Scenedesmus obliquus, Biochem. J. 316: 73-80.
Shan Qin, Guo-Xiang Liu, Zheng-Yu Hu (2008) The Accumulation and Metabolism of
astaxanthin in Scenedesmus obliquus (Chlorophyceae). Process Biochem. 43: 795-802.
Soeder, C.J., y Hegewald, E., (1988) Scenedesmus, Micro-algal biotechnology, Cambridge
University Press. 59-84 pp.

53
Umeno Daisuke,Alexander, V. Tobias & Arnold, Frances H. (2005) Diversifying
Carotenoid Biosynthetic Pathways by Directed Evolution. Microbiology and Molecular
Biology Reviews. 69: 51–78.
Vega-Estrada J., Montes-Horcasitas M.C., Domínguez-Bocanegra A.R., Cañizares-
Villanueva R.O. (2005) Haematococcus pluvialis cultivation in split-cylinder internal-loop
airlift photobioreactor under aeration conditions avoiding cell damage. Appl Microbiol
Biotechnol. 68: 31–35.
Vega, M. (2006) Caracterización fisiológica de Scenedesmus incrassatulus durante la
síntesis de pigmentos. Tesis de Licenciatura. Universidad Nacional Autónoma de México.
Facultad de Estudios Superiores Iztacala. Departamento de Ciencias. México D. F.
Wellburn, A.R. (1994) The spectral determination of chlorophylls a and b, as well as total
caroenoids, using various solvents with spectrophtometers of different resolution. Plant
Physiol. 144: 307-313.
Wiltshire, K.H., Boersma, M.M., Buhtz, A.H. (2000) Extraction of pigments and fatty acids
from the green alga Scenedesmus obliquus (Chlorophyceae). Aquat. Ecol. 34: 119–126.
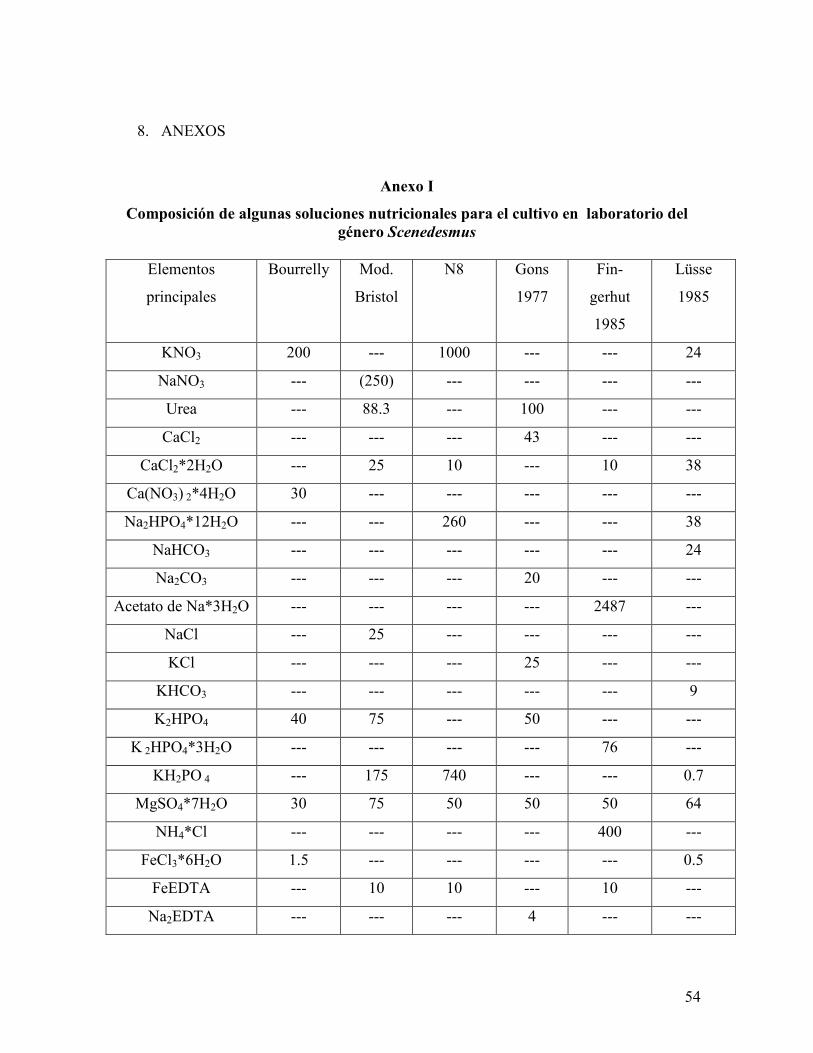
54
8. ANEXOS
Anexo I
Composición de algunas soluciones nutricionales para el cultivo en laboratorio del
género Scenedesmus
Elementos
principales
Bourrelly Mod.
