overexpression of the p80 tnf receptor leads to tnf-dependent
TRANSCRIPT

of April 9, 2019.This information is current as
Activation, and c-Jun Kinase ActivationBκTNF-Dependent Apoptosis, Nuclear Factor-
Overexpression of the p80 TNF Receptor Leads to
Bharat B. AggarwalValsala Haridas, Bryant G. Darnay, K. Natarajan, Renu Heller and
http://www.jimmunol.org/content/160/7/31521998; 160:3152-3162; ;J Immunol
Referenceshttp://www.jimmunol.org/content/160/7/3152.full#ref-list-1
, 18 of which you can access for free at: cites 40 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1998 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

Overexpression of the p80 TNF Receptor Leads toTNF-Dependent Apoptosis, Nuclear Factor-kB Activation, andc-Jun Kinase Activation1
Valsala Haridas,* Bryant G. Darnay,* K. Natarajan,* Renu Heller, † andBharat B. Aggarwal2*
Because they have distinct intracellular domains, it has been proposed that the p60 and p80 forms of the TNF receptor mediatedifferent signals. Several signaling proteins have been isolated that associate with either the p60 or the p80 receptor. By using TNFmuteins specific to the p60 and p80 receptors, we have previously shown that cytotoxicity and nuclear factor-kB (NF-kB) acti-vation are mediated through the p60 form of the endogenous receptor. What signals are mediated through the p80 receptor is lessclear. This study was an effort to answer that question. HeLa cells, which express only p60 receptors, were transfected with p80receptor cDNA and then examined for apoptosis, NF-kB activation, and c-Jun kinase activation induced by TNF and by p60 orp80 receptor-specific muteins. The p80 mutein, like TNF and the p60 mutein, induced apoptosis and activation of NF-kB and c-Junkinase in cells overexpressing recombinant p80 receptor but had no effect on cells expressing a high level of endogenous p80receptor. The apoptosis mediated through the p60 receptor was also potentiated after overexpression of the p80 receptor, sug-gesting a synergistic relationship between the two receptors. Interestingly, Abs to the p80 receptor blocked apoptosis induced byall ligands but by itself activated NF-kB in the p80-transfected cells. Overall, our results show that the p80 receptor, which lacksthe death domain, mediated apoptosis, NF-kB activation, and c-Jun kinase activation, but only when it was overexpressed, whereasendogenous p60 receptor mediated similar signals without overexpression.The Journal of Immunology,1998, 160: 3152–3162.
T NF, a highly pleiotropic cytokine, mediates all its effectsthrough two distinct receptors, referred to as p60 and p80.The existence of a similar extracellular domain (;29%
homology in amino acid sequences) but a dissimilar intracellulardomain (ICD)3 has led to the proposal that these two TNF recep-tors must transduce distinct signals (for references, see Ref. 1).This hypothesis is further strengthened by observations that theICD of the p60 receptor contains an amino acid sequence of ap-proximately 80 amino acids, referred to as the death domain (DD;due to its involvement in cell death), whereas the p80 receptor doesnot (2). Moreover, it has been shown that the ICD of the p60receptor recruits several signaling proteins distinct from that of thep80 receptor (for references, see Ref. 3), including TRADD,FADD, FLICE, RIP, TRAF-2, ICH-1, MADD, TRIP, I-FLICE,sentrin, A-20, and I-TRAF. In contrast, factors associated with
N-sphingomyelinase activation, PIP5K, TNF receptor-associatedprotein-1, and TNF receptor-associated protein-2 bind to the jux-tamembrane domain. The ICD of p80 without the DD recruitsTRAF-1, TRAF-2, IAP-1, IAP-2, and NF-kB-inducing kinase (3,4). Despite distinct ICD of the two receptors, TRAF-2 and IAP-1,which bind to the p80 receptor, also bind through secondary in-teraction to the p60 receptor, but most of the p60 receptor bindingproteins do not bind to the p80 receptor. In addition to these pro-teins, two distinct putative serine/threonine protein kinases havebeen shown to be recruited by the p60 and p80 receptors (5, 6).The roles of these proteins in TNF signaling are not entirely clear,but it appears that different proteins may mediate different TNF-induced signals. Even though TRADD, FADD, FLICE, RIP, andICH-1 have been implicated in cell death (7–13), and RIP,TRADD, and TRAF-2 have been implicated in TNF-induced ac-tivation of NF-kB and c-Jun kinase (14–17), the mechanisms areunknown.
One of the most fundamental unanswered questions in TNF bi-ology is thus what types of signals are mediated through the p80receptor compared with the p60 receptor. In general, it has beenshown that most of the TNF signals are mediated through the p60receptor. The p80 receptor has been implicated in ligand passing,in cytotoxicity, and in potentiation of p60 receptor-mediated cy-totoxicity (18–20). Recent observations using receptor-specificTNF muteins have indicated that the cytotoxicity and NF-kB ac-tivation are mediated entirely through the endogenous p60receptor (21).
In the present report we compared signaling through the endog-enous receptor with that through the recombinant p80 receptor.TNF muteins that bind exclusively to either the p60 or the p80receptor (22) were employed for these studies. In p60 receptor-specific mutein, residue 32 was altered from arginine to trypto-phan, and residue 86 was altered from serine to threonine, whereas
Cytokine Research Section, Department of Molecular Oncology, University of TexasM. D. Anderson Cancer Center, Houston, TX 77030; and *Roche Biosciences, PaloAlto, CA 94304
Received for publication July 9, 1997. Accepted for publication December 3, 1997.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by a grant from The Clayton Foundation for Research.2 Address correspondence and reprint requests to Dr. Bharat B. Aggarwal, CytokineResearch Section, Department of Molecular Oncology, University of Texas M. D.Anderson Cancer Center, Houston, TX 77030. E-mail address: aggarwal@utmdacc,mda.uth.tmc.edu3 Abbreviations used in this paper: ICD, intracellular domain; DD, death domain;FADD, Fas-associated death domain; FLICE, Fas-associated death domain-like IL-1-converting enzyme; RIP, receptor-interacting protein; TRAF, tumor necrosis factorreceptor-associated factor; IAP, inhibitor of apoptosis; NF-kB, nuclear factor-kB;JNK, c-Jun kinase; PARP, poly(A)DP ribose polymerase; GST, glutathione-S-trans-ferase; EMSA, electrophoretic mobility shift assay. TRADD, TNF-receptor associ-ated death domain; ICH-1, ICE and ced-3 homolog-1; MADD; Map kinase-activateddeath domain protein; TRIP, TRAF-interacting protein; I-FLICE, Inhibitor of FLICE;I-TRAF-Inhibitor of TRAF.
Copyright © 1998 by The American Association of Immunologists 0022-1767/98/$02.00
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

in p80 receptor-specific mutein, the carboxyl-terminus at position143 was mutated from aspartic acid to asparagine and at position145 from alanine to arginine. The p80 receptor-binding muteininduced apoptosis, activated NF-kB, and induced c-Jun kinase incells overexpressing the recombinant receptor but not in cells ex-pressing endogenous receptor. Thus, our results demonstrate thateven though the ICD of the p80 receptor is distinct from that of thep60 receptor and recruits several distinct proteins, overexpressedp80 receptor can transduce signals similar to those of the endog-enous p60 receptor but distinct from those of the endogenous p80receptor.
