on the polyps of the common jellyfish aurelia … the polyps of aurelia aurita in kagoshima bay 453...
TRANSCRIPT

451
Journal of Oceanography, Vol. 58, pp. 451 to 459, 2002
Keywords:⋅ Polyp,⋅ Aurelia aurita,⋅ artificial substrata,⋅ colony,⋅ distributionpattern,
⋅ Iδ-Index,⋅ strobila,⋅ floating pier,⋅ undersurface.
* Corresponding author. E-mail: [email protected] Present address: Japan Marine Science and Technology Center, 2-15Natsushima-cho, Yokosuka 237-0061, Japan.
Copyright © The Oceanographic Society of Japan.
On the Polyps of the Common Jellyfish Aurelia aurita inKagoshima Bay
HIROSHI MIYAKE1*, MAKOTO TERAZAKI1 and YOSHIKO KAKINUMA2
1Ocean Research Institute, University of Tokyo, 1-15-1 Minamidai, Nakano-ku, Tokyo 164-8639, Japan2Faculty of Science, Kagoshima University, 1-21-35 Korimoto, Kagoshima 890-0065, Japan
(Received 22 November 2000; in revised form 6 October 2001; accepted 10 October 2001)
There is the natural habitat of polyps of the common jellyfish Aurelia aurita in theTaniyama area, Kagoshima Bay. We examined the attachment substrata, density,colony structure and strobilation of the polyps. The polyps were observed only on thehorizontal undersurface of floating piers. They attached specifically to Mytilus shells,solitary ascidians, calcareous polychaete tubes, muddy amphipod tubes and the gapspace that fouling animals peeled off the substrata. The polyp colonies were distrib-uted in patches. Spatial distribution patterns of the polyps within their colonies wereuniform. Strobilation occurred during late December to March, when water tempera-tures were 16–17°C, and a large number of ephyrae were released. An increase inman-made structures such as floating piers in coastal areas may lead to bloomings ofAurelia aurita medusae.
and the interaction between polyps and their environment.We recently discovered polyps of Aurelia aurita in
Kagoshima Bay (Miyake et al., 1997). The purpose ofthis paper is to report the distribution pattern and colonyforms of the polyps with special regard to intra- andinterspecific interactions and the occurrence of strobila-tion.
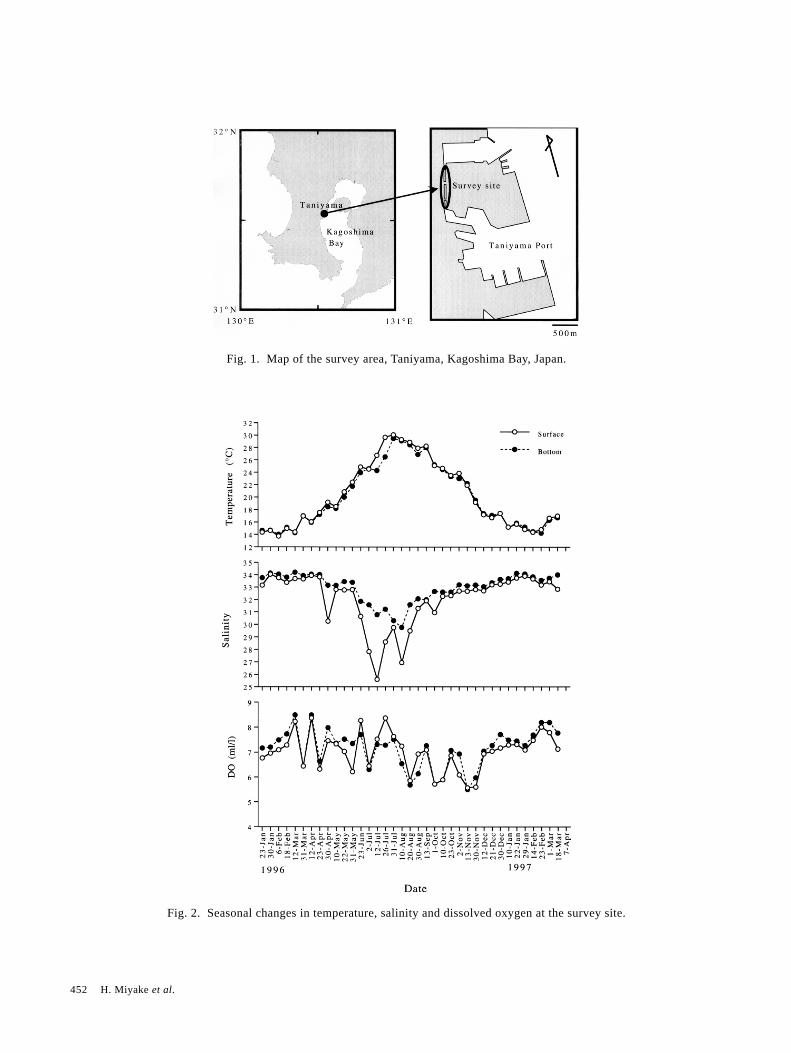
2. Materials and MethodsThe survey site was located in the Taniyama area,
Kagoshima Bay, southern end of Kyushu, Japan (Fig. 1).This site is in a man-made canal that is 1300 m long and30 m wide, 3 m deep at ebb tide and 6 m deep at hightide. The wall of the canal was a metal plate extending to4 m from the bottom and the upper part was concrete.Many vessels such as leisure boats and many floating piersare anchored in the canal.
Investigations were conducted by SCUBA at leastonce per month from April 1996 to March 1997 in orderto clarify the period of strobilation. Temperature, salinity(ALEC ELECTRONICS, MODEL ACT20-1), and dis-solved oxygen (YSI, Model 55/25 FT) were measured inthe habitat of the polyps.
