oleuropein aglycone, an olive oil compound,...
TRANSCRIPT

Oleuropein Aglycone, an Olive Oil Compound, AmelioratesDevelopment of Arthritis Caused by Injection of Collagen TypeII in Mice
Daniela Impellizzeri, Emanuela Esposito, Emanuela Mazzon, Irene Paterniti,Rosanna Di Paola, Valeria Maria Morittu, Antonio Procopio, Domenico Britti, andSalvatore CuzzocreaDepartment of Clinical and Experimental Medicine and Pharmacology, School of Medicine, University of Messina, Messina,Italy (D.I., E.E., I.P., S.C.); Istituto Di Ricovero e Cura a Carattere Scientifico Centro Neurolesi “Bonino-Pulejo,” Messina, Italy(E.M., R.D., S.C.); and Departments of Pharmacobiological Sciences (V.M.M., D.B.) and Clinical and Experimental Medicine(A.P.), University of Catanzaro Magna Graecia, Catanzaro, Italy
Received April 8, 2011; accepted August 30, 2011
ABSTRACTThe aim of this study was to investigate the effect of oleuropeinaglycone, an olive oil compound, on the modulation of theinflammatory response in mice subjected to collagen-inducedarthritis (CIA). CIA was induced in mice by an intradermalinjection of 100 �l of an emulsion containing 100 �g of bovinetype II collagen (CII) and complete Freund’s adjuvant (CFA) atthe base of the tail. On day 21, a second injection of CII in CFAwas administered. Mice developed erosive hind paw arthritiswhen immunized with CII in CFA. Macroscopic clinical evi-dence of CIA first appeared as periarticular erythema andedema in the hind paws. The incidence of CIA was 100% byday 28 in the CII-challenged mice and the severity of CIAprogressed over a 35-day period with resorption of bone. Thehistopathology of CIA included erosion of the cartilage at the
joint. Treatment with oleuropein aglycone starting at the onsetof arthritis (day 25) ameliorated the clinical signs at days 26 to35 and improved histological status in the joint and paw. Thedegree of oxidative and nitrosative damage was also signifi-cantly reduced in oleuropein aglycone-treated mice. Plasmalevels of the proinflammatory cytokines were also significantlyreduced by oleuropein aglycone. In addition, we have con-firmed the beneficial effects of oleuropein aglycone on an ex-perimental model of CIA in a therapeutic regimen of post-treatment, with treatment started at day 28, demonstrating thatoleuropein aglycone exerts an anti-inflammatory effect duringchronic inflammation and ameliorates the tissue damage asso-ciated with CIA.
IntroductionReactive oxygen species (ROS) have been considered as
risk and enhancer factors for autoimmune diseases (Filippinet al., 2008) because there is a significant relation betweenoxidative stress and such diseases (Avalos et al., 2007; Filip-pin et al., 2008). Rheumatoid arthritis (RA) is an autoim-
mune disease characterized by the sequestration of variousleukocyte subpopulations within both the developing pannusand synovial space. The chronic nature of this disease resultsin multiple joint inflammation with subsequent destructionof joint cartilage and erosion of bone. Although distribution ofthis disease is worldwide, its pathogenesis is not clearlyunderstood (Harris, 1990). Type II collagen-induced arthritis(CIA) in the mouse has proven to be a useful model of RA,because it possesses many of the cell and humoral immunitycharacteristics found in human RA (Holmdahl et al., 1990).The pathogenesis of CIA is dependent on the host’s responseto type II collagen challenge and the subsequent generationof antibodies that recognize collagen-rich joint tissue (Holm-dahl et al., 1990). The chronic activities initiated by immune
This study was supported by Istituto Di Ricovero e Cura a CarattereScientifico Centro Neurolesi “Bonino-Pulejo” and Ricerca ed Innovazione perL’olivicoltura Meridionale 2 del Ministero delle Politiche Agricole, Alimentarie Forestali e Consiglio per la Ricerca e la Sperimentazione in Agricoltura-Centro di Ricerca per l’Olivicoltura e l’Industria Olearia.
D.I. and E.E. contributed equally to this work.Article, publication date, and citation information can be found at
http://jpet.aspetjournals.org.doi:10.1124/jpet.111.182808.
ABBREVIATIONS: ROS, reactive oxygen species; RA, rheumatoid arthritis; CIA, type II collagen-induced arthritis; IL, interleukin; MIP, macrophageinflammatory protein; CII, collagen type II; iNOS, inducible nitric-oxide synthase; COX, cyclooxygenase; PGE2, prostaglandin E2; PAR, poly(ADP-ribose); CFA, complete Freund’s adjuvant; PBS, phosphate-buffered saline; ELISA, enzyme-linked immunosorbent assay; TNF-�, tumor necrosisfactor-�; MPO, myeloperoxidase; NF-�B, nuclear factor-�B; PARP, poly(ADP-ribosyl) polymerase.
0022-3565/11/3393-859–869$25.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 339, No. 3Copyright © 2011 by The American Society for Pharmacology and Experimental Therapeutics 182808/3728850JPET 339:859–869, 2011 Printed in U.S.A.
859
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

complexes trigger a variety of cell-mediated and humoralevents. Moreover, the recruitment and activation of neutro-phils, macrophages, and lymphocytes in joint tissues and theformation of the pannus are hallmarks of the pathogenesis ofboth CIA and human RA. It has been demonstrated thatinterleukin (IL)-8, macrophage inflammatory protein (MIP)-1�, MIP-1�, and regulated on activation normal T cell ex-pressed and secreted are differentially chemotactic for lym-phocyte subsets (Taub et al., 1993). Chemokines may playprominent roles in RA, because neutrophil and mononuclearcell stimulation and activation are prevalent in this disease.Concomitant with inflammation is the generation of ROS(Trichopoulou et al., 2003), which increase oxidation of pro-teins and lipids, resulting in signals that trigger more inflam-mation. Olive oil is an integral ingredient of the traditionalMediterranean diet, and several studies attribute many ofthe healthy advantages of this diet to olive oil’s unique char-acteristics (Menendez et al., 2007). Different studies havealso shown that the consumption of olive oil has a potentialprotective effect against several malignancies, especially can-cers (Menendez et al., 2007). The major constituent of theleaves and unprocessed olive drupes of Olea europaea isoleuropein and the majority of polyphenols found in olive oilor table olives are derived from its hydrolysis. Oleuropein hashigh antioxidant activity in vitro, comparable to that of ahydrosoluble analog of tocopherol (Kremastinos, 2008). Oleu-ropein scavenges superoxide anions and hydroxyl radicalsand inhibits the respiratory burst of neutrophils and hypo-chlorous acid-derived radicals (Visioli et al., 1998). In addi-tion to their antioxidant properties, polyphenolic compoundshave been shown to exhibit a range of indirect actions thatmay be beneficial to health, including the inhibition of en-zymes involved in the inflammatory process (Kohyama et al.,1997), the inhibition of platelet aggregation (Petroni et al.,1995), and the inhibition of the metabolic activation of pro-carcinogens (Stavric, 1994). The present study demonstratedthat oleuropein aglycone, a hydrolysis product obtained fromoleuropein by the action of �-glucosidase on the parent glu-coside (Walter et al., 1973) ameliorates development of ar-thritis caused by injection of collagen type II (CII) in mice.We have evaluated the following endpoints of the inflamma-tory process: 1) clinical score, 2) body weight, 3) induciblenitric-oxide synthase (iNOS) and cyclooxygenase (COX)-2 ex-pression and the serum levels of prostaglandin E2 (PGE2), 4)nitrotyrosine formation and poly(ADP-ribose) (PAR) expres-sion in the joint tissues, 5) cytokine and chemokine produc-tion, 6) neutrophil infiltration in the joint tissues, and 7) jointhistopathology.
