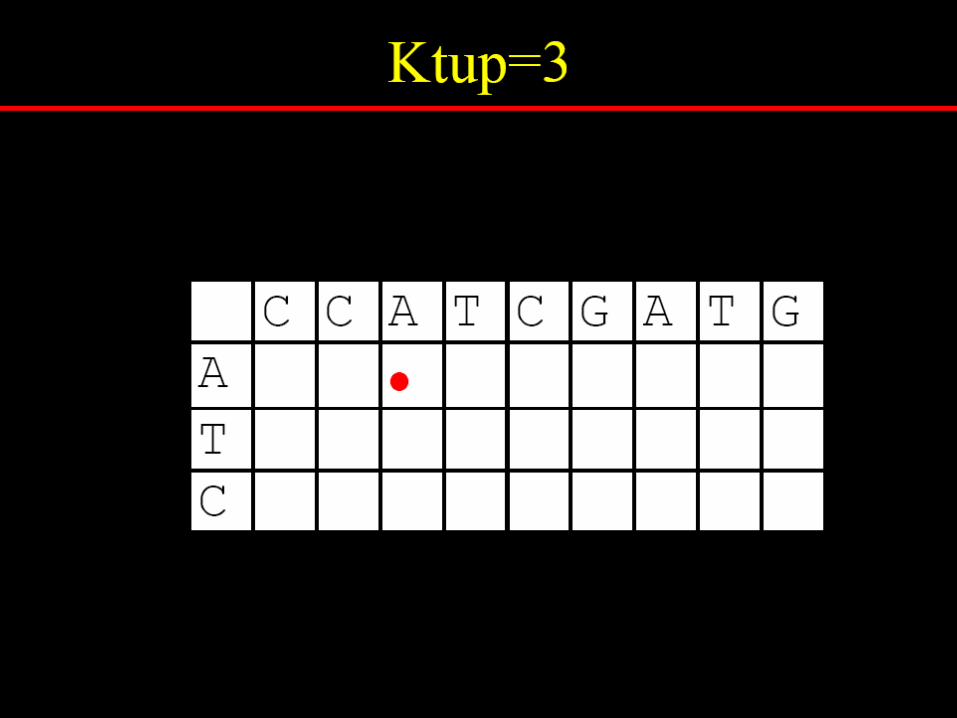
pairwise sequence alignment h.c.huang @ 2005/9/22 2005 autumn / ym / bioinformatics
Post on 18-Dec-2015
226 views
TRANSCRIPT

Pairwise Sequence Alignment
H.C.Huang @ 2005/9/22
2005 Autumn / YM / Bioinformatics

Outline
MotivationA simple alignment problemScoring alignments
Substition matrices Gap penalties
Dynamic programmingGlobal vs. local alignment

Reference
D.W. Mount / Bioinformatics / Ch.3
Slides fromProf. C.H.ChangUW / Genomic Informatics / W.S. Noble

Example

Brief History Robinson (1938) described the 1st algorithm
for Longest Common Subsequence problem Levenshtein (1966) introduced “edit distance”
between strings Min. # of edit operations needed to transform from
one string to another No algorithm described
Gibbs & McIntyre (1970): dot matrix analysis Needleman & Wunsch (1970) “re-discovered”
the algorithm for sequence alignment using DP Smith & Waterman (1981) proposed a clever
modification of DP for local alignment

Brief History
Knuth (1973) suggested a filtering idea for pattern matching高德纳 (Donald Knuth)
Dumas & Ninio (1982) first applied the filtering idea for fast sequence comparison
Lipman & Pearson (1985): FASTAAltschul et al. (1990): BLAST

Pairwise Alignment Methods
Dotplot analysis methodDynamic programming (DP)
algorithmWord / k-tuple methods
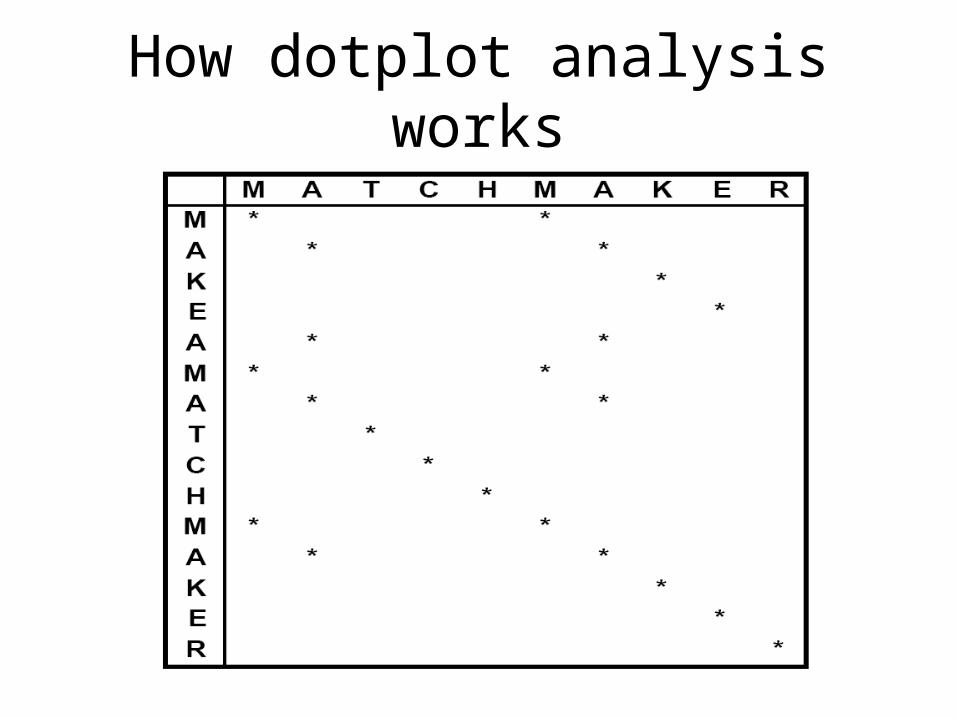
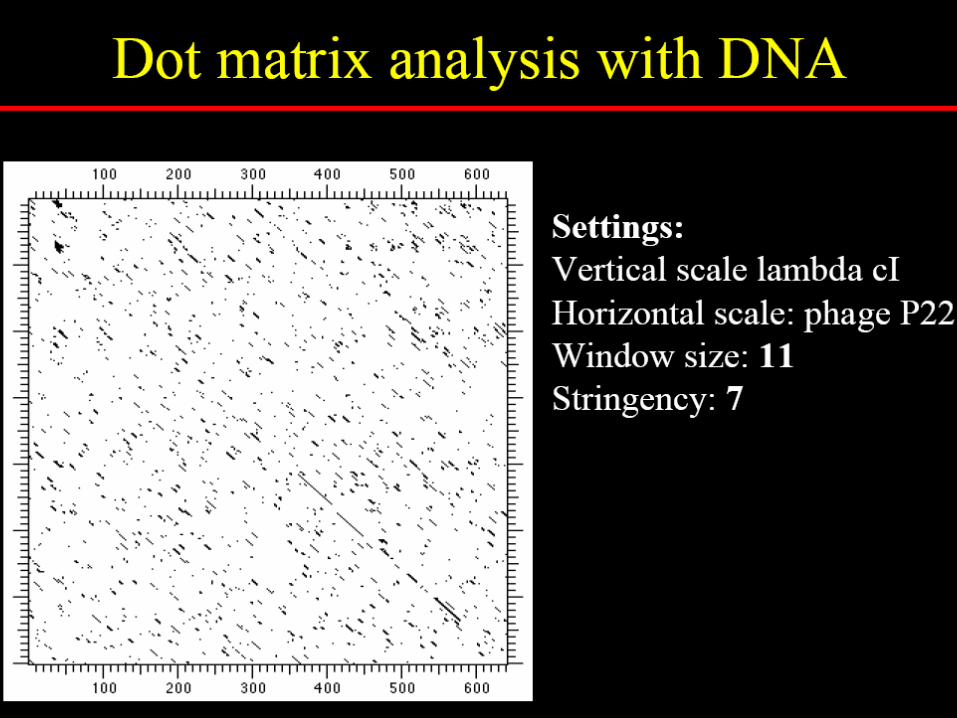
How dotplot analysis works
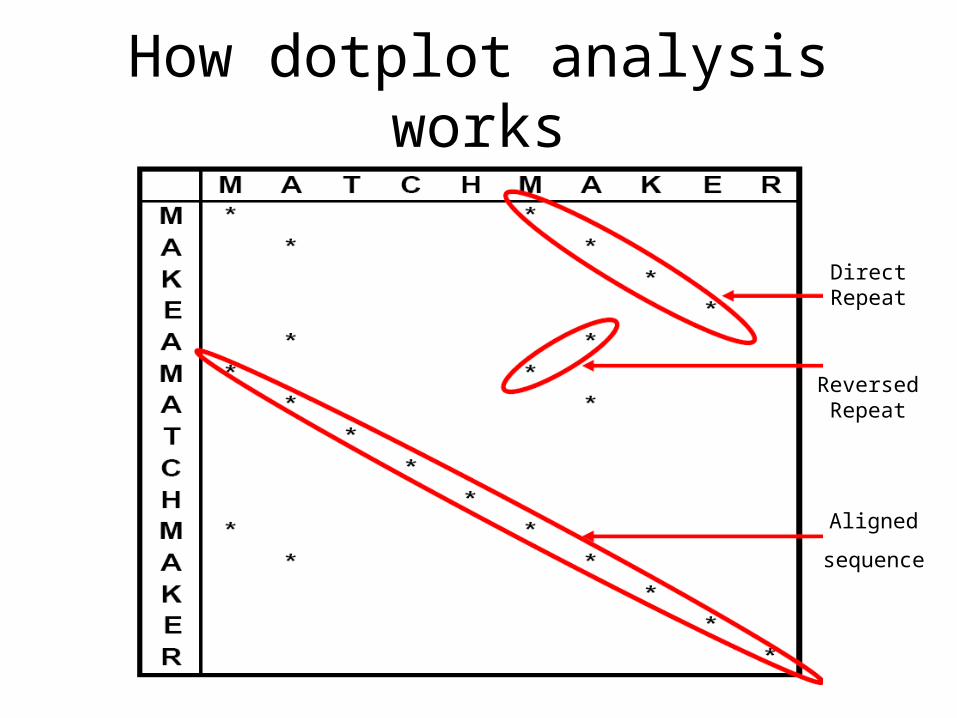
How dotplot analysis works
Direct Repeat
Reversed Repeat
Aligned
sequence

