parasites and disease - ny sea grant | welcome
TRANSCRIPT

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
49
Parasitesand
Disease

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
50
to be pathogenic to other organisms. The other parasome-containing amoebae for which sequence data areavailable belong together, and contain the type species of Neoparamoeba, N. pemaquidensis. These species,which include the lobster pathogen and other known pathogenic strains, belong in Neoparamoeba, notParamoeba.
4. Paramoeba eilhardi is a parasome-containing amoeba. Another species has been described, under thename Korotnevella nivo, that is identical to P. eilhardi in gross morphology, habitat, food preference, andultrastructure, but no parasome is present. Molecular sequence data indicate that the two entities are verysimilar, close enough so that both may be treated as species of Paramoeba, perhaps even as strains of P.eilhardi. In P. eilhardi, therefore, the parasome may not be a fixed character. So far, no parasome-freeamoebae related to species of Neoparamoeba have been found, and amoebae (genus Vexillifera) formerlyclassified in the same family (Vexilliferidae) as Neoparamoeba are not even the closest relatives ofNeoparamoeba species. Nevertheless, the possibility of parasome-free strains of Neoparamoeba cannot beignored in attempts to assess presence or absence of pathogenic amoebae.
5. Among strains of Neoparamoeba so far examined, there is considerable variation in normally conservativedomains of the nuclear-encoded small subunit ribosomal RNA (SSU rRNA) gene. The significance of thisvariation is not yet clear, but it is likely that there are many more genotypes of Neoparamoeba in the environmentthan are apparent from phenotype. If so, this complexity needs to be considered when attempting to definegene “markers” for identifying potentially pathogenic Neoparamoeba in the environment.
6. We are in the process of fingerprinting sediment and water samples collected by Mathew Lyman (CTDepartment of Environmental Protection, Bureau of Water Management) to try and identify potential pathogenicNeoparamoeba in the environment and determine their geographic distribution. As mentioned above, weare working to develop a series of specific PCR primers to amplify species from the Neoparamoeba clade,and need to take into account the atypical variations in the normally-conserved domains of the NeoparamoebaSSU rRNA.
7. It has so far not been possible to cultivate the lobster pathogen. We have very recently had success,however, in cultivating Neoparamoeba species from moribund sea urchins (Strongylocentrotus droebachiensis)
Progress in Paramoeba Research
Patrick M. Gillevet1 and Charles J. O’Kelly2
1George Mason University2Bigelow Laboratory for Ocean Sciences
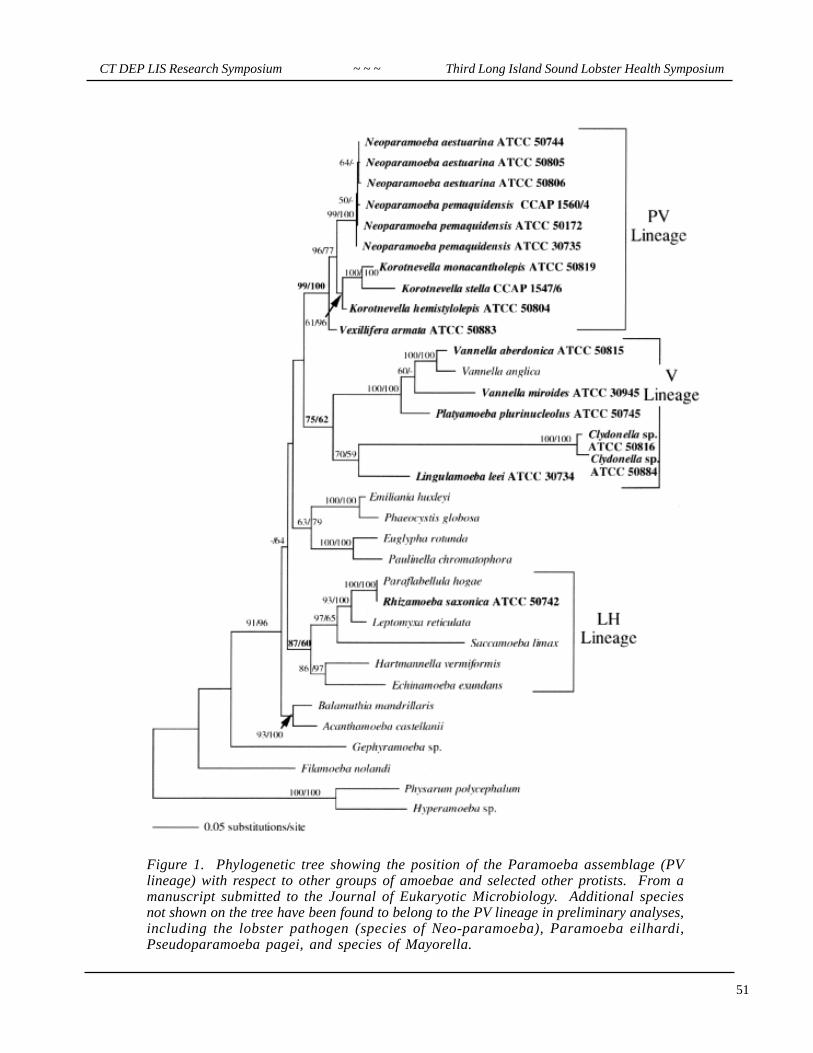
1. Paramoeba and its relatives form a clade of eukaryotes that was previously unrecognized at the molecu-lar level, though it had previously been proposed on the basis of the type of pseudopodia (“dactylopodia”)produced by amoebae in this group. This lineage, designated the PV lineage (Figure 1), includes scale bearingspecies with (Paramoeba) and without (Mayorella, Korotnevella) parasomes, and species without scalesboth with (“Paramoeba”=Neoparamoeba) and without (Vexillifera, Pseudoparamoeba) parasomes.
2. The amoebae in the PV lineage had previously been placed in one or other of three different families;Mayorellidae, Paramoebidae, and Vexilliferidae. If the topology of Figure 1 continues to be supported, all ofthese amoebae will be placed in a single family, the Paramoebidae.
3. The type species of Paramoeba, P. eilhardi, differs from other parasome-containing amoebae in bothultrastructure and molecular sequence. Paramoeba eilhardi is a large amoeba that eats diatoms and is unlikely
CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
51
Figure 1. Phylogenetic tree showing the position of the Paramoeba assemblage (PVlineage) with respect to other groups of amoebae and selected other protists. From amanuscript submitted to the Journal of Eukaryotic Microbiology. Additional speciesnot shown on the tree have been found to belong to the PV lineage in preliminary analyses,including the lobster pathogen (species of Neo-paramoeba), Paramoeba eilhardi,Pseudoparamoeba pagei, and species of Mayorella.

