pathogenesis of ankylosing … of ankylosing spondylitis and rheumatoid arthritis proceedings ofthe...
TRANSCRIPT

PATHOGENESIS OF A N K Y L O S I N G SPONDYLITIS A N D
R H E U M A T O I D ARTHRITIS
P r o c e e d i n g s o f t h e Second I n t e r n a t i o n a l Symposium Heid a t t h e M i d d l e s e x H o s p i t a l
1 4 - 1 5 A p r i l 1 9 8 7
O R G A N I Z I N G INSTITUTIONS O R G A N I Z I N G C O M M I T T E E
Department of Rheumatology Research. D R . M A R Y C O R B E T T Univcrsitv College and Middlesex Hospital Medieal School, D R . A . E B R I N G E R London W l 2PG.
D R . M . S H I P L E Y
Immunology Unit, Department of Biochemistry, King's College, Univcrsitv of London. London W8 7 A H .
D e d i c a t e d t o D r . D . C O. James, C o n s u l t a n t H a e m a t o l o g i s t a t t h e Westminster H o s p i t a l a n d
M e d i e a l D i r e c t o r of t h e A n t h o n y N o l a n L a b o r a t o r i e s

CONTENTS
List of Participants IV
Introduction by A l a n E b r i n g e r \\\\
Tuesday 14 April 1987 FIRST SESSION Morning C h a i r m a n : Mary Corbett
1. Perspectives in the Actioloev of Seronegative Polyarthritis by V. N e u m a n n a n d V. W r i q h t 1 2. Geneticsof HLA-B27/?v M . ' / L K h a n " 6 3. Molecular Biology of the HLA-B27 Locus by E l i s a b e t h H . Weiss, K a t h i B l o e m e r ,
C h r i s t a D o e m e r , W. K u o n , M a r g o t L a n g , H e i k e P o h l a , M . S c h a t t e n k i r c h n e r a n d G. Riethmüller 12
4. HLA-B7/B27 Constructs: a Tool to Understand the Molecular Mimicry Hypothesis between HLA-B27 and Gram-negative Bacterial Antigens in Ankylosing Spondylitis bv A . T o u b e r t , ./. S i t e , R. Sodoyer, B . A m o r a n d B . R. J o r d a n 19
5. Lymphocyte Response to Enterobacterial Biostructures in Scronegative Spondarthritis: Specific T-cell Mediated Immunity or Non-spccific Polyclonal B-cell Activation? by W. L . G r o s s , C. L o h m a n n , G i t t a Lüdemann a n d J'. Lüdemann 23
6. Gram-negative Bactcria, HLA-B27 and Inflammatorv Bowel Disease bv P . E . Pease, M . R. L a n e , H . C h a h a l , J . E . T a l l a c k a n d R. N . A l l a n ' 29
7. Cross-rcactivity Studics on Bactcria Believed to bc Associated with Inflammatorv Bowel Disease (IBD), Ankylosing Spondylitis (AS) and Reactivc Arthritis (ReA) bv P . E . Pease, H . C h a h a l , J . E . T a l k i c k , M . R. L a n e a n d R. N . A l l a n 32
8. Criteria for Ankylosing Spondylitis: Facts and Fallacies by J . M . H . M o l l 34 9. The Association between Ankylosing Spondylitis, Acute Anterior Uveitis and HLA-B27 :
the Results of a Swiss Family Study by S. M . v a n der L i n d e n , H . U. Rentsch, N . G e r b e r , A . C a t s a n d H . A . V a l k e n b u r g 39
10. Acute Anterior Uveitis and Faecal Carriage of Gram-negative Bactcria by R. E b r i n g e r 42
Tuesday 14 April 1987 S E C O N D SESSION Aftcrnoon C h a i r m a n : Michael Shipley
I L Factors Associated with the Development of Reactive Arthritis by A . T o i v a n e n , R. T e r t t i , R . L a h e s m a a - R a n t a l a , T. H . S t a h l b e r g a n d K . G r a n f o r s 46
12. Experimental ym7«/a-associated Arthritis in the Spontaneously Hypertensive Rat by P . T o i v a n e n , R. M e r i l a h t i - P a l o , C. G r i p e n b e r g , K . - O . Söderstrom a n d IJ.-M. J a a k k o l a . 52
13. Molecular Mimicry: Fact or Fiction? by D . T. Y. Yu 55 14. Rabbit Antisera lo Enterobacteriaceae isolated from H L A - B 2 7 Positive Patients with