We observed the substrata of polyps in situ and col-lected six to twelve colonies of polyps with the substrataduring every investigation. We examined polyp coloniesand their substrata under a stereoscopic microscope todetermine the micro-distribution of polyps, colony form,
1. IntroductionFrom matured Aurelia aurita medusae, planulae are
released and attach to some suitable substrata and developinto polyps. Each polyp reproduces asexually by budding,fission, or pedal disk (cyst) (Kakinuma, 1975) and formsa colony.
There are few reports about the polyps of Aureliaaurita in the field. Because of the lack of informationabout the habitat of polyps, it has been difficult to findpolyps in the field. Therefore, their natural history hasbeen inferred mainly from studies on polyps attached toartificial plates (Hernroth and Gröndahl, 1983, 1985a,1985b; Yasuda, 1988; Gröndahl, 1989) and those kept inaquaria (see Arai, 1997). Detailed ecological studies onpolyps in situ have been published from the MariagerFjord in Denmark (Ussing, 1927), the Kiel Fjord in Ger-many (Thiel, 1962), and the Gullmar Fjord in Sweden(Hernroth and Gröndahl, 1985a; Gröndahl, 1988a, b).These studies describe the occurrence of strobilae, pol-yps, and the physical environment in which they werefound, i.e., temperature and salinity regimes. However,little is known about the distribution pattern of polyps
452 H. Miyake et al.
Fig. 1. Map of the survey area, Taniyama, Kagoshima Bay, Japan.
Fig. 2. Seasonal changes in temperature, salinity and dissolved oxygen at the survey site.

On the Polyps of Aurelia aurita in Kagoshima Bay 453
and presence or absence of strobilae. Strobilation ratioswere based on the strobilae components of all polyps col-lected on the same day.
The density of polyps on the collected substrata wasalso measured. For the colonies on the almost flat sub-strata, spatial distribution patterns were determined bythe Iδ-Index (Morisita, 1959). Distribution patterns wereclassified into six patterns from the shape of the Iδ-quadratsize relationship: 1. Random distribution; 2. Uniform dis-tribution; 3. Aggregated distribution with small clumpsand random intra-clump distribution; 4. Aggregated dis-tribution with small clumps and uniform intra-clump dis-tribution; 5. Aggregated distribution with large clumpsand random intra-clump distribution; 6. Aggregated dis-tribution with large clumps and uniform intra-clump dis-tribution (Morisita, 1959). The size of peaks in theIδ(s)
/Iδ(2s) curve indicates the size of a clump (unit colony
size) or the size of aggregation of clumps (Morisita, 1959).
