overexploitation of coastal resources at bajamar-jatay
TRANSCRIPT

Pacific Coast Archaeological Society Quarterly, Volume 54, Numbers 3 and 4
Overexploitation of Coastal Resources at Bajamar-Jatay? Size Composition of the Mollusks Consumed Prehistorically in Baja California
Enah M. Fonseca Ibarra
Abstract
During the prehistory of Bajamar-Jatay, Baja California, the capture of mollusks was intensive and concentrated on the species Haliotis cracherodii (black abalone), Haliotis fulgens (green abalone), Lottia gigantea (owl limpet), Mytilus californianus (California mussel) and Septifer bifurcatus (bifurcate mussel). The pressure on these food resources may have triggered their overexploitation, w ccord-ing to our working hypothesis, would be reflected in a decrease in size over time. Nonparametric tests were performed to look for significant differences between the study periods. Toward the end of the Archaic period (3500–1500 BP), the smallest size ranges and a decrease in average size for all species were identified, with the exception of the abalone H. cracherodii. In the Late Prehistoric peri-od (1500–140 BP), an increase in the maximum lengths for M. cal-ifornianus and S. bifurcatus was observed. The size of L. gigantea also increased in the Late Prehistoric period but did not reach the average sizes recorded in the Archaic period (8000–3500 BP). Contrary to what was expected, a simple pattern of overexploitation was not found. Alternative explanations for changes or stability in shell size that were considered include selective harvesting, focus on alternative resources, reduction of intraspecific competition, and environmental disturbances.
Introduction
Shells are one of the most important resources of the archaeological record in coastal camps, so their anal-ysis is ideal for inferring patterns of consumption, cap-ture strategies, and seasonality in resource use of over time (Thomas 2015a, 2015b). Another line of research has used the shells as a proxy to evaluate the impact of collecting, based on variations in sizes for the consumed species (Anderson 1981; Jerardino 1997, 2010; Mannino and Thomas 2002; Braje et al. 2007,
2012; Erlandson et al. 2008, 2011, 2015; Jerardino et al. 2008; Rick and Erlandson 2008; Rivadeneira et al. 2009; Erlandson and Rick 2010; Olguín et al. 2015).
Archaeological evidence for prey size increase, accred-ited to human overharvesting, is limited. Nevertheless, studies around the world have documented evidence of economic intensification associated with an increase in shellfish size (Giovas et al. 2010, 2013; Poteate et al. 2015; Thakar 2011; Olguín et al. 2015). It is also considered that due to the pressure on some species, other taxa were incorporated into the diet, perhaps as a resource management strategy (Quitmyer and Reitz 2006; Whitaker 2008; Erlandson and Rick 2010).
The shell midden archeological sites located on Baja California’s northwest coast are characterized by the high density of marine and terrestrial resources. How-ever, the shell middens are generally dominated by rel-atively few species of mollusks, fish, marine mammals, and shorebirds (Guía 2005a, 2005b, 2010; Ovilla and García 2007, 2008; Celis 2011; Robles 2013).
For the groups of hunter-gatherer-fishers who exploit-ed the coastal environments of Bajamar-Jatay, the collection of mollusks seems to have been a major activity that went back 5,000 years (Guía 2008; Drakíc 2009). Given the great density, relatively good state of preservation, and age of the faunal assemblages recovered in this area, it has been possible to carry out

PCAS Quarterly 54(3&4)
Fonseca 58
Bajamar-Jatay is an area of great importance for the study of the hunter-gatherer-fisher groups that inhab-ited Baja California, due to the high concentration, good state of preservation, and diversity of recorded archaeological sites (Serrano 1992, 1993; Hernández and Schoenberg 1993a, 1993b; Reina 1994, 1995; Drakíc and Cuadra 2005; Drakíc et al. 2007; Ovilla and García 2007, 2008; Rojas 2009; Drakíc and Delgado 2010; García 2013; Ovilla 2013). Investigating this area from 2012 to 2014, the project “Study of Camps on the Coastline and Intermontane Valleys of Baja California” under the direction of the Instituto Nacional de Antro-pología e Historia (INAH) was focused on carrying out extensive excavations at eight shell midden camps, designated U4b, U4c, U4e, H2A, H2B, CT41d, MP3g, and MP3c (Fonseca 2013a, 2014, 2015, 2016).
Chronology
Excavation of the discussed archaeological sites was done in metric levels of 10 cm until the parent rock or sterile stratum (absence of cultural objects) was reached. The excavated areas and number of levels were not uniform, because they depended on each site’s characteristics. The degree of bioturbation in the area is high, and there was no clear stratigraphic dif-ferentiation; therefore, the malacological analysis was made from the excavation levels. Despite the disad-vantages represented by the use of metric levels within stratigraphic layers (Harris 1991), in this investigation it permitted the data’s normalization for making com-parisons and having greater control of the materials, both in the field and in the laboratory. In what follows we will use the word “stratum” or “occupation” indif-ferently to refer to an excavation level.
Although there are eight sites, some may have been occupied uninterruptedly and others intermittently, by the same or by different prehistoric groups. In order to have a comparable unit of measurement and to avoid the risk of confusing occupations from different time periods, it was decided to treat the strata as if there
an analysis of the size composition of the main mol-lusks consumed on Baja California’s northwest coast. In this article we present the results of the variation of malacological material sizes from eight archaeological sites with occupations ranging from 4908 to 486 BP. Changes in targeted species are discussed as a pos-sible strategy for the management of marine-coastal resources.
Materials and Methods
Study Area
The archaeological sites addressed in the present study are located within the Bajamar Tourist Com-plex in Ensenada, Baja California, 63 km south of the U.S. border (Figure 1). The Baja California peninsula’s Pacific coast is one of the areas world-wide with the highest primary productivity per unit area (Hernández-Ayón et al. 2007), due, in large part, to the upwelling events controlled by the California Current System. With respect to biological frontiers, the study area is located within the Californian prov-ince, which extends from Point Conception in the U.S. to Punta Eugenia in Baja California Sur (Hubbs 1960; Briggs 1974). This region is characterized by its temperate subtropical waters, with temperature records of up to 21° C (70° F) in summer and min-imums of 10° C (50° F) in winter (McLean 1978). The habitats that occur in the coastal region of this province are rocky intertidal, debris reef, sandy beach, bay and lagoon, rocky subtidal, and muddy and sandy bottom (McLean 1978).
Bajamar’s beach is part of the stretch of rocky coast-line formed by extrusive igneous rock, with contribu-tions of andesite at the base extending for more than 20 km from the beach of La Salina to the beach at Salsipuedes. The coast is highly exposed to surf, with an orientation to the southwest and an intertidal zone that extends for more than 40 m through a steep slope of more than 30° (Ramírez 2009).

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 59
were 34 different occupations. For example, in site H2B, six 2-by-2 m squares were excavated until they reached 40 cm in depth. In this way, all the material of level 1 was called H2B I and so on to H2B IV.
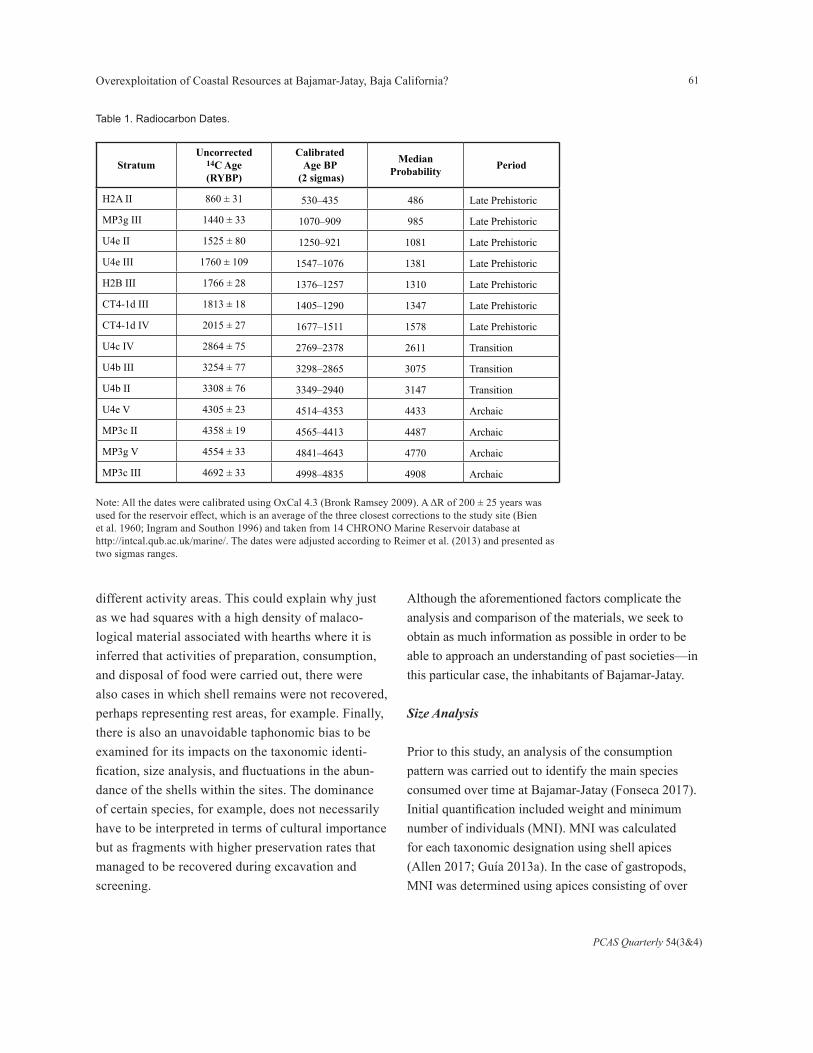
The chronological order of the strata was based on radiocarbon dating of 14 shells of the species H. cracherodii. From the dated strata, the rest of the stratigraphic layers were classified. Despite the evident postdepositional mixing (rodent burrows), no inverted radiocarbon dates were obtained except in the case of strata U4b II and U4b III. However, the
difference between them is ~50 years, and both corre-spond to the same chronological period (Transition), so it does not affect the results of the analysis.
Of the 34 initial strata, it was necessary to discard seven that did not have an absolute dating or whose assign-ment to a period was not possible, even after being ordered according to the stratigraphic sequence. That is, for example, strata H2B I and H2B II were not dated, but we know that they correspond to the Late Prehistor-ic period because they are later than stratum H2B III, whose adjusted age range was 1376–1257 BP.
Figure 1. Location of the study area, a region known as Bajamar-Jatay, located on the northwest coast of Baja California. The archaeological sites located inside the Bajamar Tourist Complex that were excavated in the 2012, 2013, and 2014 field seasons are indicated.

