oilproductionfromyarrowialipolyticapo1g using ... · pdf fileout in a soxhlet extractor with...
TRANSCRIPT

Hindawi Publishing CorporationJournal of Biomedicine and BiotechnologyVolume 2012, Article ID 378384, 10 pagesdoi:10.1155/2012/378384
Research Article
Oil Production from Yarrowia lipolytica Po1g UsingRice Bran Hydrolysate
Yeshitila Asteraye Tsigie,1 Chun-Yuan Wang,1 Novy S. Kasim,1 Quy-Do Diem,1
Lien-Huong Huynh,1, 2 Quoc-Phong Ho,2 Chi-Thanh Truong,2 and Yi-Hsu Ju1
1 Department of Chemical Engineering, National Taiwan University of Science and Technology, 43 Keelung Road, Section 4,Taipei 106, Taiwan
2 Department of Chemical Engineering, Cantho University, 3-2 Street, Cantho City, Vietnam
Correspondence should be addressed to Yi-Hsu Ju, [email protected]
Received 22 September 2011; Revised 31 October 2011; Accepted 3 November 2011
Academic Editor: Isabel Sa-Correia
Copyright © 2012 Yeshitila Asteraye Tsigie et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
The purpose of this study was to produce microbial oil from Yarrowia lipolytica Po1g grown in defatted rice bran hydrolysate. Afterremoving oil from rice bran by Soxhlet extraction, the bran is subjected to acid hydrolysis with various sulfuric acid concentrations(1–4% v/v), reaction times (1–8 h), and reaction temperatures (60–120◦C). The optimal conditions for maximum total sugarproduction from the hydrolysate were found to be 3% sulfuric acid at 90◦C for 6 h. Glucose was the predominant sugar (43.20 ±0.28 g/L) followed by xylose (4.93 ± 0.03 g/L) and arabinose (2.09 ± 0.01 g/L). The hydrolysate was subsequently detoxified byneutralization to reduce the amount of inhibitors such as 5-hydroxymethylfurfural (HMF) and furfural to increase its potential asa medium for culturing Y. lipolytica Po1g. Dry cell mass and lipid content of Y. lipolytica Po1g grown in detoxified defatted ricebran hydrolysate (DRBH) under optimum conditions were 10.75 g/L and 48.02%, respectively.
1. Introduction
The increasing industrialization and motorization of theworld has led to a steep rise for the demand of petroleum-based fuels which are obtained from limited reserves. Thesefinite reserves are highly concentrated in certain regionsof the world. Therefore, those countries not having theseresources are facing energy/foreign exchange crisis, mainlydue to the import of crude petroleum. Hence, it is necessaryto look for alternative fuels which can be produced fromresources available locally within the country such as alcohol,biodiesel, and vegetable oils [1].
Biodiesel is a clean, biodegradable, renewable, andnontoxic fuel which contributes no net carbon dioxide orsulfur to the atmosphere and emits less pollutants thanconventional diesel. However, the high cost of raw material(70–75%) for biodiesel production has become one of themajor obstacles for its development and wide applications[2]. On the other hand, consumption of a large amount ofvegetable oils as raw material for biodiesel production would
result in a shortage in edible oils and leads to the soar offood price. Adoption of animal fat, used frying oil, and wastecooking oil as feedstock is a good strategy to reduce the cost.However, these limited resources cannot meet the increasingneeds for clean renewable fuels [3, 4].
Recently, there has been an increasing interest in lookingfor new oil sources for biodiesel production. Among them,microbial oils, namely single cell oils (SCOs), have attractedgreat attention worldwide. Oils from oleaginous microor-ganisms including bacteria, yeasts, moulds, and microalgae[5] are now considered as promising candidates due to theirspecific characteristics such as being unaffected either byseasons or by climates, having high lipid contents, ability tobe produced from a wide variety of sources with short periodof time, and their similar fatty acid compositions to thatof vegetable oils [6, 7]. However, the high production costof SCO makes microbial oils less economically competitive.As a result, the production of microbial oils from wastes orrenewable materials is significantly important [8].

2 Journal of Biomedicine and Biotechnology
The oleaginous yeast Y. lipolytica is one of the mostextensively studied “nonconventional” yeasts found in envi-ronments rich in hydrophobic substrates, such as alkanesor lipids, and has developed sophisticated mechanisms forthe efficient use of hydrophobic substrates as its sole carbonsource [9]. One of the most striking features of this yeastis the presence in its genome of several multigene familiesinvolved in these metabolic pathways. The complexity andmultiplicity of these genes enable Y. lipolytica to use andvalorize a wide range of HS. Using these mechanisms,this yeast is able to accumulate large amount of lipids[10]. Studies about the uses of Y. lipolytica showed itspotential application for the production of reserve lipidswith composition resembling that of cocoa butter [11], lipidswith particular structures (e.g., oils enriched in essentialpolyunsaturated fatty acids), and nonspecific oils for use asrenewable starting materials for the synthesis of biofuels [12].
Lipid biosynthesis from sugars and related substratesis a secondary anabolic activity, conducted after essentialnutrient (usually nitrogen) depletion in the medium. Dueto this exhaustion, the carbon flow is directed towards theaccumulation of intracellular citric acid that is used as acetyl-CoA donor in the cytoplasm. Acetyl-CoA generates cellularfatty acids and subsequently triacylglycerols. SCO producedby Y. lipolytica cultivated under specific growth conditionscould be directly converted into biodiesel, as its fatty acidcomposition is similar to the one from common vegetableoils [13]. Studies related to the production of specialtylipids by the yeast Yarrowia lipolytica during growth onvarious fatty agroindustrial residues utilized as substratesshowed that the potential for growth on stearin (a low-costindustrial derivative of tallow composed of saturated freefatty acids) resulted in significant biomass production. Thiswas accompanied by notable intracellular accumulation oflipid which occurred as a primary anabolic activity regardlessof the extracellular nitrogen availability in the medium [14].
In order to reduce the cost of microbial oil productionfrom Y. lipolytica, low-cost raw materials, such as rice hull[15], industrial fats [16], industrial sugars [17], lignocellu-losic residues [18], and raw glycerol [19, 20], have been usedas substrates. Rice hull, one of the agricultural residues, isused for microbial oil production. According to Economouet al. [15], acid hydrolysis of rice hull using sulfuric acidresulted in hydrolysate that was used as feedstock for micro-bial lipids production with the oleaginous fungus Mortierellaisabellina. Results from kinetic experiments showed themaximum oil accumulation into fungal biomass to be 64.3%and suggested rice hull as cheap source of carbon.