Ankylosing Spondylitis are Lytic for the Mononuclear Cells of AS Patients by C. J . B e n k e l m a n , A . v a n L e e u w e n , P . C. A e r l s , E . v a n K r e g t e n , S. v a n B r e e , J . ./. v a n Rood, J . M . N . W i l l e r s a n d H . v a n Dijk 58
15. Enterobacterial Involvement in the Pathogenesis of Secondary Ankylosing Spondylitis by C h . G. v a n Bohemen, E . W e t e r i n g s , H . S. G o e i T h e , E . C. G r u m e t a n d H . C. Zanen 61
16. Expression of H L A Antigens on the Human Uvea by D . A b i - H a n n a a n d D . W a k e f i e l d 68 17. K l e b s i e l l a Antibodies in Ankylosing Spondylitis and P r o t e u s Antibodies in Rheumatoid
Arthritis by A . E b r i n g e r , N . L . C o x , I . A b u l j a d a y e l , M . G h u l o o m , S. K h a l a f p o u r , T. Ptaszynska, E . S h o d j a i - M o r a d i a n d C. W i l s o n 72
18. Antibodies to K l e b s i e l l a and P r o t e u s Microorganisms in Ankylosing Spondylitis and Rheumatoid Arthritis Patients Measured by E L I S A by S. K h a l a f p o u r , A . E b r i n g e r , I . A b u l j a d a y e l a n d M . C o r b e t t 86
19. Antibodies to P r o t e u s in Rheumatoid Arthritis by P . Rogers, J . H a s s a n , B . B r e s n i h a n , C. E e i g h e r y a n d A . W h e l a n 90
20. Ileocolonoscopic Findings in Seronegative Spondylarthropathies by H . M i e l a n t s . E . M . Veys, C. C u v e l i e r a n d M . D e Vos 95
21. Pathogenesis of Ankylosing Spondylitis: the State of the Art by A . C a l i n 106
ii

Wednesday 15 April 1987 T H I R D SESSION C h a i r m a n : Ivan Roitt, F.R.S.
Morning
22. Historical Aspects of the Aetiology of Rheumatoid Arthritis by E . G. L . By w a t e r s 110 23. Pathophysiology of Chronic Synovitis by J . C. W. E d w a r d s 116 24. Is there a Role for Epstein-Barr Virus in the Aetiology of Rheumatoid Arthritis? by P . M .
L y d y a r d a n d W. L . I r v i n g 120 25. Abnormal Mitochondria in Cultured Synovial Fibroblasts in Rheumatoid and Reactive
Arthritis? by E . E e r o l a , K . P u l k k i , L . J . P e l l i n i e m i , E . V u o r i o a n d A . T o i v a n e n 128 26. H L A - D R 4 and Other Genetic Markers in Rheumatoid Arthritis by P . Stastny, E . J . B a l l ,
M . A . K h a n , N a n c y J . O l s e n , T. P i n e a s a n d X . G a o 132 27. Antigens of H L A - D R Locus in Rheumatoid Arthritis by Z. Jajic a n d I . Jajic 139 28. Induced Expression of Class II Transplantation Antigens in the Cartilage-Pannus Junction
in R A : Chronic Synovitis as a Model System for Aberrant T-lymphocyte Activation by L . K l a r e s k o g a n d O. J o h n e l l 141
29. Prevalence of Familial Occurrence in Patients with Rheumatoid Arthritis by F. Wolfe, Sue M . K l e i n h e k s e l a n d M . A . K h a n 150
Wednesday 15 April 1987 F O U R T H SESSION Afternoon C h a i r m a n : J. C. W. Edwards
30. Pathogenese Factors in Rheumatoid Synovitis M . Ziff D . C a v e n d e r a n d D . H a s k a r d 153 31. Rheumatoid Factor: Structural and Genetic Studies indicating Novel Binding Sites may
confer the Specificity for IgG Fe by F. C. H a y 157 32. Rheumatoid Arthritis as a Glycosylation Disorder by R. B . P a r e k h , R. A . D w e k a n d T. W.
Rademacher 162 33. Decreased B-cell Galactosyltransferase activity in Rheumatoid Arthritis by J . S. A x f o r d 170 34. The Middlesex Hospital Prospective Study of Early Rheumatoid Disease by Mary C o r b e t t
a n d A . Y o u n g 171 35. Observations on the Causes of Rheumatoid Arthritis by S. A t k i n , D . W a l k e r , M . M a n d e r ,
A . M a l c o h n a n d W. C a r s o n D i c k 173 36. An Overview of Research into Ankylosing Spondylitis and Rheumatoid Arthritis by R. N .