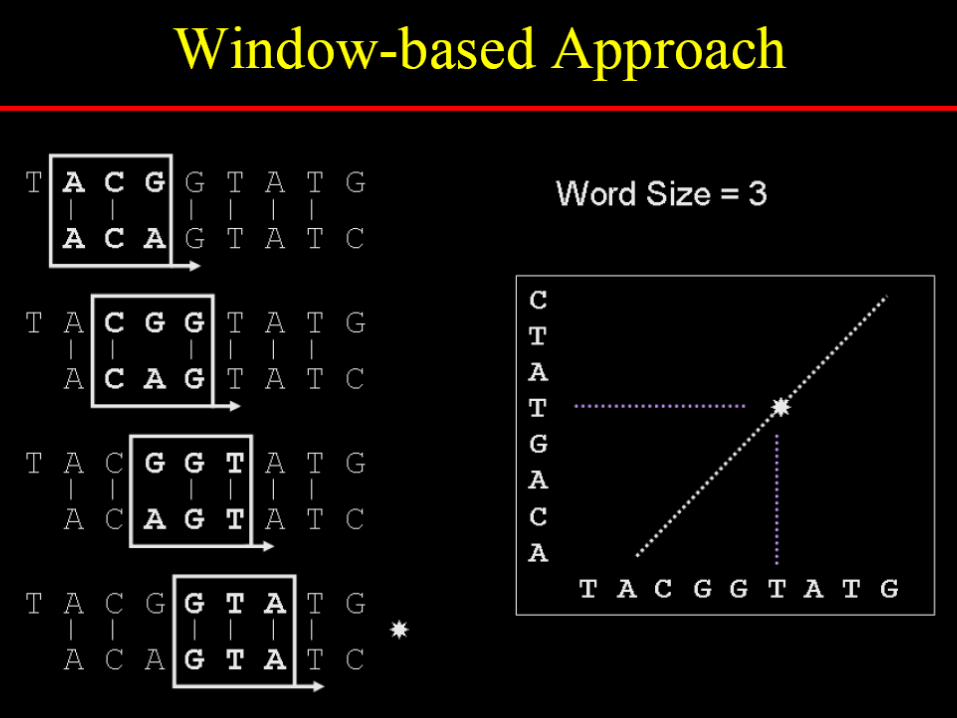
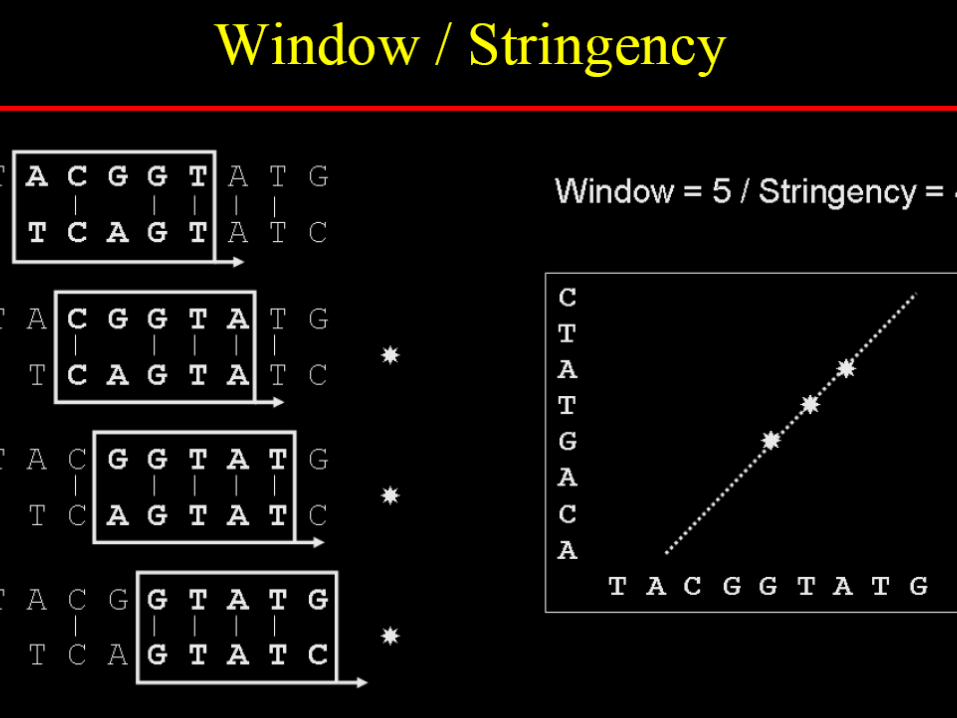
To reduce random noise in dotplots
Specify a window size, wExamine w residues at a timeCount how many matches are
among the w pairs of residues
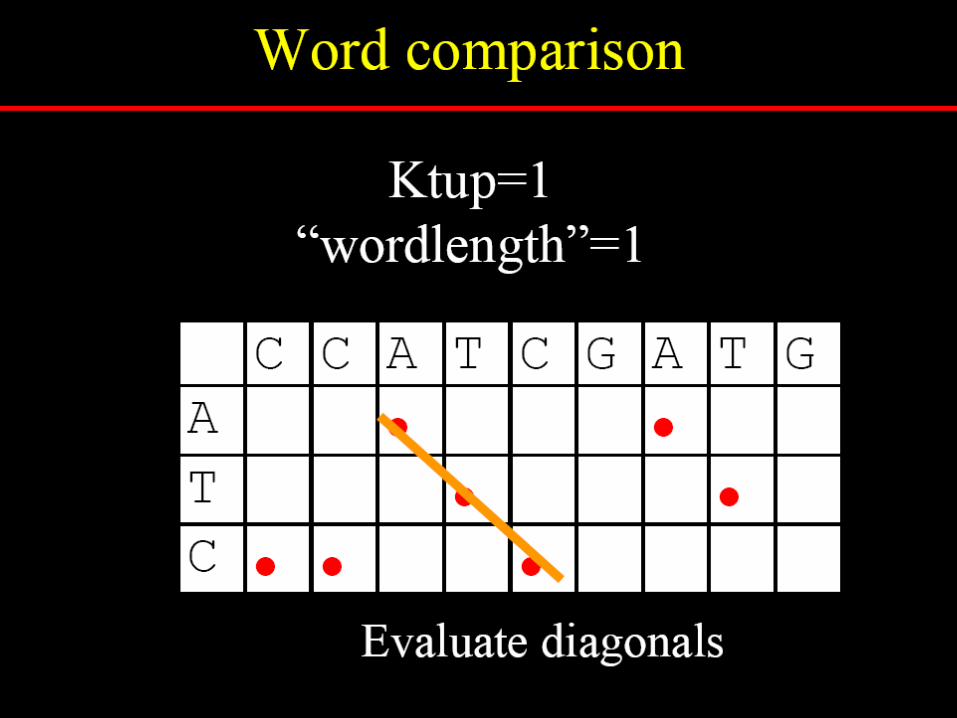
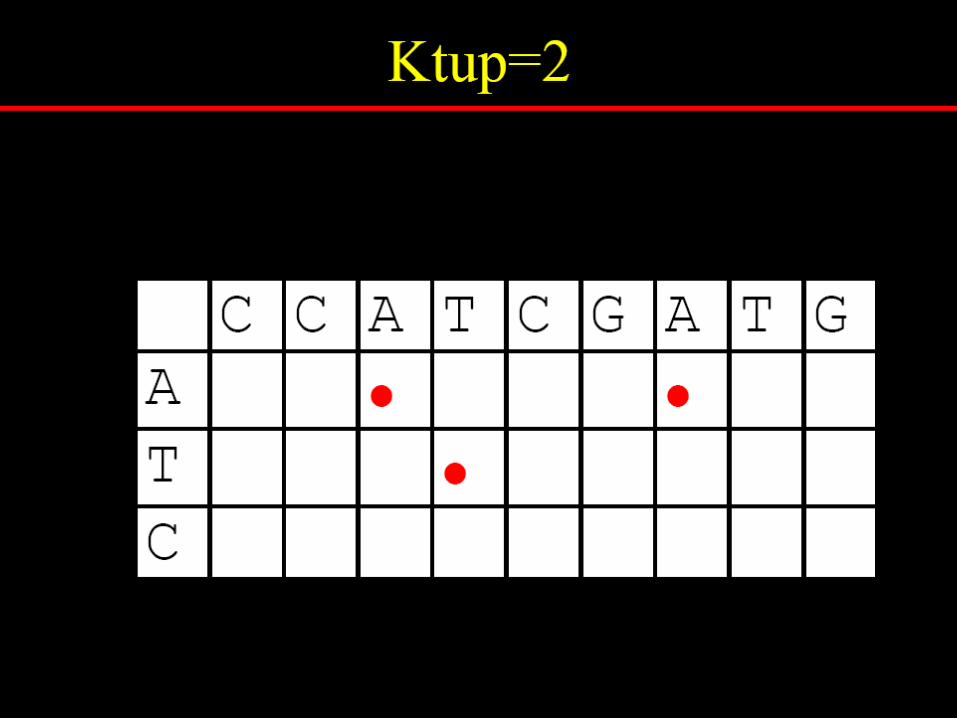