3. Results
3.1 Environmental parametersTemperature ranged from 14.2 to 30.1°C at the sur-
face and from 14.2 to 29.5°C at the bottom (3–6 m depth)(Fig. 2). Salinity varied from 25.6 to 34.0 at the surfaceand from 30.0 to 34.1 at the bottom (Fig. 2). Salinity atthe surface was low in summer, the lowest value being25.6, but salinity at the bottom was normally 30.8. The
Fig. 3. Colony of Aurelia aurita polyps (P) on Mytilus sp. There were many muddy amphipod tubes (AT) on the shell.
dissolved oxygen concentration was lower from Augustto January (Fig. 2). The maximum value of dissolvedoxygen was 8.37 mg/l in July and the minimum was 5.57mg/l in November.
3.2 SubstrataThere were many shells and remains of fouling or-
ganisms on the bottom and muddy sediment had accu-mulated. Polyps were found on the undersurface of float-ing piers and buoys. Floating piers were rectilinear, 15 mlong, 3 m wide and 1.5 m thick. These piers were madeof a polystyrene float with a concrete frame and deck.The lower one meter of the pier was submerged.
No polyps were present on the sides of the floatingpiers, the wall of the channel or on the bottom of ships.They were discovered only on the undersurface of thefloating piers and of the buoys (Fig. 3). The polyps at-tached to these substrata in patches. Most of them wereattached to solitary ascidians, barnacles, Mytilus,polychaete tubes and amphipod tubes (Table 1). They alsoattached densely to the gap spaces that fouling animalshad peeled off the substrata. In particular Mytilus, soli-tary ascidians and the gap spaces were the major sub-strata for polyp attachment. The Mytilus shells that hadattached polyps were larger than 10 mm long, and in par-ticular polyps formed large colonies on shells longer than50 mm. However, no polyps were observed on the shellsof dead Mytilus. Polyps were never found on colonial

454 H. Miyake et al.
ascidians nor soft solitary ascidians like Ciona, but onlyon hard ascidians like Styela. The polyps were few on theshells of living barnacles, but were numerous on deadbarnacles. Polyps also formed extremely large, densecolonies on freshly vacated polystyrene substratum, wherelarge fouling organisms had fallen off or were removed.In addition, a fallen leaf and the cellophane cover of acigarette package also provided substrata for polyps.
3.3 Distribution patternThe densest part in any colony reached 88 inds./cm2.
The average densities of polyps were 7.3–17.6 inds./cm2
on Mytilus and 3.1–18.5 inds./cm2 on polystyrene. Thesevariations in the densities were derived from conditionsof colonies.
Every colony had an aggregated distribution withsmall clumps and intra-clump distribution was uniformor random independently of the season (Fig. 4). Typicaldistribution patterns of polyps on major substrata (Mytilusand polystyrene substrata) are outlined below.
