occhio neuroanatomia
DESCRIPTION
Elementi di neuroanatomia dell'occhio arricchito di immagini e schemi.TRANSCRIPT

1
OCCHIO
CAVITÁ ORBITARIA
Le cavità orbitarie sono cavità ampie pari e simmetriche del cranio poste nella parte alta della faccia ai lati del naso, accolgono il bulbo oculare e gli annessi. Hanno forma simile a quella di una piramide quadrangolare, hanno una disposizione con assi che convergono posteriormente e medialmente in corrispondenza della sella turcica dell’osso sfenoide, gli assi delle due orbite formano un angolo aperto di circa 45°. Non sono complanari ma leggermente ruotate verso l’esterno. All’apice della piramide a cui si può assimilare la cavità orbitaria c’è il foro ottico da cui emerge il nervo ottico, alcune pareti non sono direttamente a contatto ma ci sono delle discontinuità, cioè delle aperture allungate in prossimità degli spigoli, quindi ci sono due angoli suturali e due angoli che presentano aperture,: fessura orbitaria superiore (tra grande e piccola ala dello sfenoide) che immette nel foro che immette nel canale ottico) e tra parete laterale e pavimento la fessura orbitaria inferiore compresa tra grande ala dello sfenoide e osso mascellare. Attraverso la fessura orbitaria superiore la cavità orbitaria comunica con la cavità cranica, attraverso la fessura orbitaria inferiore comunica con la fossa infratemporale e pterigomascellare o pterigopalatina. In corrispondenza della parete superiore fossa lacrimale che accoglie la ghiandola lacrimale e più medialmente è presente la spina trocleare o troclea che forma un anello con il contributo di tessuto connettivo all’interno del quale passa il tendine del muscolo obliquo superiore innervato dal nervo trocleare 4° paio dei nervi encefalici. Nella parete mediale tra la cresta lacrimale anteriore e posteriore c’è la fossa lacrimale che contiene il sacco lacrimale che accoglie le lacrime che verranno convogliate nel canale naso lacrimale. Nella parete inferiore c’è un forellino che fa seguito alla fossa del sacco lacrimale: è l’inizio del canale naso lacrimale che si apre in corrispondenza delle cavità nasali.
La fessura orbitaria superiore è attraversata da tutte le strutture che raggiungono o che escono dalla cavità orbitaria, ad eccezione dell’arteria oftalmica e del nervo ottico che decorrono nel canale ottico. l III, il IV ed il VI paio di nervi cranici (rispettivamente oculomotore comune, trocleare ed abducente), la vena oftalmica e la prima branca del nervo trigemino (V paio di nervi cranici), ovvero il nervo oftalmico.
La cavità orbitaria oltre a contenere il bulbo oculare contiene anche altre strutture importanti come:
- Corpo adiposo: è posto attorno al bulbo oculare e alle strutture che ad esso afferiscono. Riempie tutti gli spazi compresi tra le varie strutture, è sensibile agli ormoni tiroidei infatti in caso di ipertiroidismo il suo volume aumenta notevolmente esoftalmo.
- Capsula di Tenone: è un involucro connettivale che avvolge in parte il bulbo oculare e altre strutture annesse al bulbo come parte del nervo ottico. Si estende dalla guaina durale del nervo ottico fino all’orlo sclero-corneale, ossia fino al punto di confine tra sclera e cornea. La dura madre si estende fino a ricoprire il nervo ottico perché il bulbo oculare con il nervo ottico non è altro che un’estroflessione del diencefalo verso l’esterno, al livello del tratto iniziale del nervo ottico ritroviamo anche il liquor contenuto nello spazio sub aracnoideo. Alle volte è possibile accusare un edema della papilla, cioè un difficoltoso ristagno del sangue venoso al livello della

2
papilla del nervo ottico che causa u rialzamento della papilla stessa e ciò può essere causato da un’aumentata ipertensione endocranica. La capsula di Tenone aderisce al bulbo oculare solo al livello della sua porzione anteriore, cioè nel punto di passaggio tra sclera e cornea e al livello del nervo ottico, per il resto risulta libera. In corrispondenza dell’equatore del bulbo oculare la capsula di Tenone fornisce il passaggio ai tendini dei muscoli che si inseriscono sul bulbo oculare.
Muscoli estrinseci dell’occhio: sono muscoli striati di derivazione somitica, gli unici della testa insieme a quelli della lingua, gli altri hanno origine branchiale. Sono 7 di cui 4 sono retti, 2 sono obliqui, più l’elevatore della palpebra superiore.
- Retti: si distinguono in superiore, mediale, inferiore e laterale. Originano con un tendine comune a forma anulare, detto anello tendineo di Zinn, posto nel fondo della cavità orbitaria dove circoscrive il foro ottico e l’estremità mediale della fessura orbitaria superiore. I muscoli retti si inseriscono sulla sclera all’altezza dell’equatore e si dipartono ognuna dalla omonima porzione dell’anello tendineo. L’anello tendineo comune di Zinn Lo spazio da esso circoscritto viene diviso in due parti da un setto fibroso: la parte supero-mediale, corrispondente al foro ottico, dà passaggio al nervo ottico e all’arteria oftalmica prima diramazione intradurale dell'arteria carotide interna; la parte infero-laterale, in rapporto con l’estremità mediale della fessura orbitaria superiore, dà passaggio ai nervi oculomotore, nasociliare, abducente e alla vena oftalmica superiore. In corrispondenza della linea di inserzione sull’osso, l’anello tendineo prende rapporto con la guaina durale che avvolge il nervo ottico. (Il nervo naso-ciliare origina dal nervo oftalmico in corrispondenza della fessura orbitale superiore dello sfenoide, Il nervo oftalmico è un nervo sensitivo e rappresenta la branca di minore dimensione del nervo trigemino. Emerge dal margine anteriore convesso del ganglio semilunare del Gasser, in posizione mediale),
- Obliqui: si distinguono in superiore e inferiore. Il muscolo obliquo superiore prende origine dal contorno mediale del foro ottico, si porta in avanti, segue la parete mediale della cavità orbitaria e con il suo tendine si impegna nella troclea (struttura fibrocartilaginea che si inserisce nella fossetta trocleare dando luogo a una struttura simile a una puleggia), poi il muscolo si porta lateralmente per inserirsi sul quadrante supero-laterale della sclera. L’obliquo inferiore origina invece dall’osso mascellare, subito sotto la fossa del sacco lacrimale, si porta lateralmente passando sotto al retto inferiore e si inserisce sul quadrante infero-laterale della sclera, al di dietro dell’equatore.
- Muscolo elevatore della palpebra superiore nasce dall’anello tendineo comune e si porta alla palpebra
L’obliquo superiore n. trocleare 4°, il retto laterale n. abducente 6°. Retto mediale, inferiore, superiore, obliquo inferiore, elevatore della palpebra superiore n. oculomotore 3°.
- Congiuntiva o sacco congiuntivale è una membrana connettivale piuttosto sottile che riveste la faccia interna della palpebra e che da qui portandosi indietro si riflette sulla sclera del bulbo, una volta sulla sclera, la congiuntiva si porta in avanti terminando laddove la sclera si continua con la cornea: ciò ci

3
permette di distinguere una congiuntiva palpebrale e una congiuntiva del bulbo oculare. Il punto di riflessione della congiuntiva prende il nome di fornice congiuntivale. È la congiuntiva è normalmente vascolarizzata, nal caso di una congiuntivite,, si ha una congiuntiva iperemica. La congiuntiva quindi non deve rivestire la cornea perché essa non è trasparente in quanto ben vascolarizzata.
BULBO OCULARE
La sua forma non è perfettamente sferica, è costituito da due segmenti di sfera affrontati con diverso raggio di curvatura: si distingue un segmento anteriore e un segmento posteriore corrispondenti rispettivamente dalla cornea e alla sclera. La cornea corrisponde a 1/6 anteriore del bulbo oculare, è quel segmento con minor raggio di curvatura, quindi è più convessa rispetto alla sclera che costituendo 5/6 posteriori ha un raggio di curvatura maggiore. Nel bulbo oculare in senso antero-posteriore si riconoscono
due assi:
- Asse anatomico (24-25mm) linea che congiunge il polo anteriore al polo posteriore del bulbo)
- Asse visivo o visuale: è l’asse percorso dal raggio luminoso, segue il raggio luminoso dal momento in cui proviene dall’esterno e attraversa tutti i mezzi diottrici (dall’av. al dietro cornea, umor acqueo, cristallino e corpo vitreo) per andare al centro della corna, nella cosiddetta fovea centrali (piccola depressione della retina in cui si ha la massima discriminazione visiva a disposizione,.
- Equatore del bulbo: punto posto ortogonalmente all’asse anatomica del bulbo, qui distante dal polo anteriore e posteriore a questa altezza arrivano i muscoli retti (superiore, mediale, inferiore e laterale)
Dal bulbo oculare distinguiamo tre tonache che dall’esterno verso l’interno sono
- Tonaca fibrosa: rappresentata per 1/6 anteriore dalla cornea e per i 5/6 posteriori dalla sclera (parte dell’occhio che cambia colore in persone con ittero), cornea e sclera sono in continuità in corrispondenza del limbus o solco sclero-corneale.
- Tonaca vascolare o uvea (processo infiammatorio: uveite) si costituisce di tre parti che dall’avanti al dietro sono iride, corpo ciliare e coroidea
- Tonaca nervosa o retina: distinta a sua volta in retina visiva in cui sono presenti i fotorecettori e retina cieca a sua volta distinguibile in retina cieca ciliare (porzione di retina cieca in rapporto con il corpo ciliare) e in una retina cieca iridea, porzione di retina cieca che riveste la porzione posteriore dell’iride.

