p. brandon matheny & rachel a. swenie
TRANSCRIPT

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=umyc20
Mycologia
ISSN: 0027-5514 (Print) 1557-2536 (Online) Journal homepage: http://www.tandfonline.com/loi/umyc20
The Inocybe geophylla group in North America: arevision of the lilac species surrounding I. lilacina
P. Brandon Matheny & Rachel A. Swenie
To cite this article: P. Brandon Matheny & Rachel A. Swenie (2018) The Inocybe�geophylla groupin North America: a revision of the lilac species surrounding I.�lilacina, Mycologia, 110:3, 618-634,DOI: 10.1080/00275514.2018.1469880
To link to this article: https://doi.org/10.1080/00275514.2018.1469880
View supplementary material
Accepted author version posted online: 30Apr 2018.Published online: 22 Jun 2018.
Submit your article to this journal
Article views: 150
View Crossmark data

The Inocybe geophylla group in North America: a revision of the lilac speciessurrounding I. lilacinaP. Brandon Matheny and Rachel A. Swenie
Department of Ecology and Evolutionary Biology, University of Tennessee, Hesler 332, Knoxville, Tennessee 37996-1610
ABSTRACTThe Inocybe geophylla group is circumscribed based on phylogenetic analysis of DNA sequenceslargely sampled from North America and Europe. Twenty-nine phylogenetic species are uncov-ered after analysis of combined nuc 28S rDNA (28S) and RNA polymerase II second largest subunit(rpb2) DNA sequence data. Species in the I. geophylla group share the presence of a cortina, silky-fibrillose pileus and stipe, pruinose stipe apex, spermatic odor, thick-walled hymenial cystidia, andsmooth amygdaliform or elliptical basidiospores. Within the group, as many as five phylogeneticspecies attributable to I. lilacina and allies form a strongly supported clade based on analysis ofnuc ITS1-5.8S-ITS2 rDNA (ITS [internal transcribed spacer]), 28S, and rpb2 data. However, all lilac-colored species do not form a monophyletic group. Sufficient morphological and ecological dataare present to document four of the I. lilacina subgroup species, two of which are described fromNorth America as new: I. ionocephala and I. sublilacina. Inocybe lilacina is recircumscribed based onsequencing the holotype and is distributed in the eastern United States under pines and/orhardwoods. Inocybe pallidicremea is a widespread and common conifer associate in mostly north-ern parts of North America, to which the name I. lilacina was previously applied. Descriptions,photographs, line drawings, and a taxonomic key to lilac species in the I. lilacina subgroup fromNorth America are provided. Well-documented collections, especially notes on gross morphologyand ecology, are needed to continue to assess and describe the high taxonomic variation in the I.lilacina subgroup and its allies worldwide.
ARTICLE HISTORYReceived 21 November 2017Accepted 24 April 2018
KEYWORDSAgaricales; Inocybaceae;new species; systematics;taxonomy; 2 new taxa
INTRODUCTION
The Inocybe geophylla (Fr.: Fr.) P. Kumm. groupincludes some of the most common and easily recogniz-able species in the genus Inocybe (Fr.) Fr. Classic speciesin this group, apart from I. geophylla, produce basi-diomes that are white, lilac to lavender, or bear brown-ish-yellow or dark gray fibrils. All of these speciespossess a cortina and feature smooth basidiospores, theapices of which are rounded or obtuse in some species.The surface texture of the pileus and stipe is silky-fibril-lose, with the upper part of the stipe pruinose and thelower part at times bearing agglutinated fibrils. The odorof basidiomes in this group is characteristically sper-matic. In addition, all species have thick-walled hymenialcystidia and caulocystidia at the stipe apex. Members ofthe I. geophlla group were first recognized as a mono-phyletic entity in Matheny (2005) as “clade 1b” and laterby Ryberg et al. (2010) as clade XIIIb.
Species in the I. geophylla group have been variouslyclassified within Inocybe since 19th century works.Heim (1931) treated the group as “stirpe geophylla” in
section Viscosae R. Heim. Singer (1986) included thegroup in an unnamed section 5. Bon established sub-section Geophyllinae Bon within section Tardae Bon.Jacobsson and Larsson (2012) did not include any sub-sectional treatments of Inocybe but retained I. geophyllaand allies in section Tardae. Daniel E. Stuntz, in hisunpublished works on smooth-spored Inocybes, recog-nized “stirps Geophylla” within his large encompassingbut unpublished “section Inocybium.”
Species in this group were subjected to broad mor-phological species interpretations (Kuyper 1986).However, Ryberg et al. (2008) demonstrated that, asthen delimited, several species were not monophyletic,and that further taxonomic assessments were necessaryto untangle the polyphyletic morphological taxa in thegroup. Here, we address the systematic treatment of thespecies I. lilacina (Peck) Kauffman, originally describedfrom New York, the name of which has been broadlyapplied, based on morphological interpretations, tocollections in North America, Europe, Asia, andAustralia (Matheny and Bougher 2005).
CONTACT P. Brandon Matheny [email protected] data for this article can be accessed on the publisher’s Web site.
MYCOLOGIA2018, VOL. 110, NO. 3, 618–634https://doi.org/10.1080/00275514.2018.1469880
© 2018 The Mycological Society of America
Published online 22 Jun 2018

Inocybe lilacina was first acknowledged as Agaricusgeophyllus var. lilacinus Peck in a brief two-sentencestatement (Peck 1874). Peck wrote: “AGARICUSGEOPHYLLUS Sow. The variety with the pileus of a beau-tiful lilac color occurs in Bethlehem. It is Ag. affinisPers. and might appropriately be named var. lilacinus.”Gillet (1879), a few years later, made the new combina-tion based on Peck’s variety as I. geophylla var. lilacina(Peck) Gillet [as “lilacinus”]. Kauffman (1918) was thefirst to establish I. lilacina at species rank distinct fromthe white I. geophylla, noting that in Michigan the twospecies never co-occurred, and that the lilac-violaceous-colored basidiomes were sufficiently diagnostic. InNorth America, I. lilacina was recognized as an auton-omous morphological species in its own right(Kauffman 1918, 1924; Hesler 1936; Smith et al. 1979;Lincoff 1981) or at the rank of a variety of I. geophylla(Smith 1949; Nishida 1989; Bessette et al. 1995, 2007;Evenson 1997; Elliott and Stephenson 2018).
The purpose of our work is threefold: (i) to circum-scribe the I. geophylla group at large based on expandedsampling of North American taxa and estimate itstaxonomic richness and highlight clades or taxa thatdeserve further taxonomic revision; (ii) to determinethe identity of I. lilacina based on collecting efforts andby sequencing the holotype collected in the early 1870s;and (iii) to provide a taxonomic framework, with anevolutionary underpinning, to identify North Americanspecies of the I. lilacina species complex, a subgroupwithin the greater I. geophylla group. This work repre-sents the first step to revise members of this group, orsubsection Geophyllinae, in North America.
MATERIALS AND METHODS
Field sampling.—Collections made in the field weredocumented when fresh using Kornerup and Wanscher(1967), Munsell Soil Color Charts (1954), or Ridgway(1912). Gross morphological descriptions were madefrom notes based on fresh material or reconstructedfrom photographs. Fresh tissues were subject to para-dimethylaminobenzaldehyde (PDAB) macrochemicaltests when available (Matheny et al. 2013). Specimenswere dehydrated and preserved at herbaria,abbreviations of which follow Thiers [continuouslyupdated]. NYS provided digitized images of theholotype of Agaricus geophyllus var. lilacinus and aloan of the holotype.
Microscopy.—Microscopic examinations were madeon dried material. Sections were rehydrated in 5%KOH to study the morphology of basidiospores,
basidia, hymenial cystidia, stipitipellis, and pileipellis.Terminology regarding use of the terms cheilocystidia,paracystidia, pleurocystidia, caulocystidia, andcauloparacystidia follows that of Kuyper (1986).Basidiospore dimensions in excess of 2 standarddeviations from the mean are placed in parentheses.The number of total basidiospores measured (n) isindicated from x collections. Mean basidiosporelengths, widths, and Q values (quotients or lengthsdivided by widths measured in profile view) areitalicized. Cell measurements and line drawingsfollowed the methods of Braaten et al. (2014).
DNA extractions, PCR, and sequencing.—DNAextractions of dried material involved grinding 10–20 mgof material with a mortar and pestle in liquid nitrogen anduse of an E.Z.N.A. fungal DNA extraction kit (Omega Bio-Tek, Norcross, Georgia). For type specimens, we used a“High Performance” HP Fungal DNA Kit (Omega Bio-Tek) and filtered pipette tips in a laminar flow hood toreduce chances for DNA cross-contamination. Recentdried collections (<5 y old) were placed in 40 μL ofExtract-N-Amp solution (Sigma-Aldrich, St. Louis,Missouri) and incubated at room temperature (RT) for>10 min, then incubated at 95 C for 10 min, followed bymixing with an equal volume of 3% bovine serum albumin(BSA) dilution solution (Truong et al. 2017). After theaddition of the BSA solution, samples were ready forpolymerase chain reaction (PCR).
Gene sampling.—Regions of nuc ITS1-5.8S-ITS2rDNA (ITS [internal transcribed spacer]), regions ofnuc 28S rDNA (28S), and the most variable region ofRNA polymerase II second largest subunit (rpb2) wereamplified, purified, and sequenced following protocolsoutlined in Sánchez-García et al. (2014). ITS1 and ITS2were amplified and sequenced separately for types andhistorical collections >30 y. Sequence chromatogramswere assembled in Sequencher 5.0.1 (Gene CodesCorporation, Ann Arbor, Michigan).
DNA alignments, taxon sampling, and phylogeneticanalyses.—Taxa used in phylogenetic analyses and theircorresponding GenBank accession numbers are providedin TABLE 1. Newly produced sequences are in bold andwere added manually in MacClade 4.08 (Maddison andMaddison 2005) to curated alignments of 28S and rpb2produced originally inMatheny (2005) andMatheny et al.(2009). These alignments were supplemented with 28Ssequences produced by various works, namely, those ofRyberg et al. (2008, 2010). Taxa were pruned to those
MYCOLOGIA 619