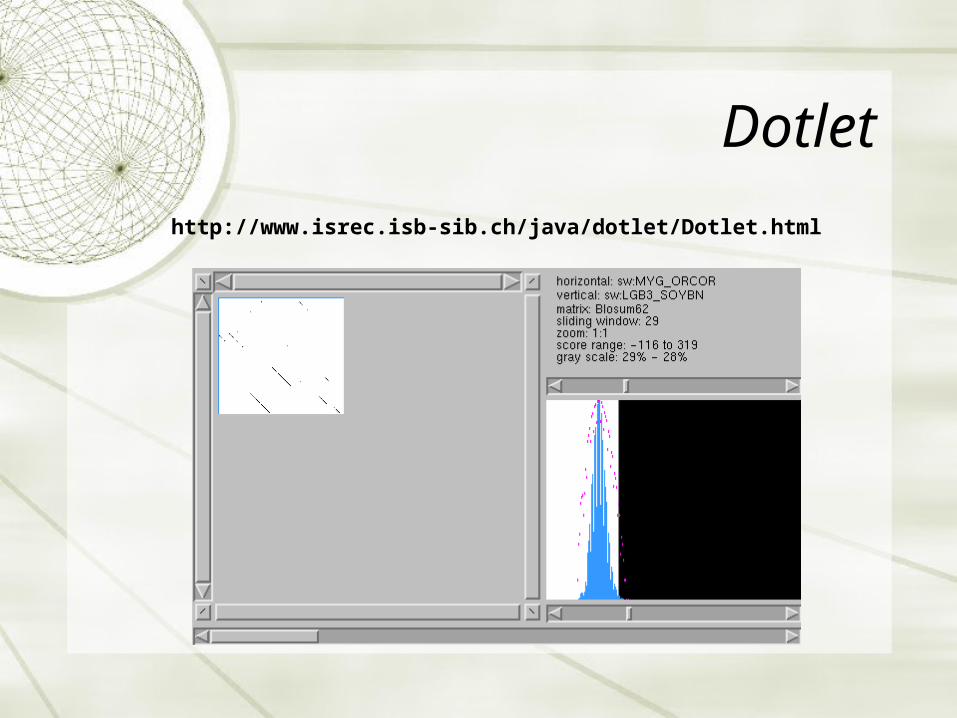
Dotlet
http://www.isrec.isb-sib.ch/java/dotlet/Dotlet.html

A simple alignment problem.
Problem: find the best pairwise alignment of GAATC and CATAC.

Scoring alignments
• We need a way to measure the quality of a candidate alignment.
• Alignment scores consist of two parts: a substition matrix, and a gap penalty.
GAATCCATAC
GAATC-CA-TAC
GAAT-CC-ATAC
GAAT-CCA-TAC
-GAAT-CC-A-TAC
GA-ATCCATA-C
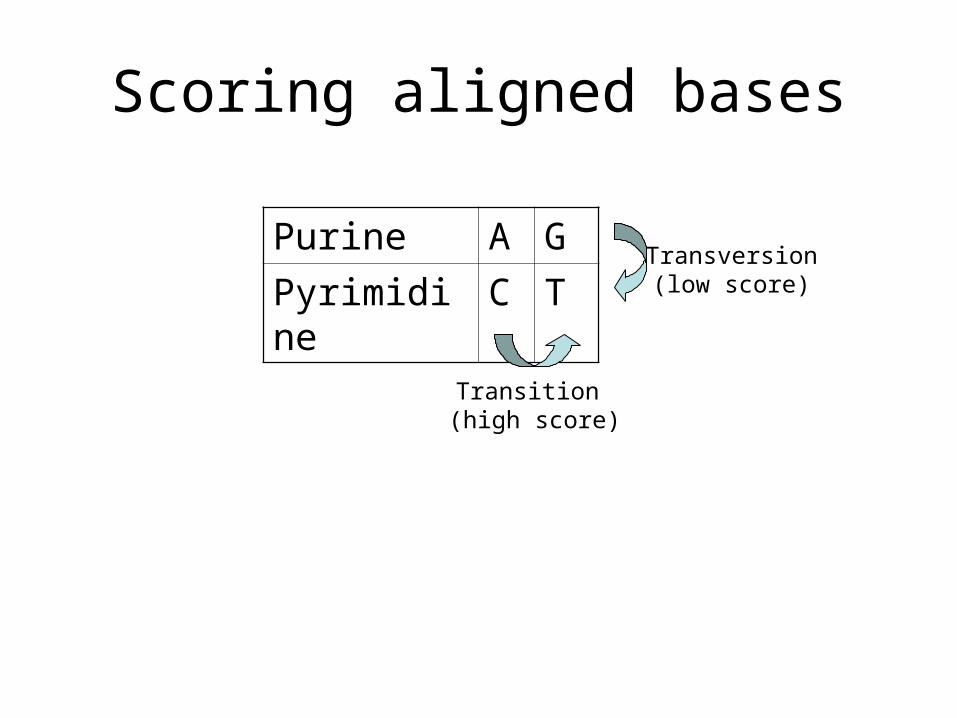
Scoring aligned bases
Purine A G
Pyrimidine C T
Transition (high score)
Transversion(low score)
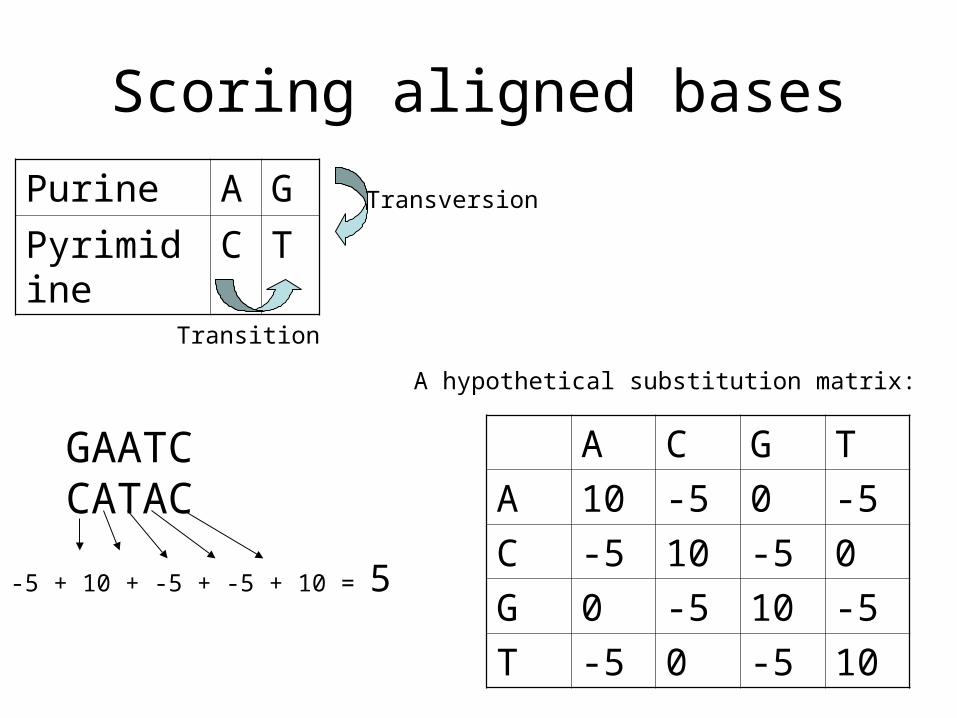
Scoring aligned bases
Purine A G
Pyrimidine C T
Transition
Transversion
A C G T
A 10 -5 0 -5
C -5 10 -5 0
G 0 -5 10 -5
T -5 0 -5 10
A hypothetical substitution matrix:
GAATCCATAC
-5 + 10 + -5 + -5 + 10 = 5
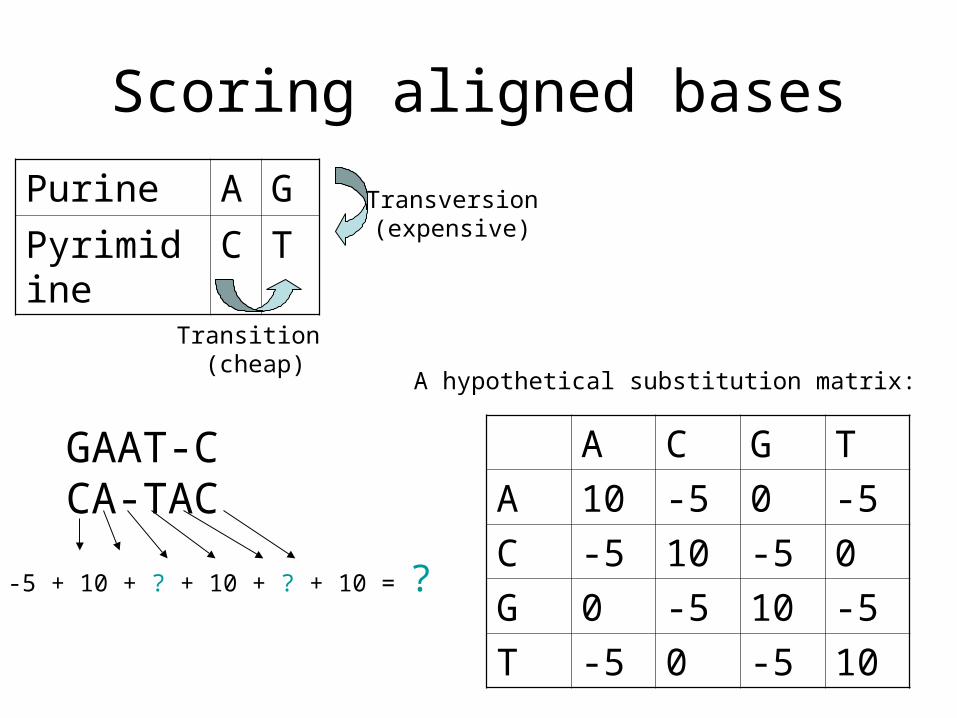
Scoring aligned bases
Purine A G
Pyrimidine C T
Transition (cheap)
Transversion(expensive)
A C G T
A 10 -5 0 -5
C -5 10 -5 0
G 0 -5 10 -5
T -5 0 -5 10
A hypothetical substitution matrix:
GAAT-CCA-TAC
-5 + 10 + ? + 10 + ? + 10 = ?