Bristol
N8 Gons
1977
Fin-
gerhut
1985
Lüsse
1985
KNO3 200 --- 1000 --- --- 24
NaNO3 --- (250) --- --- --- ---
Urea --- 88.3 --- 100 --- ---
CaCl2 --- --- --- 43 --- ---
CaCl2*2H2O --- 25 10 --- 10 38
Ca(NO3) 2*4H2O 30 --- --- --- --- ---
Na2HPO4*12H2O --- --- 260 --- --- 38
NaHCO3 --- --- --- --- --- 24
Na2CO3 --- --- --- 20 --- ---
Acetato de Na*3H2O --- --- --- --- 2487 ---
NaCl --- 25 --- --- --- ---
KCl --- --- --- 25 --- ---
KHCO3 --- --- --- --- --- 9
K2HPO4 40 75 --- 50 --- ---
K 2HPO4*3H2O --- --- --- --- 76 ---
KH2PO 4 --- 175 740 --- --- 0.7
MgSO4*7H2O 30 75 50 50 50 64
NH4*Cl --- --- --- --- 400 ---
FeCl3*6H2O 1.5 --- --- --- --- 0.5
FeEDTA --- 10 10 --- 10 ---
Na2EDTA --- --- --- 4 --- ---
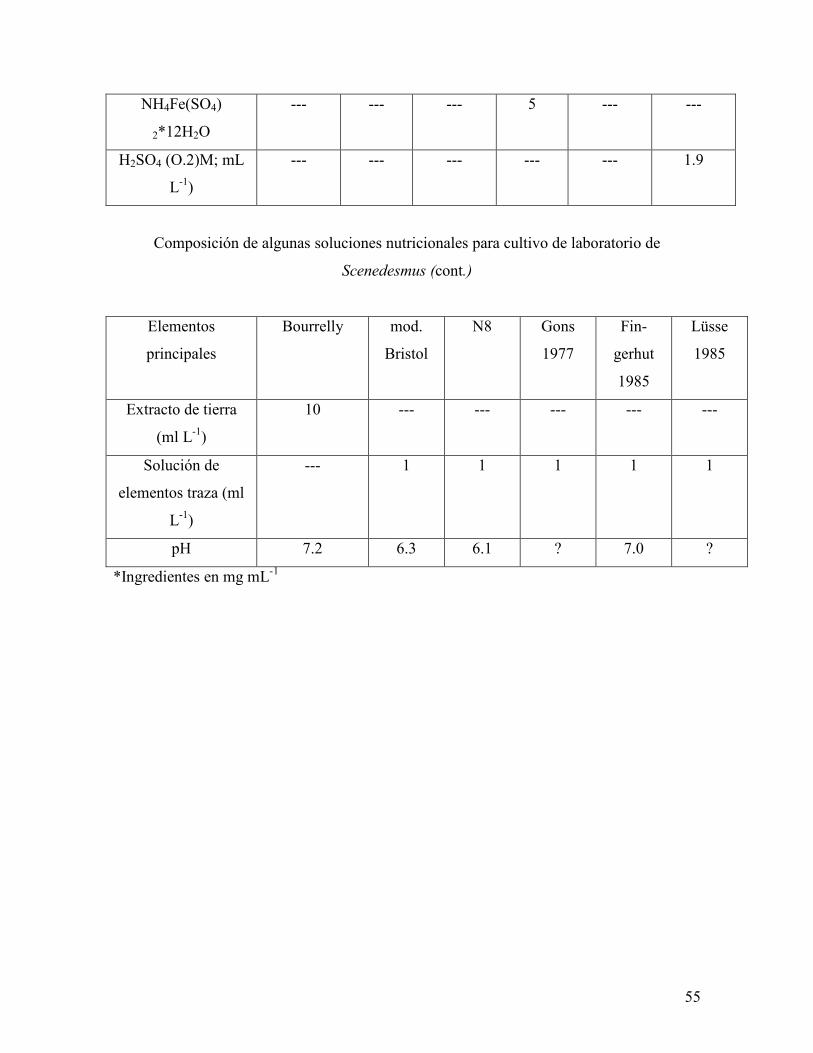
55
NH4Fe(SO4)
2*12H2O
--- --- --- 5 --- ---
H2SO4 (O.2)M; mL
L-1)
--- --- --- --- --- 1.9
Composición de algunas soluciones nutricionales para cultivo de laboratorio de
Scenedesmus (cont.)
Elementos
principales
Bourrelly mod.
Bristol
N8 Gons
1977
Fin-
gerhut
1985
Lüsse
1985
Extracto de tierra
(ml L-1)
10 --- --- --- --- ---
Solución de
elementos traza (ml
L-1)
--- 1 1 1 1 1
pH 7.2 6.3 6.1 ? 7.0 ?
