oligomycin-dependent ionophoric protein subunit … · · 2005-04-22chloroform/methanol-soluble...
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 74, No. 10, pp. 4306-4310, October 1977Biochemistry
Oligomycin-dependent ionophoric protein subunit of mitochondrialadenosinetriphosphatase
(mitochondria/adenosine triphosphate/proteolipid/ionophore/ion pump)
RICHARD S. CRIDDLE, LESTER PACKER, AND PAUL SHIEHDepartment of Biochemistry and Biophysics, University of California, Davis, Davis, California 95616; and Membrane Bioenergetics Group, Lawrence BerkeleyLaboratory and the Department of Physiology-Anatomy, University of California, Berkeley, Berkeley, California 94720
Communicated by David E. Green, July 28,1977
ABSTRACT A proteolipid isolated from yeast mitochondrialadenosinetriphospitatase (subunit 9) (ATP phosphohydrolase;EC 3.6.1.3) by chloroform/methanol extraction has been shownto discharge photo-induced potentials across a planar phos-pholipid membrane containing bacteriorhodopsin. Oligomycin,a specific inhibitor of oxidative phosphorylation which bindsto tis protein, allows the potential gradient to be reestablished.When proteolipid was isolated from an oligomycin-resistantstrain, ionophoric activity was still obtained but the effect wasnot reversed by oligom cin. These studies suggest that the hy-drophobic subunit-9 polypeptide is the ionopboric componentlinking ATP synthesis (hydrolysis) with proton translocation.
Recent studies of mitochondrial oxidative phosphorylation havedemonstrated a link between ATP hydrolysis or synthesis andthe transmembrane movement of H+ (for review see ref. 1).The terminal step of chemical synthesis of ATP or of ATP hy-drolysis is generally agreed to be associated with the ATPaseprotein complex (ATP phosphohydrolase; EC 3.6.1.3) of theinner membrane of mitochondria. Although proton translo-cation is thought to proceed through the membrane sector ofthe complex, the components involved in the proton translo-cating mechanism remain unidentified. The concept that en-dogenous protein ionophores or protein-bound ionophores inthe mitochondrial inner membrane may play an important rolein energized membrane processes has recently been advancedby Green (2) and has been examined experimentally in severallaboratories (3-10).Some support for this concept is obtained from studies of
oligomycin inhibition of ATPase activities. The soluble F1-ATPase is not inhibited by this antibiotic; yet when this enzymeis isolated with a bound membrane sector, it is oligomycinsensitive (11). The site of oligomycin binding is clearly associ-ated with the membrane sector of ATPase complex. Protonpermeability of mitochondrial vesicles is low, but upon removalof FI-ATPase, permeability is greatly increased (12). Additionof oligomycin effectively reverses this increase. One interpre-tation of such results is that removal of F1 allows protons accessto some ionophoric component in the membrane sector ofATPase and that this ionophoric activity is blocked by bindingoligomycin.
Using an affinity labeling technique, we have recently shownthat the small chloroform/methanol-soluble subunit, designatedsubunit 9 by. the nomenclature of Tzagoloff and Meagher (13),is probably the site of oligomycin binding. This conclusion isstrengthened by observations that altered subunit 9 proteins arefound in the ATPase complex isolated from one class of oli-gomycin-resistant mitochondrial gene mutants of yeast (14, 15).Since oligomycin blocks proton transport and appears to do soby binding to subunit 9, we have investigated the possibility thatThe costs of publication of this article were defrayed in part by thepaynrient of page charges. This article must therefore be hereby marked"advertisement" in accordence with 18 U. S. C. §1734 solely to indicatethis fact.
subunit 9 may actually be the proton transporter. The assay wasmade in phospholipid-impregnated Millipore filter membranescontaining bacteriorhodopsin (16). Upon illumination, suchmembranes develop a photopotential between two electrodechambers separated by the membrane. Phospholipid vesiclescontaining subunit 9 were shown to discharge the potentialgradient and correspondingly to increase membrane conduc-tance upon association with the planar membrane. This iono-phoric activity of subunit 9- was greatly altered by oligomy-cin.
MATERIALS AND METHODS
PreparationsSaccharomyces cerevtsiae, strain D243-4A (a,ade,lys) and amitochondrial gene mutant, OR4, generated from this strainwere used in all the studies described (17). Cells were grownin 10-liter batch cultures in a New Brunswick microferm toearly stationary phase in medium consisting of (wt/vol) 1%Difco Peptone, 1% Difco Yeast Extract, and either 2% glucoseor 2% ethanol. No differences in isolated protein were notedwith the different carbon sources. Cells were harvested, washed,and broken with a Braun glass bead homogenizer; mitochondriawere then prepared by differential sedimentation (18). Sub-mitochondrial preparations were obtained by the method ofTzagoloff (19). Protein concentrations were determined by theLowry method (20).