Table1.
DNAsequ
encesused
inthisstud
y.GenBank
DNAsequ
ences
Taxon
Collection(Herbariu
m)
Geographicorigin
ITS
nLSU
-rRN
Arpb2
Inocybeagglutinata
WTU
1094
Washing
ton
KY99
0521
KY99
0479
—Stz1178(W
TU)
Washing
ton
KY99
0522
KY99
0480
—PBM1352
(WTU
)Washing
ton
—AY
038312
AY509113
SAT0623820
(WTU
)New
Mexico
KY99
0523
KY99
0481
—I.armeniaca
PBM1228
(WTU
)Washing
ton
—AY
380367
AY337363
SNH6(W
TU)
Washing
ton
KY99
0524
KY99
0482
—EL24606as
I.geophylla
var.lateritia
France
FN550916
FN550916
—I.fuscicothurnata
PBM3980
(TEN
N068940)
North
Carolina
MF4
8784
4KY
9904
85MF4
1640
8
AU9919
(isotyp
eWTU
)NovaScotia
KY92
3020
KY92
3039
—I.“fuscodisca”
PBM1950
Washing
ton
—AY
380376
AY337376
I.geophylla
IEL9005
Sweden
AM882870
AM882870
—JV6374
(WTU
)Finland
—AY
380377
AY333777
I.geophylla
IIEL8003
Sweden
AM882877
AM882877
—I.“geophylla”
SAT0308001
(WTU
)California
KY99
0530
KY99
0486
—WT11
California
—DQ273437
—MTS2811
(UC)
California
KY99
0531
——
PBM3040
(TEN
N062791)
California
KY99
0532
KY99
0488
MF4
1640
9
PBM3041
(TEN
N062792)
California
KY99
0533
KY99
0489
MF4
1641
0
LG496(W
TU)
California
KY99
0534
KY99
0490
—SA
T0630802
(TEN
N071578)
Washing
ton
KY99
0535
JN974952
MF4
1641
1
I.“geophylla”
CA1882
(WTU
)Orego
nKY
9905
36JN974951
—I.“geophylla”
PBM546(W
TU)
Washing
ton
KY99
0537
KY99
0491
—040904av27
New
foun
dlandandLabrador
KY99
0538
KY99
0492
—I.“geophylla”
REH7879
CostaRica
KY99
0539
JN974953
—I.“geophylla”
110924av05
(TEN
N070836)
New
foun
dland&Labrador
KY99
0542
KY99
0496
MF4
1641
5
PBM2732a
(TEN
N062544)
Colorado
KY99
0543
KY99
0497
MF4
1641
6
I.“geophylla”
CCB147
(TEN
N068276)
Tenn
essee
KY99
0544
KY99
0498
MF4
1641
7
I.“geophylla”
100823av02
(TEN
N071463)
New
foun
dland&Labrador
KY99
0545
KY99
0499
MF4
1641
8
I.insin
uata
MGW783
(TEN
N063977)
California
KY99
0546
KY99
0500
MF4
1641
9
I.aff.insin
uata
JMB122111-04
(TEN
N066938)
Washing
ton
KY99
0547
KY99
0501
MF4
1642
0
OC031608
(TEN
N063620)
British
Columbia
KY99
0548
KY99
0502
—
I.ionocephalasp.n
ov.
MTS2488
(UC)
California
KY99
0549
JN974950
—PBM3043
(TEN
N062794)
California
KY99
0550
KY99
0503
MF4
1642
1
PBM3049
(holotyp
eTENN062799)
California
KY99
0551
KY99
0504
MF4
1642
2UC1859626
California
MF4
9043
9—
—UC1859629
California
KY99
0552
——
I.lilacina
Peck
s.n.
(holotyp
eNYSf1711)
New
York
MH02
4860
——
JM96/25
North
Carolina
—AF
042616
—Pinusmycelia
North
Carolina
EF619676
——
(Con
tinued)
620 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

Table1.
(Con
tinued).
GenBank
DNAsequ
ences
Taxon
Collection(Herbariu
m)
Geographicorigin
ITS
nLSU
-rRN
Arpb2
PBM3982
(TEN
N068443)
North
Carolina
KY99
0527
——
TFB12747
(TEN
N061204)
North
Carolina
KY99
0526
——
PBM2590
(TEN
N062429)
Tenn
essee
EU523556
——
PBM2628
(TEN
N062463)
Tenn
essee
KY99
0525
KY99
0483
MF4
1640
6
PBM2629
(TEN
N062464
Tenn
essee
KY99
0528
KY99
0484
MF4
1640
7
PBM3940
(TEN
N068443)
Tenn
essee
KY99
0529
——
VRH13
(TEN
N073103)
Tenn
essee
MG66
3234
——
I.lilacinasensuLarssonI
EL9205
Finland
AM882873
AM882873
—EL175-06
asI.geophylla
Scotland
UDB
002393
——
EL5004
asI.geophylla
var.lilacina
Sweden
AM882869
AM882869
—I.lilacinasensuLarssonII
EL12605
Sweden
AM882875
AM882875
—I.“lilacina”
PK3575
(UBC
)British
Columbia
HQ604292
HQ604292
—I.“lilacina”
Y55
China
—KU
764688
—I.pallidicrem
eaPiceasoilsample3329H20
Alaska
KF61782
——
PBM4060a
(TEN
N071254)
Arizon
aMG42
9695
——
DBG
001183
Colorado
MG42
9696
——
DBG
004919
Colorado
MG42
9697
——
DBG
014326
Colorado
MG42
9698
——
DBG
023850
Colorado
MG42
9699
——
DBG
027710
Colorado
MG42
9700
——
PBM2744
(TEN
N062552)
Maine
KY99
0553
KY99
0505
MF4
1642
3
DBG
001716
Michigan
MG42
9701
——
PBM2445
(TEN
N063879)
New
York
—KY
9905
06MF4
1642
4
PBM2448
(TEN
N062757)
New
York
HQ201357
HQ201357
MF4
1642
5
OSC1064044
asI.geophylla
Orego
nEU
525951
——
OSC1064214
asI.geophylla
Orego
nEU
525981
——
BM381#10
(TEN
N063535)
Washing
ton
KY99
0554
KY99
0507
—
PBM817(W
TU)
Washing
ton
KY99
0555
KY99
0508
—PBM2039
(WTU
)Washing
ton
—AY
380385
AY337388
DBG
018072
Wyoming
MG42
9702
——
PseudotsugaECM
British
Columbia
EU645611
——
PseudotsugaECM
British
Columbia
EF218776
——
UBC
F16253
asI.geophylla
British
Columbia
EF530936
EF530936
—
UBC
F19540
British
Columbia
HQ604296
HQ604296
—AC
AD11600(isotyp
eI.pallidicrem
ea)
NovaScotia
KY92
3033
KY92
3042
—
(Con
tinued)
MYCOLOGIA 621

Table1.
(Con
tinued).
GenBank
DNAsequ
ences
Taxon
Collection(Herbariu
m)
Geographicorigin
ITS
nLSU
-rRN
Arpb2
GM5-300
(TEN
N070835)
New
foun
dland&Labrador
—KY
9905
09MF4
1642
6
GM4-222
(TEN
N070832)
New
foun
dland&Labrador
—KY
9905
10MF4
1642
7
I.phaeodiscavar.geophylloides
MD08100106
France
—KY
9905
11—
I.posterula
EBJ051120
Sweden
AM882868
AM882868
—EM
6.1
France
EU711171
EU711171
—JV2527F(W
TU)
asI.xanthodisca
Finland
KY99
0556
KY99
0512
—
I.aff.posterula
CLC2998A
(TEN
N069518)
Mon
tana
MF4
9044
0KY
9905
13MF4
1642
8
L3BD
11California
EF417810
EF417810
—I.pudica
MGW721
(TEN
N066724)
California
KY99
0557
KY99
0514
—
PBM2732b
(TEN
N062544)
Colorado
KY99
0558
KY99
0515
—
SAT0630804
(TEN
N071580)
Washing
ton
KY99
0559
KY99
0516
—
SAT0732301
(TEN
N071579)
Washing
ton
KY99
0560
KY99
0517
—
PBM1373
(WTU
)Washing
ton
—AY
038323
AY337394
EL15905
Sweden
AM882872
AM882872
—SJ06012as
I.whitei
Sweden
FN550915
FN550915
—I.sambucella
PBM2913
(TEN
N062774)
Verm
ont
—KY
9904
93MF4
1641
2
PBM2456
(TEN
N063623)
New
York
KY99
0540
KY99
0494
MF4
1641
3
PBM2457
(TEN
N062431)
New
York
KY99
0541
KY99
0495
MF4
1641
4
Inocybesp.Q
UE03
HRL2223
(TEN
N071128)
Quebec
KX89
7427
KY99
0518
—
I.sublilacina
sp.n
ov.
PBM2716
(TEN
N062531)
Colorado
KY99
0561
JN974949
MF4
1642
9
PBM2730
(holotyp
eTENN062542)
Colorado
—KY
9905
19MF4
1643
0GM5-302
(TEN
N071464)
New
foun
dlandandLabrador
KY99
0562
KY99
0520
MF4
1643
1
New
lyreleased
sequ
encesarehigh
lighted
inbo
ld.
622 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