Rice bran is also one of the most abundant agriculturalby-products in the world. Typical rice bran is composedof about 15–19.7% lipids, 34.1–52.3% carbohydrates, 7–11.4% fiber, 6.6–9.9% ash, and 10–15% proteins. After oilis being removed, the residual defatted rice bran (DRB)powder contains significant amount of starchy and cellulosicpolysaccharides. Enzymatic and chemical hydrolysis is usedto break the polysaccharides into smaller molecules whichwill be used as carbon sources for microorganisms [21].
The purpose of this study was to investigate the possi-bility of using cheap and easily available defatted rice bran
hydrolysate as a nutrient source for Y. lipolytica Po1g formicrobial oil production. The effects of acid concentration,reaction time, and temperature on the hydrolysis of DRBwere investigated. The effects of different types of acidhydrolysates of rice bran on the growth and lipid contentwere also studied. To the best of our knowledge, this is thefirst report to use defatted rice bran hydrolysate to culture Y.lipolytica Po1g for microbial oil production.
2. Materials and Methods
2.1. Materials. All solvents and reagents were either high-performance liquid chromatography (HPLC) or analyticalreagent grade, obtained from commercial sources. ForHPLC analysis, all the standards were purchased fromAcros Organics (USA) and Sigma Aldrich (USA). Thinlayer chromatography (TLC) aluminium plates (20× 20 cm)were obtained from Merck KGaA (Darmstadt, Germany).Qualitative filter papers (grade no. 2, 0.26 mm thickness,80% collection efficiency) were acquired from Advantec,MFS Inc. (Dublin, CA).
2.2. Defatted Rice Bran Hydrolysate (DRBH) Preparation
2.2.1. Raw Material and Pretreatment. Fresh rice bran waspurchased from a local rice mill in Taoyuan County, Taiwan.Bran collected from the mill was stored in a freezer at 4◦Cbefore use. Prior to defatting, it was dried at 50◦C for 24 huntil weight was constant. Defatting of rice bran was carriedout in a Soxhlet extractor with hexane as the solvent at 60◦Cfor 6 h. The defatted rice bran was kept at 4◦C for future use.
2.2.2. Acid Hydrolysis of Defatted Rice Bran. The defatted ricebran was hydrolyzed, following the method of Zhu et al. [4]with minor modification, by using H2SO4 (1%, 2%, 3%, or4%, v/v) with a bran to acid ratio of 1 : 8 (g/mL). The effectof temperature on hydrolysis was investigated at 60, 70, 80,90, 100, and 120◦C. The effect of hydrolysis time was alsoinvestigated. After hydrolysis, the mixture was subjected tovacuum filtration to obtain the defatted rice bran hydrolysate(DRBH).
2.3. Detoxification of DRBH. To reduce the concentrationof inhibitors in DRBH, neutralization with Ca(OH)2 wasemployed. Ca(OH)2 was slowly added to the DRBH bystirring the mixture at room temperature until the pH wasadjusted to 6.5. Then, the hydrolysate was repeatedly vacuumfiltrated until the precipitated particles were removed. Afterits composition was analyzed by HPLC, the detoxifiedhydrolysate was stored in a refrigerator at 4◦C for further useas nutrient for microbial fermentation.
2.4. Fermentation
2.4.1. Microorganism. Y. lipolytica Po1g cells were obtainedfrom YEASTERN Biotech Co. Ltd. (Taipei, Taiwan). Thestrain is a derivative of the wild-type strain W29 (ATCC20460) by a series of genetic modifications. The cells were

Journal of Biomedicine and Biotechnology 3
maintained on a sterilized yeast-, peptone-, and dextrose-(YPD-) agar plate containing 10 g/L yeast extract (Bacto,France), 10 g/L peptone (Bacto, France), 20 g/L glucose(Acros Organics, USA), and 20 g/L agar (Acros Organics,USA) at 4◦C for further microbial fermentation.
2.4.2. Media, Inoculum Preparation, and Large-Scale Fermen-tation. The preculture of Y. lipolytica Po1g was incubated ona sterilized YPD-agar plate and rejuvenated by incubationin 25 mL YPD medium containing yeast extract (10 g/L),peptone (10 g/L), and D-glucose (20 g/L) for 24 h at 26◦Cin an orbital shaker incubator model LM-570 (ChemistScientific Corp, Taiwan) and then inoculated to cultures in500 mL Erlenmeyer flasks at an inoculum to medium ratioof 1 : 10 (v/v).
Fermentation was carried out in several 500 mL Erlen-meyer flasks each containing 250 mL detoxified DRBH withinitial pH 6.5. To investigate the effect of different nitrogensources on growth of cells and lipid content, either urea(5 g/L, Acros Organics, USA) or peptone (5 g/L) was addedto the DRBH. The effect of the concentration of fermentablesugars in DRBH on microbial growth and lipid contentwas studied by diluting the sugar concentration obtainedat optimum conditions (48.41 g/L) and adjusted to 20 g/L,30 g/L, or 40 g/L.
The flasks were then incubated in an orbital shakerincubator at 150 rpm and 26◦C. Cell biomass was harvestedby centrifugation at 3500 g for 30 min. The cell masswas washed twice with deionized water and dried in anoven at 50◦C until constant weight. All media and flaskswere autoclaved at 121◦C for 30 min before the microbialfermentation.
2.5. Analytical Methods
2.5.1. Cell Concentration Determination. For monitoringyeast growth, optical density (OD) of the diluted fermen-tation broth was measured at 600 nm using a UV/Vis spec-trometer V-550 model (Jasco, Japan), and cell concentrationwas determined from a calibration curve of absorbanceversus dry cell weight.
2.5.2. Analysis of Protein, Sugar, and Inhibitor Concentrationin DRBH. The solubilized protein content of the hydrol-ysates was determined by the Bradford method [22].Absorbance at 595 nm was measured by using a UV-VISspectrophotometer V-550 model (Jasco, Japan), and thencomparison to a standard curve provided a relative measure-ment of protein concentration.
Concentrations of D-glucose, D-xylose, and L-arabinosein the hydrolysates were determined by HPLC (Jasco, Japan)equipped with a PU-1580 pump, a DG-4400 degasser, anAlltech 2000 ELSD detector, and a Zorbax NH2 column(5 µm particle size, 250 mm× 4.6 mm, Agilent Technologies,USA). The sample was diluted appropriately with deionizedwater, filtered through a 0.22 µm PVDF syringe filter (Testhigh) and then injected into the HPLC under the followingconditions. The temperature of the ELSD detector was
80◦C, and nitrogen flow rate was 2 mL/min. The columntemperature was 25◦C. Acetonitrile: water (80 : 20, v/v) wasthe eluent mobile phase with a flow rate of 1 mL/min, andthe injection volume was 25 µL.