PCAS Quarterly 54(3&4)
Fonseca 60
According to Laylander (1987b), the history of human occupation on the northern peninsula of Baja Califor-nia prior to the Spaniards’ arrival can be divided into three periods: Paleoindigenous (12,000–8000 BP), Archaic (8000–1500 BP), and Late Prehistoric (1500 BP–18th century AD). However, toward the end of the Archaic period, between 3000 and 2000 BP, different cultural practices begin to be observed (Rogers 1945; Moriarty 1966; Porcayo 2007), probably associated with a phase of transition between the occupation of La Jolla groups in the Archaic period and the entry of the Yuman groups that would characterize the Late Prehistoric period.
In order to identify this transitional occupation, des-ignated by Rogers (1945) as the preceramic Yuman phase, or Yuman I, and by Moriarty (1966) as Dieg-ueño I, the occupations of this study were classified into three groups: six strata corresponding to the Archaic period (8000–3500 BP), five in the Transition (3500–1500 BP), and 16 occupations during the Late Prehistoric period (1500–140 BP).
We calibrated and modeled all dates in OxCal 4.3 (Bronk Ramsey 2009) using the most recent marine calibration curve, Marine13 (Reimer et al. 2013). A ΔR of 200 ± 25 years was used for the reservoir effect, which is an average of the three corrections closest to the study site (Bien et al. 1960; Ingram and Southon 1996) and taken from 14 CHRONO Marine Reser-voir database at http://intcal.qub.ac.uk/marine/. Table 1 provides the two-sigma calibrated ranges for each date and the modeled age for each period. Dates were placed in an ordered sequence and modeled within three phases, Archaic, Transition and Late Prehistoric (Figure 2).
Malacological Material
To carry out the present analysis, the malacological material recovered during the excavations in the 2012, 2013, and 2014 field seasons of the project “Study of
Camps on the Coastline and Intermontane Valleys of Baja California” was included (Fonseca 2013a, 2013b, 2014, 2016). The malacological material recovered in 2012 and 2013 was analyzed by the biologist Andrea Guía Ramírez, in charge of the Archaeozoology-Pa-leontology Section of Centro INAH Baja California. To obtain comparable results, the 2014 material was cleaned, separated, and identified taxonomically fol-lowing the previously used methods (Guía 2013a).
The number of squares excavated per site was not uni-form; the number of squares with analyzed malacolog-ical content also differed. The main difference lies in the decrease of material toward the deeper layers. This not only happened with mollusks but in general with all the archaeological material.
The archaeological sites were selected for their pres-ervation status and the density of materials identified on the surface. The excavation grid was drawn based on the shape and extent of the shell midden on the surface, but the camps’ characteristics may have varied over time. Greater density of materials corre-sponding to the oldest occupations might exist outside the excavated areas, and consequently we would have a decrease of materials toward the deepest excavation levels. It should also be considered that the intensity of the occupation—the number of people and the length of their stay—may have been different, and this would affect the quantity and distribution of materials. Even though mixing from the upper layers by postdeposi-tional bioturbation is a possibility, independent occu-pations, probably of less intensity, seem to us the most reasonable explanation for this phenomenon because the samples taken for dating were found associated with heated stone features, oval structures —possible foundations for small dwellings—or in association with lithic materials in situ. The dating of the rest of the strata will corroborate this hypothesis.
In each archaeological site, different activities may have been carried out in specific places, resulting in
PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 61
StratumUncorrected
14C Age (RYBP)
Calibrated Age BP
(2 sigmas)
Median Probability Period
H2A II 860 ± 31 530–435 486 Late Prehistoric
MP3g III 1440 ± 33 1070–909 985 Late Prehistoric
U4e II 1525 ± 80 1250–921 1081 Late Prehistoric
U4e III 1760 ± 109 1547–1076 1381 Late Prehistoric
H2B III 1766 ± 28 1376–1257 1310 Late Prehistoric
CT4-1d III 1813 ± 18 1405–1290 1347 Late Prehistoric
CT4-1d IV 2015 ± 27 1677–1511 1578 Late Prehistoric
U4c IV 2864 ± 75 2769–2378 2611 Transition
U4b III 3254 ± 77 3298–2865 3075 Transition
U4b II 3308 ± 76 3349–2940 3147 Transition
U4e V 4305 ± 23 4514–4353 4433 Archaic
MP3c II 4358 ± 19 4565–4413 4487 Archaic
MP3g V 4554 ± 33 4841–4643 4770 Archaic
MP3c III 4692 ± 33 4998–4835 4908 Archaic
Note: All the dates were calibrated using OxCal 4.3 (Bronk Ramsey 2009). A ΔR of 200 ± 25 years was used for the reservoir effect, which is an average of the three closest corrections to the study site (Bien et al. 1960; Ingram and Southon 1996) and taken from 14 CHRONO Marine Reservoir database at http://intcal.qub.ac.uk/marine/. The dates were adjusted according to Reimer et al. (2013) and presented as two sigmas ranges.
Table 1. Radiocarbon Dates.
different activity areas. This could explain why just as we had squares with a high density of malaco-logical material associated with hearths where it is inferred that activities of preparation, consumption, and disposal of food were carried out, there were also cases in which shell remains were not recovered, perhaps representing rest areas, for example. Finally, there is also an unavoidable taphonomic bias to be examined for its impacts on the taxonomic identi-fication, size analysis, and fluctuations in the abun-dance of the shells within the sites. The dominance of certain species, for example, does not necessarily have to be interpreted in terms of cultural importance but as fragments with higher preservation rates that managed to be recovered during excavation and screening.
Although the aforementioned factors complicate the analysis and comparison of the materials, we seek to obtain as much information as possible in order to be able to approach an understanding of past societies—in this particular case, the inhabitants of Bajamar-Jatay.
Size Analysis
Prior to this study, an analysis of the consumption pattern was carried out to identify the main species consumed over time at Bajamar-Jatay (Fonseca 2017). Initial quantification included weight and minimum number of individuals (MNI). MNI was calculated for each taxonomic designation using shell apices (Allen 2017; Guía 2013a). In the case of gastropods, MNI was determined using apices consisting of over

PCAS Quarterly 54(3&4)
Fonseca 62
50 percent to eliminate any potential of counting an individual more than once. Bivalve remains contain-ing intact umbos (i.e., over 50 percent) were sided (left or right) and counted, and MNI was calculated using whichever side contained higher values for each site. In some cases, a single element indicative of an uncommon species was counted as an individual,
whether it was a whole shell or a single fragment, as there was no possibility of overrepresentation in these instances.
As a result of the analysis of the consumption pattern, we identified Haliotis cracherodii (black abalone), Haliotis fulgens (green abalone), Lottia gigantea (owl
Figure 2. Results of OxCal 4.3 radiocarbon dates modeling (Bronk Ramsey 2009). The dates were adjusted according to the Marine13 calibration curve (Reimer et al. 2013) and presented as two sigma ranges.

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 63
limpet), Mytilus californianus (California mussel), and Septifer bifurcatus (bifurcate mussel) as the main species consumed over time at Bajamar-Jatay (Fonse-ca 2017).
The specimens were measured along their antero-pos-terior axis with a precision of 5 mm, and the mini-mum, maximum, and average values of each species are presented (Tables 2–4). In the case of bivalves, the MNI was used. We entered our measurements for 22,926 shells into Excel files and calculated the means, ranges, and standard deviations for each assemblage.
In most cases, specimens that were complete or sufficiently complete were selected to record their maximum length. Due to the high degree of fragmen-tation presented by the mussel M. californianus, when the specimens were fractured, the size was estimated from a measurement of the umbo toward the end of the ligament, which corresponds approximately to half the maximum length of the whole specimen (Andrea Guía, personal communication 2015). Since the mea-surements of the specimens from the 2012 and 2013 seasons were calculated with this methodology (Guía 2013a, 2015), for comparative purposes the same procedure was used with the materials from the 2014 season.
Recently, Singh and McKechnie (2015) developed a model to obtain the total length of fragmented M. californianus and found that the estimate based on the measurement of the umbo to the ligament offers the most reliable results (R2 = 0.93). The proposed equation (total length = 1.8074 * [measurement of the umbo at the end of the ligament] + 4.947 mm) yields results within the precision range (5 mm) that was used in this investigation.1
To test the statistical significance of changes in mean shell lengths for each stratum, the compo-nents were grouped into three temporal subgroups
corresponding to: Archaic (8000–3500 BP), Transi-tion (3500–1500 BP), and Late Prehistoric periods (1500–140 BP). The statistical analysis compared the direct shell averages assigned to each stratum (Ta-bles 2–4). The Shapiro-Wilk test for normality, Q-Q plots for outliers, and the Levene tests for homoge-neity were conducted. Then the groups were com-pared using a non-parametric Kruskal-Wallis one-way analysis of variance, followed by Games-How-ell post-hoc analysis to determine if the differences between each temporal period were statistically significant. Data were statistically evaluated using SPSS 17.0 software.
Results
Tables 2–4 and Figures 3–6 summarize and graphi-cally illustrate descriptive statistics for mean, median, standard deviations, and length ranges for 22,838 shells recovered from the Bajamar-Jatay area. The di-rect shell average by period indicates that there is not any homogeneous trend over time (Table 5).
The results of the analysis indicate that H. crachero-dii was collected in a wide range of sizes (5–145 mm) during the Archaic period, with an average size of 47.03 mm. For the Transition, the average size is 61.69 mm, and the size range is from 15 to 115 mm. For the Late Prehistoric period, the average size is 54.64 mm, and the range is from 5 to 125 mm (Table 5; Figure 3).
H. fulgens is poorly represented for the Archaic peri-od; it was found in relatively low abundance, and the specimens were very fragmented. The average size is 90.0 mm, and the range is from 35 to 125 mm. For the Transition, the average size is 79.73 mm, and the range is from 25 to 125 mm, and for the Late Prehis-toric period, the average size is 82.56 mm, and the range is from 2 to 145 mm. For this last period, there are also strata in which no shells of this species were recorded or where there were fewer specimens that