affiliated with “clade 1b” identified in Matheny (2005),which correspond to the I. geophylla and I. flocculosagroups at large. Inocybe kauffmanii A.H. Sm. was usedfor outgroup purposes. All sites were included foranalyses. After inspection for intergene conflict, the 28Sand rpb2 alignments were concatenated. Phylogeneticanalyses were conducted in RAxML 8.2.9 (Stamatakis2014) under the maximum likelihood (ML) criterionusing a GTRGAMMA model as recommended in theRAxML user manual. The concatenated alignment waspartitioned by 28S, rpb2 gene codon positions, and onerpb2 intron region. One thousand bootstrap replicateswere performed. Bootstrap values >50% are shown onresulting tree figures.
ITS+28S+rpb2 sequences of members of the I. lila-cina subgroup, as identified by the 28S+rpb2 analysis,were aligned in Clustal X 2.0.9 (Larkin et al. 2007) andinspected in MacClade. This data set was assembled toinclude nuc rDNA sequences from type collections anda selection of those available on GenBank. All sites wereincluded in this analysis. For this alignment, sequenceslabeled as I. lilacina that clustered outside the I. lilacinagroup in the 28S+rpb2 analysis were used for outgrouppurposes. Phylogenetic analyses were conducted asdescribed above but partitioned by ITS+28S+rpb2-intron 4 data and by rpb2 codon position. Thus, fourdifferent partitions were used in analyses of this dataset. A third data set was examined, including 51 taxabut only ITS+28S data. This data set incorporatednumerous ITS- or 28S-only sequences, including fromenvironmental samples. For phylogenetic analyses, asingle model partition was used.
Bayesian Inference (BI) phylogenetic analyses werealso performed on the three alignments in MrBayes3.2.6 (Ronquist et al. 2012). Models were partitionedas in the RAxML analyses, but model selection wasbased on Matheny (2005). The 28S+rpb2 data set wasrun for 5 million generations, sampling trees every 5000steps. The ITS+28S+rpb2 data set was run for 2 milliongenerations, sampling trees every 500 steps. The ITS+28S data set was run for 1 million generations, sam-pling trees every 500 steps. Convergence diagnosticswere observed and length of analyses run as recom-mended in the user manual. The first 25% of the treesfrom two independent runs for each data set werediscarded as the burn-in prior to calculation of poster-ior probabilities (PPs). PP values >0.95 are reported.Alignments and tree files are available at two locations:http://mathenylab.utk.edu/Site/Alignments_%26_Data_Sets.html and at TreeBASE as submission 21910.
Genetic distances were measured for the ITS andrpb2 loci separately using the “showdist” command inPAUP 4.0b10 for Unix (Swofford 2002). This command
produces a pairwise uncorrected or proportional (“p”)distance matrix.
RESULTS
Fifty-eight ITS, 43 28S, and 26 rpb2 sequences wereproduced during this study (127 total; see TABLE 1).No strongly supported intergene conflict was detectedwhen comparing individual gene topologies; thus, the28S and rpb2 data were combined. The 28S+rpb2 align-ment contained 79 taxa and 2185 included sites. Eachtaxon was represented by a 28S sequence, and 52 taxawere represented by rpb2 sequences. The combined ITS+28S+rpb2 alignment included 33 taxa and 2960 sites.In this data set, 28 taxa were represented by ITSsequences, 27 by 28S, and 16 by rpb2. In the BI ana-lyses, 15 002 trees were sampled to calculate PPs for the28S+rpb2 data set and 6002 trees sampled to calculatePPs for the ITS+28S+rpb2 data set after discarding thefirst 25% of trees sampled from both analyses.
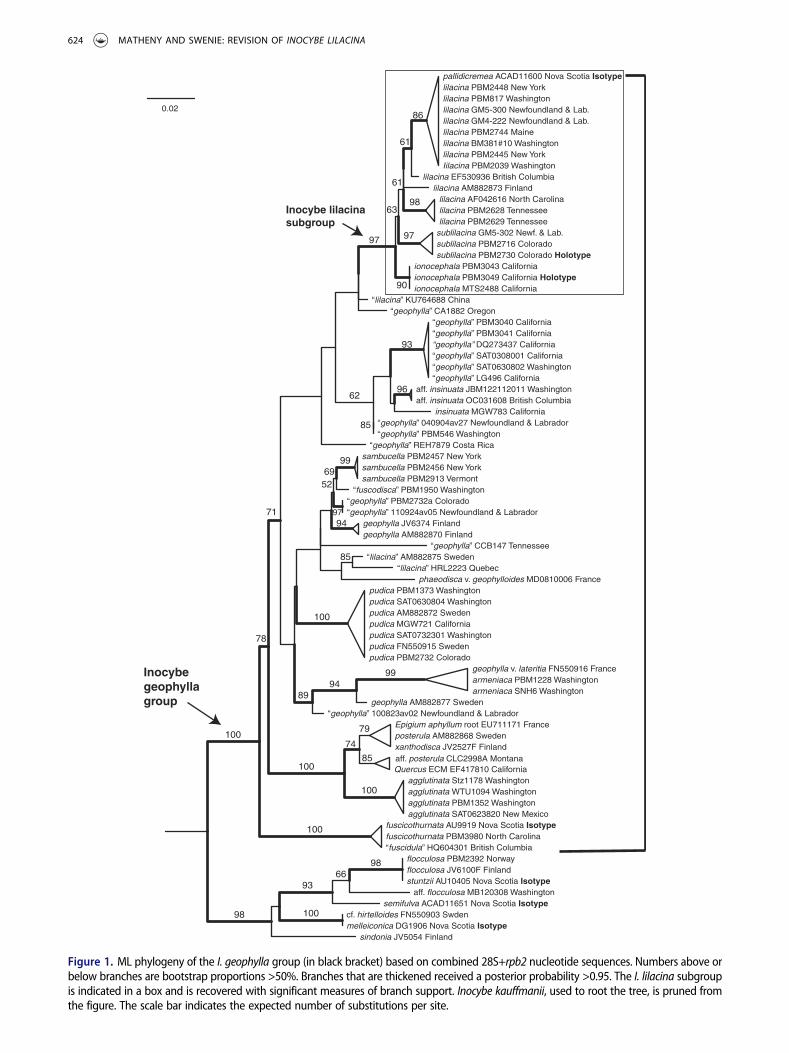
Phylogenetic analysis of the 28S+rpb2 data set (FIG. 1)resulted in strong support for the I. geophylla group orclade Ib of Matheny (2005), as indicated by the bracket.Considerable taxonomic diversity amounting to 29 phy-logenetic species in the I. geophylla group was detectedfrom samples in North America, Europe, and East Asia.At least 11 morphological taxa are recognizable membersof the I. geophylla group. These include I. agglutinataPeck, I. fuscicothurnataGrund &D.E. Stuntz, I. fuscodisca(Peck) Massee, I. geophylla, I. insinuata Kauffman, I.lilacina, I. pallidicremea Grund & D.E. Stuntz, I. phaeo-disca var. geophylloides Kühner, I. posterula (Britzelm.)Sacc., I. pudica Kühner, and I. xanthodisca Kühner. BothI. geophylla and I. lilacina, however, comprise eight tonine phylogenetic species-level lineages each and are poly-phyletic, reinforcing earlier finds of their nonmonophylyby Ryberg et al. (2008). In contrast, the species I. pudica,although sampled from a wide geographic range includ-ing western North America and northern Europe, forms acohesive monophyletic group. Inocybe fuscicothurnata,earlier names for which may be I. virgata G.F. Atk. andI. fuscodisca (Peck) Massee, is found to be sister to theremainder of the I. geophylla group with strong support.
Six separate phylogenetic lineages are recovered inwhat we call the I. lilacina subgroup, a strongly sup-ported clade of mostly lilac-colored species occurringpredominantly in North America (FIG. 1). This cladeincludes I. lilacina in the strict sense based on analysisof ITS sequences from the holotype collected in NewYork and collections from North Carolina andTennessee (FIG. 2). Two single stem lineages, repre-sented by 28S sequence data only, are labeled I. lilacinafrom British Columbia and Finland. These branch
MYCOLOGIA 623
0.02
geophylla AM882877 Sweden
aff. flocculosa MB120308 Washington
sublilacina GM5-302 Newf. & Lab.
sublilacina PBM2730 Colorado Holotype
“geophylla” 100823av02 Newfoundland & Labrador
“lilacina” AM882875 Sweden
“geophylla” PBM2732a Colorado
ionocephala MTS2488 California
“geophylla” SAT0630802 Washington
“fuscodisca” PBM1950 Washington
lilacina PBM2445 New York
“geophylla” LG496 California
geophylla v. lateritia FN550916 France
pallidicremea ACAD11600 Nova Scotia Isotype
lilacina PBM2628 Tennessee
“geophylla” 040904av27 Newfoundland & Labrador
flocculosa PBM2392 Norway
“geophylla” REH7879 Costa Rica
“geophylla” 110924av05 Newfoundland & Labrador
pudica PBM1373 Washington
“lilacina” KU764688 China
pudica SAT0630804 Washington
Quercus ECM EF417810 California
sublilacina PBM2716 Colorado
sindonia JV5054 Finland
lilacina PBM2629 Tennessee
pudica MGW721 California
aff. insinuata JBM122112011 Washington
xanthodisca JV2527F Finland
lilacina AM882873 Finland
stuntzii AU10405 Nova Scotia Isotype
sambucella PBM2456 New York
aff. posterula CLC2998A Montana
“geophylla” CCB147 Tennessee
flocculosa JV6100F Finland
pudica FN550915 Sweden
cf. hirtelloides FN550903 Swden
agglutinata Stz1178 Washington
“geophylla” PBM546 Washington
agglutinata SAT0623820 New Mexico
aff. insinuata OC031608 British Columbia
sambucella PBM2913 Vermont
lilacina GM4-222 Newfoundland & Lab.
lilacina AF042616 North Carolina
Iilacina PBM2039 Washington
ionocephala PBM3043 California
fuscicothurnata AU9919 Nova Scotia Isotype
armeniaca SNH6 Washington
geophylla AM882870 Finland
phaeodisca v. geophylloides MD0810006 France
sambucella PBM2457 New York
geophylla JV6374 Finland
pudica SAT0732301 Washington
posterula AM882868 Sweden
ionocephala PBM3049 California Holotype
“geophylla” PBM3040 California
lilacina PBM2448 New York
lilacina PBM2744 Maine
Epigium aphyllum root EU711171 France
lilacina EF530936 British Columbia
“lilacina” HRL2223 Quebec
“geophylla” DQ273437 California
“fuscidula” HQ604301 British Columbia
lilacina PBM817 Washington
semifulva ACAD11651 Nova Scotia Isotype
agglutinata WTU1094 Washington
“geophylla” CA1882 Oregon
pudica PBM2732 Colorado
agglutinata PBM1352 Washington
lilacina GM5-300 Newfoundland & Lab.
“geophylla” SAT0308001 California
armeniaca PBM1228 Washington
“geophylla” PBM3041 California
fuscicothurnata PBM3980 North Carolina
melleiconica DG1906 Nova Scotia Isotype
lilacina BM381#10 Washington
pudica AM882872 Sweden
insinuata MGW783 California
86
85
62
94
85
93
63
98
69
97
98
100
61
85
94
66
98
99
100
90
61
100
93
97
78
96
97
74
100
100
100
99
71
89
52
79
Inocybe lilacinasubgroup
Inocybe geophyllagroup
Figure 1. ML phylogeny of the I. geophylla group (in black bracket) based on combined 28S+rpb2 nucleotide sequences. Numbers above orbelow branches are bootstrap proportions >50%. Branches that are thickened received a posterior probability >0.95. The I. lilacina subgroupis indicated in a box and is recovered with significant measures of branch support. Inocybe kauffmanii, used to root the tree, is pruned fromthe figure. The scale bar indicates the expected number of substitutions per site.
624 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

