open access ec pharmacology and toxicology research ... · cronicon open access ec pharmacology and...
TRANSCRIPT

CroniconO P E N A C C E S S EC PHARMACOLOGY AND TOXICOLOGY
Research Article
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in
Sodium Valproate-Toxicity
Lucky Legbosi Nwidu1* and Yibala Ibor Oboma2
1Experimental Pharmacology and Toxicology, Faculty of Pharmaceutical Sciences, University of Port Harcourt, Choba, Rivers State, Nige-ria2Medical Laboratory Department, Faculty of Basic Medical Sciences, College of Health Sciences, Niger Delta University, Amassoma, Yenegoa, Bayelsa State, Nigeria
*Corresponding Author: Lucky Legbosi Nwidu, Experimental Pharmacology and Toxicology, Faculty of Pharmaceutical Sciences, University of Port Harcourt, Choba, Rivers State, Nigeria.
Received: April 06, 2019; Published: May 02, 2019
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Keywords: Musanga cecropioides; Sodium Valproate; Oxidative Stress; Glia Fibrillary Acidic Proteins; Neurofibrillary Tangle; Prefron-tal Cortex; Immunohistochemistry
Valproic acid (VPA) is a foremost drug in the management of epilepsy and several kinds of seizures. Cellular toxicities concerns of VPA arising from the treatment of diseases have received global attention. Oxidative stress underpins VPA-induced neurotoxicity and is caused by free radicals in the brain tissue. The aim of our investigation was to evaluate the protective effects of Musanga ce-cropioides (MCS) against VPA-induced toxicity in prefrontal cortex rat brain in vivo model of neurotoxicity. Oxidative stress such as lipid peroxidation (LPO), glutathione reductase (GSH), glutathione peroxidase (GPx), glutathione transferase (GST), catalase (CAT), superoxide dismutase. Besides, the expression of glial fibrillary acidic protein (GFAP) and neurofibrillary tangles (NTs) neurological markers were appraised. The levels of oxidative stress markers, LPO, GSH, GST, GPx, CAT, and SOD were significantly (P < 0.05 - 0.001) elevated. Valproate exposure significantly produces alteration of the activities of the glutathione metabolizing enzymes such as glu-tathione-S-transferase, glutathione peroxidase, LPO and the expression of GFAP and NTs tangles. Post-treatment with MCS reversed the VPA-induced effects in brain tissue preparation of rat and mitigated the expression of GFAP and NTs. Based on the protective and antioxidant action of MCS and abrogation of the expression of GFAP and NTs in the PFC of the brain, we propose that MCS possess potential safe active compounds that could be considered effective therapeutic approach in attenuating the adverse effects of VPA-induced neurotoxicity.
Abstract
IntroductionReactive oxygen species (ROS) play an important role in physiological functions [1]. Upregulation of reactive species, especially reac-
tive oxygen (ROS) and nitrogen (RNS) species in addition to the imbalance in the body’s antioxidant enzyme systems will results in de-struction of cellular structures, lipids, proteins, and genetic materials such as DNA and RNA [2]. Moreover, the effects of reactive species on mitochondria and their metabolic processes eventually cause a rise in ROS/RNS levels, leading to oxidation of mitochondrial proteins, lipids, and DNA. Oxidative stress has been considered to be linked to the etiopathology of many diseases, including neurodegenerative diseases (NDDs) such as Alzheimer diseases, Amyotrophic lateral sclerosis, Friedreich’s ataxia, Huntington’s disease, Multiple sclerosis,

405
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
and Parkinson’s diseases. In addition, oxidative stress causing protein misfold may turn to other NDDs include Creutzfeldt-Jakob disease, Bovine Spongiform Encephalopathy, Kuru, Gerstmann-Straussler-Scheinker syndrome, and Fatal Familial Insomnia [1,3-7].
Oxidative stress has been proven to be involved in VPA-induced toxicity [8,9]. Valproic acid (VPA) is broad spectrum antiepileptic drug used for the management of idiopathic and symptomatic generalized seizures, neurological and psychiatric illness [10-12]. Diverse molecular and cellular events underlie different types of seizures [13].
Though valproate offer protection in some neurological disease investigated [14-16], other reports indicated that valproate - induced neurodegeneration in both cultured neuronal cells and experimental animals [17-20]. Intraperitoneal valproate injected rats reveal the decimation of the proliferation of hippocampal neurons and cognitive impairment [17,21] and clinically relevant doses of VPA therapy administered to neonatal mice and rats exhibited widespread apoptotic neurodegeneration in several brain regions [22,23]. Teratogenic activity in humans induced by VPA has been established [24,25]. Besides, neural tube defects and increases the incidence of children with autism spectrum disorder (ASD) were reported when taken during pregnancy [26,27]. Free radicals generation and concomitant oxida-tive stress was reported in developmental neurotoxicity observed with valproate therapy [28] and caspase-dependent apoptosis [18,29]. In the brains of autistic children the histomorphological changes observed confirmed that VPA induces programmed cell death (PCD) [30-32]. Though, the pathways involved in the VPA-induced neurotoxicity is still poorly understood [33]. Caspase-independent PCD was also reported [34] but its exact mechanism is unknown. Valproate induced cascades of deleterious events contributing to neuronal cell death via atypical calpain-dependent necroptosis pathway [35,36]. The valproic acid activated calpain-dependent necroptosis cascade is recognized as the principal player in apoptosis [37-39].
Glial fibrillary acidic proteins (GFAP) play important physiological roles in brain functions. The presence of GFAP indicates astro-cytes or ependymal differentiation. Glial fibrillary acidic protein (GFAP) is an intermediate filament (IF) III protein exceptionally found in astrocytes and a main type of glial cells in the central nervous system [40,41]. In traumatic brain and spinal cord injuries and stroke, GFAP and its metabolites are quickly released into biological fluids, presenting them as strong candidate biomarkers for such neurologi-cal disorders. Intermediate filament (IF) in astrocytes is a crisis-command center coordinating cell responses in situations connected to cellular stress, which is a central component of many neurological diseases [40,41]. Activation of astrocytes has been implicated in the pathogenesis of a variety of neurodegenerative diseases and astrocyte-mediated pathways as targets for drugs treating the root causes of the pathology [42,43]. The activated astrocytes secrete several neurotropic factors necessary for neuronal survival; it is believed that rapid and severe activation of astrocytes causes secretion of neurotoxic substances that augments/initiates an inflammatory response, leading to neuronal death and brain injury. Thus, the injury then causes astrocyte to be express can enhanced the level of glial fibrillary acid protein (GFAP) expression which is considered a marker protein for astrogliosis [42,43]. Activation of astrocytes is characteristic morphological hallmarks and has important functional consequences in situations such as stroke, trauma, epilepsy, Alzheimer’s disease (AD) and other neurodegenerative diseases [44,45]. Neurofibrillary tangle or protein (NFP) is mostly commonly known as a primary hall marker of Alzheimer’s disease [45]. Their presence is also found in numerous other diseases known as tauopathies. These neurofibril-lary tangles are also found in the brain of normal aged humans. Similarly, neurofibrillary changes are also found in Guan-Parkinsonism dementia complex, mongolism, post encephalitic Parkinsonism and tuberous sclerosis [46].
The reference drug, vinpocetine, utilized in this study, is an apovincaminic acid ethyl ester, a semi-synthetic derivative of vincamine, an alkaloid extract derived from the periwinkle plant (Vinca minor). Vinpocetine is accessible as a prescription pharmaceutical worldwide; it, is indicated for management of cerebrovascular and cognitive disorders [47]. It is usage as dietary supplements underpin advocated use for claims of cognitive enhancement [48-52].
Finally, the medicinal importance of Musanga cecropioides has been extensively reported in M. cecropioides subchronic toxicity [53], anti-hyperglycemic effect [54], hepatoprotective effect [55], inhibition of edema [56] and antihypertensive agent [57].
Here, we investigated for the first time the neuroprotective effect of MCS extract on VPA - induced neurotoxicity in rat brain PFC. The relationship of several important molecules such as lipid peroxidation (MDA), superoxide dismutase (SOD), catalase (CAT), glutathi-one reductase (GSH), glutathione peroxidase (GSPx), glutathione-S-transferase (GST), glial fibrillary acidic proteins (GFAP) and neuro-fibrillary proteins (NFP). The significant downregulation of LPO and GFAP, NFP protein expression and upregulation of enzymatic and