PCAS Quarterly 54(3&4)
Fonseca 64
Haliotis cracherodii Haliotis fulgens
Stratum Period N Max Min Mean SD N Max Min Mean SD
MP3c IV Archaic 72 145 5 49.03 34.39 2 95 75 85 10
MP3c III Archaic 248 125 5 49.88 27.56 1 125 125 125 0
MP3c II Archaic 678 125 15 44.07 22.42 1 95 95 95 0
MP3g VI Archaic 5 65 15 43 16
MP3g V Archaic 11 95 45 72.27 16.56
U4e V Archaic 29 95 55 78.1 13.42 4 115 35 82.5 29.47
U4b III Transition 78 115 25 72.05 25.53 26 125 35 89.23 19.45
U4b II Transition 98 115 15 57.04 23.43 19 115 35 78.16 19.75
U4c IV Transition 36 105 25 65 21.34 9 95 35 67.22 16.85
U4c III Transition 38 115 25 56.05 19.57 6 125 25 80 31.49
U4c II Transition 52 95 15 56.73 19.49 14 115 25 72.14 20.5
CT4-1d IV L. Prehistoric 163 125 15 71.13 16.74 2 105 95 100 5
H2B IV L. Prehistoric 72 125 15 52.5 26.76 1 75 75 75 0
CT4-1d III L. Prehistoric 413 125 15 62.22 25.24 17 125 25 74.41 32.08
H2B III L. Prehistoric 214 125 25 54 24.3 6 115 65 105 21.6
U4e III L. Prehistoric 216 115 15 67.92 23.97 49 145 25 80.1 25.32
U4e II L. Prehistoric 172 115 15 61.1 22.19 15 115 25 81.67 23
MP3g III L. Prehistoric 215 115 5 46.07 29.84 14 125 25 94.29 28.4
H2A II L. Prehistoric 25 105 15 47.8 23.24 1 75 75 75 0
H2B II L. Prehistoric 127 125 15 48.62 22.37 1 85 85 85 0
CT4-1d II L. Prehistoric 470 125 15 57.74 23.49 7 115 65 87.86 16.66
MP3g II L. Prehistoric 143 115 15 44.72 26.76 3 115 75 101.7 18.86
U4e I L. Prehistoric 59 85 15 41.78 16.92 5 85 45 61 13.56
H2B I L. Prehistoric 168 115 15 38.93 17.49 1 65 65 65 0
CT4-1d I L. Prehistoric 284 95 15 51.58 18.06 5 105 45 79 23.32
MP3g I L. Prehistoric 76 105 15 27.11 15.59
H2A I L. Prehistoric 35 95 15 43.29 21.58
Table 2. Summary Data for Black Abalone (H. scracherodii) and Green Abalone (H. fulgens) Samples from Bajamar-Jatay Shell Middens.
Note: N = sample size,; length measurements in mm; SD = standard deviation.
could be measured due to their poor state of preserva-tion (Table 5; Figure 4).
The limpet L. gigantea has a wide size range for the Archaic period (5–55 mm), and the average size is 35.78 mm. For the Transition, the average size is
29.75 mm, and the range is from 15 to 75 mm. For the Late Prehistoric period, the average size is 36.74 mm, and the length range is from 5 to 75 mm (Figure 5).
The average size of M. californianus mussels from the Archaic period is 35.70 mm and from the

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 65
Stratum Period N Max Min Mean SD
MP3c IV Archaic 20 25 15 18 4.58
MP3c III Archaic 78 35 5 20.38 5.48
MP3c II Archaic 153 35 5 20.23 5.61
MP3g VI Archaic 3 15 15 15 0
MP3g V Archaic 2 25 15 20 5
U4e V Archaic 1 25 25 25 0
U4b III Transition 39 35 15 19.1 5.42
U4b II Transition 70 35 15 18.86 5.15
U4c IV Transition 19 25 15 18.68 4.82
U4c III Transition 14 45 15 20.71 8.21
U4c II Transition 19 25 15 19.21 4.94
CT4-1d IV L. Prehistoric 24 35 5 23.75 5.99
H2B IV L. Prehistoric 85 35 15 24.76 5.08
CT4-1d III L. Prehistoric 229 35 15 24.78 6.09
H2B III L. Prehistoric 172 45 5 25.76 6.73
U4e III L. Prehistoric 105 25 15 18.14 4.64
U4e II L. Prehistoric 64 35 15 19.38 5.56
MP3g III L. Prehistoric 58 35 5 18.79 5.82
H2A II L. Prehistoric 51 45 15 23.63 8.17
H2B II L. Prehistoric 113 35 15 25.35 5.94
CT4-1d II L. Prehistoric 225 45 5 25.31 6.76
MP3g II L. Prehistoric 32 35 15 23.75 5.99
U4e I L. Prehistoric 32 35 15 20.63 5.56
H2B I L. Prehistoric 180 45 5 23.44 6.31
CT4-1d I L. Prehistoric 95 35 15 24.16 6.27
MP3g I L. Prehistoric 12 35 15 22.5 5.95
H2A I L. Prehistoric 79 35 15 20.32 6.13
Table 3. Summary Data for Owl Limpet (L. gigantea) and California Mussel (M. califor-nianus) Samples from Bajamar-Jatay Shell Middens.
Note: N = sample size,; length measurements in mm; SD = standard deviation.
Transition is 35.50 mm. They match in the minimum size (15 mm), while the maximum size from the Archaic period strata is 115 mm, and in the Transi-tion it is 75 mm. For the Late Prehistoric period, the average size is 43.62 mm, and the length range is from 5 to 145 mm (Figure 6).
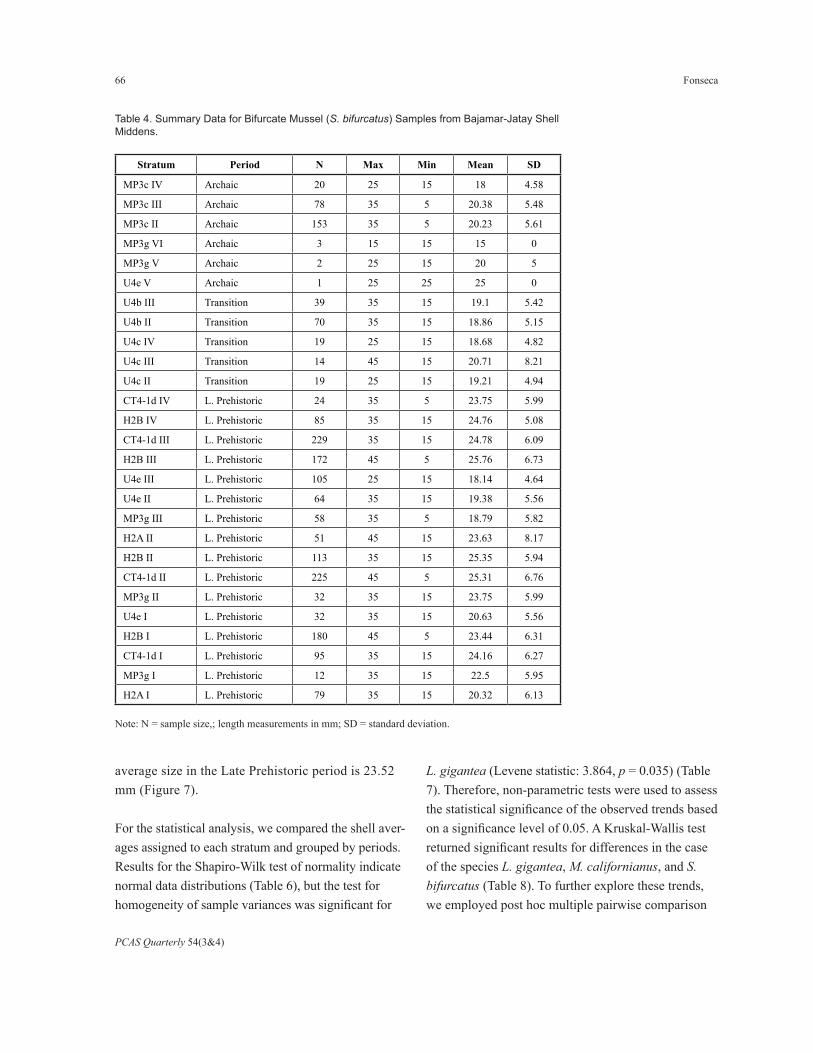
The average sizes of S. bifurcatus mussels col-lected during the Archaic period (20.06 mm) and the Transition (19.10 mm) are similar, but the size ranges are not. In the Archaic period, it is from 5 to 35 mm, in the Transition from 15 to 45 mm, and in the Late Prehistoric period from 5 to 45 mm. The
PCAS Quarterly 54(3&4)
Fonseca 66
Stratum Period N Max Min Mean SD
MP3c IV Archaic 20 25 15 18 4.58
MP3c III Archaic 78 35 5 20.38 5.48
MP3c II Archaic 153 35 5 20.23 5.61
MP3g VI Archaic 3 15 15 15 0
MP3g V Archaic 2 25 15 20 5
U4e V Archaic 1 25 25 25 0
U4b III Transition 39 35 15 19.1 5.42
U4b II Transition 70 35 15 18.86 5.15
U4c IV Transition 19 25 15 18.68 4.82
U4c III Transition 14 45 15 20.71 8.21
U4c II Transition 19 25 15 19.21 4.94
CT4-1d IV L. Prehistoric 24 35 5 23.75 5.99
H2B IV L. Prehistoric 85 35 15 24.76 5.08
CT4-1d III L. Prehistoric 229 35 15 24.78 6.09
H2B III L. Prehistoric 172 45 5 25.76 6.73
U4e III L. Prehistoric 105 25 15 18.14 4.64
U4e II L. Prehistoric 64 35 15 19.38 5.56
MP3g III L. Prehistoric 58 35 5 18.79 5.82
H2A II L. Prehistoric 51 45 15 23.63 8.17
H2B II L. Prehistoric 113 35 15 25.35 5.94
CT4-1d II L. Prehistoric 225 45 5 25.31 6.76
MP3g II L. Prehistoric 32 35 15 23.75 5.99
U4e I L. Prehistoric 32 35 15 20.63 5.56
H2B I L. Prehistoric 180 45 5 23.44 6.31
CT4-1d I L. Prehistoric 95 35 15 24.16 6.27
MP3g I L. Prehistoric 12 35 15 22.5 5.95
H2A I L. Prehistoric 79 35 15 20.32 6.13
Table 4. Summary Data for Bifurcate Mussel (S. bifurcatus) Samples from Bajamar-Jatay Shell Middens.
Note: N = sample size,; length measurements in mm; SD = standard deviation.
average size in the Late Prehistoric period is 23.52 mm (Figure 7).
For the statistical analysis, we compared the shell aver-ages assigned to each stratum and grouped by periods. Results for the Shapiro-Wilk test of normality indicate normal data distributions (Table 6), but the test for homogeneity of sample variances was significant for
L. gigantea (Levene statistic: 3.864, p = 0.035) (Table 7). Therefore, non-parametric tests were used to assess the statistical significance of the observed trends based on a significance level of 0.05. A Kruskal-Wallis test returned significant results for differences in the case of the species L. gigantea, M. californianus, and S. bifurcatus (Table 8). To further explore these trends, we employed post hoc multiple pairwise comparison