4
Nel bulbo distinguiamo oltre alle tre tonache: fibrosa, vascolare e nervosa, anche
- Cristallino: unico mezzo diottrico del bulbo di cui può essere modificato l’indice di rifrazione modificando la sua forma tramite l’azione del muscolo ciliare.
- Camera anteriore: spazio compreso tra faccia posteriore della cornea e faccia anteriore dell’iride. Occupata dall’umor acqueo.
- Camera posteriore: spazio compreso tra iride, corpo ciliare e cristallino, anch’essa è ripiena di umor acqueo. L’umor acqueo viene prodotto al livello della camera posteriore, attraversa la pupilla, cioè il margine dell’iride e raggiungerà la camera anteriore dove viene riassorbito. Il corpo ciliare ha anche una componente muscolare data dal muscolo ciliare, muscolatura intrinseca, si occupa di modificare la forma del cristallino per la messa a fuoco di oggetti vicini al di sotto dei 10metri. Esiste anche un muscolo sfinteriale capace di restringere la pupilla sotto controllo parasimpatico, il costrittore della pupilla al livello dell’iride, e un dilatatore della pupilla sotto controllo ortosimpatico.
- Camera vitreale: spazio occupato dal corpo vitreo.
TONACA FIBROSA:

5
CORNEA: rappresenta il 1/6 anteriore della tonaca fibrosa del bulbo oculare, essa è anteriormente è a contatto con l’ambiente esterno mediante la mediazione del liquido lacrimale. È priva di vascolarizzazione perché deve essere trasparente, è infatti il primo mezzo diottrico incontrato dal raggio luminoso, durante la vita embrionale risulta vascolarizzata così come altre strutture quali i cristallino e il corpo vitreo, ma mai al termine dello sviluppo. La cornea non corrisponde esattamente a una sezione di sfera, infatti essa presenta uno sviluppo orizzontale (in media 12mm) leggermente maggiore rispetto a quello verticale (in media 11mm). La faccia anteriore della cornea presenta un raggio di curvatura maggiore rispetto a quello della faccia posteriore, quindi nella parte centrale la cornea è più sottile. Lo spessore medio della cornea nell’uomo è di circa 1mm, ma al centro diminuisce a 0,8mm, mentre nella parte più laterale arriva anche ad 1,1mm.
La cornea umana è costituita da 5 strati che dall’esterno verso l’interno sono:
1) Epitelio corneale Epitelio pavimentoso composto non cheratinizzato (per non inficiare sulla trasparenza), è costituito da 5-6 strati di cellule, deve essere resistente a stress meccanici, di cui il più profondo di questi 5-6 strati è costituito da cellule basali fondamentali per il rinnovamento dell’epitelio stesso. Le cellule degli strati intermedi sono dette cellule alari o wing cells. Al livello dell’orlo sclero-corneale tale epitelio si continua con quello della congiuntiva, questa continuità anatomica tra i due epiteli non rappresenta un problema, in quanto gli epiteli non sono vascolarizzati. Il film lacrimale protegge, lubrifica e nutre l’epitelio corneale, quindi ha anche funzione trofica. Le cellule dello strato più apicale presentano dei corti microvilli necessari per ostacolare la fisiologica discesa per gravità del film lacrimale. L’epitelio cornale è innervato moltissimo dalla branca oftalmica del trigemino, in particolare dal nervo naso-ciliare.
2) Membrana limitante esterna o di Bowmann o elastica anteriore, corrisponde alla lamina basale dell’epitelio esterno. 12micron
3) Stroma corneale: è la parte preponderante della cornea, ossia l’80-90% di essa con spessore di circa 500micron. È costituito da un connettivo le cui fibre collagene e la matrice mucopolisaccaridica extracellulare hanno lo stresso indice di rifrazione, il raggio luminoso deve attraversare questa struttura e uscire integro per andare nella fovea centrale. Le fibre e la matrice si dispongono a formare circa 50 lamelle sovrapposte, in ognuna di queste lamelle le fibre si dispongono parallelamente ed in modo equidistante tra loto. Inoltre le fibre di una lamella sono disposte ortogonalmente a quelle della lamella antistante e retrostante con questa particolare struttura il raggio luminoso che attraversa lo stroma corneale non subisce alcuna deviazione.
4) Membrana limitante interna o di Descemet: corrisponde alla membrana basale dell’endotelio corneale, ha spesso inferiore rispetto a quella della membrana di Bowmann. È costituita da sottili fibre collagene.
5) Endotelio corneale: è un epitelio semplice costituito da cellule con all’interne molti organuli citoplasmatici, esse dovranno garantire gli scambi con l’umor acqueo. Anche le cellule dell’endotelio corneale possiedono al livello della citomembrana rivolta all’umor dei corti microvilli con stessa funzione di quelli disposti più superficialmente nell’epitelio corneale. L’umor acque non è un semplice filtrato del sangue, le strutture che prendono contatto con il sangue effettuano uno scambio selettivo, sono le cellule della retina e i vasi provengono dalla corioidea della vascolare.
Umor acqueo è accolto nella camera anteriore dell’occhio, lo spazio posto tra cornea e iride, deve essere trasparente per permettere il passaggio del raggio luminoso ma anche avere funzione trofica per le strutture che non possono essere vascolarizzate, una di queste è la cornea ma anche i cristallino, una trama vascolare impedirebbe il passaggio del raggio

6
luminoso. L’epitelio a contatto con l’aria viene nutrito mediante il liquido lacrimale che però non potrà nutrire gli strati sottostanti, allora lo stroma corneale ad esempio viene nutrito per diffusione tramite l’endotelio, la mancanza di giunzioni serrate dell’endotelio permette il passaggio dell’umor allo stroma corneale, le giunzioni serrate dell’epitelio corneale impediscono il passaggio del liquido lacrimale allo stroma, quindi il film lacrimale non può passare all’interno. Lo stroma tende ad assorbire acqua, la presenza di giunzioni serrate tra cellule dell’epitelio corneale impedisce il passaggio di acqua dal liquido lacrimale allo stroma, mentre la mancanza di giunzioni serrate tra le cellule dell’endotelio cornale non impedisce il passaggio di acqua dall’umor acqueo allo stroma corneale.
SCLERA: rappresenta i 5/6 posteriori della tonaca fibrosa del bulbo oculare, ha colorito biancastro ed è costituita da tessuto connettivo fibroso denso. Su di essa si inseriscono i muscoli estrinseci dell’occhio (al livello dell’equatore i due retti, al livello del quadrante supero-laterale l’obliquo superiore e nel quadrante infero laterale l’obliquo inferiore.) È spessa circa 2mm e dà attacco ai muscoli estrinseci dell’occhio ad eccezione del muscolo elevatore della palpebra superiore. La sua faccia esterna è a contatto con la capsula di Tenone e con la congiuntiva bulbare, mentre la sua faccia interna è a contatto con una parte della corioidea, in particolare con l’epitelio pigmentato della sovra corioidea (lo strato più esterno della corioidea). Osservando al di dietro del bulbo oculare si nota al livello della sclera il foro di uscita delle fibre del nervo ottico, cioè gli assoni delle cellule gangliari della retina, questa porzione è detta anche lamina cribrosa della sclera. In mezzo al nervo ottico c’è l’arteria centrale della retina, ramo dell’arteria oftalmica, intorno ci sono i fori per il passaggio i vasi e nervi ciliari posteriori. Questo punto di fuoriuscita delle fibre corrisponde a quel punto in cui il nervo ottico da intrabulbare diventa intraorbitario. Intorno ai numerosi forellini della sclera che nel complesso danno luogo al foro di uscita del nervo ottico ritroviamo anche una serie di altri forellini che invece danno passaggio ai vasi e nervi ciliari posteriori, i vasi ciliari posteriori sono l’impalcatura vascolare della tonaca vascolare.
Anche il sistema dei vasi ciliari è alimentato dall’arteria oftalmica, mediante le arterie ciliari posteriori e anteriori. Le arterie ciliari posteriori, mediale e laterale, danno origine a un certo numero di rami brevi (arterie ciliari posteriori brevi) e a un solo ramo lungo (arteria ciliare posteriore lunga). Le arterie ciliari posteriori brevi (circa 20) perforano la sclera tutto intorno al nervo ottico e, con alcuni rami, nello spessore della sclera stessa, costituiscono un cerchio arterioso attorno alla parte intrabulbare del nervo (anello di Haller).
Al livello dell’equatore la sclera presenta i fori di passaggio per le 4 vene vorticose (superiore, laterale, inferiore e mediale), importanti per il drenaggio venoso del bulbo oculare, tributarie della vena oftalmica.
Più avanti ancora la sclera presenta i fori per il passaggio dei vasi ciliari anteriori, le arterie ciliari anteriori (derivano dalle arterie muscolari provenienti dall’oftalmica, Le arterie ciliari anteriori hanno origine dai rami muscolari dell’arteria oftalmica e, in numero di 6-8, raggiungono la sclera che perforano in prossimità dell’inserzione dei muscoli retti, dopo aver dato origine ai rami episclerali. Danno rami al corpo ciliare e contribuiscono a costituire il grande cerchio arterioso dell’iride. ) sono importanti per la vascolarizzazione del bulbo e anche dei muscoli estrinseci dell’occhio, soprattutto per parte del corpo ciliare e dell’iride.
Le arterie ciliari sia anteriori che posteriori provvedono alla vascolarizzazione della tonaca vascolare.

7
Il punto di passaggio tra sclera e cornea è detto limbus o orlo sclero-corneale, non ha un profilo verticale ma obliquo in modo che l’ultima parte della sclera sormonti una piccolissima parte della cornea, è detta regione limbale di passaggio di circa 1,5mm.
TONACA VASCOLARE o UVEA (somiglia ad un chicco d’uva nera al quale è stata tolta la buccia):
- Iride- Corpo ciliare- Corioidea

8
IRIDE: è l’ultima parte dell’uvea, costituisce la parte anteriore della tonaca vascolare ed è posta a totale contatto con l’umor acqueo. Al suo interno sono presenti gli altri due muscoli intrinseci del bulbo oculare: muscolo sfintere della pupilla innervato dal parasimpatico e il dilatatore della pupilla innervato dall’ortosimpatico. (esame all’occhio si usa atropina per impedire che colpito dalla luce la pupilla si restringa per l’azione del muscolo sfinteriale della pupilla). Muscolo sfinteriale e ciliare entrano in gioco per la messa a fuoco.
L’iride è un diaframma che presenta una faccia anteriore che delimita posteriormente la camera anteriore e una faccia posteriore che delimita anteriormente la camera
posteriore. L’iride ha un margine pupillare che delimita il foro pupillare il cui diametro può variare in base alla quantità di luce presente o dalla condizione emozionale dell’individuo, oltre al margine pupillare è presente anche un margine ciliare in rapporto con il corpo ciliare, vicino all’orlo sclero-corneale.
Faccia anteriore dell’iride: di colore variabile e aspetto irregolare per la presenza di fessurazione dette cripte, diverse per ognuno di noi. La zona di passaggio tra faccia anteriore e cornea corrisponde alla parte periferica della camera anteriore ed è detta angolo irido-corneale, qui ritroviamo uno dei due apparati di riassorbimento dell’umor acqueo. Faccia posteriore dell’iride: totalmente a contatto con la parte iridea della retina cieca e ciò le conferisce un aspetto caratteristico scruto per la presenza dell’epitelio pigmentato.
Struttura dell’iride:
- Endotelio: singolo strato di cellule appiattite che si continua con quello della cornea lungo l’angolo irido-corneale. Discontinuo per mediare gli scambi con l’umor.
- Stroma: è la parte centrale dell’iride, costituito da connettivo lasso quindi fibre collagene, fibrociti, cellule pigmentate e macrofagi. In corrispondenza del margine pupillare è presente un anello di fibrocellule muscolari lisce che costituisce il muscolo sfintere della pupilla (anteriore centrale e circolare) innervato anch’esso da fibre postgangliari parasimpatiche che si dipartono dal ganglio ciliare. Anche questo muscolo di contrarsi nel riflesso di accomodazione durante il quale viene cambiato forma del cristallino e viene diminuito il diametro del foro pupillare. Il restringimento del foro pupillare è importante per due motivi
o Minore è il calibro del foro pupillare e maggiore è la profondità del campo visivo
o Con il restringimento della pupilla si evita l’aberrazione cromatica, infatti quando la pupilla si restringe, il fascio luminoso in entrata è costretto maggiormente ad attraversare la porzione centrale del cristallino e non quella laterale, questo evita la scomposizione del fascio luminoso, scomposizione che quindi è maggiore quando un fascio luminoso attraversa la porzione laterale di una lente biconvessa.
- Epitelio pigmentato: è la parte iridea della retina, continuazione della parte ciliare, formata da due strati: uno interno rivolto alla camera posteriore molto pigmentato, uno