*Ingredientes en mg mL-1
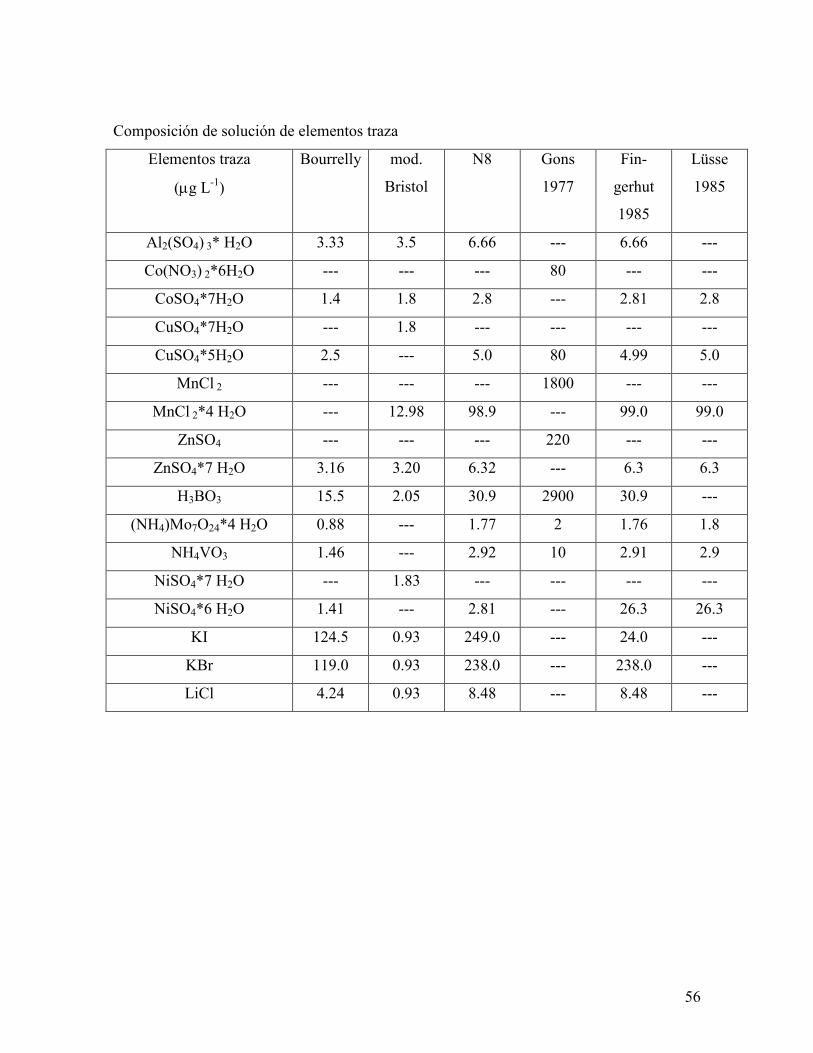
56
Composición de solución de elementos traza
Elementos traza
(µg L-1)
Bourrelly mod.
Bristol
N8 Gons
1977
Fin-
gerhut
1985
Lüsse
1985
Al2(SO4) 3* H2O 3.33 3.5 6.66 --- 6.66 ---
Co(NO3) 2*6H2O --- --- --- 80 --- ---
CoSO4*7H2O 1.4 1.8 2.8 --- 2.81 2.8
CuSO4*7H2O --- 1.8 --- --- --- ---
CuSO4*5H2O 2.5 --- 5.0 80 4.99 5.0
MnCl 2 --- --- --- 1800 --- ---
MnCl 2*4 H2O --- 12.98 98.9 --- 99.0 99.0
ZnSO4 --- --- --- 220 --- ---
ZnSO4*7 H2O 3.16 3.20 6.32 --- 6.3 6.3
H3BO3 15.5 2.05 30.9 2900 30.9 ---
(NH4)Mo7O24*4 H2O 0.88 --- 1.77 2 1.76 1.8
NH4VO3 1.46 --- 2.92 10 2.91 2.9
NiSO4*7 H2O --- 1.83 --- --- --- ---
NiSO4*6 H2O 1.41 --- 2.81 --- 26.3 26.3
KI 124.5 0.93 249.0 --- 24.0 ---
KBr 119.0 0.93 238.0 --- 238.0 ---
LiCl 4.24 0.93 8.48 --- 8.48 ---
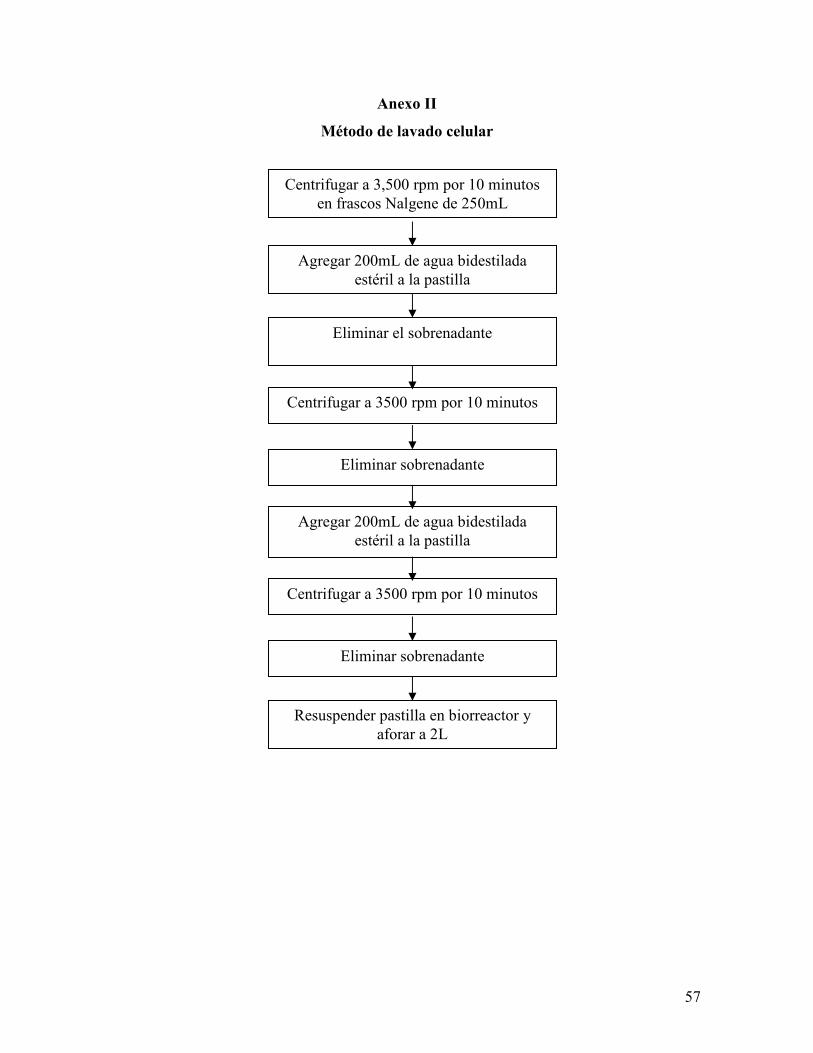