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 67
Figure 3. Maximum, minimum, and average values in the size (antero-posterior) of the species H. cracherodii in each stratum over time.
Figure 4. Maximum, minimum, and average values in the size (antero-posterior) of the species H. fulgens in each stratum over time.
Figure 5. Maximum, minimum, and average values in the size (antero-posterior) of the species L. gigantea in each stratum over time.

PCAS Quarterly 54(3&4)
Fonseca 68
Haliotis cracherodii Haliotis fulgens Lottia gigantea
Period Total Mean (mm) SD (mm) Total Mean (mm) SD (mm) Total Mean (mm) SD (mm)
Archaic 1043 47.03 25.29 8 90.00 25.50 644 35.78 9.57
Transition 302 61.69 23.63 74 79.73 22.13 476 29.75 8.58
Late Prehistoric 2852 54.64 25.06 127 82.56 26.43 2407 36.74 8.78
Mytilus californianus Septifer bifurcatus
Period Total Mean (mm) SD (mm) Total Mean (mm) SD (mm)
Archaic 3712 35.70 14.62 257 20.06 5.52
Transition 180 35.50 12.35 161 19.10 5.51
Late Prehistoric 9039 43.62 13.79 1556 23.52 6.64
Figure 6. Maximum, minimum, and average values in the size (antero-posterior) of the species M. californianus in each stratum over time.
Table 5. Shellfish Size Data by Archaic, Transition, and Late Prehistoric Periods.
Figure 7. Maximum, minimum and average values in the size (antero-posterior) of the species S. bifurcatus in each stratum over time.

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 69
using the Games-Howell test. The results appear in Tables 9–11 with statistically significant comparisons indicated.
As can be seen from these data, the Transition period presents the most evident changes for L. gigantea. The average size decreased considerably in this period, and
the size of the specimens increased again in the Late Prehistoric. In the cases of M. californianus and S. bifurcatus our results indicate significant differences between the Transition and Late Prehistoric samples. In both cases, an increased in the maximum length is observed in the Late Prehistoric.
Discussion
Changes in the size of the mollusks found in the coast-al camps may be an indicator of processes of human overexploitation, where it is expected to find larger sizes in the deepest strata and a gradual decrease toward the upper strata, at least when the changes cannot be attributed to causes of an environmental nature (Swadling 1976; Anderson 1981; Mannino and Thomas 2002; Braje et al. 2007; Erlandson et al. 2008; Jerardino et al. 2008; Yamazaki and Oda 2009; Erlandson and Rick 2010; Jerardino 2010; Erlandson et al. 2011, 2015; Thangavelu et al. 2011; Braje et al. 2012; Olguín et al. 2015). In Bajamar-Jatay, contrary to what was expected, a simple pattern of overex-ploitation was not found. Alternative explanations for changes or stability in shell size that were considered include selective harvesting, focus on alternative resources, reduction of intraspecific competition, and environmental disturbances.
Despite the high number of species identified in the ar-chaeological sites (82 different species), the diet of the
Table 6. Shapiro-Wilk Test of Normality for Each Shellfish Species by Periods.
Archaic Transition Late Prehistoric
Species s-w df p s-w df p s-w df p
H. cracherodii 0.812 6 0.075 0.811 5 0.099 0.984 16 0.985
H. fulgens 0.834 4 0.18 0.98 5 0.935 0.96 14 0.723
L. gigantea 0.939 6 0.652 0.936 5 0.635 0.918 16 0.159
M. californianus 0.949 6 0.735 0.958 5 0.791 0.897 16 0.071
S. bifurcatus 0.941 6 0.669 0.777 5 0.052 0.889 16 0.055
Note: No significant results at α = 0.05.
Table 7. Levene Test for Homogeneity.
Species F p
H. cracherodii 1.85 0.179
H. fulgens 1.239 0.311
L. gigantea 3.864 0.035*
M. californianus 0.673 0.52
S. bifurcatus 2.486 0.104
Table 8. Results of the Kruskal-Wallis Statistical Test (Non-Parametric Anova) Relative to the Average Size per Species over Time.
*Significant results at an α = 0.05.
Species H p
H. cracherodii 3.392 0.183
H. fulgens 3.944 0.139
L. gigantea 11.117 0.004*
M. californianus 9.981 0.007*
S. bifurcatus 7.048 0.029*
*Significant results at an α = 0.05 with two degrees of freedom.

PCAS Quarterly 54(3&4)
Fonseca 70
(I) Period (J) Period Mean Difference (I-J) Std. Error p95% Confidence Interval
Lower Bound Upper Bound
Archaic L. Prehistoric .1683 .22898 .752 -.5123 .8490
Transition .8433* .22053 .022 .1601 1.5266
L. Prehistoric Archaic -.1683 .22898 .752 -.8490 .5123
Transition .6750* .10572 .000 .4045 .9455
Transition Archaic -.8433* .22053 .022 -1.5266 -.1601
L. Prehistoric -.6750* .10572 .000 -.9455 -.4045
*Significant results at an α = 0.05.
Table 9. Results of the Games-Howell Post Hoc Analysis Used to Test Significant Differences between the Average Sizes of L. Gigantea in the Study Periods.
(I) Period (J) Period Mean Difference (I-J) Std. Error Sig.95% Confidence Interval
Lower Bound Upper Bound
Archaic L. Prehistoric -.6917 .24939 .058 -1.4082 .0248
Transition .0183 .28349 .998 -.7751 .8118
L. Prehistoric Archaic .6917 .24939 .058 -.0248 1.4082
Transition .7100* .20998 .025 .1045 1.3155
Transition Archaic -.0183 .28349 .998 -.8118 .7751
L. Prehistoric -.7100* .20998 .025 -1.3155 -.1045
*Significant results at an α = 0.05.
Table 10. Results of the Games-Howell Post Hoc Analysis Used to Test Significant Differences between the Average Sizes of M. Californianus in the Study Periods.
(I) Period (J) Period Mean Difference (I-J) Std. Error Sig.95% Confidence Interval
Lower Bound Upper Bound
Archaic L. Prehistoric -.3021 .14815 .170 -.7334 .1292
Transition .0447 .13864 .945 -.3874 .4768
L. Prehistoric Archaic .3021 .14815 .170 -.1292 .7334
Transition .3468* .07250 .000 .1625 .5310
Transition Archaic -.0447 .13864 .945 -.4768 .3874
L. Prehistoric -.3468* .07250 .000 -.5310 -.1625
Table 11. Results of the Games-Howell Post Hoc Analysis Used to Test Significant Differences between the Average Sizes of S. Bifurcatus in the Study Periods.
*Significant results at an α = 0.05.