Chloroform/methanol-soluble protein was isolated fromsubmitochondrial preparations by the method of Sierra andTzagaloff (21). The final steps in this procedure involve chro-matography on preparative silica gel plates (Merck, PrecoatedSilica gel F), location of components by exposure to iodinevapor, then scraping the sample from the plate and elution ofprotein from the silica gel. Alternatively, this same proteinfraction was prepared from oligomycin-sensitive ATPasepreparations solubilized from submitochondrial particles withTriton X-100 (13) and purified by chromatography on Sepha-rose 6B (22). Purified enzyme was precipitated with threevolumes of ether, and the chloroform/methanol-soluble proteinwas then isolated as described above. Thin-layer chromatog-raphy of the proteolipid fraction from both procedures indi-cated a single protein component or highly similar mixture ofproteins. This is consistent with the conclusion of Kuzela et al.(23) that only a single protein is found in this fraction from ratliver. A similar preparation from Neurospora mitochondriayields a single polypeptide for which the sequence has beenlargely determined (24). Oligomycin was obtained from Cal-biochem Corp. Venturicidin was a gift from D. E. Griffiths.Assays
Preparation of Lipid-Impregnated Filter Membrane.Millipore filters were purchased from Millipore Corp., Bedford,
4306
Proc. Natl. Acad. Sci. USA 74 (1977) 4307
Bacteriorhodopsin liposome (31 5Bjg protein/ml)plus 100 mM CaCI2 were added to innercompartment 15 mM Tris-buffer, 100 mMsucrose, pH 7.3)
Inner compartment was rinwith fresh Tris-buffer, sucr
nsedrose
Subunit-9 liposome (91Ag protein/mi) Oligomycin (2 5yug/ml) was
plus 100 mM CaC12 were added added to outer compartment.
to outer compartment. Oligomycin(2.5ug/ml) was
added to inner compartmentOuter compartment was rinsedwith fresh Tris-buffer, sucroseI I~~~~~~~~~~~~
2XI(oTmhoxcm2
ix IO8mho xC 2
0300 30 60 90
Dark incubation time, min
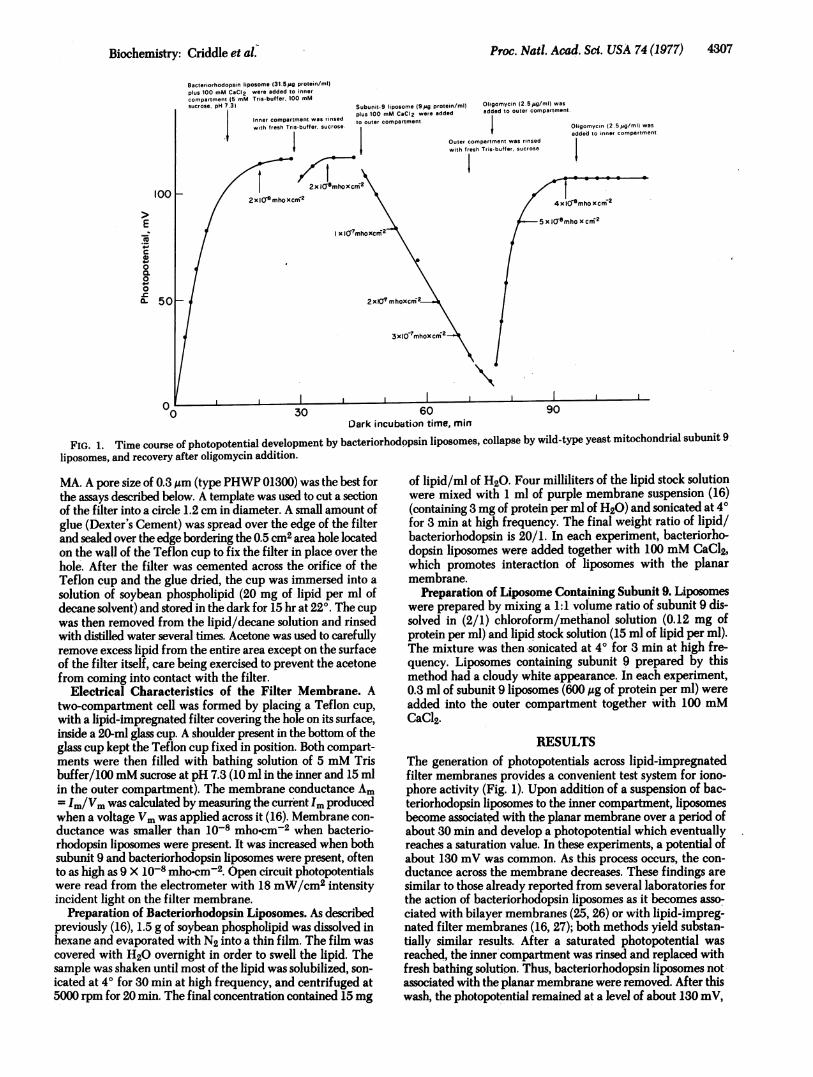
FIG. 1. Time course of photopotential development by bacteriorhodopsin liposomes, collapse by wild-type yeast mitochondrial subunit 9liposomes, and recovery after oligomycin addition.