The concentrations of 5-hydroxymethylfurfural (5-HMF) and furfural in the hydrolysate were determinedby HPLC (Jasco, Japan) equipped with a PU-2089 pumpcombined with degasser, an UV 2077 UV detector, and aLuna C-18 column (5 µm particle size, 250 mm × 4.6 mm,Phenomenex, USA). The sample was diluted appropriatelywith deionized water, filtered through a 0.22 µm PVDFsyringe filter (Test high), and then injected into the columnunder the following conditions: 25◦C column temperature,acetonitrile : water : acetic acid (11 : 88 : 1, v/v/v) with a flowrate of 0.8 mL/min, injection volume of 25 µL, and theabsorption wavelength was 276 nm. The concentrations ofthese compounds were calculated by using calibration curvesobtained from standard D-glucose, D-xylose, L-arabinose,furfural, and 5-HMF solutions.
The amount of total reducing sugars in the hydrolysatewas measured by the dinitrosalicylic acid (DNS) methodbased on a colorimetric reaction between the sugars anddinitrosalicylic acid. DNS reagent (0.5 mL) was added to theappropriately diluted sample (0.5 mL) in a capped brownbottle to prevent DNS from being affected by light. Then, themixture was heated to 100◦C for 5 min, and after cooling toroom temperature in a cold water bath, the absorbance wasmonitored with a spectrophotometer at 540 nm. The resultswere calculated based on calibration curve of standard D-glucose.
2.5.3. Lipid Analysis. Extraction of total lipid was performedby using Soxhlet extractor with hexane and methanol (1 : 1,v/v) for 4 h. The extracted lipid was subjected to silica gelthin layer chromatography (TLC) analysis to identify itsneutral lipid content. After that, the crude microbial lipidwas dewaxed and degummed according to the methodsdescribed by Rajam et al. [23], and Vandana et al. [24].Crude microbial oil was dissolved in hot water at 60◦C, andthe water soluble fraction was separated from the insolublefraction by vacuum filtration. The insoluble fraction wasthen dissolved in acetone and kept at 60◦C for 1 h toobtain clear solution. After allowing the content to coolto room temperature, the solution was then kept at 5◦Cfor 24 h to crystallize the remaining waxes. The insolublefraction was separated by vacuum filtration. The dewaxedand degummed lipid was analyzed by gas chromatography(GC-17A, Shimadzu, Japan) for its neutral lipid compositionas well as its fatty acid profile. The GC was equipped with aflame ionization detector and a DB-17HT capillary column(0.25 cm × 30 m, Agilent Technologies Inc., USA). Thecolumn temperature was programmed to increase from 80◦Cto 365◦C at 10◦C/min and kept at 365◦C for 29 min. Nitrogenwas used as the carrier gas at a flow rate of 0.80 mL/min.The split ratio was 1 : 50 (v/v). The temperatures of injectorand the detector were both maintained at 370◦C. Twentymilligrams sample was dissolved in 1 mL ethyl acetate, and0.5 µL sample was taken and injected into the GC. Standards

4 Journal of Biomedicine and Biotechnology
Table 1: Composition of DRBH with different reaction time and H2SO4 concentration at 90◦C.
H2SO4
(%v/v)Reactiontime (h)
Concentration (g/L)
Glucose Xylose Arabinose HMF Furfural
2%
2 13.36± 0.89 1.88± 0.42 1.13± 0.28 0.05± 0.02 0.004± 0.003
4 16.25± 0.94 2.34± 0.28 1.46± 0.19 0.16± 0.01 0.012± 0.002
6 21.26± 0.90 3.20± 0.28 1.97± 0.35 0.21± 0.02 0.020± 0.003
8 25.57± 0.76 3.84± 0.11 2.41± 0.16 0.28± 0.02 0.029± 0.020
3%
2 26.87± 0.64 4.61± 0.11 2.82± 0.07 0.13± 0.01 0.006± 0.002
4 36.67± 0.32 5.45± 0.043 2.23± 0.01 0.25± 0.01 0.015± 0.001
6 43.20± 0.28 4.93± 0.03 2.09± 0.01 0.32± 0.05 0.025± 0.003
8 40.12± 0.49 3.89± 0.05 1.46± 0.07 0.41± 0.02 0.041± 0.003
4%
2 28.80± 0.15 4.26± 0.02 2.83± 0.01 0.23± 0.01 0.011± 0.002
4 37.98± 0.19 4.60± 0.02 2.07± 0.81 0.40± 0.01 0.030± 0.003
6 43.64± 0.49 4.37± 0.16 1.87± 0.12 0.60± 0.08 0.053± 0.009
8 36.75± 1.79 3.78± 0.28 1.32± 0.21 0.88± 0.07 0.078± 0.005
of saturated and unsaturated fatty acids (Sigma-Aldrich,USA) were used for the identification of fatty acids in thelipid. All data were averages of triplicate determinations.
3. Result and Discussion
3.1. Acid Hydrolysis of DRB. Huang et al. [25] studied theproduction of microbial oil from T. fermentans grew insulfuric-acid-treated rice straw hydrolysate (SARSH). Thehydrolysate (SARSH) contained a maximum of 35.2 g/Lfermentable monosaccharides including glucose, xylose, andarabinose, and the concentration of pentoses was about sixtimes higher than hexose.
In this study, the composition of the defatted ricebran hydrolysate (DRBH) obtained by using 1% H2SO4
at different temperature (60◦C–120◦C) for 1 h showed thatglucose was the predominant sugar in the hydrolysate. Thisresult suggests the presence of a higher proportion of starchand glucans in DRB along with other minor pentosessuch as xylose and arabinose, which are mainly derivedfrom xylans and arabinoxylans, respectively. The maximumfermentable sugars concentration from DRBH under theseconditions was 5.96 ± 0.43 g/L out of which glucose, xylose,and arabinose are 3.16 ± 0.22 g/L, 1.36 ± 0.12 g/L, and0.74±0.01 g/L, respectively. However, the results suggest thathydrolysis of DRB for 1 h with 1% H2SO4 was not efficientto hydrolyze the DRB to produce fermentable sugars. Theoptimum temperature was 90◦C, and this was used as abasis for the next hydrolysis reactions with different acidconcentrations.
Table 1 shows the effects of H2SO4 concentrations (2%,3%, and 4%) and hydrolysis time (2–8 h) on the compositionof DRBH at 90◦C. The concentration of total fermentablesugars using 2% H2SO4 was much lower than that of 3% and4%, which shows that higher acid concentration could breakdown the polymerized structure of hemicellulose, amylase,and amylopectin more efficiently. The highest total releasedsugar concentration (50.22 ± 0.34 g/L) was obtained whenDRB was hydrolyzed with 3% H2SO4 at 90◦C for 6 h. These
optimum conditions resulted in glucose (43.20 ± 0.28 g/L),xylose (4.93 ± 0.03 g/L), arabinose (2.09 ± 0.01 g/L), HMF(0.32±0.05 g/L), and furfural (0.025±0.003 g/L). Comparedto the total fermentable sugars from SARSH (35.2 g/L), ourresults showed that rice bran has higher concentration offermentable sugars than sulfuric-acid-pretreated rice strawhydrolysate. On the other hand, the highest monosaccharidesugar concentration obtained in this study (43.20± 0.28 g/L)is glucose, while that of SARSH (25.5 g/L) is xylose. Thisindicates rice bran is a better source of glucose than rice strawfor microbial fermentation.
