opisthobranchia: a taxonomic and biological review … karlsson exjobb ifm... · undergraduate...
TRANSCRIPT

Undergraduate thesis in biology
Opisthobranchia: A taxonomic and biological review with emphasis on the families Chromodorididae and
Phyllidiidae together with field notes from South East Sulawesi, Indonesia
Lars Karlsson
LiU-IFM-Biol-Ex-892

Avdelning, Institution Division, Department
Avdelningen för biologi Institutionen för Fysik och mätteknik
Datum Date 010330
Språk Language
Rapporttyp Report category
ISBN LiU-IFM-Biol-Ex-892
Engelska/English x Examensarbete x D-uppsats
ISRN LiU-Biol-Ex-350
Handledare: Per Milberg Institution: Biology Department, IFM, University of Linköping
URL för elektronisk version
Titel Title: Opisthobranchia: A taxonomic and biological review with emphasis on the families Chromodorididae and Phyllidiidae together with field notes from South East Sulawesi, Indonesia Författare Author: Lars Karlsson
Sammanfattning Abstract This study gives an overview regarding Opisthobranchia (Mollusca, Gastropoda) and its taxonomy and some important biological facts. It also contains two parts of information regarding fieldwork conducted in Indonesia (Sulawesi) in cooperation with “Operation Wallacea” (a joint venture between a British company and an Indonesian research institute). The first of the two parts regarding fieldwork, gives a checklist of “Opisthobranchs of the Wakatobi Marine National Park” containing a total of 297 identified or suspected (provisionally identified) species (1997-1999). In the survey season 1999: 3523 opisthobranch observations were recorded, belonging to 235 species (identified or suspected) and 73 identified genera. During the whole survey 60 % of the opisthobranchs were identified to species level. The largest order was the Doridina, both in species number and number of genera. Fourteen percent of the opisthobranch species found in 1999 made up seventy-five percent of the total number of observations recorded. The second of the two parts regarding fieldwork is a description and analysis of spatial distribution of nudibranchs according to benthic structure (especially the families: Chromodorididae and Phyllidiidae since they are the most abundant). This survey was conducted at two sites, at two depths (around 3 and 12 m) and at two times a day (“am/pm”). During this survey 252 nudibranchs belonging to at least 19 species were recorded. According to a Detrended Correpondance Analysis, species composition seems to vary with benthic forms and depth.
Nyckelord Keywords: Opisthobranchia, Nudibranchia, Chromodorididae, Phyllidiidae, Operation Wallacea, Sulawesi, opisthobranch surveys, spatial distribution

1
Contents Contents 1 Abstract 2 1 Review of opisthobranch taxonomy & biology 3 1.1 Introduction 3 1.2 Taxonomy of Gastropoda/Opisthobranchia 3 1.3 Morphology, ecology and physiology in Opisthobranchia 6 1.4 Nudibranchia 11 1.5 Chromodorididae 15 1.6 Phyllidiidae 18 1.7 References 18 2 Checklist of opisthobranchs of the Wakatobi National Park, SE Sulawesi, Indonesia 21 2.1 Introduction 21 2.2 Methods 21 2.3 Results 23 2.4 Discussion 26 2.5 References 28 3 Spatial distribution of nudibranchs according to benthic structure 29 3.1 Introduction 29 3.2 Material and methods 29 3.3 Results 31 3.4 Discussion 35 3.5 References 36 4 Conclusions and recommendations 37 5 Acknowledgements 38 Plates A B Appendices I II

2
Opisthobranchia: A taxonomic and biological review with emphasis on the families Chromodorididae and Phyllidiidae together with field
notes from South East Sulawesi, Indonesia
Lars Karlsson, Biology Department, IFM, University of Linköping, 2001.
Abstract
This study gives an overview regarding Opisthobranchia (Mollusca, Gastropoda) and its taxonomy and some important biological facts. It also contains two parts of information regarding fieldwork conducted in Indonesia (Sulawesi) in cooperation with “Operation Wallacea” (a joint venture between a British company and an Indonesian research institute). The first of the two parts regarding fieldwork, gives a checklist of “Opisthobranchs of the Wakatobi Marine National Park” containing a total of 297 identified or suspected (provisionally identified) species (1997-1999). In the survey season 1999: 3523 opisthobranch observations were recorded, belonging to 235 species (identified or suspected) and 73 identified genera. During the whole survey 60 % of the opisthobranchs were identified to species level. The largest order was the Doridina, both in species number and number of genera. Fourteen percent of the opisthobranch species found in 1999 made up seventy-five percent of the total number of observations recorded. The second of the two parts regarding fieldwork is a description and analysis of spatial distribution of nudibranchs according to benthic structure (especially the families: Chromodorididae and Phyllidiidae since they are the most abundant). This survey was conducted at two sites, at two depths (around 3 and 12 m) and at two times a day (“am/pm”). During this survey 252 nudibranchs belonging to at least 19 species were recorded. According to a Detrended Correpondance Analysis, species composition seems to vary with benthic forms and depth.

3
1 Review of opisthobranch taxonomy & biology 1.1 Introduction Opisthobranchs or “sea slugs” are a large, exclusively (except for the freshwater Ancylodoris baicalensis) marine group within the Gastropoda (Wägele & Willan, 2000). They are often colourful and display several forms of crypsis, special resemblance, aposematic coloration and mimicry (Gosliner & Behrens, 1990; Tullrot, 1998). Most opisthobranchs lack the protection of a shell as adults (some of them do have a shell but it is not functioning as a protective device since it is reduced and sometimes also internal). Instead these organisms rely on other defences. Some opisthobranchs use spicules as an alternative to shell, others are cryptic, some utilize the nematocysts from their prey and a few use autotomy. One common defensive systems, though, is chemical defence (Faulkner & Ireland, 1977; Todd, 1981; Faulkner & Ghiselin, 1983; Gunthorpe & Cameron, 1987; Avila, 1995; Cimino et al., 1999). This is the case for both Chromodorididae and Phyllidiidae (Nudibranchia). Both groups prey on sponges, which often contain bad tasting or even toxic compounds. The opisthobranchs then accumulate (or convert) some of these compounds and thereby become less interesting for potential predators. This habit has made them interesting for pharmaceutical screening as many of the compounds they use are suspected to have pharmaceutical value (Bohlin, 1989). Another interesting feature is that the chemical toxicity of these species often is coupled with strong examples of aposematism. Often, the same type of strong warning coloration is used by many different opisthobranchs. If the opisthobranchs with the similar coloration are all unpalatable, then it is an example of Müllerian mimicry. On the other hand, if there are palatable species among the ones with similar coloration, then it is an example of Batesian mimicry (Ruppert & Barnes, 1994). One interesting discovery is that there is also a flatworm species, which seems to mimic a phyllidiid nudibranch in order to avoid predation (Newman et al., 1994). 1.2 Taxonomy of Gastropoda/Opisthobranchia There are many different and confusing views on Gastropod systematics. In most publications, the class Gastropoda consists of three subclasses: Prosobranchia, Pulmonata and Opisthobranchia (Ruppert & Barnes, 1994; Hayward & Ryland, 1995; Thompson, 1976). This class is not considered by some authors to reflect the phylogeny of gastropods and a newer (cladistic) classification divides the Gastropoda into two clades: Eogastropoda and Orthogastropoda. In the latter group is a clade called Apogastropoda, which contain Heterobranchia and Caenogastropoda. The former group, in turn, consists of three clades: Heterostropha, Opisthobranchia and Pulmonata (Ponder & Lindberg, 1997). Most authors agree that within the Opisthobranchia there are at least eight groups, although they have been given different taxonomic ranks. These subgroups within the Opisthobranchia are (Marshall & Willan 1999): Cephalaspidea (Bubble shells), Anaspidea (Sea hares), Notaspidea (Side-gilled slugs), Acochlidiacea (Caddis-slugs), Sacoglossa (Sap-sucking sea slugs), Thecosomata (Sea butterflies), Gymnosomata (Naked sea butterflies) and Nudibranchia (Nudibranchs). Sometimes other groups are included in the Opisthobranchia, for example Pyramidellacea (Ruppert & Barnes, 1994) Pyramidellidae (Hayward & Ryland, 1995), Entomotaeniata (Willan & Coleman, 1984), Rhodopida (Hansson, 1998) and Rhodopemorpha (Beesley et al., 1998). Many of these groups have previously been assigned different names and some of these synonyms are shown in Table 1 together with examples of different views regarding levels of taxa.

4
Table 1. Systematic review of Opisthobranchia according to different authors. Marshall & Willan,
1999 Rudman & Willan, 1998
Hansson, 1998 Hayward & Ryland, 1995
Ruppert & Barnes,
(In Beesley et al., 1998)
1994 *
Regnum Animalia Animalia Phylum Mollusca Mollusca Mollusca Subphylum Conchifera Superclassis Classis Gastropoda Gastropoda Gastropoda Gastropoda Gastropoda Subclassis Opisthobranchia Opisthobranchia Heterobranchia Opisthobranchia Opisthobranchia Infraclassis Euthyneura Superordo Opisthobranchia Ordo Cephalaspidea Cephalaspidea = Cephalaspid(e)a = Cephalaspidea Cephalaspidea Bullomorpha Bullomorpha = Tectibranchida Ordo Anaspidea Anaspidea = Anaspid(e)a = Anaspidea Anaspidea = Aplysiomorpha Aplysiomorphi(id)a Aplysiacea Ordo Notaspidea Notaspidea = Notaspidea Notaspidea Pleurobranchomorpha Ordo Acochlidia Acochlidea = Acochlidiomorph(id)a = Acochlidiacea Acochlidioidea Acochlidia Acochlidioid(e)a Ordo Sacoglossa Sacoglossa = Sacogloss(id)a = Sacoglossa Sacoglossa Ascoglossa = Ascogloss(id)a Monostichoglossa = Pellibranchiata Ordo Thecosomata Thecosomata Thecosomat(id)a Thecosomata Thecosomata Ordo Gymnosomata Gymnosomata Gymnosomat(id)a Gymnosomata Gymnosomata Ordo Rhodopemorpha Rhodopida Pyramidellidae Pyramidellacea Ordo Nudibranchia Nudibranchia = Nudibranchi(d)a Nudibranchia Nudibranchia Behrens, 1991 and Thompson, 1988 ** Willan & Coleman, Thompson, 1976 Gosliner, 1987 1984 Regnum Phylum Mollusca Subphylum Superclassis Classis Gastropoda Gastropoda Gastropoda Subclassis Opisthobranchia Opisthobranchia Opisthobranchia Opisthobranchia Infraclassis Superordo Ordo Cephalaspidea = Bullomorpha Cephalaspidea Bullomorpha Bullomorpha Ordo Anaspidea = Aplysiomorpha Anaspidea Aplysiomorpha Aplysiomorpha Ordo Notaspidea = Pleurobranchomorpha Notaspidea Pleurobranchomorpha Pleurobranchomorpha Ordo Acochlidiacea Acochlidiacea Acochlidiacea Ordo Sacoglossa = Sacoglosssa Sacoglossa Sacoglosssa Ascoglossa Ordo Thecosomata Thecosomata Ordo Gymnosomata Gymnosomata Ordo Entomotaeniata Ordo Nudibranchia Nudibranchia Nudibranchia Nudibranchia * Note: Thecosomata and Gymnosomata are sometimes called: "pteropods". ** Note: Excluding the pteropods and the pyramidellids. Going further down in the classification, a summary of opisthobranch families is presented in Table 2. (This classification is also used by a very valuable database on the Internet, “The Sea Slug Forum” (Rudman, 2000)).

5
Table 2. Summary of ophistobranch families (after Beesley et al, 1998). OPISTHOBRANCHIA Order CEPHALASPIDEA Superfamily ACTEONOIDEA Family Acteonidae Family BuIlinidae Family Hydatinidae Superfamily RINGICULOIDEA Family Ringiculidae Superfamily CYLINDROBULLOIDEA Family Cylindrobullidae Superfamily DIAPHANOIDEA Family Diaphanidae Superfamily PHILINOIDEA Family Cylichnidae Family Retusidae Family Philinidae Family Philinoglossidae Family Aglajidae Family Gastropteridae Superfamily HAMINOEOIDEA Family Haminoeidae Family Bullactidae Family Smaragdinellidae Family Incertae sedis Superfamily BULLOIDEA Family Bullidae Superfamily RUNCINOIDEA Family Runcinidae Family Ilbiidae Superfamily INCERTAE SEDlS Family Notodiaphanidae Order ACOCHLIDEA Superfamily ACOCHLIDIOIDEA Family Acochlidiidae Family Hedylopsidae Superfamily MICROHEDYLOIDEA Family Asperspinidae Family Microhedylidae Family Ganatidae Order RHODOPEMORPHA Family Rhodopidae
Order SACOGLOSSA Superfamily OXYNOOIDEA Family Volvatellidae Family Oxynoidae Family Juliidae Superfamily ELYSIOIDEA Family Plakobranchidae Family Elysiidae Family Boselliidae Family GascoigneIlidae Family Platyhedylidae Superfamily LIMAPONTIOIDEA Family Caliphyllidae Family Costasiellidae Family Hermaeidae Family Limapontiidae Order ANASPIDEA Superfamily AKEROIDEA Family Akeridae Superfamily APLYSIOIDEA Family Aplysiidae Order NOTASPIDEA Superfamily TYLODINOIDEA Family Tylodinidae Family Umbraculidae Superfamily PLEUROBRANCHOIDEA Family Pleurobranchidae Order THECOSOMATA Family Limacinidae Family Cavoliniidae Family Peraclididae Family Cymbuliidae Family Desmopteridae Order GYMNOSOMATA Suborder GYMNOSOMATA Family Pneumodermatidae Family Notobranchaeidae Family Cliopsidae Family Clionidae Suborder GYMNOPTERA Family Hydromylidae
Order NUDIBRANCHA Suborder DORIDINA Superfamily ANADORIDOIDEA Family Corambidae Family Goniodorididae Family Onchidorididae Family Polyceridae Family Gymnodorididae Family Aegiretidae Family Vayssiereidae Superfamily EUDORIDOIDEA Family Hexabranchidae Family Dorididae Family Chromodorididae Family Dendrodorididae Family Phyllidiidae Suborder DENDRONOTINA Family Tritoniidae Family Bornellidae Family Marianinidae Family Hancockiidae Family Dotidae Family Scyllaeidae Family Tethydidae Family Phylliroidae Family Lomanotidae Suborder ARMININA Family Arminidae Family Doridomorphidae Family Charcotiidae Family MadreIlidae Family Zephyrinidae Family Pinufiidae Suborder AEOLIDINA Family Flabellinidae Family Eubranchidae Family Aeolidiidae Family Glaucidae Family Embletoniidae Family Tergipedidae Family Fionidae

6
1.3 Morphology, ecology and physiology in Opisthobranchia Figure 1 shows a broad characterisation of grades of organisation in the opisthobranchs. The circle “1” is considered to represent the oldest and most “primitive” level.
Figure 1. Grades (time-development level) of organisation in the opisthobranchs (Beesley et al, 1998) (Reproduced by special permission from Richard Willan and Australian Biological Resources Study, Environment Australia) As can be seen, within each order there are usually several grades of organisation. In spite of this, it is also possible to see that the different groups of nudibranchs are considered to have reached the “highest” forms of organisation (Beesley et al., 1998). There is a strong coupling between organisation grades and morphological appearances, as for instance, when the shell is reduced or lacking completely. The first way of identifying opisthobranch orders is usually by external examination as the morphology often gives clues to which order a specimen can be placed. Figure 2-9 shows some typical members of eight opisthobranch groups.
a b Figure 2. (a) Cephalaspidea, Bulla ampulla. (b) Cephalaspidea, Chelidonura sandrana (Thompson, 1976). (Thompson, 1976).

7
Figure 3. Anaspidea, Aplysia depilans Figure 4. Notaspidea, Pleurobranchus (Thompson, 1976). membranaceus (Thompson, 1976).
a b Figure 5. (a) Acochlidiacea, Hedylopsis loricata (b) Acochlidiacea, Microhedyle sp (Thompson, 1976). (Thompson, 1976).
a b
Figure 6. (a) Sacoglossa, Stiliger fuscatus (b) Sacoglossa, Elysia marginata (Thompson, 1976). (Thompson, 1976).

8
a b Figure 7. (a) Thecosomata, Spirellatidae (b) Thecosomata, Peraclidae. (Thompson, 1976). (Thompson, 1976).
a b Figure 8. (a) Gymnosomata, Crucibranchaea (b) Gymnosomata, Clione limacina. macrochira (Thompson, 1976). (Thompson, 1976).
a b Figure 9. (a) Nudibranchia, Polycera capensis (b) Nudibranchia, Tritonia festiva (Thompson, 1976). (Thompson, 1976).