separately from the other species-level lineages. Threeother clades that correspond to the North Americanspecies I. pallidicremea, I. ionocephala, sp. nov., and I.sublilacina, sp. nov. are recovered, each with high mea-sures of branch support.
Phylogenetic analysis of the ITS+28S+rpb2 data setresulted in recovery of at least five phylogenetic speciesin the I. lilacina subgroup, all of which are character-ized by basidiomes with lilac pigmentation (FIG. 2). Ofthese, we have sufficient documentation to present four
0.0080
sublilacina PBM2716 Colorado
lilacina BM381#10 Washington
lilacina GM5-300 Newfoundland & Lab.
lilacina KP454032 British Columbia
“geophylla” PBM3040 California
pallidicremea ACAD11600 Nova Scotia (isotype)
“lilacina” HRL2223 Quebec
“lilacina” EL12605 Sweden
lilacina PBM2629 Tennessee
ionocephala PBM3049 California (holotype)
sublilacina PBM2730 Colorado (holotype)
lilacina UDB002393 Scotland
ionocephala MTS2488 California
lilacina PBM817 Washington
lilacina PBM2448 New York
lilacina KP406571 British Columbia
ionocephala PBM3043 California
lilacina NYSf1711 New York (holotype)
“lilacina” PK3575 Brit. Col.
Picea soil sample KF617872 Alaska
“geophylla” SAT0630802 Washington
“geophylla” PBM3041 California
lilacina PBM2744 Maine
lilacina PBM3940 Tennessee
lilacina EL5004 Sweden
lilacina PBM2039 Washington
sublilacina GM5-302 Newfoundland & Lab.
lilacina PBM3982 North Carolina
lilacina EL9205 Sweden
lilacina PBM2628 Tennessee
lilacina HQ604296 British Columbia
lilacina GM4-222 Nova Scotia
lilacina PBM2445 New York
100
83
100
100
89
78
78
99
81
71
100
100
Inocybepallidicremea
Inocybe lilacina sensu stricto
Inocybe lilacina sensu Larsson I
Inocybesublilacina sp. nov.
Inocybeionocephala sp. nov.
Inocybe lilacina sensu lato
Inocybe lilacina sensu Larsson II
Inocybe lilacinasubgroup
Figure 2. ML phylogeny of the I. lilacina subgroup based on combined ITS+28S+rpb2 nucleotide sequences. Numbers above branches arebootstrap proportions >70%. Branches that are thickened received a posterior probability >0.95. The scale bar indicates the expectednumber of substitutions per site.
MYCOLOGIA 625

North American species, each receiving strong branchsupport, in taxonomic detail below.
Many samples attributed to I. lilacina, mainly from thenorthern United States and western and eastern Canada,cluster with type of I. pallidicremea described from NovaScotia (FIGS. 1, 2; SUPPLEMENTARY FIG. 1). Mostcollections from the Rocky Mountains and ColoradoPlateau labeled as I. lilacina also cluster with I. pallidicre-mea (SUPPLEMENTARY FIG. 1). A distinct clade ofsoutheastern US samples labeled I. lilacina is recoveredwith strong support together with a partial ITS sequenceof the type of Peck’s Agaricus geophyllus var. lilacinusfrom New York (FIG. 2). In all three data sets, samplesof I. sublilacina from eastern Canada and the RockyMountains cluster together with strong support. In addi-tion, samples from California form a monophyletic groupwith strong support—these are labeled I. ionocephala, sp.nov. Sequences from environmental samples and basi-diomes collected in pure conifer stands suggest that I.pallidicremea is ectomycorrhizal with Picea,Pseudotsuga, and/or Tsuga (SUPPLEMENTARY FIG. 1).Similar environmental and ecological data suggest that I.lilacina in the strict sense is ectomycorrhizal at least withPinus (SUPPLEMENTARY FIG. 1) and with Fagales.TABLE 1 indicates the species assignment of all DNAsequences analyzed.
Infraspecific variation measured as uncorrected “p”distances at the rpb2 locus for I. ionocephala, I. lilacina,I. pallidicremea, and I. sublilacina ranged from 0% toless than 1%. Interspecific variation or “p” distances atthe rpb2 locus ranged from 1% to 2.5%. Infraspecificvariation at the ITS locus measured less than 1%,whereas interspecific distances ranged between 1.6%and 2.8% in all comparisons except for those betweenI. lilacina and I. pallidicremea, which exhibited low ITSvariation, between 0.2% and 0.8%.
TAXONOMY
Inocybe pallidicremea Grund & D.E. Stuntz, Mycologia69:399. 1977. FIGS. 3A–B, 4A–B
Description: Pileus 10–40 mm wide, obtusely conicalto broadly convex with a nipple-like umbo, becomingplane or even somewhat depressed with age, margindecurved; surface of umbo smooth or with agglutinatedfibrils, silky-fibrillose towards the margin, becomingrimose, velipellis not observed, dry to tacky subviscid;in youth lilac to pale lilac (14B4–B3–C4; reddish lilac tograyish lilac) in entirety but fading with age; at times orin age the umbo yellowish brown to mustard brown orbrown (5E8–E7–E6), rarely smoky gray (5YR 4/1–3/1),becoming grayish yellow to brownish orange (4B5–5C6) around the umbo, this dissipating towards the
margin into a grayish-white color, at times maintaininggrayish-lilac fibrillose streaks; context white, thin, up to3–4 mm thick under the umbo, not changing colorwhere cut or bruised, odor spermatic to strongly so;pileus surface negative with PDAB. Lamellae adnexed,uncinate, or adnate, close, with several tiers of lamellu-lae, light gray to gray when young, becoming palebrown to brown (5D4–D5) with age, subventricose,edges white-fimbriate. Stipe 35–60 × 3–6 mm at theapex, flexuous, clavate-bulbous towards the base, this7–9 mm wide, upper 1/6–1/8 pruinose; cortina present,cortinate fibrils collapsed and forming a ring zone attimes or fugacious, more or less smooth towards thebase or finely fibrillose; lilac to pale lilac at first, thissoon fading and becoming whitish overall, rarely withgrayish-brown streaks (6D3) beneath the insertion ofthe cortina and above the base, or with agglutinatedgray fibrils above the base, the base itself (pale) yellowor with brownish-orange or grayish-yellow tones (5C5–4B5); context solid, whitish.
Basidiospores (7–)7.5–9.0–10.5(–11) × 4.5–5.3–6.0μm, Q = (1.40–)1.44–1.70–1.96(–2.11) (n = 175/17),smooth, mostly amygdaliform to subamygdaliform, attimes elliptical, apices often bluntly pointed or obtuse,apiculus small but distinctive, yellowish brown, slightlythick-walled. Basidia 29–33 × 8–10 μm, 4-sterigmate,clavate, hyaline. Pleurocystidia 49–73 × 11–20 μm, fusi-form-ventricose, often with a slender basal pedicel, thick-walled (walls 1.0–3.0 μm thick), hyaline, apices often bareor weakly crystalliferous. Cheilocystidia similar to pleur-ocystidia but some shorter and more ventricose or sac-cate, densely arranged, paracystidia infrequent.Caulocystidia similar to hymenial cystidia but subfusi-form, ventricose, or clavate, these 30–110 × 9–18 μm,mixed occasionally with cauloparacystidia. Pileipellis aninterwoven cutis of smooth, hyaline hyphae, these slightlythick-walled or thin-walled, mostly 4–8 μm wide, hyalinein mass. Clamp connections present.
Ecology and distribution: Scattered singly or ingroups on soil in western states and provinces: Alaska,British Columbia, Washington, Oregon, Wyoming,Colorado, and Arizona, eastward to Michigan, NewYork, New England, and the eastern provinces ofNewfoundland and Labrador and Nova Scotia (type);associated with conifers including Pseudotsuga, Picea,Tsuga, and/or Pinus. Occurring Aug–Dec.
Illustrations: Smith (1949, as I. geophylla var. lilacina);Lincoff (1981, as I. lilacina); Baroni (2017, as I. geophylla).
Specimens examined: CANADA. NEWFOUNDLANDAND LABRADOR: Gros Morne National Park, StucklessPond (49.4299, −57.7103), in moss on ground underPicea, 4 Sep 2005, M. Prior & D. Malloch GM5-300(TENN 070835); ibid., on soil in coniferous woods, 18
626 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