57
Anexo II
Método de lavado celular
Centrifugar a 3,500 rpm por 10 minutos en frascos Nalgene de 250mL
Eliminar el sobrenadante
Agregar 200mL de agua bidestilada estéril a la pastilla
Centrifugar a 3500 rpm por 10 minutos
Eliminar sobrenadante
Agregar 200mL de agua bidestilada estéril a la pastilla
Centrifugar a 3500 rpm por 10 minutos
Eliminar sobrenadante
Resuspender pastilla en biorreactor y aforar a 2L
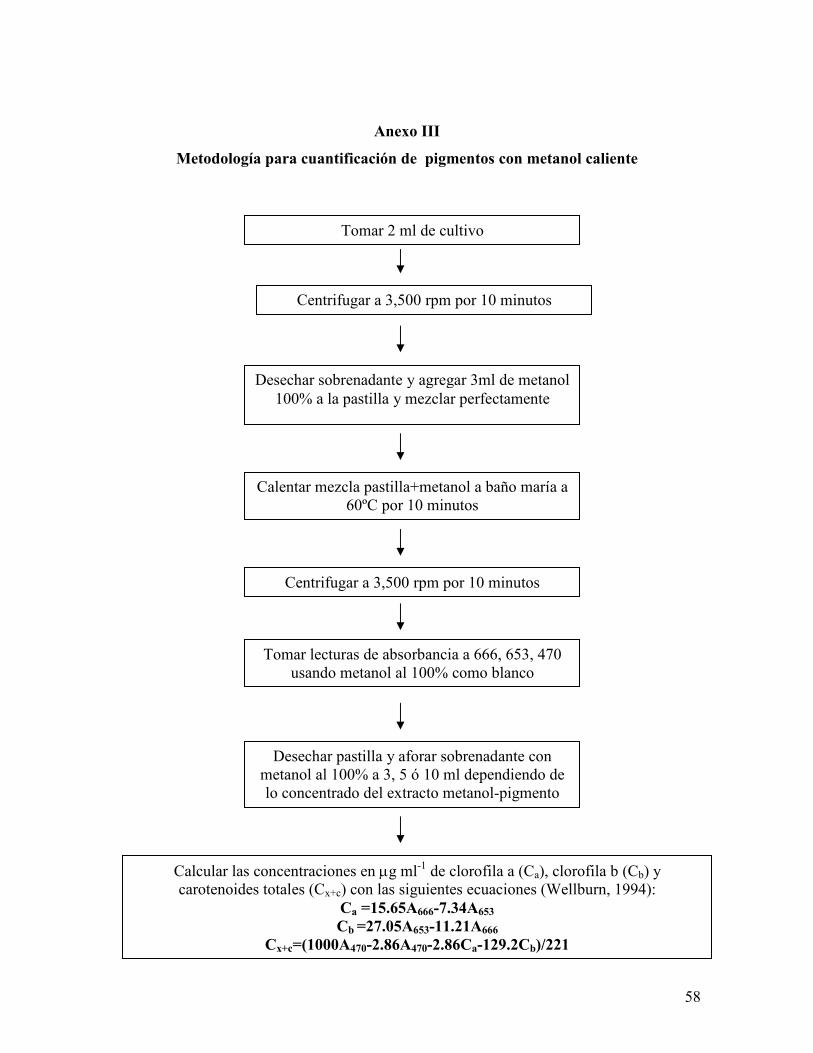
58
Anexo III
Metodología para cuantificación de pigmentos con metanol caliente
Tomar 2 ml de cultivo
Centrifugar a 3,500 rpm por 10 minutos
Desechar sobrenadante y agregar 3ml de metanol 100% a la pastilla y mezclar perfectamente
Calentar mezcla pastilla+metanol a baño maría a 60ºC por 10 minutos
Centrifugar a 3,500 rpm por 10 minutos