9
esterno al cui interno sono presenti cellule mio epiteliali che costituiscono nel complesso il muscolo dilatatore della pupilla (posteriore e disposto radialmente) innervato da fibre postgangliari ortosimpatiche che partono dal ganglio cervicale superiore e che giungono in scatola cranica decorrendo satelliti dell’arteria carotide interna (entra attraverso il foro carotico )contribuendo alla formazione del suo plesso. Tali fibre giungono in cavità cranica seguendo il nervo oftalmico, prima branca del trigemino. Si portano nella cavità orbitaria attraversando la fessura orbitaria superiore. Il ganglio cervicale riceve a sua volta fibre ortosimpatiche pregangliari che partono dalla parte superiore della colonna intermedio-laterale (lamina 7°)
- Muscolo sfintere della pupilla: il nucleo edinger- westphal (nucleo oculomotore accessorio) è il nucleo visceroeffettore parasimpatico del III paio di nervi encefalici (nervo oculomotore comune). Innerva attraverso il ganglio ciliare i muscoli costrittore della pupilla e ciliare.) è la sede dei neuroni parasimpatici pregangliari, le sue fibre seguono il nervo oculomotore e terminano nel ganglio ciliare. Le fibre parasimpatiche del ganglio ciliare raggiungono i muscoli ciliare e sfintere della pupilla tramite i nervi ciliari brevi.
- Muscolo dilatatore della pupilla: le fibre pregangliari ortosimpatiche che partono dalla parte craniale della colonna intermedio-laterale al livello di T1 e T2 ed entrano nella catena dell’ortosimpatico e si fanno ascendenti fino a raggiungere il ganglio cervicale superiore, posto in vicinanza della biforcazione della carotide comune, quindi è al livello della quarta vertebra cervicale. Dal ganglio, fibre postgangliari si accompagnano al plesso carotico interno, entrano nella cavità orbitaria tramite il nervo oftalmico. Le fibre decorrono poi nel nervo naso-ciliare, alcune attraversano il ganglio ciliare, ci transitano solo ed entrano nel bulbo tramite i nervi ciliari brevi, altre arrivano all’occhio tramite i nervi ciliari lunghi. Altre fibre postgangliari ortosimpatiche contribuiscono all’innervazione del muscolo elevatore della palpebra superiore.
CORIOIDEA rappresenta la porzione posteriore della tonaca vascolare, precisamente i 2/3 posteriore del bulbo, essi si spinge in avanti fino a un punto detto ora serrata della retina, è il punto di passaggio tra retine cieca e retina visiva. La corioidea si estende fino a dove è presente la retina visiva, infatti tra le due strutture vi è uno stretto rapporto funzionale. Il margine tra retina visiva e cieca è un margine festonato, sfrangiato.
La corioidea è altamente vascolarizzata, è costituita da 4 strati, i quali dall’esterno vero l’interno sono:
1) Lamina sovra corioidea o frusca: costituita da collagene, fibroblasti e cellule pigmentate dette melanociti con funzione sconosciuta.
2) Lamina vascolosa: è formata da vasi arteriosi, le arterie ciliari posteriori brevi e vene omonime. Nel suo ambito si riconoscono due strati sulla base del diametro dei vasi che li costituiscono, quello più esterno è detto strato di Haller ed è costituito da vasi con calibro maggiore, quello sottostante è detto strato di Sattler ed è costituito da vasi di calibro minore. Le arterie ciliari posteriori brevi danno rami per la corioidea, le ciliari posteriori lunghe danno rami per il corpo ciliare. La portata ematica della corioide è altissima.
3) Lamina corio-capillare: è costituita da un enorme e finissima capillarizzazione delle arterie ciliari posteriori brevi, è fondamentale per il trofismo dello strato dei fotorecettori della retina, trofismo mediato dall’epitelio pigmentato della retina. L’epitelio pigmentato della retina che svolge funzioni importanti si occupa della gestione dello strato dei recettori della retina fagocitando le parti recettoriali esaurite, è scuro quindi impedisce la riflessione dei raggi luminosi e in questo modo impedisce ad es. che i recettori adiacenti si stimolino a vicenda, è importante anche

10
per il trofismo per lo strato dei fotorecettori e per il riciclo dei pigmenti fotosensibili. Quindi il corio capillare è importante per il trofismo dello strato dei fotorecettori della retina perché al livello della fovea centralis la retina è costituita esclusivamente dallo strato dei recettori e siccome la fovea rappresenta il punto massimo di discriminazione visiva, certamente non può essere vascolarizzato in modo diretto, ma il suo trofismo deve avvenire per diffusione dalla corioidea. Al livello della fovea centralis il corio capillare ha uno spessore maggiore. I capillari per permettere scambi efficaci sono fenestrati e piuttosto grandi.
4) Membrana di Brunch o lamina vitrea: è costituita da connettivo, non ci sono cellule. È in rapporto diretto con l’epitelio pigmentato della retina, cioè lo strato più superficiale della retina e corrisponde a quello strato di questa ultima a diretto contatto con i fotorecettori. La membrana di brunch funge da membrana basale tra l’endotelio dei corio-capillare, è lo strato più esterno dei fotorecettori. È organizzata in 5 strati che dall’esterno verso l’interno sono: membrana basale del corio capillare, lamina esterna di collagene, strato di elastina, zona interna costituita da collagene e membrana basale per l’epitelio pigmentato della retina. Quindi la membrana di Brunch non è un unico strato connettivale. L’integrità della membrana di Brunch è fondamentale: una sua alterazione si traduce in un’alterazione dell’interfaccia tra il corio-capillare e l’epitelio pigmentato della retina, quindi comporta problemi alla diffusione di sostanze nutritive dalla corio-capillare all’epitelio pigmentato della retina e da qui allo strato dei recettori. 1) La membrana del Brunch è una membrana semipermeabile che permette il passaggio di molecole nutritive, ha anche 2) una funzione di supporto perché dà attacco alle cellule dell’epitelio pigmentato della retina, ha funzione respiratoria e partecipa alla formazione della barriera emato-retinica. 3) La membrana di brunch è anche coinvolta nella riparazione dell’epitelio pigmentato della retina. Nel corso di alcune patologie si può accumulare materiale al livello dell’epitelio pigmentato della retina prima e della membrana di Brunch dopo, ciò comporta un’anomalia del trofismo e quindi una degenerazione maculare (macula lutea, fovea, fovea centralis sono la stessa cosa). Il drenaggio venoso della corioidea avviene ad opera delle vene vorticose.
CORPO CILIARE: è la seconda delle tre parti che costituiscono la tonaca vascolare, si interpone tra corioidea e iride. Inizia dove termina la corioidea nel punto di passaggio tra retina visiva e cieca ciliare, cioè in corrispondenza dell’ora serrata, infatti il rivestimento interno cioè la faccia posteriore del corpo ciliare è rivestita da retina cieca ciliare. Ha forma circolare. Si divide in
- Parte orbicolare: è la parte posteriore, detta anche pars plana (si parla di planite), presenta una struttura simile a quella corioidea, caratterizzata da molte pieghe dette orbicolari
- Pars plicata o corona ciliare, è la parte anteriore, presenta molti processi ciliari 80-90, sono come estroflessioni, sono una specializzazione vascolare costituite da una porzione più interna dove troviamo vasi sanguigni provenienti dalle ciliari posteriori lunghe. svolgono un ruolo importante nella produzione dell’umor acqueo, per la sua produzione gioca un ruolo importante anche la retina cieca che riveste il corpo ciliare.
- Una struttura muscolare detta muscolo ciliare, è uno dei tre muscoli intrinseci dell’occhio, ha forma anulare e in sezione trasversale appare triangolare con apice prospiciente all’angolo irido-corneale. Il muscolo ciliare è posto tra la parte anteriore della faccia esterna del corpo ciliare e la parte