• Linear gap penalty: every gap receives a score of d.
• Affine gap penalty: opening a gap receives a score of d; extending a gap receives a score of e.
Scoring gaps
GAAT-C d=-4CA-TAC
-5 + 10 + -4 + 10 + -4 + 10 = 17
G--AATC d=-4CATA--C e=-1
-5 + -4 + -1 + 10 + -4 + -1 + 10 = 5

A simple alignment problem.
• Problem: find the best pairwise alignment of GAATC and CATAC.
• Use a linear gap penalty of -4.
• Use the following substitution matrix:
A C G T
A 10 -5 0 -5
C -5 10 -5 0
G 0 -5 10 -5
T -5 0 -5 10

How many possibilities?
• How many different alignments of two sequences of length N exist?
GAATCCATAC
GAATC-CA-TAC
GAAT-CC-ATAC
GAAT-CCA-TAC
-GAAT-CC-A-TAC
GA-ATCCATA-C
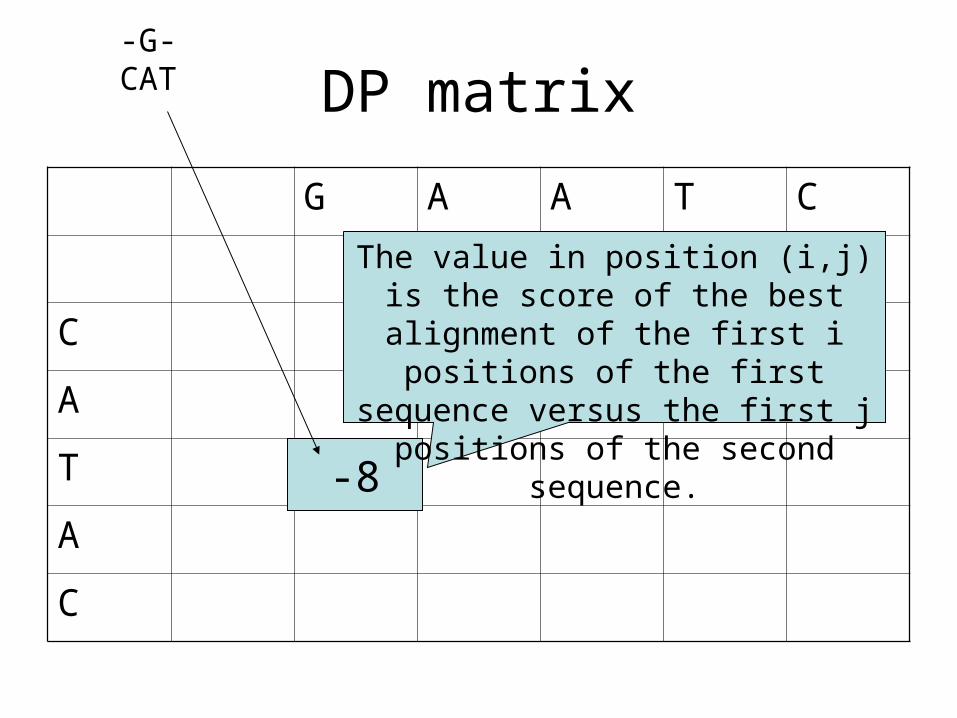
DP matrix
G A A T C
C
A
T
A
C
-8
The value in position (i,j) is the score of the best alignment of the first i
positions of the first sequence versus the first j positions of the
second sequence.
-G-CAT
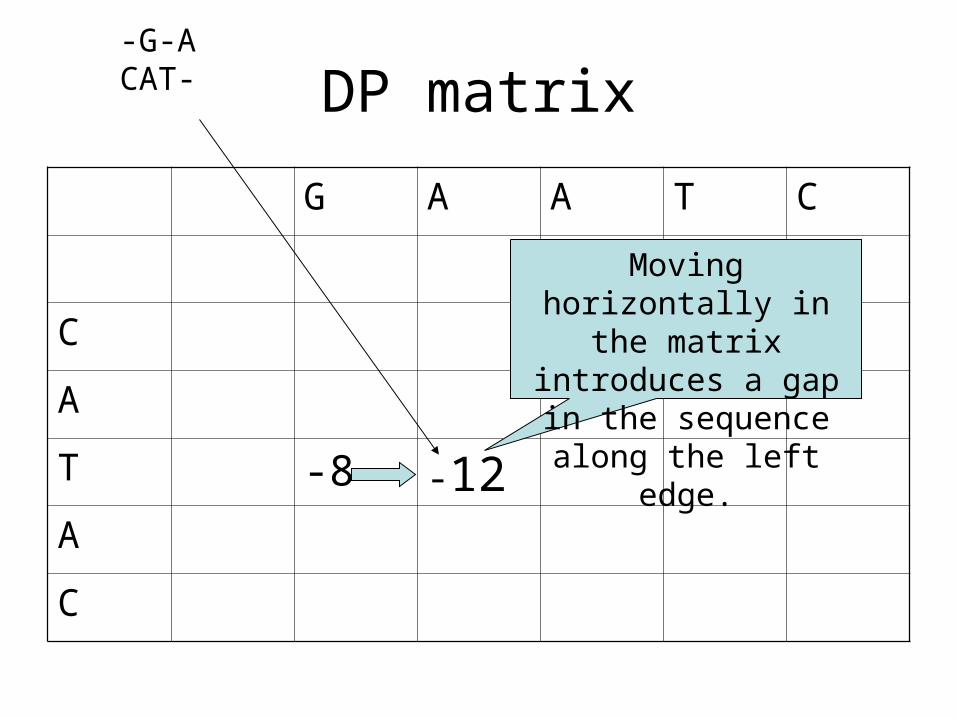
DP matrix
G A A T C
C
A
T -8 -12A
C
Moving horizontally in the matrix introduces a
gap in the sequence along the left edge.
-G-ACAT-
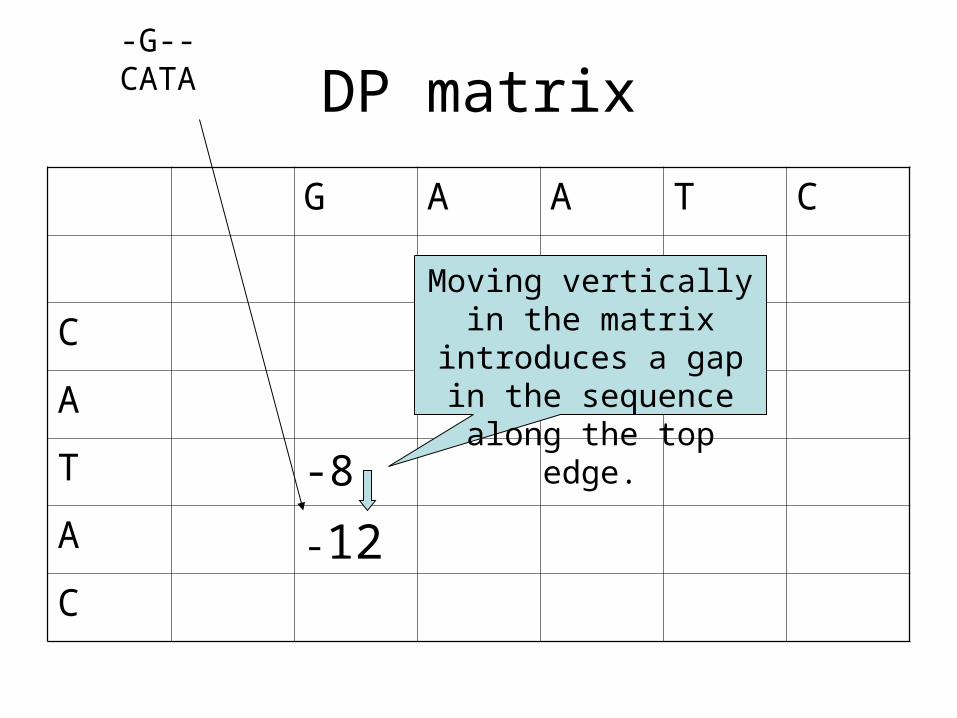
DP matrix
G A A T C
C
A
T -8
A -12C
Moving vertically in the matrix introduces a gap in the sequence along
the top edge.
-G--CATA

Initialization
G A A T C
0
C
A
T
A
C
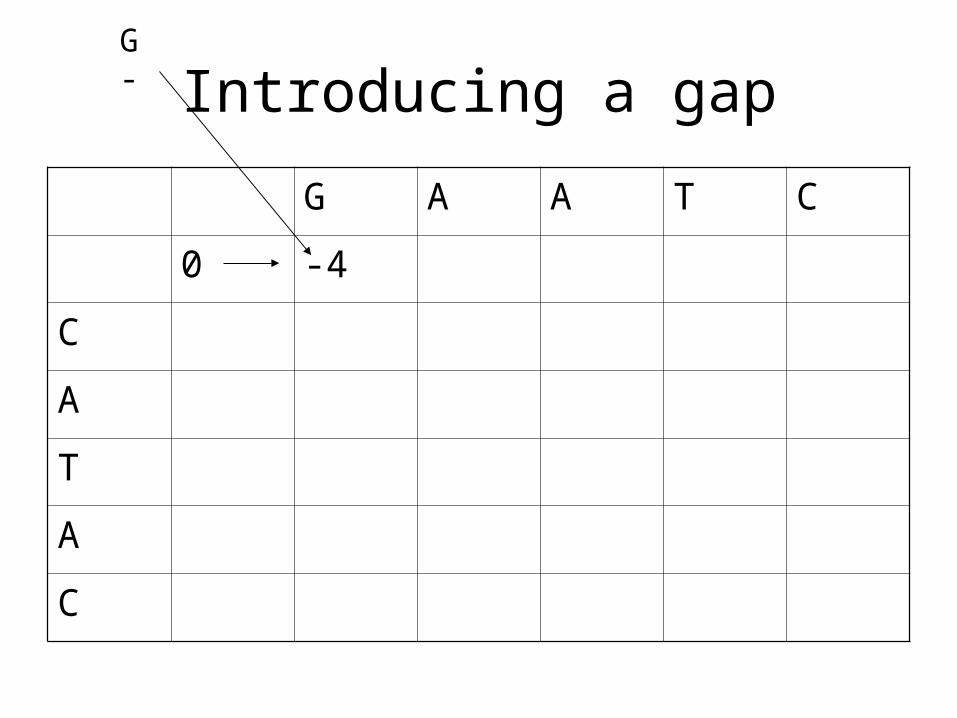
Introducing a gap
G A A T C
0 -4
C
A
T
A
C
G-