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
52
We gratefully acknowledge the contributions of:
Michael T. Peglar and Thomas A. Nerad (American Type Culture Collection)Thomas F. Mullen and Salvatore Frasca (University of Connecticut)Rebecca Gast (Woods Hole Oceanographic Institution)Linda A. Amaral Zettler and Mitchell L. Sogin (Marine Biological Laboratory)O. Roger Anderson (Columbia University)Jeffrey D. Silberman (Dalhousie University and UCLA)
in the Gulf of Maine (putative sea urchin pathogen N. invadens; Figure 2). Moreover, two strains of theurchin amoeba are presently in axenic culture; if these cultures continue to passage successfully, they willrepresent the first axenic cultivation of paramoebid amoebae known to us. The two strains isolated (bothfrom a single urchin) differ in size and in parasome number; if both are pathogenic, then the possibility thatmultiple infective amoebal strains may be present must be considered for other disease conditions involvingNeoparamoeba strains.
Figure 2. Light micrograph (phase contrast optics)of strain 1 of the Neoparamoeba species isolated(tentatively into axenic culture) from green sea urchinsin the Gulf of Maine. The cell is ca. 10 µm in length.Unpublished. Cells of strain 2 are half again as large(ca. 15 µm length) and may have up to threeparasomes.

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
53
Development of Polymerase Chain Reaction- and in situ Hybridization-basedTests for the Specific Detection of the Paramoeba Associated with EpizooticLobster Mortality by Determination of the Molecular Systematics of the GenusParamoeba
Salvatore Frasca Jr., Kathleen R. Nevis, and Thomas E. Mullen
Department of Pathobiology and Veterinary Science, University of Connecticut, Storrs, CT
IntroductionMass mortalities of American lobster (Homarus americanus) occurred in western Long Island Sound (LIS)during the autumn of 1999. Lobstermen reported dead or dying “limp” lobsters, and there were concurrentreports of increased numbers of dead or dying crabs and sea urchins from LIS. Histopathologic evaluation ofviscera and nervous tissues from dead or dying lobsters revealed hemocytic neuritis and ganglioneuritis. Aprotozoan parasite was identified in foci of neuritis, and this protozoan possessed a paranuclear body thatstained positively for DNA using the Feulgen technique. Transmission electron microscopic studies of thisparasite conducted by Dr. Thomas Burrage (USDA, Plum Island Animal Disease Center, Greenport, L.I.,NY) demonstrated ultrastructural features consistent with a parasome, or “Nebenkörper,” a feature commonto protozoa of the genus Paramoeba Schaudinn, 1896 (Page 1970). The role of this neurotropic paramoeba asa primary or secondary agent in the lobster die-off has not yet been determined, nor have Koch’s postulatesbeen fulfilled. However, species of Paramoeba have been identified as the causative agents of mass mortalitiesof several commercially relevant marine invertebrates, e.g., sea urchins (Strongylocentrotus droebachiensis)(Jones et al., 1985) and blue crabs (Callinectes sapidus) (Sprague et al., 1969). A description of theparamoeba from LIS lobsters and the results of initial investigations into the occurrence of parameobiasis inassociation with the epizootic mass mortality of American lobsters in LIS have been submitted for journalpublication (Mullen et al., In submission). Very little is known about free-living and parasitic species of thegenus Paramoeba Schaudinn, 1896. Molecular data derived from phylogenetically relevant genes (i.e.ribosomal RNA genes) are absent for this genus.
Hypothesis and ObjectivesThe principal goal of this research is to develop sensitive and specific DNA-based tests for the detection ofthis parasitic paramoeba in host tissues by determining the molecular systematics of the genus Paramoebathrough 18S SSU rRNA gene sequence analysis. Development of such DNA-based tests is predicated upondetermination of the molecular systematics of the genus, since DNA sequence data essential for primer andprobe design are currently nonexistent.
The following are the specific aims of this research:1. Determine the molecular systematics of known (i.e. previously identified) species of Paramoeba, or
morphologically related free-living or parasitic amoeba. 2. Determine the phylogenetic relationship of the lobster paramoeba to other known paramoeba species.
3. Develop molecular tests based on 18S SSU rDNA sequence data to detect this parasitic paramoeba inhost tissue.
MethodsSpecific Aim 1. In vitro cultures of strains of three Paramoebid amoebae and five Vexilliferid amoebae wereobtained for 18S SSU rRNA gene sequence analysis. DNA was isolated using silica-gel spin column extractionkits (DNeasy DNA extraction system, Qiagen, Inc., Chatsworth, CA). 18S SSU rDNA was amplified by