MA. A pore size of 0.3Mm (type PHWP 01300) was the best forthe assays described below. A template was used to cut a sectionof the filter into a circle 1.2 cm in diameter. A small amount ofglue (Dexter's Cement) was spread over the edge of the filterand sealed over the edge bordering the 0.5 cm2 area hole locatedon the wall of the Teflon cup to fix the filter in place over thehole. After the filter was cemented across the orifice of theTeflon cup and the glue dried, the cup was immersed into a
solution of soybean phospholipid (20 mg of lipid per ml ofdecane solvent) and stored in the dark for 15 hr at 22°. The cupwas then removed from the lipid/decane solution and rinsedwith distilled water several times. Acetone was used to carefullyremove excess lipid from the entire area except on the surfaceof the filter itself, care being exercised to prevent the acetonefrom coming into contact with the filter.
Electrical Characteristics of the Filter Membrane. Atwo-compartment cell was formed by placing a Teflon cup,with a lipid-impregnated filter covering the hole on its surface,inside a 20-ml glass cup. A shoulder present in the bottom of theglass cup kept the Teflon cup fixed in position. Both compart-ments were then filled with bathing solution of 5 mM Trisbuffer/100 mM sucrose at pH 7.3 (10 ml in the inner and 15 mlin the outer compartment). The membrane conductance Am= Im/Vm was calculated by measuring the current Im producedwhen a voltage Vm was applied across it (16). Membrane con-
ductance was smaller than 10-8 mho cm2 when bacterio-rhodopsin liposomes were present. It was increased when bothsubunit 9 and bacteriorhodopsin liposomes were present, oftento as high as 9 X 10-8 mho-cm-2. Open circuit photopotentialswere read from the electrometer with 18 mW/cm2 intensityincident light on the filter membrane.
Preparation of Bacteriorhodopsin Liposomes. As describedpreviously (16), 1.5 g of soybean phospholipid was dissolved inhexane and evaporated with N2 into a thin film. The film wascovered with H20 overnight in order to swell the lipid. Thesample was shaken until most of the lipid was solubilized, son-
icated at 40 for 30 min at high frequency, and centrifuged at5000 rpm for 20 min. The final concentration contained 15 mg
of lipid/ml of H20. Four milliliters of the lipid stock solutionwere mixed with 1 ml of purple membrane suspension (16)(containing 3mg of protein per ml of H20) and sonicated at 40for 3 min at high frequency. The final weight ratio of lipid/bacteriorhodopsin is 20/1. In each experiment, bacteriorho-dopsin liposomes were added together with 100 mM CaCl2,which promotes interaction of liposomes with the planarmembrane.
Preparation of Liposome Containing Subunit 9. Liposomeswere prepared by mixing a 1:1 volume ratio of subunit 9 dis-solved in (2/1) chloroform/methanol solution (0.12 mg ofprotein per ml) and lipid stock solution (15 ml of lipid per ml).The mixture was then -sonicated at 40 for 3 min at high fre-quency. Liposomes containing subunit 9 prepared by thismethod had a cloudy white appearance. In each experiment,0.3 ml of subunit 9 liposomes (600 Mg of protein per ml) wereadded into the outer compartment together with 100 mMCaCl2.