Materials and MethodsMaterials
RPMI 1640 medium, FCS, penicillin, streptomycin, and trypsin-EDTAwere obtained from Life Technologies (Grand Island, NY). Tris, glycine,NaCl, and BSA were obtained from Sigma Chemical (St. Louis, MO).Hygromycin B was purchased from Calbiochem-Novabiochem Interna-tional (La Jolla, CA). Bacteria-derived recombinant human TNF purified tohomogeneity was provided by Genentech (South San Francisco, CA).32P-labeledg-ATP with a sp. act. of 7000 Ci/mmol was obtained from Am-ersham Life Sciences (Arlington Heights, IL). Recombinant human TNFmuteins altered by site-specific mutagenesis to bind to either the p60 or p80receptor, expressed in bacteria, and purified to homogeneity were suppliedby F. Hoffmann-La Roche (Basel, Switzerland). The isolation and charac-terization of these muteins has been described previously (22). Rabbit anti-JNK polyclonal Ab was purchased from Santa Cruz Biotechnology (SantaCruz, CA). Mouse anti-poly(A)DP ribose polymerase (anti-PARP) Ab waspurchased from PharMingen (San Diego, CA). Anti-mouse horseradishperoxidase-conjugated secondary Ab was purchased from TransductionLaboratories (Lexington, KY). Protein A/G-Sepharose beads were ob-tained from Pierce Chemical (Rockford, IL). GST-Jun1–79was expressed inEscherichia coliand purified essentially as previously described (5).
Cell lines
The human histiocytic cell line U-937, human myeloid ML-1a, and humanrhabdomyosarcoma cell line KYM-1 were grown in RPMI 1640 mediumsupplemented with FBS (10%), penicillin (100mg/ml), and streptomycin(100 mg/ml). Cells were seeded at a density of 13 105/ml in T75 flasks(Falcon 3013, Becton Dickinson Labware, Lincoln Park, NJ) containing 20ml of medium and were grown at 37°C in an atmosphere of 95% air and5% CO2. Cell cultures were split every third day. KYM-1 cells weretrypsinized and seeded again every third day.
HeLa cells stably transfected with pCDM8 mammalian expression vec-tor containing the full-length TNFRp80 cDNA, as previously described(18), were used for our studies. A plasmid carrying the hygromycin phos-photransferase gene was used as a selection marker and was cotransfectedwith the pCDM18 plasmid. Transfected cells were routinely grown inMEM containing 10% FBS, penicillin (100mg/ml), streptomycin (100mg/ml), and hygromycin (200mg/ml).
Different clones were generated from the HeLa-p80 cells by two dif-ferent methods. The first method involved sorting by FACS based on anti-p80 receptor Ab and anti-rabbit FITC-labeled secondary Ab. Depending onthe level of p80 expression, four different populations of HeLa-p80 cellswere obtained by this method. The second method involved isolation ofdifferent HeLa-p80 clones by limited dilution. Briefly, HeLa-p80 cells werediluted to 10 cells/ml and plated at 1 cell/100ml/well into 96-well plates.The plates were incubated at 37°C for 2 wk, and thereafter, three differentclones (no. 1–3) were isolated and further characterized.
The effect of p80 receptor transfection was also analyzed in another cellline, breast carcinoma MCF-7 (expresses only the p60 receptor), and byusing a different vector. For this, full-length TNFRp80 cDNA (6) wascloned into pEC1214A(pTet), a modified tetracycline repressor system ex-pression vector (a gift from Dr. Hong-Ji Xu, University of Texas M. D.Anderson Cancer Center, Houston, TX) containing a cloning site for thedesired gene that is under the control of the tetracycline repressor, theconstitutive expression of tetracycline repressor, and the neomycin resis-tance gene. MCF-7 cells were transfected with the pTet and pTet-p80 vec-tor using Lipofectamine as described by the manufacturer (Life Technol-ogies). A stable pool of MCF-7 cells harboring either pTet (MCF-7) orpTet-p80 (MCF-7 (p80)) were selected by growing the cells in completemedium containing tetracycline (1mg/ml) and G418 (600mg/ml). Induc-tion of p80 receptor occurred after the withdrawal of tetracycline.
Cytotoxicity assay
The cytotoxic effects of TNF and its muteins on adherent cells (KYM-1 andHeLa) were measured by the crystal violet dye uptake assay. Briefly, 103103 cells were plated in 0.1 ml of medium in 96-well flat-bottom plates.After an overnight incubation at 37°C, the medium was removed, anddifferent concentrations of TNF were layered in 0.1 ml of fresh medium.After 72 h of incubation at 37°C, the medium was removed, and the via-bility of cells was monitored by crystal violet staining according to a pro-cedure described previously (23). The cytotoxic effects of TNF on nonad-herent cells (U-937) were determined by the amount of [3H]thymidineincorporated by the cells as described previously (24). Briefly, cells wereplated at 5000/well in 0.1 ml of medium in 96-well flat-bottom Falconplates. Different concentrations of TNF were added in an additional 0.1 mlof medium and were incubated at 37°C for 72 h. During the last 6 h beforeharvesting, [3H]thymidine (5 mCi/mmol; Amersham) was added to eachwell (0.5 mCi/well), and then cells were harvested with the aid of Filter-mate 196 harvester (Packard Instruments, Meriden, CT). Radioactivitybound to the filter was measured in a liquid scintillation counter (model1600 TR, Packard Instruments).
Immunoblot analysis of PARP degradation
TNF-induced apoptosis was also examined by proteolytic cleavage ofPARP (25). Briefly, cells (13 106/ml) were treated with TNF or the mu-teins for either 2 h (U-937 cells) or 4 h (KYM-1, HeLa and HeLa-p80 cells)in the presence of 1mg/ml of cycloheximide. After treatment, cell extractswere prepared by incubating the cells for 30 min on ice in 0.05 ml of buffercontaining 20 mM HEPES (pH 7.4), 2 mM EDTA, 250 mM NaCl, 0.1%Nonidet P-40, 2mg/ml leupeptin, 2mg/ml aprotinin, 1 mM PMSF, 0.5mg/ml benzamidine, and 1 mM DTT. The lysate was centrifuged, and thesupernatant was collected. Cell extract protein (50mg) was resolved on a7.5% SDS-polyacrylamide gel, electrotransferred onto a nitrocellulosemembrane, blotted first with mouse anti-PARP Ab and then with anti-mouse horseradish peroxidase-conjugated IgG as the secondary Ab, andthen detected by chemiluminescence (ECL, Amersham). Apoptosis wasrepresented by the cleavage of 116-kDa PARP into 85- and 41-kDa peptideproducts (25).