Desechar pastilla y aforar sobrenadante con metanol al 100% a 3, 5 ó 10 ml dependiendo de lo concentrado del extracto metanol-pigmento
Tomar lecturas de absorbancia a 666, 653, 470 usando metanol al 100% como blanco
Calcular las concentraciones en µg ml-1 de clorofila a (Ca), clorofila b (Cb) y carotenoides totales (Cx+c) con las siguientes ecuaciones (Wellburn, 1994):
Ca =15.65A666-7.34A653
Cb =27.05A653-11.21A666
Cx+c=(1000A470-2.86A470-2.86Ca-129.2Cb)/221
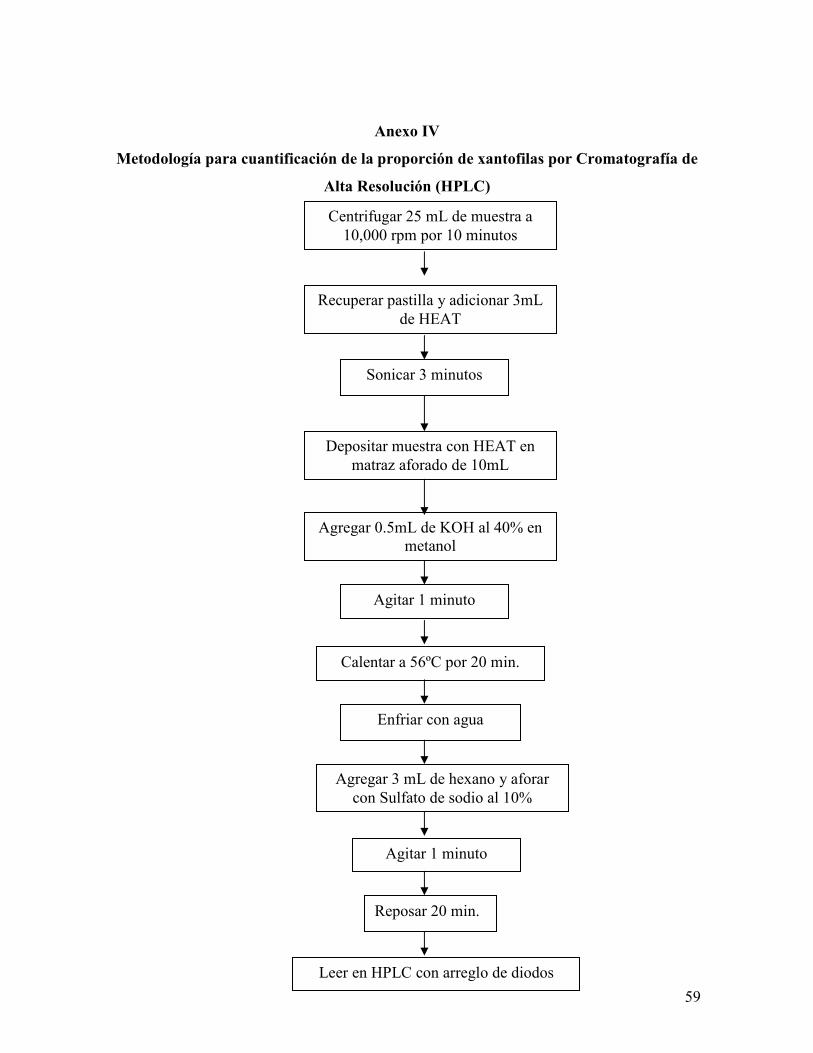
59
Anexo IV
Metodología para cuantificación de la proporción de xantofilas por Cromatografía de
Alta Resolución (HPLC)
Sonicar 3 minutos
Depositar muestra con HEAT en matraz aforado de 10mL
Centrifugar 25 mL de muestra a 10,000 rpm por 10 minutos
Recuperar pastilla y adicionar 3mL de HEAT
Agregar 0.5mL de KOH al 40% en metanol
Agitar 1 minuto
Calentar a 56ºC por 20 min.