M a i n i 176
iii

B r i t i s h J o u r n a l of Rheumatology 1988;27(suppl II): 12-18
MOLECULAR BIOLOGY OF THE HLA-B27 LOCUS B Y E L I S A B E T H H . W E I S S , K A T H I B L O E M E R , C H R I S T A D O E R N E R ,
W . K U O N , M A R G O T L A N G , H E I K E P O H L A , M . S C H A T T E N K I R C H N E R AND G . R I E T H M Ü L L E R
Correspondence to Dr. Weiss.
SUMMA RY
The molecular biology of the H L A - B 2 7 locus is rcviewed. The H L A - B 2 7 gene itself does not differ between healthy individuals and ankylosing Spondyl i t i s patients. Several u n i q u e features o f the H L A - B 2 7 molecule have been identified and one epitope was proposed to cross-react with bacterial proteins.
K F . Y W O R D S : D N A , Intron probe, Polymorphisms, Cosmid clones.
T H E human major histocompatibility complex ( M H C ) or H L A complex (human leucocytc antigen) located on the short arm of chromosome 6 contains at least 17 closely linked genes, which code for the heavy chains of the class I antigens. These glycosylated Polypeptides are expressed on the cell surfacc associated with ß 2-microglobulin. The gene produets of the highly Polymorphie loci, H L A - A , H L A -B , and H L A - C are transplantation antigens which have been shown to serve as recognition and restriction elements for cytotoxic T-lymphocytes. Because of the importance of these antigens in clinical transplantation, the genetic polymorphism of the H L A complex has been extensively studied. The finding that H L A antigens are associated with disease susceptibility was rather unexpected. The most remarkable association is that of the H L A - B 2 7 allele and ankylosing Spondylitis (AS) with a relative risk of 69 [1-3]. Although the strength of the association of the HLA-B27 antigen and AS renders the HLA-B27 gene unique among the class I gene family, the HLA-B27 gene was only recently isolated. Several groups have cloned H L A - B 2 7 gene sequences (see Table I) [4-7] and the amino acid sequences of four H L A - B 2 7 Subtypes have also been obtained by protein sequencing [8-11]. The gene Organization has been determined by isolating c D N A and genomic cosmid clones [4,5] from gene libraries construeted of D N A prepared from peripheral white blood cells (PBL) and of R N A isolated from a homozygous B-cell line (B L C L ) of persons with no history of disease. Seemann et a l . [6] reported two H L A - B 2 7 subtype sequences cloned from a B-cell line (HLA-B27.1/27.2) which differ only by four nucleotides resulting in three amino acid exchanges. A l l HLA-B27 sequences are identical in the protein coding regions, the differences found in the L G 2 - c D N A sequence being due to c D N A cloning artefacts. It is not known whether the several nucleotkle differences detected in the intron sequences are due to a low genetic variabilitv of intron sequences or might be explained by sequencing errors.
Although previous studies did not give any indication that the HLA-B27 molecule of AS patients
might differ from the antigen of healthy individuals, the possibility still remained of a pathogenic HLA-B27 variant present in affected individuals which has not so far been detected at the protein level. We therefore isolated H L A - B 2 7 specific c D N A clones from a c D N A library construeted with
T A B L E I T A B L E OF H L A - B 2 7 SEQUENCES
Library Source H L A haplotype D N A sequence AS Publication
C D genomic P B L A2 , B27/51, Cw2/4 complete gene - f Weisse/«/. [4) LG2 c D N A B L C L A2, B27, Cwl coding - m Szoets et al.[5\ B R U G genomic B L C L A3/11, B27.1/2 Cwl/2 complete gene Seemann et <//.[6]
gap in intron 3 Seemann et <//.[6]
K C A genomic B L C L A l l / w 2 4 , B27/w44, Cwl/5 exon 2 - 4 - m Coppin & McDcvitt[7]
M R W C genomic B L C L A2/32, B14/27, Cw2 exon 2 - 4 + Coppin & McDevitt[7]
M V L genomic B L C L Aw32, B27, Cw2 exon 2 - 4 — Coppin & McDevitt[7)
Correspondence to Dr. Weiss.

WEISS E T A L . : M O L E C U L A R B I O L O G Y O F T H E H L A - B 2 7 L O C U S 13
DNA from P B L of an AS patient (HLA-A2/23 , B27/w60, Cw2/3) and Coppin and McDevitt [7] cloned the genomic sequence using a B-cell line established from an affected individual. The D N A seqience data, derived from the protein coding and the 3'untranslated region, demonstrate that the HLA-B27 gene is identical in healthy persons and A S patients.