9
In most books and surveys only five orders are treated. Usually Acochlidiacea is not treated since they are minute and difficult to collect (requires sieving through sand). Thecosomata and Gymnosomata are also difficult to collect since they are pelagic. Accordingly, most estimates of opisthobranch species diversity are based on just five groups. One exception to this is Thompson (1988) who states that there is “about 3000 species” world-wide. (For more information regarding opisthobranch diversity, see Table 3.) Table 3. Review of opisthobranch diversity in chosen areas. Opisthobranchia (A = Estimations, B = Found species)
A Area Source Remarks: 3000 Worldwide Thompson, 1976 700 Tropical Western
Pacific Marshall & Willan, 1999 Not including: Acochlidea, Thecosomata, Gymnosomata
600 Pacific "Equatorial Belt"
Marshall & Willan, 1999 Not including: Acochlidea, Thecosomata, Gymnosomata
550 Great Barrier Reef
Marshall & Willan, 1999 Not including: Acochlidea, Thecosomata, Gymnosomata
350 S. Barrier Reef Marshall & Willan, 1999 Not including: Acochlidea, Thecosomata, Gymnosomata 120 Sweden Hansson, 1998 Eight orders B Area Source Remarks:
561 Madang (PNG) Gosliner (in M & W, 1999) Not including: Acochlidea, Thecosomata, Gymnosomata 462 Great Barrier
Reef Marshall & Willan, 1999 Checklist of Great Barrier Reef Opisthobranchia (eight
orders) 439 Guam Carlson & Hoff (in M & W,
1999) Not including: Acochlidea, Thecosomata, Gymnosomata
390 South Africa Gosliner, 1987 Not including Acochlidea 389 Marshall Islands Johnson & Boucher (in M
& W, 1999) Not including: Acochlidea, Thecosomata, Gymnosomata
240 Suva (Fiji) Brodie & Brodie ( in M & W, 1999)
Not including: Acochlidea, Thecosomata, Gymnosomata
262 Heron Island Marshall & Willan, 1999 Not including: Acochlidea, Thecosomata, Gymnosomata 178 Norway Moen & Svensen, 1999 156 British Isles Thompson, 1988 Not including: Thecosomata, Gymnosomata 105 New Caledonia Risbec, 1928 (in M & W,
1999) Not including: Acochlidea, Thecosomata, Gymnosomata
All opisthobranchs are hermaphrodite and copulation is normally reciprocal (Thompson, 1988; Hayward & Ryland, 1995). The reproductive organs are present in all adults on the right-hand side of the body (Hayward & Ryland, 1995). Most species spawn eggs, which hatch as free swimming, shelled, veliger larvae (Thompson, 1988; Hayward & Ryland, 1995). In Figure 10 there are some examples of typical spawn masses and in Figure 11 there are examples of veliger larvae.

10
Figure 10. Examples of opisthobranch spawn masses (Behrens, 1991).

11
Figure 11. Examples of opisthobranch veliger larva (Thompson, 1988). The shelled veliger larva is normally planktotrophic while those species that hatch as miniature adults are normally lecitotrophic. There is a greater proportion of these species in polar seas (Willan & Coleman, 1984) Internally, the opisthobranchs display detorsion and many are secondarily bilaterally symmetrical. They have one auricle and one nephridium (Ruppert & Barnes, 1994). Most opisthobranchs are carnivorous, although sacoglossans and aplysiids are herbivorous (Hayward & Ryland, 1995). 1.4 Nudibranchia The largest group within the Opisthobranchia is the “Nudibranchia”. Currently, there are conflicting views whether Nudibranchia is monophyletic (Wägele & Willan, 2000) or polyphyletic (Tholleson, 1998). Wägele & Willan (2000) goes even further and propose a new taxon “Nudipleura” as a monophyletic group, which contains Pleurobranchoidea and Nudibranchia. As with the opisthobranchs, there are different views on nudibranch systematic and a short review is presented in Table 4. (A historical summary of the nudibranch divisions is shown in Table 5.)

12
Tab
le 4
. Sys
tem
atic
revi
ew o
f Nud
ibra
nchi
a ac
cord
ing
to v
ario
us a
utho
rs.
Wäg
ele
& W
illan
, 20
00 C
lade
s!
M
arsh
all &
Will
an,
1999
R
udm
an &
Will
an,
1998
(In
Bee
sley
et
al.,
1998
)
Han
sson
, 199
8 D
ebel
ius,
199
6 Th
omps
on, 1
988
Wel
ls &
Bry
ce,
1993
B
ehre
ns, 1
991
Gos
liner
, 198
7 W
illan
& C
olem
an,
1984
Anth
obra
nchi
a Su
bord
o
Dor
idin
a An
thob
ranc
hi(n
)a =
D
orid
acea
D
orid
oide
a D
orid
acea
An
thob
ranc
hia
* C
lado
bran
chia
Dor
id(a
c)in
a *
Su
bord
o
Den
dron
otin
a C
lado
bran
chin
a #
Den
dron
otac
ea
Den
dron
otoi
dea
Den
dron
otac
ea
Cla
dobr
anch
ia #
Su
bord
o
Arm
inin
a N
otas
pide
(in)a
=
Arm
inac
ea
Arm
inoi
dea
Arm
inac
ea
Pl
euro
bran
chom
orph
(in)a
Subo
rdo
Ae
olid
ina
Ae
olid
acea
Ae
olid
oide
a
Aeol
idac
ea
Infr
aord
o
Supe
rfam
ilia
Dor
idoi
dea
* E
udor
idoi
dea
* D
orid
acea
D
endr
onot
oide
a
* Pol
ycer
oide
a
# D
endr
onot
acea
Ar
min
oide
a
* Ana
dorid
oide
a
# Ar
min
acea
Ae
olid
oide
a
* Nam
e?
#
Aeol
idac
ea
* Nam
e?
* Nam
e?
# D
endr
onot
oide
a
#
Arm
inoi
dea
# Ae
olid
oide
a
Fam
ilia
Subf
amili
a Tr
ibus
G
enus
Su
bgen
us
Spec
ies
Guv
erne
d by
ICZN
G
uver
ned
by IC
ZN
Guv
erne
d by
ICZN
G
uver
ned
by IC
ZN
Guv
erne
d by
ICZN
G
uver
ned
by IC
ZN
Guv
erne
d by
ICZN
Not
e: N
ew ta
xon
prop
osed
"N
udip
leur
a" =
N
udib
ranc
hia
+ Pl
euro
bran
choi
dea
Not
e: N
udib
ranc
hia
= An
thob
ranc
hia
(=
Euct
enid
ieac
ea =
C
teni
diac
ea) +
C
lado
bran
chia
(=
Acte
nidi
acea
)
Not
e: In
Dor
idoi
dea
all s
peci
es a
re
holo
hepa
tic.
Not
e: D
orid
acea
is
divi
ded
in th
ree
grou
ps:
cryp
tobr
anch
s,
poro
stom
es a
nd
phan
erob
ranc
hs
Not
e: S
omet
imes
th
e An
thob
ranc
hia
is d
ivid
ed in
to
cryp
tobr
anch
s an
d ph
aner
obra
nchs
Foot
note
; IC
ZN =
Inte
rnat
iona
l Cod
e of
Zoo
logi
cal N
omen
clat
ure
(Fou
rth E
ditio
n 20
00)

13
Table 5. Historical summary of the nudibranch divisions (Wägele & Willan, 2000).
The most difficult nudibranch group to characterise is the Arminina/Arminacea/Arminoidea group (Figure 12).
a b Figure 12. (a) Arminina, Armina loveni (b) Arminina, Dirona albolineata (Todd, 1981). (Thompson, 1976). The other nudibranch groups are somewhat easier to describe as seen in Figures 13-15.

14
a b Figure 13. (a) Doridina, Ceratosoma cornigerum (b) Doridina, Plocamopherus ceylonicus (Thompson, 1976). (Thompson, 1976).
a b Figure 14. (a) Dendronotina, Doto yongei (b) Dendronotina, Hancockia burni (Thompson, 1976). (Thompson, 1976).
a b Figure 15. (a) Aeolidina, Pseudovermis mortoni (b) Aeolidina, Facelina annulicornis (Thompson, 1976). (Thompson, 1976).

15
As with the opisthobranchs, the number of nudibranch species is largest in the tropics (Willan & Coleman, 1984). As most recent publications treat Nudibranchia as an order or a suborder, the corresponding lower levels are either suborders or (super) families. Regardless, Nudibranchia is usually considered to contain four subgroups: Doridina/Doridacea/Dorodoidea, Aeolidina/Aeolidacea/Aeolidoidea, Arminina/Arminacea/Arminoidea and Dendronotina/Dendronotacea/Dendronotoidea. The group with the largest number of species is the Doridina/Doridacea/Dorodoidea. In “Checklist of Great Barrier Reef Opisthobranchia” (Marshall & Willan, 1999) 281 nudibranch species are recorded totally and out of them, 204 species belong to the Doridina/Doridacea/Dorodoidea group. According to Wägele & Willan (2000) Nudibranchia consists of about 190 genera and 60 families. In Table 6, examples of various nudibranch estimations or records are shown. Table 6. Review of nudibranch diversity in chosen areas.
Nudibranchia (A = estimations, B = found species) A Area Source Remarks
3000 Worldwide Willan & Coleman 1984 800 Tropical Indo Pacific Willan & Coleman 1984 B Area Source Remarks
281 Great Barrier Reef Marshall & Willan, 1999 Checklist of Great Barrier Reef Opisthobranchia
269 Australia Willan & Coleman 1984 Checklist of Australian Nudibranchia 250 Mediterreanean Cattaneo-Vietti et al, 1990 Checklist of the Mediterranean
Nudibranchs 241 South Africa Gosliner, 1987 Southern African Opisthobranch
Species 108 British isles Picton & Morrow, 1994 British Check List 108 British Isles Thompson, 1988 British Check List 80 Sweden Hansson, 1998 Included 2 species of Notaspidea!
The life span of different nudibranch species varies between a few weeks and two years. Usually three ecological groupings are used: “Subannual“, “Annual”, and “Biennial” species. The subannual species are normally feeding on seasonally variable resources while the biennial species are feeding on stable diets. All nudibranchs appear to be semelparous (Todd, 1981). The majority of anthobranchs eat sponges or bryozoans and most cladobranchs eat coelenterates. Other nudibranch diets include ascidians, barnacles, crustaceans, and even the spawn of other opisthobranchs (Willan & Coleman, 1984). The most complete review regarding nudibranch food has been provided by McDonald & Nybakken (1997). Two families of the Doridina/Doridacea/Dorodoidea group are very common both in species number and abundance in the Indo-Pacific: Chromodorididae and Phyllidiidae. 1.5 Chromodorididae This family has a lot of species (more than 300 according to Wells & Bryce, 1993) and many are difficult to distinguish. For instance, in order to separate the genus Chromodoris and the genus Hypselodoris, the researcher usually has to dissect, prepare and compare the radula (Figure 16), which is present in all chromodorids. This takes a lot of skill and access to a good microscope or even a scanning electron microscope (SEM).
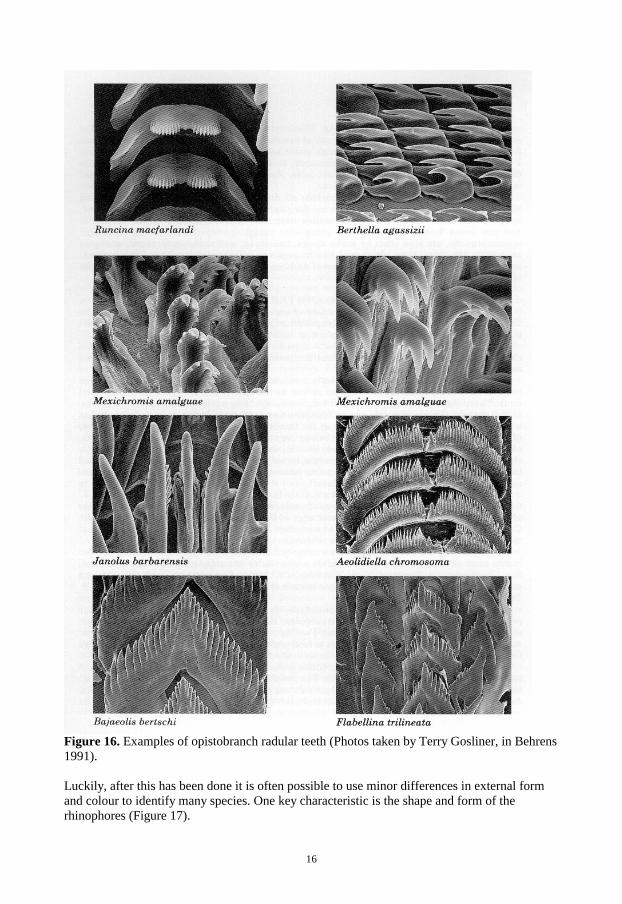
16
Figure 16. Examples of opistobranch radular teeth (Photos taken by Terry Gosliner, in Behrens 1991). Luckily, after this has been done it is often possible to use minor differences in external form and colour to identify many species. One key characteristic is the shape and form of the rhinophores (Figure 17).

17
Figure 17. Various shapes and forms of ophistobranch rhinophores (Behrens, 1991). Especially within the Chromodorididae there are many “colour groups” which are very similar in appearances (Rudman, 1973, 1977, 1982, 1984, 1991; Gosliner & Johnson 1999). Often their appearance is quite colourful and it is thought that this is an example of aposematism. As several species then share the same general colour markings it is suspected that this is also an example of Müllerian mimicry and that the species take advantage of similar coloration in order to avoid predation. The deterrent, which makes this work, is normally a substance acquired from sponges, which are the normal food for chromodorids. In some cases it is just a question of accumulating useful compounds from the sponges but there are also some chromodorids which can convert ingested substances to a more “toxic” state (Karuso, 1987).

18
1.6 Phyllidiidae Another interesting family is the Phyllidiidae, it is also a member of the Doridina/Doridacea/Dorodoidea group and according to Wells & Bryce (1993) the family consists of about 70 species. The phyllidiids lack a radula (Brunckhorst, 1993; Yonow, 1996). Instead they “suck” in pieces of sponges which they utilize as food. Together with the dendrodorids (which also lacks radula) the phyllidiids constitutes a group which is called “porostomata”. The phyllidiids can secrete a “milky” fluid when handled and this fluid can work as a poison. This was first noted when a phyllidiid accidentally killed all the lobsters in an aquarium where it was put for storage. After this discovery a lot of research have been conducted on chemical substances in different species of phyllidiids (Karuso, 1987). (Some chemical structures of these compounds are shown in Figure 18-19).
Figure 18. Toxin from Phyllidia varicosa, Figure 19. Major metabolite from 9-isocyanopupukeananae P. pulitzeri, axisonitrile-1 (Karuso, 1987). and its 2-isomer (Karuso, 1987). The phyllidiids are mainly restricted to tropical areas and there are just a few species of this family in more temperate water (Gosliner & Behrens, 1988). 1.7 References Avila, C. 1995. Natural products of opisthobranch molluscs: a biological review.
Oceanography and Marine Biology: an Annual Review 33: 487-559. Beesley, P. L., Ross, G. J. B. & Wells, A. (eds). 1998. Mollusca: The Southern Synthesis.
Fauna of Australia. Vol. 5. CSIRO Publishing. Melbourne. Behrens, D. W. 1991. Pacific Coast nudibranchs: a guide to the opisthobranchs, Alaska to
Baja California. Seachallenger. Monterey. Bohlin, L. 1989. Pharmacologically active compounds from marine organisms. Acta
Pharmacetica Nordica 1: 175-182. Brunckhorst, D. J. 1993. The systematics and phylogeny of phyllidiid nudibranchs
(Doridoidea). Australian Museum. Sydney. Cattaneo-Vietti, R., Chemello, R. and Gianuzzi-Savelli, R. 1990. Atlas of Mediterranean
nudibranchs. La Conchiglia. Roma. Cimino, G., Fontana, A. and Gavagnin, M. 1999. Marine Opisthobranch Molluscs:
Chemistry and Ecology in Sacoglossans and Dorids. Current Organic Chemistry 3: 327-372. Debelius, H. 1996. Nudibranchs and sea snails Indo Pacific field guide. IKAN –
Unterwasserarchiv. Frankfurt. Faulkner, D. J. and Ireland, C. 1977. The chemistry of some opisthobranch molluscs. Pp 23-
34 in D. J. Faulkner and W. H. Fenical (eds). Marine natural products chemistry. Plenum Press. New York.