406
non-enzymatic antioxidants alterations by MCS extract reveal its potential health benefits and probable exploitation as pharmaceuticals for mitigating VPA- induced toxicity in anticonvulsant therapy and management for neurodegenerative diseases.
Materials and Methods
Drugs, Chemicals and equipment
Sodium valproate (Epilim, Sanofi, France), Vinpocetine (Cognitol, Tyonex, Nigeria) both purchased from Luckpharm Pharmacy Int’l Ltd (Nigeria), Rivers State, Nigeria. n-hexane (extrapure 85%) (LobalChemie, Mumbai, India), Methanol 99.8% (LobalChemie, Mumbai, India), formalin (LobalChemie, Mumbai, India), Diethyl ether (LobalChemie, Mumbai, India); Avidin Biotin Complex (Bosterbioengener-ing limited, Wuhan, China). The equipment utilized includes: rotary evaporator (Shenke® R-205, ShangaiShenshun Biotechnology Co. Ltd, China), analytical balance model AR323 CN (Ohaus Corp. Pine Brook, NJ, USA), auto-hematology analyzer model MY-B002B (Maya Medical Equipment Limited, Shanghai, China), Spectrophotometer model SM-23 D (Surgifield Medical, England, UK), scientific weighing balance model TH 600 (Labscience, England, UK), centrifuge model 412B (Techmel and Techmel, MI, USA), Water bath (Techmel and Techmel, MI, USA).
Collection and authentication of plant materials
Musanga cecropioides stem-bark was collected and authenticated by, a taxonomist (Dr. Oladele Adekunle) attached to the Forestry Department at university of Port Harcourt from the University of Port Harcourt campus, Nigeria. The Herbarium specimen with voucher number UUPH 2001 had already been deposited at Department of Pharmacognosy, University of Uyo, Akwa Ibom State, Nigeria.
Preparation of Musanga cecropioides stem-bark (MCS) extract
The dried M. cecropioides stem-bark (MCS) was pulverized to fine particles using mechanical grinder. A 500g weight of MCS powder was then macerated in 2000 mL of n-hexane for defatting. After 24 hours of maceration, the extract was concentrated using a rotary evaporator and the marc submerged in 2000 mL of methanol. It was macerated for 72 hours while shaking vigorously every 2 hour for 12 hours. Rotary evaporator was used to concentrate the extract and evaporated to dryness on the water bath at 45°C. The percentage yield was then calculated to be 15.7%.
Phytochemical screening
Phytochemical screening of the M. cecropioides extract was executed at the Pharmacognosy and Phytotherapy Department laboratory, University of Port Harcourt. The bioactive agents screened include: alkaloids, triterpenoids, flavonoids, cardiac glycosides, saponins, and phlobatannins using standardized protocol [58].
Animals and drug administration
Forty two male Wistar albino rats were used. The animals (180 - 200g) were obtained from the animal house, Department of Pharma-cology and Toxicology, University of Nigeria Nsukka. The animals were acclimatized in the University of Port Harcourt Animal House for 14 days under standard laboratory conditions. The animals were randomly selected and housed in pathogen free plastic cages (n = 7) per cage under relative humidity (40 - 55%) and ambient temperature (26°C) and exposed to daily circle of night and day. The rats were pro-vided with pelleted rodent chow (Vital Feeds, Onitsha, Anambra State, Nigeria) and water ad libitum and animals were allowed unfettered access to water and food. The experimental protocol was in line with the Guide to the Care and Use of Animals in Research and Teaching (NIH, 1996) and institutional guideline for care and use of animals for experiment as specified in the University of Port Harcourt Animal ethics committee approval (No. UPHAEC/2018/008).
Rationale for dose selection (Valproic acid)
Low doses of VPA such as 200 mg/kg, s.c. (mice) [59], 350 mg/kg, i.p. (rat) [60] and 400 mg/kg (mice) [59,61] have been utilized for experimental investigations. Moderate doses of VPA such as 500 mg/kg and 600 mg/kg (i.p. /s.c.) [62-66] were used in mice. High dose of VPA (800 mg/kg) was also used [67]. Animal studies report a significant amount of VPA (900 µg/ml) in the maternal plasma within 1h of VPA administration [63,65]. Although majority of the reports followed 500 mg/kg [68], therefore this dose was utilize in the present study. Sodium valproate or valproic acid (VPA) brand Epilim® formulated as 300 mL syrup was used to induce neurotoxicity at a dose
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity

407
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
500 mg/kg daily for 28 days when administered orally by gavage in the experimental animals [68]. Each one mL contains 200 mg sodium valproate. The animals were divided randomly into six groups with 7 animals per group.
Rationale for dose selections (Vinpocetine)
The oral median lethal dose (LD50) of vinpocetine in rats (strain not specified) is approximately 500 mg/kg [69]. Toxicity of vinpo-cetine was investigated by [69] in a multiple studies in rats (strain not specified). Report indicated that subchronic gavage exposure to vinpocetine at doses between 25 and 100 mg/kg elevated salivation, liver, and thyroid weights especially with the highest dose. But with intraperitoneal injection of 5 or 25 mg/kg for 3 months, mortality was recorded. In a similar report chronic gavage exposure, at doses between 25 and 100 mg/kg, no adverse effects were stated [69]. Since the lowest dose commonly used is 25mg/kg, this those was utilize in the current study. Oral bioavailability of vinpocetine in rats was 52% suggesting extensive first pass metabolism [70].
Experimental doses (MCS, valproate and vinpocetine)
The acute toxicity (LD50) of MCS indicated that at dose above 5 g/kg did not show any sign of toxicity as reported in previous publica-tion [53]. Therefore, three dose levels, 100, 200 and 400 mg/kg were utilized for this investigation.
The MCS, valproate and vinpocetine (Cognitol®) (the reference drug) were administered orally per kg of body weight once daily for 28 days. Sodium valproate (500 mg/kg) was administered one hour prior to the administration of the control drugs or extracts respectively for animals in groups 2 to 6. The MCS extract (100, 200, 400 mg/kg) and vinpocetine 25 mg/kg were solubilized in 2% Tween 80 (Poly-sorbate 80). The experimental groups utilized for the study are as follows:
• Group 1 (negative control): The animals in this group received 2% Tween 80 with 10 ml/kg distil water.
• Group 2 (disease control group): The animals in this group received sodium valproate followed by 2% Tween 80 with 10 ml/kg distil water
• Group 3 to 5 (test groups): The animals in these groups received sodium valproate (500 mg/kg) followed by the MCS extract 100, 200 and 400 mg/kg respectively.
• Group 6 (reference control): The animals in this group received sodium valproate followed by vinpocetine 25 mg/kg.
The rats were administered valproic acid (500 mg/kg b.w.) and one hour later, either distilled water, MCS or vinpocetin was adminis-tered adopting standard procedure [68].
Neurotoxicity percentage (%) was deduced using this formula:
=
Neuroprotective activity (%) was calculated as follows:
Where, MCS, VPA, and W are mean experimental variables estimated in the rats treated with valproic acid (VPA) plus MCS (Test groups), valproic acid (VPA) only (diseases control group) and distil water treated animals (W) (negative control) respectively [43].
Tissues processing
All rats were sacrifice killed and the whole brain excised and weighed immediately. The prefrontal cortex was removed and immersed in ice-cooled physiological saline. A 10% homogenate of the PFC was prepared. The homogenate was centrifuge (4000 × g, 10 min) at - 80°C to remove debris and nuclei. The resultant supernatant was stored at -80°C for various biochemical assays.
Oxidative stress determinations
Lipid peroxidation (Malondialdehyde, MDA): Lipid peroxidation was examined by the thiobarbituric acids reactive substances (TBARS) method [71]. An aliquot of 0.4 mL of supernatant was mixed with 1.6 mL of Tris-KCl buffer and 0.5 mL of 30% trichloroacetic acid (TCA) added. Finally, 0.5 mL of 0.75% TBA was added and then placed in a boiling water bath for an hour. This was then cooled in ice and centrifuged (3000 × g, 10 min). The clear supernatant absorbance was measured against a reference blank of distilled water at 532
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity

408
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
nm and the MDA level estimated by Todorova., et al. [72] method and expressed as nmol of MDA/g of wet tissue using a molar extinction coefficient of the chromophore (1.56 × 10-5 /M/cm).
Reduced Glutathione (GSH): Sedlak and Lindsay [73] method was used to evaluate the level of reduced glutathione. To an aliquot sample (0.2 mL) was added 1.8 mL of distilled water and 3 mL of the precipitating solution (4% sulphosalicylic acid) and vortexed. The mixture was allowed to stand for approximately 5 minutes and then centrifuged (1200 × g, 10 min). To 1 mL of the filtrate was added 4 mL of 0.1M phosphate buffer and 0.5 mL of DTNB (Ellman’s reagent). Similarly, a blank was prepared by addition of 4 mL of 0.1M phosphate buffer, 1 mL of dilute precipitated solution and 0.5 mL DTNB. The solution was incubated at room temperature for 15 minutes and read at 412 nm on a spectrophotometer.
Glutathione peroxidase: The level of glutathione peroxidase (GSPx) was estimated using by adopting the method of Rotruck., et al [74]. Sodium phosphate buffer mixture (1 mL) was prepared by combing 0.2 mL sodium azide, 0.4 mL of GSH and 0.2 mL of hydrogen peroxide (H2O2) and 1 mL of 1:10 extract of cell homogenate and made up to 4 mL volume with distilled water. This was incubated for 3 minutes at 37ºC before 0.5 mL 10% TCA was added to terminate the reaction. To estimate the remaining glutathione constituent, DTNB reagent (1 mL) and phosphate solution (0.3 Mmol/L) were added. The change in color was assayed at 412 nm. Similar treatments were administered to aliquots of the standard solutions.
Catalase (CAT): Catalase level was evaluated according to the method of Clairborne [75] with slight modifications. The method is based on the ability of catalase in the sample preparation to split hydrogen peroxide which and can be measured spectrophotometrically at 240 nm. One unit of catalase equals the amount of protein that converts one micromole H2O2/min. A 0.2 mL of sample was added to 50 mM of phosphate buffer (pH 7.4) containing 100 mM (v/v) of H2O2 in a total of 1 mL. The reaction mixture was incubated for 2 minutes at 37ºC and the rate of absorbance change (AA/min) at 240 nm was recorded which indicated the decomposition of H2O2. Activities were calculated using the molar extinction coefficient of H2O2 at 240 nm. All samples were measured in quadruplicates.
Superoxide dismutase (SOD): Superoxide dismutase (SOD) level was evaluated using the auto-oxidation technique [76]. Tissue ho-mogenates were centrifuged (12, 000g for 10 minutes) and an aliquot of supernatant was diluted with water to make a 1:10 dilution. The diluted sample (200 mL) was added carbonate buffer (pH 10.2) and 0.3 mL of 0.3 mmol/L epinephrine was supplemented to the mixture and mixed by inversion swiftly. All solutions used were freshly prepared. Two and half milliliter of buffer, 0.3 mL of epinephrine (sub-strates) and 0.2 mL of distilled water were in the reference cuvette. The change in absorbance was read at 480 nm read at 30 s intervals for 2.5 minutes.
Glutathione-S-transferase (GST): Glutathione-S-transferase level was determined in accordance with the method of Habig., et al [77]. The principle is based upon the relatively high activity of GST with 1-chloro-2, 4-dinitrobenzene (CDNB) as the second substrate. When CDNB is conjugated with reduced GSH, its absorption maximum shifts to a longer wavelength. The absorption increase at the new wavelength of 340 nm provides a direct measurement of the enzymatic reaction. The medium for estimation was prepared with reduced GSH 0.1M, CDNB 20 mM and 0.1 M phosphate buffer and the reaction was allowed to run for 60 seconds each time before the absorbance of the medium containing the sample was read against the blank at 340 nm. The absorbance was measured using spectrophotometer. Proteins determination: Protein content estimation was determined from PFC of rat brain tissues by the modified method of Lowry assay [78] using bovine serum albumin (BSA) standard.
Histopathology
Hematoxylin and eosin staining
After 28 days exposure to valproic acid and MCS, all rats were sacrificed by decapitation. The prefrontal cortex was excised immedi-ately, fixed by 20% phosphate buffered formalin for 48hrs. Tissues were sectioned using rotary microtome model (Leica Microsystems, Wetzlar, Germany at 6µm and floated in water bath onto charged slides. The sections were dried on hot plate at 60°C for 2 hours. The tissue were further processed by dehydrating with ascending grades of alcohol, cleared in two changes of xylene and embedded in mol-ten paraffin wax and sectioned using rotary microtome, mounted on glass slide and stained with hematoxylin and eosin. Ultra-sections
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity

409
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
from each group were examined by light microscopy for tissue damage and neurodegenerative features such as shrinkage of the neuron, hyperchromasia, and nuclear pyknosis as revealed by hematoxylin and eosin staining techniques following the procedure of Oyinbo., et al [79]. Cortical Purkinje cells estimation was based on semi-quantitative scale described in previous study [79] was used to assess the extent of neurodegeneration in the PFC with 400× magnification.
Immunohistochemistry of GAFP and NFP
Tissue preparation
The abstracted prefrontal cortex which was excised is fixed in 4% formaldehyde in 0.01 mmol/L phosphate buffer and embedded in paraffin. Tissues were sectioned 3 µm using rotary microtome model (Leica Microsystems, Wetzlar, Germany). Slice were mounted on a clean slide and toasted at 60°C for 2 hrs. Treatment with dimethyl benzene was instituted for 20 minutes. Slices were sequentially treated with 95%, 80% and distilled water until transparency of slide was achieved.
StreptAvidin- Biotin Complex (SABC) Histochemistry
Immunohistochemistry assessment of neuronal damage was executed by evaluating the levels of glial fibrillary acid protein (GFAP) and neurofibrillary protein (NFP) biomarkers. The tissue sections were placed in 0.3% hydrogen peroxide (H2O2 in phosphate buffer solu-tion (PBS) for 30 minutes, then pre-incubated with normal goat serum to prevent nonspecific binding at 4°C for 30 minutes after which further incubation with a primary polyclonal rabbit anti-GFAP or - NFT (1:1000; Sigma) antibody at 37°C for 1 hr or overnight at 4°C. The tissue slices were washed thrice for 10 minutes with PBS; the sections were then incubated for 1 hr at room temperature with their corresponding horseradish peroxidase conjugated anti-species secondary antibody (polyclonal goat anti-rabbit IgG) immune-labelled against GFAP or NFT.
Antibody immunoreactivity or Immunoreactions were observed under an optical microscopes described by Oboma., et al. [80] was adopted. Cells with specific brown colors in the cytoplasm, cell membrane or nuclei depending on the antigenic sites were considered to be positive for glial fibrillary acid protein (GFAP) and neurofibrillary protein (NFP). The stained cells without any form of brown colors were scored as negative.
Statistical analysis
The Graph pad Prism 5.1 was used for data analysis by one-way analysis of variance (ANOVA). Multiple comparisons among groups were made according to the Turkey’s test. P values < 0.05 were considered significant. Data are represented as expressed as Mean ± stan-dard deviation (SD).
Results
Effect of MCS on MDA, antioxidant and protein levels
The effect of MCS after sub-acute intoxication with valproic acid on antioxidant activity is presented in table 1. The result indicated that VPA treated rats exhibited increased level of neuronal damage and oxidative stress witnessed by statistical significant depression (P < 0.05 - 0.001) of SOD (47%), GSH (61%), GST (84%), GPx (90%), CAT (54%) and TP (9%) compared with the normal control group. However, sub-acute treatment with MCS extract protected against the neurotoxicity induced by valproic acid. This neuroprotection was demonstrated by the elevation of SOD (121%, 124%, 189%), GSH (70%, 115%, 102%), GST (67%, 135%, 98%), GPx (78%, 60%, 95%), CAT (57%, 81%, 76%) and TP (59%, 105%, 122%); and reduction of MDA (89%, 101%, 98%) with MCS treatment at 100 mg/kg, 200 mg/kg and 400 mg/kg body weight respectively when compared with the disease control group. Vinpocetine demonstrated neuroprotective activity on antioxidant enzymes with increased in SOD, GSH, GST, GPx, CAT, and TP by 86%, 83%, 128%, 65%, 86%, and 68% but reduced lipid peroxidation in brain prefrontal cortex by 96%.
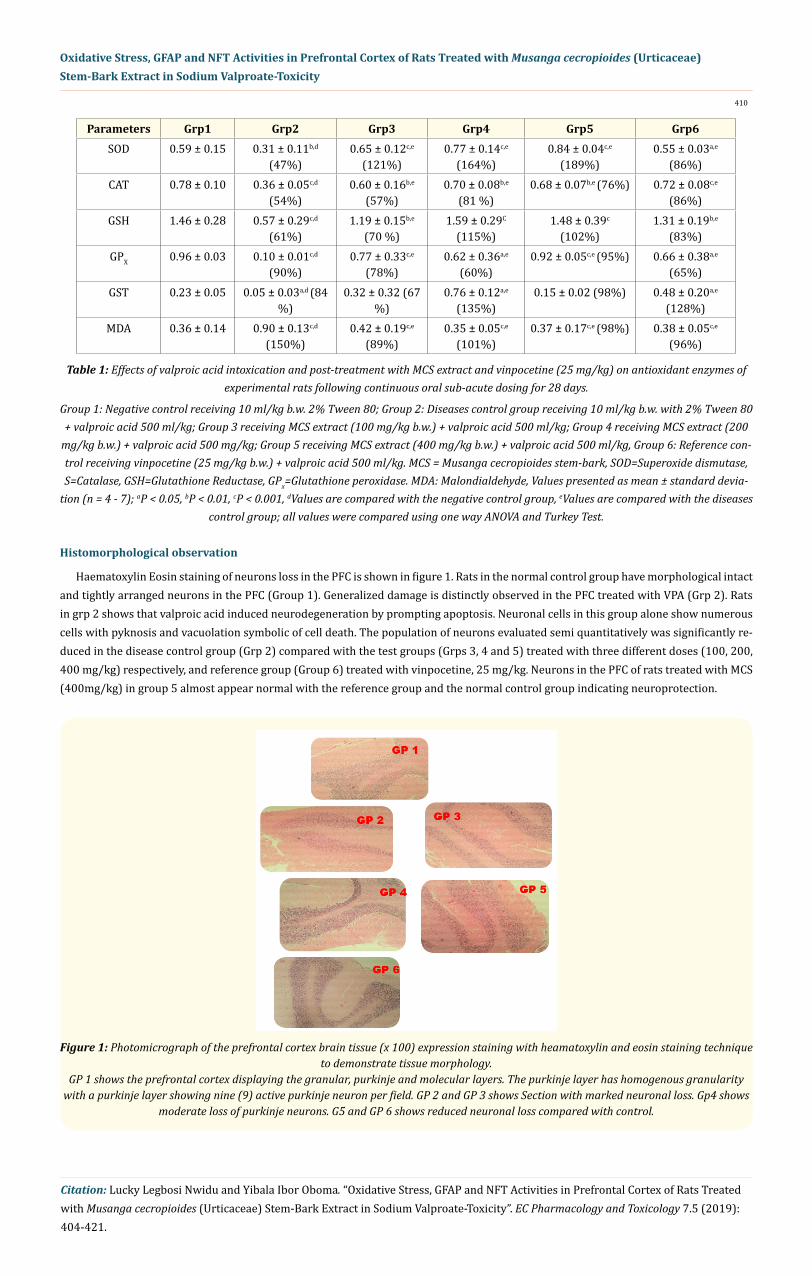
410
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
Histomorphological observation
Haematoxylin Eosin staining of neurons loss in the PFC is shown in figure 1. Rats in the normal control group have morphological intact and tightly arranged neurons in the PFC (Group 1). Generalized damage is distinctly observed in the PFC treated with VPA (Grp 2). Rats in grp 2 shows that valproic acid induced neurodegeneration by prompting apoptosis. Neuronal cells in this group alone show numerous cells with pyknosis and vacuolation symbolic of cell death. The population of neurons evaluated semi quantitatively was significantly re-duced in the disease control group (Grp 2) compared with the test groups (Grps 3, 4 and 5) treated with three different doses (100, 200, 400 mg/kg) respectively, and reference group (Group 6) treated with vinpocetine, 25 mg/kg. Neurons in the PFC of rats treated with MCS (400mg/kg) in group 5 almost appear normal with the reference group and the normal control group indicating neuroprotection.
Parameters Grp1 Grp2 Grp3 Grp4 Grp5 Grp6SOD 0.59 ± 0.15 0.31 ± 0.11b,d
(47%)0.65 ± 0.12c,e
(121%)0.77 ± 0.14c,e
(164%)0.84 ± 0.04c,e
(189%)0.55 ± 0.03a,e
(86%)CAT 0.78 ± 0.10 0.36 ± 0.05c,d
(54%)0.60 ± 0.16b,e
(57%)0.70 ± 0.08b,e
(81 %)0.68 ± 0.07b,e (76%) 0.72 ± 0.08c,e
(86%)GSH 1.46 ± 0.28 0.57 ± 0.29c,d
(61%)1.19 ± 0.15b,e
(70 %)1.59 ± 0.29C
(115%)1.48 ± 0.39c
(102%)1.31 ± 0.19b,e
(83%)GPX 0.96 ± 0.03 0.10 ± 0.01c,d
(90%)0.77 ± 0.33c,e
(78%)0.62 ± 0.36a,e
(60%)0.92 ± 0.05c,e (95%) 0.66 ± 0.38a,e
(65%)GST 0.23 ± 0.05 0.05 ± 0.03a,d (84
%)0.32 ± 0.32 (67
%)0.76 ± 0.12a,e
(135%)0.15 ± 0.02 (98%) 0.48 ± 0.20a,e
(128%)MDA 0.36 ± 0.14 0.90 ± 0.13c,d
(150%)0.42 ± 0.19c,e
(89%)0.35 ± 0.05c,e
(101%)0.37 ± 0.17c,e (98%) 0.38 ± 0.05c,e
(96%)
Table 1: Effects of valproic acid intoxication and post-treatment with MCS extract and vinpocetine (25 mg/kg) on antioxidant enzymes of experimental rats following continuous oral sub-acute dosing for 28 days.
Group 1: Negative control receiving 10 ml/kg b.w. 2% Tween 80; Group 2: Diseases control group receiving 10 ml/kg b.w. with 2% Tween 80 + valproic acid 500 ml/kg; Group 3 receiving MCS extract (100 mg/kg b.w.) + valproic acid 500 ml/kg; Group 4 receiving MCS extract (200
mg/kg b.w.) + valproic acid 500 mg/kg; Group 5 receiving MCS extract (400 mg/kg b.w.) + valproic acid 500 ml/kg, Group 6: Reference con-trol receiving vinpocetine (25 mg/kg b.w.) + valproic acid 500 ml/kg. MCS = Musanga cecropioides stem-bark, SOD=Superoxide dismutase, S=Catalase, GSH=Glutathione Reductase, GPx=Glutathione peroxidase. MDA: Malondialdehyde, Values presented as mean ± standard devia-
tion (n = 4 - 7); aP < 0.05, bP < 0.01, cP < 0.001, dValues are compared with the negative control group, eValues are compared with the diseases control group; all values were compared using one way ANOVA and Turkey Test.
Figure 1: Photomicrograph of the prefrontal cortex brain tissue (x 100) expression staining with heamatoxylin and eosin staining technique to demonstrate tissue morphology.
GP 1 shows the prefrontal cortex displaying the granular, purkinje and molecular layers. The purkinje layer has homogenous granularity with a purkinje layer showing nine (9) active purkinje neuron per field. GP 2 and GP 3 shows Section with marked neuronal loss. Gp4 shows
moderate loss of purkinje neurons. G5 and GP 6 shows reduced neuronal loss compared with control.