Figure 3. Basidiomes of the Inocybe lilacina subgroup in North America. A. Inocybe pallidicremea (PBM2448). B. Faded specimens of I.pallidicremea (PBM1743). C. Inocybe lilacina (PBM3982). D. Inocybe lilacina (PBM3940; photo, B.P. Looney). E. Inocybe ionocephala(PBM3049, holotype). F. Inocybe ionocephala (UC1859626; photo, K. Peay). G. Inocybe sublilacina (PBM2730, holotype). H. Inocybesublilacina (GM5-302; photo, R. Smith). Bars = 10 mm.
MYCOLOGIA 627

Sep 2004, N. Siegel & L.L. Norvell GM4-222 (TENN070832); Twillingate Foray, on soil under Abies in con-iferous woods, 11 Sep 2009, MS10-007 (TENN 071726);NOVA SCOTIA: Kings Co., Kentville, AgriculturalExperimental Station (45.0692, −64.4781), in woodedarea under conifers, 7 Sep 1966, D. Grund ACAD 11600(isotype of Inocybe pallidicremea); ONTARIO: York Co.,N. Summit Golf Club, 7 Oct 1936, H.S. Jackson HSG9619(TENN 010331). USA. ARIZONA: Coconino Co., LocketMeadow Trail (35.3586, −111.6208), on soil under con-ifers and Populus, 2896 m, 6 Aug 2015, P.B. MathenyPBM4060a (TENN 071254); COLORADO: White RiverNational Forest, Braille Trail on Independence Pass (long-itude, latitude not available), on soil or dead wood underPicea, 9 Aug 2013,H. Burgess (DBG 027710); Summit Co.,Keystone, River Run Lodge, 2150 m (39.60694,−105.94028); Eagle Co., White River National Forest,Sam’s Ranch (39.573878, −106.61574), in moss and litterunder Picea, 2745 m, 22 Jul 1967, S. Mitchell (DBG001183); Gilpin Co., Roosevelt National Forest, Perigo,2865 m (39.8922, −105.5303), 13 Aug 1974, A.H. Smith,Mitchell & Chapman (DBG 004919); Pitkin Co., ElkCamp, Snomass Village, 2591 m (39.1881, −106.9353),under Picea, 10 Aug 1979, V.S. Evenson (DBG 014326);similar locality as previous, 2590 m (39.2408, −106.9822),in moss under Picea andAbies, 28 Aug 1980,V.S. Evenson(DBG 017522); Teller Co., Clyde Campground (38.7308,−105.0254), on soil in grass near conifers, 10 Aug 2007, L.Barzee (DBG 023850); MAINE: Bar Harbor, Harbor Trail(44.3875, −68.20389), on ground next to trail under Piceaand Betula, 18 Sep 2005, P.B. Matheny PBM2744 (TENN062552); MICHIGAN:Montmorency Co., northwest area(45.0993, −84.2233), in boggy area, 22 Aug 1967, D.H.Mitchel & M. Wells (DBG 001716); OREGON: Linn Co.,Crescent Mountain Trail (44.42930, −122.03024), on soilunder conifers, 23 Oct 1999, P.B. Matheny PBM1743(WTU); NEW YORK: South Bethlehem, JoralemonMemorial Park, 4.6 miles south of Brewer Station(42.53222, −73.84611), on soil in mixed woods underPinus strobus, Juniperus, Carya, Quercus, 20 Sep 2006, K.Bushley PBM2445 (TENN 063879); ibid., gregarious toscattered or in small pairs or clusters on soil under Pinusstrobus on edge of mixed forest, 20 Sep 2006, P.B.Matheny PBM2448 (TENN 062757); WASHINGTON:Island Co., Whidbey Island, Ebbey’s Landing, 35 m, onsoil under Pseudotsuga, 14 Nov 1998, P.B. MathenyPBM1360 (WTU); Island Co., Whidbey Island, Ebbey’sLanding, 35 m (48.2292, −122.7073), on soil underPseudotsuga, 14 Nov 1998, P.B. Matheny PBM1353(WTU); King Co., Sammamish, Pine Lake State Park(47.58760, −122.04429), on ground under Pseudotsuga, 5Nov 2000, P.B. Matheny PBM2039 (WTU); King Co.,Lincoln Park (47.5306, −122.3960), in mossy grass under
Figure 4. Line drawings of pleurocystidia and basidiospores of fourNorth American species in the I. lilacina subgroup. A–B.Pleurocystidia and basidiospores of I. pallidicremea (PBM2448). C–D Pleurocystidia and basidiospores of I. lilacina (PBM3940). E–F.Pleurocystidia and basidiospores of I. ionocephala (PBM3049, holo-type). G–H. Pleurocystidia and basidiospores of I. sublilacina(PBM2730, holotype). Bars = 10 μm.
628 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

Abies grandis, 21 Dec 2011, J.M. Birkebak JMB122111-05(TENN 066939); Kittitas Co., Crystal Springs Campround(47.30955, −121.31370), on soil under Abies, Pseudotsuga,Thuja, 12 Oct 1997, P.B. Matheny PBM781 (WTU);Kitsap Co., Seabeck (47.6409, −122.8286), scattered togregarious on soil under Pseudotsuga, Tsuga,Rhododendron, Vaccinium, 25 Oct 1997, P.B. MathenyPBM817 (WTU); ibid., on soil under Pseudotsuga,Tsuga, 21 Nov 1998, P.B. Matheny PBM1364 (WTU);Whatcom Co., Bellingham Fall Mushroom Show, 20 Oct2007, B. McAdoo BM381#10 (TENN 063535); San JuanCo., San Juan Island, Friday Harbor (48.5343, −123.0171),12 Nov 1962 (TENN 026020); ibid., on soil under con-ifers, 3 Nov 2001, P.B. Matheny PBM2237 (WTU); samelocality and date as previous, on soil under conifers, P.B.Matheny PBM2238 (WTU); Thurston Co., Olympia,Priest Point Park (47.0723, −122.8954), on soil underPseudotsuga, Tsuga, Abies, 31 Oct 1998, P.B. MathenyPBM1334 (WTU); WYOMING: Teton Co., FlaggRanch-Ashton Rd., Teton National Forest, 2285 m, onsoil near Picea, 30 Aug 1995, V.S. Evenson (DBG 018072).
Notes: Based on phylogenetic results and currentsampling, I. pallidicremea appears to be known onlyfrom western and northern North America, associatedwith conifers, and can be misinterpreted for I. geophyllawhen faded. The species was long interpreted under thename I. lilacina or as I. geophylla var. lilacina. With ageor in dry conditions, however, basidiomes of this spe-cies become pale yellow with a darker-colored umbo(brown, brownish yellow, or even grayish), losing alllilac coloration including from the stipe. Indeed, I.pallidicremea was described based on specimens inwhich the lilac colors must have been completely lost(Grund and Stuntz 1977; as in FIG. 3B). The combina-tion of a darker-colored umbo and the yellow- tocream-colored stipe base are clues that can be used todistinguish faded forms of I. pallidicremea from white-colored basidiomes of I. geophylla in the field.
Inocybe lilacina, documented below based on thetype and southeastern specimens, can be distinguishedfrom I. pallidicremea by the dark purple or dark violetcolors that persist at the center of the pileus, smallerbasidiome size, and eastern US distribution (New York,North Carolina, Tennessee). The distribution of I. pal-lidicremea and I. lilacina, based on present collectionsconfirmed by molecular data (SUPPLEMENTARY FIG.1), overlaps only in New York. Inocybe ionocephala,described below, is known thus far only from theCoastal Redwood zone of northern California and dis-tinguished morphologically by its white stipe and stipebase and larger basidiome size. Inocybe sublilacina, asecond new species described below, is best distin-guished from I. pallidicremea by genetic divergence at
multiple loci. The basidiospores of I. sublilacina aresomewhat larger than those of I. pallidicremea on aver-age (9.4 × 5.7 μm vs. 9.0 × 5.3 μm), a difference that isprobably too subtle to detect for practical purposes.However, we did observe that the basidiospores of I.sublilacina are more often elliptical than amygdaliform,unlike those of I. pallidicremea, which is typically char-acterized by amygdaliform basidiospores. Basidiomes ofI. sublilacina are lilac throughout when young exceptfor the yellowish or cream-colored stipe base and thebrownish-yellow to yellowish-brown pileal disc. Thegeographic range of I. sublilacina overlaps with that ofI. pallidicremea, where the former has thus far onlybeen confirmed with molecular data from Coloradoand Newfoundland and Labrador. Despite their mor-phological and ecological similarities (FIGS. 3, 4), thetwo species are rather distantly related (FIGS. 1, 2).
The photo of I. lilacina under Tsuga in Lincoff(1981) resembles specimens in which the lilac colorsfaded and thus match the description of I. pallidicre-mea. The photograph of I. geophylla in Baroni (2017)probably illustrates faded specimens of I. pallidicremea.
Two morphological variants among the specimensexamined stand out. PBM1334 was observed with pallidto pale vinaceous drab lamellae when young, andPBM2238 had a whitish stipe throughout with no yel-low or cream to the stipe base. In the overwhelmingnumber of collections we studied of I. pallidicremea, theyoung lamellae are light gray to gray when young, andthe stipe base is yellowish or cream-colored.
Inocybe lilacina (Peck) Kauffman, The Agaricaceaeof Michigan I:466. 1918. FIGS. 3C–D, 4C–D
≡ Agaricus geophyllus var. lilacinus Peck, Ann RepNY St Mus Nat Hist 26:90. 1874 [1873].
≡ Inocybe geophylla var. lilacina (Peck) Gillet [as‘lilacinus’], Les Hyménomycètes ou Description de tousles Champignons quie Croisssent en France:520. 1876.
Description: Pileus 8–18(–30) mm wide, obtuselyconical in youth, expanding to plane with age, devel-oping a small subacute umbo, margin incurved inyouth becoming decurved; surface tacky, subviscid,margin entire; intense dark purple, dark violet, toblackish purple in youth (15F8–F7), this remaining soover the disc (“Raisin Black” to “Dull Violet-Black” orvery dark purple), with purplish or dark violet fibrillosestreaks towards the margin over a whitish background;context up to 2 mm thick, whitish but may be dullviolet above the lamellae when water soaked, not chan-ging color where cut or bruised, odor strongly sper-matic. Lamellae adnexed to subsinuate, moderatelyclose with several tiers of lamellulae, light gray inyouth, becoming pale brown to brown at maturity,edges pallid-fimbriate or indistinctly so with age,
MYCOLOGIA 629