The total sugar obtained in this work was also muchhigher than that of enzymatic hydrolysis of DRB whichresulted in 37 g/L total sugars when defatted rice bran wassubjected to saccharification with a mixture of amylaseand cellulase [26]. Therefore, dilute acid hydrolysis breaksdown the lignocelluloses better than enzymes and can beconsidered as an efficient method to hydrolyze the DRB toobtain sugars for utilization in microbial fermentation.
The concentration of monosaccharides increases as timeincreases from 2 h to 8 h using 2% acid concentration.However, at an acid concentration of 3% or 4%, glucoseconcentration increased with time until 6 h and then startedto decrease. The same trend was observed for xylose and ara-binose. The rapid increase in the concentration of inhibitorswith longer hydrolysis time indicates the decomposition ofmonosaccharides into less desirable compounds under thesevere acid hydrolysis condition (4% H2SO4). A similar workalso reported that sugar concentration increases with longerhydrolysis time and higher acid concentration until theoptimum conditions were reached and decreases thereafter[27].
3.2. Effect of Detoxification on the Composition of DRBH.The presence of inhibitory substances in the fermentationmedium can restrict both sugar utilization and growthof microorganisms. It has been reported that oleaginousmicroorganisms could not grow and reproduce well in therice straw hydrolysate obtained by acid hydrolysis without
Journal of Biomedicine and Biotechnology 5
Table 2: Chemical composition of DRBH before and after detoxification. The hydrolysate used for detoxification was obtained by using 3%H2SO4 at 90◦C for 6 h.
Type ofDRBH
Concentration (g/L)
Glucose Xylose Arabinose HMF Furfural
2%Before detoxification 25.57± 0.76 3.84± 0.11 2.41± 0.16 0.28± 0.02 0.029± 0.020
After detoxification 25.40± 0.87 3.53± 0.43 2.03± 0.11 0.22± 0.01 0.023± 0.001
3% Before detoxification 43.20± 0.28 4.93± 0.03 2.09± 0.01 0.32± 0.05 0.025± 0.003
After detoxification 42.15± 0.39 4.71± 0.14 1.99± 0.09 0.24± 0.04 0.019± 0.005
4%Before detoxification 43.64± 0.49 4.37± 0.16 1.87± 0.12 0.60± 0.08 0.053± 0.009
After detoxification 42.99± 0.44 4.16± 0.75 1.70± 0.33 0.47± 0.16 0.044± 0.003
detoxification. Low lipid content was obtained after culturingT. fermentans for 8 days owing to both sugar utilizationand lipid accumulation being inhibited by various inhibitors[25]. Therefore, detoxification of DRBH is necessary toreduce the amount of inhibitors and achieve higher fer-mentability of sugars. Various methods such as overliming,neutralization, ion exchange, steam stripping, and treatmentwith activated carbon have been applied to detoxify thelignocellulosic hydrolysate. However, the most economicalway to detoxify lignocellulosic hydrolysate was neutralizationby calcium hydroxide [28].
Detoxification of DRBH by using Ca(OH)2 was appliedsince it can easily remove the residual SO4
2− by formingCaSO4 precipitate and efficiently reducing the level ofinhibitors simultaneously. The decrease in the concentra-tions of HMF and furfural is due to the reaction betweencalcium ions and furans, producing complex ions [29].
During the hydrolysis of DRB, the concentrations ofboth HMF and furfural increased with increasing acidconcentrations. Using 2% sulfuric acid, the concentrations ofHMF and furfural were 0.28 g/L and 0.029 g/L, respectively,and when the acid concentration increased from 3% to4%, the concentration of HMF and furfural also increasedfrom 0.32 g/L to 0.60 g/L and from 0.025 g/L to 0.053 g/L,respectively.
The maximum concentration of total released sugarsfrom nondetoxified DRBH at optimum conditions was50.22 ± 0.34 g/L, out of which the concentration of glucosewas 8.76 times higher than that of xylose. HMF andfurfural were the main inhibitors in the hydrolysate beforeneutralization. As shown in Table 2, the concentration ofmonosaccharides before and after detoxification by Ca(OH)2
did not show a significant change. However, the concen-trations of HMF and furfural decreased by 25.0% and24.0% (in the 3% DRBH), respectively, which indicates thatdetoxification by lime can effectively reduce the amount ofinhibitors and can provide a potential alternative medium forbetter microbial fermentation.
3.3. Growth of Y. lipolytica Po1g in Different Types of DRBHMedia. DRB is rich in carbohydrates, fiber, and proteins andcan be regarded as a potential nutrient source for microbialoil production [21]. Therefore, detoxified DRBH can be apotential nutrient source for growing Y. lipolytica Po1g.