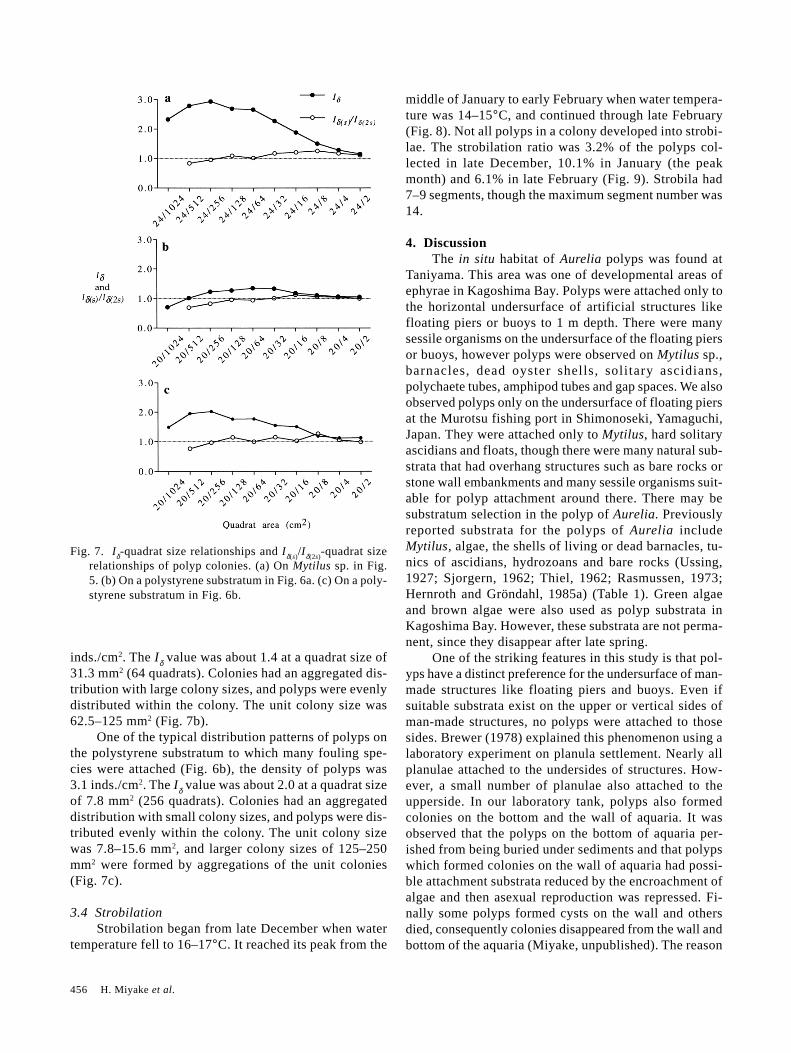
One of the typical distribution patterns of polyps onMytilus (Fig. 5), the density of polyps was 7.4 inds./cm2,and the Iδ-value was about 3 at a quadrat size of 9.4 mm2
(256 quadrats). According to the shape of the Iδ-quadrat-size graph, colonies were in an aggregated distributionpattern with small colonies, and polyps were evenly dis-
tributed within the colony. The unit colony size on Mytiluswas 9.4–18.8 mm2, and larger colonies (150–300 mm2)were formed by aggregations of the unit colonies (Fig.7a).
One of the typical distribution patterns of polyps onthe polystyrene substratum to which amphipod tubes wereattached (Fig. 6a), the density of polyps was 6.1
Table 1. Substratum for Aurelia aurita polyp colonies. : occurred commonly; �: occurred; ×: did not occur.
Fig. 4. Iδ-quadrat size relationships of polyp colonies on somesubstrata.

On the Polyps of Aurelia aurita in Kagoshima Bay 455
Fig. 5. Polyp distribution on both sides of a Mytilus shell attached to the undersurface of a floating pier.
Fig. 6. Polyp distribution on a polystyrene substratum on the undersurface of a floating pier. (a) A substratum where few speciesexisted. (b) A substratum where many other species existed.