411
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
Histochemical Evaluation
Glial fibrillary acid protein (GFAP) immunohistochemistry
Immunohistochemistry labelling of GFAP on PFC brain tissues showing glial cells (brownish) expression in valproic acid induced neu-rodegeneration AND treated with MCS extract (Figure 2). Glial cells (astrocytes) help to maintain hemostasis and provide support and protection for neurons. A number of positive glial neurons revealing GFAP protein expression in PFC is consistent with normal histomor-phology. AS observed in control group (Grp 1). Increased protein expression of GFAP is observed in the diseases control group (Grp 2). Mild protein expression of GFAP in PFC of rats treated with 100 mg /kg-b.wt of MCS extract (Grp 3). Moderate protein expression of GFAP in PFC of rats treated with 200 mg/ kg. b.wt of extract (Grp 4). Reduced protein expression of GFAP and increase proliferation of astrocytes in PFC of rat brain treated with 400 mg/kg b.wt compared with negative control (Grp 5). Similarly, the reference control grp 6 treated with 25 mg/kg. b.wt of vinpocetine demonstrate the reduce protein expression of GFAP and astrocytes proliferation. Higher doses of extract lowered neuronal damage. The expression pattern shows the mechanism of protection by the extract which is by astrocytes proliferation.
Figure 2: Photomicrograph showing immunohistochemistry labelling of glial fibrillary acid protein (GFAP) expression (x 400). Gp1 shows photomicrograph of GFAP expression within the cerebellum. The Astrocytes processes are stained brown and are unremark-
able consistent with normal brain tissue. GP 2 section shows moderate expression of GFA protein compared to the control consistent with neuronal inflammation. Gp 3 section shows mild expression of the Glia fibrillary acid protein on the astrocytes processes. Gp4 Section shows moderate expression of the astrocytes processes. GP 5 section shows mild expression of GFAP while GP6 section shows moderate expression
of GFAP. Gp 3 and GP 5 exhibited higher neuronal protection with GP 5 competing strongly with compared to the reference drug (GP 6).
Neurofibrillary protein (NFP) immunohistochemistry
The effect of MCS ON NFP immunohistochemistry is shown in figure 3. Few NFP positive neurons were observed in the control group (Grp 1). Great numbers of NFP positive neurons were noted in the PFC of rats treated with VPA indicating generalized p53 positive neu-rons were over expressed in the disease control rat brain (Grp 2). In contrast, administration of MCS in three doses (100, 200, 400 mg/kg) progressively downregulate NFP positive neurons observed in the PFC dose dependently (Grp 3 - 5). Similarly, vinpocetine 25 mg/kg markedly reduced the number of NFP positive neurons in the PFC (Grp 6). Generally, the immunohistochemistry reveal that the extract demonstrated neuronal protection by suppression of NFP and activation of astrocytes as expressed by NFP immunostain.
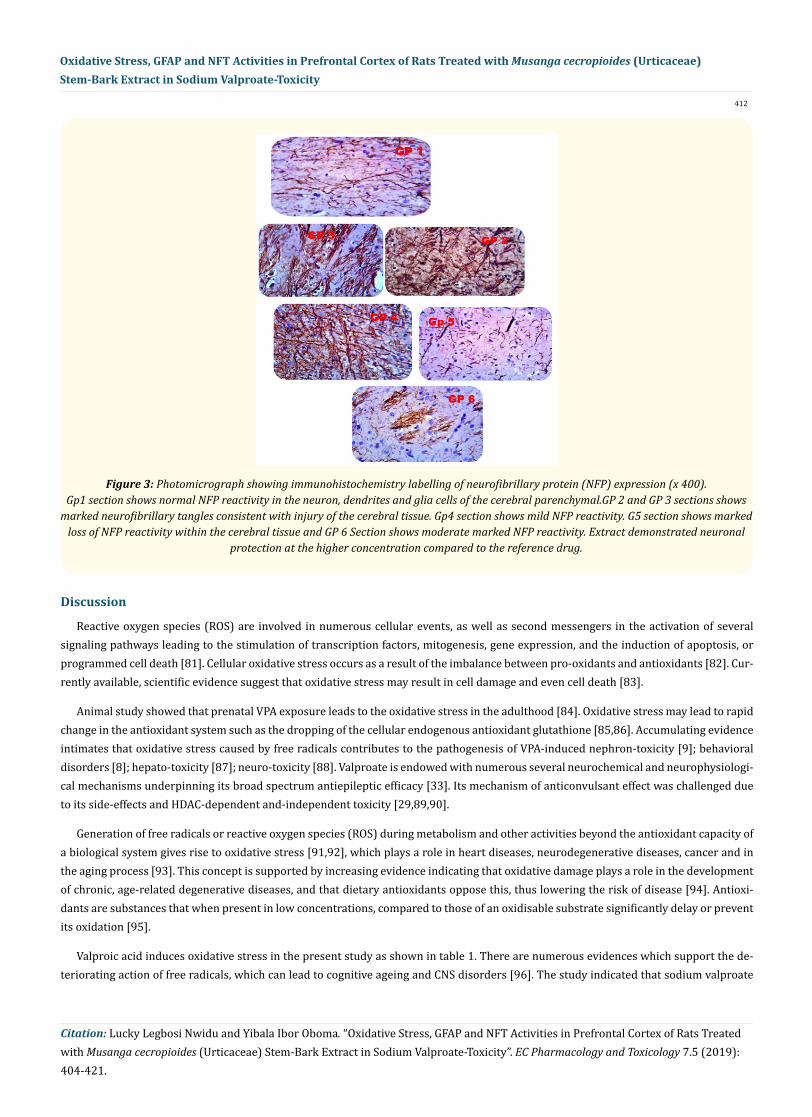
412
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
Discussion
Reactive oxygen species (ROS) are involved in numerous cellular events, as well as second messengers in the activation of several signaling pathways leading to the stimulation of transcription factors, mitogenesis, gene expression, and the induction of apoptosis, or programmed cell death [81]. Cellular oxidative stress occurs as a result of the imbalance between pro-oxidants and antioxidants [82]. Cur-rently available, scientific evidence suggest that oxidative stress may result in cell damage and even cell death [83].
Animal study showed that prenatal VPA exposure leads to the oxidative stress in the adulthood [84]. Oxidative stress may lead to rapid change in the antioxidant system such as the dropping of the cellular endogenous antioxidant glutathione [85,86]. Accumulating evidence intimates that oxidative stress caused by free radicals contributes to the pathogenesis of VPA-induced nephron-toxicity [9]; behavioral disorders [8]; hepato-toxicity [87]; neuro-toxicity [88]. Valproate is endowed with numerous several neurochemical and neurophysiologi-cal mechanisms underpinning its broad spectrum antiepileptic efficacy [33]. Its mechanism of anticonvulsant effect was challenged due to its side-effects and HDAC-dependent and-independent toxicity [29,89,90].
Generation of free radicals or reactive oxygen species (ROS) during metabolism and other activities beyond the antioxidant capacity of a biological system gives rise to oxidative stress [91,92], which plays a role in heart diseases, neurodegenerative diseases, cancer and in the aging process [93]. This concept is supported by increasing evidence indicating that oxidative damage plays a role in the development of chronic, age-related degenerative diseases, and that dietary antioxidants oppose this, thus lowering the risk of disease [94]. Antioxi-dants are substances that when present in low concentrations, compared to those of an oxidisable substrate significantly delay or prevent its oxidation [95].
Valproic acid induces oxidative stress in the present study as shown in table 1. There are numerous evidences which support the de-teriorating action of free radicals, which can lead to cognitive ageing and CNS disorders [96]. The study indicated that sodium valproate
Figure 3: Photomicrograph showing immunohistochemistry labelling of neurofibrillary protein (NFP) expression (x 400). Gp1 section shows normal NFP reactivity in the neuron, dendrites and glia cells of the cerebral parenchymal.GP 2 and GP 3 sections shows
marked neurofibrillary tangles consistent with injury of the cerebral tissue. Gp4 section shows mild NFP reactivity. G5 section shows marked loss of NFP reactivity within the cerebral tissue and GP 6 Section shows moderate marked NFP reactivity. Extract demonstrated neuronal
protection at the higher concentration compared to the reference drug.