Materials and MethodsAnimals. Male DBA/1J mice (9 weeks; Harlan Nossan, Milan,
Italy) were used for these studies. Mice were housed in individualcages (two for each group) and maintained under a 12-h light/darkcycle at 21 � 1°C and 50 � 5% humidity. The animals were accli-mated to their environment for 1 week and had ad libitum access totap water and a standard rodent diet. All animal experiments com-plied with regulations in Italy (Ministerial Decree 116192), Europe(Official Journal of Ethical Committee L 358/1 12/18/1986), and theUnited States (Animal Welfare Assurance No. A5594-01, U.S. De-partment of Health and Human Services). All behavioral testing wasconducted in compliance with the National Institutes of Health lab-oratory animal care guidelines (Institute of Laboratory Animal Re-
sources, 1996) and with protocols approved by the institutional an-imal care and use committee (Council Directive No. 87-848, October19, 1987, Ministere de l’Agriculture et de la Foret, Service Veteri-naire de la Sante et de la Protection Animale, permission no. 92-256to S.C.). The study was approved by the University of Messina reviewboard for the care of animals (PRIN ID 1042).
Experimental Groups. Mice were divided into the following sixexperimental groups:
1. CIA-control. Mice were subjected to collagen-induced arthritis(as described below) and administered 200 �l of 10% ethanolsolution (intraperitoneally, vehicle for oleuropein aglycone) every24 h, starting from day 25 to day 35 (n � 20).
2. CIA-oleuropein aglycone (pretreatment I). Mice subjected tocollagen-induced arthritis (as described below) were administeredoleuropein aglycone (molecular mass 394 kDa; 40 �g/kg, 10%ethanol i.p.) every 24 h, starting from day 25 to day 35 (n � 20).
3. CIA-oleuropein aglycone (pretreatment II). Mice subjectedto collagen-induced arthritis (as described below) were adminis-tered oleuropein aglycone (molecular mass 394 kDa; 20 �g/kg,10% ethanol i.p.) every 24 h, starting from day 25 to day 35(n � 20).
4. CIA-oleuropein aglycone (post-treatment). Mice subjected tocollagen-induced arthritis (as described below) were administeredoleuropein aglycone (molecular mass 394 kDa; 40 �g/kg, 10%ethanol i.p.) every 24 h, starting from day 28 to day 35 (n � 20).
5. Sham-control. Mice subjected to an intradermal injection at thebase of the tail of 100 �l of 0.01 M acetic acid instead of theemulsion containing 100 �g of CII were treated with 200 �l of 10%ethanol solution (i.p., vehicle for oleuropein aglycone), every 24 hstarting from day 25 to day 35 (n � 20).
6. Sham-oleuropein aglycone. Mice subjected to an intradermalinjection at the base of the tail of 100 �l of 0.01 M acetic acidinstead of the emulsion containing 100 �g of CII were adminis-tered oleuropein aglycone (40 �g/kg, 10% ethanol i.p.), every 24 hstarting from day 25 to day 35 (n � 20).
The dose and the route of oleuropein aglycone administration usedhere to reduce joint injury were chosen on the basis of previousstudies (Procopio et al., 2009; Impellizzeri et al., 2011).
Induction of CIA. Chicken type II collagen was dissolved in 0.01M acetic acid at a concentration of 2 mg/ml by stirring overnight at4°C. Dissolved CII was frozen at �70°C until use. Complete Freund’sadjuvant (CFA) was prepared by the addition of Mycobacteriumtuberculosis H37Ra at a concentration of 2 mg/ml. Before injection,CII was emulsified with an equal volume of CFA. Collagen-inducedarthritis was induced as described previously (Szabo et al., 1998). Onday 1, mice were injected intradermally at the base of the tail with100 �l of the emulsion containing 100 �g of CII. On day 21, a secondinjection of CII in CFA was administered.
Clinical Assessment of CIA. The development of arthritis inmice in all experimental groups was evaluated daily starting fromday 20 after the first intradermal injection by using a macroscopicscoring system: 0, no signs of arthritis; 1, swelling and/or rednessof the paw or one digit; 2, two joints involved; 3, more than twojoints involved; and 4, severe arthritis of the entire paw and digits(Szabo et al., 1998). The arthritic index for each mouse wascalculated by adding the four scores of individual paws. Clinicalseverity was also determined by quantitating the change in thepaw volume using plethysmometry (model 7140; Ugo Basile, Com-erio, Italy).
Histological Examination. On day 35, animals were sacrificedwhile they were under anesthesia (sodium pentobarbital, 45 mg/kgi.p.), and paws and knees were removed and fixed in 10% formalin.The paws were trimmed and decalcified in EDTA for 7 to 8 days asdescribed previously (Cuzzocrea et al., 2003a) and, after being dehy-drated in different graded alcohol concentrations, were embedded inparaffin, sectioned at 5 �m, stained with hematoxylin and eosin and
860 Impellizzeri et al.
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

Masson’s trichrome stain, and studied using light microscopy (LeitzDialux 22). Arthritis damage (histological damage score) was evalu-ated and scored by an investigator blinded for the treatment regi-men. The following morphological criteria were considered: score 0,no damage; score 1, edema; score 2, inflammatory cell presence; andscore 3, bone resorption.
Immunohistochemical Localization of Nitrotyrosine, PAR,iNOS, and COX-2. On day 35, the joints were trimmed and placedin a decalcifying solution of EDTA for 7 to 8 days. Then 8-�m sectionswere prepared from paraffin-embedded tissues. After deparaffiniza-tion, endogenous peroxidase was quenched with 0.3% H2O2 in 60%methanol for 30 min. The sections were permeabilized with 0.1%Triton X-100 in PBS for 20 min. Nonspecific adsorption was mini-mized by incubating the section in 2% normal goat serum in phos-phate-buffered saline for 20 min. Endogenous biotin- or avidin-bind-ing sites were blocked by sequential incubation for 15 min withavidin and biotin. Sections were incubated overnight with 1) anti-rabbit polyclonal antibody directed at iNOS (1:1000 in PBS, v/v)(DBA, Milan, Italy) or 2) anti-COX-2 goat polyclonal antibody (1:500in PBS, v/v) or 3) anti-nitrotyrosine rabbit polyclonal antibody (1:1000 in PBS, v/v) or 4) with anti-PAR goat polyclonal antibody rat(1:500 in PBS, v/v). Controls included buffer alone or nonspecific
purified rabbit IgG. Specific labeling was detected with a biotin-conjugated goat anti-rabbit IgG (for nitrotyrosine, iNOS, PAR, andCOX-2) and avidin-biotin peroxidase complex. To confirm that theimmunoreaction for the nitrotyrosine was specific, some sectionswere also incubated with the primary antibody (anti-nitrotyrosine)in the presence of excess nitrotyrosine (10 mM) to verify the bindingspecificity. To verify the binding specificity for PAR, COX-2, andiNOS, some sections were also incubated with only the primaryantibody (no secondary antibody) or with only the secondary anti-body (no primary antibody). In these situations, no positive stainingwas found in the sections, indicating that the immunoreaction waspositive in all the experiments performed. Immunocytochemistryphotographs (n � 5) were assessed by densitometry by using OptilabGraftek software on a Macintosh personal computer (Cuzzocrea etal., 2003b).