19
Faulkner, D. J. and Ghiselin, M. T. 1983. Chemical defence and evolutionary ecology of dorid nudibranchs and some other opisthobranch gastropods. Marine Ecology – Progress Series 13: 295-301.
Gosliner, T. 1987. Nudibranchs of Southern Africa. Seachallenger. Monterey. Gosliner, T. M. and Behrens, D. W. 1988. A Review of the Generic Divisions Within the
Phyllidiidae with the Description of a New Species of Phyllidiopsis (Nudibranchia: Phyllidiidae) from the Pacific Coast of North America. The Veliger 30: 305-314.
Gosliner, T. M. and Behrens, D. W. 1990. Special Resemblance, Aposematic Coloration and Mimicry in Opisthobranch Gastropods. Pp 127-138 in M. Wicksten (ed). Symposium on the Adaptive Significance of Colour in Invertebrates. University Press Texas A & M.
Gosliner, T. M. and Johnson, R. F. 1999. Phylogeny of Hypselodoris (Nudibranchia: Chromodorididae) with a review of the monophyletic clade of Indo Pacific species, including descriptions of twelve new species. Zoological Journal of the Linnean Society 125: 1-114.
Gunthorpe, L. and Cameron, A. M. 1987. Bioactive properties of extracts from Australian dorid nudibranchs. Marine Biology 94: 39-43.
Hansson, H. G. 1998. Sydskandinaviska marina flercelliga evertebrater, utgåva 2. Länsstyrelsen Västra Götaland. Göteborg.
Hayward, P. J. & Ryland, J. S. (eds). 1995. Handbook of the Marine Fauna of North – West Europe. Oxford University Press. Oxford, New York, Tokyo.
ICZN, Fourth Edition. 1999. International Code of Zoological Nomenclature. The International Trust for Zoological Nomenclature. London.
Karuso, P. 1987. Chemical Ecology of the Nudibranchs. Pp 32-60 in Scheuer, P. J. (ed). Bioorganic Marine Chemistry, Volume 1. Springer- Verlag. Berlin, Heidelberg.
McDonald, G. & Nybakken, J. 1997. A Worldwide Review of the food of Nudibranch Mollusks. The Veliger 40: 157-159.
Marshall, J. G. and Willan, R. C. 1999. Nudibranchs of Heron Island, Great Barrier Reef. Backhuys Publishers. Leiden.
Moen, F. E. & Svensen, E. 1999. Dyreliv i havet. KOM forlag, Kristiansund. Newman, L. J., Cannon, L. R. G. and Brunckhorst, D. J. 1994. A new flatworm
(Platyhelminthes: Polycladida) which mimics a Phyllidiid Nudibranch (Mollusca, Nudibranchia). Zoological Journal of the Linnean Society 110: 19-25.
Picton, B. E. & Morrow, C. C. 1994. A Field Guide to the Nudibranchs of the British Isles. Immel Publishing. London.
Ponder, W. F. and Lindberg, D. R. 1997. Towards a phylogeny of gastropod molluscs: an analysis using morphological characters. Zoological Journal of the Linnean Society 119: 83-265.
Rudman, W. B. 1973. Chromodorid opisthobranch Mollusca from the Indo-West Pacific. Zoological Journal of the Linnean Society 52: 175-199.
Rudman, W. B. 1977. Chromodorid opisthobranch Mollusca from East Africa and the tropical West Pacific. Zoological Journal of the Linnean Society 61: 351-397.
Rudman, W. B. 1982. The Chromodorididae (Opisthobranchia: Mollusca) of the Indo-West Pacific: Chromodoris quadricolour, C. lineolata and Hypselodoris nigrolineolata colour groups. Zoological Journal of the Linnean Society 76: 183-241.
Rudman, W. B. 1984. The Chromodorididae (Opisthobranchia: Mollusca) of the Indo –West Pacific: a review of the genera. Zoological Journal of the Linnean Society 81: 115-273.
Rudman, W. B. 1991. Purpose in pattern: the evolution of colour in chromodorid nudibranchs. Journal of Molluscan Studies 57: 5-21.
Rudman, W. B. 2000. The Sea Slug Forum, www.austmus.gov.au/science/invert/mal/forum Ruppert, E. E. & Barnes, R. D. 1994. Invertebrate Zoology. Saunders College Publishing.
Fort Worth, Philadephia, San Diego.

20
Thollesson, M. 1998. Nudibranch systematics and molecular data. PhD Thesis, Göteborgs Universitet.
Thompson, T. E. 1976. Biology of opisthobranch molluscs. Volume I. The Ray Society. London.
Thompson, T. E. 1988. Molluscs: Benthic Opisthobranchs. E. J. Brill / Dr W. Backhuys. Leiden, New York, København, Köln.
Todd, C. D. 1981. The ecology of nudibranch molluscs. Oceanography and Marine Biology: an Annual Review 19: 141-234.
Tullrot, A. 1998. Evolution of warning coloration in the nudibranch Polycera quadrilineata. PhD Thesis, Göteborgs Universitet.
Wells, F. E. & Bryce, C. W. 1993. Sea slugs of Western Australia. Western Australian Museum. Perth.
Willan, R. C. & Coleman, N. 1984. Nudibranchs of Australasia. Australasian Marine Photographic Index. Sydney.
Wägele, H. & Willan, R. C. 2000. Phylogeny of the Nudibranchia. Zoological Journal of the Linnean Society 130: 83-181.
Yonow, N. 1996. Systematic revision of the family Phyllidiidae in the Indian Ocean province: Part 1 (Opisthobranchia: Nudibranchia: Dorodoidea). Journal of Conchology 35: 483-516.

21
2 Checklist of opisthobranchs of the Wakatobi National Park, SE Sulawesi, Indonesia
2.1 Introduction Operation Wallacea (“Opwall”) is a joint venture between the Indonesian “Wallacea Development Institute” and a British company: Ecological Surveys Ltd. The co-operation started 1995 and the purpose was to promote conservation work in a remote Indonesian archipelago, Tukang Besi (Chart 1). The Operation Wallacea works with both land and marine projects, concentrating mainly on field surveys regarding diversity issues but it has also been involved in some projects promoting sustainable use of natural resources. This means, for instance, changing the fishing methods and promoting ecotourism in the area. A significant result of this work was achieved when “The Wakatobi Marine National Park” was created in July 1996. It is currently the second largest Indonesian “National Marine Park” (13000 km2) stretching between 05º15’S, 123º23’E and 06º08’S, 124º37’E (Stanzel & Newman, 1997). The name Wakatobi comes from the first letters of the four largest islands in the area (Wanci, Kaledupa, Tomea and Binongko). One aim of Operation Wallacea is to survey and publish checklists of the marine life in the national park. During 1997-99 a lot of work was geared towards a checklist of Opisthobranchia. This paper presents results from this work.
Chart 1. Tukang Besi archipelago, Indonesia (Expedia, 2001). 2.2 Methods The marine projects, run by Operation Wallacea, are concentrated around a small island, Hoga (Chart 2), situated near Kaledupa. During my visit 1999, two major projects were conducted, one regarding correlation between butterfly fish diversity and coral diversity and another one regarding opisthobranch diversity. Paying volunteers did nearly all work but there was also some scientific staff supervising the projects. The opisthobranch project started in 1997 (two

22
weeks) and continued for 10 months in 1998 and for 7 months in 1999. The main effort was done during 1999 when a lot of work was concentrated at opisthobranchs.
Chart 2. Hoga Island, Tukang Besi archipelago, Indonesia (Expedia, 2001). In the opisthobranch project, groups of diving volunteers concentrated on finding as many species of opisthobranchs as possible (down to a maximum depth of 30 m). For each recording they noted numbers of individuals, dates, depths and sizes. All species were photographed and unidentifiable species were also drawn by hand. The main identification books used at Hoga were: Allen & Steene, 1996; Behrens, 1991; Coleman, 1989; Debelius, 1996; Gosliner, 1987, Gosliner, et al, 1996; and Wells & Bryce, 1993. The collected data for 1999 (handwritten notes) were then taken to England and later also copied and sent to Sweden where the compilation (Appendix I) was made during the year of 2000. During this work it was possible to identify some more species with the help of the Internet, especially as the site “The Sea Slug Forum” proved to be very valuable (Rudman, 2000). At least one specimen of each species was conserved at Hoga for later shipment to the Bogor Zoological Museum. Also, from many species, a small tissue sample (large species) or a whole individual (small species) was taken and conserved. The aim with this material is to compare the DNA from the species and to do a cladistic analysis later on (Warren, 2000 and personal communication, October 1999*). Notes of the taxa found during 1997 or 1998 but not 1999 are added to “Appendix I” but these taxa are not included in calculations regarding “observations” or “occasions” due to lack of data. The reason for choosing the word “observations” instead of “individuals” is because each individual could have been recorded more than once. The term “occasions” is used since not all survey work was done diving. In a few cases surveys were made in the intertidal area during low tide. * Lindsay Warren, personal communication, October 1999 (Hoga Island).
Hoga Island

23
2.3 Results A total of 60 different species were found during two weeks in September 1997. Between March and December 1998 another 115 opisthobranch species were found. The total species number for the whole period 1997-1999 is 297 species (Warren, 2000). During the season of 1999 (May-November), 3523 opisthobranchs were found during the survey. They belonged to a total of 235 identified or suspected (provisionally identified) species and to 73 identified genera (Appendix I). Table 1. Summary of “The 1999 Opisthobranch Survey – Operation Wallacea” (Indonesia) Anaspidea Cephalaspidea Notaspidea Nudibranchia Sacoglossa Species 9 36 5 161 23 Observations 116 316 42 2888 159
Two additional observations were of Colpodaspis thompsoni, which probably belong to Cephalaspidea but its current taxonomic status is uncertain (Marshall & Willan, 1999). The results regarding discovered species and genera for the periods 1997-1999, 1997-1998 and 1999 are shown in Figure 1 and 2.
Operation Wallacea
Anaspidea
Cephalaspidea
Notaspidea
Nudibranchia
Sacoglossa
0 50 100 150 200 250
Cat
egor
y
Number of species
1997-199919991997-1998
Figure 1. Summary of species, “Operation Wallacea - Opisthobranchia survey” 1997-1999 (Indonesia).

24
Operation Wallacea
Anaspidea
Cephalaspidea
Notaspidea
Nudibranchia
Sacoglossa
0 20 40 60
Cat
egor
y
Number of identified genera
1997-199919991997-1998
Figure 2. Summary of identified genera, “Operation Wallacea - Opisthobranchia survey” 1997-1999 (Indonesia). In Figure 3 there is a summary of the results of the identification effort.
Identification of Opisthobranchs 1997-99
Genus + speciesGenus Only higher taxa
Figure 3. Result of identification effort regarding 295 opisthobranch taxa, “Operation Wallacea - Opisthobranchia survey” 1997-1999 (Indonesia). The proportions (species and genera) between the nudibranch suborders for the period 1997-1999 are presented in Figure 4 and 5.

25
Nudibranchia 1997-99
AeolidinaArmininaDendronotinaDoridina
Figure 4. Proportion of species within nudibranch suborders, Operation Wallacea survey 1997-1999 (Indonesia). Totally 208 nudibranch species.
Nudibranch genera 1997-99
AeolidinaArmininaDendronotinaDoridina
Figure 5. Proportion of genera within nudibranch suborders, Operation Wallacea survey 1997-1999 (Indonesia). Totally 56 nudibranch genera.

26
Table 2 presents a list of the most common species. Some of these species (and a few others) are illustrated in Plate A and B. Table 2. The most common opisthobranch species during Operation Wallacea survey 1999 (Indonesia). Order Species Observations Occasions Cephalaspidea Chelidonura amoena 24 17 Notaspidea Pleurobranchus forskalii 20 11 Nudibranchia Phidiana indica 17 17 Nudibranchia Phyllodesmium briareum 398 27 Nudibranchia Pteraeolidia ianthina 25 19 Nudibranchia Dermatobranchus sp (OWN 101) 20 13 Nudibranchia Chromodoris annae 215 109 Nudibranchia Chromodoris coi 35 30 Nudibranchia Chromodoris dianae 106 60 Nudibranchia Chromodoris geometrica 49 40 Nudibranchia Chromodoris kuniei 22 17 Nudibranchia Chromodoris leopardus 13 11 Nudibranchia Chromodoris lochi 132 81 Nudibranchia Chromodoris magnifica 119 71 Nudibranchia Chromodoris willani 106 63 Nudibranchia Hypselodoris bullocki 18 13 Nudibranchia Jorunna funebris 29 26 Nudibranchia Nembrotha kubaryana 36 ? Nudibranchia Phyllidia coelestis 113 82 Nudibranchia Phyllidia elegans 164 115 Nudibranchia Phyllidia ocellata 19 19 Nudibranchia Phyllidia varicosa 131 96 Nudibranchia Phyllidiella pustulosa 604 289 Nudibranchia Phyllidiella rudmani 30 24 Nudibranchia Phyllidiopsis pipeki 58 52 Nudibranchia Phyllidiopsis shireenae 12 11 Nudibranchia Phyllidiopsis striata 34 28 Nudibranchia Reticulidia fungia 15 14 Nudibranchia Roboastra gracilis 16 16 Sacoglossa Elysia sp (OWN 99) 15 12 Sacoglossa Thuridilla bayeri 36 25 Sacoglossa Thuridilla hoffae 13 11 Total number 2644
2.4 Discussion Appendix I is the summary of more than 450 hand-written pages with opisthobranch information from the 1999 survey. The material was delivered in three batches between January and April 2000. As numerous people wrote the material, a continuing obstacle was to decipher the notes accurately. Another difficulty was the uses of many synonyms as this made the compilation work very time consuming. A third problem was the fact that the most complete photographic handbook (Debelius, 1996) contains many misidentifications and lacks information about authors.

27
As seen from Figure 1 most of the species (in all orders) were found during 1999. The reason for this is probably because of the increased effort under the 1999 season (May – November). Regrettably, there is no information regarding search effort, which makes it difficult to compare the three years. When looking at the number of genera instead (Figure 2) the picture is less clear. Fewer Cephalaspidean genera were found 1999 than 1997-1998. This is probably because of less search effort during 1999 than 1997-1998. (During my own visit, no actual digging was conducted in search of the burrowing Cephalaspidea.) It is interesting to note that, despite a massive identifying effort, only about 60 % of the opisthobranchs were identified to species level (Figure 3). The majority of the observations involved members of the Nudibranchia, which is not surprising since Nudibranchia is the largest order in Opisthobranchia. As seen from Figures 4 and 5, the most common nudibranch suborder is the Doridina whether looking for species number or generic number. There are very few similar surveys to compare with. The most recent one is the “Nudibranchs of Heron Island, Great Barrier Reef” (Marshall & Willan, 1999). This survey was run between November 1980 and December 1998 and a total of 262 species was found during that period. In comparison 295 species were found during the Operation Wallacea surveys 1997-1999 (Figures 1 and 3). This number will probably be somewhat lower after final examinations but it is still an impressive figure after only three years of surveys.( According to Warren (2000) there are 297 species but I have excluded Colpodaspis thompsoni and Tritonopsilla australis. In the first case because of its uncertain taxonomic status and in the later case because I believe it to be a case of mistaken identity.) The highest count of opisthobranchs for an ongoing survey in the Indo Pacific is 561 species (Madang, Papua New Guinea) but that is for the period 1986 – March 1997 (Gosliner in Marshall & Willan, 1999). If not just looking at surveys then Gosliner & Draheim (1996) give the highest count of Indo Pacific opisthobranchs in a study. They state that there are more than 3400 opisthobranch species in the region and that the locality with the most species, 646, is Papua New Guinea. No doubt, these figures shows that the region of Papua New Guinea and Indonesia is the leading diversity centre for Opisthobranchia. According to Table 2, 75 % of the opisthobranch observations were of the 14 % most common species. One problem is to categorise the found species. Is a species always common when found in large numbers? What if the species is found only a few times but in large number? In my case I choose to regard all species, which were found on more than 10 separate occasions (during the 1999 season) as common. In “Nudibranchs of Heron Island, Great Barrier Reef” (Marshall & Willan, 1999) there is an attempt to give a relative abundance scale. This could function but since the authors do not give information about the time duration of sampling periods it was not possible to use their abundance scale in this report.