11
anteriore della faccia interna della sclera, è costituito da mio cellule con andamento diverso e per questo distinguiamo:
o Porzione esterna : le fibre si dispongono in modo longitudinale, dette fibre meridiane e rappresentano circa il 90% delle fibre totali. Queste fibre costituiscono il muscolo di Brucke. Originano dallo sperone della sclera si portano indietro, la loro contrazione determina lo spostamento in avanti del corpo ciliare.
o Una porzione interna in cui le fibre si dispongono in modo circolare, corrispondono al muscolo di Muller, la loro contrazione determina un restringimento del muscolo e quindi un rilassamento delle fibre della zonula ciliare di Zinn con conseguente aumento dell’indice di rifrazione del cristallino. Il muscolo di Muller è fondamentale per l’accomodazione, le sue fibre di sviluppano verso i due anni, quindi all’inizio della vita la vista non è perfetta.
Il muscolo ciliare è innervato da fibre parasimpatiche postgangliari provenienti dal ganglio ciliare il quale è presente in cavità orbitaria e riceve fibre pregangliari dal nucleo di Edginger-Westphal, è un nucleo posto nel mesencefalo che fa riferimento al nervo oculomotore (3°)
o Sul corpo ciliare, sia sulla pars plicata quindi tra gli spazi compresi tra i processi ciliari, sia nella pars plana, si inseriscono le fibre zonulari, che nel complesso costituiscono l’apparato sospensore del cristallino o detto apparato zonulare, cioè quella struttura che permette al cristallino di mantenere il suo rapporto con il corpo ciliare. Le fibre sono anulari hanno composizone diversa dal collegene, si inseriscono radialmente sul cristallino. Il muscolo ciliare è fondamentale per l’accomodazione: processo di adattamento del cristallino necessario per la visione distinta degli oggetti, la contrazione del muscolo ciliare causa un rilassamento delle fibre della zonula ciliare di Zinn che determina il cambiamento del raggio di curvatura delle facce del cristallino, soprattutto quella anteriore che diventa più convessa. La faccia esterna del corpo ciliare è in diretto rapporto con la sclera, lo strato più interno del corpo ciliare è ricoperto da una porzione della retina cieca detta in questo caso retina ciliare, è responsabile della produzione dell’umor acqueo ed è formata da due strati che dall’esterno verso l’interno sono:
Epitelio pigmentato che poggia direttamente sulla superficie interna del corpo ciliare, in continuità con la retina visiva
Strato di cellule non pigmentate che guardano direttamente verso la camera posteriore.
UMOR ACQUEO: l’umor acqueo è uno dei quattro mezzi diottrici del bulbo oculare, è quindi trasparente, è chiaro e leggermente alcalino con indice di rifrazione di 1,33. ma svolge anche un ruolo nel mantenimento della pressione endo-oculare. È prodotto dall’epitelio ciliare della retina, al livello della camera posteriore del bulbo( esistono tre camere anteriore, posteriore e camera vitreale), viene riassorbito in corrispondenza dalle camera anteriore. Viene accolto nella

12
camera posteriore e anteriore del bulbo oculare. L’umor acqueo viene costantemente prodotto e riassorbito, è importante anche per il trofismo del cristallino e della cornea. Viene prodotto a partire dal plasma sanguigno ma rispetto ad esso ha una composizione diversa, esiste infatti una barriera emato-acquea e per questa sono fondamentali le cellule dell’epitelio non pigmentato che hanno giunzioni serrate.
Per la sua produzione esiste una modalità passiva e una modalità attiva. Per modalità passiva viene prodotto mediante diffusione e ultrafiltrazione del capillare attraverso lo stroma per giungere poi all’epitelio pigmentato. Dall’epitelio pigmentano non può giungere direttamente nella camera posteriore a causa delle giunzioni serrate, da qui in poi la modalità di produzione dell’umor sarà attiva, con dispendio di energia da parte delle cellule dello strato non pigmentato della retina cieca che riveste il corpo ciliare. Rispetto al plasma c’è una minor concentrazione di proteine per mantenere una certa trasparenza, è alta anche la concentrazione di vitC (nei pazienti con cataratta, opacizzazione del cristallino, hanno livelli minori di acido ascorbico al livello ematico, quindi anche al livello dell’umor acqueo, vitC è un antiossidante e se ce ne ho meno contribuisce alla catarattogenesi.)
Per il suo riassorbimento, questo avviene al livello della camera anteriore del bulbo oculare, precisamente nell’angolo irido-corneale posto nella prossimità del limbus (punto di passaggio tra sclera e cornea). A questo livello è presente un sistema trabecolare sclero-corneale, cioè un insieme di trabecole che delimitano degli spazi detti spazi di Fontana, gli spazi di Fontana convogliano nel canale di Schlemm o detto seno venoso della sclera ( si va verso le vene). Dal canale di Schlemm l’umor acqueo che deve essere riassorbito si insinua prima nelle vene acquee (sono strutturalmente delle vene ma non passa sangue, ma solo umor acqueo perché a questo livello la pressione dell’umor supera quella del sangue. Dalle vene acquee si passa alle vene episclerali poste sopra la sclera.
La pressione endo-oculare serve per garantire la turgidità dell’occhio, il flusso di umor acqueo viene regolato per differenza di pressione e in condizioni normali la pressione endonucleare è 20mmHg, mentre quella del sangue contenuta nelle vene episclerali che ricevono l’umor è di circa 13mmHg, quindi ho 7mmHg di vantaggio, se tale pressione aumenta si ha la base del glaucoma, generalmente non è considerato patologico un valore compreso tra 10 e 21 mmHg. In un paziente in cui l’equilibrio tra produzione e riassorbimento viene a mancare può avere ipertensione endo-oculare con conseguente glaucoma, l’aumento della quantità di umor acqueo causa problemi soprattutto alla retina perché il sistema spinge verso questa. Esistono due tipi di glaucoma
- Ad angolo aperto angolo sclero-corneale ad una sorta di invecchiamento del sistema trabecolare di deflusso: l’umore acqueo raggiunge il trabecolato, ma non viene sufficientemente filtrato in quanto quest’ultimo è strutturalmente alterato (ostruzione dei canali di scarico).
- Ad angolo chiuso posizionamento anomale dell’iride. Nel glaucoma ad angolo chiuso l’umore acqueo non riesce a raggiungere il sistema trabecolare di deflusso perchè l’angolo formato da iride e cornea è troppo stretto: l’iride si addossa lentamente alla cornea ostruendo completamente il passaggio.

13
Sebbene non possa essere curato, il glaucoma può essere ben controllato con una terapia adeguata.
Il drenaggio dell’umor avviene sia nel sistema trasecolare per circa 85%, il resto viene drenato dalla cosiddetta via uveo-sclerale la quale si compone di un sistema trabecolare con delle lacune in comunicazione con la camera anteriore, questi spazi sono attraversati dall’umor,esso raggiunge il tessuto uveale che riveste la faccia esterna del muscolo ciliare e viene riassorbimento ma a differenza di quello precedente, il sistema di riassorbimento uveo-sclerale è pressione indipendente e in questo punto è stato osservato un dispositivo simile dal punto di vista immuno-istochimico al tessuto linfatico.
- Sistema trabecolare riassorbimento pressione dipendente.- Sistema uveo-sclerale riassorbimento pressione indipendente.
CRISTALLINO: forma di lente biconvessa asimmetrica, la sua faccia anteriore è meno convessa della posteriore. In corrispondenza della regione equatoriale del cristallino si inseriscono le fibre zonulari che si dispongono sia in corrispondenza di esso che dietro, esse sono il collegamento con il muscolo ciliare, questo è detto apparato sospensore del cristallino e tali fibre sono tese tra la corona ciliare (pars plicata) e la parte anteriore dell’orbicolo ciliare da una parte e la regione equatoriale del cristallino dall’altra. Tali fibre sono disposte radialmente intorno al cristallino, si distungono in anteriori e posteriori, derivano dall’epitelio della parte ciliare della retina e sono digerite dall’elastasi e non dalla collagenasi. Il cristallino deve modificarsi quindi deve essere per forza costituito da un materiale elastico, messo in soluzione fisiologica i cristallino assume forma sferica, ma deve essere comunque resistente.

14
Il cristallino è l’unico mezzo diottrico in cui possiamo modificare l’indice di rifrazione, la forma del cristallino è modificabile grazie alla presenza del muscolo ciliare, un muscolo liscio. La modificazione del cristallino avviene tramite il riflesso di accomodazione, tale riflesso è fondamentale per permettere ai raggi luminosi di convergere nel punto di fissazione della retina corrispondente a una regione della stessa, caratterizzata da un diametro di circa 350micron.
Nel paziente miope i raggi luminosi convergono prima di giungere sul punto di fissazione, per cui il suo occhio necessita di una lente divergente accessoria per spostare più indietro il punto di convergenza. Un paziente ch ha perso l’elasticità del cristallino è detto presbite, calo del potere di accomodazione dell'occhio, si manifesta come difficoltà a mettere a fuoco da vicino.
termine ametropia indica un'anomalia di rifrazione dell'occhio. In un occhio otticamente normale, ad accomodazione rilassata (in condizione cioè di rifrazione statica), i raggi paralleli, provenienti da un punto oggetto posto all'infinito, convergono sulla retina a formare un punto immagine, che in questo caso coincide con il fuoco immagine del sistema ottico oculare. Quando questa condizione ottica ideale è rispettata siamo in presenza di emmetropia. Le ametropie possono essere di tre tipi fondamentali: Ipermetropia (dal greco: υπερ, in eccesso, μετρον, misura, ωψ, occhio, dove la misura in eccesso è riferita alla lunghezza focale), è la condizione in cui il fuoco immagine si trova oltre il piano retinico. Miopia (dal greco: μυω, chiuso ωψ, occhio, dall'abitudine dei miopi di fessurare le palpebre per vedere meglio), è la condizione in cui il fuoco immagine si trova davanti al piano retinico.[2] Astigmatismo (dal greco: α, privativo στιγμα, punto), è la condizione in cui non si ha la presenza di un singolo punto focale. Da questo elenco è esclusa una condizione oculare affine, la presbiopia (dal greco: πρεσβνς, vecchio, e ωψ, occhio), che rappresenta invece un difetto di accomodazione.
Durante il riflessi di accomodazione si ha la contrazione del muscolo di Muller con conseguente rilassamento delle fibre zonulari di Zinn, i cristallino non subendo più la tensione delle fibre assume forma maggiormente sferica, soprattutto la faccia anteriore del cristallino diventa più convessa, in questo modo il cristallino aumento il suo indice di rifrazione. La forma del cristallino non cambia però in continuazione, infatti la contrazione del muscolo ciliare avviene solo se si osserva un oggetto posto massimo a 5m di distanza, quindi il riflesso di accomodazione si innesca quando si osserva un oggetto da vicino, ad esempio quando si legge.
Il cristallino è l’unico mezzo diottrico dell’occhio di cui è possibile modificare l’indice di rifrazione, è una lente biconvessa di consistenza duro-elastica, l’immagine che attraversa il cristallino al livello della retina sarà ribaltata, cioè ciò che è posto superiormente a destra, sulla retina sarà rappresentato inferiormente e a sinistra. lungo la regione equatoriale del cristallino si inseriscono le fibre zonulari dello Zinn.
La faccia anteriore del cristallino è meno convessa della posteriore, è in rapporto con la camera posteriore, con l’iride e la pupilla ed è quella che si modifica maggiormente. La faccia posteriore è più convessa e poggia sulla fossa ialoide. Il cristallino è avvolto dal cristalloide, una capsula che rappresenta una membrana basale piuttosto spessa costituita da collagene soprattutto 4, connettivo, laminina ecc. la cristalloide è una barriera contro batteri e virus, quando il cristallino non è ancora sviluppato la rosolia può colpirlo e dare cataratta congenita. La cristalloide è anche una riserva di fattori di crescita, non è dunque solo una barriera. Al di sotto della capsula detta cristalloide si trova l’epitelio del cristallino formato da cellule cubiche che divengono cilindriche verso l’equatore. Tali cellule si trasformano nelle fibre del cristallino, cioè in cellule allungate che vanno a costituire la maggior parte della sostanza propria del cristallino, tra le fibre non è