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
54
PCR from genomic DNA extracts using universal 18S SSU oligonucleotide primers adapted from Medlin etal., 1988; Hillis and Dixon, 1991; and Weekers et al., 1994. For each strain, PCR products from threeseparate reactions were treated independently of one other. PCR products were purified, ligated into TAcloning vectors (TOPO TA Cloning Vector for Sequencing, Invitrogen Corp., Carlsbad, CA), and cloned.Multiple clones from each of three independent cloning reactions per strain were screened, plasmids wereisolated, and one representative clone from each cloning reaction was sequenced in the sense direction. Sequenceswere determined by oligonucleotide-directed dideoxynucleotide chain-termination sequencing reactions, sequenceABI files were assembled, and consensus sequences were generated from three separate sense sequences foreach strain using Sequencher 4.1.1 for Macintosh. The 18S SSU rRNA gene sequences were then alignedagainst a set of pre-aligned eukaryotic rDNA sequences available through the Michigan State RibosomalDatabase Project II (Maidak et al., 2001) and Genbank (Benson et al., 2002) using ClustalX v1.81 (Thompsonet al. 1994). Phylogenetic trees were inferred by distance and parsimony optimality criteria using PAUP*(Swofford, 2002). Distance-based phylogenetic trees were created using the minimum evolution optimalitycriterion, and a maximum parsimony tree was obtained via random stepwise addition and tree-bisection-reconnection branch-swapping algorithm. The confidence of branching in each case was assessed by 1,000bootstrap re-samplings of the data set, and 200 random sequence addition replicates were used to search forthe most parsimonious tree.
Specific Aim 2. To amplify 18S SSU rDNA from the lobster parasite, paramoeba-infected lobster tissueswere used to provide genomic DNA. Lobsters were either collected by trawl survey by the CT DEP orsubmitted independently by fisherman or biologists. Lobster tissues were processed for paraffin-embedding,evaluated microscopically, and frozen tissue samples from specimens diagnosed as infected by histopathologicalexamination were subjected to nucleic acid extraction (DNeasy DNA extraction system, Qiagen, Inc.,Chatsworth, CA). Order-based and genus-based primers were identified from multiple sequence alignmentsinvolving the 18S SSU rRNA gene sequences of Paramoebid and Vexilliferid amoebae from the first stage inthis project, along with 18S SSU rDNA sequence data from American lobster available through GenBank.Sequences of oligonucleotide primers were selected so as to be conserved among Paramoebid and Vexilliferidgenes and not homologous to lobster 18S SSU rDNA using comparative sequence alignments through ClustalXv1.81 and DNAMAN (Linnon Biosoft, Quebec, Canada). Three sets of inner and outer primer pairs forthree separate nested PCR protocols were identified in order to amplify three overlapping regions of the 18SSSU rRNA gene of the parasitic paramoeba. Primers were tested for specificity by using purified plasmidSSU rDNA of Paramoebid and Vexilliferid amoebae as positive controls and genomic DNA of Americanlobster (Homarus americanus), blue crab (Callinectes sapidus) and green sea urchin (Strongylocentrotusdraebachiensis) as negative controls. Multiple tissues from infected lobsters were tested in triplicate usingone or more of the three, separate, nested PCR protocols. PCR products were treated independently of eachother, and each was purified, ligated into a TA cloning vector, and cloned. One clone from each independentcloning reaction, i.e. representative of an individual PCR product, was sequenced in the sense and antisensedirections, sequences were assembled using Sequencher 4.1.1 for Macintosh, and consensus sequenceswere constructed. Consensus sequences for each of the three overlapping regions of 18S SSU rDNA werein turn aligned to construct a final overall consensus representative of the nucleotide sequence of the 18SSSU rRNA gene of the parasite. To infer the relationships of the lobster parasite to other taxonomically andpathobiologically relevant species, phylogenetic trees were construct based on distance and parsimony optimalitycriteria using PAUP* (Swofford ,2002).
Specific Aim 3. Comparative sequence alignments of 18S SSU rDNA were constructed through ClustalXv1.81 and DNAMAN using sequence data from the lobster parasite, Paramoebid and Vexilliferid amoebae,and American lobster in order to identify target sites for nucleic acid-based diagnostic tests. Oligonucleotideprimers were designed to variable regions of the 18S SSU rRNA gene of the parasite that were not homologousto lobster SSU rDNA. Primer pairs were evaluated theoretically using DNAMAN sequence analysis software