DP matrix
G A A T C
0 -4
C -4
A
T
A
C
-C

DP matrix
G A A T C
0 -4
C -4 -8
A
T
A
C
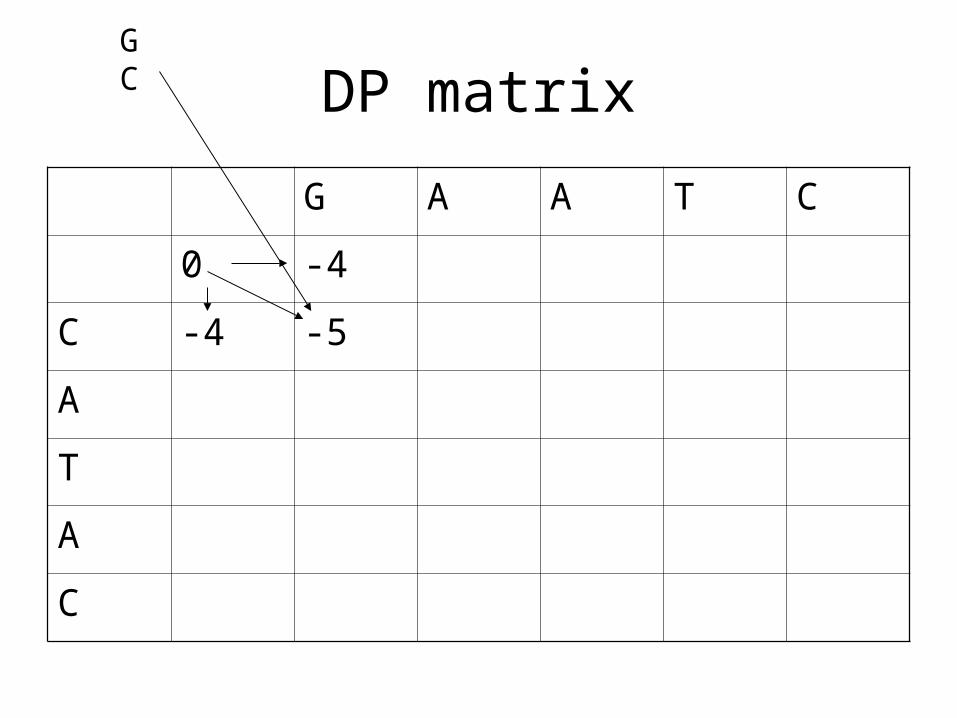
DP matrix
G A A T C
0 -4
C -4 -5
A
T
A
C
GC

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5
A -8
T -12
A -16
C -20
-----CATAC

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5
A -8 ?
T -12
A -16
C -20

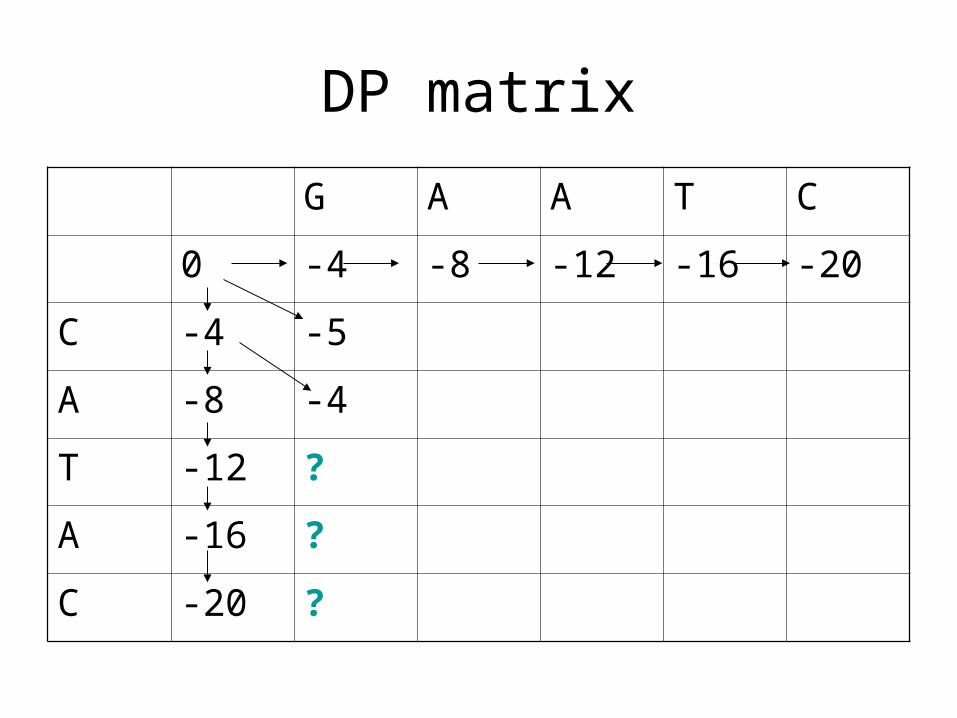
DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5
A -8 -4
T -12
A -16
C -20
-4
0 -4
-GCA
G-CA
--GCA-
-4 -9 -12
DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5
A -8 -4
T -12 ?
A -16 ?
C -20 ?
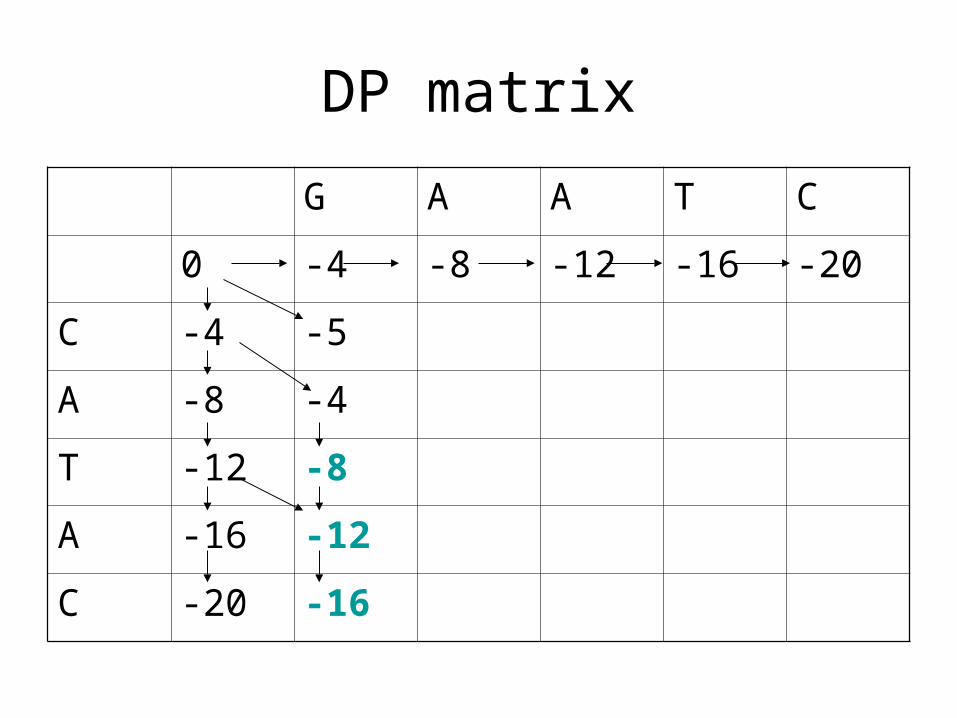
DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5
A -8 -4
T -12 -8
A -16 -12
C -20 -16
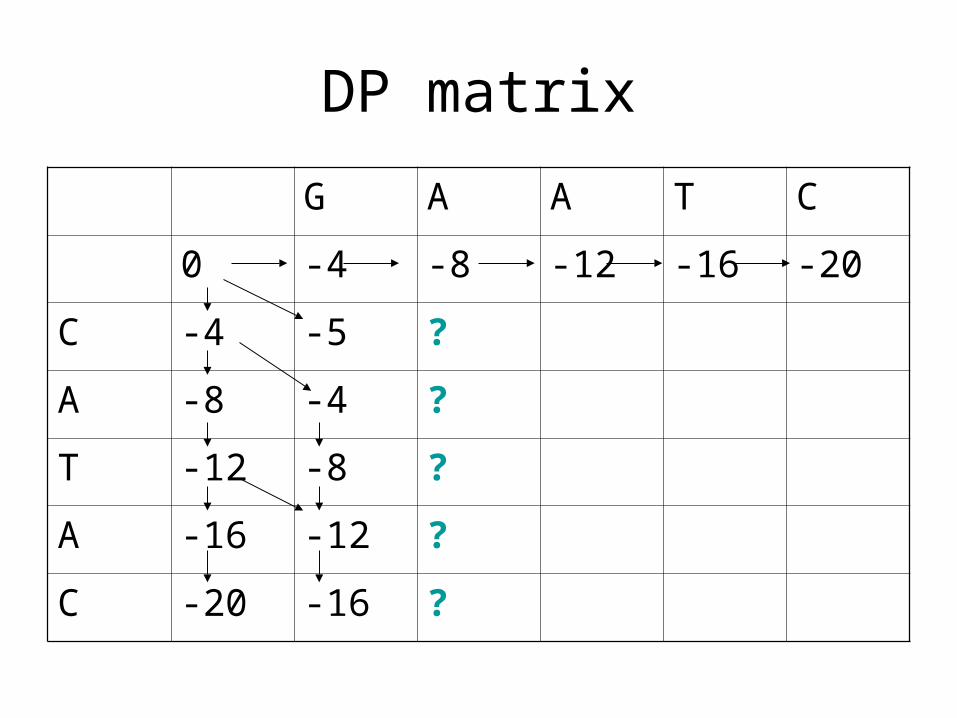
DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 ?
A -8 -4 ?
T -12 -8 ?
A -16 -12 ?
C -20 -16 ?