ventricose. Stipe 20–30 × 2–3 mm, even but with aswollen or slightly rounded bulbous base; cortinamixed violet and whitish with the interior silky white;surface dark violet like the pileus with an underlyingwhite background, white where expanding upwards,leaving streaks of dark violet glutinous fibrils, cream-colored at the extreme base mixed with white at pointof attachment; context soft, interior white with a violetcortex when young, white in the base.
Basidiospores 8–9–9.5(–10) × 4.5–5.2–5.5 μm, Q =1.45–1.74–1.90(–2.00) (n = 31/2), smooth, subamygdali-form or elliptical with bluntly pointed or rounded apices,apiculus small but distinctive, yellowish brown in KOH.Basidia 24–30 × 8–9 μm, 4-sterigmate, clavate, hyaline.Pleurocystidia 45–60 × 14–18 μm, fusiform to fusiform-ventricose, apices obtuse and sparsely crystalliferous,tapered towards the base, thick-walled (walls 1.0–3.0μm thick), hyaline. Cheilocystidia similar to pleurocysti-dia but often shorter, mixed with paracystidia that arepyriform to clavate, hyaline, and thin-walled.Caulocystidia restricted to the extreme apex or upper1/8 of stipe surface, similar to cheilocystidia, at timesmixed with shorter, thin-walled, hyaline cells; stipe sur-face covered with an interwoven superficial layer of hya-line hyphae, these thin-walled, smooth, and mostly 5–10μm wide. Pileipellis an interwoven cutis, pale pinkish inmass, hyphae smooth, cylindrical, thin-walled, mostly5–10 μm wide. Clamp connections present.
Ecology and distribution: Scattered singly or in smallclusters on soil in the eastern United States—New York(type), North Carolina, Tennessee; in woods of Pinus,in mixed forests containing Tsuga, Pinus, Fagus, Betula,Quercus, Juglans, Carya, or in beech-oak-hickory for-ests (Fagus, Quercus, Carya). Occurring Aug–Oct.
Illustration: Elliott and Stephenson (2018, as I. geo-phylla var. lilacina).
Specimens examined: USA. NEW YORK: Bethlehem,Albany Co., no date, C.H. Peck (holotype NYSf1711);NORTH CAROLINA: Highlands (35.0525, −83.19694),31 Aug 1939, L.R. Hesler (TENN 012367); Highlands,Franklin, Wayah Bald (35.18028, −83.56055), in deciduouswoods, 21 Aug 1955, L.R. Hesler (TENN 022115); GreatSmoky Mountains National Park, Kephart Prong Trail(35.58667, −83.35972), 17 Aug 2005, E. Lickey TFB12747(TENN 061204); Blue Ridge Parkway, Little Switzerland,0.2 miles from the Little Switzerland Tunnel (35.82472,−82.10639), on soil under Quercus and Betula, 13 Sep2013, J.M. Birkebak & M. Sánchez-García PBM3982(TENN 068443); TENNESSEE: Anderson Co., NorrisDam State Park, near Clear Creek Trail (36.2352,−84.1032), on soil in hardwood forest under Fagus,Quercus, Carya, 23 Oct 2017, V.R. Harpe VRH13 (TENN073103); Knoxville, New Hopewell (35.96055, −83.92083),
on soil in pine woods, 21 Oct 1934, L.R. Hesler (TENN006434); Knoxville, New Hopewell (35.92833, −83.80055),on soil in pine woods, 17 Nov 1935, L.R. Hesler (TENN008431); ibid., on soil in pine woods, 17 Oct 1936, L.R.Hesler (TENN 009562); ibid., on soil in pine woods, 20 Oct1937, L.R. Hesler (TENN010926); Great SmokyMountainsNational Park, near LeConte (35.655, −83.44111), in mixedwoods, 29 Sep 1956, T.H. Campbell & L.R. Hesler (TENN022423); Great Smoky Mountains National Park, CadesCove (35.60194, −83.81138), on soil in pine woods, 23 Sep1966, L.R. Hesler (TENN029412); Great SmokyMountainsNational Park, Greenbrier (35.73028, −83.40611), onground under Tsuga, Pinus, Liquidambar, and other hard-woods (Quercus absent), 6 Sep 2004,M.C. Aime&E. LickeyPBM2590 (TENN 062429); Great Smoky MountainsNational Park, Cades Cove, Little Baptist Church area(35.60194, −83.81333), on soil under Tsuga, Fagus,Quercus, Juglans, Carya, 9 Sep 2004, M.C. Aime & E.Lickey PBM2628 (TENN 062463); ibid., 9 Sep 2004, E.Lickey PBM2629 (TENN 062464); Great SmokyMountains National Park, Greenbrier area, GrapeyardRidge Trail (35.70778, −83.38361), 25 Sep 2006, E. LickeyTFB13384 (TENN 061647); Great Smoky MountainsNational Park, Greenbrier area, Ramsey Cascades PicnicArea (35.71056,−83.38306), inmixed forest, 13Oct 2012,E.Harrower EH212 (TENN 067734); Union Co., Big RidgeState Park, on ground in mixed hardwood forest (Fagus,Carya, Quercus) with Pinus virginiana, 27 Sep 2011, J.W.Bills JWB17 (UTEEB351 teaching collection); Great SmokyMountains National Park, Cherokee Orchard Loop, OglePlace (35.6829, −83.4898), on soil in mixed forest of Tsuga,Pinus, Fagus, Betula, 30 Aug 2013, B.P. Looney PBM3940(TENN 068443).
Notes: Inocybe lilacina differs morphologically fromI. pallidicremea by the smaller basidiome size, moreintense dark violet pigmentation, and pinkish colorsthat tend to persist after drying. It is comparativelymuch less frequent that I. pallidicremea, which is com-mon in northern and western parts of North Americabased on molecular confirmation of specimens andenvironmental sequences. The two species are sistergroups with strong measures of support (FIGS. 1, 2).
Inocybe lilacina has been recorded “in pine woods”but also in habitats without any Pinaceae. L.R. Heslermade several collections in the greater Knoxville areaunder pines (Hesler 1936). An environmental sequence(EF619676) that corresponds to I. lilacina was sampledfrom a Pinus taeda dominated forest in North Carolina(Parrent and Vilgalys 2007) (SUPPLEMENTARY FIG.1). Associations with other trees (Tsuga, Quercus,Fagus, Carya), however, cannot be ruled out, and onecollection (VR13) was made in a pure oak-hickory-beech forest.
630 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

ITS variation between I. lilacina and I. pallidicremeais very low (0.2–0.8%). Operational approaches that relyon ITS sequence dissimilarity (genetic distance thresh-olds) are not sufficient to distinguish these two species.The genetic distance at the rpb2 locus is higher than inthe ITS comparison (1.2–1.8%). Phylogenetic analyses ofmultiple loci serve to delineate the two species.
The collection HRL2223 referred to as I. “lilacina”(FIG. 1) was recorded from Montreal, Quebec, at theMorgan Arboretum under planted Abies, Salix, andPopulus. Despite its lilac coloration, the species is notclosely related to the I. lilacina subgroup. We cannotexclude the possibility that this lilac-colored taxon wasintroduced from outside North America. Phylogeneticanalysis of molecular data from the sample stronglysuggests a close relationship with a sample labeled I.“lilacina” from Sweden (EL12605).
A rare secondary metabolite, an ergostane triterpe-noid, was reported from a culture of I. lilacina by Liuet al. (2014). The collection from which the culture wasproduced should be examined and placed within aphylogenetic context to confirm the identification.Inocybe ionocephala Matheny, sp. nov.
FIGS. 3E–F, 4E–FMycoBank MB822202
Typification: USA. CALIFORNIA: MendocinoWoodlands Camp area, Mycological Society of SanFrancisco Foray (39.3311, −123.7349), on soil inmixed woods including Pseudotsuga, Notholithocarpus,Tsuga, Arbutus, Sequoia, 15 Nov 2008, P.B. MathenyPBM3049 (holotype TENN 062799). GenBank: ITS =KY990551; 28S = KY990504; rpb2 = MF416422.
Etymology: ionocephala (Greek), ion- purple,-cephala head, in reference to the pale lilac to lightgrayish lilac margin of the pileus.
Diagnosis: Inocybe ionocephala differs from I. lila-cina, I. pallidicremea, and I. sublilacina by the largerbasidiomata, less intense lilac coloration of the pileus,and the white stipe with a white base.
Description: Pileus 15–35 mm wide, obtusely conicalwith an obtuse (to broad) umbo, expanding with age butretaining umbo, margin incurved to decurved; surfaceslightly sticky or subviscid, smooth an unbroken over thedisc, silky-fibrillose towards the margin, velipellis absent;center cream, brownish yellow, to light brown (5B4), mar-gin pale lilac to light grayish lilac (14B2–B1); context white,thick, odor spermatic. Lamellae adnexed to sinuate, closewith several tiers of lamellulae, light gray to avellaneous,becoming brown with age, edges pallid-fimbriate, ventri-cose. Stipe 25–45 × 4–7 mm at the apex, even or swollen atthe base and up to 10 mm wide, cortina fugacious, apexpruinose, elsewhere finely fibrillose, white throughout,apparently white even when young; context not observed.
Basidiospores 7.5–8.5–9.5(–10) × 4.5–5.0–5.5 μm, Q =1.45–1.68–1.80 (n = 31/2), smooth, subamygdaliform orelliptical with bluntly pointed or rounded apices, apiculussmall but distinctive, yellowish brown in KOH. Basidia25–30 × 8–9 μm, 4-sterigmate, clavate, hyaline.Pleurocystidia 58–70 × 12–19 μm, fusiform to fusiform-ventricose, less often subcylindrical, apices obtuse andcrystalliferous, tapered towards the base, thick-walled(1.5–2.5 μm) or occasionally only slightly thick-walled(ca. 1.0 μm), hyaline. Cheilocystidia similar to pleurocys-tida, often shorter, mixed with hyaline paracystidia.Pileipellis a compact interwoven cutis of cylindricalhyphae, these thin-walled, smooth, hyaline, mostly 5–10μm wide. Caulocystidia 79–108 × 10–11 μm, present onupper 1/8 of stipe surface (absent below), long and narrowranging from slenderly fusiform to cylindrical, mixed withcauloparacystida and other intermediate clavate to subcy-lindrical cells; lower part of stipe covered with a superficiallayer of hyaline, cylindrical, thin-walled hyphae, mostly 5 ×12 μm wide. Clamp connections present.
Ecology and distribution: In small clusters or scatteredsingly on soil in the coastal redwood zone betweenMendocino and San Mateo counties, northern California;associated with Pseudotsuga, Notholithocarpus, Tsuga, and/or Arbutus. Occurring Nov–Jan.
Other specimens examined: USA. CALIFORNIA: SanMateo Co., Butano State Park, Jackson Flat Trail(37.2227, −122.3033), on soil under Pseudotsuga inmixed woods, 2 Jan 1988, M.T. Seidl MTS2488 (UC);Mendocino Co., Caspar, Caspar Little Lake Rd. (39.3600,−123.7860), scattered singly on soil under Pseudotsuga,Notholithocarpus, Rhododendron, 26 Dec 2001, P.B.Matheny PBM2275 (WTU); Mendocino Co.,Mendocino Woodlands Camp Area, MycologicalSociety of San Francisco Foray (39.3311, −123.7349),on soil in mixed woods including Pseudotsuga,Notholithocarpus, Tsuga, Arbutus, Sequoia, 15 Nov2008, P.B. Matheny PBM3043 (TENN 062794); PointReyes National Seashore, Point Reyes Mycoblitz, collec-tion route Stew (38.0666, −122.8844), 10 Dec 2005,PtR21 (UC1859629); Point Reyes National Seashore,Point Reyes Mycoblitz, collection route OLEMA2(38.0435, −122.7905), 10 Dec 2005, PtR96 (UC1859626).
Notes: Inocybe ionocephala can be distinguished fromother North American species in the I. lilacina subgroupby the light grayish-lilac pileus with a brownish disc andoften robust stipe that is white throughout. Other speciesin the group feature a lilac-colored stipe when young butdiffer especially by the yellow- to cream-colored stipebase. Collections cited by Nishida (1989) as I. geophyllavar. lilacina most likely represent I. ionocephala, as theseoriginated from northern California under conifers, butmolecular confirmation is lacking.
MYCOLOGIA 631