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
55
and in PCR experiments for thermodynamic compatibility, heterodimer and homodimer formation, spuriousreactivity, and product length, and an inner and outer set of primer pairs was selected for a nested PCRprotocol. PCR primers were tested for specificity using purified plasmid SSU rDNA of Paramoebid andVexilliferid amoebae as positive controls and genomic DNA of American lobster, blue crab and green seaurchin as negative controls. Genomic DNA extractions from fresh lobster tissue samples and from formalin-fixed paraffin-embedded tissue sections, examined histologically for the presence of the parasite, are beingprepared to test the application of this nested PCR protocol for the identification of parasite DNA in lobstertissue.
Results Specific Aim 1. The 18S SSU rRNA genes of 3 Paramoebid and 5 Vexilliferid amoebae were amplified,cloned and sequenced, and the contig assemblies of independently derived clones for each particular strain ofamoeba demonstrated less than 1.5% nucleotide ambiguity. Molecular systematic studies undertaken to describethe taxonomic relationships of these amoebae generated a rooted phylogenetic tree with bootstrap values fromdistance and maximum parsimony analyses. In both instances, the Paramoebid-Vexilliferid (P-V) clade aroseas a distinct line of descent separate from genera belonging to the subclass Gymnamoebia. The P-V cladearose before the crown taxa of higher eukaryotes and after basal radiations by eukaryotes such as Vannellaanglica. In addition within the P-V clade, there is a distinct branching that separates the Vexilliferid species,e.g. Neoparamoeba spp, from the Paramoebid species, e.g. Paramoeba sp and Korotnevella spp. The resultsof these SSU rDNA sequence data and phylogenetic interpretations have been contributed to a manuscriptthat is currently in review by the Journal of Eukaryotic Microbiology and represents a collaborative researcheffort with Drs. P. Gillevet, T. Nerad, and C. O’Kelly [Peglar M.T., L.A. Amaral Zettler, R. Anderson, T.A.Nerad, P.M. Gillevet, T.E. Mullen, S. Frasca, Jr., J.D. Silberman, C. O’Kelly, and M. Sogin. Two new small-subunit ribosomal RNA gene lineages within the morphologically defined subclass Gymnamoebia]. Specific Aim 2. Amplification of SSU rDNA of the parasite from genomic DNA extractions of paramoeba-infected lobster tissue was accomplished by using three separate nested PCR protocols targeted at overlappingregions of the 5’-end, internal 1.3-kilobase segment, and 3’-end. For each target region, PCR products amplifiedonce or in duplicate from one or more tissue sources from a minimum of three different lobsters were visualized,purified and cloned. Consensus sequences for each target region were assembled from cloned PCR productsfrom a minimum of three separate lobster hosts, representing a minimum of six double-stranded sequences.An overall consensus SSU rDNA sequence representing the nucleotide sequence of the 18S SSU rRNA geneof the parasite was generated by aligning the separate consensus sequence of each target region. Phylogeneticrelatedness inferred by distance and parsimony analyses and assessed by bootstrap re-samplings of the dataset revealed a very high relatedness of the 18S SSU rRNA gene sequence of the lobster parasite with those ofthe Vexilliferid amoebae, particularly Neoparamoeba pemaquidensis. Branching of the lobster parasite withspecies of N. pemaquidensis was supported by very high distance and parsimony bootstrap values. Specific Aim 3. Comparative sequence analyses, along with theoretical and applied evaluations of primerpairings, generated a nested PCR protocol targeted at a variable region of Paramoebid and Vexilliferid SSUrDNA. In preliminary studies, this nested PCR protocol yielded a 144-base pair product from the 3 Paramoebidand 5 Vexilliferid amoebae tested in specificity experiments, as well as from paramoeba-infected lobstertissue, without amplifying genomic DNA from lobster, blue crab or green sea urchin. Sequence analysis ofthis 144-base pair product distinguishes the lobster parasite from other Vexilliferid or Paramoebid amoebae.DNA extractions and formalin-fixed paraffin-embedded tissue sections are being prepared to test the applicabilityof this nested PCR protocol for the detection of parasite DNA in lobster samples from a large sample groupin comparison to other means of parasite detection, e.g. histopathological evaluation. In addition, probes arebeing prepared to begin in situ hybridization experiments to develop techniques to label the parasite in tissuesections.

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
56
Acknowledgements This research is supported by a grant from the University of Connecticut Research Foundation and by theLong Island Sound Research Fund, Connecticut Department of Environmental Protection, under Grant CWF333-R (to SF).
ReferencesBenson D.A., Karsch-Mizrachi I., Lipman D.J., Ostell J., Rapp B.A., Wheeler D.L.. 2002. GenBank. Nucleic
Acids Res 30: 17-20.Hillis D.M., and Dixon M.T. 1991. Ribosomal RNA:Molecular evolution and phylogenetic inference. Quart.
Rev. Biol. 66: 411-446.Jones G.M., Hebda A.J., Scheibling R.E., and Miller R.J. 1985. Histopathology of the Disease Causing Mass
Mortality of Sea Urchins (Stronglyocentrotus droebachiensis) in Nova Scotia. J. Invertebr. Pathol. 45:260-271.
Maidak B.L., Cole J.R., Lilburn T.G., Parker C.T., Saxman P.R., Farris R.J., Garrity G.M., Olsen G.J., SchmidtT.M., Tiedje J.M. 2001. The RDP-II (Ribosomal Database Project). Nucleic Acids Res. 29: 173-174.
Medlin L., Elwood H.J., Stickel S., Sogin M.L. 1988. The characterizationof enzymatically amplified eukaryotic16S-like rRNA-codong regions. Gene 71: 491-499.
Mullen TE, Russell S, Tucker MT, Maratea JL, Koerting C, Hinkley L, Perkins C, Burrage TG, De Guise S,Frasca Jr. S., and French R.A. Paramoebiasis associated with mass mortality of American lobster(Homarus americanus) in Long Is.land Sound, USA. J. Aquat. Anim. Health. In submission.
Page F.C. 1970. Two new species of Paramoeba from Maine. J. Protozool. 17(3): 421-427.Sprague V., Beckett R.L., Sawyer T.K. 1969. A New Species of Paramoeba (Amoebida, Paramoebidae)
Parasitic in the Crab (Callinectes sapidus). J. Invertebr. Pathol. 14:167-174.Swofford D.L. 2002. Phylogenetic analysis using parsimony (*and other methods). Version 4. Sinauer
Associates, Sutherland, Massachusetts.Thompson J.D., Higgins D.G., and Gibson T.J. 1994. CLUSTAL W: improving the sensitivity of progressive
multiple sequence alignment through sequence weighting, position specific gap penalties and weightmatrix choice. Nucleic Acids Res. 22:4673-4680.
Weekers P.H.H, Gast R.J., Fuerst P.A., and Byers T.J. 1994. Sequence variations in small-subunit ribosomalRNAs of H.artmanella verniformis and their phylogenetic implications. Mol. Biol. and Evol. 11: 684-690.