456 H. Miyake et al.
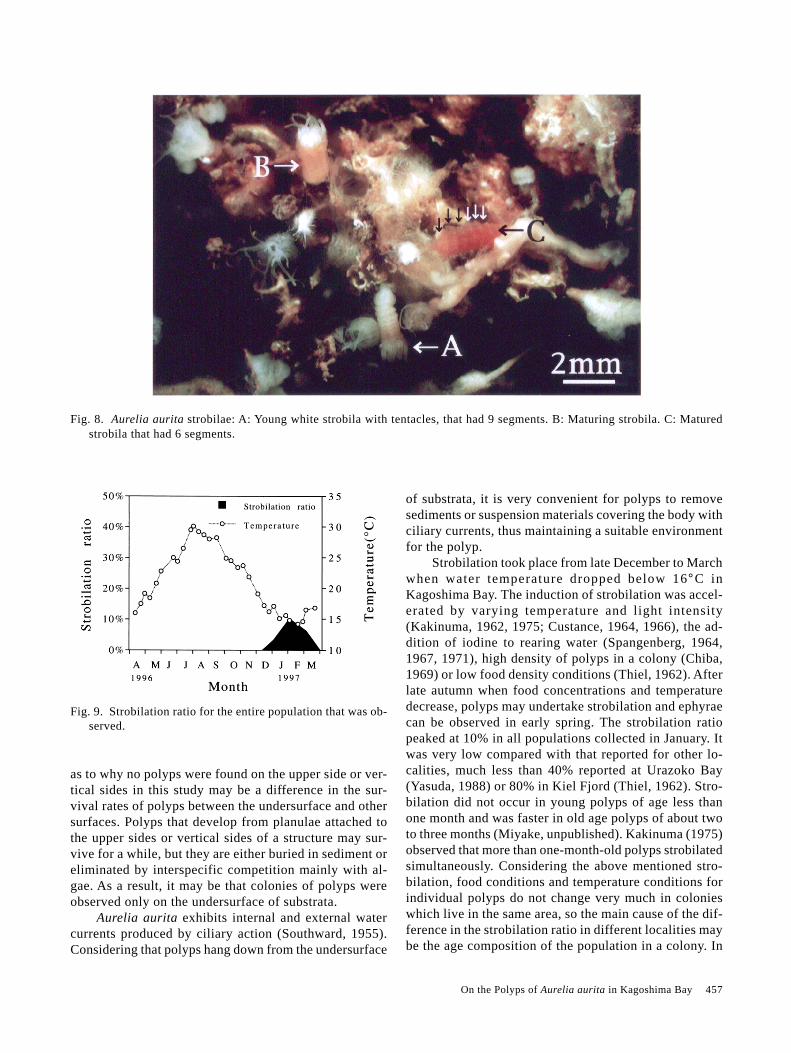
middle of January to early February when water tempera-ture was 14–15°C, and continued through late February(Fig. 8). Not all polyps in a colony developed into strobi-lae. The strobilation ratio was 3.2% of the polyps col-lected in late December, 10.1% in January (the peakmonth) and 6.1% in late February (Fig. 9). Strobila had7–9 segments, though the maximum segment number was14.
4. DiscussionThe in situ habitat of Aurelia polyps was found at
Taniyama. This area was one of developmental areas ofephyrae in Kagoshima Bay. Polyps were attached only tothe horizontal undersurface of artificial structures likefloating piers or buoys to 1 m depth. There were manysessile organisms on the undersurface of the floating piersor buoys, however polyps were observed on Mytilus sp.,barnacles, dead oyster shells, solitary ascidians,polychaete tubes, amphipod tubes and gap spaces. We alsoobserved polyps only on the undersurface of floating piersat the Murotsu fishing port in Shimonoseki, Yamaguchi,Japan. They were attached only to Mytilus, hard solitaryascidians and floats, though there were many natural sub-strata that had overhang structures such as bare rocks orstone wall embankments and many sessile organisms suit-able for polyp attachment around there. There may besubstratum selection in the polyp of Aurelia. Previouslyreported substrata for the polyps of Aurelia includeMytilus, algae, the shells of living or dead barnacles, tu-nics of ascidians, hydrozoans and bare rocks (Ussing,1927; Sjorgern, 1962; Thiel, 1962; Rasmussen, 1973;Hernroth and Gröndahl, 1985a) (Table 1). Green algaeand brown algae were also used as polyp substrata inKagoshima Bay. However, these substrata are not perma-nent, since they disappear after late spring.