Measurement of PGE2 in the Serum. The amount of PGE2
present in the serum was measured with an ELISA kit (R&D Sys-tems, Milan, Italy). Blood serum samples were diluted 5 to 10 timesin assay diluents (BioFX, Milan, Italy), and 100 �l of the dilutionwere added in 96-well plates. No special treatment was necessary forstandard series. The rest of the process followed the kit provider’sprotocol.
Fig. 1. Effect of oleuropein aglycone(OLE AGLYCONE) on the clinical ex-pression of CIA and on body weight.A, no clinical signs were observed insham mice. CIA developed rapidly inmice immunized with CII and clinicalsigns such as periarticular erythemaand edema (B) were seen with a 100%incidence of CIA at day 28 (D). E, hindpaw erythema and swelling increasedin frequency and severity in a time-dependent mode. CIA-oleuropein agly-cone mice demonstrated a significantreduction in the clinical signs of CIA(C), leading to a decrease in the inci-dence of arthritis in a dose-dependentmanner (D). Swelling of hind paws(F) over time was measured at 2-dayintervals. G, beginning on day 25, theCII-challenged mice gained signifi-cantly less weight and this trend con-tinued through day 35. CIA-oleuropeinaglycone mice demonstrated a signifi-cant reduced incidence of weight loss(G) as well as less paw edema in a dose-dependent manner (F). The figure isrepresentative of all the animals ineach group. Values are means � S.E.M.of 20 animals for each group. ��, P �0.01 versus sham-control; °, P � 0.01versus CIA.
Oleuropein Aglycone and Arthritis 861
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

Measurement of Cytokines. TNF-�, IL-1�, and IL-6 levels wereevaluated in the plasma from CIA and sham mice as describedpreviously (Cuzzocrea et al., 2003b). The assay was performed usinga colorimetric commercial ELISA kit (Calbiochem-Novabiochem Cor-poration, Milan, Italy) with a lower detection limit of 10 pg/ml.
Measurement of Chemokines. Levels of chemokines MIP-1�and MIP-2 were measured in the aqueous joint extracts. In brief,joint tissues were prepared by first removing the skin and separatingthe limb below the ankle joint. Joint tissues were homogenized on icein 3 ml of lysis buffer [PBS containing 2 mM phenylmethylsulfonylfluoride and 0.1 mg/ml (final concentration) each of aprotinin, anti-pain, leupeptin, and pepstatin A] using a Polytron homogenizer(Brinkmann Instruments, Westbury, NY). The homogenized tissueswere then centrifuged at 2000g for 10 min. Supernatants were ster-ilized with a Millipore filter (0.2 �m) and stored at �80°C untilanalyzed. The extracts usually contained 0.2 to 1.5 mg of protein/ml,as measured by a protein assay kit (Thermo Fisher Scientific,Waltham, MA). The levels of MIP-1� and MIP-2 were quantifiedusing a modification of a double ligand method, as described previ-ously (Kasama et al., 1994). In brief, flat-bottomed 96-well microtiterplates were coated with 50 �l/well of rabbit anti-cytokine antibodies(R&D Systems, Milan, Italy) (1 �g/ml in 0.6 M NaCl, 0.26 M H3BO4,and 0.08 N NaOH, pH 9.6) for 16 h at 4°C and then were washed withPBS, pH 7.5, and 0.05% Tween 20 (wash buffer). Nonspecific bindingsites on microtiter plates were blocked with 2% bovine serum albu-min in PBS and incubated for 90 min at 37°C. Plates were rinsed fourtimes with wash buffer, and diluted aqueous joint samples (50 �l)were added, followed by incubation for 1 h at 37°C. After washing ofplates, chromogen substrate was added. The plates were incubatedat room temperature to the desired extinction, after which the reac-tion was terminated with 50 �l/well of 3 M H2SO4 solution. Theplates were then read at 490 nm in an ELISA reader. This ELISAmethod consistently had a sensitivity limit of �30 pg/ml.
Myeloperoxidase Assay. Neutrophil infiltration to the inflamedjoints was indirectly quantitated using an MPO assay, as describedpreviously for neutrophil elicitation (Mullane et al., 1985). Tissue
was prepared as described above and placed in 50 mM phosphatebuffer (pH � 6.0) with 5% hexadecyltrimethyl ammonium bromide(Sigma-Aldrich, Milan, Italy). Joint tissues were homogenized, son-icated, and centrifuged at 12,000g for 15 min at 4°C. Supernatantswere assayed for MPO activity using a spectrophotometric reactionwith O-dianisidine hydrochloride (Sigma-Aldrich) at 460 nm.
Materials. Oleuropein aglycone was obtained from the con-trolled hydrolysis of oleuropein extracted from olive leaves bymeans of the patented method reported by Procopio et al., 2009.Unless otherwise stated, other compounds were obtained fromSigma-Aldrich. All chemicals were of the highest commercialgrade available. All stock solutions were prepared in nonpyrogenicsaline (0.9% NaCl; Baxter Healthcare Ltd., Thetford, Norfolk, UK)or 10% ethanol (Sigma-Aldrich).
Data Analysis. All values in the figures and text are expressed asthe mean � S.E.M. of n observations. For the in vivo studies, nrepresents the number of animals studied. In the experiments in-volving histology or immunohistochemistry, the figures shown arerepresentative of at least three experiments (histological or immu-nohistochemistry coloration) performed on different experimentaldays on the tissue sections collected from all the animals in eachgroup. Data sets were examined by one- or two-way analysis ofvariance, and individual group means were then compared with aStudent’s unpaired t test. For the arthritis studies, a Mann-Whitney U test (two-tailed, independent) was used to comparemedians of the arthritic indices (Szabo et al., 1998). P � 0.05 wasconsidered significant.
ResultsEffect of Oleuropein Aglycone on Joint Injury dur-
ing Experimental Arthritis. To imitate the clinical sce-nario of RA, mice were subjected to CIA. CIA developedrapidly in mice immunized with CII, and clinical signs (peri-articular erythema and edema) (Fig. 1B) of the disease first
Fig. 2. Morphological changes of CIA.Representative hematoxylin and eo-sin-stained section of joint was exam-ined by light microscopy. The histo-logical evaluation (on day 35) of jointfrom CIA-control mice (B and G) re-vealed signs of severe arthritis, withinflammatory cell infiltration andbone erosion. The histological altera-tions of the joint were significantlyreduced in the tissues from CIA-oleu-ropein aglycone (OLE AGLYCONE)(40 �g/kg)-treated mice (C and G).Masson’s trichrome stain reveals de-creased collagen in bone and cartilageof arthritic joint due to bone erosionand cartilage degradation in CIA-con-trol mice (E and G). The alterations ofjoint were significantly reduced in oleu-ropein aglycone (40 �g/kg)-treated mice(F and G). There was no evidence ofpathology in the sham-control mice (A,D, and G). The histological score (G)was made by an independent observer.The figure is representative of at leastthree experiments performed on differ-ent experimental days. Values aremeans � S.E.M. of 20 animals for eachgroup. ��, P � 0.01 versus sham-con-trol; °, P � 0.01 versus CIA.