0
2
4
6
8
10
12
14
16
18
20
22
0
2
4
6
8
10
Bio
mas
s (g
/L)
0 1 2 3 4 5 6
Fermentation time (day)
Suga
r co
nce
ntr
atio
n (
g/L
)
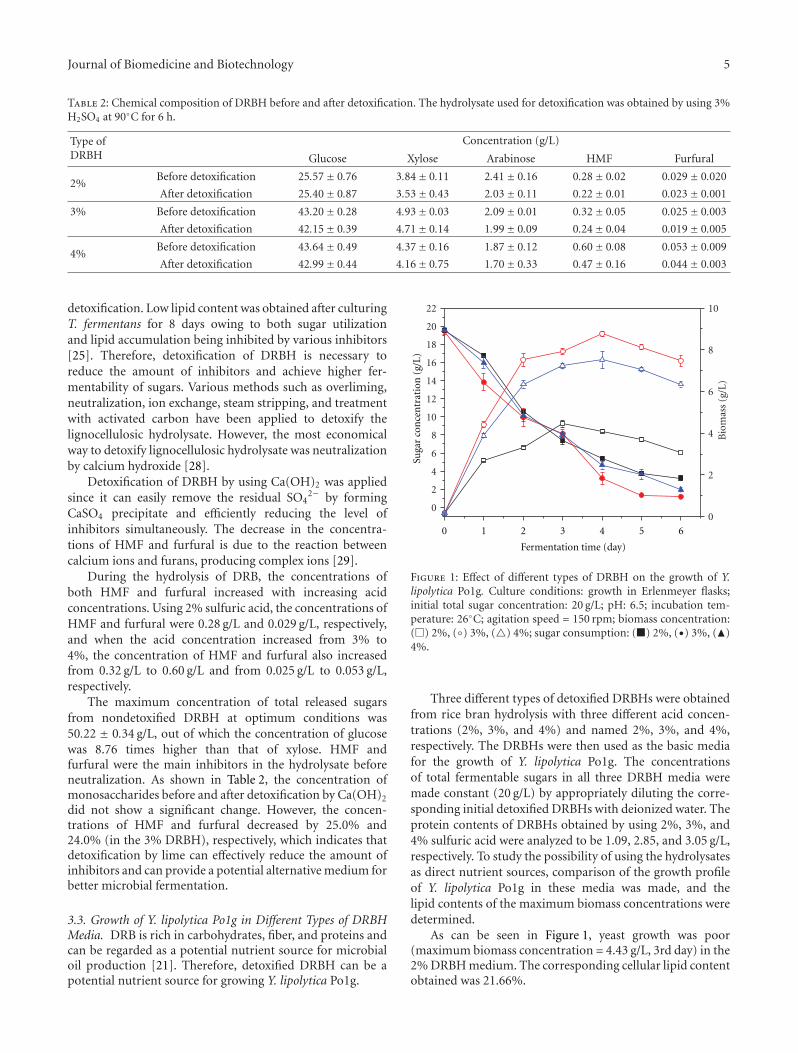
Figure 1: Effect of different types of DRBH on the growth of Y.lipolytica Po1g. Culture conditions: growth in Erlenmeyer flasks;initial total sugar concentration: 20 g/L; pH: 6.5; incubation tem-perature: 26◦C; agitation speed = 150 rpm; biomass concentration:(�) 2%, (◦) 3%, (�) 4%; sugar consumption: (�) 2%, (•) 3%, (�)4%.
Three different types of detoxified DRBHs were obtainedfrom rice bran hydrolysis with three different acid concen-trations (2%, 3%, and 4%) and named 2%, 3%, and 4%,respectively. The DRBHs were then used as the basic mediafor the growth of Y. lipolytica Po1g. The concentrationsof total fermentable sugars in all three DRBH media weremade constant (20 g/L) by appropriately diluting the corre-sponding initial detoxified DRBHs with deionized water. Theprotein contents of DRBHs obtained by using 2%, 3%, and4% sulfuric acid were analyzed to be 1.09, 2.85, and 3.05 g/L,respectively. To study the possibility of using the hydrolysatesas direct nutrient sources, comparison of the growth profileof Y. lipolytica Po1g in these media was made, and thelipid contents of the maximum biomass concentrations weredetermined.
As can be seen in Figure 1, yeast growth was poor(maximum biomass concentration = 4.43 g/L, 3rd day) in the2% DRBH medium. The corresponding cellular lipid contentobtained was 21.66%.

6 Journal of Biomedicine and Biotechnology
Increases in biomass concentration were observed whenY. lipolytica Po1g grew in the DRBH obtained by hydrol-ysis using 3% H2SO4. The highest biomass concentration(8.76 g/L) was observed on the 4th day. The cellular lipidcontent obtainable is 30.13% and is relatively higher than theone from the medium that contained the DRBH from 2%H2SO4. The sugar concentration in the medium preparedby hydrolysis of rice bran with 3% H2SO4 also decreasedfaster due to an increase in biomass. As the biomass increases,sugar consumption also increases. On the other hand, bothbiomass and lipid contents decreased slightly to 7.51 g/L and27.73%, respectively, when Y. lipolytica Po1g was cultivatedin the DRBH obtained by hydrolysis with 4% H2SO4.
According to a study conducted by Economou et al. [15],rice hulls hydrolysate was used as the sole carbon and energysource for oleaginous fungus M. isabellina. In their study,an increase in acid concentration from 0.03 M to 0.09 Mincreased the extractable oil amount from 36% to 64.3%, andit was observed that in the rice hulls hydrolysate obtainedfrom sulfuric acid concentration higher than 1 M, the fungusfailed to grow, probably due to the high concentrations oftoxic compounds in the culture medium. The same trend wasobserved in our study. The highest acid concentration (4%)might have led to higher concentration of inhibitors duringhydrolysis. This shows that neutralization of this hydrolysatewith Ca(OH)2 alone did not reduce the inhibitors concen-tration, and these inhibitors affected the growth of the yeastcells.
A review about lipid accumulation properties of Y.lipolytica shows that the study of de novo lipid accumulationas a result of the conversion of carbohydrate substrates,such as glucose, provides insight into the regulation of theentire lipid synthesis pathway. The accumulation of lipidsproduced from these substrates is triggered by nutrientlimitation. Nitrogen limitation is the principal type oflimitation governing lipid accumulation [30]. Our resultsalso suggested that limited nitrogen content in the media(3% DRBH) led to better growth and lipid accumulationwhile the protein content (1.09 g/L) in the medium from 2%H2SO4 was not enough for the growth of the cells, whichlimited biomass concentration to 4.43 g/L.
3.4. Effect of DRBH Sugar Concentration on Biomass andMicrobial Oil Content. To investigate the effect of sugarconcentrations on microbial growth and lipid accumulation,three different concentrations of total fermentable sugars(20 g/L, 30 g/L, and 40 g/L) were prepared by appropriatedilution of the detoxified DRBH (type 3%, Table 2) withan initial total sugar concentration of 48.41 g/L. The totalprotein content in each of these media was adjusted to 5 g/Lby addition of appropriate amount of yeast extract.
As shown in Table 3, the lowest cellular lipid content(30.13%) was obtained when the Y. lipolytica Po1g wasincubated in the medium with the lowest sugar concentra-tion (20 g/L). However, the highest lipid content (48.02%)and lipid yield (5.16 g/L) were observed when the sugarconcentration in the medium was 30 g/L. Both biomassconcentration and cellular lipid contents decreased when thetotal sugar concentration was 40 g/L.
Table 3: Effects of sugar concentration on microbial oil productionby Y. lipolytica Po1g.
Sugarconcentration(g/L)
Maximumbiomass (g/L)
Lipid content(%)
Lipid yield(g/L)
20 8.76± 0.13 30.13± 0.56 2.64± 0.08
30 10.75± 0.19 48.02± 0.61 5.16± 0.15
40 9.33± 0.16 40.97± 0.74 3.82± 0.13
Zhu et al. [4] investigated cell growth and lipid accumu-lation of T. fermentans in molasses with different total sugarconcentrations ranging from 5% to 30%. Both biomass andlipid content increased with the total sugar concentration,and the maximum biomass (29.9 g/L) and lipid content(31.8%) were attained at 15% total sugar concentration.Further rise in the total sugar concentration beyond 15%led to the decrease in biomass and lipid content. The sametrend was observed when Y. lipolytica Po1g was cultivated inDRBH with different sugar concentrations which increasedboth biomass and lipid content when sugar concentrationwas raised from 20 g/L to 30 g/L. But further increase to40 g/L resulted in lowering of biomass concentration andlower lipid content. This may be due to substrate inhibitionin which excess sugar concentration in the DRBH mediainhibits the growth of the microorganism during the earlystages of growth.