The genomic Organization of the H L A - B 2 7 gene, that is the excon/intron structure, is similar to thatof other human class I genes (Fig .L) . The overall homology, between the HLA-B27 gene and other H L A - B alleles, e.g. H L A - B 7 , is the same as is observed when comparing two H L A - A alleles. A hgh degree of sequence divergence is found outside of the gene itself, in the 3' flanking region, whei the HLA-B27 is compared to H L A - B 7 f 12] or HLA-B51 [13], alleles for which D N A sequence data in this region are availablc. The genomic sequence of the HLA-B27 gene does not provide an obvhus explanation for the disease association of this H L A - B allele.
H L A - B 27.1
m RNA
Protein
Fic;. 1. — Structure of the HLA-B27.1 locus.
C0M>ARIS0N OF HLA-B27 SEQUENCES U1TH OTHER HLA-B ALLELES:
EBV. BLRF 60 80 90 KLEB.PNEU
B27 1 U D R E T 0 1 C K A K A Q T D R E D L R T L L R Y Y N Q S E A B27 2 N . . I A B27 3 S B7 . . . N . . . Y . . Q S . . N . R G B8 . . . N . . . F . T N T S . . N . R G B44 S . T N T . . Y . . N . . . A A B51 . . . N . . . F . T N T . . Y . . N . . . R B58 . . G . . R N M . . S . . . Y . . N . . I A Bu6: S . T N T . . Y . . S . . N . R G
* 100 130" 150 B27 1 G S H T L Q N M Y G C - - L S U T A A D T A A - - A R V A E Q L B27 3 . . E . . . . B7 S . . . . . R . . E . . . R B8 S . . . . . R D B44 . . . I I . R . . . . D B51 . . . . U . T . . . . . . E . . . . B58 . . . I I . R . . . . Bu6! R . . . . . . E . . . U
Fic . 2 .— Comparison of H L A - B 2 7 sequences with other H L A - B antigens.

14 P A T H O G E N E S I S O F A N K Y L O S I N G S P O N D Y L I T I S A N D R A
The protein nature of the HLA-B27 molecule is similar to other class I antigens, as the Polymorphie amino acid substitutions are found in the Polymorphie segments of the class I heavy chain (Fig. 2). Three amino acid residues (aa) are present which are unique to the HLA-B27 antigen and its Subtypes, and are not found in any other H L A molecule sequeneed so far. Two of these changes can be valued as unremarkable as they represent conservative substitutions: lysine ( K ) in aa position 70 and asparagine (N) in 97. Other H L A antigens also contain a basic amino acid residuc in these positions. These conservative substitutions are unlikely to influencc the structure or biological activity of the molecule in a unique fashion. One replacemcnt, however, seems unusual; position 67 is occupied by a potentially reactive cysteinc residuc (C) that might influencc the intcrmolecular interactions of HLA-B27 , due to its location in an area that is probably exposed on the surfacc of the membranc molecule. We are currently using site directed mutagenesis to investigate what influencc the HLA-B27 unique amino acid substitutions have on the structure of the molecule as detected by serological reagents or by allospecific cytotoxic T-lymphocytes. The structural uniqueness of H L A -B27 which might be rcsponsible for its disease association. may be ercated by several contiguous amino acid residues in the Polymorphie regions. On the other hand it also might be a unique conformational epitope of the HLA-B27 molecule. It can be postulated that the HLA-B27 unique substitutions conserved among the Subtypes might be involved in the major antigenic determinants of HLA-B27 and might serve as a molecular site of correlation with the disease.
Oldstone et cd. [14] have described a linear sequence of six amino acids present both in H L A -B27.1 and in K l e b s i e l l a p n e u m o n i a e (Fig. 2) which, they propose, represents the epitope responsiblc for the cross-reactivity of HLA-B27 and K l e b s i e l l a , first described by Ebringer [15]. As no Single HLA-B27 subtype shows a preferential correlation with A S , the putative cross-reactive epitope should be common to all HLA-B27 variants. Only five amino acids of the K l e b s i e l l a sequence are conserved in HLA-B27.2 and HLA-B27.3 . The HLA-B27.3 subtype sequence in this region is identical to the sequence present in the H L A - B 7 [12] and the H L A - B 8 [16] antigen. It therefore remains to be studied if the proposed amino acid sequence of K l e b s i e l l a p n e u m o n i a e is responsible for the cross-reactivity observed between H L A - B 2 7 and certain microbial agents, and therefore the induetion of A S through molecular mimicry.