Enfriar con agua
Agregar 3 mL de hexano y aforar con Sulfato de sodio al 10%
Agitar 1 minuto
Reposar 20 min.
Leer en HPLC con arreglo de diodos

60
Soluciones
Disolvente A: Acetonitrilo: Metanol: Tris-HCl 0.1 M pH 7.8(72:8:3)
Disolvente B: Metanol: Hexano (40:10)
Disolución extractora: HEAT (Hexano: Etanol: Acetona: Tolueno; 10: 6: 7 :7)
Disolución acuosa de sulfato de sodio al 10%
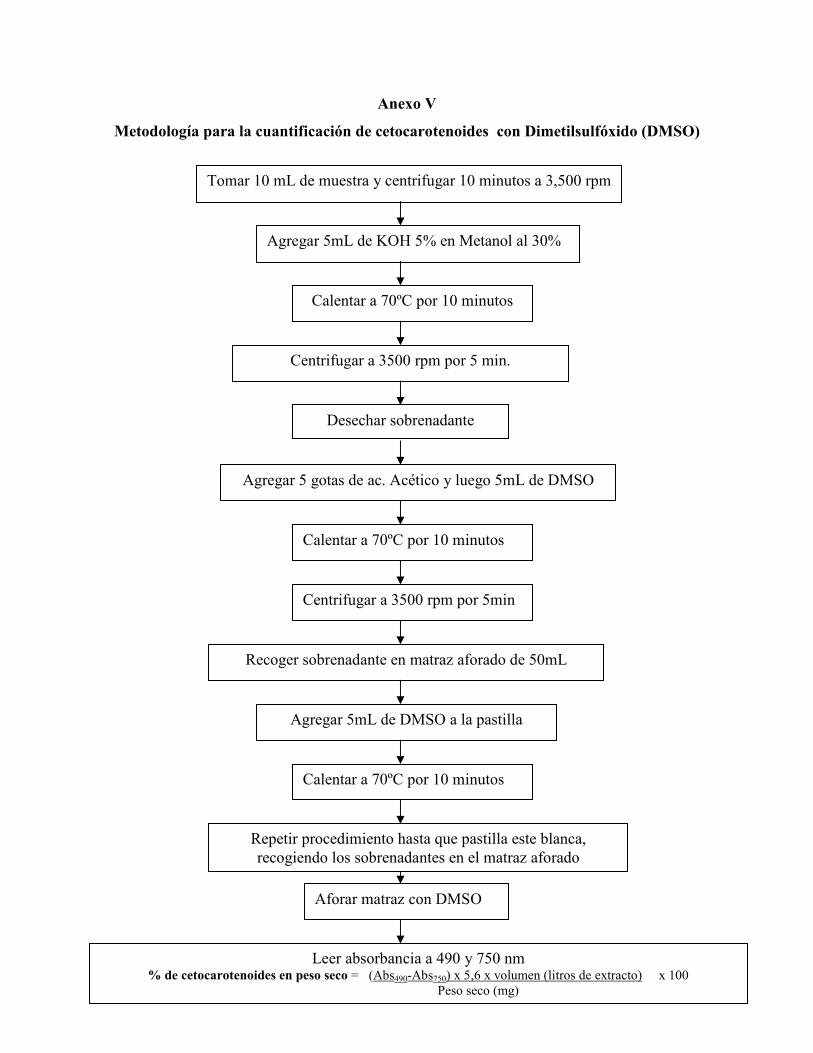
61
Anexo V
Metodología para la cuantificación de cetocarotenoides con Dimetilsulfóxido (DMSO)
Tomar 10 mL de muestra y centrifugar 10 minutos a 3,500 rpm
Agregar 5mL de KOH 5% en Metanol al 30%
Calentar a 70ºC por 10 minutos
Centrifugar a 3500 rpm por 5 min.