862 Impellizzeri et al.
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from
appeared in hind paws between 24 and 26 days postchallenge(Fig. 1E), leading to a 100% incidence of CIA at day 28 (Fig.1D). Hind paw erythema and swelling increased in frequencyand severity in a time-dependent mode with maximal scorevalues of approximately 12 that can be seen in Fig. 1E fromdays 32 to 35 after immunization in CIA-control mice. Oleu-ropein aglycone treatment (40 �g/kg, 10% ethanol and 20�g/kg, 10% ethanol) demonstrated a significant reduction ofjoint inflammation, as identified by a significant reduction inthe incidence of arthritis in a dose-dependent manner (Fig.1D). CIA-oleuropein aglycone mice showed a 40% reductionin the development of arthritis and a significantly lowerarthritis index compared with CIA-control mice (Fig. 1E).There was no macroscopic evidence of either hind paw ery-thema or edema in the sham-control group mice (Fig. 1A).
The data in Fig. 1F demonstrate a time-dependent in-crease in hind paw volume (each value represents the meanof both hind paws). The CIA-oleuropein aglycone miceshowed a significant reduction of paw edema formation in adose-dependent manner compared with the CIA-control mice
(Fig. 1F). No increase in hind paw volume over time wasobserved in the sham-control mice (data not shown).
The rate and the absolute gain in body weight were com-parable in sham-control and in CIA-control mice in the 1stweek (data not shown). From day 25, the CII-challenged micegained significantly less weight than the sham-control mice(data not shown), and this trend continued through to day 35.Oleuropein aglycone treatment determined a significant in-crease of the weight gain compared with the vehicle treat-ment in CIA-control mice (Fig. 1G).
The histological evaluation (on day 35) of the tibiotarsaljoint from CIA-control mice (Fig. 2B) revealed signs of severearthritis, with inflammatory cell infiltration and bone ero-sion. The histological alterations of the tibiotarsal joint weresignificantly reduced in oleuropein aglycone-treated mice (40�g/kg) (Fig. 2C). Moreover, Masson’s trichrome stain revealsdecreased collagen in bone and cartilage of arthritic jointsdue to bone erosion and cartilage degradation in CIA-controlmice (Fig. 2E). The alterations of the tibiotarsal joint weresignificantly reduced in oleuropein aglycone-treated mice (40
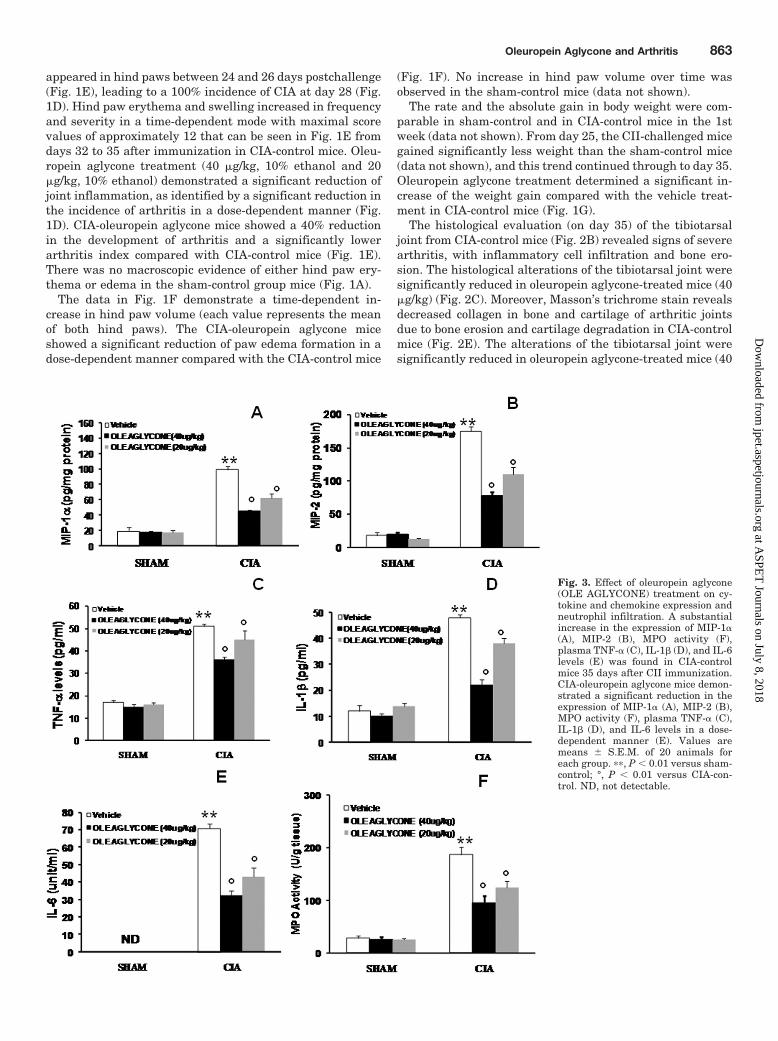
Fig. 3. Effect of oleuropein aglycone(OLE AGLYCONE) treatment on cy-tokine and chemokine expression andneutrophil infiltration. A substantialincrease in the expression of MIP-1�(A), MIP-2 (B), MPO activity (F),plasma TNF-� (C), IL-1� (D), and IL-6levels (E) was found in CIA-controlmice 35 days after CII immunization.CIA-oleuropein aglycone mice demon-strated a significant reduction in theexpression of MIP-1� (A), MIP-2 (B),MPO activity (F), plasma TNF-� (C),IL-1� (D), and IL-6 levels in a dose-dependent manner (E). Values aremeans � S.E.M. of 20 animals foreach group. ��, P � 0.01 versus sham-control; °, P � 0.01 versus CIA-con-trol. ND, not detectable.
Oleuropein Aglycone and Arthritis 863
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

�g/kg) (Fig. 2F). There was no evidence of pathology in thesham-control mice (Fig. 2, A and D). The histological score(Fig. 2G) was determined by an independent observer.
Effect of Oleuropein Aglycone on Chemokine andCytokine Expression and Neutrophil Infiltration. Weinitiated studies to assess the effect of oleuropein aglycone onthe expression of chemokines in the aqueous joint extractsduring the development of CIA. As shown in Fig. 3, A and B,the expression of MIP-1� and MIP-2, measured by ELISA,was significantly increased in the joint 35 days after CIIimmunization. MIP-1� and MIP-2 levels in CIA-oleuropeinaglycone mice on day 35 were significantly reduced in adose-dependent manner in comparison with those in vehicle-treated CIA-control mice. Assessment of neutrophil infiltra-tion into the inflamed joint tissue was performed by measur-ing the activity of MPO. It was significantly elevated 35 daysafter CII immunization in vehicle-treated CIA-control mice(Fig. 3F), whereas in the CIA-oleuropein aglycone group,
MPO activity was markedly reduced in a dose-dependentmanner (Fig. 3F). To test whether oleuropein aglycone mod-ulates the inflammatory process through the regulation ofcytokine secretion, we analyzed the plasma levels of theproinflammatory cytokines TNF-�, IL-1�, and IL-6. A sub-stantial increase in TNF-� (Fig. 3C), IL-1� (Fig. 3D), andIL-6 (Fig. 3E) production was found in CIA-control mice 35days after CII immunization. Levels of TNF-� (Fig. 3C),IL-1� (Fig. 3D), and IL-6 (Fig. 3E) were significantly reducedin a dose-dependent manner in CIA-oleuropein aglycone micein comparison to CIA-control mice.