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 71
prehistoric groups was concentrated in five species: Haliotis cracherodii, Haliotis fulgens, Lottia gigantea, Mytilus californianus, and Septifer bifurcatus (Fonse-ca 2017). The data presented here indicate temporal patterning of shellfish size fluctuation, particularly during the Late Prehistoric. A statistically signifi-cant increase in average shell size for L. gigantea, M. californianus, and S. bifurcatus from the Transi-tion (3500–1500 BP) to the Late Prehistoric period (1500–140 BP) was observed.
A range of environmental factors may influence size patterns, including microhabitat and small-scale oceanographic variations such as wave action, storms, and upwelling activity (Claassen 1986; Jerardino et al. 2008; Thakar et al. 2017; Flores 2017), as well as intraspecific competition and relative resilience of different species to harvesting (Coe and Fox 1942; Perry 2002; Bailey and Milner 2008; Ortega 2015). Additional environmental conditions, such as re-configuration of the coastal morphology or changes in water turbidity due to sea level changes, can also in-fluence shellfish growth rates (Jeradino 1997; Yamaza-ki and Oda 2009). Considering that the occupations of Bajamar-Jatay occurred during the Late Holocene, it is not likely that the paleogeographic conditions of the locality have been extremely different from those of its current configuration (Clark et al. 2014; Reed-er-Myers et al. 2015).
The growth rates of open-coast mollusk species are in-fluenced by the sea surface temperature (SST), relative-ly low-temperature conditions being physiologically more efficient, while conversely relatively warm SST would slow growth rates (Jerardino 1997; Jerardino et al. 2008). Since there are no paleotemperature records for northern Baja California, high-resolution SST and marine productivity curves developed for the Channel Islands and southern California were considered in order to evaluate the influence of climatically driven environmental changes on assemblage composition
(Kennett and Kennett 2000; Barron et al. 2003; Ken-nett et al. 2007; Barron and Anderson 2011).
Paleoceanographic reconstructions indicate that during the middle to late Holocene (~4000 BP), southern California SSTs were 2° C colder than today (Barron et al. 2003; Barron and Anderson 2011). Between 3000 and 1500 BP, water temperatures were relatively warm and stable, marked by low upwelling (Kennett and Kennett 2000). There is evidence for a higher frequency of El Niño Southern Oscillation (ENSO) events since then (Barron and Anderson 2011). During the Late Holocene, specifically the Medieval Climate Anomaly (MCA) (1500–600 BP), temperatures were 1° C colder compared to current temperatures (Ken-nett and Kennett 2000; Barron et al. 2003; Barron and Anderson 2011). After 600 BP, during Little Ice Age (LIA), temperatures were warmer and more stable (Kennett and Kennett 2000).
The greatest decreases in average shell size occur after 3000 BP. This coincided with a period of warm water and low marine productivity (Kennett and Kennett 2000). There were changes in the selection of mollusks; for example, H. fulgens, a species that required more effort for its capture, was collected in a higher propor-tion (Fonseca 2017), and the size range of the main species decreased. This may have reflected a strategy to maximize the available resources (Raab 1992).
An increase in shell size of the mussels and the owl limpet during Late Prehistoric period in Bajama-Jatay occurred in a period marked by generally cold SST and high marine surface productivity (Kennett and Kennett 2000). Evidence for economic expansion, population growth, proliferation of sedentary commu-nities, and technological improvements coincided with an emphasis on fishing and the greatest decreases in average shell size in the Santa Barbara Channel region (Kennett and Kennett 2000; Braje et al. 2007; Rick 2007; Jazwa et al. 2012).

PCAS Quarterly 54(3&4)
Fonseca 72
In the case of Bajamar-Jatay, we found a change to less highly ranked species but an increase in shell sizes. We consider that this phenomenon may have been the result of a combination of environmental and human factors. During the Transition, climatic events may have caused a decrease of mollusk populations or a displacement of the species most sensitive to temperature changes. With time and new variations in climate and marine productivity, populations may have recovered (Jerardino 1997). The Late Prehistor-ic period was characterized by an intensification of ENSO events (Barron and Anderson 2011) and two general climatic phases in water temperatures; cold and unstable periods in the MCA, and warmer and stable periods during the LIA (Kennett and Kennett 2000). Perhaps these environmental changes had an effect on the forms of organization of northern Baja California’s inhabitants.
An increase in mollusk consumption was observed in the Bajamar-Jatay area in the Late Prehistoric period (Guía 2008, 2013a; Fonseca 2017), but there was also an increase in the capture of shallow-water and rocky-bottom fish (Guía 2008, 2013b). This pattern suggests an increase in the use of fish, as occurred in the Channel Islands (Kennett and Kennett 2000; Vellanoweth et al. 2002; Rick 2007; Braje et al. 2007; Ainis et al. 2011; Jazwa et al. 2012), but not necessar-ily a decline in or replacing of shellfish (Fonseca et al. 2018). In comparison to mollusks, vertebrate groups, both marine and terrestrial, appear to have been sec-ondary resources in the coastal areas of northern Baja California (Fonseca et al. 2018). During this period, a demographic increase, new groups, and new organiza-tion may possibly have occurred, and if so, they might explain the harvesting changes (Rogers 1945; Moriar-ty 1966; Laylander 1987a, 1987b).
If there was an increase in mollusk consumption in the Late Prehistoric period, why was there not a decrease in mollusk sizes? We believe that the pattern could be explained by changes in the capture strategies,
triggered by population growth and/or a change in col-lection strategies that were derived from groups with new forms of organization. Human population growth is one of the most import-ant causes of increased predation pressure and overex-ploitation of shellfish. Broadening diet to include less profitable and lower-ranked shellfish species (Raab 1992; Broughton 1994) is a strategy that has been em-ployed to counteract the effects of population increase and the impact of overexploitation of local resources (Raab 1992; Mannino and Thomas 2002; Braje et al. 2007; Smith et al. 2014). This may explain the increase in the consumption of S. bifurcatus, a species of lower rank (Braje et al. 2007).
Shifts in selective strategies in settlements located on the Gulf of California have previously been observed (Celis 2011). In conjugation, ecological and envi-ronmental data indicated that shell middens from the middle to late Holocene (2299 BP) were formed under conditions when the Colorado River was not flowing into the Gulf. During this period, the resources collect-ed were significantly more diverse, and collection was less selective compared to more recent settlements (1498 BP). This variation is explained by Celis (2011) as a change in subsistence organization, and, over time, by human groups focusing their efforts on only some of the most profitable shellfish. Although it is a different pattern than the one we observed, it is anoth-er example of changes in collection strategies aimed at making the most of resources.
As in this analysis, no overexploitation trends were observed in archaeological assemblages elsewhere on the northwest coast of Baja California. At La Jovita, 5 km south of our study area, Robles (2013) reports no evidence of overexploitation of either California mus-sel or owl limpet, based on a homogenous pattern of shell sizes between two Late Prehistoric occupations.
On Baja California’s northwest coast, the resource use implied strong pressure on the mollusk populations,

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 73
since they were exploited for more than 5,000 years and evidence of year-round consumption is suggested (Fonseca 2017). A degree of chronological control is not available that could confirm that their use was continuous and prolonged, and it is possible that their use was only episodic and had no real impact on the populations, an alternative that cannot be entirely ex-cluded, at least at present. However, evidence supports an increase in harvest and changes in consumption patterns during the Late Prehistoric period.
Resource management strategies may have been conducted by the inhabitants of Bajamar-Jatay to counteract the pressure on the resource. Overexploita-tion did not occur at the levels discussed by Bailey et al. (2008), in which prey numbers decrease because human predation exceeds the level at which a targeted species can sustain its population.
During the Late Prehistoric period, new social or-ganization styles were developed. It is believed that Yuman groups had a different subsistence pattern than that of their predecessors, with a more seasonal pattern to their harvesting. The previous marine diet was changed to a diet based on the consumption of terrestrial mammals and seeds such as acorns or pine nuts, as evidenced by the proliferation of mortars and grinding stones. During summer, the groups moved from the coast to the mountains for a seed-collect-ing season, and they returned to the coast during the winter (Drucker 1937; Rogers 1945; Laylander 1987b; Christenson 1992; Hohenthal 2001; Guía 2005a; Oviedo and Guía 2009). When El Niño events inten-sified during the late Holocene period (Barron et al. 2003), human communities must have increased their reliance on terrestrial resources as marine productivity decreased (Davis 2010).
In contrast, the results of seasonality allocation of mussel harvesting activities for the MCA period sug-gest that around 1000 BP, shellfish harvesting was car-ried out throughout the year. This finding contradicts
the hypothesis that after the arrival of Yuman groups harvesting activities were seasonal (Fonseca 2017). It is important to consider that although the Yumans had a seasonal livelihood pattern, it was sometimes more convenient for them to inhabit coasts on a permanent basis during the MCA period, as a result of the high marine productivity that characterized the region and the extreme inland droughts (Kennett and Kennett 2000). It is possible that after this period, the Yumans resumed their nomadic or seminomadic settlement pattern. Another possibility is parallel subsistence pat-terns from two different group (Jones et al. 2008). One group may have occupied the coastal area permanent-ly, while the other harvested coastal resources only during certain times of the year.
In either of the two scenarios mentioned above, chang-es in mollusk capture strategies due to new forms of organization in the region could explain the increase in the size of mussels and limpets collected. Jones and Richman (1995) analyzed two possible mussel har-vesting strategies: the en masse harvesting of complete mussel beds (“stripping”), and the selection of larger individuals (“plucking”).
Finding larger mussel sizes from the Late Prehistoric period may be related to a change in capture strategy (from collecting the entire cluster of mussels to select-ing certain individuals) (Jones and Richman, 1995). This action may reduce intraspecific competition and stimulate the growth of individuals (Whitaker 2008; Giovas et al. 2010; Thangavelu et al. 2011; Giovas et al 2013; Poteate et al. 2015; Olguín et al. 2015; Flores 2017). This does not rule out the possibility that other factors influenced field processing and transport of dif-ferent mollusk prey. Foraging strategies may vary also between individuals, men and women, family groups, and age groups (Bird and Bird 2000; Bird et al. 2002; Bird 2007; Bird and Codding 2015).
If the Yumans had a seasonal settlement pattern with a more varied diet based on marine and terrestrial