15
presente matrice extracellulare si evita il problema della rifrazione Le proteine che costituiscono la parte interna delle cellule sono dette proteine cristalline, sono una componente peculiare delle fibre e rappresentano il 90% circa delle proteine del cristallino. Il potere di rifrazione varia dalla periferia 1,38 al centro 1,5 perché varia la concentrazione delle proteine cristalline dal 15% nelle fibre corticali al 70% in quelle centrali.
CORPO VITREO: è un o dei quattro mezzi diottrici, quello posto più posteriormente. Occupa la camera vitreale, quella più ampia tra le camere del bulbo. Aderisce alla retina visiva specialmente in corrispondenza dell’ora serrata, si discosta leggermente dalla retina ciliare. Il corpo vitreo presenta al davanti la fossa ialoidea su cui poggia la faccia posteriore (+convessa) ha indice di rifrazione di 1,334. Il corpo vitreo è una massa gelatinosa, incolore e amorfa formata per metà circa da acquea, in esso si possono distinguere una fase liquida formata acido ialuronico e acqua e una fase solida formata da una sottile rete di fibre collagene. Alla periferia sono presenti cellule detti ialociti deputati alla produzione di acido ialuronico e fibre collagene. In seguito a fenomeni ossidativi le fibre collagene che costituiscono il corpo vitreo tendono a frammentarsi formando filmaneti di forme diverse che fluttuano all’interno del liquido e proiettano un’ombra sulla retina, questi filamenti sono chiamati dagli inglese mosche volanti. Con l’età può ridursi, tirare la retina e provocarne il distacco. Non è vascolarizzato al termine dello sviluppo.
canale ialoideo (o canale di Cloquet o hyaloid canal) è un canale che percorre il corpo vitreo dell'occhio, dalla papilla ottica (punto in cui le fibre nervose convergono per formare il nervo ottico) fino alla fossa ialoidea (parte posteriore del cristallino). Nel periodo fetale conteneva l'arteria ialoidea che portava nutrimento al cristallino e che, tranne in alcuni casi dove può rimanere e creare problemi, è assente in età adulta.
TONACA NERVOSA
Retina: le informazioni provenienti dalla retina non sono tutte destinate a produrre una visione. Il termine via ottica e via visiva hanno diverso significato. Appartengono alle vie visive anche circuiti nervosi importanti per il riflesso di accomodazione, per il riflesso pupillare e per la regolazione dei cicli circadiani, sono sistemi che si basano non sull’informazione proveniente da una certa immagine ma piuttosto dalla quantità di luce che giunge alla retina.
È la tonaca nervosa del bulbo, la più interna delle tre. Aderisce alla tonaca vascolare fino al margine pupillare, l’ora serrata è una linea di confine in corrispondenza della quale la retina modifica la sua struttura (scomparsa dei recettori) che separa la retina in due parti: retina visiva e retina cieca. La retina cieca si suddivide a sua volte in retina ciliare

16
(importante per la capacità dell’epitelio pigmentano e non di produrre umor) e retina iridea. Sia la retina cieca che la retina visiva sono costituiti da due foglietti a contatto tra loro:
- Un foglietto esterno detto epitelio pigmentato: è a contatto con lo strato interno della tonaca vascolare, o meglio con la sua lamina vitrea di Brunch. È formata da un singolo strato di cellule ricche di pigmento, la melanina. Ha varie funzioni: impedisce la riflessione dei raggi luminosi, agisce come una camera scura dell’occhio e può svolgere questa funzione non tanto perché scura ma grazie al rapporto che contrae con il segmento esterno dei fotorecettori, infatti l’epitelio pigmentato si frappone tra i vari recettori. Altre funzioni importanti del foglietto esterno è la rimozione apicale dei segmenti esterni dei fotorecettori che sono andati incontro a esaurimento funzionale, infatti ha anche attività fagocitaria. Recupera allo stesso tempo in parte il materiale fotosensibile, è ricco infatti di vitamina A che rappresenta un precursore, quella non recuperabile diventa lipofuscina. Il foglietto esterno media gli scambi tra corio capillare e fotorecettori.
- Un foglietto interno o retina propriamente detta: aderisce molto allo strato esterno solo in corrispondenza dell’ora serrata e papilla ottica, per questo la retina può distaccarsi in seguito a traumi, nel punto i ncui si stacca si perde la visione. La rinite pigmentosa avviene se i processi di fagocitosi sono insufficienti perché la retina non +è adesa saldamente all’epitelio pigmentato. il foglietto interno della retina è costituito da 10 strati. Ne ricordiamo solo 3 1) strato di fotorecettori 2) strato di cellule bipolari 3) strato di cellule gangliari
Il retinale, o aldeide della vitamina A, è una delle molte forme della vitamina A. Il retinale, il cui nome deriva dalla sua presenza nella retina dell'occhio, è un cromoforo polienico che, legato a delle proteine dette opsine, costituisce la base chimica della vista.
I fotorecettori si distinguono in coni (3 tipi) importanti per la visione cromatica e bastoncelli (1 tipo) importanti per la visione in bianco e nero. Tutti e 4 i tipi sono in contatto con le cellule bipolari le quali entrano in contatto con le cellule gangliari, dalle cellule gangliari parte la via visiva.
Dei recettori possiamo individuare varie parti, importante è soprattutto il segmento esterno, essendo la parte più esterna del recettore è l’unica dove può avvenire la trasduzione del segnale.
Bastoncelli: è caratterizzato da un segmento esterno di 30-50micron. Sono i più numerosi circa 100milioni, sono assenti nella foveola e praticamente anche nella fovea ad eccezione della sua parte più periferica. Nella regione extrafoveale e periferica della retina sono molto più abbondanti rispetto ai coni. Sono più sensibili alla luce rispetto ai coni e sono maggiormente concentrati al livello della zona periferica della retina, rispetto ai coni. I bastoncelli hanno bassa soglia di attivazione, quindi è sufficiente poca luce per attivarli e avere una trasduzione del segnale. I bastoncelli hanno la capacità di adattarsi all’oscurità, infatti possono scaricare meglio dopo un certo arco di tempo nelle stesse condizioni di luce, anche se dopo un certo periodo la visione rimane la stessa. i bastoncelli hanno la caratteristica di consentire la visione cromatica dei colori. Sinaptano con le cellule bipolari. Il loro pigmento è la rodopsina.
Coni: hanno un segmento esterno di 12micron. Si trovano nella fovea. Hanno una maggior soglia di eccitazione, consentano la visione cromatica e permettono di avere una visione distinta cogliendo i dettagli più fini. Sono maggiormente concentrati nella fovea centralis dove si ha il massimo grado di discriminazione visiva, e infatti qui ci sono concentrati altamente e inoltre sono presenti dei coni foveali che sono leggermente più sottili del normali rispetto agli extrafoveali. Nella foveola abbiamo solo i coni che sono in rapporto 1:1 coni le cellule

17
bipolari gangliari per permette una maggior capacità discriminativa (se su un recettore convergono tante fibre si ha maggior sensibilità ma minor capacità discriminativa, nei coni foveali accade l’opposto cioè un’unica fibra va su un recettore quindi la capacità discriminativa è altissima). Sinaptano con le cellule bipolari.
Le porzione più importante del fotorecettore è il segmento esterno che è dato da numerose pieghe della citomembrana, le parti più nuove del segmento esterno sono quelle poste più in basso, mentre quelle più apicali sono quelle più vecchie che quando andranno incontro a esaurimento funzionale verranno fagocitate dalle cellule dell’epitelio pigmentato della retina. Questa struttura del segmento esterno consente una maggior superficie sulla quale esporre il pigmento fotosensibile.
Quando il recettore viene raggiunto che succede? In condizioni di oscurità il fotorecettore è depolarizzato, cioè i canali sono aperti, mentre è iperpolarizzato quando viene esposto alla luce cioè i canali si chiudono. Si tratta di una novità perché di solito quando un recettore viene stimolato si depolarizza, nel caso dei fotorecettori invece accade l’opposto. Nei Bastoncelli abbiamo inoltre numerosi canali per il Na+ aperti in condizione di buio, perché nella cellula ci sono alte concentrazioni di cGMP che tiene aperti i canali. Il GMP ciclico gioca un ruolo fondamentale. Appena la rodopsina viene colpita dal raggio (formata da opsina e 11-cis retinale) diventa 11-transretinale, stimola l’opsina che attivata induce l’attivazione della trasducina che a sua volta attiva la 5-fosfodiesterasi che causa l’idrolisi del GMP ciclico presente e ciò induce una diminuzione della sua concentrazione intracellulare che induce la chiusura dei canali ionici sodio, quindi il fotorecettore si iperpolarizza.
La luce viene assorbita in primo luogo da un cromoforolegato alla proteina recettrice. Tale cromoforo, che può appartenere tanto ai coni che ai bastoncelli, è l'11-cis-retinale.[9] Esso, legandosi alla proteina opsina con formazione di una base di Schiff, forma il pigmento visivo rodopsina. Quest'ultima, quando viene a contatto con un fotone di luce, subisce una fotodecomposizione, o imbianchimento, che porta alla dissociazione della molecola con formazione di retinale tutto-trans e opsina. Nella fotodecomposizione viene generato il potenziale d'azione che si propaga attraverso il nervo ottico fino all'encefalo. Quando la luce interagisce con l'11-cis-retinale, catalizza la sua conversione a tutto-trans-retinale, che determina un cambiamento conformazionale nella "opsina" (parte proteica della rodopsina) trasformandola in opsina attiva. Questa proteina attivata, attiva una seconda proteina, una proteina Gt, la trasducina, che a sua volta attiva una specifica GTP Fosfodiesterasi, che scinde il cGMP in GMP e determina un abbassamento della concentrazione del cGMP: i canali per il Na+ lungo la membrana della cellula a bastoncello si chiudono, si determina una variazione del potenziale di membrana (iperpolarizzazione). La membrana essendo polarizzata non rilascia il GABA (γ-amminobutirrato, un neurotrasmettitore inibitorio), in questo modo le cellule del nervo ottico non essendo inibite possono generare l'impulso nervoso, che arriverà al nostro cervello tramite il nervo ottico.
In sintesi. Esiste un solo tipo di bastoncello tre tipi diversi di coni in base alla lunghezza d’onda captata. I tipi di coni sono rossi, verdi e blu. I due tipi di recettore hanno diversa capacità di adattamento alla condizione di scarsa illuminazione, infatti i bastoncelli sono più sensibili dei coni a queste condizioni perché hanno bassa soglia di stimolazione e hanno una maggiore sommazione spaziale rispetto ai coni, cioè più bastoncelli fanno riferimento ad un’unica cellula gangliare, mentre il rapporto tra coni e cellule gangliari è 1:1. In condizioni di scarsa illuminazione, essendo i bastoncelli recettori a bassa soglia di stimolazione e facendo riferimento a un certo numero di essi ad un’unica fibra, lo stimolo percepito da ogni singolo bastoncello del gruppo si somma a quello percepito dagli altri bastoncelli che fanno riferimento alla stessa fibra, in questo modo la sommatoria è in grado di eccitare l fibra. Il