Electrophoretic mobility shift assays (EMSAs)
An EMSA to examine NF-kB activation was conducted as described pre-viously (26, 27). Briefly, cells (23 106/ml) were treated separately withdifferent concentrations of TNF and its muteins at 37°C for 30 min. Nu-clear extracts were then prepared as described previously (27). EMSA wasperformed by incubating 4mg of nuclear extracts with 16 fmol of32Pend-labeled 45-mer double-stranded NF-kB oligonucleotide from theHIV-1 long terminal repeat, 59-TTGTTACAAGGGACTTTCCGCTGGGGACTTTCCAGGGAGGCGTGG-39, in the presence of 2 to 5mg ofpoly(dI-dC) in a binding buffer (25 mM HEPES (pH 7.9), 0.5 mM EDTA,0.5 mM DTT, 1% Nonidet P-40, 5% glycerol, and 50 mM NaCl) for 15min at 37°C. The DNA/protein complex formed was separated from freeoligonucleotide on a 7.5% native polyacrylamide gel. A double-strandedmutant oligonucleotide was used as a control to examine the specificity ofbinding of NF-kB to the DNA. The specificity of binding was also checkedby competition with the unlabeled oligonucleotide, by using mutant oligo-nucleotide, and by supershift of the band with Abs against NF-kB p50 andp65 proteins. The radioactive bands from dried gels were visualized on aPhosphorImager (Molecular Dynamics, Sunnyvale, CA) and quantitatedusing ImageQuant software.
Receptor binding assays
Receptor binding assays were performed as described previously (28). TNFwas labeled with Na125I, using the Iodogen method, to an approximate sp.act. of 40 mCi/mg. Binding assays were performed in flexible 96-wellplates. Cells (0.53 106/0.1 ml) were incubated in a binding medium(RPMI 1640 containing 10% FBS) with125I-labeled TNF in the absence(total binding) or the presence of 100 nM unlabeled ligand (nonspecificbinding) for 1 h at 4°C. Thereafter, cells were washed three times withice-cold medium (PBS containing 0.1% BSA) at 4°C, and the cell-boundradioactivity was determined in a gamma counter (Cobra-AutoGamma,Packard Instrument). All determinations were performed in triplicate. Spe-cific binding of TNF was calculated by subtraction of the amount (countsper minute) of nonspecific binding from the total binding.
To determine the levels of p60 and p80 forms of TNF receptors, affin-ity-purified Abs specific for each type of receptor were employed (21).Cells (0.53 106/0.1 ml) were preincubated with the Ab (5mg/ml) for 1 hat 4°C and then examined for specific binding of labeled TNF as describedabove. TNF muteins were also used to compete for TNF binding sites.
3153The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

Specific binding of TNF observed on competition with TNF (p60) or oncells pretreated with anti-p60 Ab would be due to the p80 receptor andvice versa.
c-Jun kinase assay
The c-jun kinase assay was performed using a modified method as de-scribed previously (29). Briefly, after treatment of cells (33 106/ml) withTNF for 15 min, cell extracts were prepared by lysing cells in buffer con-taining 20 mM HEPES (pH 7.4), 2 mM EDTA, 250 mM NaCl, 0.1%Nonidet P-40, 2mg/ml leupeptin, 2mg/ml aprotinin, 1 mM PMSF, 0.5mg/ml benzamidine, and 1 mM DTT. Cell extracts (150–250mg) weresubjected to immunoprecipitation with 0.03mg of anti-JNK Ab for 30 minat 4°C. Immune complexes were collected by incubation with protein A/G-Sepharose beads for 30 min at 4°C. The beads were collected by centrif-ugation and washed extensively with lysis buffer (four times, 400ml eachtime) and kinase buffer (twice, 400ml each time; 20 mM HEPES (pH 7.4),1 mM DTT, and 25 mM NaCl). Kinase assays were performed for 15 minat 30°C with 2mg GST-Jun1–79 in 20 ml containing 20 mM HEPES (pH7.4), 10 mM MgCl2, 1 mM DTT, and 10mCi [g-32P]ATP. Reactions werestopped with 15ml of SDS-sample buffer, boiled for 5 min, and subjectedto SDS-PAGE. GST-Jun1–79 was visualized by staining with CoomassieBlue, and the dried gel was analyzed by a PhosphorImager (MolecularDynamics) and quantitated by ImageQuant Software (MolecularDynamics).
ResultsWe examined the role of the p80 receptor in TNF-mediated apo-ptosis, NF-kB activation, and c-Jun kinase activation. To distin-guish the signaling transduced by the p80 receptor from that ofp60, we used genetically engineered TNF muteins that bind toeither the p60 or the p80 receptor. The results were confirmed bythe use of affinity-purified Abs specific to each receptor. We alsocompared the signaling of the recombinant p80 receptor with thatof the endogenous p80 receptor.
Characterization of endogenous vs recombinant p80 receptors
We used U-937 and KYM-1 cells, which have been shown toexpress a high density of high affinity (;5000 binding sites with aKd of 0.1 nM) TNF receptors (30). Furthermore, the level of p80receptors on these cells, determined using receptor-specific Abs,was approximately 3000 sites/cell (30, 31). In the present reportwe further examined the levels of p60 and p80 receptors by usingTNF muteins specific to each receptor as a competitor. TNF (p60)inhibited TNF binding on U-937 cells by approximately 46%, andTNF (p80) inhibited it by 56% (Fig. 1,upper panel), suggestingthat they expressed almost equal numbers of p60 and p80 recep-tors. Like U-937 cells, KYM-1 (Fig. 1,lower panel) cells ex-pressed a high level of TNF receptors, and their binding wasequally inhibited by p60- and p80-specific muteins. Comparedwith p60, U-937 cells expressed slightly lower p80, and KYM-1cells expressed slightly higher p80 receptor levels.
To examine the role of recombinant p80 receptors, we usedHeLa cells transfected with cDNA to overexpress p80 receptor asdescribed previously (18). Radioreceptor results (Fig. 2A) indicatethat nontransfected HeLa cells expressed p60 receptor, as TNFbinding could be blocked by anti-p60 Ab only (upper right) andinhibited by TNF (p60) mutein and not by TNF (p80) mutein (up-per left). HeLa cells, when transfected with p80 receptor cDNA,bound 20 to 30 times more labeled TNF, and this binding could beblocked primarily by anti-p80 Abs (lower right) and inhibited byTNF (p80) mutein. Anti-p60 Ab or TNF (p60) had a minimal effecton binding of TNF to transfected cells. These results clearly indi-cate that transfected HeLa cells overexpressed p80 receptors. Scat-chard analysis revealed that transfected HeLa cells express approx-imately 190,000 binding sites/cell (18).
The expression of endogenous and recombinant p80 receptors wasalso confirmed by Western blot analysis using anti-p80 receptor Abs(Fig. 2B). U-937, ML-1a, and KYM-1 expressed variable amounts of
endogenous p80 receptor protein, with the highest being expressed byKYM-1. As expected, nontransfected HeLa cells expressed no detect-able p80 receptor, whereas after transfection these cells expressed ahigh level. A small amount of a low m.w. band was also found intransfected cells. The origin of this band is unclear at present, but mayrepresent the degraded form of the p80 receptor.