Initially it was suggested that there might be another disease susceptibility gene for AS in strong linkage disequilibrium with the HLA-B27 locus (the 'two gene theory'). As it has not been possible to prove unequivocally that the HLA-B27 gene or its produet is directly involved in disease pathogenesis ( lone gene theory') attempts were made to identify a second Polymorphie gene in the HLA-B27 subregion. Several groups used class I specific probes to analyse the configuration of the HLA-B27 gene by genomic Southern blot analyses in order to identify H L A - B 2 7 specific restriction fragment length polymorphisms (RFLP) which might indicate a unique genetic arrangement at the HLA-B27 locus or detect a polymorphism varying in the presence of AS . Using a class I c D N A probe Trapani et a i described a Polymorphie 3.5 kb (kilobases) Taql fragment specific for genomic D N A of HLA-B27 positive individuals [17]. This fragment is present irrespective of the disease Status. Ness and Grumct [18] searched for HLA-B27 specific polymorphisms employing an intragenic probe, derived from intron 7 of the H L A - B 7 gene on Taql digested genomic D N A . The Taql restriction enzyme is prcferentially used in R F L P studies, as it recognizes a four base pair sequence, thus cleaving quitc frequently, which contains a CpG in its recognition sequence. The C in C p G dinucleotides can be methylated and is thus prone to mutations. Using the intron probe four Polymorphie Taql restriction fragments of 2.5, 3.4, 3.8 and 4.0 or 8.0 kb were detected for HLA-B27 . The most common fragment among Caucasians is the 3.4 kb band, as first described by Trapani et cd. [17]. The 3.8 kb band has been found in onc individual and the 4.0/8.0 kb fragment in three unrelated individuals. The 2.5 kb Taql fragment which is characteristic for the H L A - B 7 gene [12], can also be detected in a few H L A -B27 positive normal Caucasians but almost never appears in AS patients, suggesting a trend that is not yet statistically significant. Reveille et a l . reported on a 9.2 kb PvuII fragment which is present in HLA-B27 positive AS patients and less frequently found in HLA-B27 positive healthy individuals [19,20]. The presence of both HLA-B27 and the 9.2 kb PvuII R F L P in an individual conferred a relative risk for AS of 297, compared to 119 with HLA-B27 alone. This polymorphism which can also be found in non-HLA-B27 haplotypes, was detected using either a füll length H L A - B c D N A probe or the shorter H L A - B locus specific fragment, encoding the 3' untranslated region of the H L A - B m R N A . We tested a large panel of HLA-typed unrelated healthy individuals, AS patients and three families with at least one AS patient using the HLA-B27 füll length c D N A clone p B l [5] and the 3' Pstl fragment of p B l as a H L A - B locus specific probe on genomic Southern blot analyses. The majority of the approximately 50 individuals investigated so far possess the 9.2 kb PvuII fragment

WEISS E T A L . . M O L E C U L A R B I O L O G Y O F T H E H L A - B 2 7 L O C U S 15
jr T T i 1 2 3 4 5 6 7 8 I H M • •• n m
10C0 bp HLA-B27
F I G . 3. — T a q l restriction map of H L A - B 2 7 locus.
described by Revcille et cd. The discrepancies between the two studies may be explained by the fact that two different populations were tested. The 9.2 kb PvuII fragment is not derived from the 3' end of the H L A - B 2 7 gene, as this region is contained on a 540 bp (base pair) PvuII fragment characteristic for all H L A - B alleles tested so far.
Steere et cd. [21] used two adjacent D N A probes derived from the 5' flanking region of the H L A -B locus just upstream of the H L A - B 7 gene which cross-hybridize with the H L A - C gene. They identified a 4.0 kb Taql fragment specific for the HLA-B27 gene in AS patients and controls. These results and the intragenic Taql cleavage sites derived from the HLA-B27 gene sequence [4] allowed us to construct a Taql restriction map of the HLA-B27 locus as shown in Fig. 3.
20 40 6 0 80
I I I I
H L A - B
— ; A B C D E
F I G . 4. — Schematic presentation of the H L A - B subregion. The position of the H L A - B gene and the 3' class I half gene are shown as boxes. The orientation of the H L A - B gene 5' to 3' is indicated by the arrow. The scale above the cloned D N A scamenl is given in kilobases (kb). The origins of the probes used for genomic Southern blot analyses are shown. ( A ) 5.0 kb KpnI/Hindll l fragment: (B) 800 bp Sall/Xhol fragment of CD2.6 [4]; ( C ) 2.3 kb Pstl/EcoRI fragment of the 6.8 kb HLA-B27.1 EcoRI subclone; (D) 1.15 kb Taq I fragment: (E) 800 bp Pstl/Xhol fragment.