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
57
Oligonucleotide-based Detection of Pathogenic Neoparamoeba Species
Rebecca J. Gast
Woods Hole Oceanographic Institution,Woods Hole, MA
Introduction Monitoring the environmental distribution of the Neoparamoeba species that infects lobsters is an essentialcomponent to understanding how infections occur and predicting their spread. For example, if the amoeba isan opportunistic pathogen that can normally exist as a free-living organism in the same place as its host, thepotential for repeated infections may be more likely than if the organism needed to be reintroduced fromanother area. Unfortunately, a good understanding of the etiology and distribution of paramoebiasis has beendifficult to attain, due largely to the inability to reliably detect and identify the parasitic species of interest.The identification of Neoparamoeba species is based primarily upon culture of the organism and/or theanalysis of morphologic characters by light or electron microscopy. Issues of culture bias, morphologic variability, level of infectivity and the potential for dormancy have ledus to propose the utilization of gene sequences for the detection and identification of the organism infectinglobsters. Small subunit ribosomal gene (srDNA) sequences contain both invariant and variable regions thatcan be used as templates for the design of oligonucleotide primers (very short pieces of DNA) with specificitiesranging from kingdom to individual isolate levels. The very large number of srDNA sequences available indatabases, such as GenBank, also make this molecule useful because of the large volume of comparativesequence information available. In the past 12 years, the use of ribosomal sequences in ecological studies hasallowed researchers to detect an organism of interest, to determine natural abundances and to follow theorganism’s occurrence over time (for examples, see Amann et al.,1990; Lim et al.,1999; Manz et al.,1993).Coupled with polymerase chain reaction amplification (PCR), the results obtained are not only specific, butthey can also be sensitive.
Objectives
Detection Currently there is no simple and reliable method for the discrimination of the Neoparamoeba thatinfects lobsters from other parasitic, and non-parasitic, paramoebae. Therefore, one of the objectives of myproject is to develop a method for the detection of the lobster parasite that can be used to analyze environmentalsamples for the presence of the parasite, as well as to analyze tissue samples. I have been working to developprimers and protocols for denaturing gradient gel electrophoresis (DGGE) that will permit detection of thelobster parasite (and eventually for the parasitic species from fish, crab, and urchin). PCR amplification of aportion of the small subunit ribosomal gene is employed to generate fragments of DNA from the sample ofinterest. These fragments are separated on a gel with a gradient of denaturant, and the bands DGGE in my labto study the genetic diversity of protists in environmental samples, but the method is also useful for detectinga particular organism. To increase the sensitivity of the method, I have designed primers that are specific forNeoparamoeba species and use them in amplifications prior to amplification with the DGGE primers.
Environmental monitoring The second objective in my project is to use the DGGE method to examine thenatural occurrence of paramoebae in Long Island Sound. I will analyze sediment and water samples that havebeen collected, on a monthly basis, for 18 months. This will allow me to determine whether the lobsterNeoparamoeba parasite is present year-round in the Sound, and will potentially help us to predict the likelihoodfor future outbreaks of infection.

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
58
Progress to Date
This project was initiated in July, 2001. With the assistance of an undergraduate summer student in mylaboratory, we have established protocols for extracting DNA from lobster tissues, and developed fiveamplification primers that would be selective for paramoeba and established amplification parameters torecover neoparamoeba-like ribosomal gene amplification products. We have tissue samples from lobsters andcrabs with confirmed and putative paramoebic infections, and have established an unofficial collaborationwith the other research groups working on the molecular biology of paramoebae in order to make the exchangeof samples and sequences easier. We have been successful in recovering amplifiable DNA from lobster andcrab tissues, and have been able to recover a neoparamoeba product from the lobster tissues (Figure 1).
Figure 1. PCR amplification and reamplificationof samples using Neoparamoeba-specific primersP1, P2 and P3R. Lane 1, 8 – uninfected lobsterDNA; lane 2, 9 – LIS Neoparamoeba isolate A4S;lane 3, 10 – lobster DNA 1268; lane 4, 11 – lobsterDNA 1280; lane 5, 12 – lobster DNA 1352; lane 6,13 – lobster DNA 1697; lane 7, 14 – Neoparamoebapemaquidensis DNA. Samples in lanes 1-7 wereamplified with P1 and P3R. Samples in lanes 8-14were P1/P3R reactions reamplified with P2 and P3R.M = size marker; N = negative control.
Crab samples have not yet yielded neoparamoeba-like products with any of our primers. The crab parasitesequences would be useful for determining whether the lobster amoeba is the same as the one infecting crabs,and for developing specific probes to be used in the detection method, but they are not necessary for furtherdevelopment of the DGGE method. DGGE gels have recently been run using products from infected lobstersamples to identify the Neoparamoeba band (Figure 2), and we are currently working on determining the levelof sensitivity that we can expect for this method.
Figure 2. PCR amplification of bands pickedfrom DGGE gel. Lanes 1 - 4 are productsthat are the correct size for the fragment ofthe Neoparamoeba small subunit ribosomalgene. Lanes 5 – 7 are products that are thecorrect size for the products from the lobstergene. Sequencing will be used to confirmthe identity of the fragments. Lane 8 –negative control; M = size markers.