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9
A -8 -4 5
T -12 -8 1
A -16 -12 2
C -20 -16 -2
What is the alignment associated with this
entry?
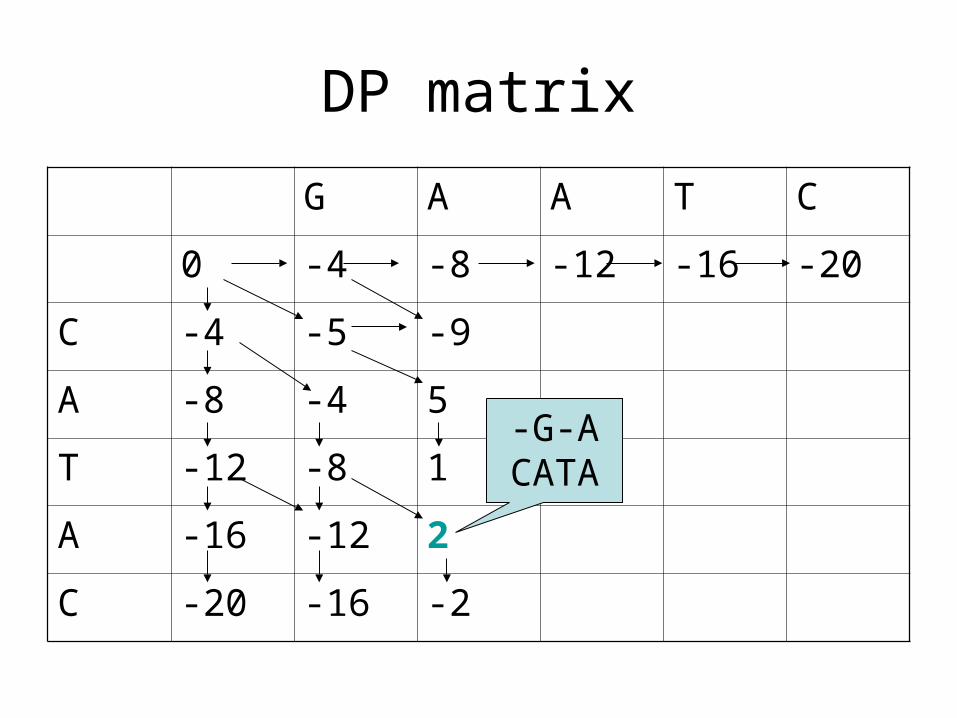
DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9
A -8 -4 5
T -12 -8 1
A -16 -12 2
C -20 -16 -2
-G-ACATA

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9
A -8 -4 5
T -12 -8 1
A -16 -12 2
C -20 -16 -2 ?
Find the optimal alignment, and
its score.

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9 -13 -12 -6
A -8 -4 5 1 -3 -7
T -12 -8 1 0 11 7
A -16 -12 2 11 7 6
C -20 -16 -2 7 11 17

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9 -13 -12 -6
A -8 -4 5 1 -3 -7
T -12 -8 1 0 11 7
A -16 -12 2 11 7 6
C -20 -16 -2 7 11 17
GA-ATCCATA-C

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9 -13 -12 -6
A -8 -4 5 1 -3 -7
T -12 -8 1 0 11 7
A -16 -12 2 11 7 6
C -20 -16 -2 7 11 17
GAAT-CCA-TAC

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9 -13 -12 -6
A -8 -4 5 1 -3 -7
T -12 -8 1 0 11 7
A -16 -12 2 11 7 6
C -20 -16 -2 7 11 17
GAAT-CC-ATAC

DP matrix
G A A T C
0 -4 -8 -12 -16 -20
C -4 -5 -9 -13 -12 -6
A -8 -4 5 1 -3 -7
T -12 -8 1 0 11 7
A -16 -12 2 11 7 6
C -20 -16 -2 7 11 17
GAAT-C-CATAC

Multiple solutions
When a program returns a sequence alignment, it may not be the only best alignment.
GA-ATCCATA-C
GAAT-CCA-TAC
GAAT-CC-ATAC
GAAT-C-CATAC

DP in equation form
• Align sequence x and y.
• F is the DP matrix; s is the substitution matrix; d is the linear gap penalty.
djiF
djiF
yxsjiF
jiF
F
ji
1,
,1
,1,1
max,
00,0

DP in equation form
1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,

Dynamic programming
Yes, it’s a weird name.DP is closely related to recursion
and to mathematical induction.We can prove that the resulting
score is optimal.

Dynamic Programming
A method for solving recursive problemBreak a problem into smaller
subproblemsSolve subproblems optimally,
recursivelyUse these optimal solutions to
construct an optimal solution for the original problem

Summary
Scoring a pairwise alignment requires a substition matrix and gap penalties.
Dynamic programming is an efficient algorithm for finding the optimal alignment.
Entry (i,j) in the DP matrix stores the score of the best-scoring alignment up to those positions.
DP iteratively fills in the matrix using a simple mathematical rule.

A simple exercise
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
A
G
C 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal alignment of AAG and AGC.Use a gap penalty of d=-5.

A simple exercise
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0
A
G
C 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal alignment of AAG and AGC.Use a gap penalty of d=-5.

A simple exercise
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 -5 -10 -15
A -5
G -10
C -15 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal alignment of AAG and AGC.Use a gap penalty of d=-5.

A simple exercise
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 -5 -10 -15
A -5 2 -3 -8
G -10 -3 -3 -1
C -15 -8 -8 -6 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal alignment of AAG and AGC.Use a gap penalty of d=-5.

Traceback
Start from the lower right corner and trace back to the upper left.
Each arrow introduces one character at the end of each aligned sequence.
A horizontal move puts a gap in the left sequence.
A vertical move puts a gap in the top sequence.
A diagonal move uses one character from each sequence.

Start from the lower right corner and trace back to the upper left.
Each arrow introduces one character at the end of each aligned sequence.
A horizontal move puts a gap in the left sequence.
A vertical move puts a gap in the top sequence.
A diagonal move uses one character from each sequence.
A simple example
A A G
0 -5
A 2 -3
G -1
C -6
Find the optimal alignment of AAG and AGC.Use a gap penalty of d=-5.

Start from the lower right corner and trace back to the upper left.
Each arrow introduces one character at the end of each aligned sequence.
A horizontal move puts a gap in the left sequence.
A vertical move puts a gap in the top sequence.
A diagonal move uses one character from each sequence.
A simple example
A A G
0 -5
A 2 -3
G -1
C -6
Find the optimal alignment of AAG and AGC.Use a gap penalty of d=-5.
AAG- AAG--AGC A-GC
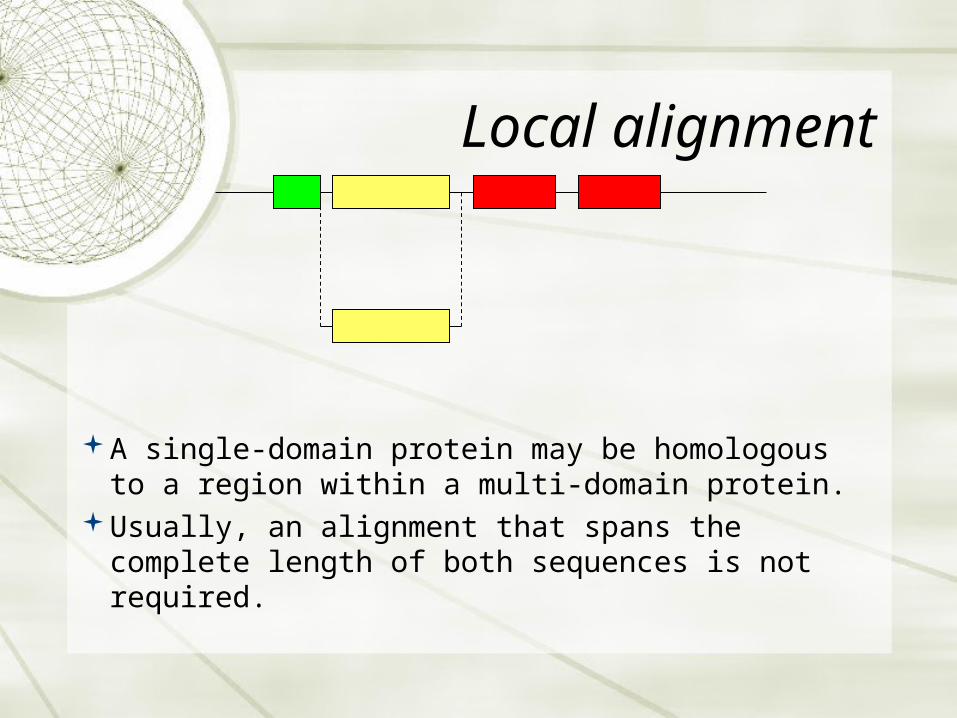
Local alignment
A single-domain protein may be homologous to a region within a multi-domain protein.
Usually, an alignment that spans the complete length of both sequences is not required.
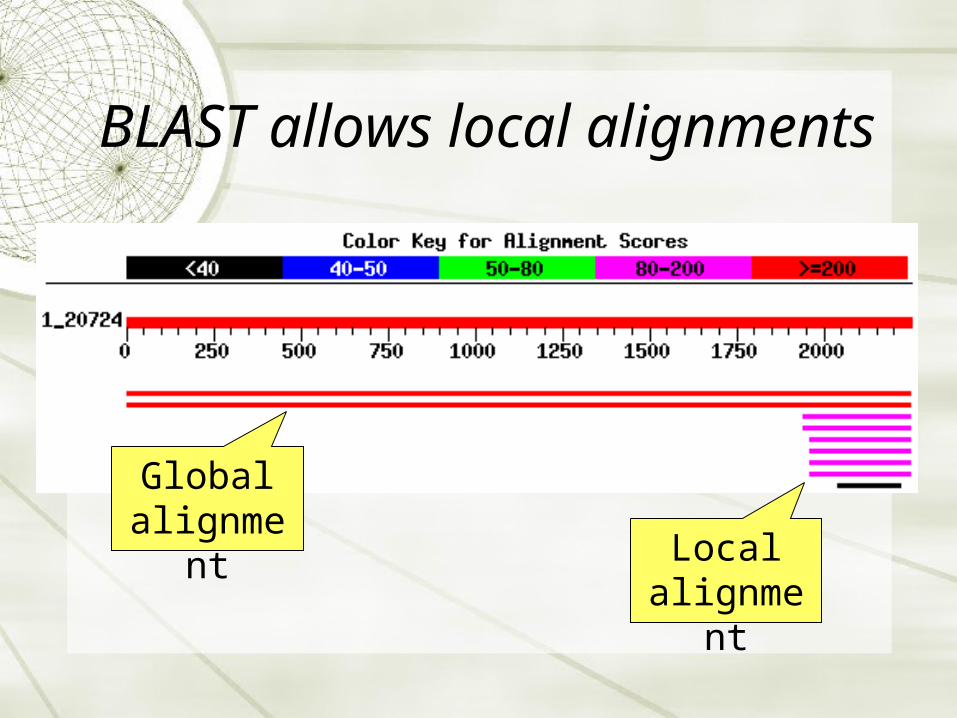
BLAST allows local alignments
Global alignment
Local alignment