Effect of Oleuropein Aglycone Treatment on iNOS,COX-2, PGE2, Nitrotyrosine, and PAR Formation. Im-munohistochemical analysis of the tibiotarsal joint sectionsobtained from CIA-control mice revealed positive staining foriNOS (Fig. 5, A and A1) and COX-2 (Fig. 4, A and A1), whichwere primarily localized in inflammatory cells. In contrast,staining for iNOS (Fig. 5B) and COX-2 (Fig. 4B) was mark-
Fig. 4. Effect of oleuropein aglycone(OLE AGLYCONE) treatment onCOX-2 immunostaining and on serumPGE2 levels. A marked increase inCOX-2 (A and in particular A1) stain-ing was evident in the paw 35 daysafter initiation of CIA. There was amarked reduction in the immuno-staining for COX-2 (B) in the paw ofCIA-oleuropein aglycone (40 �g/kg)mice. To verify the binding specificityfor COX-2, some sections were alsoincubated with only the secondary an-tibody (no primary antibody). No pos-itive staining for COX-2 was found inthe sections indicating that the immu-noreaction was positive (see negativecontrol C). In addition, a marked in-crease of PGE2 levels was found in theserum of CIA control mice 35 daysafter CII immunization (E). The treat-ment with oleuropein aglycone alsocaused a significant reduction in adose-dependent manner of the serumlevels of the metabolite of COX-2 (E).The figure is representative of at leastthree experiments performed on dif-ferent experimental days. Densitome-try analysis of immunocytochemistryphotographs (n � 5) for COX-2 frompaw section was assessed (D). The as-say was performed by using OptilabGraftek software on a Macintosh per-sonal computer (CPU G3-266). Dataare expressed as a percentage of totaltissue area. ��, P � 0.01 versus sham-control; °, P � 0.01 versus CIA. ND,not detectable.
864 Impellizzeri et al.
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

edly reduced in the tibiotarsal joints of CIA-oleuropein agly-cone (40 �g/kg) mice. No staining for either iNOS or COX-2was detected in the tibiotarsal joints obtained from sham-control mice (data not shown). Moreover, we also evaluatedthe levels of PGE2, the metabolite of COX-2, in the serumduring the development of CIA. A substantial increase inPGE-2 production was found in CIA-control mice 35 daysafter CII immunization (Fig. 4E). Levels of PGE-2 were sig-nificantly reduced in CIA-oleuropein aglycone mice in a dose-dependent manner compared with those in CIA-control mice(Fig. 4E).
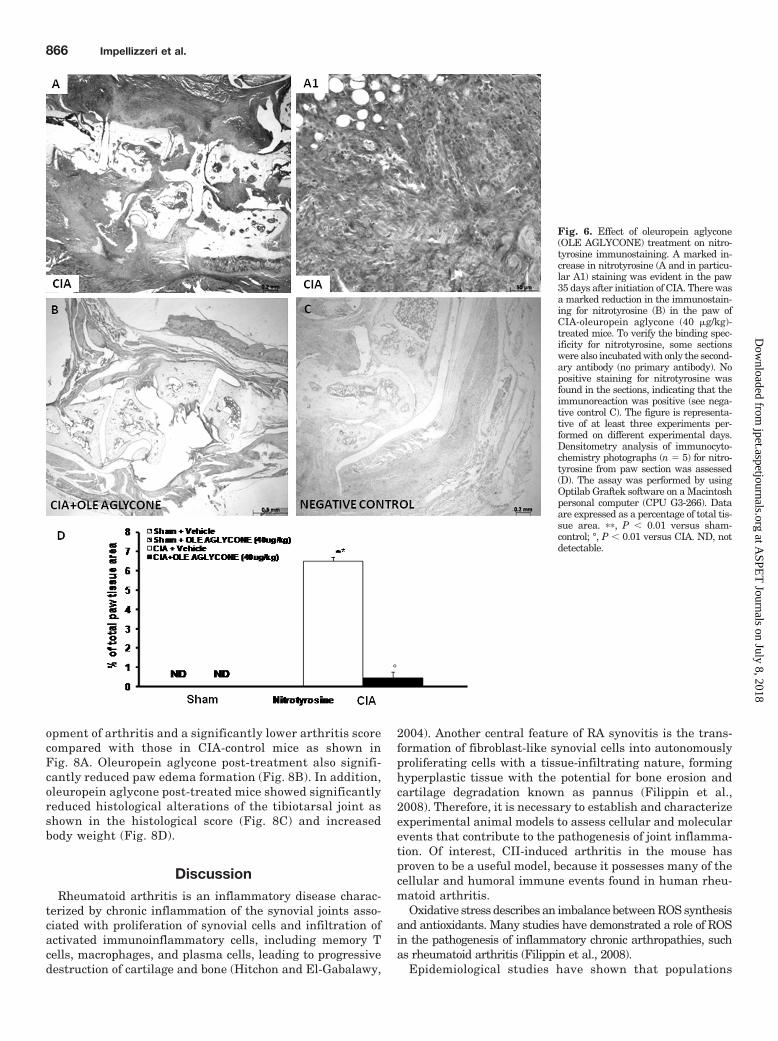
The release of free radicals and oxidant molecules duringchronic inflammation has been suggested to contribute sig-nificantly to the tissue injury (Cuzzocrea et al., 2001). On day35, positive staining for nitrotyrosine, a marker of nitrosativeinjury, was found in the tibiotarsal joints of vehicle-treatedCIA-control mice (Fig. 6, A and A1). Oleuropein aglycone (40
�g/kg) treatment significantly reduced the formation of ni-trotyrosine (Fig. 6B).
Immunohistochemical analysis of joint sections obtainedfrom CII-challenged mice revealed positive staining forPAR (Fig. 7, A and A1). In contrast, no positive PAR wasfound in the tibiotarsal joints of CII-challenged micetreated with oleuropein aglycone (40 �g/kg) (Fig. 7B).There was no staining for either nitrotyrosine or PAR inthe tibiotarsal joints obtained from sham-control mice(data not shown).
Oleuropein Aglycone Inhibits the Progression ofEstablished Arthritis. To confirm that oleuropein agly-cone exerts beneficial effects in the experimental model ofcollagen-induced arthritis, we have also evaluated its ef-fect in a therapeutic regimen of post-treatment (40 �g/kg),starting the treatment at day 28. CIA-oleuropein aglyconepost-treatment mice also showed a reduction in the devel-
Fig. 5. Effect of oleuropein aglycone(OLE AGLYCONE) treatment oniNOS immunostaining. A marked in-crease in iNOS (A and in particularA1) staining was evident in the paw35 days after initiation of CIA. Therewas a marked reduction in the immu-nostaining for iNOS (B) in the paw ofCIA-oleuropein aglycone (40 �g/kg)mice. To verify the binding specificityfor iNOS, some sections were also in-cubated with only the secondary anti-body (no primary antibody). No posi-tive staining for iNOS was found inthe sections, indicating that the im-munoreaction was positive (see nega-tive control C). The figure is represen-tative of at least three experimentsperformed on different experimentaldays. Densitometry analysis of immu-nocytochemistry photographs (n � 5)for iNOS from paw section was as-sessed (D). The assay was performedby using Optilab Graftek software ona Macintosh personal computer (CPUG3-266). Data are expressed as a per-centage of total tissue area. ��, P �0.01 versus sham-control; °, P � 0.01versus CIA. ND, not detectable.