PCAS Quarterly 54(3&4)
Fonseca 74
resources (Drucker 1937; Rogers 1945; Laylander 1987b; Christenson 1992; Hohenthal 2001; Guía 2005b; Oviedo and Guía 2009), it is possible that, as has been observed in other hunter-gatherer econo-mies (Bird and Bird 2000; Whitaker and Byrd 2014), the tasks associated with the collection of mollusks were developed by the elderly or children who stayed in the base camps. For now, these ideas are highly speculative, but they are hypotheses that could guide future work in which it will be important to analyze the evidence in light of other capture and processing strategies related to the groups’ mobility (Mannino and Thomas 2002; Bird et al. 2002; Jazwa et al. 2015; Thangavelu et al. 2011; Jerardino 2014; Smith et al. 2014; Thakar et al. 2017).
Conclusions
During the prehistory of Bajamar-Jatay, Baja Califor-nia, the capture of mollusks was intensive and concen-trated on the species Mytilus californianus, Septifer bifurcatus, Haliotis cracherodii, Haliotis fulgens, and Lottia gigantea. The pressure on these food resources might have triggered their overexploitation, which, ac-cording to our working hypothesis, would be reflected in a decrease in size through time.
According to the analysis of the size composition of the principal mollusks consumed, the most important changes are seen for the Transition period (3500–1500 BP). The smallest size intervals and a decrease in the average size of all species were identified, with the exception of the abalone H. cracherodii. Were the ob-served changes due to climatic factors? It will be im-portant to analyze in future work whether the observed phenomenon was related to an increase in temperature or low marine productivity that might have forced the groups to optimize the available resources to the maximum.
For the Late Prehistoric period (1500–140 BP), an increase in the maximum lengths of M. californianus
and S. bifurcatus was observed. The size of L. gi-gantea decreased for the Transition and increased for the Late Prehistoric period, but it did not again reach the average sizes recorded for the Archaic period (8000–3500 BP).
Diet broadening to include less profitable and low-er-ranked shellfish species is a strategy employed to counteract the effects of population increase and the impact of overexploitation of local resources (Raab 1992; Broughton 1994; Mannino and Thomas 2002; Braje et al. 2007; Smith et al. 2014). This could ex-plain the increase in the consumption of S. bifurcatus.
We do not think that the Bajamar-Jatay inhabitants reached overexploitation in terms of surpassing the capacity for recovery, but we can see signs of intensifi-cation of the resources’ use in a different consumption pattern over time. There was a succession in the species that were part of the diet, and there was an increase in the size of the mollusks collected in late prehistory.
It is necessary to emphasize that changes in size of certain species may have been the result of a combina-tion of factors. On the one hand, climatic events may have caused a decrease of the mollusk populations or the displacement of the most sensitive species due to the changes in temperature. With time and new variations in climate and marine productivity, popula-tions may have recovered. On the other hand, a human demographic increase and/or the arrival of groups with new forms of organization on the northwest coast of Baja California during the Late Prehistoric period may have been responsible.
Endnote
1. Even though the Campbell and Braje (2015) formula is more accurate and localized for southern California, Singh and McKechnie’s regressions are likely more suitable for application to archaeological assemblages where growing conditions may vary over

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 75
archaeological time scales and apply to a wider size range of mussels than Campbell and Braje’s method (McKechnie et al. 2015).
Acknowledgments
I would like to acknowledge the Instituto Nacional de Antropología e Historia (INAH), especially Julia Bendímez Patterson, Director of the Centro INAH Baja California. My sincere appreciation to Andrea Guía and Miguel Téllez for sharing helpful feed-back. Thanks to Don Laylander and two anonymous reviewers for criticisms and suggestions that helped to improve this article.
References Cited
Ainis, Amira F., René L. Vellanoweth, Troy W. Davis, Jon M. Erlandson, and Torben C. Rick
2011 Changes in Marine Subsistence on San Miguel Island from 8,500 to 2,400 Years Ago: Analysis of Bulk Samples from Cave of the Chimneys (CA-SMI-603). Journal of California and Great Basin Anthropology 31(1):59–79.
Allen, Michael J. (editor)2017 Molluscs in Archaeology: Methods, Approach-
es and Applications. Oxbow Books, Oxford. Anderson, A. J.1981 A Model of Prehistoric Collecting on the
Rocky Shore. Journal of Archaeological Science 8(2):109–120.
Bailey, Geoff, James Barrett, Oliver Craig, and Nicky Milner
2008 Historical Ecology of the North Sea Basin: An Archaeological Perspective and Some Problems of Methodology. In Human Impacts on Ancient Ecosystems: A Global Perspec-tive, edited by Torben C. Rick and Jon M. Er-landson, pp. 215–42. University of California Press, Berkeley.
Bailey, Geoff, and Nicky J. Milner2008 Molluscan Archives from European Pre-
history. In Early Human Impact on Mega-molluscs, edited by Andrzej Antczak and Roberto Cipriani, pp. 111–34. Archaeopress, Oxford.
Barron, John A., and Lesleigh Anderson2011 Enhanced Late Holocene ENSO/PDO
Expression along the Margins of the East-ern North Pacific. Quaternary International 235(1&2):3–12.
Barron, John A., Linda Heusser, Timothy Herbert, and Mitch Lyle
2003 High-resolution Climatic Evolution of Coastal Northern California during the Past 16,000 years. Paleoceanography 18(1):1020–1035.
Bien, G. S., N. W. Rakestraw, and H. E. Suess1960 Radiocarbon Concentration in Pacific Ocean
Water. Tellus 12(4):436–443. Bird, Douglas W., and Rebecca Bliege Bird
2000 The Ethnoarchaeology of Juvenile Foragers: Shellfishing Strategies among Meriam Chil-dren. Journal of Anthropological Archaeolo-gy 19(4):461–476.
Bird, Douglas W., Jennifer L. Richardson, Peter M. Veth, and Anthony J. Barham
2002 Explaining Shellfish Variability in Middens on the Meriam Islands, Torres Strait, Aus-tralia. Journal of Archaeological Science 29(5):457–469.
Bird, Rebecca Bliege2007 Fishing and the Sexual Division of Labor
among the Meriam. American Anthropologist 109(3):442–451.
Bird, Rebecca Bliege, and Brian F. Codding2015 The Sexual Division of Labor. In Emerging
Trends in the Social and Behavioral Sci-ences, edited by Robert Scott and Stephan Kosslyn, pp. 1–16. John Wiley & Sons, New York.

PCAS Quarterly 54(3&4)
Fonseca 76
Braje, Todd J., Douglas J. Kennett, Jon Erlandson, and Brendan J Culleton
2007 Human Impacts on Nearshore Shellfish Taxa: A 7, 000 Year Record from Santa Rosa Island. American Antiquity 72(4):735–756.
Braje, Todd J., Torben C. Rick, and Jon M. Erlandson2012 A Trans-Holocene Historical Ecological
Record of Shellfish Harvesting on Califor-nia’s Northern Channel Islands. Quaternary International 264:109–120.
Briggs, John Carmon1974 Marine Zoogeography. McGraw-Hill, New
York.Bronk Ramsey, Christopher 2009 Bayesian Analysis of Radiocarbon
Dates. Radiocarbon 51(1):337–360.Broughton, Jack M.1994 Late Holocene Resource Intensification in the
Sacramento Valley, California: The Verte-brate Evidence. Journal of Archaeological Science 21(4): 501–14.
Campbell, Breana, and Todd J. Braje2015 Estimating California Mussel (Mytilus
californianus) Size from Hinge Fragments : A Methodological Application in Historical Ecology. Journal of Archaeological Science 58:167–174.
Celis Hernández, Ana Katalina2011 Explotación de recursos costeros y condi-
ciones ambientales en el delta del Río Colorado durante la prehistoria. Unpublished Master’s thesis, Universidad Autónoma de Baja California, Ensenada.
Christenson, Lynne E.1992 The Late Prehistoric Yuman Settlement and
Subsistence System: Coastal Adaptation. In Essays on the Prehistory of Maritime Cali-fornia, edited by Terry L. Jones, pp. 217–230. Center for Archaeological Research at Davis No. 10, University of California, Davis.
Claassen, Cheryl1986 Temporal Patterns in Marine Shellfish
Species Use along the Atlantic Coast in the Southeastern United State. Southeastern Archaeology 5(2):120–137.
Clark, Jorie, Jerry X. Mitrovica, and Jay Alder 2014 Coastal Paleogeography of the California-Or-
egon-Washington and Bering Sea Continental Shelves during the Latest Pleistocene and Holocene: Implications for the Archaeo-logical Record. Journal of Archaeological Science 52:12–23.
Coe, Wesley R., and Dennis L Fox 1942 Biology of the California sea-mussel (Myti-
lus californianus). I. Influence of tempera-ture, food supply, sex and age on the rate of growth. Journal of Experimental Zoology 90(1):1–30.
Davis, Loren G.2010 Baja California´s Paleonvironmental Context.
In La Prehistoria de Baja California. Avances en la arqueología de la península olvidada, edited by Don Laylander, Jerry Moore, and Julia Bendímez Patterson, pp. 19–30. Centro INAH Baja California, Mexicali.
Drakíc, Danilo 2009 Initial Interpretations of the La Punta Site,
Ensenada, Baja California. Proceedings of the Society for California Archaeology 22.
Drakíc, Danilo, I. Aquino, and Luis Delgado. 2007 Salvamento arqueológico Cañada Del Águi-
la, sitio La Punta, Conchero 40, informe final de la primera etapa de excavación y análisis, mayo-noviembre 2007. Centro INAH Baja California, Ensenada.
Drakíc, Danilo, and Oswaldo Cuadra2005 Salvamento arqueológico Costa Azul, Lote
20. Centro INAH Baja California, Ensenada.Drakíc, Danilo, and Luis Delgado2010 Salvamento arqueológico Cañada del Águila,
segunda temporada, informe final de la