RESULTSThe generation of photopotentials across lipid-impregnatedfilter membranes provides a convenient test system for iono-phore activity (Fig. 1). Upon addition of a suspension of bac-teriorhodopsin liposomes to the inner compartment, liposomesbecome associated with the planar membrane over a period ofabout 30 min and develop a photopotential which eventuallyreaches a saturation value. In these experiments, a potential ofabout 130 mV was common. As this process occurs, the con-
ductance across the membrane decreases. These findings are
similar to those already reported from several laboratories forthe action of bacteriorhodopsin liposomes as it becomes asso-
ciated with bilayer membranes (25, 26) or with lipid-impreg-nated filter membranes (16, 27); both methods yield substan'-tially similar results. After a saturated photopotential was
reached, the inner compartment was rinsed and replaced withfresh bathing solution. Thus, bacteriorhodopsin liposomes notassociated with the planar membrane were removed. After thiswash, the photopotential remained at a level of about 130 mV,
100
E
.XaC0
0
0-c0.CL
Biochemistry: Criddle et al.
i
4308 Biochemistry: Criddle et al.
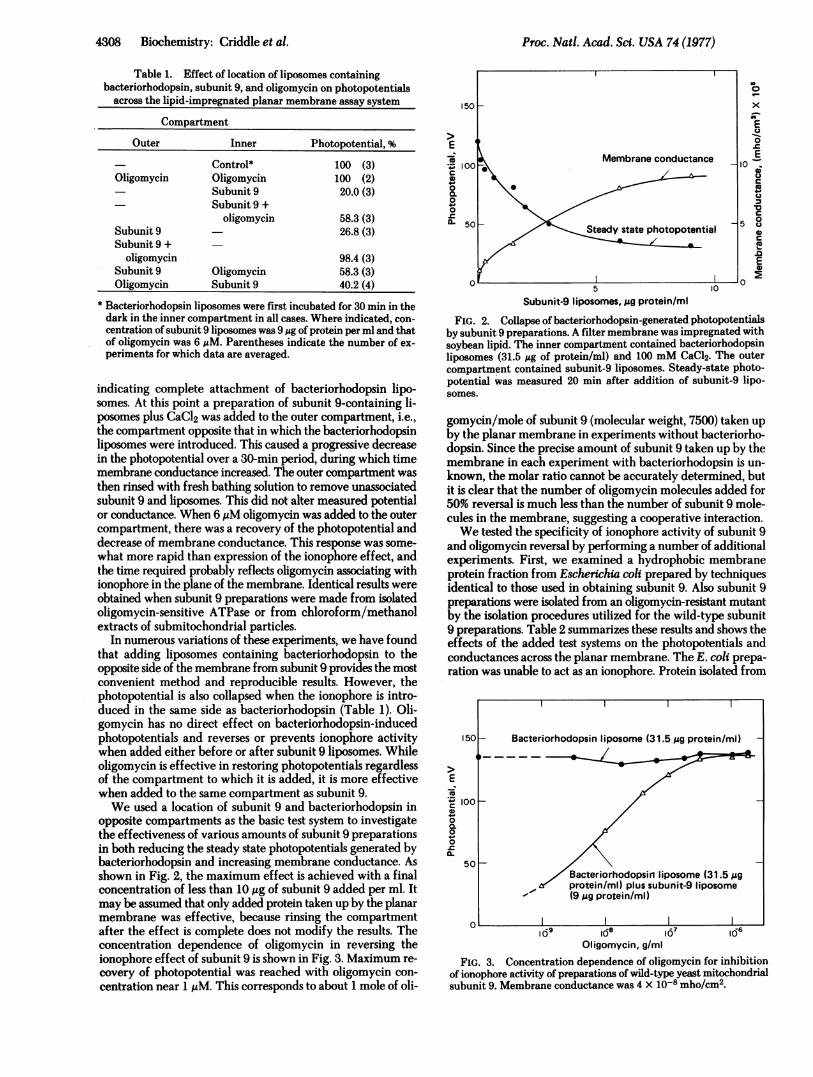
Table 1. Effect of location of liposomes containingbacteriorhodopsin, subunit 9, and oligomycin on photopotentials
across the lipid-impregnated planar membrane assay system
Compartment
Outer Inner Photopotential, %
Control* 100 (3)Oligomycin Oligomycin 100 (2)
Subunit 9 20.0 (3)Subunit 9 +
oligomycin 58.3 (3)Subunit 9 26.8 (3)Subunit 9 +
oligomycin 98.4 (3)Subunit 9 Oligomycin 58.3 (3)Oligomycin Subunit 9 40.2 (4)
* Bacteriorhodopsin liposomes were first incubated for 30 min in thedark in the inner compartment in all cases. Where indicated, con-centration of subunit 9 liposomes was 9 ug of protein per ml and thatof oligomycin was 6 MM. Parentheses indicate the number of ex-periments for which data are averaged.