28
2.5 References Allen, G. R. & Steene, R. 1996. Indo-Pacific coral reef field guide. Tropical Reef Research.
Singapore. Behrens, D. W. 1991. Pacific Coast nudibranchs: a guide to the opisthobranchs, Alaska to
Baja California. Seachallenger. Monterey. Coleman, N. 1989. Nudibranchs of the South Pacific. Neville Coleman’s Sea Australia
Resource Centre. Springwood. Debelius, H. 1996. Nudibranchs and sea snails Indo Pacific field guide. IKAN –
Unterwasserarchiv. Frankfurt. Expedia, 2001. Internet site: www.expedia.com (March 2001). Gosliner, T. 1987. Nudibranchs of Southern Africa. Seachallenger. Monterey. Gosliner, T. M., Behrens, D. W. and Williams, G. C. 1996. Coral Reef Animals of the Indo-
Pacific. Seachallengers. Monterey. Gosliner, T. M. and Draheim, R. 1996. Indo-Pacific opisthobranch gastropod biogeography:
how do we know what we don’t know? American Malacological Bullentin 12: 37-43. Marshall, J. G. & Willan, R. C. 1999. Nudibranchs of Heron Island, Great Barrier Reef.
Backhuys Publishers. Leiden. Rudman, B. 2000. The Sea Slug Forum. (www.austmus.gov.au/science/invert/mal/forum) Stanzel, K. B. & Newman, H. 1997. Progress report on the 1996 marine survey of the Tukang
Besi (Wakatobi) archipelago, South East Sulawesi, Indonesia. (Internal report: Operation Wallacea).
Warren, L. 2000. Opisthobranchs of the Tukang Besi archipelago, S.E. Sulawesi, 1999 survey report & results. (www.operationwallacea.win-uk.net/opisthob/opisthob.htm (2000-11-15))
Wells, F. E. & Bryce, C. W. 1993. Sea slugs of Western Australia. Western Australian Museum. Perth.

29
3 Spatial distribution of nudibranchs according to benthic structure 3.1 Introduction Why do we know so little about nudibranch ecology? One reason is that nudibranch populations are both scarce and transient (Todd, 1981, Marshall & Willan, 1999). This means that finding places where one can do nudibranch research are difficult. One potential site for such research is at Hoga Island (Indonesia) as the nudibranchs are rather abundant at this area. There are also many nudibranch species around Hoga that are quite common and not so shortlived and that makes it possible to do interesting scientific research at this locality. The research facility is run by Operation Wallacea, a joint venture between The Wallacea Development Institute (Indonesia) and Ecosurveys Ltd (United Kingdom). Hoga Island is situated in the Tukang Besi archipelago, South East of Sulawesi. In July 1996 the area was designated as the Wakatobi Marine National Park (Stanzel & Newman, 1997) and since then there has been ongoing research at this locality. My hypothesis was: Nudibranch distribution depends on benthic structure. The reason for the hypothesis is that I wanted to see if it was possible to find environment preferences among some of the more common nudibranchs in the area. 3.2 Material and methods The dive sites, chosen for research, were “Sampela” and “Buoy 3” (Chart 1). Sampela was considered an example of a more polluted area (from a nearby village) and Buoy 3 an example of a more “unspoilt” area. The survey was conducted in 1999 between the eight and twelfth of November (five consecutive days). On both locations we put down two stationary lines, that each were 30 m long. One line was put at “+12 m” and another one at “+3 m”. The “+” sign indicates that the lines were put down on an estimated depth of 3 or 12 m below the reef crest. This was done in accordance with other ongoing projects and relevant literature at hand (English et al. 1994). The actual survey depth varied with the tidal state but we recorded the depth of the starting point on each dive so that we could adjust our measurements later if necessary. On each dive we aimed to spend 20 minutes searching along each line. The search area was roughly 2.5 m above and under the line, thus having a search area of about 150 m2. An alternative search description is to say that we surveyed a volume of 300 m3 (this figure is calculated from the volume of a half cylinder with a radius of 2.5 m). We dived at each line twice a day (before and after lunch), recording observed species, depths and numbers and measuring individual sizes. The number of divers, on each dive, varied between two and five.

30
Chart 1. Location of the dive sites Buoy 3 and Sampela (Tukang Besi, Indonesia) In order to measure the benthic structure, the length of the benthic forms along the line transect was recorded. Standardised codes were used to describe the benthic forms, see Table 1. Collected data were later analysed by using statistical methods such as ANOVA (Analysis of Variance) and DCA (Detrended Correspondence Analysis). Comparable indices regarding benthic structure and diversity of nudibranchs for the two sites were computed by using the Shannon-Weaver Index formula: H = - Σ pi ln pi (Fowler et al., 1998). (The term pi is the proportion of a particular species in a sample which is multiplied by the natural logarithm of itself. H is derived by summing the product for all species in the sample. The minus sign is to make the final value of H positive.)
Buoy 3
Sampela

31
Table 1. Standardised benthic codes. After English et al., 1994. Hard coral Algea Other
ACB = Acropora Branching AA = Algal assembly OT = Other ACE = Acropora Encrusting CA = Coralline algea SC = Soft coral ACD = Acropora Digitate HA = Halimeda SP = Sponge ACS = Acropora Submassive MA = Macro algea SPB = Sponge Branching ACT = Acropora Tabulate TA = Turf algea SPD = Sponge Digitate CB = Coral Branching Abiotic SPE = Sponge Encrusting CD = Coral Digitate DDD = Data missing SPF = Sponge Foliose CE = Coral Encrusting R = Rubble SPM = Sponge Massive CF = Coral Foliose RCK = Rock SPO = Sponge Other form CM = Coral Massive S = Sand SPS = Sponge Submassive CMR = Coral Mushroom SI = Silt SPT = Sponge Tubular, Barrel, CS = Coral Submassive WA = Water Vase DC = Dead coral ZO = Zoanthids DCA = Dead coral With algea CHL = Heliopora CME = Millepora
3.3 Results The results from measuring benthic structure are shown in Table 2. Shannon-Weaver Indices for the four transects were calculated according to Fowler et al., 1998. Table 2. Benthic structure at Buoy 3 and Sampela (Tukang Besi, Indonesia). Benthic code (See Table 1)
Buoy 3 (“+ 3 m”) % Cover
Buoy 3 (“+ 12 m”) % Cover
Sampela (“+ 3 m”) % Cover
Sampela (“+ 12 m”) % Cover
AA 1 2 1 3 ACB 3 CA 7 32 9 20 CB 10 1 CD 10 1 CE 13 15 7 7 CF 2 1 1 CM 1 CMR 3 4 CS 10 DCA 3 3 DC 7 MA 1 OT 2 1 R 12 2 25 11 S 3 38 SC 3 7 13 12 SP 2 5 7 3 SPE 1 4 WA 26 32 3 2 Shannon-Weaver Index
2.251 1.746 2.422 1.780

32
The average line depth and search time (and their standard deviation = SD) for Buoy 3 and Sampela are given in Table 3. Table 3. Average search time and line depth for Buoy 3 and Sampela (Tukang Besi, Indonesia). Buoy 3
AM “+3m”
Buoy 3 PM “+3m”
Buoy 3 AM “+12m”
Buoy 3 PM “+12m”
Sampela AM “+3m”
Sampela PM “+3m”
Sampela AM “+12m”
Sampela PM “+12m)
Average search time (min)
27 27 30 31 32 28 33 31
SD 8 3 11 14 7 6 13 14 Average line depth (m)
4.7 4.7 11.9 14.0 5.5 6.8 12.2 13.3
SD 0.78 0.18 0.22 0.48 0.24 0.05 0.41 0.25 During 40 dives we managed to find a total of 252 nudibranchs belonging to at least 19 species (two nudibranch taxa were not identified to species level). At Sampela we found 138 nudibranchs and at Buoy 3 we found 114. The search record regarding observations and species is presented in Table 4. The complete search record is available in Appendix II. Table 4. Search record for Buoy 3 and Sampela (Tukang Besi, Indonesia). Species Buoy 3 Sampela Chromodoris annae 15 7 Chromodoris boucheti 3 0 Chromodoris coi 0 5 Chromodoris dianae 6 0 Chromodoris elisabethina 5 0 Chromodoris geometrica 1 2 Chromodoris kunei 0 2 Chromodoris leopardus 8 0 Chromodoris magnifica 19 0 Jorunna funebris 17 21 Nembrotha kubaryana 6 0 Reticulidia fungia 3 0 Phyllidiopsis pipeki 5 7 Phyllidiopsis striata 0 9 Phyllidia coelestis 3 15 Phyllidia sp 0 2 Phyllidia varicosa 5 10 Phyllidiella pustolosa 18 55 Unidentified 0 1 Pteraolidia ianthina 0 1 Thecacera picta 0 1
The Shannon-Weaver Index (Fowler et al., 1998) regarding nudibranch species for Buoy 3 is 2.377 and for Sampela 1.976.

33
The effects of the sites (Sampela, Buoy 3), depths (+3m, +12m) and times (“am”, “pm”) were analysed by an ANOVA (By the computer program MANOVA). Both nudibranch observations (Figure 1) and species number (Figure 2) were tested. The lowest p-value (0.08) was found for interaction of sites and depths regarding number of species. In no cases “am/pm” seemed to affect species number or observations.
�������������� ����� ��������
������������ �������� �������
�������������� �����
����������������
������������ �����
��������������
B u o y 3 - S a m p e la
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
Tota
lly (+
3m)
Chr
omod
orid
idae
(+3m
)
Phyl
lidiid
ae
(+3m
)
Tota
lly (+
12m
)
Chr
omod
orid
idae
(+12
m)
Phyl
lidiid
ae
(+12
m)
Num
ber o
f obs
erva
tion s
T o ta l B u o y 3�����A v e r a g e B o u y 3T o ta l S a m p e la�����A v e r a g e S a m p e la
Figure 1. Buoy 3 and Sampela (Tukang Besi, Indonesia): Nudibranch observations and average (with SE bars) at two depths.
������������������������������������
���������������
������������
����������������������������
������������������������
����������������
������������������������
������������
���������������������
������������������������ ��������
���������������
B u o y 3 - S a m p e la
0
2
4
6
8
1 0
1 2
1 4
1 6
Tota
lly (+
3m)
Chr
omod
orid
idae
(+3m
)
Phyl
lidiid
ae
(+3m
)
Tota
lly (+
12m
)
Chr
omod
orid
idae
(+12
m)
Phyl
lidiid
ae
(+12
m)
Num
ber o
f spe
cies
B uo y 3������A ve ra g e B uo y 3S a m p e la������A ve ra g e S a m p e la
Figure 2. Buoy 3 and Sampela (Tukang Besi, Indonesia): Species number and average (with SE bars) of nudibranchs, at two depths.

34
B u o y 3 - S a m p e la
0
2
4
6
8
1 0
1 2
1 4
Tota
lly (B
uoy
3)
Chr
omod
orid
idae
(Buo
y 3)
Phyl
lidiid
ae(B
uoy
3)
Tota
lly (S
ampe
la)
Chr
omod
orid
idae
(Sam
pela
)
Phyl
lidiid
ae(S
ampe
la)
Spec
ies
num
ber
To ta lA ve ra g e
Figure 3. Buoy 3 and Sampela (Tukang Besi, Indonesia): Total species number and average (with SE bars) of nudibranchs. Collected data was also used in a Detrended Correspondence Analysis (DCA). This is an indirect gradient analysis technique which can be used for explanatory analysis (Gauch, 1984). The analysis was made with the computer program CANOCO 4.0 (Ter Braak, 1998). The result from this analysis is shown in Figure 4.

35
Figure 4. Results from a “Detrended Correspondence Analysis” (DCA). The data are taken from the sites: Buoy 3 and Sampela (Tukang Besi, Indonesia) 3.4 Discussion Totally, the average number of observed nudibranchs was higher at Sampela (7) than Buoy 3 (6). We found approximately the same average number (6) of nudibranchs around Buoy 3 regardless of depth. At Sampela there was a difference in nudibranch abundance between the
Nembrotha kubyaryana
Reticulidia fungia
Chromodoris leopardus
Chromodoris dianae
Chromodoris magnifica
Chromodoris elisabethinaChromodoris boucheti
Chromodoris annae
Phyllidiopsis pipeki
Jorunna funebris
Chromodoris geometrica
Sp
Thecacera picta
Phyllidia varicosa
Phyllidiella pustulosa
Phyllidia coelestis
Ptereoalidia ianthina
Phyllidia spPhyllodiopsis striataChromodoris coi
Chromodoris kunei
0
2
4
0 2 4 6
Cor. massive
Other
Water
Macro algae
Cor. digitate
Dead cor. with algaeCor. foliose
Cor. branching
Cor. algeaSponge
Algal assemblyRubble
Soft coralSponge encrusting
Sand
Bouy 3+3
PMAM
+12Sampela
1.2
1.4
1.6
1.8
1 2 3
[Acropora encr,Cor. submassive,Dead coral ]
[Cor. encr.,Cor. mushroom ]

36
depths. We found on average more nudibranchs at “+3m” then at “+12m” (Figure 1). The lower number for “+12m” is probably due to the fact that there was a large sandy area at a part of this depth (very few nudibranchs live on sand). At both depths at Buoy 3 the normal pattern was that we would find more chromodorids than phyllidiids. This was not the case at Sampela however, where the situation was reversed. As this reversal occurs whether we look at individual number (Figure 1) or number of species (Figure 2), it would be interesting to check more similar sites and see if there is a larger pattern. There is a possibility that this reversal could work as an environment indicator since we consider Sampela more polluted than Buoy 3. The highest number of species, 14, was found at Buoy 3 but Sampela was not far behind with 12 species. This was in total numbers, when looking at the average values per dive there was no difference between the sites. On each dive we could expect to find about 4 species whether we were diving at Buoy 3 or Sampela (Figure 3). When comparing the “Shannon-Weaver Index” for the two sites (Sampela, 1.976 and Buoy 3, 2.377) there is a cause to say that Buoy 3 seemed to be more diverse than Sampela. As for my hypothesis that “Nudibranch distribution depends on benthic structure”, it is difficult to get a clear picture of what kind of benthic structure the different nudibranchs prefer but according to Figure 4 some of the surveyed nudibranch species are affiliated with specific benthic forms, as Chromodoris dianae, C. annae and C. magnifica occurred with macro algea, digitate and branching coral. Another indication is that Chromodoris dianae, C. annae, C. magnifica, C. coi, C. kunei, Phyllidiella pustolosa, Phyllidia sp and Phyllidiopsis striata seemed to prefer “+3m” to “+ 12m”. Since most nudibranchs are supposed to settle near food sources, I had expected a much more clear and close affiliation between Chromodoridae/Phyllidiidae and the “sponge” categories, as sponges are supposed to be their food source. This connection is not so easy to see in Figure 4. Nevertheless, I still think it should be possible to find out more facts regarding nudibranchs and benthic structure, by doing more survey attempts like the one presented here. 3.5 References English, S., Wilkinson, C. and Baker, V. (eds). 1994. Survey Manual for Tropical Resources.
Australian Institute of Marine Science. Townsville. Fowler, J., Cohen, L. and Jarvis, P. 1998. Practical Statistics for Field Biology. John Wiley
& Sons. Chichester, New York, Weinheim, Brisbane, Singapore, Toronto. Gauch, H. G. Jr. 1984. Multivariate analysis in community ecology. Cambridge University
Press. Cambridge, London, New York, New Rochelle, Melbourne, Sidney. Marshall, J. G. & Willan, R. C. 1999. Nudibranchs of Heron Island, Great Barrier Reef.
Backhuys Publishers. Leiden. Stanzel, K. B. & Newman, H. 1997. Progress report on the 1996 Marine Survey of the
Tukang Besi (Wakatobi) archipelago, South East Sulawesi, Indonesia. (Internal report: Operation Wallacea).
Ter Braak, C. J. F. 1998. Canoco 4.0 (Computer program). Centre for Biometry Wageningen. Wageningen.
Todd, C. D. 1981. The Ecology of Nudibranch Molluscs. Oceanography and Marine Biology: an Annual Review 19: 141-234.