Another study on the biochemical behavior of wild-typeor genetically modified Y. lipolytica strains cultivated oncommercial glucose and nitrogen-limited cultures showedthat carbon-excess conditions favored the secretion oforganic acids into the growth medium [17]. Cultivation ofY. lipolytica Po1g using 40 g/L in this study also resultedin reduction of biomass concentration, which may mostprobably be due to the secretion of organic acids.
3.5. Effect of Addition of Different Nitrogen Sources on Growthand Microbial Oil Production. Factors such as carbon andnitrogen sources have significant influences on cell growthand lipid accumulation of oleaginous microorganisms [31].To study the effect of addition of different nitrogen sourceson biomass concentration and microbial oil production,three different DRBH media with the same total fermentablesugar concentration (30 g/L) and the same initial proteincontent (2.82 g/L) were prepared. No external nitrogensource was added to the first one, but either peptone (5 g/L)or urea (5 g/L) was added to the other two media.
As it can be seen from Figure 2, the medium withoutadditional nitrogen source was the best for yeast cells toaccumulate the highest lipid content (48.02%) with a lipidyield of 5.16 g/L. The addition of urea did not significantlyaffect (P-value = 0.27) biomass production (10.85 g/L) butcaused reduction in cellular lipid accumulation to 14.19%.Compared to urea, Y. lipolytica Po1g grew better, and themaximum biomass concentration was 13.53 g/L when thenitrogen source was peptone. The cellular lipid content(26.43%) obtained when cells grew in peptone was muchhigher than the one with urea, indicating that urea is not a

Journal of Biomedicine and Biotechnology 7
0
2
4
6
8
10
0
10
20
30
40
50
Biomass Lipid content Lipid yield
Bio
mas
s (g
/L),
lipi
d co
nte
nt
(%)
Urea PeptoneNo external nitrogenadded
Lip
id y
ield
(g/
L)
Figure 2: Effects of nitrogen addition on microbial oil productionby Y. lipolytica Po1g grown in DRBH medium. Culture conditions:growth in Erlenmeyer flasks; initial total sugar concentration:30 g/L; pH: 6.5; incubation temperature: 26◦C; agitation speed =150 rpm.
good nitrogen source for microbial oil production from Y.lipolytica Po1g grown in detoxified DRBH medium.
Microbial oil production with both nitrogen supple-ments showed similar tendencies in which the microorgan-isms did not accumulate more lipids but produced morebiomass than the one without external nitrogen source.Nitrogen is essential for the syntheses of proteins and nucleicacids which are required for cellular proliferation. However,this synthesis will be repressed when nitrogen source is lim-ited, and carbon source will be channeled to lipid synthesiscausing accumulation of lipid in the cells [32]. Therefore,nitrogen limitation is effective in enhancing the accumula-tion of lipid in Y. lipolytica Po1g grown in DRBH media.
Similar results by Zhu et al. [4] also showed that thecellular lipid content of T. fermentans grown in a mediumwith urea as the nitrogen source is 2.4 times lower than thatwhen peptone was the nitrogen source. The result of thisstudy shows that the yeast tended to channel sugar into cellu-lar lipid production when it was cultivated on the nitrogen-limited detoxified DRBH resulting in the highest cellularlipid content. On the other hand, according to Papanikolaouet al. [17], growth of either wild-type or genetically modifiedstrains of Y. lipolytica under nitrogen-limited culture con-ditions, with glucose or similarly metabolized compoundsused as substrates, was not accompanied by significantlipid accumulation. Carbon-excess conditions favored thesecretion of organic acids into the growth medium.
In general, various Y. lipolytica strains, despite being inca-pable of producing lipid via the de novo lipid accumulation
mechanism, are able to accumulate huge amounts of lipidwhen growth is carried out on various hydrophobic carbonsources as substrates, since accumulation of lipid from fatmaterials is a completely different process from that fromsugars [13].
3.6. Cellular Lipid Analysis. Crude microbial oil from Y.lipolytica Po1g cells grown in a medium with detoxifiedDRBH (3%) was investigated by TLC, and the contents weredetermined gravimetrically. After dewaxing and degum-ming, the neutral lipid content and the fatty acid profile weredetermined by GC 17A.
The crude microbial oil consisted of gum (52.77%),neutral lipids (41.92%), and wax (5.31%). The results ofanalysis of lipids isolated from the biomass of the yeast C.lipolytica grown in medium with methanol or glucose asthe only carbon source also showed that polar lipids makemore than half of the total cell lipids for both carbon sources(52.3 and 64.2%, resp.) [32]. This is in agreement with theresults of our work. On the other hand, Makri et al. [19]found that neutral lipids are the major components in Y.lipolytica when grown in a glycerol medium. According totheir study, lipid composition in Y. lipolytica showed specifictrends with time that largely reflect the physiological roleof individual lipids. The increase of neutral lipids duringlipogenic phase and their subsequent decrease in citric acidproduction phase may explain the physiological role ofneutral lipids, which is energy storage in times of plenty andenergy provision in times of shortage. In fact, during lipidturnover phase, neutral lipids are preferentially degraded,while some quantities of polar lipids are synthesized.
The GC chromatograms (Figures 3(a) and 3(b)) showthat the major components of neutral lipid are free fattyacids (FFA, 79.56%), monoacyl glycerides (MAG, 8.91%),diacyl glycerides (DAG, 2.15%), and triacyl glycerides (TAG,8.59%). Media, type of strain, and cultivation time havesignificant effect on the amount of FFA in lipids from Y.lipolytica. As reported by Papanikolaou et al. [16], growth ofY. lipolytica on medium containing hydrophobic compoundsas (co)substrates resulted in the accumulation of storage lipidcontaining considerable quantities of FFA (30 to 40 wt% oftotal lipids). They also found that large quantities of FFA didnot result in lethality or growth deficit, which is similar toour result. According to Juliano [21], rice bran is composedof about 15–19.7% lipids, which depends on factors such astype and origin of rice. The solvent used or extraction timeused in this study might have an effect in which some amountof oil has not been removed from rice bran. The excess oilmay remain in the medium and contribute to the increasein the FFA amount that can be extracted from Y. lipolyticaPo1g. The reason for the high FFA content needs furtherinvestigation.