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
59
Sampling of Long Island Sound sediment and water began in August 2001 in conjunction with the CTDEP Water Quality Monitoring program. Samples are collected once a month at stations A4, B3, C2, D3,E1, H4, J2 and N3 (Figure 3). We completed our sampling this fall and are continuing the processing of thesesamples. DNA is extracted from water within two weeks of collection, and the samples are tested to confirmgeneral amplification competence. Sediment samples are being stored at –80 oC until extraction this year.Although we have successfully recovered DNA from sediments in the past, we are currently testing severalnew protocols that may provide cleaner nucleic acids in less time. All isolated DNA is archived at –20 oCuntil analysis by the DGGE method. In addition to collecting samples for DNA extraction from Long Island Sound, we have collected waterand sediment samples for enrichment culture of amoebae to determine what paramoeba-like species are present.Many amoebae have been cultured, including several paramoeba-like isolates. Three of these have beenconfirmed as Neoparamoeba species based upon their small subunit ribosomal sequences. Two are verysimilar to N. pemaquidensis, and the other is most similar to P. eilhardi. Neither of the N. pemaquidensisisolates has a sequence that is the same as that of the lobster parasite, although they are very similar to eachother. These confirmed sequences come from isolates recovered from sediment samples, but we have alsoobtained paramoeba-like isolates from the water column samples.
Project Significance
Once developed, the Neoparamoeba-specific DGGE will be a simple and relatively quick method fordetecting the presence of the lobster parasite from environmental or tissue samples. Despite the impact ofparasitic paramoebae on several different marine fisheries, we know very little about their natural distribution.The reliable identification of parasitic paramoebae from natural samples would represent a huge step forwardin our ability study these organisms. We will use our method to survey Long Island Sound water and sedimentsamples to determine whether the lobster parasite is endemic to the region. Our results on the natural distributionof the organism may eventually help researchers to predict the potential for outbreaks of disease and theimpact on host populations.
Figure 3. Map of Long Island Sound showing a subset of the 18 sampling stationsmonitored monthly throughout the year by the Connecticut DEP Long Island SoundWater Quality Monitoring Program. The sites indicated on this map are the oneswhere we collect water and sediment samples. (Image modified from the 1998 SummerHypoxia Survey, online at <http://www.epa.gov/region01/eco/lisLis98tot.pdf>.)

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
60
ReferencesAmann, R.I., Binder, B.J., Olson, S.W., Devereux, R. and Stahl, D.A. (1990) Combination of 16S rRNA
targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appliedand Environmental Microbiology, 56, 1919-1925.
Lim, E.L., Caron, D.A. and Dennett, M.R. (1999) The ecology of Paraphysomonas imperforata based onstudies employing oligonucleotide probe identification in coastal water samples and enrichment culture.Limnology and Oceanography, 44, 37-51.
Manz, W., Szewzyk, U., Ericsson, P., Amann, R., Schleifer, K.-H. and Stenström, T.-A. (1993) In situidentification of bacteria in drinking water and adjoining biofilms by hybridization with 16S and 23SrRNA-directed fluorescent oligonucleotide probes. Applied and Environmental Microbiology, 59,2293-2298.

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
61
Bacterial Assemblages involved in the Development and Progression of ShellDisease in the American Lobster
Andrei Chistoserdov1, Roxanna Smolowitz2, and Andrea Hsu2
1University of Louisiana, Lafayette, LA2Marine Biological Laboratory, Woods Hole, MA
Sampling. 25 lobsters with lesions and 6 healthy lobsters were collected from the Eastern Long IslandSound (ELIS) by Connecticut DEP and were made available for this research. Five lobsters, provided by NYDEC, were collected from Long Island coastal waters (LICW). Ten lobsters with shell disease collectedfrom the Buzzards Bay (BB) and one lobster with lesions from the Vineyard Sound (VS) was a gift fromBruce Estrella (MA Division of Marine Fisheries). All lobsters had various degrees of severity of cuticularlesions and primarily substages C3 and C4 of cuticular development. The lobsters were sacrificed and usedfor (1) collection of hemolymph for microbiological analysis, (2) collection of lesion material for microbiologicalanalysis, (3) histological examination. A half of the carapace lesion(s) was used to collect bacterial biomass and a half was preserved in 4%formaldehyde for further histopathological examination. The scraped material from healthy carapaces wasalso suspended in sterile seawater to optical densities similar to those of lesion material suspensions and wasused for DNA isolation. Typically, we collected material only from carapace lesions. However, for ninelobsters (eight from ELIS and one from LICW) we collected material from both carapace and tail lesions.Approximately, 5 ml of hemolymph were drawn directly from hearts of each lobster into sterile Vacutainer®
tubes and refrigerated. Materials collected for gross and microscopic histopathological examination included: fragments of shellwith lesions, hepatopancreas, nerve cord, portions of stomach, gonads, heart, green gland and antennae. Thismaterial was fixed in 10 % formalin in seawater. Fixed tissues were trimmed, decalcified, processed inparaffin and hematoxylin and eosin slides were prepared for examination using standard histological methods.
Culture-dependent microbiological analyses of shell lesion material. Our experiments have shownthat Seawater Agar II (SAII; seawater with 1.7% of agar, 0.1% peptone, 0.01% Tween 80 and vitamin mix)and Marine Agar 2216 are the most adequate media for isolation of chitinolytic and non-chitinolytic bacteriaassociated with shell lesions. Chitinolytic bacteria were selectively cultured in media containing crude chitinpowder from crab shells, which is an adequate imitation of lobster shell material in its biochemical composition,since it contains not only chitin but also proteins and lipids. We could successfully isolate from each lobster three to eight unrelated bacterial strains. Bacterial strainsisolated from different lobsters, colonies of which appeared very similar, turn out to be either identical orclosely related (based on 16S rDNA analysis). Generally, shell lesion material from many but not all lobsterscontained chitinolytic bacteria. However, their isolation in pure cultures was difficult, due to a gliding motilityof non-chitinolytic bacteria and a long time response in the development of positive reactions (sometimes upto two weeks). During this time, gliding bacteria completely engulf colonies of chitinolytic bacteria. Throughmultistage re-streaking, however, we managed to isolated pure cultures of chitinolytic bacteria from fivelobsters one of which was from ELIS and four from BB and one horseshoe crab.
Culture-dependent microbiological analyses of hemolymph. 5 µl of hemolymph from all lobstersand a horseshoe crab was streaked on rabbit or sheep blood agar plates. One set of the plates was incubatedat room temperature and another at 37 oC. No growth was observed on plates incubated at 37 oC. Somebacterial growth was detected on plates incubated at room temperature with plated hemolymph from ELISlobsters #2, 3, 6, 12 and 13 (Table 1). We concluded that there is no correlation between shell disease and