Overexpression of p80 receptor mediates apoptosis
To examine signaling through the endogenous p80 receptor, weexamined the cytotoxic responses to TNF, TNF (p60), and TNF(p80). Both TNF and TNF (p60) were cytotoxic to U-937 cells(Fig. 3A,upper panel) and to KYM-1 cells (Fig. 3A, lower panel)in a dose-dependent manner. As little as 0.01 ng/ml of the ligand
FIGURE 1. Displacement of the binding of labeled TNF to U937 (up-per panel) and KYM-1 (lower panel) cells by unlabeled TNF and TNF(p60) and TNF (p80) muteins. Cells (0.53 106/0.1 ml) were incubatedwith labeled TNF (0.23 106 cpm) in the presence or the absence of 100nM excess cold TNF, TNF (p60), or TNF (p80) for 1 h at4°C. Thereafter,cells were washed and counted as described inMaterials and Methods. Allmeasurements were conducted in triplicate.
3154 TNF SIGNALING THROUGH THE p80 RECEPTOR
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

showed a significant effect. TNF (p80) mutein, however, had noeffect on these cells, even when used at 200 ng/ml, suggesting thatthe p80 receptor is inert in these cells. As will be shown later, thelack of this effect could not be attributed to the possibility that TNF(p80) mutein was not biologically active.
We investigated the effect of overexpressed recombinant p80 re-ceptors on TNF-mediated cytotoxicity. TNF had severalfold highercytotoxic effects on p80-transfected cells than control, endogenousp60-expressing HeLa cells (Fig. 3B, upper panel). These results im-ply that the p80 receptor, when overexpressed, could also mediate
FIGURE 2. A, Inhibition of the binding of labeled TNF to HeLa (upper panel) and HeLa-p80 cells (lower panel) cells by unlabeled TNF and TNF (p60)and TNF (p80) muteins (left panel) and by Abs to p60 and p80 receptors (right panel). Cells (0.53 106/ml) were either preincubated with 2mg/ml ofthe Ab at 4°C for 1 h followed by labeled TNF (0.23 106 cpm) for another 1 h orthey were coincubated with labeled TNF (0.23 106 cpm) in the presenceor the absence of 100 nM excess unlabeled TNF, or TNF (p60) or TNF (p80) muteins for 1 h at4°C. Thereafter, cells were washed and counted as describedin Materials and Methods. All measurements were conducted in triplicate.B, Western blot analysis of p80 receptor protein expression on U-937, ML-1a,KYM-1, HeLa, and HeLa-80 cells. One hundred micrograms of cell extracts were electrophoresed, transferred onto a nitrocellulose filter, blotted withanti-p80 Ab, and visualized by the ECL method.
3155The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

TNF-induced cell killing. To determine whether TNF could kill thecells without the assistance of the endogenous p60 receptor, we usedTNF muteins. Interestingly, we found that p80 receptor transfectionalso enhanced the activity of the mutein specific to the p60 receptor(Fig. 3B, middle panel), suggesting a synergistic effect in the signalingmediated through the p60 and p80 receptors. TNF (p80) mutein,which was inactive on cells expressing endogenous receptor, was alsoinactive on nontransfected HeLa cells (which express only the p60receptor; Fig. 3B, bottom panel). However, when examined on cellstransfected to overexpress the p80 receptor, TNF (p80) mutein was asactive as TNF, indicating that unlike the p60 receptor, the p80 recep-tor can mediate cytotoxicity only when overexpressed. These results
also indicate that p80 not only can mediate cytotoxicity but can alsoenhance p60-mediated cytotoxicity.
Besides cytotoxicity, which required 72-h treatment to be ex-pressed, we also examined TNF-induced cleavage of PARP, an-other early characteristic feature of apoptosis (25). After treatmentof KYM-1 cells for 4 h, both TNF and the TNF (p60) muteininduced cleavage of PARP; the TNF (p80) mutein, however, hadno effect (Fig. 4). Similarly, in U-937 cells the TNF (p60) muteininduced PARP cleavage within 2 h, whereas the TNF (p80) muteindid not (Fig. 4). These results clearly indicate that apoptosis ismediated through the endogenous p60 receptor but not the endog-enous p80 receptor. We also found that the TNF (p80) mutein
FIGURE 3. A, Cytotoxic effect of TNF, and TNF (p60) and TNF (p80) muteins on U937 (upper panel) and KYM-1 (lower panel) cells. Cells (5000/0.1ml) were cultured with different concentrations of TNF, TNF (p60), and TNF (p80) for 72 h at 37°C. During the last 6 h, cells were pulsed with thymidine(U-937), harvested, and counted or checked for cell viability by the crystal violet dye exclusion method (KYM-1) as described inMaterials and Methods.All measurements were made in triplicate.B, Cytotoxic effects of TNF, and TNF (p60) and TNF (p80) muteins on HeLa and HeLa-p80 cells. Cells (103103/0.1 ml) were cultured with different concentrations of TNF, TNF (p60), and TNF (p80) for 24 h at 37°C. Cytotoxicity was estimated by crystal violetstaining as described inMaterials and Methods. All measurements were made in triplicate.
3156 TNF SIGNALING THROUGH THE p80 RECEPTOR
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

induced PARP cleavage in p80-transfected HeLa cells but not incells expressing endogenous p60 receptor. Thus, as was the casefor cytotoxicity, PARP cleavage was mediated through the recom-binant p80 receptor but not through the endogenous receptor.
Overexpression of p80 TNF receptor mediates NF-kB activation
Both TNF and TNF (p60) activated NF-kB in U-937, ML-1a, andKYM-1 cells, but the TNF (p80) mutein had no effect on any ofthese cells (Fig. 5A). Even TNF (p80) at 10 nM (100-fold higher)failed to activate NF-kB in KYM-1 cells (Fig. 5B). Thus, endog-enous p80 receptor appears to be inactive for induction of bothapoptosis and NF-kB activation. Both TNF and TNF (p60) acti-vated NF-kB within 30 min in a dose-dependent manner in HeLacells, whereas TNF (p80) was inactive even when tested at a 100-fold excess (Fig. 5C, upper panel). In contrast, on p80-transfectedcells, TNF (p80) mutein was as active as TNF or TNF (p60) forthis cellular response (Fig. 5C,middle panel). To determine thespecificity of the TNF-mediated response, we used anti-p80 Abs,which block endogenously expressed p80-mediated signaling. Ser-endipitously, we found that when these Abs to the p80 receptorwere used to treat cells that overexpress recombinant p80 receptor,NF-kB was activated through the p80 receptor (Fig. 5C,lowerpanel). Thus, the Ab acted as a TNF agonist. These results indicate
FIGURE 4. Cleavage of PARP by TNF and muteins in different celltypes. Western blot analysis of PARP proteins in U-937, KYM-1, HeLa,and HeLa-p80 cells treated with TNF (1 nM) for 4 h (except U-937 cellsfor 2 h) at 37°C in the presence or the absence of cycloheximide (1mg/ml).Cell extracts (50mg) were electrophoresed, transferred onto nitrocellulosefilter, blotted with anti-PARP Ab, and visualized by the ECL method asdescribed inMaterials and Methods.