413
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
intoxication significantly reduces the level of all the antioxidant enzymes (SOD, CAT, GSH, GPx and GST) and markedly upregulated lipid peroxidation dose dependently in experimental rats. However, the post-treatment of MCS extract at all doses upregulated all the anti-oxidant enzymes and down regulated lipid peroxidation. The percentage of VPA- induced neurotoxicity on antioxidant enzymes profile range from 47 to 84% and the lipid peroxidation was as high as 150%. However, the neuroprotection offered by MCS for the antioxidant enzymes range from 121 - 189% (SOD), 57 - 81% (CAT), 70 - 115% (GSH), 60 - 95% (GPx), 67 - 135% (GST); while the alleviation of lipid peroxidation range from 89 - 101% (MDA). Non-enzymatic serum GSH is a sensitive biomarker of the antioxidant status was markedly increased, it plays a pivotal defensive role against oxidative insults as an endogenous scavenger of free radicals. GSH is considered crucial in cellular function and viability [97]. Alteration of glutathione metabolizing enzymes, GST, GPx, and Glutathione reductase (GSSH) as part of the armamentarium of antioxidant defense system has been elucidated [98].
Similarly, lipid peroxidation was significantly (P < 0.001) evident by increased formation of MDA (150%) in the intoxicated rats. The augmentation in LPO observed in our study may be as a result of elevated ROS production or decimation in the status of antioxidants and membrane fluidity [99]. Understandably, MCS diminished oxidative stress marker LPO by scavenging ROS/ RNS and MCS extract could underpin a beneficial action against LPO, providing additional compensatory mechanism for restoring cellular integrity and protection against free radicals- induced damage [8,100]. Probably, LPO is regarded as an excellent biomarker of oxidative stress because it is the most singular largely considered product produced by free radicals [101]. It is mediated by oxygen free radicals and is thought to be sig-nificant cause of destruction and damage to cell membranes and suggested to be a causative factor to the development of VPA-mediated tissue damage [8,102]).
Moreover, enzymatic antioxidants such as SOD and CAT were depleted significantly (P < 0.01 - 0.001). Similarly, glutathione was re-duced significantly (P< 0.001). Taking together, all these exemplify exacerbation of oxidative stress that provoke impairment or damage to nervous tissues and development in the VPA exposed rats. The disequilibrium in the action of ROS /RNS production and scavenging sys-tem result in oxidative stress and concomitant brain insults. The susceptibility of brain tissues to oxidative stress has been demonstrated by Cechetti., et al [103]. Both ROS/ RNS cause neuronal damage including lipid peroxidation and optimizes the fluidity of membrane dis-rupting essential biochemical and physiological function as well as down-regulating oxidized phospholipids and cholesterol [103-105]. Therefore, the level of glutathione is depleted in oxidative stress as observed in the VPA- intoxicated rats.
Conflicting results also exist with respect to the effect of VPA on the levels of glutathione metabolizing enzymes or non-enzymatic an-tioxidants. All glutathione metabolizing enzymes may serve as antioxidants [9]. The neurotoxicity of VPA was established by a significant (P < 0.05) declined in the activity of GST in prefrontal tissue of rat brain. GST is an all-important multifunctional enzyme, which plays a role in toxic electrophiles detoxification and catalyzing their conjugation with GSH [106]. The subacute supplementation of MCS along with VPA in PFC of brain tissue of rats restore significantly (P < 0.05) the diminished GST level only at MCS 200 mg/kg and vinpocetine, 25 mg/kg. The findings of this study did not corroborate earlier published reports that the concentration of GSH is upregulated during xenobiotic intoxication [107,108], as VPA-induced significant down regulation of GSH in this investigation. However, protection against the VPA-induced neurotoxicity was achieved through repeated supplementation of antioxidant, MCS. The present study confirms that subacute exposure of MCS significantly abrogated neuronal VPA-oxidative stress induction. Besides, GPx is selenium-containing enzymes, which contribute a noticeable role in the reduction of hydrogen peroxide (H2O2) and hydroxide and elimination of excess free radicals to non-toxic products [109].
In our results, VPA treatment showed a significantly (P < 0.01) increase in activity of GPx in PFC tissues of the brain. But MCS extract administration was able to reverse significantly (P < 0.05 - 0.001) the activities of GPx in PFC cells of VPA exposed rats. The restoration might in part be due to alteration of substrate (GSH) availability [110]. Reduced glutathione (GSH) is produced from the activity of Gluta-thione reductase (GR) which is an enzyme responsible for the reduction of oxidized glutathione (GSSG) to GSH. The activity of GR directly influence the protection and reparation of sulfhydryl (-SH) protein under oxidative stress [111]. In our study, VPA treatment showed increased activity of GSH in kidney. It may be due to the protection from oxidative stress formed by over production of free radical [112]. The significant prevention in alteration in the activity of GR in PFC can possibly be attributed to its antioxidant effect. Antioxidant enzymes most times become inactive due to VPA exposure, which is due to direct binding of VPA to their active sites [113]. In the present study, the