indicating complete attachment of bacteriorhodopsin lipo-somes. At this point a preparation of subunit 9-containing li-posomes plus CaCl2 was added to the outer compartment, i.e.,the compartment opposite that in which the bacteriorhodopsinliposomes were introduced. This caused a progressive decreasein the photopotential over a 3S-min period, during which timemembrane conductance increased. The outer compartment wasthen rinsed with fresh bathing solution to remove unassociatedsubunit 9 and liposomes. This did not alter measured potentialor conductance. When 6 ,uM oligomycin was added to the outercompartment, there was a recovery of the photopotential anddecrease of membrane conductance. This response was some-what more rapid than expression of the ionophore effect, andthe time required probably reflects oligomycin associating withionophore in the plane of the membrane. Identical results wereobtained when subunit 9 preparations were made from isolatedoligomycin-sensitive ATPase or from chloroform/methanolextracts of submitochondrial particles.
In numerous variations of these experiments, we have foundthat adding liposomes containing bacteriorhodopsin to theopposite side of the membrane from subunit 9 provides the mostconvenient method and reproducible results. However, thephotopotential is also collapsed when the ionophore is intro-duced in the same side as bacteriorhodopsin (Table 1). Oli-gomycin has no direct effect on bacteriorhodopsin-inducedphotopotentials and reverses or prevents ionophore activitywhen added either before or after subunit 9 liposomes. Whileoligomycin is effective in restoring photopotentials regardlessof the compartment to which it is added, it is more effectivewhen added to the same compartment as subunit 9.We used a location of subunit 9 and bacteriorhodopsin in
opposite compartments as the basic test system to investigatethe effectiveness of various amounts of subunit 9 preparationsin both reducing the steady state photopotentials generated bybacteriorhodopsin and increasing membrane conductance. Asshown in Fig. 2, the maximum effect is achieved with a finalconcentration of less than 10 ,g of subunit 9 added per ml. Itmay be assumed that only added protein taken up by the planarmembrane was effective, because rinsing the compartmentafter the effect is complete does not modify the results. Theconcentration dependence of oligomycin in reversing theionophore effect of subunit 9 is shown in Fig. 3. Maximum re-covery of photopotential was reached with oligomycin con-centration near 1 ,M. This corresponds to about 1 mole of oli-
E
IZco
Q0.000
0
x
E0
0E0)
C0w
40
u'U80
ES)
Subunit-9 liposomes,,g protein/ml
FIG. 2. Collapse ofbacteriorhodopsin-generated photopotentialsby subunit 9 preparations. A filter membrane was impregnated withsoybean lipid. The inner compartment contained bacteriorhodopsinliposomes (31.5 gg of protein/ml) and 100 mM CaCl2. The outercompartment contained subunit-9 liposomes. Steady-state photo-potential was measured 20 min after addition of subunit-9 lipo-somes.
gomycin/mole of subunit 9 (molecular weight, 7500) taken upby the planar membrane in experiments without bacteriorho-dopsin. Since the precise amount of subunit 9 taken up by themembrane in each experiment with bacteriorhodopsin is un-known, the molar ratio cannot be accurately determined, butit is clear that the number of oligomycin molecules added for50% reversal is much less than the number of subunit 9 mole-cules in the membrane, suggesting a cooperative interaction.We tested the specificity of ionophore activity of subunit 9
and oligomycin reversal by performing a number of additionalexperiments. First, we examined a hydrophobic membraneprotein fraction from Escherichia coli prepared by techniquesidentical to those used in obtaining subunit 9. Also subunit 9preparations were isolated from an oligomycin-resistant mutantby the isolation procedures utilized for the wild-type subunit9 preparations. Table 2 summarizes these results and shows theeffects of the added test systems on the photopotentials andconductances across the planar membrane. The E. coli prepa-ration was unable to act as an ionophore. Protein isolated from
150 -
E
C0,
00.
0-C0o
100I
50-
%
109 10 8 17 106Oligomycin, g/ml
FIG. 3. Concentration dependence of oligomycin for inhibitionof ionophore activity of preparations of wild-type yeast mitochondrialsubunit 9. Membrane conductance was 4 X 10-8 mho/cm2.