37
4 Conclusions and recommendations In section 3, I had another hypothesis: Chromodorids have larger home ranges than Phyllidids. The background for this hypothesis was that there was a general field observation among the volunteers that nudibranchs of the family Phyllidiidae were much more stationary than nudibranchs of the family Chromodorididae. Accordingly, I wanted to objectively test this observation. We first tried to use a coordinate system and write down the co-ordinates for every nudibranch we found, hoping to be able to follow their movement. It seemed to work at first but later a storm ruined our efforts by moving the lines a little bit and thus made it impossible to continue using co-ordinates. We then tried to mark the nudibranch position with a string of rope and a buoyant bamboo marker. This also seemed to work at first but we later got problem as the bamboo markers lost their buoyancy and that made it very difficult to find them. It was also difficult to recognise the specific nudibranchs and I believe that we would have needed an identification system in order to successfully follow the nudibranchs. Perhaps a “fish marking” system with tags would work? Another problem is to keep a functioning “dive relay race” going on for a long enough period. When doing my own survey we also discovered how difficult it is to do scientific work underwater. As we had to be several divers sharing the workload we always had to adjust for the least experienced and/or the diver with the highest air consumption. This makes underwater surveys with volunteers extremely difficult due to the differences in diving experience, biological knowledge and air consumption. It takes many dives before you can find a system that is workable. Another problem with working in a remote area like Wakatobi is the lack of equipment and the logistic transport problems when needing repairs or spare parts. This is especially hard since the climate affects and wears down all kinds of equipment, for instance, diving equipment, boats, recording devices and of course computer equipment. There is always the need to come up with new repair ideas, using very simple tools or equipment. We also found out how hard the tropical environment affects the personnel, usually about one third are on the sick list due to infections (stomach illness, ear infections, badly infected wounds, etc). This means that you always have to have a lot more people helping you than you would normally expect. In the future I think it would be worthwhile to do a more structured opisthobranch study at Hoga. One main mistake, when working with the checklist (section 2), is that there has been no registration of search effort. This makes it impossible to correlate the opisthobranch abundance with, for instance, dive time or search area. The easiest way to register the search effort is to keep track of the “active search time” but another idea could be to cooperate with other projects where the search area is defined by transects. In these cases an estimation of search area is given for “free”!

38
5 Acknowledgements I would like thank: Per Milberg, for all the help with this report. - I could not have done it without your help. Lindsay Warren and Krzystof Brzkieta, for trying to help out at Hoga when my appointed (Opwall) supervisor disappeared! All the volunteers at Hoga who graciously tried to help me, especially my dive buddies: Peter Bächi, Rob Saunders, Nikki Rowland, Jenny Hill, Elaine Worley, Gail Young, Sarah Yeates and Tony Gatcombe. Richard Willan, Pam Beesley, The Ray Society, David Behrens and the Linnean Society of London for kind permissions to reproduce appropriate illustrations in my thesis. My parents, for financial and moral support. Åsa Nilsson, my fiancé, for helping me, all the time. – Thank You!

Plate A
Chelidonura amoena Bergh, 1905 Pleurobranchus grandis Pease, 1868 Note: White squid cleaning the opisthobranch!
Cyerce elegans Bergh, 1870 Nembrotha sp (lineolata? Bergh, 1905)
Jorunna funebris (Keelart, 1858) Hexabranchus sanguineus (Rüppell & Leuckart, 1828) Note: The identification details are taken from “The Sea Slug Forum” (www.seaslugforum.net 2001-04-22). Copyright holder and photographer is the author (Lars Karlsson). The photos are all from Hoga Island (1999).

Plate B
Chromodoris annae Bergh, 1877 Chromodoris magnifica (Quoy & Gaimard, 1832)
Chromodoris sp (boucheti? Rudman, 1982) Phyllidiella pustulosa (Cuvier, 1804)
Phyllidia sp (willani? Brunckhorst, 1993) Phyllidiopsis striata Bergh, 1888 Note: The identification details are taken from “The Sea Slug Forum” (www.seaslugforum.net 2001-04-22). Copyright holder and photographer is the author (Lars Karlsson). The photos are all from Hoga Island (1999).

A
ppen
dix
I, 1(
6)
Com
pila
tion
over
Opi
stho
bran
chs
foun
d du
ring
Ope
ratio
n W
alla
cea
surv
ey 1
997-
1999
.
Fam
ilyF
amily
Gen
us
Sp
ecie
sA
uth
ors
O
bse
r-O
cca-
Fir
stF
irst
Las
tO
WN
Rem
arks
"SS
F"
*"C
GB
RO
" **
(If
iden
tifi
ed)
(If
iden
tifi
ed)
vati
on
ssi
on
s97
/98
9999
***
Apl
ysiid
aeA
plys
iidae
Apl
ysia
extra
ordi
naria
Alla
n 19
32
22
9906
1699
06cf
ext
raor
dina
ria. I
n "C
GB
RO
": (
Alla
n, 1
932)
Apl
ysia
sp3
299
0730
9909
2515
6A
plys
iasp
22
9908
1699
0825
172
Apl
ysia
sp1
199
0621
121
Apl
ysiid
aeA
plys
iidae
Dol
abel
laau
ricul
aria
(Lig
htfo
ot, 1
786)
218
9808
0199
0612
9909
28A
plys
iidae
Phy
llapl
ysia
sp. 1
.x
398
9908
2899
0911
184
sp 9
9091
1 ?
Apl
ysiid
aeA
plys
iidae
Sty
loch
eilu
slo
ngic
auda
(Qou
y &
Gai
mar
d, 1
825)
31
9907
3099
0730
In "
CG
BR
O":
(Q
uoy
& G
aim
ard,
182
4)A
plys
iidae
Apl
ysiid
aeS
tylo
chei
lus
stria
ta(Q
ouy
& G
aim
ard,
183
2)42
699
0905
9911
1616
420
+, 9
9110
5A
plys
iid3
399
0610
9908
2511
0(1
16?)
Ord
er =
An
asp
idea
116
Ham
inoe
idae
Ham
inoe
idae
Aty
s cy
lindr
ica
/ cyl
indr
icus
(Hel
blin
g, 1
779)
9809
01cy
lindr
icus
Aty
s cf
. cyl
indr
icus
11
9909
0317
8H
amin
oeid
aeA
tys
debi
lisP
ease
, 186
011
299
1103
9911
05H
amin
oeid
aeH
amin
oeid
aeA
tys
nauc
um(L
inna
eus,
175
8)1
198
0901
9907
17In
"G
BR
O":
(Li
nné,
175
8)A
tys
cf n
aucu
m2
298
0524
9905
2499
0309
32H
amin
oeid
aeH
amin
oeid
aeA
tys
sem
istri
ata
/ sem
istri
atus
Pea
se, 1
860
9805
3033
sem
istr
iatu
sA
tys
cf s
emis
triat
us
11
9907
0814
1(s
emis
tria
ta?)
Aty
s sp
84
9909
0699
1102
182
Bul
lidae
Bul
lidae
Bul
laam
pulla
Linn
aeus
, 175
810
399
1103
9911
16In
"G
BR
O":
Lin
né, 1
758
Bul
lasp
374
9911
0299
1116
202
Bul
lasp
11
9907
3016
6B
ullid
aeB
ulla
vern
icos
aG
ould
, 185
998
0901
Agl
ajid
aeA
glaj
idae
Che
lidon
ura
amoe
naB
ergh
, 190
524
1797
0920
9905
1599
1101
Agl
ajid
aeC
helid
onur
acf
. am
oena
x1
199
1027
200
Agl
ajid
aeA
glaj
idae
Che
lidon
ura
hiru
ndin
ina
(Quo
y &
Gai
mar
d, 1
832)
75
9709
2099
0702
9909
29In
"G
BR
O":
(Q
uoy
& G
aim
ard,
183
3)A
glaj
idae
Agl
ajid
aeC
helid
onur
ain
orna
taB
aba,
194
914
598
1028
9909
1799
1105
Che
lidon
ura
lineo
lata
9805
01 P
roba
bly
Phi
linop
sis
lineo
lata
A
glaj
idae
Che
lidon
ura
sand
rana
Rud
man
, 197
340
398
0519
9907
2599
0929
26C
helid
onur
asp
9805
1928
Che
lidon
ura
sp98
0519
29C
helid
onur
asp
11
9906
1511
3C
helid
onur
asp
11
9906
1511
4C
helid
onur
asp
. 121
499
0511
9909
2989
Agl
ajid
aeC
helid
onur
asp
. 2x
11
9811
2499
0730
73A
glaj
idae
Che
lidon
ura
sp. 3
.x
11
9910
0120
9O
S07
75?
Agl
ajid
aeA
glaj
idae
Che
lidon
ura
tsur
ugen
sis
Bab
a &
Abe
, 195
91
198
0621
9907
2538
Agl
ajid
aeC
helid
onur
ava
rians
Elio
t, 19
035
599
0614
9910
31G
astr
opte
ron
sp.
11
9911
0320
4H
amin
oea
sp4
399
0524
9909
1798
Ham
inoe
asp
11
9905
1997
Ham
inoe
asp
11
9906
2112
2H
amin
oea
sp1
199
0729
152
Ham
inoe
asp
5
22
9906
1699
0621
115
Agl
ajid
aeH
amin
oea
sp. 1
.x
148
9811
2499
0617
9909
3178
See
SS
F u
nder
Ham
inoe
a sp
. 3.
Agl
ajid
aeH
amin
oea
sp. 2
.x
9811
2479
See
SS
F u
nder
Ham
inoe
a sp
. 3.
Ilbid
aeIlb
iam
aria
naH
off &
Car
lsso
n, 1
990
9811
1970
Agl
ajid
aeA
glaj
idae
Odo
ntog
laja
guam
ensi
sR
udm
an, 1
978
9810
01S
mar
agdi
nelli
dae
Sm
arag
dine
llida
eP
hane
ropt
halm
ussm
arag
dinu
s(R
uppe
ll &
Leu
ckar
t, 18
28)
854
9809
0799
0611
9911
1452
Som
bano
lake
(83
).In
"G
BR
O"
(Rüp
pell
& L
euck
art,
1830
)A
glaj
idae
Agl
ajid
aeP
hilin
opsi
sga
rdin
eri
(Elio
t, 19
03)
11
9709
2099
0528
Agl
ajid
aeA
glaj
idae
Phi
linop
sis
pils
bryi
(Elio
t, 19
00)
22
9806
2099
0728
9907
15P
hilin
opsi
ssp
.1
199
0524
95P
hilin
opsi
ssp
.1
199
0617
116
Gas
trop
terid
aeG
astr
opte
ridae
Sag
amin
opte
ron
psyc
hede
licum
Car
lsso
n &
Hof
f, 19
749
897
0920
9906
0499
1101

A
ppen
dix
I, 2(
6)
Com
pila
tion
over
Opi
stho
bran
chs
foun
d du
ring
Ope
ratio
n W
alla
cea
surv
ey 1
997-
1999
.S
ipho
pter
onbr
uneo
mar
gina
tum
11
9811
1799
0621
In "
WB
" sp
ecie
s 18
. Car
lsso
n &
Hof
f, 19
74C
helid
onur
idsp
31
9905
1188
Phi
linop
sis/
Che
lidon
ura?
Ord
er =
Cep
hal
asp
idea
316
Dia
phan
idae
Dia
phan
idae
Col
poda
spis
thom
pson
iB
row
n, 1
979
22
9906
0399
0622
107
Cep
hala
spid
ea in
SS
F b
ut "
unre
solv
ed"
in C
LO
rder
= C
eph
alas
pid
ea ?
2P
leur
obra
nchi
dae
Ple
urob
ranc
hida
eB
erth
ella
m
arte
nsi
(Pils
bry,
189
6)4
397
0920
9907
0999
1023
Ple
urob
ranc
hida
eP
leur
obra
nchi
dae
Ple
urob
ranc
hus
fors
kalii
(Rup
pel &
Leu
ckar
t, 18
28)
2011
9709
2099
0613
9911
17(f
orsk
ali)
In "
GB
RO
": R
üppe
ll &
Leu
ckar
t, 18
28P
leur
obra
nchi
dae
Ple
urob
ranc
hus
gran
dis
Pea
se, 1
868
165
9806
2099
0527
9911
20B
uoy
3 -1
5 sp
ecim
ens
Ple
urob
ranc
hida
eP
leur
obra
nchi
dae
Ple
urob
ranc
hus
pero
niC
uvie
r, 1
804
11
9908
24P
leur
obra
nchu
ssp
.1
199
0525
96O
rder
= N
ota
spid
ea42
Aeo
lidiid
aeA
eolid
iella
alba
Ris
bec,
192
84
498
1119
9906
2999
0826
Aeo
lidie
llasp
11
9908
0316
0A
eolid
iidae
Cer
beril
laan
nula
ta(Q
uoy
& G
aim
ard,
183
2)2
298
0908
9907
0899
0725
53an
nula
ta ?
Cut
hona
cf a
nnul
ata
11
9908
0616
5C
. ann
ulat
a in
"G
" sp
ecie
s 23
2. (
Bab
a, 1
949)
Gla
ucid
aeF
acel
inid
aeC
rate
nalin
eata
(Elio
t, 19
05)
11
9905
1187
In "
GB
RO
": (
Elio
t, 19
04)
Cra
tena
sp1
199
0925
192
Cut
hona
sp.
9803
257
Eub
ranc
hus
?sp
11
9908
1817
3G
lauc
idae
Fac
elin
idae
Fav
orin
usja
poni
cus
Bab
a, 1
949
11
9908
11(ja
poni
cus?
)G
lauc
idae
Fac
elin
idae
Fav
orin
usts
urug
anus
Bab
a &
Abe
, 196
43
398
1027
9908
1199
0926
Fla
belli
nida
eF
labe
llini
dae
Fla
belli
nabi
colo
r(K
elaa
rt, 1
858)
11
9908
16F
labe
llina
delic
ata
(Gos
liner
& W
illan
, 199
1)98
1114
In G
oslin
er, 1
987
(for
mer
Cor
yphe
lla s
p)F
labe
llini
dae
Fla
belli
nida
eF
labe
llina
exop
tata
Gos
liner
& W
illan
, 199
110
898
0315
9906
0599
1117
Fla
belli
naor
nata
9810
27F
labe
llini
dae
Fla
belli
nariw
oG
oslin
er &
Will
an, 1
991
43
9906
1799
1115
118
Fla
belli
nida
eF
labe
llini
dae
Fla
belli
naru
brol
inea
ta(O
´Don
oghu
e, 1
929)
11
9906
0914
8&
OW
N 1
48F
labe
llini
dae
Fla
belli
nasp
. 1.
x1
198
1112
9906
0567
Aeo
lidiid
aeLi
men
andr
ano
dosa
Hae
felfi
nger
& S
tam
m, 1
958
11
9811
2499
0906
77G
lauc
idae
Fac
elin
idae
Mor
idill
abr
ocki
Ber
gh, 1
888
9803
256
Gla
ucid
aeF
acel
inid
aeP
hidi
ana
indi
ca(B
ergh
, 189
6)17
1698
1019
9905
1599
1116
61(C
alor
ia)
Gla
ucid
aeF
acel
inid
aeP
hyllo
desm
ium
bria
reum
(Ber
gh, 1
896)
398
2798
0916
9905
1799
1121
(bria
reus
)G
lauc
idae
Fac
elin
idae
Phy
llode
smiu
mlo
ngic
irrum
(Ber
gh, 1
905)
11
9709
2099
0721
Phy
llode
smiu
msp
33
9906
2399
0816
124
sim
ilar
to O
WN
85
Phy
llode
smiu
msp
9709
2085
Aeo
lidiid
aeP
rota
eolid
iella
atra
Bab
a, 1
955
9811
0465
Gla
ucid
aeF
acel
inid
aeP
tera
eolid
iaia
nthi
na(A
ngas
, 186
4)25
1997
0920
9905
0599
1111
Aeo
lid98
1202
82A
eolid
9812
0283
Aeo
lid1
199
0617
117
Aeo
lid1
199
0630
126
Aeo
lid2
199
0630
127
Aeo
lid1
199
0617
129
Aeo
lid1
199
0816
174
Su
bo
rder
Aeo
lidin
aA
rmin
idae
Der
mat
obra
nchu
scf
. gon
atop
hora
x97
0920
D. g
onat
opho
ra in
"D
" 19
96 p
295
.D
erm
atob
ranc
hus
sp3
398
0907
9905
1599
0917
51D
erm
atob
ranc
hus
sp20
1399
0603
9906
1510
1D
erm
atob
ranc
hus
sp2
199
1111
206
Der
mat
obra
nchu
ssp
9811
1368
Der
mat
obra
nchu
ssp
11
9905
1490
Der
mat
obra
nchu
ssp
11
9906
2111
9D
erm
atob
ranc
hus
sp1
199
0805
165
Der
mat
obra
nchu
ssp
11
9910
2219
8D
erm
atob
ranc
hus
sp1
199
0624
cf O
WN
101