18
singolo stimolo percepito dal singolo bastoncello non può eccitare la fibra. Questo implica però scarsa discriminazione dello stimolo.
Nella percezione della visione le cellule gangliari svolgono un ruolo sia nella percezione dei movimenti sia nella percezione dei dettagli, essi sono infatti sensibili agli stimoli luminosi e dispongono di un pigmento fotosensibile detto melanopsina.
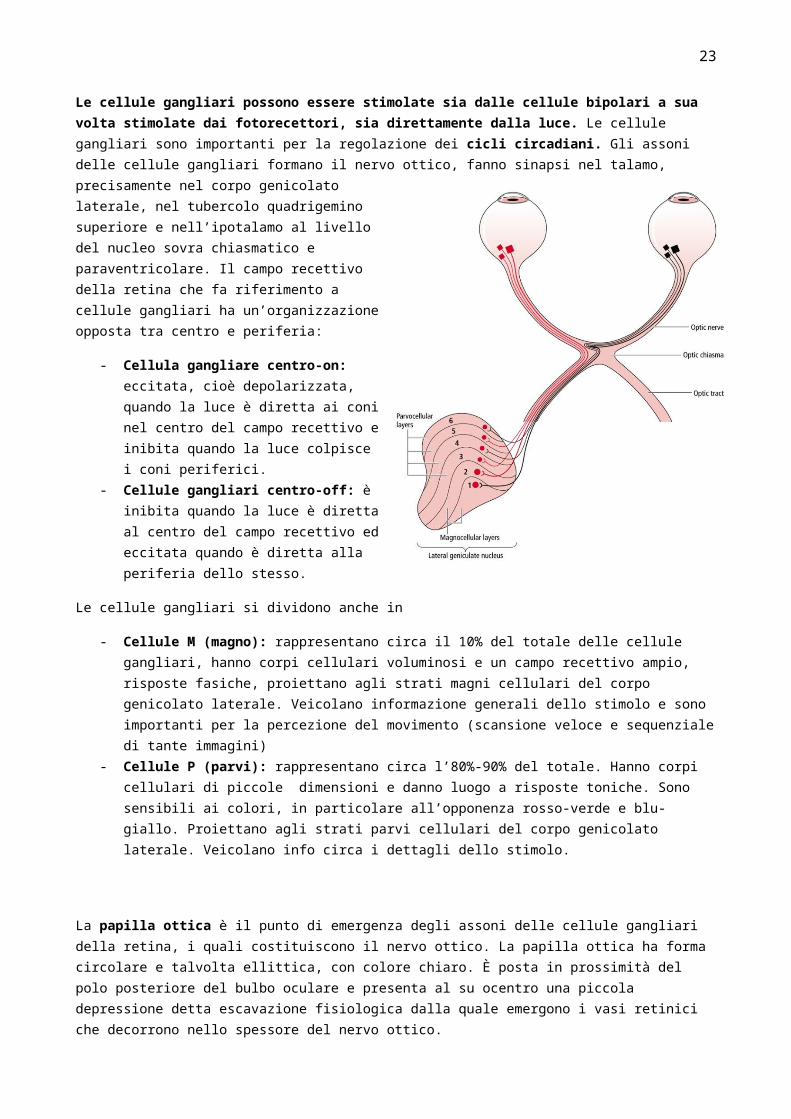
Le cellule gangliari possono essere stimolate sia dalle cellule bipolari a sua volta stimolate dai fotorecettori, sia direttamente dalla luce. Le cellule gangliari sono importanti per la regolazione dei cicli circadiani. Gli assoni delle cellule gangliari formano il nervo ottico, fanno sinapsi nel talamo, precisamente nel corpo genicolato laterale, nel tubercolo quadrigemino superiore e nell’ipotalamo al livello del nucleo sovra chiasmatico e paraventricolare. Il campo recettivo della retina che fa riferimento a cellule gangliari ha un’organizzazione opposta tra centro e periferia:
- Cellula gangliare centro-on: eccitata, cioè depolarizzata, quando la luce è diretta ai coni nel centro del campo recettivo e inibita quando la luce colpisce i coni periferici.
- Cellule gangliari centro-off: è inibita quando la luce è diretta al centro del campo recettivo ed eccitata quando è diretta alla periferia dello stesso.
Le cellule gangliari si dividono anche in
- Cellule M (magno): rappresentano circa il 10% del totale delle cellule gangliari, hanno corpi cellulari voluminosi e un campo recettivo ampio, risposte fasiche, proiettano agli strati magni cellulari del corpo genicolato laterale. Veicolano informazione generali dello stimolo e sono importanti per la percezione del movimento (scansione veloce e sequenziale di tante immagini)
- Cellule P (parvi): rappresentano circa l’80%-90% del totale. Hanno corpi cellulari di piccole dimensioni e danno luogo a risposte toniche. Sono sensibili ai colori, in particolare all’opponenza rosso-verde e blu-giallo. Proiettano agli strati parvi cellulari del corpo genicolato laterale. Veicolano info circa i dettagli dello stimolo.
La papilla ottica è il punto di emergenza degli assoni delle cellule gangliari della retina, i quali costituiscono il nervo ottico. La papilla ottica ha forma circolare e talvolta ellittica, con colore chiaro. È posta in prossimità del polo posteriore del bulbo oculare e presenta al su ocentro una piccola depressione detta escavazione fisiologica dalla quale emergono i vasi retinici che decorrono nello spessore del nervo ottico.

19
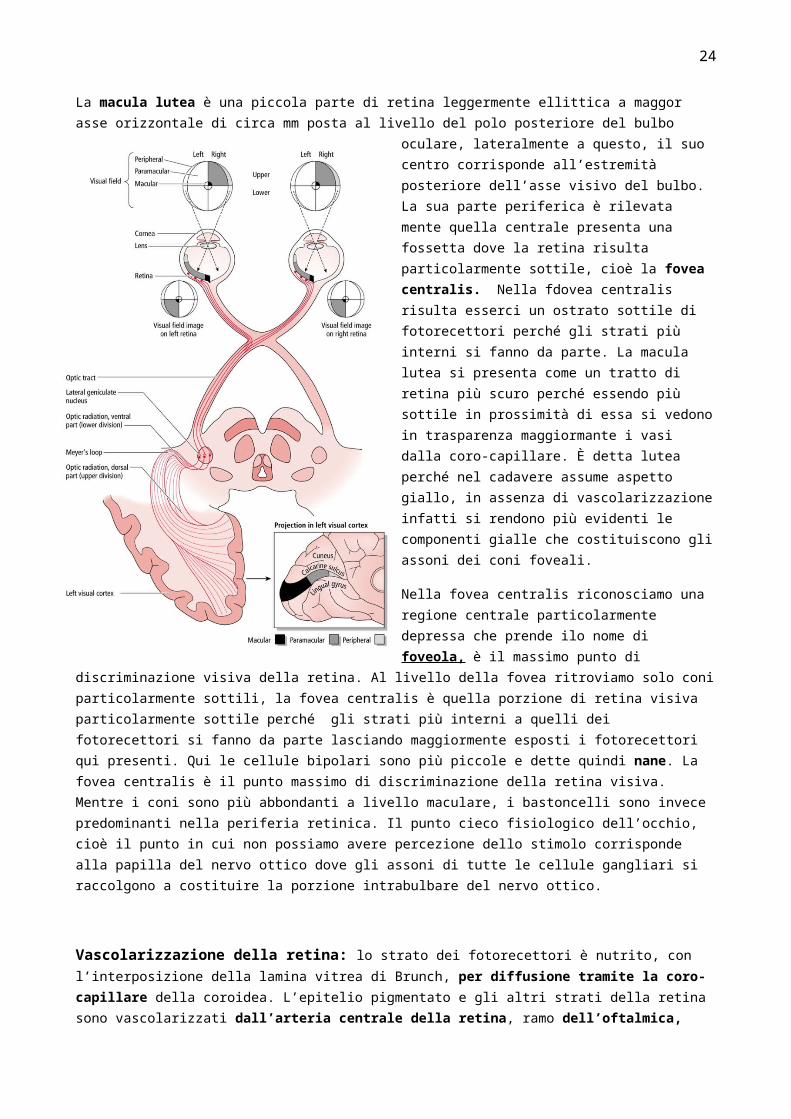
La macula lutea è una piccola parte di retina leggermente ellittica a maggor asse orizzontale di circa mm posta al livello del polo posteriore del bulbo oculare, lateralmente a questo, il suo centro corrisponde all’estremità posteriore dell’asse visivo del bulbo. La sua parte periferica è
rilevata mente quella centrale presenta una fossetta dove la retina risulta particolarmente sottile, cioè la fovea centralis. Nella fdovea centralis risulta esserci un ostrato sottile di fotorecettori perché gli strati più interni si fanno da parte. La macula lutea si presenta come un tratto di retina più scuro perché essendo più sottile in prossimità di essa si vedono in trasparenza maggiormante i vasi dalla coro-capillare. È detta lutea perché nel cadavere assume aspetto giallo, in assenza di vascolarizzazione infatti si rendono più evidenti le componenti gialle che costituiscono gli assoni dei coni foveali.
Nella fovea centralis riconosciamo una regione centrale particolarmente depressa che prende ilo nome di foveola, è il massimo punto di discriminazione visiva della retina. Al livello della fovea ritroviamo solo coni particolarmente sottili, la fovea centralis è quella porzione di retina visiva particolarmente sottile perché gli strati più interni a quelli dei fotorecettori si fanno da parte lasciando maggiormente esposti i fotorecettori qui presenti. Qui le cellule bipolari sono più piccole e dette quindi nane. La fovea centralis è il punto massimo di discriminazione della retina visiva. Mentre i
coni sono più abbondanti a livello maculare, i bastoncelli sono invece predominanti nella periferia retinica. Il punto cieco fisiologico dell’occhio, cioè il punto in cui non possiamo avere percezione dello stimolo corrisponde alla papilla del nervo ottico dove gli assoni di tutte le cellule gangliari si raccolgono a costituire la porzione intrabulbare del nervo ottico.
Vascolarizzazione della retina: lo strato dei fotorecettori è nutrito, con l’interposizione della lamina vitrea di Brunch, per diffusione tramite la coro-capillare della coroidea. L’epitelio pigmentato e gli altri strati della retina sono vascolarizzati dall’arteria centrale della retina, ramo dell’oftalmica, ramo staccato dalla carotide interna. l’arteria oftalmica entra in cavità orbitaria attraverso il canale ottico insieme al nervo ottico. L’arteria centrale decorre per un breve tratto insieme al nervo, poi in prossimità del bulbo, affonda nello stesso nervo, così raggiunge il bulbo dove si rende visibile al livello della papilla del nervo ottico. In corrispondenza della papilla del nervo ottico si divide in 4-5 rami che giungono alla retina, ad eccezione dello strato dei fotorecettori. L’arteria centrale è l’unica osservabile per mezzo di strumenti appropriati dall’esterno, la sua valutazione è alla base di valutazioni cliniche per alcune patologie come diabete.