We extended the cloned D N A of the H L A - B locus isolated in the cosmid CD2.6 containing the HLA-B27.1 gene, by chromosomal Walking experiments and have characterized 90 kb of this region (Fig. 4). A 3' half gene is localized on a 1.6 kb H ind l l l fragment 5' to the H L A - B gene which can be detected in all haplotypes studied so far. Several fragments were derived from the H L A - B subregion which can serve as single or low copy probes. The restriction fragments used in further analyses as H L A - B region probes are shown in Fig. 4. We searched for polymorphisms either specific for the H L A - B 2 7 gene or specific for AS patients. Examples of genomic restriction enzyme digests hybridized with the various probes are shown in Fig. 5. The probes B , D and E did not detect any Polymorphie restriction sites in this region. The fragment B cross-hybridized with four additional bands in the genomic D N A and probe E with one. Polymorphie Variation was observed at the very 5' end of the cloned H L A - B subregion with probe A and in the H L A - B 3' f lanking region with fragment C. The HLA-B27 3' flanking probe was particularly promising because it detects Polymorphie restriction sites with ajmost every enzyme tested, e.g. B g l l l , H i n d l l l , PvuII, Rsa, and Taql. This large number of observed RFLPs in the H L A - B 3' flanking region was not totally unexpected as a high level of divergence was observed at the D N A sequence level. The reason for this great sequence divergence in the H L A - B 3' flanking region is not known. No additional R F L P besides the known 3.5 kb Taql polymorphism [17,18] was observed with the seven restriction enzymes tested which segregated with the H L A - B 2 7 allele. In addition, none of the Polymorphie restriction sites correlated with the disease Status. Further experiments are needed to study this highly Polymorphie D N A segment in more detail.
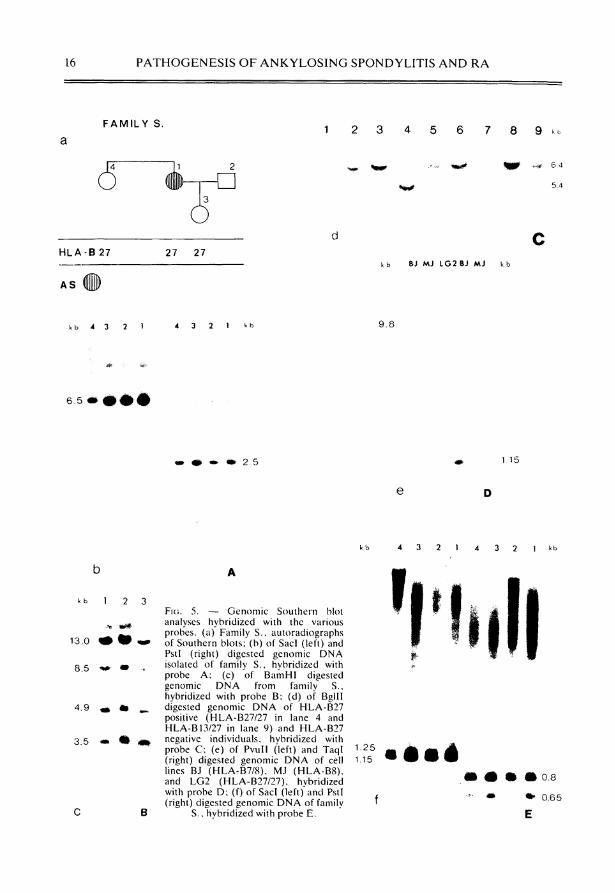
16 P A T H O G E N E S I S O F A N K Y L O S I N G S P O N D Y L I T I S A N D R A
F A M I L Y S. 1 2 3 4 5 6 7 8 9
Ö H L A - B 27 27 27
A S
6 4
5.4
d C
k b B J M J IG2 B J M J k b
k b 4 3 2 1 4 3 2 1 9.8
6.5
2 . 5 1.15
1 2 3
13.0 m ü • w*
8.5 m .