Bacteriorhodopsin liposome (31.5,g protein/ml)
Bacteriorhodopsiri liposome (31.5 Mgprotein/ml) plus subunit-9 liposome(9,ug protein/ml)
Proc. Natl. Acad. Sci. USA 74 (1977)

Proc. Nati. Acad. Sci. USA 74 (1977) 4309
Table 2. Effects of subunit 9 preparation on wild-type andoligomycin-resistant mutants and an E. coli hydrophobic protein
fraction on membrane potentials and conductances
Steady-statephotopotential, mV Membrane
Proteoliposomes* Averaget % conductance$
Control 108(100-110) 100 4 0.03Wild-type subunit 9 21(17-25) 19.8 39 f 1Wild-type subunit 9+ oligomycin 63 (55-75) 59.4 7.6 0.9
(OR4) mutant subunit 9 52(48-58) 49 10 + 1(OR4) mutant subunit 9+ oligomycin 29(25-33) 27.3 13 I 1
E. coli fraction 106(100-110) 100 4 i 0.06E. coli fraction +
oligomycin 106(100-108) 100 4 1 0.03
* 4.5 ,ug of protein per ml added.t Numbers in parentheses give the range; n = 3.t (mho X cm-2) X 108. Values are mean + SEM; n = 3.
the oligomycin-resistant mutant OR4 was nearly as good anionophore as preparations from wild-type; however, in this case,6 uM oligomycin was unable to reverse the ionophore action.To evaluate whether the presence of bacteriorhodopsin li-
posomes or contaminants in these liposomes might affect thenature of the ionophore activity and the action of oligomycin,we ran control experiments in which a pH gradient was createdacross the membrane in the absence of bacteriorhodopsin. Theaddition of subunit 9 still caused an increase in conductance anda reduction of the potential across the planar membrane. Thiseffect was also reversed by addition of oligomycin. It is note-worthy that venturicidin (1 MM) was somewhat more effectivein reversing the ionophore effects.
Yet another control was to test protein fractions that wereinitially soluble in chloroform/methanol but became insolubleafter ether precipitation during the subunit 9 preparation (seeMaterials and Methods). Such preparations were found tomanifest a somewhat reduced ionophore activity compared topurified subunit 9 preparations, but these fractions failed torespond to oligomycin reversal.
DISCUSSIONWorkers in several laboratories in recent years have attemptedto identify protein fractions in mitochondria that exhibit protonionophore activity (3-10). Racker has shown that mitochondrialand chloroplast proteolipids may act as ionophores in collapsingproton gradients generated by bacteriorhodopsin (4), but it wasnot shown that these proteolipids were in fact associated withATPase nor was any specific relation between ionophoric ac-tivity and biological function of this protein demonstrated. Thislatter question is important since observations of general pro-tein-induced conductivity changes in black lipid membranesand liposomes have been reported recently (28). Therefore, thesimple observation that proteolipids increase conductivity oflipid membranes is not in itself highly significant.
In this investigation we have established that a proteolipidfraction isolated from the mitochondrial inner membrane actsas an ionophore in an oligomycin-reversible fashion whichcoincides with the observed effect of oligomycin in vvo on ATPsynthesis or hydrolysis. Fig. 4 diagrammatically illustratesphotopotentials generated by bacteriorhodopsin, their collapsein the presence of subunit 9 liposome preparations, and thenearly complete restoration of the photopotentials broughtabout by oligomycin. Oligomycin does not exert a measurableeffect on phospholipid membrane conductivity nor does it seem
I50m1hk
60 secI
FIG. 4. Summary scheme.
to have an effect on conductivity changes induced by mem-brane proteins in general. This was shown by our studies withE. coli membrane protein and with added nonmembraneprotein. However, the best demonstration of specificity of oli-gomycin action is shown by comparison of subunit 9 prepara-tions of wild-type and mutant (OR4) proteins. Oligomycinreversed ionophore activity only with wild-type protein. Thus,the oligomycin effect on membrane conductivity resembles thein vivo effects of this inhibitor of oxidative phosphorylation.
In order for oligomycin to block proton conductivity viasubunit 9, it must bind to the protein and either interfere with,the ability to accept protons or alter the ability of protein totransport protons once bound. Oligomycin may work by"locking" subunit 9 into a nontransporting form. In this regardit is interesting to note that Schubert et al. demonstrated thataggregated membrane proteins had a markedly lowered effecton the conductivity of black lipid membranes (28). Consider-ation of Fig. 3 indicates that oligomycin may affect more thanone protein molecule and suggests an oligomycin-inducedaggregation. Previous studies have shown that subunit 9 mayexist in a monomer or aggregated form (29), and that the mo-nomer form predominates in the oligomycin-resistant mutant(14, 15).Our preparations of subunit 9 behave as a homogeneous
preparation of one ionophoric component. If there is more thana single ionophore in the preparation, all must be sensitive toinhibition by oligomycin since collapsed photopotentials arecompletely restored. Furthermore, all must lose this sensitivity
Biochemistry: Criddle et al.