The fatty acid profile of the neutral lipid consisted ofoleic acid (C18 : 1, 59.91%), palmitic acid (C16 : 0, 20.46%),palmitoleic acid (C16 : 1, 11.41%), and stearic acid (C18 : 0,5.39%), with 2.83% as unidentified fatty acids. The maincellular fatty acid of Y. lipolytica lipid produced duringgrowth on glucose or glycerol media also showed similarresults, and oleic acid, palmitic acid, palmitoleic acid, and

8 Journal of Biomedicine and Biotechnology
FFA
MAG DAGTAG
5 7.5 10 12.5 15 17.5 20 22.5 25 27.5
Retention time (min)
FID
res
pon
se
(a)
C16:0
C16:1
C18:1
5 7.5 10 12.5 15 17.5 20 22.5 25 27.5
Retention time (min)
FID
res
pon
se
OO
C18:0
O = others
(b)
Figure 3: GC chromatograms of neutral lipid before saponification (a) and fatty acid profile after saponification with KOH and ethanol (b).
Table 4: Lipid contents of different strains of Y. lipolytica grown in different agroindustrial sources.
Strain Carbon sourceBiomass
(g/L)
Lipidcontent
(%)
Lipid yield(g/L)
Reference
Y. lipolytica Industrial derivative of animal fat 12.5 54 3.8 [16]
C. lipolytica 8% molasses NA 59.9 NA [33]
Y. lipolytica LGAM (7)1 Teucrium polium L. aqueous extract 9.3 33 3.12 [34]
Y. lipolytica LGAM (7)1 Glucose 9.3 25 2.3 [34]
Y. lipolytica Industrial fats 8.7 44.0 3.8 [11]
Y. lipolytica ACA-YC 5033 Commercial glucose 5.5 14 NA [17]
C. lipolytica Mixtures of stearin, glucose, and glycerol 11.4 30 3.4 [35]
C. lipolytica 1% methanol NA 4.9 NA [32]
Y. lipolytica Po1g Sugarcane bagasse hydrolysate 11.42 58.5 6.68 [18]
Y. lipolytica Po1g Defatted rice bran hydrolysate 10.75 48.02 5.16 This study
stearic acid were the major fatty acids [17, 19]. The combinedcontent of saturated and monounsaturated fatty acid ismore than 97.17%, making neutral lipids obtained from theculturing of Y. lipolytica Po1g in rice bran hydrolysate idealfeedstock for biodiesel production.
Table 4 gives comparison of maximum biomass concen-tration, lipid content, and lipid yield of Y. lipolytica Po1ggrown in DRBH media with those from different strains ofY. lipolytica grown in different nutrient sources. In terms ofbiomass concentration, DRBH is a better nutrient sourcefor the Y. lipolytica Po1g compared to commercial glucoseused to cultivate Y. lipolytica ACA-YC 5033. This indicatesthe potential of defatted rice bran as a nutrient source foroleaginous microorganisms.
According to Karatay and Donmez [33], C. lipolytica canaccumulate lipids (59%, w/w) when grown in 8% molassesmedium. However, the maximum lipid concentration of Y.lipolytica Po1g in detoxified DRBH is 48.02%. On the otherhand, our work shows better results compared to that of Y.lipolytica cultivated in methanol and industrial fats whichresulted in a lipid content of 4.9% and 44%, respectively.Lipid yield of Y. lipolytica Po1g (5.6 g/L) is more promising
when compared to the results of different strains of Y.lipolytica.
In our previous study, Y. lipolytica Po1g cells werecultivated in sugarcane bagasse hydrolysate, and the maxi-mum biomass, lipid content, and lipid yield attainable were11.42 g/L, 58.5%, and 6.68 g/L, respectively [18]. Therefore,under different media, different strains of Y. lipolytica havedifferent lipid contents and lipid yields.
4. Conclusion
Defatted rice bran is an undervalued agricultural by-productwhich can be hydrolyzed by dilute acids not only torelease carbon sources from starch and lignocellulose butalso enough nitrogen sources for growing microorganisms.Detoxification of the defatted rice bran hydrolysate isrequired to reduce the level of inhibitors in the fermentationmedium. Detoxified DRBH was found to be an effectivemedium for cultivating Y. lipolytica Po1g, and high cellularlipid content was obtained. The composition of the neutrallipid obtained is similar to those of most staple vegetable oilsindicating its potentials for biodiesel production.

Journal of Biomedicine and Biotechnology 9
Acknowledgment
This work was supported by a Project (NSC 98-2221-E-011-046-MY3) from the National Science Council of Taiwan.
References
[1] A. K. Agarwal, “Biofuels (alcohols and biodiesel) applicationsas fuels for internal combustion engines,” Progress in Energyand Combustion Science, vol. 33, no. 3, pp. 233–271, 2007.
[2] F. Ma and M. A. Hanna, “Biodiesel production: a review,”Bioresource Technology, vol. 70, no. 1, pp. 1–15, 1999.
[3] M. J. Hass and T. A. Foglia, Eds., Alternative Feedstocks andTechnologies for Biodiesel Production, AOCS Press, Champaign,Ill, USA, 2005.
[4] L. Y. Zhu, M. H. Zong, and H. Wu, “Efficient lipid productionwith Trichosporon fermentans and its use for biodiesel prepa-ration,” Bioresource Technology, vol. 99, no. 16, pp. 7881–7885,2008.
[5] C. Ratledge, “Fatty acid biosynthesis in microorganisms beingused for Single Cell Oil production,” Biochimie, vol. 86, no. 11,pp. 807–815, 2004.
[6] Y. Li, Z. Zhao, and F. Bai, “High-density cultivation ofoleaginous yeast Rhodosporidium toruloides Y4 in fed-batchculture,” Enzyme and Microbial Technology, vol. 41, no. 3, pp.312–317, 2007.
[7] S. Papanikolaou, S. Sarantou, M. Komaitis, and G. Aggelis,“Repression of reserve lipid turnover in Cunninghamellaechinulata and Mortierella isabellina cultivated in multiple-limited media,” Journal of Applied Microbiology, vol. 97, no. 4,pp. 867–875, 2004.
[8] F. Xue, J. Miao, X. Zhang, H. Luo, and T. Tan, “Studieson lipid production by Rhodotorula glutinis fermentationusing monosodium glutamate wastewater as culture medium,”Bioresource Technology, vol. 99, no. 13, pp. 5923–5927, 2008.
[9] “Nonconventional yeasts in biotechnology,” in YarrowiaLipolytica, G. Barth and C. Gaillardin, Eds., pp. 313–388,Springer, New York, NY, USA, 1996.
[10] C. Ratledge, Ed., Single Cell Oils for the 21st Century, AOCSPress, Champaign, III, USA, 2005.
[11] S. Papanikolaou, I. Chevalot, M. Komaitis, G. Aggelis, andI. Marc, “Kinetic profile of the cellular lipid compositionin an oleaginous Yarrowia lipolytica capable of producinga cocoa-butter substitute from industrial fats,” Antonie vanLeeuwenhoek, vol. 80, no. 3-4, pp. 215–224, 2001.