Desechar sobrenadante
Agregar 5 gotas de ac. Acético y luego 5mL de DMSO
Centrifugar a 3500 rpm por 5min
Recoger sobrenadante en matraz aforado de 50mL
Agregar 5mL de DMSO a la pastilla
Calentar a 70ºC por 10 minutos
Calentar a 70ºC por 10 minutos
Repetir procedimiento hasta que pastilla este blanca, recogiendo los sobrenadantes en el matraz aforado
Aforar matraz con DMSO
Leer absorbancia a 490 y 750 nm % de cetocarotenoides en peso seco = (Abs490-Abs750) x 5,6 x volumen (litros de extracto) x 100
Peso seco (mg)

62
Anexo VI
Fotografía de los fotobiorreactores
Fig 1. Fotografías de los fotobiorreactores con luz continua al día 4 (a) y fotoperiodo al día 5 (b) de fase de crecimiento
Fig 2. Fotografías de los fotobiorreactores con LC con estrés nutricional (día 40) y con
estrés nutricional y lumínico (día 25).
(a) (b)
(a)
(b)

63
Fig 3. Fotografías de los fotobiorreactores con luz continua sin inducción (izquierda) y
fotoperiodo sin inducción (derecha) a 40 días de inoculación
Fig 4. Fotografías de los fotobiorreactores con FP con estrés nutricional (a), con estrés lumínico (b) y con estrés nutricional y lumínico (c).
(a)
(b)
(c)
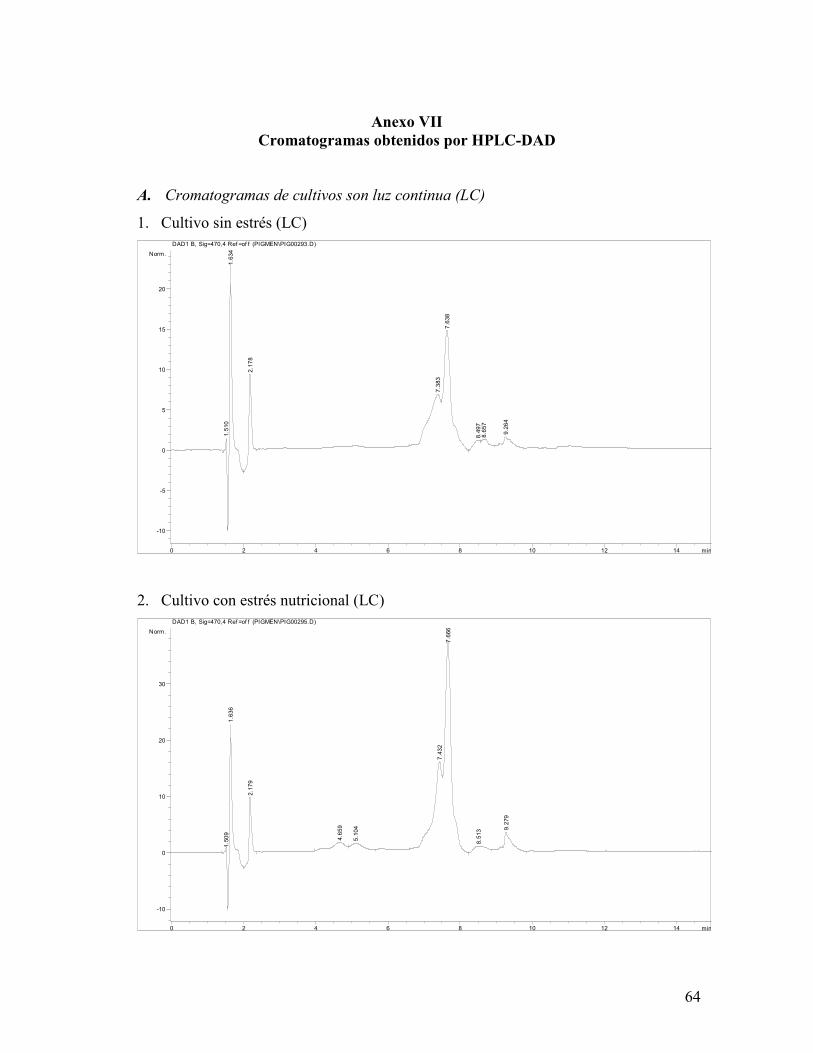
64
Anexo VII
Cromatogramas obtenidos por HPLC-DAD
A. Cromatogramas de cultivos son luz continua (LC)
1. Cultivo sin estrés (LC)
min0 2 4 6 8 10 12 14
Norm.
-10
-5
0
5
10
15
20
DAD1 B, Sig=470,4 Ref =of f (PIGMEN\PIG00293.D)
1.510
1.634
2.178
7.383
7.638
8.497
8.657
9.264
2. Cultivo con estrés nutricional (LC)
min0 2 4 6 8 10 12 14
Norm.