Oleuropein Aglycone and Arthritis 865
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from
opment of arthritis and a significantly lower arthritis scorecompared with those in CIA-control mice as shown inFig. 8A. Oleuropein aglycone post-treatment also signifi-cantly reduced paw edema formation (Fig. 8B). In addition,oleuropein aglycone post-treated mice showed significantlyreduced histological alterations of the tibiotarsal joint asshown in the histological score (Fig. 8C) and increasedbody weight (Fig. 8D).
DiscussionRheumatoid arthritis is an inflammatory disease charac-
terized by chronic inflammation of the synovial joints asso-ciated with proliferation of synovial cells and infiltration ofactivated immunoinflammatory cells, including memory Tcells, macrophages, and plasma cells, leading to progressivedestruction of cartilage and bone (Hitchon and El-Gabalawy,
2004). Another central feature of RA synovitis is the trans-formation of fibroblast-like synovial cells into autonomouslyproliferating cells with a tissue-infiltrating nature, forminghyperplastic tissue with the potential for bone erosion andcartilage degradation known as pannus (Filippin et al.,2008). Therefore, it is necessary to establish and characterizeexperimental animal models to assess cellular and molecularevents that contribute to the pathogenesis of joint inflamma-tion. Of interest, CII-induced arthritis in the mouse hasproven to be a useful model, because it possesses many of thecellular and humoral immune events found in human rheu-matoid arthritis.
Oxidative stress describes an imbalance between ROS synthesisand antioxidants. Many studies have demonstrated a role of ROSin the pathogenesis of inflammatory chronic arthropathies, suchas rheumatoid arthritis (Filippin et al., 2008).
Epidemiological studies have shown that populations
Fig. 6. Effect of oleuropein aglycone(OLE AGLYCONE) treatment on nitro-tyrosine immunostaining. A marked in-crease in nitrotyrosine (A and in particu-lar A1) staining was evident in the paw35 days after initiation of CIA. There wasa marked reduction in the immunostain-ing for nitrotyrosine (B) in the paw ofCIA-oleuropein aglycone (40 �g/kg)-treated mice. To verify the binding spec-ificity for nitrotyrosine, some sectionswere also incubated with only the second-ary antibody (no primary antibody). Nopositive staining for nitrotyrosine wasfound in the sections, indicating that theimmunoreaction was positive (see nega-tive control C). The figure is representa-tive of at least three experiments per-formed on different experimental days.Densitometry analysis of immunocyto-chemistry photographs (n � 5) for nitro-tyrosine from paw section was assessed(D). The assay was performed by usingOptilab Graftek software on a Macintoshpersonal computer (CPU G3-266). Dataare expressed as a percentage of total tis-sue area. ��, P � 0.01 versus sham-control; °, P � 0.01 versus CIA. ND, notdetectable.
866 Impellizzeri et al.
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

consuming a predominantly olive oil-based Mediterra-nean-style diet exhibit lower incidences of breast cancerand other chronic diseases (Menendez et al., 2007). A fun-damental characteristic of olive oil is the presence of alarge number of phenolic compounds such as oleuropein, aglucoside with hydroxyaromatic functionality, which hasattracted considerable attention because of its antidia-betic, antiatherosclerotic (Miles et al., 2005), and anti-inflammatory (Covas, 2008) properties. Several studieshave also shown that olive oil phenols possess potent an-tioxidant activity (Vissers et al., 2004) and prevent reac-tive oxygen species-mediated cell injury (Salvini et al.,2006). A more efficient anti-inflammatory role of the agly-conic compared with the glycosidic form of oleuropein pos-sibly derives from the greater lipophilicity of the former, aproperty that should allow better cell membrane incorpo-ration and/or interaction with other lipids (Saija et al.,1998). In this regard, we investigate here the effects of
oleuropein aglycone, a hydrolysis product of oleuropein, ina mouse model of CII-induced arthritis.
Although T-cell and antibody responses against CII area crucial event for the initiation of CIA (Holmdahl et al.,1989), it has been demonstrated that several cytokinesalso appear to direct cell-to-cell communication in a cas-cade fashion during the progression of CIA such as IL-1(Hom et al., 1992), TNF-� (Dong et al., 2010), and IL-6(Ferraccioli et al., 2010). In addition, it has been demon-strated that monocyte chemotactic protein-1, MIP-1�,MIP-1�, and regulated on activation normal T cell ex-pressed and secreted are differentially chemotactic forlymphocyte subsets and are expressed in tissue from theinflamed joints of patients with rheumatoid arthritis(Koch et al., 1994). In this study, we have confirmed thatthe cytokines (TNF-�, IL-1�, and IL-6) as well as thechemokines (MIP-1� and MIP-2) are expressed at sites ofinflamed joints and probably contribute in different capac-
Fig. 7. Effect of oleuropein aglycone(OLE AGLYCONE) treatment onPARP immunostaining. A marked in-crease in PARP (A and in particularA1), staining was evident in the paw35 days after initiation of CIA. Therewas a marked reduction in the immu-nostaining for PARP (B) in the paw ofCIA-oleuropein aglycone (40 �g/kg)-treated mice. To verify the bindingspecificity for PARP, some sectionswere also incubated with only the sec-ondary antibody (no primary anti-body). No positive staining for PARPwas found in the sections, indicatingthat the immunoreaction was positive(see negative control C). The figure isrepresentative of at least three exper-iments performed on different experi-mental days. Densitometry analysisof immunocytochemistry photographs(n � 5) for PARP from paw sectionwas assessed (D). The assay was per-formed by using Optilab Graftek soft-ware on a Macintosh personal computer(CPU G3-266). Data are expressed as apercentage of total tissue area. ��, P �0.01 versus sham-control; °, P � 0.01 ver-sus CIA. ND, not detectable.
Oleuropein Aglycone and Arthritis 867
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

ities to the progression of chronic joint inflammation. Sev-eral cytokines, including TNF-� and IL-1�, are knowninitiators of the nuclear factor (NF-B) activation cascade(Filippin et al., 2008) and are under its transcriptionalcontrol, constituting a positive feedback loop. Recent stud-ies have observed that the acute consumption of olive oildecreased the activation of the NF-B system on mononu-clear cells from healthy men (Perez-Martinez et al., 2007)and that oleuropein aglycone, trans-resveratrol, and hy-droxytyrosol incubated with human umbilical vein endo-thelial cells inhibit lipopolysaccharide-triggered NF-Band activator protein-1 activation (Carluccio et al., 2003).Of interest, using oleuropein aglycone, we have demon-strated an inhibition of the release of proinflammatory cyto-kines and chemokines and a reduction of leukocyte infiltrationmeasured by MPO activity. Several studies also showed thatthe potential cardioprotective activity of oleuropein in acutecardiotoxicity induced by doxorubicin treatment was deter-mined in vivo in rats (Andreadou et al., 2007) by inhibiting lipidperoxidation products, decreasing oxidative stress, and reduc-ing iNOS in cardiomyocytes and that the olive oil polyphenolsare capable of down-regulating COX-2 expression in coloniccancer cells by a mechanism involving the early inhibition ofp38 mitogen-activated protein kinase and downstream inhibi-tion of the transcription factor cAMP response element-bindingprotein (Corona et al., 2007). We show here that oleuropeinaglycone decreased iNOS and COX-2 expression by immuno-histochemical staining and also reduced the levels of the me-tabolite of COX-2, PGE2, in the serum of oleuropein aglycone-treated mice.