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 77
segunda etapa de excavación y análisis, noviembre 2008–marzo 2009. Centro INAH Baja California, Ensenada.
Drucker, Philip A.1937 Culture Element Distributions: V Southern
California. Anthropological Records 1:1–52. University of California, Berkeley.
Erlandson, Jon M., Amira F. Ainis, Todd J. Braje, Nicholas P. Jew, Marlene Mcvey, Torben C. Rick, René L. Vellanoweth, and Jack Watts
2015 12,000 Years of Human Predation on Black Turban Snails (Chlorostoma funebralis) on Alta California’s Northern Channel Islands. California Archaeology 7(1):59–91.
Erlandson, Jon M., Tood J. Braje, Torben C. Rick, Nicholas P. Jew, Douglas J. Kennett, Nicole Dwyer, Amira F. Ainis, René L. Vellanoweth, and Jack Watts
2011 10,000 Years of Human Predation and Size Changes in the Owl Limpet (Lottia gigantea) on San Miguel Island, Califor-nia. Journal of Archaeological Science 38(5):1127–1134.
Erlandson, Jon M., and Torben C. Rick2010 Archaeology Meets Marine Ecology: The
Antiquity of Maritime Cultures and Human Impacts on Marine Fisheries and Ecosystems. Annual Review of Marine Science 2(1):231–251.
Erlandson, Jon M., Torben C. Rick, Todd J. Braje, Alexis Steinberg, and René L. Vellanoweth
2008 Human Impacts on Ancient Shellfish: A 10,000 Year Record from San Miguel Island, California. Journal of Archaeological Science 35(8):2144–2152.
Flores, Carola2017 Importance of Small-Scale Paleo-Ocean-
ographic Conditions to Interpret Chang-es in Size of California Mussel (Mytilus californianus), Late Holocene, Santa Cruz Island, California. Quaternary International 427:137–150.
Fonseca Ibarra, Enah2013a Baja California Shell Midden Camps:
Similarities and Differences, Research in Process in the Northern Coast of the Peninsu-la. Proceedings of the Society for California Archaeology 27:73–83.
2013b Estudio de campamentos en la línea costera y valles intermontanos de Baja California: informe de excavación temporada 2012, informe de análisis de materiales temporada 2012 y propuesta de trabajo 2013. Centro INAH Baja California, Ensenada.
2014 Estudio de campamentos en la línea costera y valles intermontanos de Baja California: informe de excavación temporada 2013 y propuesta de trabajo 2014. Centro INAH Baja California, Ensenada.
2015 Estudio de campamentos en la línea costera y valles intermontanos de Baja California: informe de análisis de materiales temporada 2013. Centro INAH Baja California, Ensenada.
2016 Estudio de campamentos en la línea costera y valles intermontanos de Baja California: in-forme de excavación temporada 2014. Centro INAH Baja California, Ensenada.
2017 Patrón de asentamiento y explotación de los recursos marinos costeros durante la prehis-toria en Jatay, Baja California. Unpublished Master’s thesis, Universidad Autónoma de Baja California, Ensenada.
Fonseca Ibarra, Enah, Andrea Guía Ramírez, and Amira F. Ainis
2018 Marine Subsistence and Cooking Features of Coastal Hunter-Gatherer-Fishers in Baja California, Mexico. Journal of California and Great Basin Anthropology. Submitted.
García Lozano, Rubén Fabián 2013 Tipología y cadenas operatorias de puntas de
proyectil de la costa noroeste de Baja Califor-nia: estudio de caso del sitio Bajamar-Jatay, Ensenada, B.C. Unpublished Master’s thesis, Universidad Veracruzana, Xalapa, Veracruz.

PCAS Quarterly 54(3&4)
Fonseca 78
Giovas, Christina M., Meagan Clark, Scott M. Fitzpat-rick, and Jessica Stone
2013 Intensifying Collection and Size Increase of the Tessellated Nerite Snail (Nerita tessella-ta) at the Coconut Walk Site, Nevis, Northern Lesser Antilles, AD 890–1440. Journal of Archaeological Science 40 (11):4024–38.
Giovas, Christina M., Scott M. Fitzpatrick, Meagan Clark, and Mira Abed
2010 Evidence for Size Increase in an Exploited Mollusc: Humped Conch (Strombus gibbe-rulus) at Chelechol Ra Orrak, Palau from ca. 3000–0 BP. Journal of Archaeological Science 37(11):2788–2798.
Guía Ramírez, Andrea2005a Diversidad faunística en los concheros
de Costa Azul: la vida junto a las costas. Memorias: Balances y Perspectivas de la Antropología e Historia de Baja California 6:1–13.
2005b Human-Faunal Relationships in Northwest-ern Baja California. Proceedings of the Soci-ety for California Archaeology 18:274–280.
2008 Indigenous Fishing Practices in Ancient Cali-fornia. Paper presented at the annual meeting of the Society for California Archaeology, Burbank.
2010 Informe análisis de materiales arqueozo-ológicos en Baja California: estudio com-parativo de arqueofaunas al norte de BC, un enfoque temporal. Centro INAH Baja California, Ensenada.
2013a Informe del análisis de materiales mala-cológicos del sitio Bajamar, temporada 2012. Centro INAH Baja California, Ensenada.
2013b Informe del análisis de los restos faunísticos del sitio Bajamar, temporada 2012, Centro INAH Baja California, Ensenada.
2015 Informe del análisis de materiales mala-cológicos del sitio Bajamar, temporada 2013. Centro INAH Baja California, Ensenada.
Harris, Edward C1991 Principios de estratigrafía arqueológica.
Crítica, Barcelona.Hernández-Ayón, José Martín, Víctor F.
Camacho-Ibar, Adán Mejía-Trejo, and Ale-jandro Cabello-Pasini
2007 Variabilidad del CO2 total durante eventos de surgencia en Bahía San Quintín, Baja California, México. In Carbono en ecosiste-mas acuáticos de México, edited by Benigno Hernández de la Torre and Gilberto Gaxiola Castro, pp. 187–200. Instituto Nacional de Ecología, Mexico City.
Hernández Muñoz, Cynthia, and Daniel Schoenberg1993a Reporte de recorrido: Bajamar 93. Centro
INAH Baja California, Ensenada.1993b Reporte de salvamento: proyecto Bajamar,
sitios concheros, Ensenada, B.C. BM93. Centro INAH Baja California, Ensenada.
Hohenthal, William D., Jr.2001 Tipai Etnographic Notes. A Baja California
Indian Community at Mid-Century. Edited by Thomas Blackburn. Ballena Press Anthropo-logical Papers No. 48. Menlo Park, California.
Hubbs, Carl L.1960 The Marine Vertebrates of the Outer Coast.
Systematic Zoology 9(3):134–147.Ingram, B. Lynn, and John R. Southon 1996 Reservoir Ages in Eastern Pacific Coastal and
Estuarine Waters. Radiocarbon 38(3):573–582.
Jazwa, Christopher S., Todd J. Braje, Jon M. Erland-son, and Douglas J. Kennett
2015 Central Place Foraging and Shellfish Process-ing on California’s Northern Channel Islands. Journal of Anthropological Archaeology 40:33–47.
Jazwa, Christopher, Douglas Kennett, and Danielle Hanson
2012 Late Holocene Subsistence Change and Marine Productivity on Western Santa Rosa

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 79
Island, Alta California. California Archaeolo-gy 4(1):69–98.
Jerardino, Antonieta1997 Changes in Shellfish Species Composition
and Mean Shell Size from a Late-Holocene Record of the West Coast of Southern Africa. Journal of Archaeological Science 24(11):1031–1044.
2010 Large Shell Middens in Lamberts Bay, South Africa: A Case of Hunter-Gatherer Resource Intensification. Journal of Archaeological Science 37(9):2291–2302.
2014 Stranded Rocky Shore Mussels and Their Possible Procurement during Prehistory on the West Coast of South Africa. Journal of Archaeological Science 49 (1):536–45.
Jerardino, Antonieta, George M. Branch, and Rene Navarro
2008 Human Impact on Precolonial West Coast Marine Environments of South Africa. In Human Impacts on Marine Environments: A Global Perspective, edited by Torben C. Rick and Jon M. Erlandson, pp. 279–296. Univer-sity of California Press, Berkeley.
Jones, Terry L., Douglas J. Kennett, James P. Kennett, and Brian F. Codding
2008 Seasonal Stability in Late Holocene Shell-fish Harvesting on the Central California Coast. Journal of Archaeological Science 35(8):2286–2294.
Jones, Terry L., and Jennifer R. Richman1995 On Mussels: Mytilus californianus as a Pre-
historic Resource. North American Archaeol-ogist 16(1):33–58.
Kennett, Douglas J., B. Lynn Ingram, Jon M. Erland-son, and Phillip Walker
1997 Evidence for Temporal Fluctuations in Ma-rine Radiocarbon Reservoir Ages in the Santa Barbara Channel, Southern California. Jour-nal of Archaeological Science 24(11):1051–1059.
Kennett, Douglas J., and James P. Kennett2000 Competitive and Cooperative Responses to
Climatic Instability in Coastal Southern Cali-fornia. American Antiquity 65(2):379–395.
Laylander, Don1987a Sources and Strategies for the Prehistory of
Baja California. Unpublished Master’s thesis, San Diego State University.
1987b Una exploración de las adaptaciones cultura-les prehistóricas en Baja California. Estudios Fronterizos 5(14):117–124.
Mannino, Marcello A., and Kenneth D. Thomas2002 Depletion of a Resource? The Impact of
Prehistoric Human Foraging on Intertidal Mollusc Communities and Its Significance for Human Settlement, Mobility and Disper-sal. World Archaeology 33(3):452–474.
Mckechnie, Iain, Singh, G. Gerald, Todd J. Braje, and Breana Campbell
2015 Measuring Mytilus californianus: An Adden-dum to Campbell and Braje (2015) and Singh and McKechnie (2015) Including Commen-tary and an Integration of Data. Journal of Archaeological Science 58:184–186.
McLean, James H.1978 Marine Shells of Southern California. Natu-
ral History Museum of Los Angeles County, Los Angeles.
Moriarty, James R.1966 Cultural Phase Divisions Suggested by Typo-
logical Change Coordinated with Stratigraph-ically Controlled Radiocarbon Dating at San Diego. Anthropological Journal of Canada 4:20–30.
Olguín, Laura, Carola Flores, and Diego Salazar 2015 Aprovechamiento humano de moluscos mari-
nos en conchales arqueológicos del Holoceno Temprano y Medio (12.000–5.500 años cal AP): costa meridional del Desierto de Ataca-ma, Chile. In Arqueomalacología: abordajes metodológicos y casos de estudio en el Cono