FIGURE 5. A, Activation of NF-kB in U937, ML-1a, and KYM-1 cellsby TNF, and TNF (p60) and TNF (p80) muteins. Cells (23 106/ml) wereincubated with 0.1 nM TNF, TNF (p60), or TNF (p80) for 30 min at 37°C.Nuclear extracts were prepared and tested for activation of NF-kB as de-scribed inMaterials and Methods.B, Dose response of induction of NF-kBin KYM-1 cells by TNF and muteins. Cells were treated and assayed foractivation of NF-kB as described above.C, Dose response of induction ofNF-kB in HeLa (upper panel) and HeLa-p80 (middle panel) cells by TNF,TNF (p60), and TNF (p80) and by anti-p80 Ab (lower panel). Cells (23106/ml) were incubated with different concentrations of TNF, TNF (p60),TNF (p80), and anti-p80 Ab for 30 min at 37°C. Nuclear extracts wereprepared and tested for activation of NF-kB as described inMaterials andMethods.
3157The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

that in contrast to endogenous p80 receptor, recombinant p80 re-ceptor can also activate NF-kB.
Overexpression of p80 TNF receptor mediates c-Jun kinaseactivation
Recently, it has been reported that overexpression of TRAF2,which associates with the p80 receptor, can activate c-Jun kinase(13, 15). Therefore, we examined the role of endogenous p80 re-ceptor in mediating c-Jun kinase activation by using TNF muteins.In U-937 cells, both TNF and TNF (p60) activated c-Jun kinase,whereas TNF (p80) had no effect (Fig. 6). Both TNF and TNF(p60) activated JNK on control HeLa cells and transfected cells,whereas TNF (p80) activated JNK only on transfected cells (Fig.6). These results demonstrate that the p80 receptor can mediatec-Jun kinase activation only when overexpressed. Our results todate suggest that endogenous p80 receptor lacks the ability to me-diate apoptosis, NF-kB activation, and c-Jun kinase activation.
Evidence of a cross-talk between p60 receptor and p80 receptor
Why there are two distinct TNF receptors and whether there is anycommunication between the two receptors with respect to signal-ing have not been established. Since both receptors appeared torespond independently of each other in our system, we examinedpossible cross-talk between the two receptors. Given that our ear-lier results indicated that these receptors may function in a syner-gistic manner (Fig. 3B, middle panel), we examined the effect ofanti-p80 Ab on TNF-, TNF (p60)-, and TNF (p80)-mediated cy-totoxicity in p80-transfected cells. The results show that anti-p80blocked not only TNF- and TNF (p80)-mediated cytotoxicity, butalso the TNF (p60)-mediated effect (Fig. 7,upper panel). Inhibi-tion of TNF (p60)-induced cytotoxicity by anti-p80 occurred in adose-dependent manner (Fig. 7,lower panel). The results withPARP cleavage also showed that the TNF (p60)-mediated effectwas blocked by anti-p80 (Fig. 4). These results provide furtherevidence in HeLa-p80 cells that to mediate its effects, TNF (p60)mutein requires the p80 receptor in addition to the p60 receptor.
It is also intriguing to note that the same anti-p80 Ab that dis-played an agonistic activity on HeLa-p80 cells for NF-kB activa-
tion (Fig. 5, lower panel) and for c-Jun kinase activation (Fig. 6,lower panel) showed antagonistic activity for cytotoxicity (Fig. 7).
Different clones of HeLa-p80 cells mediate TNF-inducedsignaling through the p80 receptor
All the results shown above were conducted with a single clone ofp80-transfected HeLa cells. To ascertain that signaling throughp80 receptor is not clone specific, we generated different clones ofHeLa-p80 both by FACS sorting and by single cell cloning. HeLa-p80 cells were sorted into different populations based on the levelof p80 receptor expression by FACS analysis. The different pop-ulations were designated A (untransfected), B (very low), C (low),D (medium), E (high), and F (unsorted). The level of p80 receptorexpression on these cells was evaluated by labeled TNF binding(Fig. 8A) and by Western blot (Fig. 8B). In cells with medium andhigh levels of p80 expression (populations D and E), TNF (p80)mutein could induce cytotoxicity (Fig. 8C), activation of NF-kB(Fig. 8D), and activation of c-Jun kinase (Fig. 8E). Thus, thesedata suggest that there is a direct correlation between increasingexpression of the p80 receptor in HeLa cells and its ability totransduce various signals. Similarly, when different clones wereisolated by single cell cloning, the level of expression of p80 (Fig.
FIGURE 6. Effect of TNF, TNF (p60), TNF (p80), and anti-p80 Ab onthe activation of c-Jun kinase in U937, HeLa, and HeLa-p80 cells. Cells(3 3 106) were incubated with 1 nM TNF, TNF (p60), TNF (p80), and 2mg/ml anti-p80 at 37°C for 10 min (U-937) or 15 min (HeLa and HeLa-p80). Cytoplasmic extracts were prepared and assayed for JNK activity asdescribed inMaterials and Methods.
FIGURE 7. Neutralization of the cytotoxic effect of TNF, TNF (p60),and TNF (p80) by anti-p80 Ab in HeLa-p80 cells (upper panel). Cells(10 3 103/0.1 ml) were cultured with 200 ng/ml of either form of TNFalong with 2mg/ml of anti-p80 Ab for 24 h at 37°C. Thelower panelshowsa dose response of anti-p80 Ab in neutralizing the TNF (p60)-inducedcytotoxicity. Cytotoxicity was estimated by crystal violet staining as de-scribed inMaterials and Methods.
3158 TNF SIGNALING THROUGH THE p80 RECEPTOR
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

9A) again correlated with NF-kB activation (Fig. 9B) and cyto-toxicity (Fig. 9C) induced by TNF (p80) mutein.