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
62
hemolymph infection. Hemolymph of only four lobsters contained some bacterial contamination and only onelobster (#3) was heavily infected. 16S rRNA analysis of bacterial isolates. 16S rRNA genes from bacteria isolated from hemolymph andchitinolytic bacteria isolated shell lesions were amplified as described by Borneman et al. (Appl. Environ.Microbiol. 62:1935-1943.). On average a 700 bp portion of 16S rRNA gene was sequenced (approximatelybases 600 through 1300, E. coli numbering). The sequence information was used to identify close relatives ofour isolates in the GenBank and Ribosomal DNA project II databases. Based on the similarity with 16SrRNA sequences of closest relatives, the isolates were given either genus (less than 98% identity) or species(98% or more of identity) designation. For the hemolymph isolates H12.1-12.4 and H13.1-13.3 and the shelllesion isolate BA2, identification was inconclusive. The 16S rRNA analysis data are summarized in Tables 1and 2. No typical bacterial pathogens (Aerococcus viridans or Vibrio fluvialis) were found among hemolymphisolates. Chitinolytic bacteria forming yellow colonies, belong to one of the four closely related strains ofCytophaga sp., indicating that these bacteria are ubiquitous in shell lesions. Identical bacteria, i.e. Cytophagasp. strain 23c1, was isolated from ELIS and BB lobsters as well as the horseshoe crab. All Pseudoalteromonasgracilis isolates were identical, indicating that this bacterium is also ubiquitous in shell lesions. It appears thatmicrobial communities found in lesions of lobsters from ELIS and BB are similar to each other. However, themicrobial community in lesions of the Vineyard Sound lobster was different.
Table 1. Bacteria in hemolymph of lobsters:
Table 2. 16S rDNA analysis of isolates from lesions of lobsters with shell disease.

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
63
Culture-independent microbiological analyses. Denaturing gradient gel electrophoresis (DGGE) wasa method of choice to compare microbial communities in the lesions and individual isolates. PCR and DGGEwere carried out as described by Ferris et al. (AEM, 1996, 62:340-346) under conditions, which we optimizedearlier. The best results with PCR products of DNA from shell disease lesions were achieved using thefollowing conditions: temperature of run 60 oC, 14 hours at 100 v, 40-55% gradient of the UF solution, and 9%acrylamide gel. The gel shown in Figure 1 depicts a DGGE gel of the whole community from the lobster #11along with individual isolates from the lesions of this animal (C - Cytophaga sp., P - P. gracilis, A -Alteromonas arctica; S - Shewanella frigidimarina). Both Cytophaga sp and P. gracilis strains used inthese experiments are chitinolytic and clearly present as members of the community. P. gracilis appears tobe a dominant component of the community. Overall, the composition of bacterial communities in the lesionsof BB and ELIS lobsters are very similar in that they have few individual bacterial strains and at least two orthree of these strains are present in all analyzed animals. The composition of the microbial community in thelobster #5 (from LICW), however, was very different from those of ELIS and BB lobsters.
Histopathological assessment. Gross examination of affected animals show moderate to deep erosionsin the hard cuticle. Lesions are most common along the dorsum of the cephalothorax and abdomen, but insevere cases may extend to the claws and lateral and ventral hard carapace. Early lesions appear symmetrically,but further work needs to be done to verify this. Deeper lesions are often brown to black and result insoftened carapace tissue overlying internal connective tissues of the lobster. Histopathologically, carapace erosions are of variable depth; but deep, extensive erosions are common.Early erosions into the epicuticle and exocuticle show invasion primarily around the carapace pores andvertical extension deep into the exocuticle. These erosive areas are melanized. As the lesions deepen, and theexocuticle is eroded, they leave behind cuticular matrix that forms skeletonal pillars in the holes (Figure 2).This contrasts with impoundment shell disease, where erosions do not routinely occur as thin vertical erosionsinto the carapace, but rather appear as progressive areas of scooped out cuticle that leave no cuticular matrixbehind. More severe lesions show erosions that extend deeper into the calcified and uncalcified endocuticle.Melanization of the affected tissue is diffuse and in the deeper lesion the pillars of cuticle break off. Variablebut often extensive layers of new uncalcified carapace (endocuticle/membranous layer) are produced by theintact hyperplastic epithelium underling the eroded site. This mechanism appears to prevent eventual ulcerationinto the underlying soft tissues of the animals body that could result from progressively deepening erosions. Inflammation in epizootic shell disease is composed of increased numbers of mixed populations of hemocytetypes in the underlying connective tissues and accumulations of usually necrotic hemocytes between layersof carapace. Inflammation, and cuticular proliferation, as well as melanization, of the affected cuticle are alsoseen in impoundment shell disease and are general responses to erosion of the cuticle for any reason.
Figure 1. A DGGE gel of thewhole bacterial community fromlobster #1, along with individualisolates from the lesions of thisanimal.

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
64
Figure 2. Early erosions into theepicuticle and exocuticle show invasionprimarily around the carapace poresand vertical extension deep into theexocuticle. These erosive areas aremelanized. As the lesions deepen, andthe exocuticle is eroded, they leavebehind cuticular matrix that formsskeletonal pillars in the holes.
Ulceration, focal loss of all carapace and invasion into underlying lobster connective tissues is very rare intissues examined to date. Once ulceration occurs, intense inflammatory reaction produces a melanizedpseudomembrane that covers the lesions. These foci can inhibit molting by causing attachments between oldand new carapace. No other disease has consistantly been identified to date in animals affected by epizootic shell disease(determined by examination of other body tissues), thus indicating epizootic shell disease is not secondary tosome other primary disease (such as parameoba infections or gaffkemia). Various organisms are identified in the shell erosions. Nematodes, sponge, algae and ciliates are occasionallyseen. An as yet unidentified smaller protozoan is often seen in lesions and may be secondarily important inlesion development. But, by far the predominate organisms found at the interface of necrotic and live shell inboth shallow and deep erosions into the cuticle are bacteria. Gram staining shows Gram negative bacteria inthis position.
Infection experiments. A series of infection experiments has been commenced at the Flax Pond MarineLaboratory. Groups of healthy lobsters (5 individual each) were exposed to isolated P. gracilis and individualCytophaga strains (106 cells of each per liter of seawater) for 24 hours. The carapace of two out of the fivelobsters in each experiment was mechanically breached. To elucidate transmittance of shell disease, in aseparate experiment, four healthy lobsters (epicuticle of two of them was mechanically damaged) are kept inthe same tank with two lobsters with shell disease. No transmission of infection occurred after 6 month ofincubations.