20
Altre arterie importanti: arterie ciliari brevi e lunghe, rami dell’oftalmica. Le posteriori brevi circondano il canale sclerale del nervo ottico formando il cerchio arterioso di Zinn-Haller e sono impalcatura vascolare della coroidea. Il drenaggio venoso è svolto dalle 4 vene vorticose. Le posteriori lunghe,2, si portano in corrispondenza del corpo ciliare per dare vasi destinati a diventare i processi ciliari. Alla costituzione dell’impalcatura vascolare del corpo ciliare e dell’iride partecipano anche la arterie ciliari anteriori che si staccano da rami a
destino muscolare dell’art. oftalmica.
VIA VISIVA: dalla retina alla corteccia visiva
primaria.
Il campo visivo è cosa diversa dal campo retinico, il raggio luminoso attraversa una lente biconvessa quindi sul campo retinico l’immagine risulta ribaltata, ciò che sta nel campo visivo in alto e a destra corrisponde sul campo retinico in basso e a sinistra. L’uomo è cacciatore, grande parte del nostro campo visivo è binoculare per cui si ha migliore percezione della distanza. Si parla di emiretine: nasale e temporale. La retina viene anche divisa in quadranti che si distinguono in nasali, temporali, superiore e inferiori.
Le fibre della via visiva sono organizzate dalla retina fino alla corteccia di riferimento in modo retino-tipicamente organizzata. Ogni campo retinico proietta a una parte precisa dell’area visiva primaria. La macula ha a disposizione una grande superficie cerebrale.
Il chiasmo ottico corrisponde all’incrociamento parziale sulla linea mediana delle fibre di ciascun nervo ottico provenienti dai quadranti nasali della retina, al chiamo ottico seguono i due tratti ottici che sono dati dalla mescolanza dei due nervi ottici. La parte laterale del campo visivo è proiettata verso l’emiretina nasale omolaterale e emiretina temporale contro laterale. In caso di perdita totale o compressione del chiasmo ottico come in caso di adenoma ipofisario si ha la perdita delle porzioni laterali del campo visivo. L'emianopsia consiste nella perdita di metà del campo visivo. Può colpire la metà destra/sinistra o la metà alta/bassa del campo visivo. Il tratto ottico veicola prevalentemente info provenienti dal campo visivo contro laterale, essendo
costituito per metà da fibre provenienti dall’emiretina temporale omolaterale che riceve informazioni dal campo visivo anch’esso contro laterale. Le fibre del tratto ottico giungono al corpo genicolato laterale omolaterale, cioè a un nucleo talamico di relay sensitivo specifico per la via visiva, esso è suddiviso in strati. Nel corpo genicolato laterale ci sono

1) Cellule gangliari corpo genicolato laterale del talamo o via della percezione per forma e volore o via della percezione dei movimenti corteccia visiva primaria.
2) Area visiva primaria aree visive secondarie di secondo ordine e ordine superiore o corpo calloso per corteccia visiva primaria contro laterale x unificare le info prov dai due occhi o mesencefalo per controllare movimento oculari e messa a fuoco., precisamente collicolo superiore per i movimenti saccadici e orientamento degli occhi e testa versi stimoli, organizz,, retinotopicamente, oppure nuclei area pretatta per riflesso pupillare.
3) Retina ipotalamo.
21
strati magni cellulari e strati parvi cellulari nei quali giungono rispettivamente i prolungamenti centrali delle cellule gangliari M e P della retina. Il corpo genicolato laterale appartiene al talamo dorsale ed è costituito da sei lamine, gli strati parvi cellulari vanno dal 3° al 6°, mentre gli strati magni cellulari vanno dal 1° al 2°. Al di sotto di ogni lamina ci sono strati conio cellulari.
Ad ogni corpo genicolato laterale giungono fibre provenienti dall’emiretina temporale omolaterale e all’emiretina nasale contro laterale.
1. Strato magnicellulare giungono fibre delle cellule gangliari M dell’emiretina nasale controlaterale
2. Strato magnicellulare giungono fibre delle cellule gangliari M dell’emiretina temporale omolaterale
3. Strato parvi cellulare giungono fibre delle cellule gangliari P dell’emiretina temporale omolaterale.
4. Strato parvi cellulare giungono fibre delle cellule gangliari P dell’emiretina nasale controlaterale.
5. Strato parvi cellulare giungono le fibre delle cellule gangliari P dell’emiretina temporale omolaterale.
6. Strato parvi cellulare giungono fibre delle cellule gangliari P dell’emiretina nasale controlaterale.
Dal corpo genicolato laterale partono fibre che giungono alla corteccia visiva primaria area 17 di Brodmann, posta in corrispondenza del labbro superiore e inferiore della scissura calcarina, è corteccia etero tipica granulare spessa circa 1,5mm. Tale proiezione talamo-corticale: proiezione genicolo-calcarina, essa costituisce quasi la totalità delle porzioni sottolenticolare e retro lenticolare della capsula interna. Dopo aver costituito queste porzioni di capsula interna, le fibre della proiezione genicolo-calcarina si irradiano formando un’ampia banda che abbraccia il ventricolo laterale, si portano così indietro verso il lobo occipitale dove è presente l’area visiva primaria giungendo alla porzione di corteccia visiva primaria posta nel labbro superiore della scissura calcarina. Alcune fibre della radiazione vanno a costituire l’ansa di Meyer, un insieme di fibre provenienti dalla proiezione genicolo-calcarina che prima si porta in avanti nel lobo temporale al si sopra del corno inferiore del ventricolo laterale, poi curva indietro e raggiunge la corteccia visiva nel lobo occipitale, queste fibre giungono alla porzione di corteccia visiva primaria presente al livello del labbro inferiore della scissura calcarina.
Una lesione del lobo temporale che coinvolge l’ansa di Meyer provoca un difetto nel campo visivo superiore del lato opposto, mentre una lesione al lobo parietale che coinvolge quelle fibre della proiezione genicolo-calcarina che passano al di sopra della scissura calcarina provoca un difetto nel campo visivo inferiore contro laterale.
Esiste una rappresentazione retino-tipica nella corteccia visiva primaria:
- Sul labbro superiore giungono info provenienti dall’emriretina superiore, cioè quelle provenienti dai campi visivi inferiori.