4.9 m m ^
3.5 • • ü
F I G . 5. — Genomic Southern blot analyses hybridized with the various probes. (a) Family S., autoradiographs of Southern blots; (b) of Sacl (left) and Pstl (right) digested genomic D N A isolated of family S., hybridized with probe A ; (c) of BamHI digested genomic D N A from family S., hybridized with probe B : (d) of Bgl l l digested genomic D N A of H L A - B 2 7 positive (HLA-B27/27 in lane 4 and H L A - B 13/27 in lane 9) and H L A - B 2 7 negative individuals, hybridized with probe C ; (e) of PvuII (left) and Taql (right) digested genomic D N A of cell lines BJ (HLA-B7/8 ) , M J ( H L A - B 8 ) . and L G 2 (HLA-B27/27) , hvbridized with probe D ; (f) of Sacl (left)'and Pstl (right) digested genomic D N A of family
S., hybridized with probe E .
1 . 2 5 1.15
• • • 0.8
• *• 0.65
E

WEISS E T A L . : M O L E C U L A R B I O L O G Y O F T H E H L A - B 2 7 L O C U S 17
The tumour necrosis factor (TNF) genes a and ß (lymphotoxin) have recently been localized in the M H C to a region encompassing the complement factor genes and the H L A - B locus [22], These Polypeptide hormones are potent pleiotropins with a wide ränge of bioactivities and a major importance to the pathogenesis of inflammatorv responses. As the T N F genes might constitute the hypothetical gene X , the disease susceptibility gene for A S in strong linkage disequilibrium with the HLA-B27 gene, we isolated the human T N F locus in overlapping cosmid clones. In preliminary experiments we used a human TNF-a probe to investigate the locus in H L A - B 2 7 positive and negative individuals and in A S patients. No difference was found for the three groups tested.
C O N C L U S I O N S
In summary, several hypotheses have been proposed to explain the association of the H L A - B 2 7 allele with spondylarthropathies. The isolation and expression of the gene encoding the HLA-B27 antigen allows investigation of the proposed mechanisms at the single gene level independent of the human genetic background. The H L A - B 2 7 gene itself does not differ between healthy individuals and A S patients. Several unique features of the H L A - B 2 7 molecule have been identified and one epitope was proposed to cross-react with bacterial proteins. The isolated gene will be helpful in a study of the possible receptor function of this class I molecule and for an investigation of the molecular mimicry hypothesis. In an attempt to understand the A S - H L A - B 2 7 association the extended H L A - B region and the T N F genes have also been extensively investigated. Polymorphie restriction sites in this region associated with H L A - B 2 7 and the presence of A S could indicate a disease susceptibility gene linked to H L A - B 2 7 . The D N A located 5' and 3' of the H L A - B gene is Polymorphie. However, no R F L P was found to be significantly increased in frequency in H L A - B 2 7 positive A S patients compared to the H L A - B 2 7 positive control group. The failure to detect polymorphisms which correlate with ankylosing Spondylitis might suggest that genes important in the development of A S are not present in these regions.
A C K N O W L E D G E M K N T S
This work was supported by the Genzentrum, Munich and bv a grant of the Deutsche Forschungsgemeinschaft (SFB 217).
Rl I I R H N O S
1. Schlosstein L , Terasaki PI, Bluestone R, Pearson C M . High association of an H L - A antigen, W27, with ankylosing Spondyl i t i s . .V Engl J Med 1973:288:704-6.
2. Brewerton D A . Caffev M , Hart F D . James D C O , Nicholls A , Slurrock R D . Ankvlosing Spondyli t is a n d H L A 27. Lance! 1973;i:904-7.
3. Tiwari JL , Terasaki PI. Ankvlosing Spondyl i t i s . In H L A und disease associations. Heidelberg: Springer Verlag. 1985:85-100.
4. Weiss E H , Kuon W, Doerner C, Lang M , Riethmueller G . Organization, sequence and expression of the HLA-B27 gene: a molecular approach to analvze H L A a n d disease associations. I m m u n o b i o l o g v 1985:170:367-80.
5. Szoets H , Riethmueller G , Weiss E H . Meo T. Complete sequence o f H L A - B 2 7 c D N A identified through the characterizalion o f stuctural markers unique t o t h e H L A - A . - B . a n d -C allelic series. Proc Natl Acad Sei USA 1986:83:1428-32.
6. Seemann O H A , Rein RS, Brown CS. Plocgh H L . Gene conversion-like mechanisms may generate polymorphism in human class I genes. EMBO J 1986:5:547-52.
7. Coppin H L , McDcvitt, H O . Absence of polymorphism between H L A - B 2 7 genomic exon sequences isolated from normal donors a n d ankylosing S p o n d y l i t i s patients. J h n m u n o l 1986:137:2168-72.