-10
0
10
20
30
DAD1 B, Sig=470,4 Ref =of f (PIGMEN\PIG00295.D)
1.509
1.636
2.179
4.659
5.104
7.432
7.666
8.513 9.279
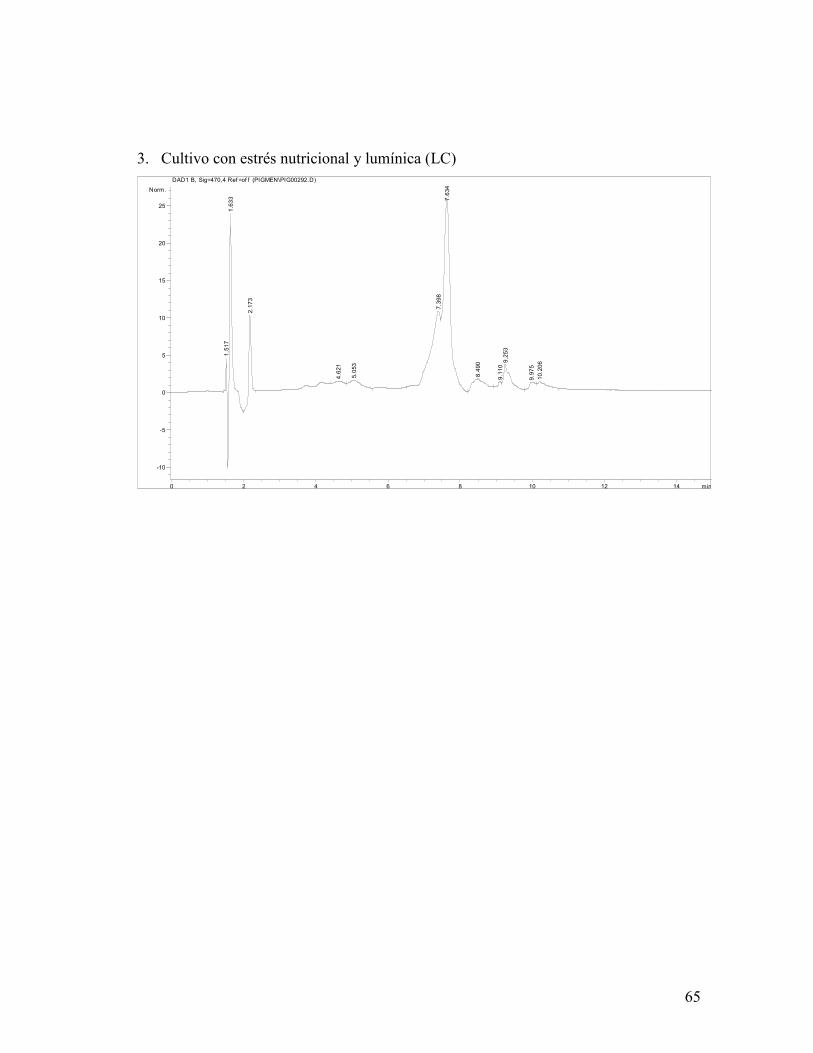
65
3. Cultivo con estrés nutricional y lumínica (LC)
min0 2 4 6 8 10 12 14
Norm.
-10
-5
0
5
10
15
20
25
DAD1 B, Sig=470,4 Ref =of f (PIGMEN\PIG00292.D)
1.517
1.633
2.173
4.621
5.053
7.398
7.634
8.490
9.110 9.253
9.975
10.206
66
B. Cromatogramas de cultivos con fotoperiodo (FP)
1. Cultivo sin estrés (FP)
min0 2 4 6 8 10 12 14
Norm.
-10
0
10
20
30
40
50
60
DAD1 B, Sig=470,4 Ref =of f (PIGMEN\PIG00294.D)
1.515
1.636
2.177
4.664
5.083
5.802
7.423
7.655
8.512
8.639 9.273
2. Cultivo con estrés nutricional (FP)
min0 2 4 6 8 10 12 14
Norm.
0
20
40
60
80
100
120
140
DAD1 A, Sig=474,4 Ref =of f (CINVES\XTC00005.D)
1.119
1.526 1.633
2.095 2.228
3.613
4.214
4.684
5.629
6.201
6.400
7.145
7.502
8.059
8.542
8.806
9.142
9.291
9.535
9.701
10.054
10.458
10.897
67
3. Cultivo con estrés lumínico (FP)
min0 2 4 6 8 10 12 14
Norm.
0
2
4
6
8
10
12
DAD1 A, Sig=474,4 Ref =of f (CINVES\XTC00007.D)
0.434
0.959
1.332 1.396
1.544 1.666
1.941
2.046
4.272
5.113
7.263
7.450
7.542
7.712
8.288
9.954
4. Cultivo con estrés nutricional y lumínico
min0 2 4 6 8 10 12 14
Norm.
0
1
2
3
4
5
6
DAD1 A, Sig=474,4 Ref =of f (CINVES\XTC00008.D)
1.334
1.396
1.547
1.671
2.050
4.301
4.999
7.212
7.663
8.337
9.908