4310 Biochemistry: Criddle et al.
by a mutation that alters the properties of ATPase subunit 9.The possibility that a small, nonprotein ionophore copurifieswith subunit 9 through chloroform/methanol extraction, etherprecipitation, and thin-layer chromatography also seems un-likely, particularly since a mitochondnal gene mutation alteringsubunit 9 changes sensitivity of the ionophore to oligomycin.This does not rule out a covalently bound ionophoric componenton the protein.
Fillingame (6, 7) has found thatE. coli membrane fractionscontain a dicyclohexylcarbodiimide-binding protein; however,in our studies a chloroform/methanol-soluble protein extractedfrom anE. coli membrane fraction by the same method usedfor mitochondria did not serve as an ionophore. However, Gitleret al. (30) have reported that proteolipid fractions isolated fromchloroplasts with butanol manifest sensitivity to dicyclohexyl-carbodiimide. Folch-Pi and Stoffyn (31) noted large solvent-dependent differences in the conformation of extracted pro-teolipids. Further study of the characteristics of these varioushydrophobic protein fractions should help to elucidate theirspecificity and molecular basis of ionophore activity.
We thank E. Rembold, T. Edwards, and S. Scharf for preparationof protein fractions and the Energy Research and Development Ad-ministration and U.S. Public Health Service for Grant GM1007 forresearch support.
1. Quagliariello, E., Papa, S. & Rosi, C. S. (1971) Energy Trans-duction in Respiration and Photosynthesis, Bari: AdriaticaEditrice.
2. Green, D. E. (1974) "The electromechanochemical model forenergy coupling in mitochondria," Biochim. Biophys. Acta 346,27-78.
3. Racker, E. & Eytan, E. (1975) "A coupling factor from sarco-plasmic reticulum required for the translocation of Ca2+ ions ina reconstituted Ca2+ ATPase pump," J. Biol. Chem. 250,7533-7534.
4. Racker, E. (1976) A New Look at Mechanisms in Bioenergetics(Academic Press, New York), pp. 128-135.
5. Kagawa, Y., Ohno, K., Yoshida, M., Takeuchi, Y. & Sone, N.(1977) "Proton translocation by ATPase and bacteriorhodopsin,"Fed. Proc. 36,1815-1818.
6. Fillingame, R. H. (1976) "Purification of the carbodiimide-re-active protein component of the ATP energy-transducing systemof E. colh," J. BMol. Chem. 251, 6630-6637.
7. Fillingame, R. H. (1975) "Identification of the dicyclohexyl-carbodiimide reactive protein component of the adenosine 5'-triphosphate energy transducing system in E. coi," j. Bacteriol.124,870-883.
8. Shamoo, A. E., Rayn, T. E., Stewart, P. S. & MacLennan, D. H.(1976) "Localization of ionophoric activity in a 20,000-daltonfragment of the adenosine triphosphatase of sarcoplasmic retic-ulum," J. Biol. Chem. 251, 4147-4154.
9. Shamoo, A. E. & MacLennan, D. H. (1975) "Separate effects ofmercurial compounds on the ionophoric and hydrolytic functionsof the Ca2+ + Mg2+-ATPase of sarcoplasmic reticulum," J.Membr. Biol. 25,65-74.
10. Shamoo, A. E. & MacLennan, D. H. (1974) "A Ca2+-dependentand -selective ionophore as part of the Ca2+ + Mg2+-dependentadenosine triphosphatase of sarcoplasmic reticulum," Proc. Nati.Acad. Sci. USA 71, 3522-3526.
11. Lardy, H. A., Johnson, D. & McMurray, W. C. (1958) "Antibioticsas tools for metabolic studies. I. A survey of toxic antiobiotics inrespiratory, phosphorylative and glycolytic systems," Arch.Biochem. Biophys. 78,587-597.
12. Mitchell, P. (1973) "Cation-translocating adenosine triphos-phatase models: How direct is the participation of adenosine
triphosphate and its hydrolysis products in cation translocation?,"FEBS Lett. 33,267-274.
13. Tzagoloff, A. & Meagher, P. (1971) "Assembly of the mito-chondrial membrane system. V. Properties of a dispersed prep-aration of the rutamyin-sensitive ATPase of yeast mitochondria,"J. Biol. Chem. 246,7328-7338.