One of the striking features in this study is that pol-yps have a distinct preference for the undersurface of man-made structures like floating piers and buoys. Even ifsuitable substrata exist on the upper or vertical sides ofman-made structures, no polyps were attached to thosesides. Brewer (1978) explained this phenomenon using alaboratory experiment on planula settlement. Nearly allplanulae attached to the undersides of structures. How-ever, a small number of planulae also attached to theupperside. In our laboratory tank, polyps also formedcolonies on the bottom and the wall of aquaria. It wasobserved that the polyps on the bottom of aquaria per-ished from being buried under sediments and that polypswhich formed colonies on the wall of aquaria had possi-ble attachment substrata reduced by the encroachment ofalgae and then asexual reproduction was repressed. Fi-nally some polyps formed cysts on the wall and othersdied, consequently colonies disappeared from the wall andbottom of the aquaria (Miyake, unpublished). The reason
Fig. 7. Iδ-quadrat size relationships and Iδ(s)/Iδ(2s)-quadrat sizerelationships of polyp colonies. (a) On Mytilus sp. in Fig.5. (b) On a polystyrene substratum in Fig. 6a. (c) On a poly-styrene substratum in Fig. 6b.
inds./cm2. The Iδ value was about 1.4 at a quadrat size of31.3 mm2 (64 quadrats). Colonies had an aggregated dis-tribution with large colony sizes, and polyps were evenlydistributed within the colony. The unit colony size was62.5–125 mm2 (Fig. 7b).
One of the typical distribution patterns of polyps onthe polystyrene substratum to which many fouling spe-cies were attached (Fig. 6b), the density of polyps was3.1 inds./cm2. The Iδ value was about 2.0 at a quadrat sizeof 7.8 mm2 (256 quadrats). Colonies had an aggregateddistribution with small colony sizes, and polyps were dis-tributed evenly within the colony. The unit colony sizewas 7.8–15.6 mm2, and larger colony sizes of 125–250mm2 were formed by aggregations of the unit colonies(Fig. 7c).
3.4 StrobilationStrobilation began from late December when water
temperature fell to 16–17°C. It reached its peak from the
On the Polyps of Aurelia aurita in Kagoshima Bay 457
of substrata, it is very convenient for polyps to removesediments or suspension materials covering the body withciliary currents, thus maintaining a suitable environmentfor the polyp.
Strobilation took place from late December to Marchwhen water temperature dropped below 16°C inKagoshima Bay. The induction of strobilation was accel-erated by varying temperature and light intensity(Kakinuma, 1962, 1975; Custance, 1964, 1966), the ad-dition of iodine to rearing water (Spangenberg, 1964,1967, 1971), high density of polyps in a colony (Chiba,1969) or low food density conditions (Thiel, 1962). Afterlate autumn when food concentrations and temperaturedecrease, polyps may undertake strobilation and ephyraecan be observed in early spring. The strobilation ratiopeaked at 10% in all populations collected in January. Itwas very low compared with that reported for other lo-calities, much less than 40% reported at Urazoko Bay(Yasuda, 1988) or 80% in Kiel Fjord (Thiel, 1962). Stro-bilation did not occur in young polyps of age less thanone month and was faster in old age polyps of about twoto three months (Miyake, unpublished). Kakinuma (1975)observed that more than one-month-old polyps strobilatedsimultaneously. Considering the above mentioned stro-bilation, food conditions and temperature conditions forindividual polyps do not change very much in colonieswhich live in the same area, so the main cause of the dif-ference in the strobilation ratio in different localities maybe the age composition of the population in a colony. In
Fig. 9. Strobilation ratio for the entire population that was ob-served.
Fig. 8. Aurelia aurita strobilae: A: Young white strobila with tentacles, that had 9 segments. B: Maturing strobila. C: Maturedstrobila that had 6 segments.
as to why no polyps were found on the upper side or ver-tical sides in this study may be a difference in the sur-vival rates of polyps between the undersurface and othersurfaces. Polyps that develop from planulae attached tothe upper sides or vertical sides of a structure may sur-vive for a while, but they are either buried in sediment oreliminated by interspecific competition mainly with al-gae. As a result, it may be that colonies of polyps wereobserved only on the undersurface of substrata.
Aurelia aurita exhibits internal and external watercurrents produced by ciliary action (Southward, 1955).Considering that polyps hang down from the undersurface

458 H. Miyake et al.
short, colonies including many old age polyps may ex-hibit a high ratio of strobilation.