[12] A. Beopoulos, J. Cescut, R. Haddouche, J. L. Uribelarrea, C.Molina-Jouve, and J. M. Nicaud, “Yarrowia lipolytica as amodel for bio-oil production,” Progress in Lipid Research, vol.48, no. 6, pp. 375–387, 2009.
[13] S. Papanikolaou and G. Aggelis, “Lipids of oleaginous yeasts—part I: biochemistry of single cell oil production,” EuropeanJournal of Lipid Science and Technology, vol. 113, no. 8, pp.1031–1051, 2011.
[14] S. Papanikolaou and G. Aggelis, “Yarrowia lipolytica: a modelmicroorganism used for the production of tailor-made lipids,”European Journal of Lipid Science and Technology, vol. 112, no.6, pp. 639–654, 2010.
[15] C. N. Economou, G. Aggelis, S. Pavlou, and D. V. Vayenas,“Single cell oil production from rice hulls hydrolysate,”Bioresource Technology, vol. 102, no. 20, pp. 9737–9742, 2011.
[16] S. Papanikolaou, I. Chevalot, M. Komaitis, I. Marc, and G.Aggelis, “Single cell oil production by Y. lipolytica growingon an industrial derivative of animal fat in batch cultures,”
Applied Microbiology and Biotechnology, vol. 58, no. 3, pp. 308–312, 2002.
[17] S. Papanikolaou, A. Chatzifragkou, S. Fakas et al., “Biosynthe-sis of lipids and organic acids by Yarrowia lipolytica strainscultivated on glucose,” European Journal of Lipid Science andTechnology, vol. 111, no. 12, pp. 1221–1232, 2009.
[18] Y. A. Tsigie, C. Y. Wang, C. T. Truong, and Y. H. Ju, “Lipidproduction from Yarrowia lipolytica Po1g grown in sugarcanebagasse hydrolysate,” Bioresource Technology, vol. 102, no. 19,pp. 9216–9222, 2011.
[19] A. Makri, S. Fakas, and G. Aggelis, “Metabolic activitiesof biotechnological interest in Yarrowia lipolytica grown onglycerol in repeated batch cultures,” Bioresource Technology,vol. 101, no. 7, pp. 2351–2358, 2010.
[20] A. Andre, A. Chatzifragkou, P. Diamantopoulou et al.,“Biotechnological conversions of bio-dieselderived crude glyc-erol by Yarrowia lipolytica strains,” Engineering in Life Sciences,vol. 9, no. 6, pp. 468–478, 2009.
[21] B. O. Juliano, Ed., Rice: Chemistry and Technolog, AACC, St.Paul, Minn, USA, 1985.
[22] M. M. Bradford, “A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein dye binding,” Analytical Biochemistry, vol.72, no. 1-2, pp. 248–254, 1976.
[23] L. Rajam, D. R. S. Kumar, A. Sundaresan, and C. Arumughan,“A novel process for physically refining rice bran oil throughsimultaneous degumming and dewaxing,” Journal of theAmerican Oil Chemists’ Society, vol. 82, no. 3, pp. 213–220,2005.
[24] V. Vandana, M. S. L. Karuna, P. Vijayalakshmi, and R. B. N.Prasad, “A simple method to enrich phospholipid contentin commercial soybean lecithin,” Journal of the American OilChemists’ Society, vol. 78, no. 5, pp. 555–556, 2001.
[25] C. Huang, M. H. Zong, H. Wu, and Q. P. Liu, “Microbialoil production from rice straw hydrolysate by Trichosporonfermentans,” Bioresource Technology, vol. 100, no. 19, pp. 4535–4538, 2009.
[26] T. Tanaka, M. Hoshina, S. Tanabe, K. Sakai, S. Ohtsubo, andM. Taniguchi, “Production of D-lactic acid from defattedrice bran by simultaneous saccharification and fermentation,”Bioresource Technology, vol. 97, no. 2, pp. 211–217, 2006.
[27] R. Gupta, K. K. Sharma, and R. C. Kuhad, “Separatehydrolysis and fermentation (SHF) of Prosopis juliflora, awoody substrate, for the production of cellulosic ethanolby Saccharomyces cerevisiae and Pichia stipitis-NCIM 3498,”Bioresource Technology, vol. 100, no. 3, pp. 1214–1220, 2009.
[28] T. D. Ranatunga, J. Jervis, R. F. Helm, J. D. McMillan, and R. J.Wooley, “The effect of overliming on the toxicity of dilute acidpretreated lignocellulosics: the role of inorganics, uronic acidsand ether-soluble organics,” Enzyme and Microbial Technology,vol. 27, no. 3-5, pp. 240–247, 2000.
[29] R. Purwadi, C. Niklasson, and M. J. Taherzadeh, “Kinetic studyof detoxification of dilute-acid hydrolyzates by Ca(OH)2,”Journal of Biotechnology, vol. 114, no. 1-2, pp. 187–198, 2004.
[30] C. Ratledge and J. P. Wynn, “The biochemistry and molecularbiology of lipid accumulation in oleaginous microorganisms,”Advances in Applied Microbiology, vol. 51, pp. 1–44, 2002.
[31] S. Wu, X. Zhao, H. Shen, Q. Wang, and Z. K. Zhao, “Microbiallipid production by Rhodosporidium toruloides under sulfate-limited conditions,” Bioresource Technology, vol. 102, no. 2, pp.1803–1807, 2011.
[32] J. Rupcic, B. Blagovic, and V. Maric, “Cell lipids of the Candidalipolytica yeast grown on methanol,” Journal of Chromatogra-phy A, vol. 755, pp. 75–80, 1996.

10 Journal of Biomedicine and Biotechnology
[33] S. E. Karatay and G. Donmez, “Improving the lipid accumula-tion properties of the yeast cells for biodiesel production usingmolasses,” Bioresource Technology, vol. 101, no. 20, pp. 7988–7990, 2010.
[34] G. Aggelis and M. Komaitis, “Enhancement of single cell oilproduction by Yarrowia lipolytica growing in the presence ofTeucrium polium L. aqueous extract,” Biotechnology Letters,vol. 21, no. 9, pp. 747–749, 1999.
[35] S. Papanikolaou, L. Muniglia, I. Chevalot, G. Aggelis, and I.Marc, “Accumulation of a cocoa-butter-like lipid by Yarrowialipolytica cultivated on agro-industrial residues,” CurrentMicrobiology, vol. 46, no. 2, pp. 124–130, 2003.

Submit your manuscripts athttp://www.hindawi.com
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporation http://www.hindawi.com
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
The Scientific World JournalHindawi Publishing Corporation http://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttp://www.hindawi.com
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
International Journal of
Microbiology