14. GrootObbink, D. J., Hall, R. M., Linnane, A. W., Lukins, H. B.,Monk, B. C., Spithill, T. & Trembath, M. K. (1976) in The GeneticFunction of Mitochondrial DNA, eds. Kroon, A. M. & Saccone,C. (North-Holland Pub. Co., Amsterdam), pp. 163-173.
15. Tzagoloff, A., Akai, A. & Foury P. (1976) "Assembly of themitochondrial membrane system. XVI. Modified form of theATPase proteolipid in oligomycin-resistant mutants of Saccha-romyces cerevisiae," FEBS Lett. 65,391-395.
16. Shieh, P., Packer, L. & Lanyi, J. (1977) "Biomembranes Part D,"in Methods in Enzymology, eds. Fleischer, S. & Packer, L. (Ac-ademic Press, New York), in press.
17. Shannon, G., Enns, R., Wheelis, L., Burchiel, K. & Criddle, R.S. (1973) "Alterations in mitochondrial adenosine triphosphataseactivity resulting from mutation of mitochondrial DNA," J. Biol.Chem. 248,3004-3011.
18. Criddle, R. S. & Schatz, G. (1969) "Promitochondria of anaero-bically grown yeasts. I. Isolation and biochemical properties,"Biochemistry 8,322-334.
19. Tzagoloff, A. (1969) "Assembly of the mitochondrial membranesystem. I. Characterization of some enzymes of the inner mem-brane of yeast mitochondria," J. Biol. Chem. 244,5020-5033.
20. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, P. J.(1951) "Protein measurements with the Folin phenol reagent,"J. Biol. Chem. 193,265-275.
21. Sierra, M. F. & Tzagoloff, A. (1973) "Assembly of the mito-chondrial membrane system. Purification of a mitochondrialproduct of the ATPase," Proc. Nati. Acad. Sci. USA 70, 3155-3159.
22. Enns, R. & Criddle, R. S. (1977) "Affinity labeling of yeast mi-tochondrial ATPase by reduction with [3H]borohydride," Arch.Biochem. Biophys., 182,587-600.
23. Kuzela, S., Kolarov, J. & Krempasky, V. (1973) "Solubility of theintramitochondrially synthesized protein and other membraneproteins of rat liver mitochondria in acids or neutral chloro-form:methanol," Biochem. Biophys. Res. Commun. 54,9-16.
24. Machleidt, W., Michel, R., Neupert, W. & Wachter, E. (1976)in Genetics and Biogenesis of Chloroplasts and Mitochondria,eds. Bucher, Th., Neupert, W. & Werner, S. (North-Holland Pub.Co., Amsterdam), pp. 195-198.
25. Drachev, L. A., Jasaitis, A. A., Kaulen, A. D., Kondrashin, A. D.,Liberman, E. A., Nemecek, L. B., Ostroumov, S. A., Semenov,A., Yu, A. & Skulachev, V. P. (1974) "Direct measurement ofelectric current generation by cytochrome oxidase, H+-ATPaseand bacteriorhodopsin," Nature 249, 321.
26. Shieh, P. & Packer, L. (1976) "Photo-induced potentials acrossa polymer stabilized planar membrane in the presence of bac-teriorhodopsin," Biochem. Biophys. Res. Commun. 71, 603-609.
27. Skulachev, V. P. (1977), "Biomembranes Part D," in Methodsin Enzymology, eds. Fleischer, S. & Packer, L. (Academic Press,New York), in press.
28. Schubert, D., Blend, H., Domning, B. & Wiedner, G. (1977)"Proton-induced conductivity changes in black lipid membranesand protein aggregation," FEBS Lett. 74, 47-49.
29. Tzagoloff, A. & Akai, A. (1972) "Assembly of the mitochondrialmembrane system. VIII. Properties of the products of mito-chondrial protein synthesis," J. Biol. Chem. 247, 6517-6523.
30. Nelson, N., Eytan, E., Notsani, B., Sigrist, H., Sigrist-Nelson, K.& Gitler, C. (1977) "Isolation of a chloroplast N,N'-dicyclohex-ylcarbodiimide-binding proteolipid, active in proton transloca-tion," Proc. Natl. Acad. Sci. USA 74, 2375-2378.
31. Folch-Pi, J. & Stoffyn, J. (1972) "Proteolipids from membranesystems," Ann. N.Y. Acad. Sci. 195,86-107.
Proc. Natl. Acad. Sci. USA 74 (1977)