A
ppen
dix
I, 3(
6)
Com
pila
tion
over
Opi
stho
bran
chs
foun
d du
ring
Ope
ratio
n W
alla
cea
surv
ey 1
997-
1999
.A
rmin
idae
Der
mat
obra
nchu
ssp
. 2.
x98
1124
74S
ub
ord
er A
rmin
ina
Bor
nelli
dae
Bor
nelli
dae
Bor
nella
angu
illa
John
son,
198
32
297
0920
9905
1599
0517
Bor
nelli
dae
Bor
nelli
dae
Bor
nella
stel
lifer
(Ada
ms
& R
eeve
, 184
8)33
599
1010
9911
24In
"G
BR
O":
(A
. Ada
ms
& R
eeve
in A
. Ada
ms,
184
8)D
endr
onot
idae
Den
dron
otus
spx
33
9709
2099
0610
9908
2516
8T
riton
idae
Mar
ioni
asp
.x
9803
2513
1O
WN
13/1
Mar
ioni
avi
rides
cens
22
9805
0199
0607
9907
03T
ethy
dida
eM
elib
een
geli
Ris
bec,
193
71
199
1102
203
Tet
hydi
dae
Mel
ibe
fimbr
iata
Ald
er &
Han
cock
, 186
43
299
0903
9909
15S
cylla
eida
eS
cylla
eida
eS
cylla
eape
lagi
ca(L
inne
aeus
, 175
8)1
199
0730
In "
GB
RO
": L
inne
, 175
8Tr
itoni
asp
.98
0325
12Tr
itoni
asp
.1
199
0708
137
Trit
oniid
aeT
riton
iops
is
alba
(Bab
a, 1
949)
11
9811
0399
0816
(Trit
onop
silla
)(T
riton
opsi
llaau
stra
lis)
9811
21P
roba
bly
a m
isst
ake!
Su
bo
rder
Den
dro
no
tin
aC
hrom
odor
idid
aeC
hrom
odor
idid
aeA
rdea
doris
egre
ttaR
udm
an, 1
984
87
9805
0199
0603
9911
20D
orid
idae
Ast
eron
otus
cesp
itosu
s(H
asse
lt, 1
824)
11
9709
2099
0621
Chr
omod
orid
idae
Cer
atos
oma
mia
mira
na(B
ergh
, 187
5)2
299
0717
?14
6C
hrom
odor
idid
aeC
erat
osom
asi
nuat
a(v
an H
asse
lt, 1
824)
11
9911
05E
arlie
r: M
iam
ira s
inua
taC
hrom
odor
idid
aeC
erat
osom
ate
nue
Abr
aham
, 187
61
199
0824
Chr
omod
orid
idae
Chr
omod
orid
idae
Cer
atos
oma
trilo
batu
m(J
. E. G
ray,
182
7)98
0901
In "
GB
RO
": (
Gra
y, 1
827)
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
albo
punc
tata
(Gar
ret,
1879
)1
199
0731
157
Chr
omod
oris
aliu
s1
199
0914
In "
WB
" sp
ecie
s 14
7. R
udm
an 1
987
Chr
omod
orid
idae
Chr
omod
oris
anna
eB
ergh
, 187
721
510
997
0920
9905
0599
1117
Chr
omod
orid
idae
Chr
omod
oris
bouc
heti
Rud
man
, 198
28
698
0501
9907
0399
1001
(bou
chet
ti)C
hrom
odor
idid
aeC
hrom
odor
idid
aeC
hrom
odor
isco
i(R
isbe
c, 1
956)
3530
9709
2099
0524
9911
05C
hrom
odor
idid
aeC
hrom
odor
isdi
anae
Gos
liner
& B
ehre
ns, 1
998
106
6097
0920
9905
1499
1117
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
elis
abet
hina
Ber
gh, 1
877
98
9709
2099
0524
9909
10(e
lizab
ethi
na)
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
fidel
is(K
elaa
rt, 1
858)
33
9905
2199
0722
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
geom
etric
aR
isbe
c, 1
928
4940
9709
2099
0603
9911
20C
hrom
odor
idid
aeC
hrom
odor
idid
aeC
hrom
odor
ishi
ntua
nens
is97
0920
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
kuite
riR
udm
an, 1
982
11
9905
22ku
iteri
? O
S 0
184
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
kuni
eiP
ruvo
t-F
ol, 1
930
2217
9709
2099
0518
9911
09C
hrom
odor
idid
aeC
hrom
odor
idid
aeC
hrom
odor
isle
opar
dus
Rud
man
, 198
713
1197
0920
9906
0999
1104
Chr
omod
orid
idae
Chr
omod
orid
idae
Chr
omod
oris
loch
iR
udm
an, 1
982
132
8197
0920
9905
0599
1117
Chr
omod
oris
cf. l
ochi
11
9906
2312
3C
hrom
odor
idid
aeC
hrom
odor
idid
aeC
hrom
odor
ism
agni
fica
(Quo
y &
Gai
mar
d, 1
832)
119
7197
0920
9905
2099
1117
Chr
omod
orid
idae
Chr
omod
oris
prec
iosa
(Kel
aart
, 185
8)1
199
0704
Chr
omod
oris
quad
ricol
our
9808
01In
Deb
eliu
s 19
96 p
200
Chr
omod
oris
retic
ulat
a2
298
0315
9911
0199
1111
In "
GB
W"
spec
ies
577
(Quo
y &
Gai
mar
d, 1
832)
(=t
inct
oria
)C
hrom
odor
issp
32
9907
1199
1007
142
Chr
omod
oris
sp98
1126
81C
hrom
odor
issp
11
9906
0310
0C
hrom
odor
issp
11
9907
1514
5C
hrom
odor
issp
11
9907
2414
9C
hrom
odor
idid
aeC
hrom
odor
idid
aeC
hrom
odor
isst
rigat
aR
udm
an, 1
982
54
9803
1899
0802
9911
172
Chr
omod
orid
idae
Chr
omod
oris
verr
ieri
(Cro
sse,
187
5)2
298
0831
9907
0499
0730
48ve
rrie
ri? a
lbon
ares
Chr
omod
orid
idae
Chr
omod
oris
will
ani
Rud
man
, 198
210
663
9804
0199
0505
9911
16D
endr
odor
idid
aeD
endr
odor
isca
rbun
colo
sa(Q
ouy
& G
aim
ard,
183
2)1
199
0906
179
Den
drod
orid
idae
Den
dodo
ridid
aeD
endr
odor
isni
gra
(Stim
pson
, 185
5)98
0801
Den
drod
oris
cf. n
igra
11
9910
1819
6D
endr
odor
idid
aeD
endo
dorid
idae
Den
drod
oris
tube
rcul
osa
(Quo
y &
Gai
mar
d, 1
832)
9804
04(D
endr
ois)
Den
drod
oris
?98
1216
86D
endr
odor
is ?
11
9911
0120
1D
iaph
orod
oris
mits
ui(B
aba,
193
8)98
1119
69In
Gos
liner
,198
7 (f
orm
er D
. Sp)
Dor
idid
aeD
isco
doris
boho
liens
isB
ergh
, 187
71
199
0728

A
ppen
dix
I, 4(
6)
Com
pila
tion
over
Opi
stho
bran
chs
foun
d du
ring
Ope
ratio
n W
alla
cea
surv
ey 1
997-
1999
.D
orid
idae
Dis
codo
ris?
litur
ata
Ber
gh, 1
905
21
9808
1599
0721
45(D
isco
doris
? li
tura
ta)
Dor
idid
aeD
oris
?sp
. 10.
x2
299
0702
9907
1713
1D
oris
sp.
10
Chr
omod
orid
idae
Dur
ville
doris
pusi
llax
11
9907
1814
7C
hrom
odor
idid
aeD
urvi
lledo
rissi
mila
risx
11
9908
1116
9Fr
yeria
men
indi
eB
runc
khor
st, 1
993
87
9709
2099
0603
9909
01(m
eini
ndae
)Fr
yeria
ruep
pelii
Ber
gh, 1
869
11
9906
21(c
f Fry
eria
rup
peli)
Glo
ssod
oris
albo
cinc
ta98
0620
Chr
omod
orid
idae
Chr
omod
orid
idae
Glo
ssod
oris
cinc
ta(B
ergh
, 188
8)98
0620
Chr
omod
orid
idae
Chr
omod
orid
idae
Glo
ssod
oris
hiku
eren
sis
(Pru
vot-
Fol
, 195
4)2
298
1024
9906
0599
0806
Chr
omod
orid
idae
Chr
omod
orid
idae
Glo
ssod
oris
rufo
mar
gina
ta(B
ergh
, 189
0)98
1216
Gym
nodo
ridid
aeG
ymno
doris
alba
(Ber
gh, 1
877)
11
9909
11G
ymno
dorid
idae
Gym
nodo
ridid
aeG
ymno
doris
ceyl
onic
a(K
elaa
rt, 1
858)
53
9911
0299
1105
In "
SS
F"
auth
or n
ot g
iven
.G
ymno
doris
sp.
11
9906
2812
5G
ymno
doris
sp.
11
9907
0413
3G
ymno
doris
sp.
11
9908
0315
8G
ymno
doris
sp.
11
9908
2517
6G
ymno
doris
sp.
11
9909
1418
6G
ymno
doris
sp.
11
9911
0320
5G
ymno
doris
sp.
11
9911
2020
8G
ymno
dorid
idae
Gym
nodo
rissp
. 1.
x98
1016
58D
orid
idae
Dor
idid
aeH
alge
rda
carls
oni /
aff.
car
lson
iR
udm
an, 1
978
11
9709
2099
1111
carls
oni
Dor
idid
aeH
alge
rda
mal
esso
Car
lson
& H
off,
1993
22
9806
2099
1001
9910
14as
in D
ebel
ius
1996
=H
alge
rda
sp. 1
.H
alge
rda
cf. m
ales
so1
199
0605
104
Hal
gerd
asp
9709
20H
exab
ranc
hida
eH
exab
ranc
hida
eH
exab
ranc
hus
sang
uine
us(R
uppe
ll &
Leu
ckar
t, 18
30)
55
9810
0199
0527
9911
1619
7In
"G
BR
O":
(R
üppe
ll &
Leu
ckar
t, 18
30)
Chr
omod
orid
idae
Chr
omod
orid
idae
Hyp
selo
doris
bullo
cki
(Col
lingw
ood,
188
1)18
1397
0920
9905
2499
0906
135
smal
l pin
k va
riety
+ p
urpl
e an
d w
hite
var
iety
Chr
omod
orid
idae
Chr
omod
orid
idae
Hyp
selo
doris
emm
a / e
mm
aeR
udm
an, 1
977
9709
20em
ma
Chr
omod
orid
idae
Hyp
selo
doris
infu
cata
(Rup
pel &
Leu
ckar
t, 18
28)
11
9810
1699
0825
57H
ypse
lodo
riskr
akat
oaG
oslin
er &
Joh
nsso
n, 1
999
11
9907
2815
4In
ZJL
S 1
25:1
-114
Chr
omod
orid
idae
Chr
omod
orid
idae
Hyp
selo
doris
mac
ulos
a(P
ease
, 187
1)5
599
0520
9909
15C
hrom
odor
idid
aeH
ypse
lodo
riscf
. mac
ulos
ax
11
9907
0713
4C
hrom
odor
idid
aeH
ypse
lodo
rispu
rpur
eom
acul
osa
Ham
atan
i, 19
951
198
0421
9908
0920
Chr
omod
orid
idae
Hyp
selo
doris
rudm
ani
x1
199
0910
183
Hyp
selo
doris
sp.
11
9908
1117
0C
hrom
odor
idid
aeH
ypse
lodo
rissp
. 1.
x98
1103
66C
hrom
odor
idid
aeH
ypse
lodo
risw
hite
i(A
dam
s &
Ree
ve, 1
850)
11
9908
10w
hite
i ex
mou
aci,
OS
062
3D
orid
idae
Dor
idid
aeJo
runn
afu
nebr
is(K
elaa
rt, 1
858)
2926
9709
2099
0513
9911
18Jo
runn
asp
.1
199
1026
199
Pol
ycer
idae
Nem
brot
hach
ambe
rlain
iG
oslin
er &
Beh
rens
, 199
797
0920
Pol
ycer
idae
Pol
ycer
idae
Nem
brot
hacr
ista
taB
ergh
, 187
710
9709
2099
0525
9910
26P
olyc
erid
aeP
olyc
erid
aeN
embr
otha
kuba
ryan
aB
ergh
, 187
736
9709
2099
0524
9911
11P
olyc
erid
aeP
olyc
erid
aeN
embr
otha
lineo
lata
Ber
gh, 1
905
497
0920
9910
1599
1117
Pol
ycer
idae
Nem
brot
haru
tilan
s(P
ruvo
t-F
ol, 1
931)
297
0920
9911
0999
1116
Nem
brot
hasp
. 97
0920
Pol
ycer
idae
Nem
brot
hasp
. 4.
x1
9811
2476
Aeg
iritid
aeA
egiri
dae
Not
odor
isga
rdin
eri
Elio
t, 19
0398
0915
Aeg
iritid
aeA
egiri
dae
Not
odor
ism
inor
Elio
t, 19
0498
0324
Not
odor
isse
rena
e1
9709
2099
0606
In "
GB
W"
spec
ies
610.
(G
oslin
er &
Beh
rens
199
8)C
hrom
odor
idid
aeC
hrom
odor
idid
aeN
oum
eacr
ocea
Rud
man
, 198
697
0920
Chr
omod
orid
idae
Nou
mea
varia
ns
(Pea
se, 1
871)
11
9908
1617
1va
rians
/ no
rba
Chr
omod
orid
idae
Nou
mea
sp
. 2.
x2
299
0630
9907
3012
8G
onio
dorid
idae
Oke
nia
sp.
x97
0920
(=H
opki
nsia
)P
ecte
nodo
risau
rora
9810
2964
(Mik
e M
iller
s S
lug
Site
)C
hrom
odor
idid
aeC
hrom
odor
idid
aeP
ecte
nodo
ristri
linea
ta(A
. Ada
ms
& R
eeve
185
0)2
298
0706
9906
0699
0806
103
& O
WN
103,
OW
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
aba
bai
Bru
nckh
orst
, 199
33
297
0920
9907
2799
0802
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
aco
eles
tisB
ergh
, 190
511
382
9709
2099
0514
9911
17