PCAS Quarterly 54(3&4)
Fonseca 80
Sur, edited by Heidi Hammond and Miguel A. Zubimendi, pp. 13–34. Vazquez Mazzini, Buenos Aires.
Ortega, Jimena 2015 Respuesta del mejillón Mytilus californianus
sometido a variaciones de temperatura en condiciones de laboratorio y trasplantados en campo. Unpublished Master’s thesis, Universidad Autónoma de Baja California, Ensenada.
Oviedo García, Fernando, and Andrea Guía Ramírez2009 Characteristics of a Shellfish Collecting
Camp at Costa Azul, Baja California. Proceedings of the Society for California Archaeology 23:1–5.
Ovilla Rayo, Gengis Judith2013 Las tradiciones funerarias en los campamen-
tos prehistóricos de Bajamar-Jatay, B.C. y su contexto regional. Unpublished Master’s thesis, Universidad Veracruzana, Xalapa, Veracruz.
Ovilla Rayo, Gengis Judith, and Rubén Fabián García Lozano
2007 Rescate arqueológico Bajamar 38-A1. Centro INAH Baja California, Ensenada.
2008 Salvamento arqueológico Bajamar-Jatay B.C. 2006–2007. Centro INAH Baja Califor-nia, Ensenada.
Perry, Jennifer E. 2002 Resource Intensification and Environmental
Variability: Subsistence Patterns in Middle and Late Period Deposits at CA-SBA-225, Vandenberg Air Force Base, California. Jour-nal of California and Great Basin Archaeolo-gy 24(1):81–102.
Porcayo Michelini, Antonio2007 Programa arqueológico del estado de Baja
California en el tercer milenio. In Memoria 2007–2012, edited by Nelly Robles, pp. 1438–1456. Instituto Nacional de Antro-pología e Historia, Mexico City.
Poteate, Aaron S., Scott M. Fitzpatrick, Meagan Clark, and Jessica H. Stone
2015 Intensified Mollusk Exploitation on Nevis (West Indies) Reveals ~Six Centuries of Sustainable Exploitation. Archaeological and Anthropological Sciences 7(3):361–74.
Quitmyer, Irvy R., and Elizabeth J. Reitz2006 Marine Trophic Levels Targeted between
AD 300 and 1500 on the Georgia Coast, USA. Journal of Archaeological Science 33(6):806–822.
Raab, L. Mark1992 An Optimal Foraging Analysis of Prehis-
toric Shellfish Collecting on San Clemente Island, California. Journal of Ethnobiology 12(1):63–80.
Ramírez Valdez, Juan Arturo2009 Ecología y biogeografía de las comunidades
de peces del intermareal rocoso en la costa occidental de la península de Baja California. Unpublished Master’s thesis, Universidad Autónoma de Baja California, Ensenada.
Reeder-Myers, Leslie, Jon M. Erlandson, Daniel R. Muhs, and Torben C. Rick
2015 Sea Level, Paleogeography, and Archeology on California’s Northern Channel Islands. Quaternary Researh 83(2):263–72.
Reimer, Paula J., Edouard Bard, Alex Bayliss, J. Warren Beck, Paul G. Blackwell, Christopher Bronk Ramsey, Caitlin E. Buck, Hai Cheng, R. Law-rence Edwards, Michael Friedrich, Pieter M. Grootes, Thomas P. Guilderson, Haflidi Hafl-idason, Irka Hajdas, Christine Hatté, Timothy J. Heaton, Dirk L. Hoffmann, Alan G. Hogg, Konrad A. Hughen, K. Felix Kaiser, Bernd Kromer, Sturt W Manning, Mu Niu, Ron W. Reimer, David A. Richards, E. Marian Scott, John R. Southon, Richard A. Staff, Christian S. M. Turney, and Johannes van der Plicht
2013 IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0–50,000 Years Cal BP. Radiocarbon 55(4):1869–1887.

PCAS Quarterly 54(3&4)
Overexploitation of Coastal Resources at Bajamar-Jatay, Baja California? 81
Reina, Magdalena1994 Trabajos arqueológicos de sondeo en el
conchero B3 del sitio Bajamar: informe preliminar, 1a temporada. Centro INAH Baja California, Ensenada.
1995 Jatay: un sitio conchero. In Memoria 1994: Seminario de Historia de Baja California, pp. 101–104. Universidad Autónoma de Baja California, Ensenada.
Rick, Torben C.2007 The Archaeology and Historical Ecology of
Late Holocene San Miguel Island. UCLA Co-tsen Institute of Archaeology, Los Angeles.
Rick, Torben C., and Jon M. Erlandson2008 Archaeology, Historical Ecology, and the Fu-
ture of Ocean Ecosystems. In Human Impacts on Ancient Marine Ecosystems: A Global Perspective, edited by Torben C. Rick and Jon M. Erlandson, pp. 297–308. University of California Press, Berkeley.
Rivadeneira, Marcelo M., Calogero M. Santoro, and Pablo A. Marquet
2009 Reconstructing the History of Human Impacts on Coastal Biodiversity in Chile: Constraints and Opportunities. Aquatic Conservation Marine Freshwater Ecosystems 20(1):74–82.
Robles Montes, Mayra del Carmen2013 Temporalidad, patrones de explotación y
significado ambiental de los moluscos en el conchero de La Jovita, Baja California. Unpublished Master’s thesis, Universidad Autónoma de Baja California, Ensenada.
Rogers, Malcolm J. 1945 An Outline of Yuman Prehistory. Southwest-
ern Journal of Anthropology 1(2):157–198.Rojas, Martín2009 Trabajo y género en la antigua Baja Califor-
nia: análisis osteobiográfico de los esqueletos del Jatay. Camino Real Misionero de Las California 6:34–37.
Serrano, Jorge1992 Informe de actividades del proyecto de reg-
istro de sitios arqueológicos en Baja Cali-fornia: temporada 1991. Centro INAH Baja California, Ensenada.
1993 Proyecto de investigación arqueológica en el sitio conchero Bajamar, 30 de julio de 1993. Centro INAH Baja California, Ensenada.
Singh, Gerald G., and Iain McKechnie2015 Making the Most of Fragments: A Method for
Estimating Shell Length from Fragmentary Mussels (Mytilus californianus and Myti-lus trossulus) on the Pacific Coast of North America. Journal of Archaeological Science 58:175–183.
Smith, Carley B., Claire E. Ebert, and Douglas J. Kennett
2014 Human Ecology of Shellfish Exploitation at a Prehistoric Fishing-Farming Village on the Pacific Coast of Mexico. Journal of Island and Coastal Archaeology 9(2):183–202.
Swadling, Pamela1976 Changes Induced by Human Exploitation in
Prehistoric Shellfish Populations. Mankind 10(3):156–162.
Thakar, Heather B. 2011 Intensification of Shellfish Exploitation:
Evidence of Species-Specific Deviation from Traditional Expectations. Journal of Archae-ological Science 38(10):2596–2605.
Thakar, Heather B., Michael A. Glassow, and Carol Blanchette
2017 Reconsidering Evidence of Human Impacts: Implications of Within-Site Variation of Growth Rates in Mytilus californianus along Tidal Gra-dients. Quaternary International 427:151–159.
Thangavelu, Anbarasu, Bruno David, Bryce Barker, Jean-Michel Geneste, Jean-Jacques Delannoy, Lara Lamb, Nick Araho, and Robert Skelly
2011 Morphometric Analyses of Batissa violacea Shells from EMO (OAC), Gulf Province,

PCAS Quarterly 54(3&4)
Fonseca 82
Papua New Guinea. Archaeology in Oceania 46(2):67–75.
Thomas, Kenneth D.2015a Molluscs Emergent, Part II: Themes and
Trends in the Scientific Investigation of Molluscs and Their Shells as Past Human Re-sources. Journal of Archaeological Science 56:159–167.
2015b Molluscs Emergent, Part I: Themes and Trends in the Scientific Investigation of Mol-lusc Shells as Resources for Archaeological Research. Journal of Archaeological Science 56:133–140.
Vellanoweth, René L., Torben C. Rick, and Jon M. Erlandson
2002 Middle and Late Holocene Maritime Adap-tations on Northeastern San Miguel Island, California. In Proceedings of the Fifth California Islands Symposium: 29 March to 1 April 1999, edited by David R. Browne, Kathryn L. Mitchell, and Henry W. Chaney, vol. 2, pp. 607–614. Santa Barbara Museum of Natural History, Santa Barbara, California.
Whitaker, Adrian R. 2008 Incipient Aquaculture in Prehistoric Califor-
nia? Long-Term Productivity and Sustainabil-ity vs. Immediate Returns for the Harvest of Marine Invertebrates. Journal of Archaeolog-ical Science 35(4):1114–1123.
Whitaker, Adrian R., and Brian F. Byrd2014 Social Circumscription, Territoriality, and the
Late Holocene Intensification of Small-Bod-ied Shellfish along the California Coast. Journal of Island and Coastal Archaeology 9(2):150–68.
Yamazaki, Takeshi, and Sen-ichi Oda2009 Changes in Shell Gathering in an Early
Agricultural Society at the Head of Ise Bay, Japan. Journal of Archaeological Science 36(9):2007–2011.