Global alignment DP
• Align sequence x and y.
• F is the DP matrix; s is the substitution matrix; d is the linear gap penalty.
djiF
djiF
yxsjiF
jiF
F
ji
1,
,1
,1,1
max,
00,0

Local alignment DP
• Align sequence x and y.
• F is the DP matrix; s is the substitution matrix; d is the linear gap penalty.
0
1,,1
,1,1
max,
00,0
djiFdjiF
yxsjiF
jiF
F
ji

Local DP in equation form
1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
0

Local alignment
Two differences with respect to global alignment:No score is negative.Traceback begins at the highest score in the
matrix and continues until you reach 0.Global alignment algorithm:
Needleman-Wunsch.Local alignment algorithm:
Smith-Waterman.

A simple example
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
A
G
C 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and AGC.Use a gap penalty of d=-5.
0

A simple example
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 0 0 0
A 0
G 0
C 0 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and AGC.Use a gap penalty of d=-5.
0
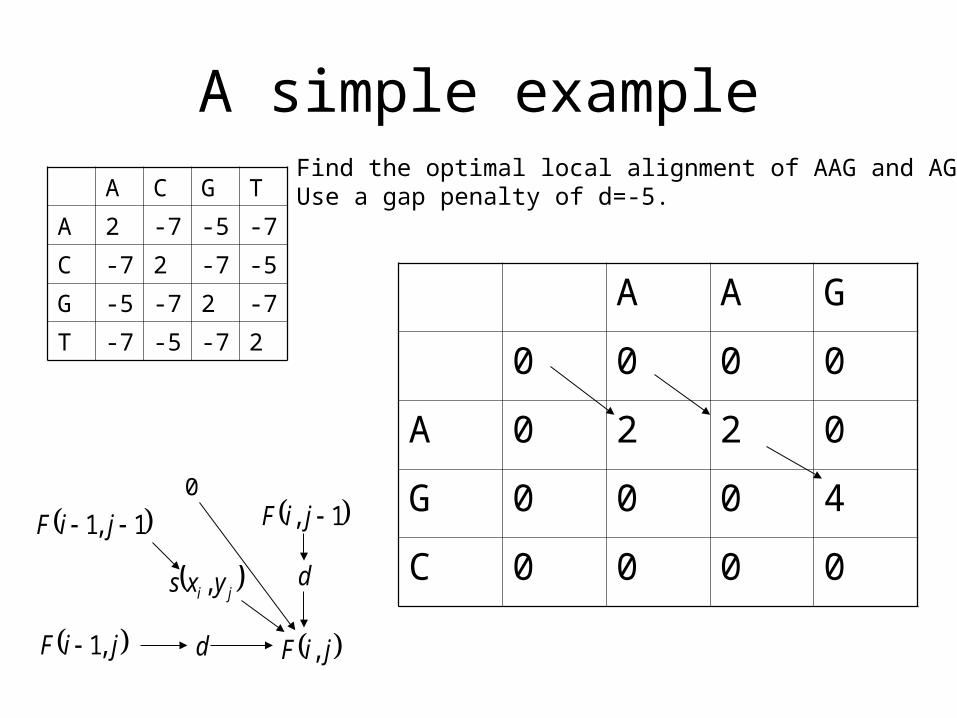
A simple example
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 0 0 0
A 0 2 2 0
G 0 0 0 4
C 0 0 0 0 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and AGC.Use a gap penalty of d=-5.
0
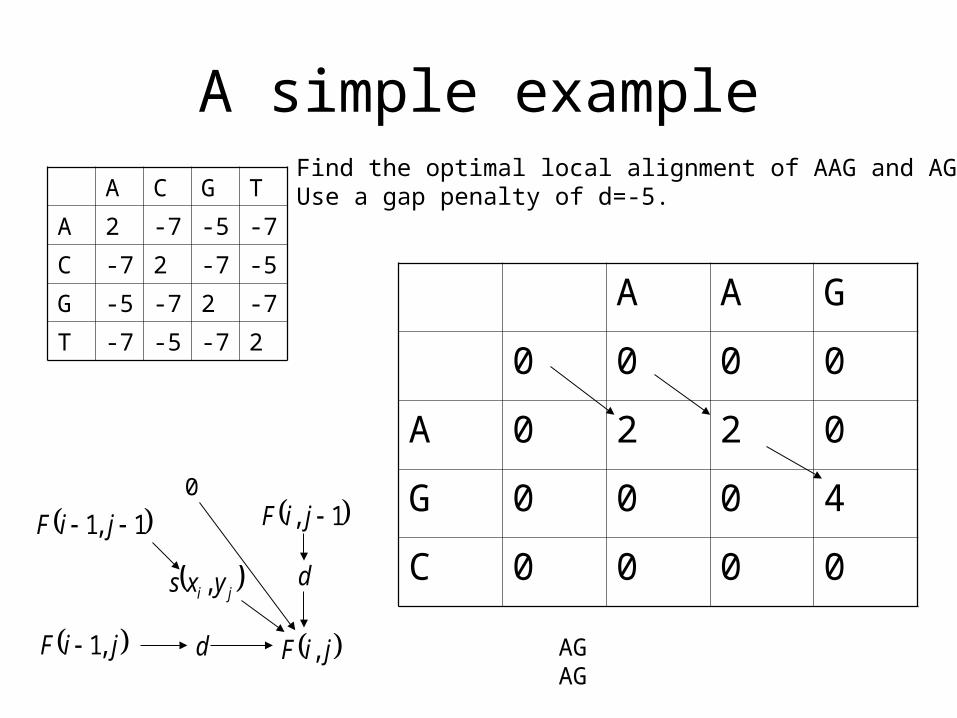
A simple example
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 0 0 0
A 0 2 2 0
G 0 0 0 4
C 0 0 0 0 1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and AGC.Use a gap penalty of d=-5.
0
AGAG

Local alignment
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 0 0 0
G 0
A 0
A 0
G 0
G 0
C 0
1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and GAAGGC.Use a gap penalty of d=-5.
0

Local alignment
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 0 0 0
G 0 0 0 2
A 0 2 2 0
A 0 2 4 0
G 0 0 0 6
G 0 0 0 1
C 0 0 0 0
1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and GAAGGC.Use a gap penalty of d=-5.
0