Colony form showed an aggregated distribution.Polyps settled regularly or randomly within the colony.This suggests that there is an intraspecific spatial rela-tionship in polyps. The Iδ value was higher on substratawhere many other species existed than on substrata wherefew species existed.
The densest part of a colony in this study, up to 88inds./cm2, was very high compared with the 6–40inds./cm2 reported in Gullmar Fjord, Sweden (Gröndahl,1988a, b). These observations, including ours, suggest thatpolyps form large colonies using a variety of asexual re-productive modes in competition with other fouling spe-cies. There were some variations in density on every sub-stratum in every investigation throughout the year. It maybe that those variations did not derive from seasonal vari-ation, but from the structure of the colony such as the agecomposition, the number of planulae recruited on the sub-strata and the existence of other species. It was very dif-ficult to set up the quadrat on the undersurface of a float-ing pier. Moreover, it was impossible to replace thecolony, which was observed under a stereoscopic micro-scope in order to observe the colony structure, to the sameplace to which it attached in nature. Accordingly we wereunable to follow the development of the same colony atthe same place continually. It was also very difficult toobtain quantitative data on colony density, because thepolyp colonies were distributed in patches and the den-sity value would change according to the method used todecide the total area of the colony substratum. To clarifythe population dynamics of colonies, an experiment us-ing a flat float made of polystyrene in the same area andalso an experiment on colony formation in the laboratorymust be conducted.
In this study, it was clear that favorable conditionsfor the settlement of Aurelia polyps were the presence ofgap spaces and sessile organisms with hard bodies, onthe undersurface of structures. The undersurfaces of natu-ral structures are not common in the field. Almost all ofthe structures that have a flat undersurface are artificialstructures such as floating piers, buoys, quay walls andembankments. Artificial structures are common in thecoastal area of Japan. Recently, because of the great in-fluence of Aurelia aurita on the coastal marine ecosys-tem and on our coastal economic activities like fishingactivities and electric power plant cooling (Matsueda,1969), field studies of the Aurelia medusa stage have beenincreasing. The increase in underwater structure of an-thropogenic origin could be related to recent increases injellyfish biomass in pelagic ecosystems. Thus, we needto research not only the medusa stage, but also the polypstage to understand the population dynamics of Aureliaaurita.
AcknowledgementsWe would like to thank Dr. Mary N. Arai (Pacific
Biological Station, Nanaimo, B.C., Canada), Dr. RichardD. Brodeur (Northwest Fisheries Science Center, HatfieldMarine Science Center, NOAA, USA), Dr. Dhugal J.Lindsay and Dr. James C. Hunt (JAMSTEC) for criticalreading and improvement of the manuscript. We thankalso Drs. Toshihiro Ichikawa and Masanori Sato atKagoshima University for providing everything for ourstudy. We thank students from the Department of Biol-ogy, Faculty of Science, Kagoshima University for help-ing with the field investigations. Heartfelt thanks go toDr. Ueno at the National Fisheries University who gaveus the chance to SCUBA dive at the Murotsu fishing port.Finally, we appreciate useful comments and criticism ofthe manuscript from the anonymous reviewers.
ReferencesArai, M. N. (1997): A Functional Biology of Scyphozoa.
Chapman & Hall, London, 293 pp.Brewer, R. H. (1978): Larval settling behavior in the jellyfish
Aurelia aurita (Linneaus) (Cnidaria: Scyphozoa). Estuar-ies, 1, 120–122.
Chiba, Y. (1969): Ecological factors affecting the strobilationof Aurelia aurita (Coelenterata: Scyphomedusae). Bull. Mar.Biol. Stat. Asamushi, 13, 173–178.
Custance, D. R. N. (1964): Light as an inhibitor of strobilationin Aurelia aurita. Nature, 204, 1219–1220.
Custance, D. R. N. (1966): The effect of a sudden rise in tem-perature on strobilae of Aurelia aurita. Experientia, 22, 588–589.