The description of I. lilacina in Siegel andSchwarz (2016), which corresponds well with I.ionocephala in terms of the white stipe base, basi-diome size, and geographic distribution, states thatthe stipe is pale lilac-gray and the lamellae palelilac-gray in youth. These features should be care-fully examined in young material of I. ionocephala,as our documentation of multiple collections foundthe stipe to be consistently white throughout(including the base) and the young lamellae aslight gray.
The photo of I. lilacina depicted under Pinus inDesjardin et al. (2015) in their California mycofloramore closely resembles the concept of I. pallidicremeabecause of the smaller basidiome size and lilac-tingedstipe compared with I. ionocephala, which produceslarger basidiomes and has an entirely white stipe.Inocybe sublilacina Matheny & A. Voitk, sp. nov.
FIGS. 3G–H, 4G–HMycoBank MB822203
Typification: USA. COLORADO: Routt Co.,Keystone, Montezuma Rd. (39.6081, −105.9228), onsoil under Picea, 2775 m, 9 Aug 2005, P.B. MathenyPBM2730 (holotype TENN 062542). GenBank: 28S =KY990519; rpb2 = MF416430.
Etymology: sublilacina (Latin), sub- almost, -lilacinalilac, so named because of the similar morphologicalappearance to I. lilacina.
Diagnosis: Most similar in morphology to I. pallidi-cremea but best distinguished from it by mostly some-what larger and elliptical basidiospores and geneticdivergence at ITS, 28S, and rpb2 loci.
Description: Pileus 10–28 mm wide, conical at firstbut expanding with age, developing an obtuse to sub-acute umbo, margin incurved at first becomingdecurved; surface dry to slightly tacky, finely fibrilloseat first and smooth and unbroken over the center,becoming silky-fibrillose to weakly rimose towards themargin with age; lilac at first (14C6–C5–C4), mostlythroughout except at the disc that may be tinged yel-lowish or brownish yellow, the umbo becoming brown-ish with age and the margin fading to purplish white(14A2–B2), or the center pallid due to a heavy velipellisand the margin fading to brown; context not observed,odor spermatic. Lamellae adnexed to sinuate, moder-ately close; whitish to grayish with lilac tints whenyoung, becoming grayish to brown; edges indistinctlypallid-fimbriate, ventricose. Stipe 17–35 × 3–6 mm atthe apex, base slightly swollen and up to 7–8 mm wide;cortina fugacious; surface silky-fibrillose, tacky or sub-viscid, pruinose at the apex; colored like the pileuswhen young except for the stipe base, which is
yellowish or cream-colored, in age fading to whitishmixed with streaks of pinkish lilac; context notobserved.
Basidiospores 8.0–9.4–10.5 × 5.0–5.7–6.5 μm, Q =1.38–1.65–1.91 (n = 63/3), smooth, elliptical withrounded apices or (sub)amygdaliform with bluntlypointed apices, at times with a slight ventral depression,apiculus small but distinctive, yellowish brown. Basidia24–28 × 7–10 μm, 4-sterigmate (occasionally 2-sterig-mate), clavate, hyaline. Pleurocystidia 50–75 × 11–14μm, fusiform to subcylindrical, apices obtuse and crystal-liferous; thick-walled (mostly 2.0–3.0 μm), pale yellow tohyaline. Cheilocystidia similar to pleurocystida, oftenshorter, mixed with hyaline paracystidia. Pileipellis acompact interwoven cutis of cylindrical hyphae, thesethin-walled, smooth, hyaline, mostly 5–12 μm wide,subhyaline to light ochraceous-buff in mass.Caulocystidia 40–60 × 14–19 μm, similar to hymenialcystidia but slightly wider, fusiform to ventricose, attimes thin-walled, mixed with cauloparacystidia orshorter clavate to cylindrical cells, present at only theextreme apex of the stipe. Clamp connections present.
Ecology and distribution: Singly on soil under Picea,Abies, Pinus at high elevation or high latitude,Newfoundland and Labrador and Colorado (type).Occurring Aug–Sep.
Other specimens examined: CANADA. NEW-FOUNDLAND AND LABRADOR: Gros MorneNational Park, Stuckless Pond (49.4299, −57.7103), 8Sep 2004, A. Voitk GM5-302 (TENN 071464). USA.COLORADO: Keystone, River Run Lodge, on soilunder Picea, ca. 2800 m, 8 Aug 2005, P.B. MathenyPBM2716 (TENN 062531).
Notes: Inocybe sublilacina is molecularly and microsco-pically distinct from I. pallidicremea, which it otherwiseclosely resembles. Phylogenetic analyses of three loci (ITS,28S, rpb2), however, readily separate I. sublilacina and I.pallidicremea as different phylogenetic species. The two arevery similar morphologically; however, the basidiosporesof I. sublilacina are slightly larger on average than those ofI. pallidicremea, and the basidiospores are more oftenelliptic in outline. Evenson (1997, as I. geophylla var. lila-cina) ascribes lilac tints to the immature or young lamellaeof Colorado specimens, but we are fairly certain that thisspecies lacks this feature after examination of numerousspecimens from DBG. Both I. sublilacina and I. pallidicre-mea occur in Newfoundland and Labrador and Colorado,as confirmed by DNA sequencing. Examination of addi-tional interior western US collections attributed to I. lila-cina confirms their status as I. pallidicremea(SUPPLEMENTARY FIG. 1). Comparatively, I. sublilacinais rare.
632 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA

DISCUSSION
This is the first treatment to revise the taxonomy ofspecies in the I. geophylla group or subsectionGeophyllinae from North America within a molecularphylogenetic framework. Overall, nearly 30 species-level lineages that correspond to phylogenetic specieswere detected, mainly from North America andEurope, increasing considerably the taxonomic diver-sity of the group. One particularly well-supported sub-group surrounds the putative cosmopolitan species I.lilacina that became the focus of this study.Phylogenetic analysis of multigene data (FIG. 1) recov-ered up to nine phylogenetic species within the broadlymorphologically recognized but polyphyletic I. lilacina.Several of these have been shown to represent distinctmorphological and phylogenetic species that occupydifferent geographical regions in North America.
European works have reported the occurrence of I.lilacina or I. geophylla var. lilacina since the late 1800s;however, our molecular analyses based on current sam-pling have yet to confirm the presence of I. lilacina inEurope. At least two different European clades ascribedto I. lilacina have been detected, neither of whichgroups with North American collections of I. lilacina(FIGS. 1, 2). New investigations of violet- or lilac-pig-mented taxa described from Europe, such as I. geo-phylla var. violacea (Pat.) Sacc. and I. geophylla var.amethystina Overeem, as well as taxa referred to byHeim (1931), are needed.
Future systematic revisions are also required to clar-ify the taxonomy and status of North American mate-rials referred to as I. geophylla, which are highlypolyphyletic, and unique but poorly known taxa suchas I. agglutinata, I. armeniaca Huijsman, I. fuscicothur-nata, I. fuscodisca, I. insinuata, I. pudica, I. virgata, I.sambucella G.F. Atk., and I. whitei (Berk. & Broome)Sacc., as well as lilac-pigmented taxa outside the I.lilacina subgroup.
KEY TO NORTH AMERICAN SPECIES OF THEINOCYBE LILACINA SUBGROUP
1. Stipe and stipe base white, robust, up to 10 mmwide; occurring in northern California in thecoastal redwood zone ..................... I. ionocephala
1′. Stipe lilac or with lilac streaks, fading to whitishwith age, base yellowish or cream-colored, moreslender than above, up to 6 mm wide; occurringelsewhere and in different habitats .................... 2
2. Pileus center persistently dark violet to dark purplewhen fresh, these colors not completely fading; stipe2–3 mm thick at the apex........................... I. lilacina
2′. Pileus center yellowish or becoming brownishyellow, yellowish brown, brown, or pallid, lilaccompletely fading; stipe 3–6 mm wide at theapex ....................................................................... 3
3. Basidiospores on average <5.5 μm wide, mostlyamygdaliform in outline; widely distributed, innorthern and western regions of NorthAmerica, common......................... I. pallidicremea
3′. Basidiospores on average >5.5 μm wide, mostlyelliptical in outline; patchily distributed, knownonly from eastern Canada and the RockyMountains, uncommon..................... I. sublilacina
ACKNOWLEDGMENTS
Laboratory support was provided by Christine Braaten, EmilyGiles, Alicia Hobbs, Hailee Korotkin, Sarah Sprague, andAaron Wolfenbarger. The following individuals are thankedfor donation of materials or supporting information: JoeAmmirati, Tom Bruns, Cathy Cripps, Vera Evenson, DavidGiblin, Kabir Peay, Roy Halling, Karen Hughes, Renée Lebeuf,Lorinda Leonardi, Brian Looney, David Malloch, BuckMcAdoo, Steve Trudell, Andy Wilson, Mike Wood, AndrusVoitk, and Maria Yousoufian. We thank the following herbariaand staff for providing loans of materials: DBG, MONT, NY,NYS, TENN, UC, and WTU. The authors thank Keith Seifertand two anonymous reviewers for their helpful comments andfeedback on an earlier version of the manuscript.
FUNDING
Funding for this work was provided by grants to P.B.M. fromthe Daniel E. Stuntz Memorial Foundation and the HeslerEndowment Fund at the University of Tennessee.
ORCID
P. Brandon Matheny http://orcid.org/0000-0003-3857-2189
LITERATURE CITED
Baroni TJ. 2017. Mushrooms of the northeastern UnitedStates and eastern Canada. Portland, Oregon: TimberPress. 599 p.
Bessette AE, Miller OK Jr, Bessette AR, Miller HH. 1995.Mushrooms of North America in color: a field guidecompanion to seldom-illustrated fungi. Syracuse, NewYork: Syracuse University Press. 172 p.
Bessette AE, Roody WC, Bessette AR, Dunaway DL. 2007.Mushrooms of the southeastern United States. Syracuse,New York: Syracuse University Press. 373 p.
Braaten CC, Matheny PB, Vies DL, Wood MG, Williams JH,Bougher NL. 2014. Two new species of Inocybe fromAustralia and North America that include novel secotioidforms. Botany 92:9–22.
MYCOLOGIA 633

Desjardin DE, Wood MG, Stevens FA. 2015. Californiamushrooms: the comprehensive identification guide.Portland, Oregon: Timber Press. 559 p.
Elliott TF, Stephenson SL. 2018. Mushrooms of the southeast.Portland, Oregon: Timber Press. 407 p.
Evenson VS. 1997. Mushrooms of Colorado and the southernRocky Mountains. Denver, Colorado: Denver BotanicalGardens, Denver Museum of Natural History: WestcliffePublishers. 207 p.
Heim R. 1931. Le genre Inocybe, encyclopedic mycologique 1.Paris: Paul Lechevalier & Fils. 420 p.
Hesler LR. 1936. Notes on southern Appalachian fungi.Journal of the Tennessee Academy of Sciences 11:107–122.
Gillet CC. 1879. Champignons de France: les Hyménomycètes.Alençon, France: E. de Broise. 828 p.
Grund DW, Stuntz DE. 1977. Nova Scotian Inocybes: IV.Mycologia 69:392–408.
Jacobsson S, Larsson E. 2012. Inocybe (Fr.) Fr. In: KnudsenH, Versterholt J, eds. Funga nordica: agaricoid, boletoid,clavarioid, cyphelloid and gastroid genera. 2nd ed.Copenhagen, Denmark: Nordsvamp. p. 981–1021.
Kauffman CH. 1918. The Agaricaceae of Michigan. Vol. 1.Lansing, Michigan: Wynkoop Hallenbeck Crawford Co.State Printers. 924 p.
Kauffman CH. 1924. Inocybe. North American Flora 10:227–260.Kornerup A, Wanscher JH. 1967. Methuen handbook of
colour. 2nd ed. London: Methuen & Co. Ltd. 243 p.Kuyper TW. 1986. A revision of the genus Inocybe in Europe:
I. Subgenus Inosperma and the smooth-spored species ofsubgenus Inocybe. Persoonia 3(Supplement):1–247.
Larkin MA, Blackshields G, Brown NP, Chenna R,McGettigan PA, McWilliams H, Valentin F, Wallace IM,Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG.2007. Clustal W and Clustal X version 2.0. Bioinformatics23:2947–2948.
Lincoff GH. 1981. National Audubon Society field guide to NorthAmerican Mushrooms. New York: Alfred A. Knopf. 926 p.
Liu DZ, Liu Q, Yang P, Jiang Wx. 2014. A new ergostanetriterpenoid from cultures of the basidiomycete Inocybelilacina. Natural Products Communications 9:369–370.
Maddison DR, Maddison WP. 2005. MacClade 4.08.Sunderland, Massachusetts: Sinauer Associates.
Matheny PB. 2005. Improving phylogenetic inference ofmushrooms with RPB1 and RPB2 nucleotide sequences(Inocybe; Agaricales). Molecular Phylogenetics andEvolution 35:1–20.
Matheny PB, Aime MC, Bougher NL, Buyck B, Desjardin DE,Horak E, Kropp BR, Lodge DL, Trappe JM, Hibbett DS.2009. Out of the palaeotropics? Historical biogeographyand diversification of the cosmopolitan mushroom familyInocybaceae. Journal of Biogeography 36:577–592.
Matheny PB, Bougher NL. 2005. A new violet species ofInocybe (Agaricales) from urban and rural landscapesin Western Australia. Australasian Mycologist 24:7–12.
Matheny PB, Norvell LL, Giles EC. 2013. A common newspecies of Inocybe in the Pacific Northwest with a diag-nostic PDAB reaction. Mycologia 105:436–446.
Munsell Soil Color Charts. 1954. Baltimore, Maryland:Munsell Color Company, Inc.
Nishida F. 1989. Key to the species of Inocybe in California.Mycotaxon 34:181–196.
Parrent JL, Vilgalys R. 2007. Biomass and compositionalresponses of ectomycorrizal fungal hyphae to elevated CO2
and nitrogen fertilization. New Phytologist 176:164–174.Peck CH. 1874. Report of the botanist. Annual Report on the
New York State Museum of Natural History 26:36–91.Ridgway R. 1912. Color standards and color nomenclature.
Washington, DC: published by the author. 43 p., 53 pl.Ronquist F, Teslenko M, van der Mark P, Ayres L, Darling A,
Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP.2012. MrBayes 3.2: efficient Bayesian phylogenetic infer-ence and model choice across a large model space.Systematic Biology 61:539–542.
Ryberg M, Larsson E, Jacobsson S. 2010. An evolutionaryperspective on morphological and ecological characters inthe mushroom family Inocybaceae (Agaricomycotina,Fungi). Molecular Phylogenetics and Evolution 55:431–442.
Ryberg M, Nilsson RH, Kristiansson E, Töpel M, Jacobsson S,Larsson E. 2008. Mining metadata from unidentified ITSsequence in GenBank: a case study in Inocybe(Basidiomycota). BMC Evolutionary Biology 8:50.
Sánchez-García M, Matheny PB, Palfner G, Lodge DL. 2014.Deconstructing the Tricholomataceae (Agaricales) andintroduction of the new genera Albomagister, Corneriella,Pogonoloma and Pseudotricholoma. Taxon 63:993–1007.
Siegel N, Schwarz C. 2016. Mushrooms of the RedwoodCoast: a comprehensive guide to the fungi of coastalNorthern California. Berkeley, California: Ten SpeedPress. 601 p.
Singer R. 1986. The Agaricales in modern taxonomy. 4th ed.Koenigstein, Germany: Koeltz Scientific Books. 981 p.
Smith AH. 1949. Mushrooms in their natural habitats.Portland, Oregon: Sawyer’s Inc. 626 p.
Smith AH, Smith HV, Weber NS. 1979. How to know thegilled mushrooms. Dubuque, Iowa: Wm. C. BrownCompany Publishers. 334 p.
Stamatakis A. 2014. RAXML 8: a tool for phylogenetic analysisand post-analysis of large phylogenies. Bioinformatics30:1312–1313.
Swofford DL. 2002. PAUP*. Phylogenetic Analysis UsingParsimony (*and other methods). Version 4. Sunderland,Massachusetts: Sinauer Associates.
Thiers B. [continuously updated]. Index herbariorum: a globaldirectory of public herbaria and associated staff. New YorkBotanical Garden’s Virtual Herbarium. [cited 2018 Apr 16].Available from: http://sweetgum.nybg.org/ih/
Truong C., Mujic AB, Healy R, Kuhar F, Furci G. Tores D,Niskanen T, Sandoval-Leiva PA, Fernández N, Escobar JM,Moretto A, Palfner G, Pfister D, Houhra E, Swenie R,Sánchez-García, M, Matheny PB, Smith ME. 2017. Howto know the fungi: combining field inventories and DNA-barcoding to document fungal diversity. New Phytologist214:913–919.
634 MATHENY AND SWENIE: REVISION OF INOCYBE LILACINA