A
ppen
dix
I, 5(
6)
Com
pila
tion
over
Opi
stho
bran
chs
foun
d du
ring
Ope
ratio
n W
alla
cea
surv
ey 1
997-
1999
.P
hylli
dia
sp. c
f. co
eles
tis1
199
0807
167
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
ael
egan
sB
ergh
, 186
916
411
597
0920
9905
1999
1117
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
aex
quis
itaB
runc
khor
st, 1
993
98
9709
2099
0721
9911
05P
hylli
diid
aeP
hylli
diid
aeP
hylli
dia
ocel
lata
Cuv
ier,
180
419
1997
0920
9906
2499
1105
Phy
llidi
asp
33
9907
0999
0812
140
Phy
llidi
asp
11
9906
2313
0P
hylli
dia
sp1
199
0708
136
Phy
llidi
asp
66
9907
1299
1115
138
9907
08 +
1 ?
Phy
llidi
asp
11
9907
0913
9P
hylli
dia
sp1
199
0713
143
Phy
llidi
asp
11
9907
1414
4P
hylli
dia
sp1
199
0729
153
Phy
llidi
idae
Phy
llidi
atu
laM
arcu
s &
Mar
cus,
197
01
199
0707
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
ava
ricos
aLa
mar
ck, 1
801
131
9697
0920
9905
1599
1117
Phy
llidi
idae
Phy
llidi
aw
illan
iB
runc
khor
st, 1
993
76
9810
1799
0628
9911
0959
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
ella
nigr
a(v
an H
asse
lt, 1
824)
154
9809
0199
0515
9909
17In
"G
BR
O":
(H
asse
lt, 1
824)
. Lah
oa C
hann
elP
hylli
diid
aeP
hylli
diid
aeP
hylli
diel
lapu
stul
osa
(Cuv
ier,
180
4)60
428
997
0920
9905
0599
1124
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
ella
rudm
ani
Bru
nckh
orst
, 199
330
2498
1002
9906
1999
1028
Phy
llidi
idae
Phy
llidi
ella
zeyl
anic
a(K
elaa
rt, 1
859)
108
9906
2299
1112
Phy
llidi
idae
Phy
llidi
opsi
san
nae
Bru
nckh
orst
, 199
32
299
0802
9908
05P
hylli
diid
aeP
hylli
diop
sis
daut
zenb
ergi
(Vay
ssie
re, 1
911)
11
9910
1819
5P
hylli
diid
aeP
hylli
diop
sis
fissu
ratu
sB
runc
khor
st, 1
993
11
9909
3019
3P
hylli
diid
aeP
hylli
diid
aeP
hylli
diop
sis
krem
pfi
Pru
vot-
Fol
, 195
72
299
0917
9911
0118
5P
hylli
diid
aeP
hylli
diid
aeP
hylli
diop
sis
pipe
kiB
runc
khor
st, 1
993
5852
9709
2099
0519
9911
24P
hylli
diid
aeP
hylli
diid
aeP
hylli
diop
sis
shire
enae
Bru
nckh
orst
, 199
312
1197
0920
9906
2699
1113
(shi
rinae
)P
hylli
diid
aeP
hylli
diop
sis
sphi
ngis
Bru
nckh
orst
, 199
31
199
0828
Phy
llidi
idae
Phy
llidi
idae
Phy
llidi
opsi
sst
riata
Ber
gh, 1
888
3428
9709
2099
0604
9911
18D
orid
idae
Pla
tydo
risfo
rmos
a(A
lder
& H
anco
ck, 1
864)
52
9905
2099
1103
Pla
tydo
rissc
abra
(Cuv
ier,
180
4)7
499
0716
9910
27P
laty
doris
sp2
199
0820
190
Pol
ycer
idae
Trio
phid
aeP
loca
mop
heru
sce
ylon
ica
/ cey
loni
cus
(Kel
aart
, 185
8)3
299
0906
9909
2518
1ce
ylon
icus
Pol
ycer
idae
Pol
ycer
asp
. 3.
x1
199
0920
188
Pol
ycer
a ?
9812
0284
Phy
llidi
idae
Phy
llidi
idae
Ret
icul
idia
fung
iaB
runc
khor
st &
Gos
liner
, 199
315
1497
0920
9906
1999
1118
In "
GB
RO
": B
runc
khor
st &
Gos
liner
in B
runc
khor
st, 1
993
Phy
llidi
idae
Phy
llidi
idae
Ret
icul
idia
halg
erda
Bru
nckh
orst
& B
urn,
199
01
197
0920
9906
09In
"G
BR
O":
Bru
nckh
orst
& G
oslin
er in
Bru
nckh
orst
, 199
3C
hrom
odor
idid
aeR
isbe
cia
impe
rialis
(Pea
se, 1
860)
9809
19C
hrom
odor
idid
aeC
hrom
odor
idid
aeR
isbe
cia
tryo
ni(G
arre
tt, 1
873)
97
9803
1999
0609
9911
13P
olyc
erid
aeR
oboa
stra
grac
ilis
(Ber
gh, 1
877)
1616
9709
2099
0515
9911
12D
orid
idae
Seb
ador
isnu
bilo
saP
ease
, 189
11
199
0521
94P
olyc
erid
aeP
olyc
erid
aeT
ambj
am
oros
a(B
ergh
, 187
7)97
0920
In "
SS
F"
auth
or n
ot g
iven
.P
olyc
erid
aeT
ambj
asp
.x
9709
20P
olyc
erid
aeT
heca
cera
paci
fica
(Ber
gh, 1
883)
9709
20P
olyc
erid
aeP
olyc
erid
aeT
heca
cera
pict
aB
aba,
197
22
297
0920
9910
2299
1111
Pol
ycer
idae
The
cace
rasp
x97
0920
Chr
omod
orid
idae
Tho
runn
aau
stra
lis(R
isbe
c, 1
928)
9811
21C
hrom
odor
idid
aeC
hrom
odor
idid
aeT
horu
nna
furti
vaB
ergh
, 187
813
898
1119
9906
0399
1116
71T
rapa
nia
sp.
9805
2630
Gon
iodo
ridid
aeT
rapa
nia
sp. 2
.x
9811
2475
Gon
iodo
ridid
aeG
onod
orid
idae
Tra
pani
ato
ddi
Rud
man
, 198
71
199
0617
Tra
pani
a ?
11
9907
0413
2T
rapa
nia
?1
199
0728
151
Dor
idid
aeD
orid
idae
Trip
pa
inte
cta
(Kel
aart
, 185
8b)
22
9809
0999
0613
9911
16T
rippa
sp
32
9906
1499
0903
112
Trip
pa
sp98
0530
34C
hrom
odor
id98
0624
39C
hrom
odor
id1
199
0515
91

A
ppen
dix
I, 6(
6)
Com
pila
tion
over
Opi
stho
bran
chs
foun
d du
ring
Ope
ratio
n W
alla
cea
surv
ey 1
997-
1999
.S
ub
ord
er D
ori
din
aO
rder
= N
ud
ibra
nch
ia28
88(A
eolid
ina
= 48
3, A
rmin
ina
= 30
, Den
dro
no
tin
a =
47, D
ori
din
a =
2328
)
Cos
tasi
ella
sp1
198
0621
9906
2112
0C
osta
siel
lidae
Cos
tasi
ella
sp. 1
.x
9811
2480
Cos
tasi
ellid
aeC
osta
siel
la ?
sp. 2
x1
199
0518
93C
alip
hylli
dae
Pol
ybra
nchi
idae
Cye
rce
eleg
ans
Ber
gh, 1
870
11
9911
1720
7In
"G
BR
O":
Ber
gh, 1
871
Ely
sia
bryc
ei3
299
0716
9911
01In
"W
B"
spec
ies
70. (
Jens
en &
Wel
ls, 1
990)
+ "
D"
1996
Ely
siid
aeP
lako
bran
chiid
aeE
lysi
aor
nata
(Sw
ains
on, 1
840)
54
9808
0199
0617
9909
10E
lysi
idae
Ely
sia
pilo
sa ?
x98
1019
62E
lysi
asp
11
9810
2999
0804
63E
lysi
asp
1512
9905
1599
1117
99E
lysi
asp
32
9906
499
0615
105
Ely
sia
sp2
299
0616
9906
1711
1E
lysi
asp
11
9907
2815
0cf
filic
auda
Ely
sia
sp.
9810
0554
Ely
sia
sp.
11
9908
0715
9E
lysi
idae
Ely
sia
sp. 1
.x
9810
0855
Ely
siid
aeE
lysi
asp
. 8.
x1
199
0906
180
Ely
siid
aeE
lysi
avi
ridis
(Mon
tagu
, 180
4)98
0620
Not
hern
spe
cies
!?E
lysi
idae
Pla
kobr
anch
iidae
Ely
siel
lapu
silla
Ber
gh, 1
872
42
9906
0999
0701
Pla
kobr
anch
ussp
494
9808
2599
0524
9911
1047
Lim
apon
tiida
eS
tilig
ercr
emon
iana
(Trin
ches
e, 1
893)
52
9909
2599
0925
191
Ely
siid
aeP
lako
bran
chiid
aeT
hurid
illa
baye
ri(M
arcu
s, 1
965)
3625
9803
1599
0515
9911
15In
"G
BR
O":
(E
r. M
arcu
s, 1
965)
Pla
kobr
anch
iidae
Thu
ridill
aca
rlson
iG
oslin
er, 1
995
11
9809
0799
0910
Ely
siid
aeP
lako
bran
chiid
aeT
hurid
illa
hoffa
eG
oslin
er, 1
995
1311
9810
2499
0603
9909
17ho
ffae
(livi
da ?
)T
hurid
illa
kath
ae5
398
0507
9907
2199
0824
25T
hurid
illa
lineo
lata
43
9806
2099
0512
9909
14In
"D
" 19
96 p
169
.T
hurid
illa
sp4
499
0517
9906
2192
Thu
ridill
asp
22
9906
0999
0610
109
Thu
ridill
aun
dula
11
9811
1699
0805
In "
GB
W"
spec
ies
550.
Gos
liner
199
5O
rder
= S
aco
glo
ssa
159
* =
Fam
ily d
esig
natio
n ac
cord
ing
to "
Sea
Slu
g F
orum
"35
23**
= F
amily
des
igna
tion
acco
rdin
g to
"C
heck
list o
f the
Gre
at B
arrie
r R
eef O
phis
tobr
anch
ia"
(in M
& W
, 199
9)
***=
Ope
ratio
n W
alla
cea
Nud
ibra
nchi
a (I
nter
nal n
umbe
r)

App
endi
x II,
1(4
)
S
earc
h re
cord
s fr
om "
Spa
tial d
istr
ibut
ion
of n
udib
ranc
hs a
ccor
ding
to b
enth
ic s
truc
ture
"
Bu
oy
3 "+
3m"
(Lat
itu
de:
S 0
5 28
386
, Lo
ng
itu
de:
E 1
23 4
5 43
9) S
earc
har
ea 1
50 s
qm
Mo
rnin
g d
ives
("A
M")
Aft
ern
oo
n d
ives
("P
M")
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
08LK
/PB
/JH
204,
599
1108
RS
/NR
/SY
244,
6S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rC
hrom
odor
is e
lisab
ethi
na25
6,6
300
290
CA
Chr
omod
oris
ann
ae40
719
064
SP
SO
rang
eC
hrom
odor
is m
agni
fica
506,
143
023
0D
CA
Chr
omod
oris
ann
ae36
719
064
SP
SO
rang
eJo
runn
a fu
nebr
is20
4,6
1480
-190
SP
LP
hylli
diel
la p
ustu
losa
205
680
95S
PE
Gre
yJo
runn
a fu
nebr
is30
4,6
1480
-190
SP
LP
hylli
diel
la p
ustu
losa
205
680
95S
PE
Gre
yD
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1109
LK/P
B27
4,7
9911
09LK
/PB
244,
5S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rD
ista
nce
(cm
)N
embr
otha
kub
arya
na56
5,8
1080
100
Sea
cuc
umbe
r!C
hrom
odor
is a
nnae
256,
419
05
AA
0Jo
runn
a fu
nebr
is35
5,2
1490
-50
SP
Chr
omod
oris
ann
ae25
6,4
190
5A
A0
Joru
nna
fune
bris
205,
214
90-5
0S
PC
hrom
odor
is m
agni
fica
607,
220
01
SP
0C
hrom
odor
is e
lisab
ethi
na28
6,1
1640
56C
EB
row
nC
hrom
odor
is m
agni
fica
404,
242
010
She
ll5
NP
hylli
dia
varic
osa
354,
824
60-5
6C
AP
ink
Joru
nna
fune
bris
305,
114
8511
SP
9 N
Joru
nna
fune
bris
205,
114
8511
SP
9 N
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
10R
S/J
H/S
Y/E
W40
9911
10R
S/J
H/S
Y/E
W27
4,9
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Chr
omod
oris
ann
ae30
5,9
130
18C
AP
ink
18 N
EC
hrom
odor
is m
agni
fica
354,
242
010
SP
T40
WC
hrom
odor
is e
lisab
ethi
na40
5,8
130
17D
C22
NP
hylli
diel
la p
ustu
losa
255,
475
020
OT
29 S
EC
hrom
odor
is m
agni
fica
354,
242
010
35 S
WJo
runn
a fu
nebr
is30
5,4
1485
11C
EB
row
n26
NW
Phy
llidi
ella
pus
tulo
sa25
5,1
750
20S
PS
Ora
nge
7 S
WJo
runn
a fu
nebr
is20
5,4
1485
11C
EB
row
n26
NW
Joru
nna
fune
bris
305,
114
8511
CE
Bro
wn
26 N
WP
hylli
dia
varic
osa
306,
422
5023
7 N
EJo
runn
a fu
nebr
is20
5,1
1485
11C
EB
row
n26
NW
Phy
llidi
a va
ricos
a30
5,5
2250
23S
8 N
Joru
nna
fune
bris
604,
529
5022
RC
K9
ND
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1111
LK/P
B/G
Y/T
G20
3,8
9911
11LK
/PB
/GY
/TG
284,
8S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)C
hrom
odor
is a
nnae
407,
16
73D
C10
NE
Chr
omod
oris
mag
nific
a35
520
032
DC
10 N
Cho
mod
oris
elis
abet
hina
304,
911
017
MA
70 N
EP
hylli
diel
la p
ustu
losa
595
012
R10
NE
Chr
omod
oris
mag
nific
a30
3,1
380
8037
NW
Joru
nna
fune
bris
4520
602
CE
0C
hrom
odor
is m
agni
fica
453,
867
069
60 N
WP
hylli
dia
varic
osa
2250
239
NW
Phy
llidi
a co
eles
tis30
523
0023
DC
8 E
Phy
llidi
ella
pus
tulo
sa60
6,4
2980
3610
SW
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
12R
S/J
H/S
Y/E
W28
5,7
9911
12R
S/J
H/S
Y/E
W32
4,9
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Chr
omod
oris
ann
ae40
7,2
073
MA
Bro
wn
42 E
Chr
omod
oris
ann
ae40
7,6
5C
S22
0 W
Chr
omod
oris
ann
ae40
6,7
240
5C
SB
row
n23
0 S
WC
hrom
odor
is a
nnae
438,
35
DC
214
SW
Chr
omod
oris
mag
nific
a3,
844
010
SP
GG
rey
67 S
Chr
omod
oris
mag
nific
a15
4,4
80C
T90
WP
hylli
diel
la p
ustu
losa
284,
667
072
SP
66 N
Chr
omod
oris
mag
nific
a44
4,4
10C
T62
EP
hylli
diel
la p
ustu
losa
304,
769
020
DC
20 W
Phy
llidi
ella
pus
tulo
sa28
5,2
72/6
9C
A94
NE
Chr
omod
oris
dia
nae
355
36M
A15
NE
Chr
omod
oris
ann
ae40
6,7
17T
A13
0 E
Phy
llidi
ella
pus
tulo
sa38
238
NC
hrom
odor
is a
nnae
406,
718
TA
130
EC
hrom
odor
is a
nnae
258
73C
SB
row
n80
EC
hrom
odor
is d
iana
e36
70 N
E

App
endi
x II,
2(4
)
S
earc
h re
cord
s fr
om "
Spa
tial d
istr
ibut
ion
of n
udib
ranc
hs a
ccor
ding
to b
enth
ic s
truc
ture
"
Bu
oy
3 "+
12m
" (L
atit
ud
e: S
05
28 3
86, L
on
git
ud
e: E
123
45
439)
Sea
rch
area
150
sq
mM
orn
ing
div
es (
"AM
")A
fter
no
on
div
es (
"PM
")D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1108
LK/P
B/J
H37
11,8
9911
08R
S/N
R/S
Y56
12,1
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rS
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Ret
icul
idia
fung
ia20
1313
011
0P
hylli
diel
la p
ustu
losa
3013
,517
580
RC
KJo
runn
a fu
nebr
is40
9,3
532
-315
MA
Bro
wn
Phy
llidi
a co
eles
tis55
15,6
340
137
CA
Pur
ple
Nem
brot
ha k
ubar
yana
2511
,976
5-3
3M
AC
hrom
odor
is g
eom
etric
a13
13,8
460
74R
CK
Chr
omod
oris
leop
ardu
s25
10,3
1000
-190
SP
EP
urpl
eR
etic
ulid
ia fu
ngia
2012
,556
012
0C
hrom
odor
is d
iana
e30
13,5
1050
160
CA
Ret
icul
idia
fung
ia5
12,5
560
120
Chr
omod
oris
mag
nific
a40
11,9
1105
0Li
neN
embr
otha
kub
arya
na20
12,8
885
-40
SC
Joru
nna
fune
bris
7011
,718
6040
CA
Chr
omod
oris
leop
ardu
s25
11,3
1000
-150
SP
EP
urpl
eC
hrom
odor
is m
agni
fica
459
2060
-325
CA
Pin
kN
embr
otha
kub
arya
na11
,329
8010
TA
Red
Phy
llidi
opsi
s pi
peki
2010
,430
00-8
0C
AP
ink
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
09LK
/PB
3512
,299
1109
LK/P
B25
12S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rD
ista
nce
(cm
)P
hylli
dia
varic
osa
6015
,845
025
0A
EG
rey/
Gre
enC
hrom
odor
is le
opar
dus
2511
900
7C
A11
NE
Chr
omod
oris
leop
ardu
s25
10,9
880
-190
TA
Pur
ple
Chr
omod
oris
mag
nific
a35
13,5
1800
6M
AR
ed0
Phy
llidi
opsi
s pi
peki
5010
,918
00-1
20C
AP
hylli
diop
sis
pipe
ki50
1119
208
CA
7 N
EP
hylli
dia
coel
estis
2412
,723
0065
SP
EP
hylli
diop
sis
pipe
ki15
10,4
3000
9C
A7
WP
hylli
diop
sis
pipe
ki15
10,4
3000
-50
RC
KD
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1110
RS
/JH
/SY
/EW
4099
1110
RS
/JH
/SY
/EW
2813
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Nem
brot
ha k
ubar
yana
3513
,513
527
AA
11 S
EN
embr
otha
kub
arya
na25
14,2
135
27A
A11
SE
Phy
llidi
ella
pus
tulo
sa20
1417
019
CA
6 S
WC
hrom
odor
is le
opar
dus
3011
,590
07
SP
E10
NE
Phy
llidi
ella
pus
tulo
sa17
1130
09
RC
K7
WC
hrom
odor
is m
agni
fica
4013
1795
6R
CK
95 E
Chr
omod
oris
leop
ardu
s30
1190
07
SP
EP
urpl
e13
EC
hrom
odor
is b
ouch
eti
4512
,613
4516
SP
EG
rey
4 W
Chr
omod
oris
mag
nific
a40
1317
956
RC
K90
ED
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1111
LK/P
B/G
Y/T
G15
11,7
9911
11LK
/PB
/GY
/TG
2212
,7S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)P
hylli
diel
la p
ustu
losa
2010
.00
33A
A14
NE
Phy
llidi
ella
pus
tulo
sa20
1017
019
1 S
EP
hylli
diel
la p
ustu
losa
2512
,717
019
CA
5 S
WC
hrom
odor
is le
opar
dus
1511
900
7C
A1
EC
hrom
odor
is b
ouch
eti
2012
,113
4516
SP
E30
NC
hrom
odor
is a
nnae
2011
1340
16S
PE
18 N
Chr
omod
oris
mag
nific
a40
1216
4017
1 S
Joru
nna
fune
bris
7012
,523
8027
SP
MD
ark
Chr
omod
oris
bou
chet
i15
1228
8035
SP
13 N
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
12R
S/J
H/S
Y/E
W21
1299
1112
RS
/JH
/SY
/EW
2613
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Phy
llidi
ella
pus
tulo
sa12
,817
019
CA
6 S
EP
hylli
diel
la p
ustu
losa
2813
,817
019
DC
50 S
WC
hrom
odor
is e
lisbe
thin
a13
12,2
1346
16D
C56
SE
Chr
omod
oris
leop
ardu
s11
,490
07
SP
E35
WC
hrom
odor
is e
lisbe
thin
a26
1315
8034
CA
42 S
EC
hrom
odor
is a
nnae
13,2
16H
YY
ello
w69
SE
Joru
nna
fune
bris
4512
2400
37A
A85
NE
Chr
omod
oris
elis
beth
ina
13,4
1615
34S
PG
59 S
EC
hrom
odor
is m
agni
fica
3011
1800
6M
A21
7 E
Joru
nna
fune
bris
3718
9 N
EC
hrom
odor
is d
iana
e32
143
268
NE
Chr
omod
oris
mag
nific
a30
623
0 E