Gröndahl, F. (1988a): Interactions between polyps of Aureliaaurita and planktonic larvae of scyphozoans: an experimen-tal study. Mar. Ecol. Prog. Ser., 45, 87–93.
Gröndahl, F. (1988b): A comparative ecological study on thescyphozoans Aurelia aurita, Cyanea capillata and C.lamarckii in the Gullmar Fjord, western Sweden, 1982 to1986. Mar. Biol., 97, 541–550.
Gröndahl, F. (1989): Evidence of gregarious settlement ofplanula larvae of the scyphozoan Aurelia aurita: an experi-mental study. Mar. Ecol. Prog. Ser., 56, 119–126.
Hernroth, L. and F. Gröndahl (1983): On the biology of Aureliaaurita (L.) 1. Release and growth of Aurelia aurita (L.)ephyrae in the Gullumar fjord, Western Sweden, 1982–83.Ophelia, 22, 189–199.
Hernroth, L. and F. Gröndahl (1985a): On the biology of Aureliaaurita (L.) 2. Major factors regulating the occurrence ofephyrae and young medusen in the Gullmar fjord, WesternSweden. Bull. Mar. Sci., 37, 567–576.
Hernroth, L. and F. Gröndahl (1985b): On the biology of Aureliaaurita (L.) 3. Predation by Coryphella verrucosa(Gastropoda, Opisthobranchia), A majour factor regulatingthe development of Aurelia populations in the Gullmar fjord,western Sweden. Ophelia, 24, 37–45.
Kakinuma, Y. (1962): On some factors for the differentiationsof Cladonema utidai and Aurelia aurita. Bull. Mar. Biol.Stat. Asamushi, 11, 81–85.

On the Polyps of Aurelia aurita in Kagoshima Bay 459
Kakinuma, Y. (1975): An experimental study of the life cycleand organ differentiation of Aurelia aurita Lamarck. Bull.Mar. Biol. Stat. Asamushi, 15, 101–112.
Matsueda, N. (1969): Presentation of Aurelia aurita at thermalpower station. Bull. Mar. Biol. Stat. Asamushi, 13, 187–191.
Miyake, H., K. Iwao and Y. Kakinuma (1997): Life history andenvironment of Aurelia aurita. South Pacific Study, 17, 273–285.
Morisita, M. (1959): Measuring of the dispersion of individualand analysis of the distributional patterns. Mem. Fac.Kyushu Univ., Ser. E, 2, 215–235.
Rasmussen, E. (1973): Systematics and ecology of the Ise fjordmarine fauna (Denmark). Ophelia, 11, 41–46.
Sjorgern, L. (1962): Scyphistoma-stage of the jellyfish, Aureliaaurita, on Fucus at Tvarminne Zoological Station. Mam.Soc. Fauna Flora Fenn., 37, 3–4.
Southward, A. J. (1955): Observations on the ciliary currentsof the jelly-fish Aurelia aurita L. J. Mar. Biol. Ass. U.K.,
34, 201–216.Spangenberg, D. B. (1964): A study of strobilation in Aurelia
aurita under controlled conditions. J. Exp. Zool., 160, 1–10.
Spangenberg, D. B. (1967): Iodine induction of metamorpho-sis in Aurelia. J. Exp. Zool., 165, 441–450.
Spangenberg, D. B. (1971): Thyroxine induced metamorphosisin Aurelia. J. Exp. Zool., 178, 183–194.
Thiel, H. (1962): Untersuchungen uber die strobilisation vonAurelia aurita Lam. an einer population der Kieler Forde.Kieler Meeresforsch., 13, 198–230.
Ussing, H. (1927): Bidrag til Aurelia aurita’s biologiiMariagerfjord. Vid. Meddr dansk naturhist. Foren, 84, 91–106.
Yasuda, T. (1988): Studies on the Common Jelly-Fish, Aureliaaurita (Linné). Japan Fisheries Resource Conservation As-sociation, Tokyo, 139 pp. (in Japanese).