CT DEP LIS Research Symposium ~ ~ ~ Third Long Island Sound Lobster Health Symposium
65
Calcinosis in LIS Lobsters During Summer 2002
Alistair D.M. Dove1, Carl P. LoBue2 and Paul R. Bowser3
1Department of Microbiology and Imunology, Cornell College of Veterinary Medicine, Ithaca, NY
2NY Department of Environmental Conservation, East Setauket, NY
3Department of Microbiology and Imunology, Cornell College of Veterinary Medicine, Ithaca, NY
A significant number of moribund and dead lobsters were reported to state authorities by lobster fishers inLong Island Sound, New York, during the summer of 2002. The first of these were reported by a lobsterfisher operating out of Mattituck who reported them to Cornell Cooperative Extension. Samples from thisfisher were submitted to the new Marine Disease Laboratory at SUNY Stony Brook where they werenecropsied and examined for infectious and other diseases. Morbid lobsters were characterized by anorange discoloration of the abdomen, lethargy, an excess of epibionts and poor post-capture survival. Mostaffected lobsters were in late intermolt or premolt stages. On necropsy, severe extensive multifocal ordiffuse granulomatous inflammation of the gills and antennal glands was the most striking pathology. In thegills, granulomas were frequently seen to be embolised in filaments, resulting in congestion, ischemia (circulatoryblockage) and coagulative necrosis of gill tissues. In the antennal glands, granulomas were concentratedalong the border between the filtration and resorption zones of the organ. Affected lobsters lacked observablereserve inclusion cells (energy storage cells) and thus appeared to be either malnourished or metabolicallyexhausted (we suspect the latter). No significant pathogens were recovered from diseased individuals,suggesting that the disease is of metabolic origin. In prechronic individuals, however, it was evident thatgranulomas were focused around calcium carbonate (aragonite) crystals. Aragonite crystals were identifiedby their spheroid shape, radial striations, clear to golden brown coloration and strong birefringence. In earlystage individuals, naked aragonite crystals were observed, whereas in later stage individuals, aragonite crystalswere observed to be at the centre of granulomas. In most cases, the granulomas had continued to mineralizein an amorphous fashion. As far as we are aware this is the first report of such a disease in lobsters. While microgranulomas of thetype we observed are not unusual in lobsters, the number observed in our study, their focus around inorganicmineral crystals and their potential to cause what appear to be fatal pathologies in the gills and antennalglands has not been reported previously. While it is not yet clear why this disease occurs, our best hypothesis of etiology is as follows. Calcinosisis probably caused by anomalously high sea bottom temperatures in Long Island Sound (~23 ºC) during thesummer of 2002 and associated disruptions of the calcium chemistry of lobsters in favor of deposition ofcalcium minerals in soft tissues. We hypothesise that temperature-related respiratory stress results inhypercapnia (excess CO
2 in the hemolymph). Some of this CO
2 enters to carbonate/bicarbonate hemolymph
buffering system of the lobster and becomes available as anionic conjugates for circulating calcium ions. Wesuspect that the excess anions combined with high hemolymph calcium concentrations during some phases ofthe molt cycle push the reaction in favor of deposition of calcium as crystalline CaCO
3 (aragonite). The
concentration of the calculi in the two main filtration and excretion organs of the lobster to the exclusion ofmost other tissues suggests that the crystals form in the hemolymph and are subsequently filtered out in theantennal gland and (later) the gills, where they lodge and become the focus of granulomatous inflammation.Exactly why the calculi should elicit an inflammatory response is not clear, given that calcium carbonate isoften exposed to lobster tissues in the form of the exoskeleton and gastroliths; this phenomenon may representa tissue specific reaction since neither the gills nor the antennal glands would be exposed to crystallinearagonite in a healthy lobster. We suspect that under appropriate conditions the disease represents a fatalpositive feedback loop in which continuing obstruction of the gills reduces the lobsters’ ability to rid itself ofCO
2, thus increasing the hypercapnia and favoring the disease progression. There may be thresholds of

Third Long Island Sound Lobster Health Symposium ~ ~ ~ CT DEP LIS Research Symposium
66
Top Left: Healthy (L) and calcinotic (R) lobster abdomens showing distinctiveorange colour in affected lobster. Top Right: gill tissue with multiplegranulomatous lesions (brown areas) and characteristic abundance of epizooticcolonial bryozoan Triticella sp. Bottom Left: brightfield (L) and darkfield (R)images of the same granuloma resolving crystalline aragonite nucleus (arrow)under darkfield illumination. Bottom Right: Histological section of mineralizedgranulomas in antennal gland tissues.
temperature and hypoxia that trigger the disease state, and we suspect that even moderate hypoxia may beclinically significant for calcinotic lobsters at hypermetabolic temperatures. The ultimate cause of death for affected lobsters is probably respiratory failure due to reduced effectivesurface area, exacerbated by the effects of hyperthermia on metabolic rate and an excess of epibionts. Highamplitude climate cycles or permanent climatic change may play a role in the emergence of this new fataldisease of lobsters.