App
endi
x II,
3(4
)
S
earc
h re
cord
s fr
om "
Spa
tial d
istr
ibut
ion
of n
udib
ranc
hs a
ccor
ding
to b
enth
ic s
truc
ture
"
Sam
pel
a "+
3m"
(Lat
itu
de
S 0
5 29
041
, Lo
ng
itu
de
E 1
23 4
5 27
1) S
earc
har
ea 1
50 s
qm
Mo
rnin
g d
ives
("A
M")
Aft
ern
oo
n d
ives
("P
M")
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
) L
ine
dep
th (
m)
9911
08R
S/N
R/S
Y39
5,7
9911
08LK
/PB
/JH
376,
9S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rP
hylli
diel
la p
ustu
losa
103,
915
0-1
80R
Phy
llidi
a va
ricos
a20
6,3
80-1
86D
CP
hylli
diop
sis
pipe
ki25
4,8
330
-120
RP
hylli
diel
la p
ustu
losa
206,
634
5-1
00C
AP
ink
Phy
llidi
ella
pus
tulo
sa25
4,3
900
-240
CA
Pin
kC
hrom
odor
is a
nnea
157,
112
45-3
1S
PE
Gre
yP
hylli
diel
la p
ustu
losa
205,
612
20-4
0S
PB
Pin
kP
hylli
diel
la p
ustu
losa
359
2390
140
CA
Bro
wn
Chr
omod
oris
ann
ea20
612
280
SP
EG
rey
Joru
nna
fune
bris
558,
724
8010
0C
EG
reen
Phy
llidi
ella
pus
tulo
sa34
824
1545
RC
KJo
runn
a fu
nebr
is40
8,7
2480
100
SP
EG
rey
Joru
nna
fune
bris
708,
324
1570
SP
ER
edP
hylli
diel
la p
ustu
losa
257,
129
10-5
0C
AJo
runn
a fu
nebr
is50
8,3
2415
70S
PE
Red
Phy
llidi
ella
pus
tulo
sa35
8,6
2950
80C
AP
ink
Phy
llidi
opsi
s pi
peki
308,
629
5080
CA
Pin
kD
ate
Div
ers
Sea
rch
time
(min
) L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1109
RS
/JH
/SY
2899
1109
RS
/JH
/SY
25S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rP
hylli
diel
la p
ustu
losa
206,
831
0-7
5S
PE
Y
ello
wP
hylli
dia
varic
osa
409,
195
017
0R
CK
Chr
omod
oris
ann
ea15
7,2
1220
-32
AE
Phy
llidi
opsi
s pi
peki
158,
619
7014
0R
CK
Joru
nna
fune
bris
507,
724
5010
0C
MJo
runn
a fu
nebr
is50
7,7
2450
100
CM
Joru
nna
fune
bris
407,
724
5010
0C
MJo
runn
a fu
nebr
is40
7,7
2450
100
CM
Phy
llidi
ella
pus
tulo
sa26
7,5
2880
75P
hylli
diel
la p
ustu
losa
257,
829
20-4
0S
PS
Ora
nge
Phy
llidi
ella
pus
tulo
sa40
7,5
2965
50S
PD
ate
Div
ers
Sea
rch
time
(min
) L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1110
LK/P
B/G
Y28
5,3
9911
10LK
/PB
/GY
/TG
236,
8S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)P
hylli
dia
varic
osa
357,
710
0046
SP
EG
rey
5 S
Phy
llidi
a co
eles
tis18
6,3
630
3927
SP
hylli
diel
la p
ustu
losa
258,
121
8047
SP
M
Yel
low
7 S
EP
hylli
dia
varic
osa
359
1000
46C
A26
NW
Joru
nna
fune
bris
707,
324
7048
CM
41 N
WP
hylli
diel
la p
ustu
losa
309,
610
0051
SP
DB
lack
14 S
WJo
runn
a fu
nebr
is40
7,3
2470
48C
A28
NE
Pte
reoa
lidia
iant
hina
356,
112
0026
SP
EG
rey
14 S
W?
257
2680
38D
C?
Phy
llidi
opsi
s pi
peki
256,
729
4049
CA
Pin
k12
SD
ate
Div
ers
Sea
rch
time
(min
) L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1111
RS
/JH
/SY
405,
699
1111
RS
/JH
/SY
316,
8S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)P
hylli
dia
coel
estis
204,
411
060
OT
40 N
EP
hylli
dia
coel
estis
6,5
610
39S
P24
SP
hylli
dia
coel
estis
115,
762
039
SP
24 S
Phy
llidi
a va
ricos
a10
0046
50 W
Phy
llidi
a va
ricos
a40
7,9
1000
46A
A38
WC
hrom
odor
is a
nnea
116,
511
7743
SP
EW
hite
11 S
WC
hrom
odor
is a
nnea
116,
511
7743
MA
11 S
WP
hylli
diop
sis
pipe
ki30
6,9
2120
74S
PE
Ora
nge
36 N
EP
hylli
diel
la p
ustu
losa
258
2180
4710
SE
Phy
llidi
ella
pus
tulo
sa21
8047
15 W
Chr
omod
oris
geo
met
rica
306,
323
6041
SP
EB
row
n4
WC
hrom
odor
is g
eom
etric
a30
7,3
2360
41C
E16
EP
hylli
diel
la p
ustu
losa
407,
924
6024
CM
18 E
Phy
llidi
ella
pus
tulo
sa24
6024
18 E
Joru
nna
fune
bris
7024
7048
53 S
WJo
runn
a fu
nebr
is24
7048
18 N
Joru
nna
fune
bris
4024
7048
42 N
WJo
runn
a fu
nebr
is24
7048
36 N
WP
hylli
diop
sis
pipe
ki30
5,7
2720
74C
A12
SW
Phy
llidi
ella
pus
tulo
sa22
3000
4210
EP
hylli
diop
sis
pipe
ki25
2940
4927
SP
hylli
diel
la p
ustu
losa
1630
0042
10 E
Phy
llidi
ella
pus
tulo
sa60
3000
49?
RC
K8
SE
The
cace
ra p
icta
To "
labo
rato
ry"
SP
Phy
llidi
ella
pus
tulo
sa22
3000
49?
RC
K8
SE
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
) L
ine
dep
th (
m)
9911
12LK
/PB
/GY
/TG
255,
299
1112
LK/P
B/G
Y/T
G23
6,8
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Chr
omod
oris
ann
ea25
611
3043
SP
E60
SW
Chr
omod
oris
ann
ea20
711
3043
SP
EG
rey
60 S
EP
hylli
diel
la p
ustu
losa
2530
24S
PE
10 E
Phy
llidi
ella
pus
tulo
sa25
6,9
3000
42C
A6
EP
hylli
diel
la p
ustu
losa
205,
326
8074
SP
E7
NW
Phy
llidi
ella
pus
tulo
sa20
6,9
3000
42C
A6
EP
hylli
diel
la p
ustu
losa
255,
630
0042
8 S
EP
hylli
diel
la p
ustu
losa
358
2530
24D
C31
SE
Phy
llidi
ella
pus
tulo
sa25
5,6
3000
428
SE
Phy
llidi
ella
pus
tulo
sa21
8047
CA
16 E
Joru
nna
fune
bris
2470
4810
5 N
WJo
runn
a fu
nebr
is45
825
3048
40 W
Joru
nna
fune
bris
40 W
Joru
nna
fune
bris
825
3048
40 W
Phy
llidi
a co
eles
tis15
6,5
620
39C
A40
SE
Phy
llidi
ella
pus
tulo
sa27
2074
CA
8 N
WP
hylli
diel
la p
ustu
losa
1000
4695
WP
hylli
diel
la p
ustu
losa
46C
E97
W

App
endi
x II,
4(4
)
S
earc
h re
cord
s fr
om "
Spa
tial d
istr
ibut
ion
of n
udib
ranc
hs a
ccor
ding
to b
enth
ic s
truc
ture
"
Sam
pel
a "+
12m
" (L
atit
ud
e S
05
29 0
41, L
on
git
ud
e E
123
45
271)
Sea
rch
area
150
sq
mM
orn
ing
div
es (
"AM
")A
fter
no
on
div
es (
"PM
")D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1108
RS
/NR
/SY
4412
,799
1108
LK/P
B/J
H32
13,6
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rS
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Phy
llidi
ella
pus
tulo
sa14
11,9
250
10S
Phy
llidi
ella
pus
tulo
sa30
13,7
100
30S
PE
Gre
yP
hylli
diel
la p
ustu
losa
4013
,332
516
7C
AP
hylli
diel
la p
ustu
losa
3014
,823
011
0S
PM
B
row
n/Y
ello
wP
hylli
dia
coel
estis
3011
,664
2-8
5S
PD
Yel
low
Phy
llidi
ella
pus
tulo
sa30
14,5
310
80S
Phy
llidi
a va
ricos
a17
14,2
670
110
SP
MB
row
n/ye
llow
Phy
llidi
a co
eles
tis35
13,2
625
-140
SP
TY
ello
wP
hylli
diel
la p
ustu
losa
4010
1270
-270
CA
Pin
kP
hylli
diop
sis
stria
ta10
15,1
1410
150
CA
Phy
llidi
opsi
s st
riata
1214
1470
170
CA
Pur
ple
Phy
llidi
a va
ricos
a25
14,6
1595
-70
DC
Phy
llidi
ella
pus
tulo
sa28
12,8
2450
0C
AP
ink
Phy
llidi
ella
pus
tulo
sa20
11,5
1730
-280
SP
CO
rang
eP
hylli
diel
la p
ustu
losa
2013
,322
50-7
0C
AD
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)99
1109
RS
/JH
/SY
4999
1109
RS
/JH
/SY
52S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)V
-C (
cm)
Su
rfac
eC
olo
ur
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
V-C
(cm
)S
urf
ace
Co
lou
rP
hylli
diel
la p
ustu
losa
3312
,80
42S
Phy
llidi
a sp
2614
030
SP
hylli
diel
la p
ustu
losa
2513
,128
080
RC
KP
hylli
dia
varic
osa
409,
115
017
9R
CK
Phy
llidi
a sp
2613
,341
090
AE
Phy
llidi
ella
pus
tulo
sa25
13,7
270
120
RP
hylli
dia
coel
estis
3011
,862
0-1
60S
PE
Dar
k re
dP
hylli
diel
la p
ustu
losa
3014
,556
580
SP
hylli
dia
coel
estis
1113
,171
093
SP
Bro
wn
Phy
llidi
a co
eles
tis30
13,2
620
-160
SP
E"D
ark"
Phy
llidi
a co
eles
tis30
14,4
1695
-130
SP
hylli
dia
coel
estis
1113
,871
093
SP
Bro
wn
Chr
omod
oris
coi
2614
2255
133
SP
hylli
diop
sis
stria
ta12
14,5
1340
-60
DC
Phy
llidi
ella
pus
tulo
sa30
12,6
2422
-130
SP
SO
rang
eP
hylli
diel
la p
ustu
losa
1812
,628
20-2
90C
AP
ink
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
10LK
/PB
/GY
1612
,299
1110
LK/P
B/G
Y/T
G16
13,3
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Phy
llidi
ella
pus
tulo
sa35
11,9
2450
40C
A3
NN
o O
pist
hobr
anch
s fo
und
Phy
llidi
ella
pus
tulo
sa24
7031
26 S
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
11R
S/J
H/S
Y30
12,1
9911
11R
S/J
H/S
Y32
13,4
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Sp
ecie
s:S
ize
(mm
)D
epth
(m
)H
-C (
cm)
Mar
ker
Su
rfac
eC
olo
ur
Dis
tan
ce (
cm)
Phy
llidi
opsi
s st
riata
1210
,624
052
SP
SO
rang
e3
EP
hylli
diop
sis
stria
ta12
12,1
240
52O
rang
e2
SE
Phy
llidi
a co
eles
tis12
1469
044
SP
Yel
low
13P
hylli
diel
la p
ustu
losa
2513
765
6516
SE
P.s
?30
11,9
1190
67D
C60
SP
hylli
dia
coel
estis
1214
689
44S
PB
Yel
low
13 N
EP
hylli
diop
sis
stria
ta15
11,4
1325
53S
PE
Ora
nge
12 N
Joru
nna
fune
bris
814
,612
9012
SP
B5E
Chr
omod
oris
coi
5013
,316
2025
AA
10 N
EP
hylli
dia
varic
osa
1815
1890
68M
A16
WP
hylli
diel
la p
ustu
losa
3012
,424
3031
OT
60 E
Phy
llidi
ella
pus
tulo
sa22
1524
2031
OT
8 E
Dat
eD
iver
sS
earc
htim
e (m
in)
Lin
e d
epth
(m
)D
ate
Div
ers
Sea
rch
time
(min
)L
ine
dep
th (
m)
9911
12LK
/PB
/GY
/TG
2811
,799
1112
LK/P
B/G
Y/T
G23
13S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)S
pec
ies:
Siz
e (m
m)
Dep
th (
m)
H-C
(cm
)M
arke
rS
urf
ace
Co
lou
rD
ista
nce
(cm
)C
hrom
odor
is c
oi35
1015
3025
CA
Ora
nge
100
NP
hylli
dia
coel
estis
1814
,569
044
SP
DG
rey
12 S
WJo
runn
a fu
nebr
is12
Chr
omod
oris
coi
3512
1395
12?
SP
E76
S o
f tap
eJo
runn
a fu
nebr
is12
Chr
omod
oris
coi
3512
1620
2520
0 N
EP
hylli
diop
sis
stria
ta10
1112
0053
AE
44 N
WC
hrom
odor
is k
unei
6013
,518
9068
40 S
EP
hylli
diel
la p
ustu
losa
2012
,324
2031
/40
SP
ER
ed6
NC
hrom
odor
is k
unei
5013
,518
9068
40 S
EP
hylli
diop
sis
stria
ta10
12,8
1325
53C
A39
NW
Phy
llidi
ella
pus
tulo
sa25
13,5
2420
31/4
0C
A5
N