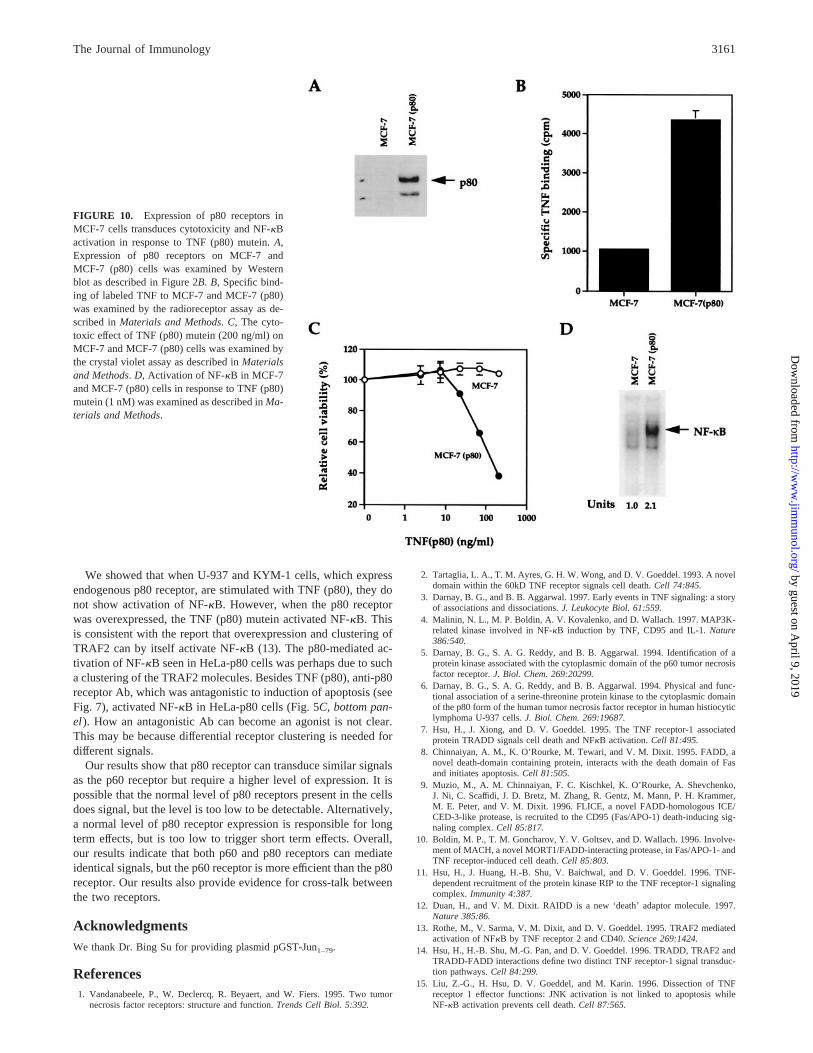
Overexpression of p80 receptor in MCF-7 cells mediatescytotoxicity and NF-kB activation
To ascertain that signaling through the recombinant p80 receptor isnot restricted to HeLa cells, we also investigated the effect of p80receptor transfection in MCF-7 cells, which normally express onlyp60 receptor. For this, MCF-7 cells were stably transfected with aninducible expression vector containing the p80 receptor (MCF-7(p80)). The p80 receptor expression was induced as evaluated byWestern blot (Fig. 10A) and by labeled TNF binding (Fig. 10B).While MCF-7 cells were completely resistant to increasing dosesof the TNF (p80) mutein, expression of the p80 receptor in MCF-7(p80) cells caused cytotoxicity in a dose-dependent manner whenexposed to TNF (p80) mutein (Fig. 10C). The expression of thep80 receptor in MCF-7 (p80) also led to activation of NF-kB by
the TNF (p80) mutein (Fig. 10D). These results suggest that over-expression of the p80 receptor leads to signaling not only in HeLacells, but also in MCF-7 cells.
DiscussionThe aim of this study was to investigate the functional role of thep80 receptor in mediating TNF-induced signaling. To examine therole of endogenous p80 receptors, we used U937 and KYM-1cells, which are known to express the maximum number of highaffinity receptors among cell lines described to date. We also usedHeLa cells and MCF-7 (which express only p60 receptor) that hadbeen transfected with p80 receptor cDNA and thus engineered toexpress higher numbers of receptors than U-937 and KYM-1 cells.In addition, to distinguish the role of the p80 receptor from that ofthe p60 receptor in TNF-mediated signaling, we used TNF mu-teins, made by site-specific mutagenesis, to bind either the p60
FIGURE 8. Correlation between the levels of p80 receptor overexpression and TNF-mediated cytotoxicity, NF-kB activation, and c-Jun kinase activationin FACS-sorted cells. Based on the level of p80 receptor expression in different populations (A, untransfected;B, very low;C, low; D, medium;E, high;F, unsorted) of HeLa-p80 cells were sorted as described inMaterials and Methods. A, Specific binding of labeled TNF to different populations examinedin triplicate as described inMaterials and Methods.B, The level of p80 receptor expression on the different cell populations examined by Western blottingas described in Figure 2B.C, The sensitivity of the different clones to TNF (p80) mutein (200 ng/ml) examined by the crystal violet method as detailedin Figure 3B.D, Activation of NF-kB by TNF (p80) mutein in the different cell populations examined after treating the cells with 1 nM p80 mutein for15 min and performing EMSA as described inMaterials and Methods. E, Activation of c-Jun kinase by TNF (p80) mutein (1 nM) in different cellpopulations examined as described inMaterials and Methods.
3159The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

(mutated at R32W and S86T) or the p80 (mutated at D143N andA145R) receptor. We studied effects on apoptosis and activation ofNF-kB and of c-Jun kinase, all previously known to be mediatedthrough the p60 receptor.
In the present report, we demonstrate that the endogenous p80receptor is incapable of inducing these three signals. However, p80receptor can, when overexpressed, mediate cytotoxicity, NF-kBactivation, and induction of c-Jun kinase in a ligand-dependentmanner. In addition, we found that overexpression of p80 receptorpotentiates p60 receptor-mediated cytotoxicity in a synergisticmanner, and p60 receptor-mediated cytotoxicity was blocked byAbs to the p80 receptor. The p80 Abs, which was antagonistic onendogenous receptor acted as an agonist on the cells overexpress-ing p80 receptor for NF-kB activation.
Although several reports indicate that the endogenous p60 re-ceptor can mediate cytotoxicity, NF-kB activation, and inductionof c-Jun kinase, very few indicate that endogenous p80 receptorcan mediate similar responses (31–37). This has led to suggestionsof an alternate role of p80 receptor including ligand passing to thep60 receptor, the receptor for transmembrane form of TNF, and thereceptor required for immunomodulation by TNF (19, 32, 33).Only when overexpressed could the p80 receptor signal for all thecellular responses examined. Induction of apoptosis by overex-pression of p80 is consistent with the findings of a previousreport (18).
Why overexpression of p80 receptor is needed for its signaling,whereas the p60 receptor can signal without overexpression is notclear. It has been shown that receptor clustering is needed forcytokine signaling, so it is possible that receptor clustering is moreefficient for the p60 receptor than for the p80 receptor. That theICD of the p60 receptor contains a DD that itself has a strongtendency to self-associate, whereas the p80 receptor lacks this do-
main, may explain the differential behavior of the two receptors.This may also explain why overexpression of the p60 receptorleads to ligand-independent signaling, whereas overexpression ofthe p80 receptor, as noted in our studies, still requires activation bythe ligand to signal.
The DD of the p60 receptor has been shown to recruit severalproteins that are involved in TNF-mediated cytotoxicity, NF-kBactivation, and induction of c-Jun kinase. How p80 receptor sig-nals without the DD binding proteins is not known. TRAF-2 andcellular IAP-1 have been shown to bind to both receptors (38–40).The potential role of TRAF-2 in NF-kB activation and induction ofJNK has been demonstrated (13–17). However, TRAF-2 plays norole in TNF-induced apoptosis. Thus, how p80 receptor can me-diate cytotoxicity independent of the p60 receptor is not clear. Ourresults also showed that p60 receptor-mediated cytotoxicity is po-tentiated by p80 receptor (see Fig. 3B) and that p60 receptor-me-diated cytotoxicity is blocked by anti-p80 (Fig. 7). How p80 re-ceptor potentiates the p60 receptor-mediated signaling is not clear.Our results, however, are in agreement with those of a recent re-port that showed enhancement of p60 receptor-mediated cytotox-icity by p80 receptor (20). The ability of p80 receptor to cooperatewith the activation of p60 receptor was found to be due to TRAF-2(20). Overexpression of TRAF-2 alone, however, does not inducecytotoxicity. The role of TRADD in p60 receptor-mediated cyto-toxicity has been demonstrated (14). It is possible that the p80receptor recruits proteins similar to TRADD that can also mediatecytotoxicity. Alternatively, it is also possible that both TRADDand TRAF2 are needed for cell killing by TNF. Ligand-inducedformation of a heterocomplex between p60 and p80 has recentlybeen demonstrated (41). In any case, our results provide evidenceof cross-talk not only at the receptor level, but also for downstreamsignaling.