Reactive nitrogen species, such as the peroxynitrite radical(ONOO�) generated by the reaction between O2
. and nitric oxide,can also cause oxidative damage (Soneja et al., 2005). The additionof ONOO� to body cells, tissues, and fluids leads to fast pro-tonation, which may result in the depletion of �SH groupsand other antioxidants, oxidation and nitration of lipids,DNA disruption, and nitration and deamination of DNAbases (Filippin et al., 2008). In this report, an intenseimmunostaining of nitrotyrosine formation also suggestedthat a structural alteration of joint had occurred, mostprobably due to the formation of highly reactive nitrogenderivatives.
ROS produce strand breaks in DNA, which triggers en-ergy-consuming DNA repair mechanisms and activates thenuclear enzyme poly(ADP-ribosyl) polymerase (PARP).There is various evidence that the activation of PARP mayalso play an important role in inflammation (Genovese etal., 2005). Continuous or excessive activation of PARPproduces extended chains of ADP-ribose (PAR) on nuclearproteins and results in a substantial depletion of intracel-lular NAD and subsequently, ATP, leading to cellulardysfunction and, ultimately, cell death (Chiarugi, 2002).We demonstrate here that oleuropein aglycone treatmentreduced the activation of PARP with a decrease in PARexpression in the joint during CIA. In this regard, severalstudies demonstrated that hydroxytyrosol, a hydrolysisproduct of oleuropein aglycone, also exerts an inhibitoryeffect on peroxynitrite-dependent DNA base modificationsand tyrosine nitration (Deiana et al., 1999). Likewise, Sal-vini et al. (2006) showed a 30% reduction of oxidative DNAdamage in peripheral blood lymphocytes during interven-tion in postmenopausal women with virgin olive oil con-taining high amounts of phenols. Thus, oleuropein agly-cone, given at the onset of the disease, reduced pawswelling, clinical score, and the histological severity of thedisease when injected after the onset of clinical arthritis.Amelioration of joint disease was associated with near tofull inhibition of cytokines as well as inhibition of neutro-phil infiltration, which is a key player in RA.
Therefore, oleuropein aglycone was also administeredfrom day 28 after collagen immunization, targeting thisearly initiation phase of CIA. Then, with treatment start-ing at day 28, oleuropein aglycone post-treatment caused asignificant reduction of inflamed joints collected at day 35.
In conclusion, RA is a complex chronic inflammatorydisease dependent on multiple interacting environmentaland genetic factors, making it difficult to understand itspathogenesis and thereby to find effective therapies.Taken together, the results of the present study enhanceour understanding of the role of ROS generation in thepathophysiology of CII-induced arthritis, implying thatolive oil compounds such as oleuropein aglycone may beuseful in the therapy of inflammation.
Fig. 8. Effect of oleuropein aglycone (OLE AGLYCONE)post-treatment on joint inflammation. Starting the treat-ment at day 28, we have also demonstrated that oleuropeinaglycone post-treatment (40 �g/kg) caused a significantlylower arthritis score (A) and a reduction of foot increase (B)compared with the CIA-control. In addition, we have alsoshown a reduction in the histological damage (C) and in-creased body weight (D) in oleuropein aglycone-treatedmice. Data are expressed as a percentage of total tissuearea. ��, P � 0.01 versus sham-control; °, P � 0.01 versusCIA.
868 Impellizzeri et al.
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from

Acknowledgments
We thank Carmelo La Spada for excellent technical assistanceduring this study, Caterina Cutrona for secretarial assistance, andValentina Malvagni for editorial assistance with the manuscript.
Authorship Contributions
Participated in research design: Britti and Cuzzocrea.Conducted experiments: Impellizzeri, Mazzon, Paterniti, Di Paola,
and Morittu.Contributed new reagents or analytic tools: Morittu and Procopio.Performed data analysis: Di Paola, Morittu, Britti, and Cuzzocrea.Wrote or contributed to the writing of the manuscript: Impellizzeri,
Esposito, and Cuzzocrea.
ReferencesAndreadou I, Sigala F, Iliodromitis EK, Papaefthimiou M, Sigalas C, Aligiannis N,
Savvari P, Gorgoulis V, Papalabros E, and Kremastinos DT (2007) Acute doxoru-bicin cardiotoxicity is successfully treated with the phytochemical oleuropeinthrough suppression of oxidative and nitrosative stress. J Mol Cell Cardiol 42:549–558.
Avalos I, Chung CP, Oeser A, Milne GL, Morrow JD, Gebretsadik T, Shintani A, YuC, and Stein CM (2007) Oxidative stress in systemic lupus erythematosus: rela-tionship to disease activity and symptoms. Lupus 16:195–200.
Carluccio MA, Siculella L, Ancora MA, Massaro M, Scoditti E, Storelli C, Visioli F,Distante A, and De Caterina R (2003) Olive oil and red wine antioxidant polyphe-nols inhibit endothelial activation: antiatherogenic properties of Mediterraneandiet phytochemicals. Arterioscler Thromb Vasc Biol 23:622–629.
Chiarugi A (2002) Poly(ADP-ribose) polymerase: killer or conspirator? The ‘suicidehypothesis’ revisited. Trends Pharmacol Sci 23:122–129.
Corona G, Deiana M, Incani A, Vauzour D, Dessi MA, and Spencer JP (2007)Inhibition of p38/CREB phosphorylation and COX-2 expression by olive oil poly-phenols underlies their anti-proliferative effects. Biochem Biophys Res Commun362:606–611.
Covas MI (2008) Bioactive effects of olive oil phenolic compounds in humans: reduc-tion of heart disease factors and oxidative damage. Inflammopharmacology 16:216–218.
Cuzzocrea S, Mazzon E, Dugo L, Genovese T, Di Paola R, Ruggeri Z, Vegeto E, CaputiAP, Van De Loo FA, Puzzolo D, et al. (2003a) Inducible nitric oxide synthasemediates bone loss in ovariectomized mice. Endocrinology 144:1098–1107.
Cuzzocrea S, Mazzon E, Dugo L, Patel NS, Serraino I, Di Paola R, Genovese T, BrittiD, De Maio M, Caputi AP, et al. (2003b) Reduction in the evolution of murine typeII collagen-induced arthritis by treatment with rosiglitazone, a ligand of theperoxisome proliferator-activated receptor �. Arthritis Rheum 48:3544–3556.
Cuzzocrea S, Riley DP, Caputi AP, and Salvemini D (2001) Antioxidant therapy: anew pharmacological approach in shock, inflammation, and ischemia/reperfusioninjury. Pharmacol Rev 53:135–159.
Deiana M, Aruoma OI, Bianchi ML, Spencer JP, Kaur H, Halliwell B, Aeschbach R,Banni S, Dessi MA, and Corongiu FP (1999) Inhibition of peroxynitrite dependentDNA base modification and tyrosine nitration by the extra virgin olive oil-derivedantioxidant hydroxytyrosol. Free Radic Biol Med 26:762–769.
Dong J, Gao Y, Liu Y, Shi J, Feng J, Li Z, Pan H, Xue Y, Liu C, Shen B, et al. (2010)The protective antibodies induced by a novel epitope of human TNF-alpha couldsuppress the development of collagen-induced arthritis. PLoS One 5:e8920.
Ferraccioli G, Bracci-Laudiero L, Alivernini S, Gremese E, Tolusso B, and DeBenedetti F (2010) Interleukin-1� and interleukin-6 in arthritis animal models:roles in the early phase of transition from acute to chronic inflammation andrelevance for human rheumatoid arthritis. Mol Med 16:552–557.