414
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
alteration of the activities of the antioxidant enzymes probably reflect the inability of the antioxidant defense to over whelmed the influx of ROS on VPA exposure. Sub-chronic MCS extract supplementation along with VPA markedly restored the levels of antioxidant enzymes and non-enzymatic antioxidants, which might be due to the ability of MCS to diminish the accumulation of free radical generation during VPA exposure. The role of SOD as an antioxidant is to convert superoxide to hydrogen peroxide, thereby protecting against the pervasive harmful effects of superoxide.
Apart from oxidative stress, many and varied neurotoxic insults occur such as the proliferation and hypertrophy of astrocytes and the accumulation of neurofibrillary proteins (NFP). The hallmark to these responses are creation of imbalance between the endogenous an-tioxidant and oxidants moiety (upregulation of oxidants and down regulation of antioxidants); heightened expression of the major inter-mediate filament protein of astrocytes, glial fibrillary acidic protein (GFAP) and formation of aggregates of hyperphosphorylated proteins also called neurofibrillary tangles respectively. These are aetiopathological biomarkers of neurodegenerative disease mechanisms and are foci for therapeutic target [1,6,40-45,114]. This study observed VPA - induced elevation in the expression of NFT and GFAP which were diminished by subacute supplementation with antioxidant, MCS extract for 28 days. The down-regulation of VPA - mediated increased of NFT and GFAP in experimental animals by administration of MCS extract thereby in part alludes to its neuroprotective potential.
The VPA - induced astrogliosis and increased neurofibrillary protein activity in PFC tissue of experimental rat model is shown in figure 2 group 2. Astrogliosis is a prominent neuropathological feature of many diseases of the cerebral tissue [115,116]. However, evidence indicates that the role played by gliosis in pathological situations may not be restricted to its “housekeeping” function but may also in-clude actions that significantly and actively contribute to the demise of neurons, especially in neurodegenerative diseases like Parkinson disease. Interestingly, several lines of evidence demonstrate that gliosis may actually exert very different effects in the diseased brain, be-cause, depending upon the situation, it may mediate either beneficial or harmful events [117]. The gliosis in the present work was evident with neuronal loss evidences in the haematoxylin and eosin stain. Also [117,118] reported that the density of GFAP-positive astrocytes appears to be inversely related to the magnitude of dopaminergic neuronal loss across the different main dopaminergic areas of the brain in PD post mortem samples as it is obtainable in the present animal model. Furthermore [119] reported that there is neuronal expression of GFAP in patients with Alzheimer pathology in the human hippocampus. Surprisingly, the GFAPþ1 immunopositive cells observed in AD patients were primarily neurons, although GFAPþ1-expressing neurons were mainly restricted to AD and DS patients [119].
Hol., et al. [119] provide strong evidence that GFAP is expressed in neurons that are no ghost tangles, since the GFAP antibodies stain neurons in which a nucleus is visible. Furthermore, the presence of GFAP mRNA in neurons proves that the GFAP immunostaining ob-served in the neurons is indeed the result of GFAP expression [119]. In another study by Kamphuis., et al. (2014) aimed at characterizing the expression pattern of different GFAP isoforms in normal human hippocampal tissue and in conditions with AD-related gliosis, the number of GFAPþ1-expressing astrocytes significantly increases during the progress of AD with lesser effects of gender and age. The in vitro studies showed that treatment of AD rats with Ab1e42 oligomers and fibrils increases the number of GFAPþ1- positive cells, suggest-ing that elevated Ab level is a causative factor of neuronal injury [120].
Besides, neurofibrillary protein activity was higher in the disease group compared to control and treated group and this is in line with findings by Irwin., et al. [121], study that define the regional distribution of acetylated K280 immunoreactive tau pathology in Alzheimer’s disease, corticobasal degeneration and progressive supranuclear palsy. Acetylated-tau was present throughout all stages of Alzheimer’s disease pathology but was more prominently associated with pathological tau epitopes in moderate to severe-stage cases. Acetylated K280 also co-localized with N- and C-terminal specific anti-tau epitopes, indicating it is present in neurofibrillary tangles prior to subse-quent tau truncation [121].
The reference drug, vinpocetine, lowered neurofibrillary protein and glial fibrillary acid protein activity in rats exposed to valproic acid thereby reducing neuronal loss compared to the disease group. Vinpocetine has been shown to exert a brain neuroprotective effect by a combined action on cerebral circulation, brain metabolism and rheological properties of the blood. This boost in the cerebral metabolism thus enhances both oxygen and glucose utilization and consequently improving cerebral functions and protection even in conditions of hypoxia and ischemia [122]. Vinpocetine has been shown to inhibit a cyclic GMP phosphodiesterase, an inhibition which enhances cyclic GMP levels in the vascular smooth muscle, leading to reduced resistance of cerebral vessels and increase of cerebral flow [123].

415
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
Bibliography
The highest concentration of MCS extract demonstrated high neuroprotective potentials compared with the reference drug in the present animal model. The MCS extract was able to lower the tau tangles formation by preventing apoptosis of neuron. Tangles forma-tion has been reported as hallmark of neuronal degeneration and significant acetylated-tau pathology in a distribution pattern similar to hyperphosphorylated-tau [121]. Similar findings have been reported for conformational and truncation tau epitopes that are thought to represent intermediate stages of tangle progression [124]. The study also showed that MCS extract reduces GFAP expression collabo-rating with other reports that neuronal degeneration initiates glial cell regeneration by triggering various growth factors that result in increased astrocytes activity [118,120]. These results suggest that GFAP persist within degenerating astrocytes in valproic brain injury and of purkinje neurons to accentuate excitotoxic damage. An effect that was abrogated by MCS extracts supplementation. Antioxidants like MCS protect biological targets against ROS, therefore, they have been considered as attractive therapeutic potential to counteract ROS-mediated neuronal damage [125].
Conclusion
The present study confirmed that VPA - mediated oxidative stress compromised the antioxidants status of the brain PFC. Subsequently, the treatment with VPA along with MCS extract mitigated this effects. This study is evident that MCS extract could be utilized in the man-agement of VPA- induced neurotoxicity and lowered VPA - induced oxidative stress, tau formation and gliosis. The neuroprotective effect observed is greater than that of vinpocetine, a standard drug, known to decrease neuronal death thus suggesting a neuroprotective effect by the MCS extract at this concentration. However, further investigation is needed to unravel the mechanism of MCS abrogation of VPA- induced neurotoxicity.
Acknowledgement
We the authors are grateful to the following laboratory staff: Mr. Wanosike Ahamefuram, Mr. Stanley Oyegham, and Mrs. Emakkoye Ighirigi, animal house technical staff Mr. Madighiode Oghaaphemugh, and Mr. Njoku Ebere, and administrative staff Ruth Dickson who were available with technical and administrative support.
Disclosure of Conflict of Interest
No conflict of interest declared.
1. Islam MT. “Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders”. Neurology Research 39.1 (2017): 73-82.
2. Vara D and Pula G. “Reactive oxygen species: physiological roles in the regulation of vascular cells”. Current Molecular Medicine 14.9 (2014): 1103-1125.
3. Bhat AH., et al. “Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight”. Biomedicine and Pharmacotherapy 74 (2015): 101-110.
4. Jiang T., et al. “Oxidative stress: A major pathogenesis and potential therapeutic target of antioxidative agents in Parkinson’s disease and Alzheimer’s disease”. Progress in Neurobiology 147 (2016): 1-19.
5. Tönnies E and Trushina E. “Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s disease”. Journal of Alzheimer’s Disease 57.4 (2017): 1105-1121.
6. Salim S. “Oxidative Stress and the Central Nervous System”. Journal of Pharmacology and Experimental Therapeutics 360.1 (2017): 201-205.
7. Liu Z., et al. “Oxidative Stress in Neurodegenerative Diseases: From Molecular Mechanisms to Clinical Applications”. Oxidative Medi-cine and Cell Longevity (2017): 2525967.
8. Al-Amin MM., et al. “Astaxanthin improves behavioral disorder and oxidative stress in prenatal valproic acid-induced mice model of autism”. Behaviour and Brain Research 286 (2015): 112-121.

416
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
9. Chaudhary S., et al. “Nephroprotective activities of quercetin with potential relevance to oxidative stress induced by valproic acid”. Protoplasma 252.1 (2015): 209-217.
10. Verrotti A., et al. “Valproic Acid Metabolism and its Consequences on Sexual Functions”. Current Drug Metabolism 17.6 (2016): 573-581.
11. Nevitt SJ., et al. “Antiepileptic drug monotherapy for epilepsy: a network meta-analysis of individual participant data”. Cochrane Da-tabase of Systematic Reviews 6 (2017): CD011412.
12. Tseng JH., et al. “Valproic acid inhibits glioblastoma multiforme cell growth via paraoxonase 2 expression”. Oncotarget 8.9 (2017): 14666-14679.
13. Umka J., et al. “Valproic acid reduces spatial working memory and cell proliferation in the hippocampus”. Neuroscience 166.1 (2010): 15-22.
14. Chen S., et al. “Valproic acid: a new candidate of therapeutic application for the acute central nervous system injuries”. Neurochemical Research 39.9 (2014): 1621-1633.
15. Carriere CH., et al. “Neuroprotection by valproic acid in an intrastriatal rotenone model of Parkinson’s disease”. Neuroscience 267 (2014): 114-121.
16. Zhang C., et al. “Neuroprotective and anti-apoptotic effects of valproic acid on adult rat cerebral cortex through ERK and Akt signaling pathway at acute phase of traumatic brain injury”. Brain Research 1555 (2014): 1-9.
17. Tung EW and Winn LM. “Valproic acid increases formation of reactive oxygen species and induces apoptosis in post-implantation embryos: a role for oxidative stress in valproic acid-induced neural tube defects”. Molecular Pharmacology 80.6 (2011): 979-987.
18. Fujiki R., et al. “A proapoptotic effect of valproic acid on progenitors of embryonic stem cell-derived glutamatergic neurons”. Cell Death Disease 4.6 (2013): e677.
19. Arvidsson Y., et al. “Cytotoxic Effects of Valproic Acid on Neuroendocrine Tumour Cells”. Neuroendocrinology 103.5 (2016): 578-591.
20. Gilardini Montani MS., et al. “Histone deacetylase inhibitors VPA and TSA induce apoptosis and autophagy in pancreatic cancer cells”. Cell Oncology (Dordr) 40.2 (2017): 167-180.
21. Takuma K., et al. “Chronic treatment with valproic acid or sodium butyrate attenuates novel object recognition deficits and hippocam-pal dendritic spine loss in a mouse model of autism”. Pharmacology Biochemistry and Behaviour 126 (2014): 43-49.
22. Bittigau P., et al. “Antiepileptic drugs and apoptosis in the developing brain”. Annals of the New York Academy of Sciences 993 (2003): 103-114.
23. Yochum CL., et al. “VPA-induced apoptosis and behavioral deficits in neonatal mice”. Brain research 1203 (2008): 126-132.
24. McAuley JW and Anderson GD. “Treatment of epilepsy in women of reproductive age: pharmacokinetic considerations”. Clinical Phar-macokinetics 41.8 (2002): 559-579.
25. Giavini E and Menegola E. “Teratogenic activity of HDAC inhibitors”. Current Pharmaceutical Design 20.34 (2014): 5438-5442.
26. Christensen J., et al. “Prenatal valproate exposure and risk of autism spectrum disorders and childhood autism”. JAMA: The Journal of the American Medical Association 309.16 (2013): 1696-1703.
27. Konopko MA., et al. “Sexually Dimorphic Epigenetic Regulation of Brain-Derived Neurotrophic Factor in Fetal Brain in the Valproic Acid Model of Autism Spectrum Disorder”. Developmental Neuroscience 39.6 (2017): 507-518.
28. Chaudhary S and Parvez S. “Valproic Acid Induced Neurotoxicological Manifestations and its Mitigation by Melatonin in Rat Brain Synaptosomes”. Archives of Medical Research 49.7 (2019): 441-450.
29. Han BR., et al. “Valproic acid inhibits the growth of HeLa cervical cancer cells via caspase-dependent apoptosis”. Oncology Reports 30.6 (2013): 2999-3005.