FIGURE 9. Correlation between the levelsof p80 receptor overexpression and TNF-me-diated cytotoxicity and NF-kB activation indifferent clones isolated by single cell cloningof HeLa-p80 cells.A, The level of p80 receptorexpression on the different clones was exam-ined by Western blotting as described inMa-terials and Methods.B, Activation of NF-kBby TNF (p80) mutein (1 nM) was examinedafter treating the cells for 15 min and then wasanalyzed by EMSA as described inMaterialsand Methods.C, The sensitivity of differentclones to TNF (p80) mutein (200 ng/ml) wasexamined by the crystal violet assay as detailedin Materials and Methods.
3160 TNF SIGNALING THROUGH THE p80 RECEPTOR
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
We showed that when U-937 and KYM-1 cells, which expressendogenous p80 receptor, are stimulated with TNF (p80), they donot show activation of NF-kB. However, when the p80 receptorwas overexpressed, the TNF (p80) mutein activated NF-kB. Thisis consistent with the report that overexpression and clustering ofTRAF2 can by itself activate NF-kB (13). The p80-mediated ac-tivation of NF-kB seen in HeLa-p80 cells was perhaps due to sucha clustering of the TRAF2 molecules. Besides TNF (p80), anti-p80receptor Ab, which was antagonistic to induction of apoptosis (seeFig. 7), activated NF-kB in HeLa-p80 cells (Fig. 5C,bottom pan-el). How an antagonistic Ab can become an agonist is not clear.This may be because differential receptor clustering is needed fordifferent signals.
Our results show that p80 receptor can transduce similar signalsas the p60 receptor but require a higher level of expression. It ispossible that the normal level of p80 receptors present in the cellsdoes signal, but the level is too low to be detectable. Alternatively,a normal level of p80 receptor expression is responsible for longterm effects, but is too low to trigger short term effects. Overall,our results indicate that both p60 and p80 receptors can mediateidentical signals, but the p60 receptor is more efficient than the p80receptor. Our results also provide evidence for cross-talk betweenthe two receptors.
AcknowledgmentsWe thank Dr. Bing Su for providing plasmid pGST-Jun1–79.
References1. Vandanabeele, P., W. Declercq, R. Beyaert, and W. Fiers. 1995. Two tumor
necrosis factor receptors: structure and function.Trends Cell Biol. 5:392.
2. Tartaglia, L. A., T. M. Ayres, G. H. W. Wong, and D. V. Goeddel. 1993. A noveldomain within the 60kD TNF receptor signals cell death.Cell 74:845.
3. Darnay, B. G., and B. B. Aggarwal. 1997. Early events in TNF signaling: a storyof associations and dissociations.J. Leukocyte Biol. 61:559.
4. Malinin, N. L., M. P. Boldin, A. V. Kovalenko, and D. Wallach. 1997. MAP3K-related kinase involved in NF-kB induction by TNF, CD95 and IL-1.Nature386:540.
5. Darnay, B. G., S. A. G. Reddy, and B. B. Aggarwal. 1994. Identification of aprotein kinase associated with the cytoplasmic domain of the p60 tumor necrosisfactor receptor.J. Biol. Chem. 269:20299.
6. Darnay, B. G., S. A. G. Reddy, and B. B. Aggarwal. 1994. Physical and func-tional association of a serine-threonine protein kinase to the cytoplasmic domainof the p80 form of the human tumor necrosis factor receptor in human histiocyticlymphoma U-937 cells.J. Biol. Chem. 269:19687.
7. Hsu, H., J. Xiong, and D. V. Goeddel. 1995. The TNF receptor-1 associatedprotein TRADD signals cell death and NFkB activation.Cell 81:495.
8. Chinnaiyan, A. M., K. O’Rourke, M. Tewari, and V. M. Dixit. 1995. FADD, anovel death-domain containing protein, interacts with the death domain of Fasand initiates apoptosis.Cell 81:505.
9. Muzio, M., A. M. Chinnaiyan, F. C. Kischkel, K. O’Rourke, A. Shevchenko,J. Ni, C. Scaffidi, J. D. Bretz, M. Zhang, R. Gentz, M. Mann, P. H. Krammer,M. E. Peter, and V. M. Dixit. 1996. FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to the CD95 (Fas/APO-1) death-inducing sig-naling complex.Cell 85:817.
10. Boldin, M. P., T. M. Goncharov, Y. V. Goltsev, and D. Wallach. 1996. Involve-ment of MACH, a novel MORT1/FADD-interacting protease, in Fas/APO-1- andTNF receptor-induced cell death.Cell 85:803.
11. Hsu, H., J. Huang, H.-B. Shu, V. Baichwal, and D. V. Goeddel. 1996. TNF-dependent recruitment of the protein kinase RIP to the TNF receptor-1 signalingcomplex.Immunity 4:387.
12. Duan, H., and V. M. Dixit. RAIDD is a new ‘death’ adaptor molecule. 1997.Nature 385:86.
13. Rothe, M., V. Sarma, V. M. Dixit, and D. V. Goeddel. 1995. TRAF2 mediatedactivation of NFkB by TNF receptor 2 and CD40.Science 269:1424.
14. Hsu, H., H.-B. Shu, M.-G. Pan, and D. V. Goeddel. 1996. TRADD, TRAF2 andTRADD-FADD interactions define two distinct TNF receptor-1 signal transduc-tion pathways.Cell 84:299.
15. Liu, Z.-G., H. Hsu, D. V. Goeddel, and M. Karin. 1996. Dissection of TNFreceptor 1 effector functions: JNK activation is not linked to apoptosis whileNF-kB activation prevents cell death.Cell 87:565.
FIGURE 10. Expression of p80 receptors inMCF-7 cells transduces cytotoxicity and NF-kBactivation in response to TNF (p80) mutein.A,Expression of p80 receptors on MCF-7 andMCF-7 (p80) cells was examined by Westernblot as described in Figure 2B. B, Specific bind-ing of labeled TNF to MCF-7 and MCF-7 (p80)was examined by the radioreceptor assay as de-scribed inMaterials and Methods. C, The cyto-toxic effect of TNF (p80) mutein (200 ng/ml) onMCF-7 and MCF-7 (p80) cells was examined bythe crystal violet assay as described inMaterialsand Methods.D, Activation of NF-kB in MCF-7and MCF-7 (p80) cells in response to TNF (p80)mutein (1 nM) was examined as described inMa-terials and Methods.