Substitution matrices
A C G T
A 2 -7 -5 -7
C -7 2 -7 -5
G -5 -7 2 -7
T -7 -5 -7 2
A A G
0 0 0 0
G 0 0 0 2
A 0 2 2 0
A 0 2 4 0
G 0 0 0 6
G 0 0 0 1
C 0 0 0 0
1,1 jiF
jiF , jiF ,1
1, jiF
d
d ji yxs ,
Find the optimal local alignment of AAG and GAAGGC.Use a gap penalty of d=-5.
0
Where did this substitution matrix
come from?
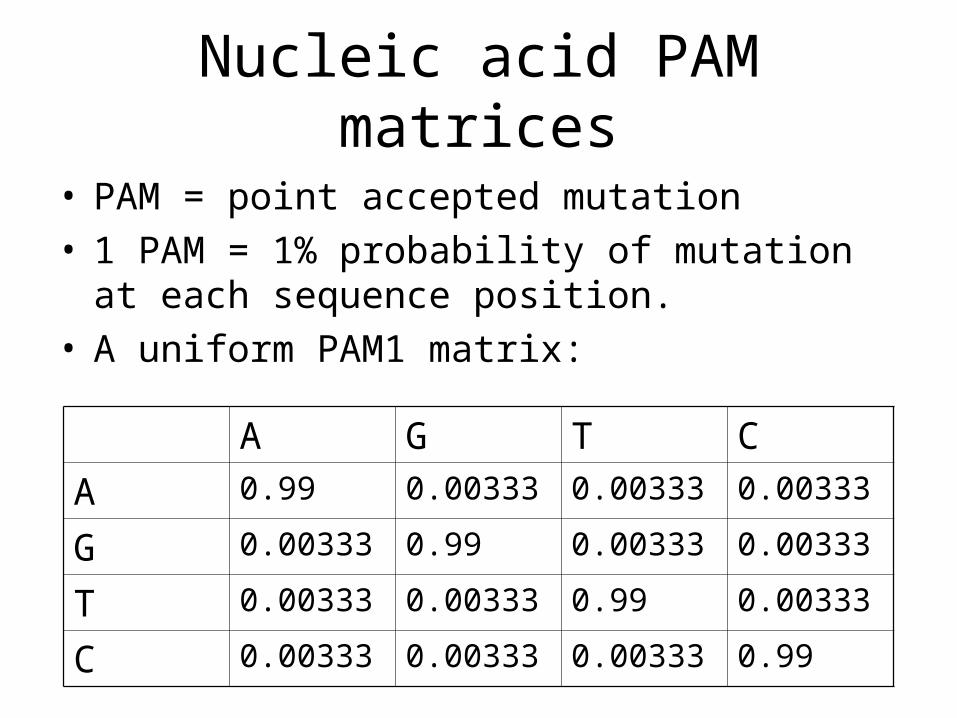
Nucleic acid PAM matrices
• PAM = point accepted mutation• 1 PAM = 1% probability of mutation at each
sequence position.• A uniform PAM1 matrix:
A G T C
A 0.99 0.00333 0.00333 0.00333
G 0.00333 0.99 0.00333 0.00333
T 0.00333 0.00333 0.99 0.00333
C 0.00333 0.00333 0.00333 0.99
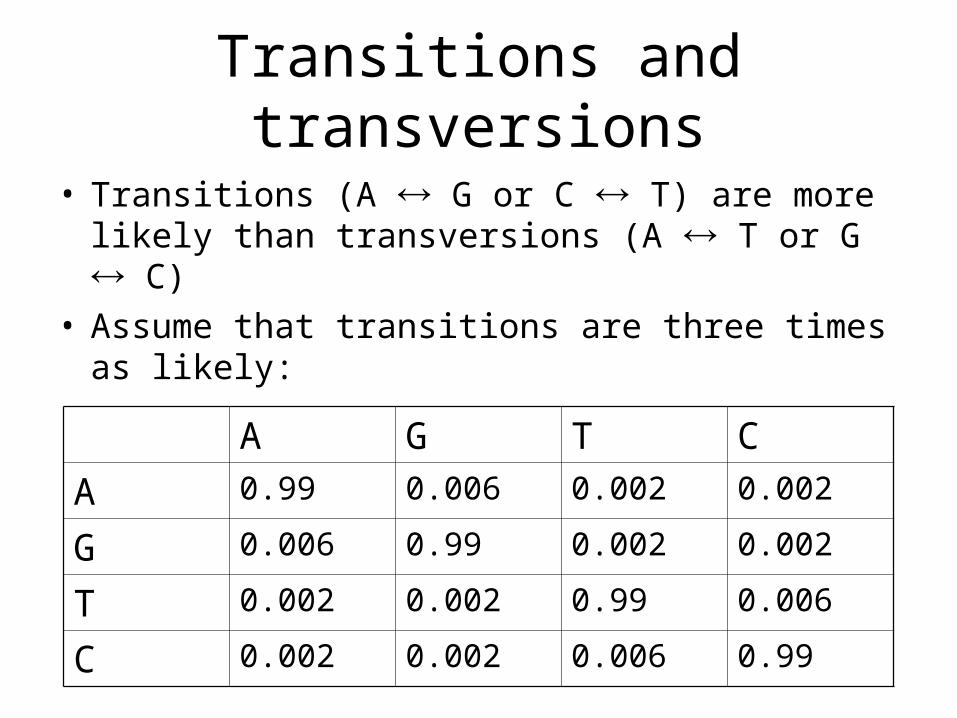
Transitions and transversions
• Transitions (A G or C T) are more likely than transversions (A T or G C)
• Assume that transitions are three times as likely:
A G T C
A 0.99 0.006 0.002 0.002
G 0.006 0.99 0.002 0.002
T 0.002 0.002 0.99 0.006
C 0.002 0.002 0.006 0.99
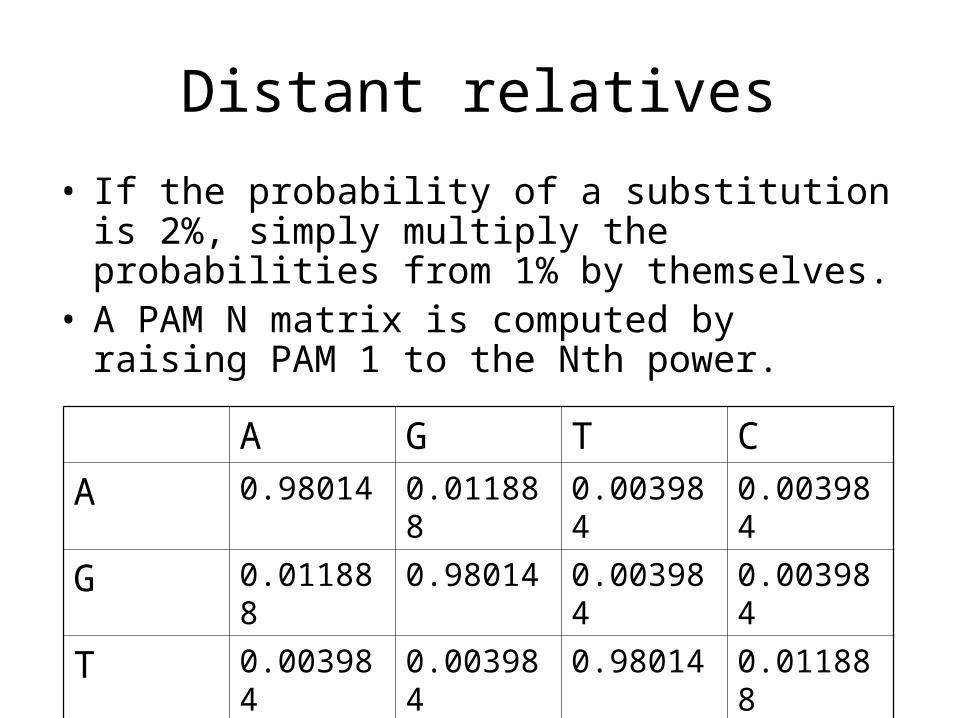
Distant relatives
• If the probability of a substitution is 2%, simply multiply the probabilities from 1% by themselves.
• A PAM N matrix is computed by raising PAM 1 to the Nth power.
A G T C
A 0.98014 0.011888 0.003984 0.003984
G 0.011888 0.98014 0.003984 0.003984
T 0.003984 0.003984 0.98014 0.011888
C 0.003984 0.003984 0.011888 0.98014
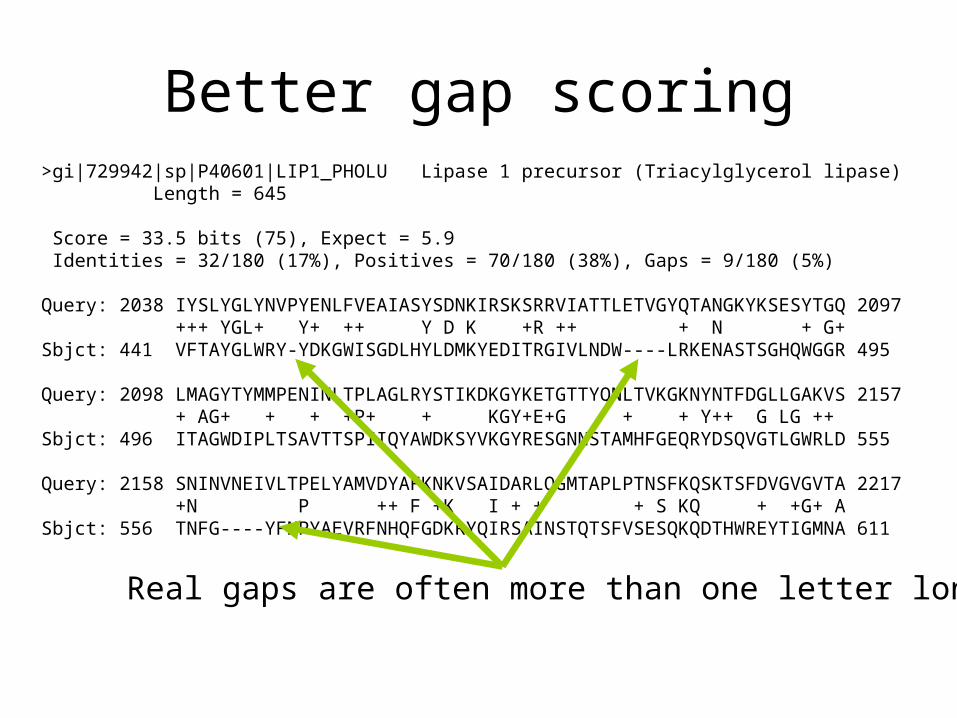
Better gap scoring>gi|729942|sp|P40601|LIP1_PHOLU Lipase 1 precursor (Triacylglycerol lipase) Length = 645
Score = 33.5 bits (75), Expect = 5.9 Identities = 32/180 (17%), Positives = 70/180 (38%), Gaps = 9/180 (5%)
Query: 2038 IYSLYGLYNVPYENLFVEAIASYSDNKIRSKSRRVIATTLETVGYQTANGKYKSESYTGQ 2097 +++ YGL+ Y+ ++ Y D K +R ++ + N + G+Sbjct: 441 VFTAYGLWRY-YDKGWISGDLHYLDMKYEDITRGIVLNDW----LRKENASTSGHQWGGR 495
Query: 2098 LMAGYTYMMPENINLTPLAGLRYSTIKDKGYKETGTTYQNLTVKGKNYNTFDGLLGAKVS 2157 + AG+ + + +P+ + KGY+E+G + + Y++ G LG ++Sbjct: 496 ITAGWDIPLTSAVTTSPIIQYAWDKSYVKGYRESGNNSTAMHFGEQRYDSQVGTLGWRLD 555
Query: 2158 SNINVNEIVLTPELYAMVDYAFKNKVSAIDARLQGMTAPLPTNSFKQSKTSFDVGVGVTA 2217 +N P ++ F +K I + + + S KQ + +G+ ASbjct: 556 TNFG----YFNPYAEVRFNHQFGDKRYQIRSAINSTQTSFVSESQKQDTHWREYTIGMNA 611
Real gaps are often more than one letter long.

Affine gap penaltyLETVGYW----L
-5 -1 -1 -1
Separate penalties for gap opening and gap extension.
This requires modifying the DP algorithm to store three values in each box.

Summary
Local alignment finds the best match between subsequences.
Smith-Waterman local alignment algorithm:No score is negative.Trace back from the largest score in the
matrix.Substitution matrices represent the
probability of mutations.Affine gap penalties include a large gap
opening penalty and small gap extension penalty.