417
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
30. Yochum CL., et al. “Animal model of autism using GSTM1 knockout mice and early post-natal sodium valproate treatment”. Behaviour Brain Research 210.2 (2010): 202-210.
31. Wang C., et al. “Valproic acid induces apoptosis in differentiating hippocampal neurons by the release of tumor necrosis factor-α from activated astrocytes”. Neuroscience Letter 497.2 (2011): 122-127.
32. Gao J., et al. “Neuroprotective effects of docosahexaenoic acid on hippocampal cell death and learning and memory impairments in a valproic acid-induced rat autism model”. International Journal of Development in Neuroscience 49 (2016): 67-78.
33. Johannessen CU and Johannessen SI. “Valproate: past, present, and future”. CNS Drug Review 9.2 (2003): 199-216.
34. Forgione N and Tropepe V. “Histone deacetylase inhibition promotes Caspase-independent cell death of ventral midbrain neurons”. Molecular and Cellular Neurosciences 48.2 (2011): 117-128.
35. Bollino D., et al. “Valproic acid induces neuronal cell death through a novel calpain-dependent necroptosis pathway”. Journal of Neu-rochemistry 133.2 (2015): 174-186.
36. Cheng SY., et al. “Regulatory role of calpain in neuronal death”. Neural Regenerative Research 13.3 (2018): 556-562.
37. Danial NN and Korsmeyer SJ. “Cell death: critical control points”. Cell 116.2 (2004): 205-219.
38. Storr SJ., et al. “The calpain system and cancer”. Nature reviews Cancer 11.5 (2011): 364-374.
39. Pasupuleti N., et al. “5-Benzylglycinyl-amiloride kills proliferating and nonproliferating malignant glioma cells through caspase-in-dependent necroptosis mediated by apoptosis-inducing factor”. The Journal of Pharmacology and Experimental Therapeutics 344.3 (2013): 600-615.
40. Hol EM and Pekny M. “Glial fibrillary acidic protein (GFAP) and the astrocyte intermediate filament system in diseases of the central nervous system”. Current Opinion in Cell Biology 32 (2015): 121-130.
41. Yang Z and Wang KK. “Glial fibrillary acidic protein: from intermediate filament assembly and gliosis to neurobiomarker”. Trends in Neuroscience 38.6 (2015): 364-374.
42. Finsterwald C., et al. “Astrocytes: New Targets for the Treatment of Neurodegenerative Diseases”. Current Pharm Design 21.25 (2015): 3570-3581.
43. Singh S and Joshi N. “Astrocytes: inexplicable cells in neurodegeneration”. International Journal of Neuroscience 127.3 (2017): 204-209.
44. Pekny M., et al. “The dual role of astrocyte activation and reactive gliosis”. Neuroscience Letters 565 (2014): 30-38.
45. Ahmad MH., et al. “Influence of microglia and astrocyte activation in the neuro-inflammatory pathogenesis of Alzheimer’s disease: Rational insights for the therapeutic approaches”. Journal of Clinical Neuroscience 59 (2019): 6-11.
46. Hirano A. “Neurofibrillary changes in conditions related to Alzheimer’s disease”. Alzheimer’s Disease And Related Conditions. Ed. Wolstenholme, G., and O’Connor, M. London, J and A Churchill Ltd. 1 (1970): 185-207.
47. Bereczki D and Fekete I. “Vinpocetine for acute ischemic stroke”. Cochrane Database of Systematic Reviews 1 (2008): CD000580.
48. Szatmári S and Whitehouse P. “Vinpocetine for cognitive impairment and dementia”. Cochrane Database of Systematic Reviews 1 (2003): CD003119.
49. Feigin VL., et al. “Vinpocetine treatment in acute ischaemic stroke: a pilot single-blind randomized clinical trial”. European Journal Neurology 8.1 (2001): 81-85.
50. Truss MC., et al. “Initial clinical experience with the selective phosphodiesterase-I isoenzyme inhibitor vinpocetine in the treatment of urge incontinence and low compliance bladder”. World Journal of Urology 18.6 (2000): 439-443.

418
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
51. Thorne., et al. “Monograph: vinpocetine”. Alternative Medical Review 7.3 (2002): 240-243.
52. Sitges M., et al. “Effect of the anti-depressant sertraline, the novel anti-seizure drug vinpocetine and several conventional antiepilep-tic drugs on the epileptiform EEG activity induced by 4-aminopyridine”. Neurochemistry Research 41.6 (2016): 1365-1374.
53. Nwidu LL and Teme RE. “Sub-Chronic Toxicity of Hydromethanolic Stem Bark Extract of Musanga cecropioides (Urticaceae) in Rat”. EC Pharmacology and Toxicology 6.3 (2018): 76-95.
54. Nyemb N., et al. “Anti hyperglycemic and Antioxidant Potential of Water-Ethanol Extract of Musanga cecropioides Stem Bark”. Interna-tional Journal of Pharmaceutical Sciences and Drug Research 8.1 (2016): 43-49.
55. Nwidu LL., et al. “Hepatoprotective effect of hydromethanolic leaf extract of Musanga cecropioides (Urticaceae) on carbon tetrachlo-ride-induced liver injury and oxidative stress”. Journal of Taibah University Medical Science 13.4 (2018): 344-354.
56. Abimbola S., et al. “Musanga cecropioides leaf extract exhibits anti-inflammatory and anti-nociceptive activities in animal models”. Revista Brasileira de Farmacognosia 25.5 (2015): 506-512.
57. Adeneye A., et al. “The hypotensive mechanisms for the aqueous stem bark extract of Musanga cecropioides in Sprague-Dawley rats”. Journal of Ethnopharmacology 106.2 (2006): 203-207.
58. Trease GE and Evans WC. “A Textbook of Pharmacognosy, 13th edition”. Bailliere Tindall Ltd., London (1989): 89-100.
59. Wagner GC., et al. “A new neuro-behavioural model of autism in mice: pre-and postnatal exposure to sodium valproate”. Journal of Autism Developmental Disorders 36.6 (2006): 779-793.
60. Cohen OS., et al. “Acute prenatal expo-sure to a moderate dose of valproic acid increases social behaviour and alters gene expression in rats”. International Journal of Development in Neuroscience 31.8 (2013): 740-750.
61. Banji D., et al. “Amelioration of behavioural aberrations and oxidative markers by green tea extract in valproate-induced autism in animals”. Brain Research 1410 (2011): 141-151.
62. Bambini-Junior V., et al. “Ani-mal model of autism induced by prenatal exposure to valproate: behavioural changes and liver param-eters”. Brain Research 1408 (2011): 8-16.
63. Schneider T and Przewlocki R. “Behavioural alterations in rats prenatally exposed to valproic acid: animal model of autism”. Neuro-psychopharmacology 30.1 (2005): 80-89.
64. Schneider T., et al. “Gender-specific behavioural and immunological alterations in an animal model of autism induced by prenatal exposure to valproic acid”. Psychoneuroendocrinology 33.6 (2008): 728-740.
65. Sandhya T., et al. “Bacopa monniera (L.) Wettst ameliorates behavioural alterations and oxidative markers in sodium valproate in-duced autism in rats”. Neurochemistry Research 37.5 (2012): 1121-1131.
66. Kerr DM., et al. “Alterations in the endo-cannabinoid system in the rat valproic acid model of autism”. Behaviour Brain Research 249 (2013): 124-132.
67. Kolozsi E., et al. “Prenatal exposure to valproic acid leads to reduced expression of synaptic adhesion molecule neuroligin 3 in mice”. Neuroscience 163.4 (2009): 1201-1210.
68. Niaraki MS., et al. “Protective effect of ghrelin on sodium valproate-induced liver injury in rat”. Journal of Stress Physiology and Bio-chemistry 9 (2013): 97-105.
69. Cholnoky E and Dömök LI. “Summary of safety tests of ethyl apovincaminate”. Arzneimittel Forschung 26.10 (1976): 1938-1944.
70. Vereczkey L., et al. “Kinetic metabolism of vinpocetine in the rat”. Arzneimittel Forschung 29.6 (1979): 953-956.
71. Ohkawa H., et al. “Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction”. Analytical Biochemistry 95.2 (1979): 351-358.