3161The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

16. Reinhard, C., B. Shamoon, V. Shyamala, and L. T. Williams. 1997. Tumor ne-crosis factor-1 induced activation of c-Jun N-terminal kinase mediated byTRAF2. EMBO J. 16:1080.
17. Natoli, G., A. Costanzo, A. Ianni, D. J. Templeton, J. R. Woodgett, C. Balsano,and M. Levrero. 1997. Activation of SAPK/JNK by TNF receptor 1 through anoncytotoxic TRAF2-dependent pathway.Science 275:200.
18. Heller, R. A., K. Song, N. Fan, and D. J. Chang. 1992. The p70 tumor necrosisfactor mediated cytotoxicity.Cell 70:47.
19. Tartaglia, L. A., D. Pennica, and D. V. Goeddel. 1993. Ligand passing: the p80tumor necrosis factor (TNF) receptor recruits TNF for signalling by the 60-kDaTNF receptor.J. Biol. Chem. 268:18542.
20. Weiss, T., M. Grell, B. Hessabi, S. Bourteele, G. Muller, P. Scheurich, andH. Wajant. 1996. Enhancement of TNF receptor p60 cytotoxicity by TNF recep-tor p80.J. Immunol. 158:2398.
21. Chainy, G. B. N., S. Singh, U. Raju, and B. B. Aggarwal. 1996. Differentialactivation of the nuclear factor-kB by TNF muteins specific for the p60 and p80TNF receptors.J. Immunol. 157:2410.
22. Loetscher, H., D. Stueber, D. Banner, F. Mackay, and W. Lesslauer. 1993. Hu-man tumor necrosis factor-a (TNF-a) mutants with exclusive specificity for the55-kDa or 75-kDa TNF receptors.J. Biol. Chem. 268:26350.
23. Aggarwal, B. B. 1985. Human lymphotoxin.Methods Enzymol. 116:441.24. Totpal, K., R. LaPushin, T. Kohno, B. G. Darnay, and B. B. Aggarwal. 1994.
TNF and its receptor antibody agonist differ in mediation of cellular responses.J. Immunol. 153:2248.
25. Tewari, M., L. T. Quan, K. O’Rourke, S. Desnoyers, Z. Zeng, D. R. Beidler,G. G. Poirier, G. S. Salvesan, and V. M. Dixit. 1995. Yama/CPP32, a mammalianhomolog ofced-3, is acrm-A inhibitable protease that cleaves the death substratepoly(ADP-ribose) polymerase.Cell 81:801.
26. Schreiber, E., M. Matthias, M. Muller, and W. Schaffner. 1989. Rapid detectionof octamer binding proteins with mini-extracts, prepared from a small number ofcells.Nucleic Acids Res. 17:6419.
27. Singh, S., and B. B. Aggarwal. 1995. Protein tyrosine phosphatase inhibitorsblock tumor necrosis factor-dependent activation of the nuclear transcription fac-tor NF-kB. J. Biol. Chem. 270:10631.
28. Higuchi, M., and B. B. Aggarwal. 1992. Microtiter plate radioreceptor assay fortumor necrosis factor and its receptors in large numbers of samples.Anal. Bio-chem. 204:53.
29. Derijard, B., M. Hibi, I.-H. Wu, T. Barrett, B. Su, T. Deng, M. Karin, andR. J. Davis. 1994. JNK 1: a protein kinase stimulated by UV light and Ha-ras thatbinds and phosphorylates the c-jun activation domain.Cell 76:1025.
30. Zhang, L., M. Higuchi, K. Totpal, M. M. Chaturvedi, and B. B. Aggarwal. 1994.Staurosporine induces the cell surface expression of both forms of human tumor
necrosis factor receptors on myeloid and epithelial cells and modulates ligand-induced cellular response.J. Biol. Chem. 269:10270.
31. Higuchi, M., and B. B. Aggarwal. 1994. Differential roles of two types of theTNF receptor in TNF-induced cytotoxicity, DNA fragmentation and differentia-tion. J. Immunol. 152:4017.
32. Grell, M., E. Douni, H. Wajant, M. Lohden, M. Clauss, B. Maxeiner,S. Georgopoulos, W. Lesslauer, G. Kollias, K. Pfizenmaier, and P. Scheurich.1995. The transmembrane form of tumor necrosis factor is the prime activatingligand of the 80 kDa tumor necrosis factor receptor.Cell 83:793.
33. Mason, A. T., D. W. McVicar, C. A. Smith, H. A. Young, C. F. Ware, andJ. R. Ortaldo. 1995. Regulation of NK cells through the 80-kDa TNFR (CD120b).J. Leukocyte Biol. 58:249.
34. Jacobsen, F. W., M. Rothe, L. Rusten, D. V. Goeddel, E. B. Smeland,O. P. Veiby, L. Slordal, and S. E. W. Jacobsen. 1994. Role of the 75-kDa tumornecrosis factor receptor: inhibition of early hematopoiesis.Proc. Natl. Acad. Sci.USA 91:10695.
35. Vandenabeele, P., W. Declercq, D. Vercammen, M. Van de Craen, J. Grooten,H. Loetscher, M. Brockhaus, W. Lesslauer, and W. Fiers. 1992. Functional char-acterization of the human tumor necrosis factor receptor p75 in a transfectedrat/mouse T cell hybridoma.J. Exp. Med. 176:1015.
36. Laegreid, A., A. Medvedev, U. Nonstad, M. P. Bombara, G. Ranges, A. Sundan,and T. Espevik. 1994. Tumor necrosis factor receptor p75 mediates cell-specificactivation of nuclear factorkB and induction of human cytomegalovirus en-hancer.J. Biol. Chem. 269:7785.
37. Bigda, J., I. Beletsky, C. Brakebusch, Y. Varfolomeev, H. Engelmann, J. Bigda,H. Holtmann, and D. Wallach. 1994. Dual role of the p75 tumor necrosis factor(TNF) receptor in TNF cytotoxicity.J. Exp. Med. 180:445.
38. Rothe, M., S. C. Wong, W. J. Henzel, and D. V. Goeddel. 1994. A novel familyof putative signal transducers associated with the cytoplasmic domain of the 75kDa tumor necrosis factor receptor.Cell 78:681.
39. Rothe, M., M.-G. Pan, W. J. Henzel, T. M. Ayers, and D. V. Goeddel. 1995. TheTNFR2-TRAF signalling complex contains two novel proteins related to bacu-lovirus inhibitor of apoptosis proteins.Cell 83:1243.
40. Shu, H. B., M. Takeuchi, and D. V. Goeddel. 1996. The tumor necrosis factorreceptor 2 signal transducers TRAF2 and c-IAP1 are components of the tumornecrosis factor receptor 1 signaling complex.Proc. Natl. Acad. Sci. USA 93:13973.
41. Pinckard, J. K., K. C. Sheehan, and R. D. Schreibe. 1997. Ligand-induced for-mation of p55 and p75 tumor necrosis factor receptor heterocomplex on intactcells.J. Biol. Chem. 269:10784.
3162 TNF SIGNALING THROUGH THE p80 RECEPTOR
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from