Filippin LI, Vercelino R, Marroni NP, and Xavier RM (2008) Redox signalling andthe inflammatory response in rheumatoid arthritis. Clin Exp Immunol 152:415–422.
Genovese T, Mazzon E, Muia C, Patel NS, Threadgill MD, Bramanti P, De Sarro A,Thiemermann C, and Cuzzocrea S (2005) Inhibitors of poly(ADP-ribose) polymer-ase modulate signal transduction pathways and secondary damage in experimen-tal spinal cord trauma. J Pharmacol Exp Ther 312:449–457.
Harris ED Jr (1990) Rheumatoid arthritis. Pathophysiology and implications fortherapy. N Engl J Med 322:1277–1289.
Hitchon CA and El-Gabalawy HS (2004) Oxidation in rheumatoid arthritis. ArthritisRes Ther 6:265–278.
Holmdahl R, Andersson M, Goldschmidt TJ, Gustafsson K, Jansson L, and Mo JA(1990) Type II collagen autoimmunity in animals and provocations leading toarthritis. Immunol Rev 118:193–232.
Holmdahl R, Mo J, Nordling C, Larsson P, Jansson L, Goldschmidt T, Andersson M,and Klareskog L (1989) Collagen induced arthritis: an experimental model for
rheumatoid arthritis with involvement of both DTH and immune complex medi-ated mechanisms. Clin Exp Rheumatol 7(Suppl 3):S51–S55.
Hom JT, Cole H, Estridge T, and Gliszczynski VL (1992) Interleukin-1 enhances thedevelopment of type II collagen-induced arthritis only in susceptible and not inresistant mice. Clin Immunol Immunopathol 62:56–65.
Impellizzeri D, Esposito E, Mazzon E, Paterniti I, Di Paola R, Bramanti P, MorittuVM, Procopio A, Britti D, and Cuzzocrea S (2011) The effects of oleuropein agly-cone, an olive oil compound, in a mouse model of carrageenan-induced pleurisy.Clin Nutr 30:533–540.
Institute of Laboratory Animal Resources (1996) Guide for the Care and Use ofLaboratory Animals, 7th ed, Institute of Laboratory Animal Resources, Commis-sion on Life Sciences, National Research Council, Washington DC.
Kasama T, Strieter RM, Lukacs NW, Burdick MD, and Kunkel SL (1994) Regulationof neutrophil-derived chemokine expression by IL-10. J Immunol 152:3559–3569.
Koch AE, Kunkel SL, Harlow LA, Mazarakis DD, Haines GK, Burdick MD, Pope RM,and Strieter RM (1994) Macrophage inflammatory protein-1�. A novel chemotacticcytokine for macrophages in rheumatoid arthritis. J Clin Invest 93:921–928.
Kohyama N, Nagata T, Fujimoto S, and Sekiya K (1997) Inhibition of arachidonatelipoxygenase activities by 2-(3,4-dihydroxyphenyl)ethanol, a phenolic compoundfrom olives. Biosci Biotechnol Biochem 61:347–350.
Kremastinos DT (2008) Olive and oleuropein. Hellenic J Cardiol 49:295–296.Menendez JA, Vazquez-Martin A, Colomer R, Brunet J, Carrasco-Pancorbo A, Gar-
cia-Villalba R, Fernandez-Gutierrez A, and Segura-Carretero A (2007) Olive oil’sbitter principle reverses acquired autoresistance to trastuzumab (Herceptin) inHER2-overexpressing breast cancer cells. BMC Cancer 7:80.
Miles EA, Zoubouli P, and Calder PC (2005) Differential anti-inflammatory effects ofphenolic compounds from extra virgin olive oil identified in human whole bloodcultures. Nutrition 21:389–394.
Mullane KM, Kraemer R, and Smith B (1985) Myeloperoxidase activity as a quan-titative assessment of neutrophil infiltration into ischemic myocardium. J Phar-macol Methods 14:157–167.
Perez-Martinez P, Lopez-Miranda J, Blanco-Colio L, Bellido C, Jimenez Y, MorenoJA, Delgado-Lista J, Egido J, and Perez-Jimenez F (2007) The chronic intake of aMediterranean diet enriched in virgin olive oil, decreases nuclear transcriptionfactor B activation in peripheral blood mononuclear cells from healthy men.Atherosclerosis 194:e141–146.
Petroni A, Blasevich M, Salami M, Papini N, Montedoro GF, and Galli C (1995)Inhibition of platelet aggregation and eicosanoid production by phenolic compo-nents of olive oil. Thromb Res 78:151–160.
Procopio A, Alcaro S, Nardi M, Oliverio M, Ortuso F, Sacchetta P, Pieragostino D,and Sindona G (2009) Synthesis, biological evaluation, and molecular modeling ofoleuropein and its semisynthetic derivatives as cyclooxygenase inhibitors. J AgricFood Chem 57:11161–11167.
Saija A, Tomaino A, Lo Cascio R, Rapisarda P, and Dederen JC (1998) In vitroantioxidant activity and in vivo photoprotective effect of a red orange extract. IntJ Cosmet Sci 20:331–342.
Salvini S, Sera F, Caruso D, Giovannelli L, Visioli F, Saieva C, Masala G, Ceroti M,Giovacchini V, Pitozzi V, et al. (2006) Daily consumption of a high-phenol extra-virgin olive oil reduces oxidative DNA damage in postmenopausal women. Br JNutr 95:742–751.
Soneja A, Drews M, and Malinski T (2005) Role of nitric oxide, nitroxidative andoxidative stress in wound healing. Pharmacol Rep 57:108–119.
Stavric B (1994) Role of chemopreventers in human diet. Clin Biochem 27:319–332.Szabo C, Virag L, Cuzzocrea S, Scott GS, Hake P, O’Connor MP, Zingarelli B,
Salzman A, and Kun E (1998) Protection against peroxynitrite-induced fibroblastinjury and arthritis development by inhibition of poly(ADP-ribose) synthase. ProcNatl Acad Sci USA 95:3867–3872.
Taub DD, Conlon K, Lloyd AR, Oppenheim JJ, and Kelvin DJ (1993) Preferentialmigration of activated CD4 and CD8 T cells in response to MIP-1� and MIP-1�.Science 260:355–358.
Trichopoulou A, Costacou T, Bamia C, and Trichopoulos D (2003) Adherence to aMediterranean diet and survival in a Greek population. N Engl J Med 348:2599–2608.
Visioli F, Bellomo G, and Galli C (1998) Free radical-scavenging properties of olive oilpolyphenols. Biochem Biophys Res Commun 247:60–64.
Vissers MN, Zock PL, and Katan MB (2004) Bioavailability and antioxidant effectsof olive oil phenols in humans: a review. Eur J Clin Nutr 58:955–965.
Walter WM Jr, Fleming HP, and Etchells JL (1973) Preparation of antimicrobialcompounds by hydrolysis of oleuropein from green olives. Appl Microbiol 26:773–776.
Address correspondence to: Prof. Salvatore Cuzzocrea, Department of Clin-ical and Experimental Medicine and Pharmacology, School of Medicine, Uni-versity of Messina, Torre Biologica, Policlinico Universitario, Via C. Valeria,Gazzi, 98100 Messina, Italy. E-mail: [email protected]
Oleuropein Aglycone and Arthritis 869
at ASPE
T Journals on July 8, 2018
jpet.aspetjournals.orgD
ownloaded from