22
- 1/3 della corteccia visiva primaria, a partire dal polo posteriore fa riferimento alle info provenienti dalla macula, proseguendo in avanti le porzioni di corteccia visiva primaria fanno riferimento a porzioni sempre più periferiche (o anche anteriori considerando che si parla di una struttura sferica) della retina visiva.
Oltre alla corteccia visiva primaria area17, è presente anche la corteccia visiva associativa che è rappresentata dalla maggior parte della corteccia della faccia inferiore del lobo temporale, è corteccia importante per il riconoscimento di colore e forme, e dalla parte della corteccia associativi dei lobi parietale e occipitale importanti per percepire il movimento degli oggetti. Le cellule gangliari M, sono fondamentali per percepire i movimenti, le cellule gangliari P per percepire colori e dettagli, così anche nelle cortecce associative visive vi è una rappresentazione precisa, le proiezioni che partono dalle cellule gangliari e che giungono alle aree associative visive si distinguono in due vie.1
1) Via ventrale o via del what pathway o via del cosa: serve per il riconoscimento di un oggetto, è una via costituita da quelle fibre che veicolano le info trasportate all’origine dalle cellule gangliari P e che giungono alla corteccia associativa della faccia inferiore del lobo temporale la quale è importante per il riconoscimento dei dettagli (colore e forma di un oggetto). Nella sindrome di Gruber-Groussy si ha agnosia visiva cioè non si riconoscono gli oggetti e il paziente li porta alla bocca per studiarli.
2) Via dorsale o via del where pathway o via del dove: è costituita da fibre che veicolano le info trasportate in partenza dalle fibre delle cellule gangliari M che si portano in aree associative visive del lobo parietale occipitale le quali sono quindi fondamentali per la percezione del movimento. Tra queste aree associative visive dei lobi occipitale e parietale vi è in particolare l’area v5, è un’area di alto livello discriminativo, un’alterazione bilaterale dell’area v5 causa achinetopsia, cioè incapacità di percepire i movimenti degli oggetti. Verso l’acqua, vedo solo prima la brocca piena e poi il bicchiere pieno e brocca vuota alla fine.
Da ricordare l’area del Wernicke è un’area associativa sia per le vie uditive che visive, l’area di Broca collabora con l’area di Wernicke e grazie a queste correlazioni posso dire il nome di un oggetto quando lo vedo.
VIE OTTICHE RIFLESSE
Riflesso di accomodazione: è un riflesso necessario per modificare il cristallino affinchè la messa a fuoco dell’oggetto che stiamo fissando si mantenga nello strato dei fotorecettori e soprattutto in corrispondenza del segmento esterno di questi ultimi. È
23
diverso parlare del riflesso di accomodazione di un occhio e del riflesso di accomodazione di entrambi. Il riflesso di accomodazione si esplicita solo quando è necessario mantenere a fuoco la visione di un oggetto a distanza ravvicinata, massimo 5 metri, per maggiori distanze non c’è necessità di agire su cristallino. La capacità di accomodazione diminuisce nella vecchiaia a causa di una progressiva perdita di elasticità del cristallino.
Accomodazione:
- Il cristallino si sposta in avanti a causa della contrazione del muscolo di Brucke costituito dalle fibre meridiane del muscolo ciliare, nel mentre diminuisce il grado di tensione dell’apparato sospensore del cristallino per una contrazione del muscolo di Muller, quella parte di muscolo ciliare costituito da fibre circolari. Il muscolo ciliare si contrae in seguito alla stimolazione di fibre postgangliari parasimpatiche provenienti dal ganglio ciliare, nel quale fanno sinapsi le fibre parasimpatiche pregangliari che partono dal nucleo di Edinger-westphal, queste fibre fanno riferimento al nervo oculomotore 3°
- Il cristallino poi aumenta il suo grado di convessità soprattutto a carico della sua faccia anteriore
- Aumenta quindi l’indice di rifrazione del cristallino con conseguente mantenimento del punto focale al livello dello strato dei fotorecettori presenti nella fovea, soprattutto in corrispondenza del segmento esterno degli stessi fotorecettori.
- Diminuzione del diametro pupillare, agisce il muscolo sfintere della pupilla innervato dal parasimpatico, questa diminuzione del calibro è funzionale sia ad aumentare la profondità di campo sia per evitare l’aberrazione cromatica da sfericità.
- Convergenza degli occhi: ossia rotazione dei bulbi in direzione opposta, si muovono i bulbi per contrazione dei retti mediali dei due occhi, retto mediale innervato dall’oculomotore, ma nel mentre si ha un’ inibizione della muscolatura
antagonista quindi retto laterale innervato dall’abducente.- Ruolo importante della corteccia visiva e collicolo superiore che è correlato con la
sua parte più esterna con le info che riceve dalla retina e con la sua parte più profonda con info correlate a funzioni motorie per la gestione della motilità oculare.
Il collicolo superiore è improntante nel controllo dei movimenti oculari e orientamento dello sguardo, alcuni nuclei del collicolo superiore proiettano al nucleo talamico latero-psoteriore e al pulvinar, tali nuclei proiettano ad aree di ordine superiore.
Organizzazione retino topica della cort visiva: macula caudalmente, parte periferica della retina cranialmente,. Il campo visivo di sinistra combacia con area visiva di destra
24
Riflesso pupillare: è quel riflesso che regola il diametro pupillare in risposta dell’intensità della luce che colpisce la retina. Presenta un braccio afferente e uno efferente.
- Braccio afferente: parte della retina, giunge ai nuclei pretettali, sono nuclei mesencefalici, questi nuclei nell’insieme costituiscono l’area pretettale del mesencefalo detta così perché vicina al tetto del mesencefalo rappresentata dalla lamina quadrigemina. L’area pretettale invia info al nucleo di Edger-Westhphal, dal quale parte il braccio efferente del riflesso. L’area pretettale proietta sia l nucleo di Edger-Westphal omolaterale checontro laterale anche se prevalentemente la proiezione è ipsilaterale, quindi se punto la luce in un occhio ottengo una risposta diretto mediante il restringimento del diametro pupillare di quell’occhio ma anche una risposta consensuale nell’occhio dove non ho puntato la luce.
- Braccio efferente: rappresentato da fibre pregangliari che partono dal nucleo di Edger-Westphal e che giungono al ganglio ciliare dal quale si dipartono fibre parasimaptiche postgangliari responsabili dell’innervazione dello sfintere della pupilla.
Midriasi: La midriasi è la condizione fisiologica di dilatazione della pupilla dell'occhio in assenza di luce. L'instillazione di alcune sostanze nell'occhio (uso topico) può provocare dilatazione del diametro pupillare, d es. atropina. Miosi:riduzione diametro pupillare.
Un ruolo importante nel riflesso pupilla è svolto dalle cellule gangliari della retina, esse sono infatti responsabili della costrizione pupillare che si verifica durante quel breve periodo che segue subito dopo lo spegnimento di una luca puntata per più di trenta secondi nell’occhio. Quando si punta una luce per un certo tempo si ha un restringimento pupillare che permane anche quando la luce viene spenta, in questo breve arco di tempo non sono i coni e i bastoncelli a scaricare ma direttamente le cellule gangliari che esprimono melanopsina.
È importante anche anche la lunghezza d’onda della luce, per lunghezze d’onda lunghe, cioè luci rosse si stimolano i coni e quindi delle cellule gangliari con conseguente costrizione, mentre per lunghezze d’onda brevi si ha una stimolazione dirette delle cellule gangliari con conseguente costrizione.
La permanenza della costrizione dopo lo spegnimento della luce si ha perché con determinate lunghezze d’onda brevi si sono stimolate le cellule gangliari che esprimono melanopsina. la permanenza della costrizione causa il fenomeno dell’abbagliamento a causa del quale non si riesce a vedere subito in modo ottimale.

25
Dilatatore della pupilla: è innervato da fibre postgangliari ortosimpatiche che appartengono al plesso carotico, prendono origine dal ganglio cervicale superiore al quale giungono fibre pregangliari provenienti dal segmento craniale della colonna intermedio-laterale del midollo.
Le fibre postgangliari originate dal ganglio cervicale superiore si accompagnano alla carotide interna contribuendo alla costituzione del plesso carotico. Con essa le fibre postgangliari entrano in cavità cranica attraversando dapprima il canale carotico, decorrono poi insieme alla carotide interna finchè essa raggiunge il nervo oftalmico. Dopo che si accompagnano al nervo oftalmico, in particolare al nervo naso ciliare, alcune attraverseranno il ganglio ciliare (il ganglio ciliare è parasimpatico, attraversano solo) andando a costituirei nervi ciliari brevi, altri invece giungono al bulbo andando a costituire i nervi ciliari lunghi. Altre fibre postgangliari ortosimpatiche provvedono poi all’innervazione del muscolo elevatore della palpebra superiore, muscolo che da un punto di vista motorio viene innervato dal nervo oculomotore.
Figura 1the pupillary dilation pathway.
MOVIMENTI OCULARI
Tra i muscoli estrinseci dell’occhio ritroviamo anche il muscolo elevatore della palpebra superiore, se tale muscolo non si contrae correttamente si ha la cosiddetta ptosi palpebrale, ovvero un abbassamento patologico della palpebra superiore. Il muscolo elevatore superiore della palpebra è innervato dall’oculomotore, terzo paio dei nervi encefalici. Il nucleo dell’oculomotore è posto nel mesencefalo in prossimità della sostanza grigia periacqueduttale, tra il collicolo superiore e il nucleo rosso. Il nervo oculomotore ha un nucleo costituito da un complesso nucleare essendo responsabile dell’innervazione di diversi muscoli, tale suddivisioneè alla base della possibilità di modulare la contrzione dei muscoli innervati dallo stesso nervo, es: se contraggo il retto superiore allo stesso tempo ho una inibizione modulata della contrazione della muscolatura antagonista quindi del retto inferiore, sempre innervato dall’oculomotore. L’origine apparente del nervo si ha al livello della fossa interpeduncolare, in

26
prossimità della sostanza perforata posteriore, quindi sulla faccia ventrale del mesencefalo. Il nervo oculomotore emerge e si porta in cavità orbitaria attraversando dapprima il seno cavernoso, uno dei seni della dura made, ai lati del corpo dello sfenoide, poi passa nella fessura orbitaria superiore e giunge in cavità orbitaria dove si divide in:
- Ramo superiore per retto superiore ed elevatore superiore della palpebra- Ramo inferiore per il retto mediale, retto inferiore e obliquo ingeriore. Insieme alle sue
fibre decorrono delle fibre parasimpatiche pregangliari destinate al ganglio ciliare e originate dal nucleo di edger-westphal (il nervo oculomotore è il primo ad avere fibre parasimpatiche, gli altri sono 7,9,10)
Il nervo trocleare è presente nel mesencaflo, in vininanza al collicolo superiore, la sua origine reale si osserva al di sotto della lamina quadrigemina, sulla faccia dorsale del mesencefalo. Il nervo trocleare è l’unico dei ervi encefalici ad avere origine apparente sulla faccia dorsale del tronco. Emerso il nerbo, circonda il tronco encefalico, entra in cavità prbitaria passando dalla fessura orbitaria superiore, in cavità orbitaria passa all’interno dell’anello tendineo di Zinn e si porta al muscolo obliquo superiore.
Nervo abducente: il suo nucleo è posto nella parte inferiore del ponte, è circondato dal primissimo tratto delle fibre del nervo facciale. L’origine apparente del nervo abducente si ha al livello del solco bulbo-pontino, sopra le piramidi bulbari. Emerso, attraversa il seno cavernoso, passa attraverso la fessura orbitaria superiore e si distribuisce al solo muscolo retto laterale.
CAMPI OCULARI FRONTALI:area 8. Essi proiettano direttamente alla formazione reticolare pontina mediana e direttamente al collicolo superiore, lo stesso collicolo superiore proietterà alla formazione reticolare pontina mediana, quindi si può dire che i campi oculari frontali proiettano sia direttamente che indirettamente tramite il collicolo superiore alla formazione reticolare pontina mediana. La formazione reticolare pontina mediana è il centro per il controllo dei movimenti oculari sul piano orizzontale e proietterà ai nuclei del nervi abducente (retto laterale) e oculomotore (retto mediale).
La funzione del collicolo superiore invece è fondamentale per il controllo dei movimenti saccadici degli occhi, fini movimenti che servono a mantenere la fissazione su un oggetto in movimento, riceve anche impulsi direttamente dai centri corticali del lobo parietale (aree associative come la v5) che forniscono informazioni sul movimento degli oggetti nel campo visivo, e riceve impulsi direttamente da centri corticali del lobo frontale per il controllo dei movimenti degli occhi. I neuroni degli strati superficiali ricevono impulsi dalla retina, pare che il collicolo superiore sia coinvolto anche nell’accomodazione, i neuroni degli strati più interni proiettano alla formazione reticolare paramediana, tuttavia gli strati più profondi del collicolo superiore proiettano anche ad altri nuclei presenti nel tronco encefalico, come il nucleo interstiziale rostrale e il nucleo interstiziale di Cajal che sono importanti per il movimento degli occhi sul piano verticali, per cui essi proiettano anche ai nuclei dei nervi oculomotore e trocleare.
Danno area 17 areteria cerebrale posteriore, difetto nel campo visivo contro laterale.
Area visiva primaria 17= V1 detta corteccia striata è divisa i nsei strati principali il 4 è ulteriormente sottotiviso in sottolamine una di queste è la stria di gennari. Al 4° strato arriva la maggior parte delle proiezioni del nucleo genicolato laterale.

27
area visiva di ordine superiore 18= V2, area visiva 19= V3,V4,V5