419
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
72. Todorova I., et al. “Reference values of oxidative stress parameters (MDA, SOD, CAT) in dogs and cats”. Comprehensive Clinical Pathol-ogy 13.4 (2005): 190 -194.
73. Sedlak J and Lindsay RH. “Estimation of total, protein-bound, and non-protein sulfhydryl groups in tissue with Ellman’s reagent”. Anal of Biochemistry 25.1 (1968): 192 -205.
74. Rotruck JT., et al. “Selenium: biochemical role as a component of glutathione peroxidase”. Science 179.4073 (1975): 588-590.
75. Claiborne A. “Catalase activity”. In: Greenwald RA, editor. Handbook of methods for oxygen radical research. Florida: CRC Press (1995): 237-242.
76. Wilson CB., et al. “Inflammation and Oxidative Stress Are Elevated in the Brain, Blood, and Adrenal Glands during the Progression of Post-Traumatic Stress Disorder in a Predator Exposure Animal Model”. PLoS One 8.10 (2013): e76146.
77. Habig WH., et al. “Glutathione S-transferases. The first enzymatic step in mercapturic acid formation”. Journal of Biological Chemistry 249.22 (1974): 7130 -7139.
78. Lowry OH., et al. “Protein measurement with the folin phenol”. Journal of Biological Chemistry 193.1 (1951): 265-275.
79. Oyinbo CA., et al. “Landolphia owariensis Attenuates Alcohol-induced Cerebellar Neurodegeneration: Significance of Neurofilament Protein Alteration in the Purkinje Cells”. Folia Medicine (Plovdiv) 58.4 (2016): 241-249.
80. Oboma YI., et al. “Breast cancer biomarkers at Niger delta university hospital: Comparisons with national and international trends and clinical significance”. Pathophysiology 24.3 (2017): 191-196.
81. Birben E., et al. “Oxidative stress and antioxidant defense”. World Allergy Organisation Journal 5.1 (2012): 9-19.
82. Sung CC., et al. “Oxidative stress and nucleic acid oxidation in patients with chronic kidney disease”. Oxidative Medicine and Cellular Longevity (2013): 301982.
83. Kern JK and Jones AM. “Evidence of toxicity, oxidative stress, and neuronal insult in autism”. Journal Toxicology and Environmental Health 9.6 (2006): 485-499.
84. Pragnya B., et al. “Ameliorating effect of piperine on behavioural abnormalities and oxidative markers in sodium valproate induced autism in BALB/C mice”. Behaviour Brain Research 270 (2014): 86-94.
85. Chauhan A and Chauhan V. “Oxidative stress in autism”. Pathophysiology 13.3 (2006): 171-181.
86. Rose S., et al. “Evidence of oxidative damage and inflammation associated with low glutathione redox status in the autism brain”. Translational Psychiatry 2 (2012): e134.
87. Ahangar N., et al. “Zinc Deficiency and Oxidative Stress Involved in Valproic Acid Induced Hepatotoxicity: Protection by Zinc and Sele-nium Supplementation”. Biology of Trace Element Research 179.1 (2017): 102-109.
88. Zhang Y., et al. “Sulindac attenuates valproic acid-induced oxidative stress levels in primary cultured cortical neurons and ameliorates repetitive/stereotypic-like movement disorders in Wistar rats prenatally exposed to valproic acid”. International Journal Molecular Medicine 35.1 (2015): 263-270.
89. Chateauvieux S., et al. “Molecular and Therapeutic Potential and Toxicity of Valproic Acid”. Journal Biomedicine and Biotechnology (2010): 479364.
90. Shah RD., et al. “Sodium valproate potentiates staurosporine-induced apoptosis in neuroblastoma cells via Akt/survivin indepen-dently of HDAC inhibition”. Journal of Cellular Biochemistry 114.4 (2013): 854-863.
91. Zheng W and Wang SY. “Antioxidant activity and phenolic compounds in selected herbs”. Journal of Agricultural and Food Chemistry 49.11 (2001): 5165-5170.
92. Tepe B., et al. “Screening of the antioxidant potentials of six Salvia species from Turkey”. Food Chemistry 95.2 (2006): 200-204.

420
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
93. Astley S B. “Dietary antioxidants past, present and future”. Trends Food Science Technology 14.3 (2003): 93-98.
94. Atoui AK., et al. “Tea and herbal infusions: their antioxidant activity and phenolic profile”. Food Chemistry 89.1 (2005): 27-36.
95. Halliwell B and Gutteridge JMC. Free Radicals in Biology and Medicine, 2nd edition; Clarendon Press: Oxford, UK (1989).
96. Golden TR., et al. “Oxidative stress and aging: beyond correlation”. Aging Cell 1.2 (2002): 117-123.
97. Nabavi SM., et al. “Ameliorative effects of quercetin on sodium fluoride-induced oxidative stress in rat’s kidney”. Renal Failure 34.7 (2012): 901-906.
98. Zhu Y., et al. “Age-related changes in glutathione and glutathione-related enzymes in rat brain”. Brain Research 1090.1 (2006): 35-44.
99. Paul MV., et al. “Protective effects of α-tocopherol against oxidative stress related to nephrotoxicity by monosodium glutamate in rats”. Toxicological Mechanism and Methods 22.8 (2012): 625-630.
100. Renugadevi J and Prabu SM. “Quercetin protects against oxidative stress-related renal dysfunction by cadmium in rats”. Experimental and Toxicologic Pathology 62.5 (2010): 471-481.
101. Messaoudi I., et al. “Protective effects of selenium, zinc, or their combination on cadmium-induced oxidative stress in rat kidney”. Biol-ogy Trace Elements Research 130.2 (2009): 152-161.
102. Chaudhary S and Parvez S. “An in vitro approach to assess the neurotoxicity of valproic acid-induced oxidative stress in cerebellum and cerebral cortex of young rats”. Neuroscience 225 (2012): 258-268.
103. Cechetti F. et al. “Environmental enrichment prevents behavioural deficits and oxidative stress caused by chronic cerebral hypoperfu-sion in the rat”. Life Science 91.1-2 (2012): 29-36.
104. Yazar E., et al. “Effects of drugs used in endotoxic shock on oxidative stress and organ damage markers”. Free Radical Research 44.4 (2010): 397-402.
105. Prabhu HR and Nandini M. “Inhibition of selenium dependent glutathione peroxidase and superoxide dismutase in rats by diethyldi-thiocarbamate: effect of pre-administration of alpha-tocopherol”. Indian Journal of Experimental Biology 45.5 (2007): 465-468.
106. Prabu SM., et al. “Amelioration of cadmium-induced oxidative stress, impairment in lipids and plasma lipoproteins by the combined treatment with quercetin and α-tocopherol in rats”. Journal of Food Science 75.7 (2010): T132-T140.
107. Zafeer MF. “Cadmium -induced hepatotoxicity and its abrogation by thymoquinone”. Journal of Biochemical and Molecular Toxicology 26.5 (2012): 199-205.
108. Waseem M., et al. “Mitochondria-mediated mitigatory role of curcumin in cisplatin-induced nephrotoxicity”. Cell Biochemistry and Function 31.8 (2013): 678-684.
109. Abdel-Raheem IT., et al. “Protective effect of quercetin against gentamicin-induced nephrotoxicity in rats”. Biological and Pharmaceu-tical Bulletin 32.1 (2009): 61-67.
110. Coskun O., et al. “Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and beta-cell dam-age in rat pancreas”. Pharmacological Research 51.2 (2005): 117-123.
111. Tabassum H., et al. “Nephrotoxicity and its prevention by taurine in tamoxifen induced oxidative stress in mice”. Human and Experi-mental Toxicology 26.6 (2007): 509-518.
112. Park HK., et al. “Protective effect of quercetin against paraquat-induced lung injury in rats”. Life Science 87.5-6 (2010): 181-186.
113. Kiang TK., et al. “Glutathione depletion by valproic acid in sandwich cultured rat hepatocytes: role of biotransformation and temporal relationship with onset of toxicity”. Toxicology and Applied Pharmacology 252.3 (2011): 318-324.
114. Colangelo AM., et al. “Astrogliosis as a therapeutic target for neurodegenerative diseases”. Neuroscience Letters 565 (2014): 59-64.

421
Citation: Lucky Legbosi Nwidu and Yibala Ibor Oboma. “Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity”. EC Pharmacology and Toxicology 7.5 (2019): 404-421.
Oxidative Stress, GFAP and NFT Activities in Prefrontal Cortex of Rats Treated with Musanga cecropioides (Urticaceae) Stem-Bark Extract in Sodium Valproate-Toxicity
115. Guillamón-Vivancos T., et al. “Astrocytes in neurodegenerative diseases (I): function and molecular description”. Neurologia 30.2 (2015): 119-129.
116. Osborn LM., et al. “Astrogliosis: An integral player in the pathogenesis of Alzheimer’s disease”. Progress in Neurobiology 144 (2016): 121-141.
117. Duet al. “Glial cell response: A pathogenic factor in Parkinson’s disease”. Journal of NeuroVirology 8.6 (2002): 551-558.
118. Damier P., et al. “Glutathione peroxidase, glial cells and Parkinson’s disease”. Neuroscience 52.1 (1993): 1-6.
119. Hol E., et al. “Neuronal expression of GFAP in patients with Alzheimer pathology and identification of novel GFAP splice forms”. Mo-lecular Psychiatry 8.9 (2003): 786-796.
120. Kamphuis W., et al. “Glial fibrillary acidic protein isoform expression in plaque related astrogliosis in Alzheimer’s disease”. Neurobiol-ogy of Aging 35.3 (2014): 492-510.
121. Irwin DJ., et al. “Acetylated tau, a novel pathological signature in Alzheimer’s disease and other tauopathies”. Brain 135.3 (2012): 807-818.
122. Nicholson CD. “Pharmacology of nootropics and metabolically active compounds in relation to their use in dementia”. Psychopharmacology (Berlin) 101.2 (1990): 147-159.
123. Preeti K and Vijay J. “Vinpocetine: a step towards memory enhancement”. International Journal of Pharma Research and Development 2.13 (2011): 99-108.
124. Guillozet-Bongaarts AL., et al. “Tau truncation during neurofibrillary tangle evolution in Alzheimer’s disease”. Neurobiology of Aging 26.7 (2005): 1015-1022.
125. Carvalho AN., et al. “Oxidative Stress and Antioxidants in Neurological Diseases: Is There Still Hope?” Current Drug Targets 18.6 (2017): 705-718.
Volume 7 Issue 5 May 2019©All rights reserved by Lucky Legbosi Nwidu and Yibala Ibor Oboma.