8. Ezquerra A , Bragado R, Vega M A . Strominger J L , Woody J N , Lopez de Castro J A . Primary structure o f papain-solubilized human histocompatibilitv antigen HLÄ-B27. Biochemistry 1985;24:1733-41.
9. Vega M A , Wallace L , Rojo S, Bragado R, Aparicio P, Lopez de Castro J A . Delinealion o f functional sites in HLA-B27 antigens. Molecular analvsis o f H L A - B 2 7 varianl Wewak I defined bv cvtolvtic T lvmphocvtes. J hnmunol 1985:135:3323-32.
10. Vega M A , Ezquerra A , Rojo S, Aparicio P, Bragado R. Lopez de Castro J A . Structural analvsis o f an H L A -B27 functional varianl: identification o f residues that contribute to the speeificity of recognition bv cytolyticTlymphocyles. Proc Natl Acad Sei USA 1985:82:7394-8.
11. Vega M A , Bragado R, ivanyi P, Pelacz J L , Lopez de Castro J A . Molecular analysis o f a functional subtype o f HLA-B27 : a possible evolutionary pathway for H L A - B 2 7 polymorphism. J hnmunol 1986; 137:3557—65.
12. Biro P A , Pan J , Sood A K . Kole R, Reddy V B , Weissman S M . Sequences of human repetitive D N A , n o n -globulin genes, a n d major histocompatibilitv locus genes. III. The major histocompatibilitv complex. Cold

18 P A T H O G E N E S I S O F A N K Y L O S I N G S P O N D Y L I T I S A N D R A
S p r i n g H a r b o r Svtnp Q u a n t B i o } 1983:47:1079-86. (An updated sequence was kindlv provided bv P .A. Biro.j
13. Weiss E H . Unpublished results. 14. Oldsione M B . Schwimmbeck P, DyrbergT, Fujinami R. Mimicry by virus of host molccules: implicalions for
autoimmune disease. In: Progress of Immunology V I . New York: Academic Press. Inc.. 1986:787-95. 15. Ebringer A , Cowling P, Ngwa-Suh N , James D C O , Ebringer RW. Crossreactivity between K l e b s i e l l a
aerogenes species and B27 lvmphocvte antigens. In: Dausset J . Svejgaard A . eds. H L A a n d disease. Paris-I N S E R M , 1976:58:27.
16. Pohla H , Weiss E H . Manuscript in preparation. 17. Trapani J A . Mickelson C A . McKenzie 1FC. A 3.5 kilobase Taql restriction fragment of genomic D N A
segregates with HLA-B27 . hnmunogenetics 1985;24:189-92. 18. Ness D B , Grumei FC. New polymorphisms of H L A - B 2 7 and other B locus antigens detected bv R F L P using a
locus-speeifie probe. H u m a n h n m u n o l 1987;18:65-73. 19. Reveille JD , McDaniel D O , Barger B O , Koopman WJ, Acton RT. Restriction fragment length polymorphism
(RFLP) in ankvlosing Spondylitis (AS): association with a 9.2 kb PvuII f ragment ."Ar thr i t i s Rheum 1986;29:S26.
20. Reveille JD , McDaniel D O , Barger B O , Koopman WJ . Acton RT. Restriction fragment length polvmorphism (RFLP) in the seronegative spondyloarthropalhies (SNSA). A r t h r i t i s Rheum 1986;29:S32.
21. Steerc K, Sidwell B , Leach R, Ward F E . Taurog JD, Orr HT. Use of D N A probes from the 5' flanking region of the H L A - B gene to examine polymorphism at the H L A - B locus. H u m a n h n m u n o l 1986:16:137-^47."
22. Spics T. Morton C C . Nedospasov S A . Fiers W. Pious D . Strominger JL . Genes for the tumor necrosis factors <x and ß are linked to the human major histocompatibilitv complex. P r o c N a t l A c a d Sei USA 1986:83:8699-702.
23. Kottmann A H , Seemann G H A , Guessow H D , Roos M H . D N A sequence of the coding region of the H L A -B44 gene. Immunogenetics 1986:23:396-400.
24. Wavs JP, Coppin H L , Parham P. The complete primarv structure of HLA-Bw58. J B i o l Chem '1985;260:11924-33.
DISCUSSION
Yu: The fragments you are using as probes are perhaps only parts of the genes responsible for the disease susceptibility. The conditions used for hybridization may vary in different centres, which could explain the discrepancies.
Did you use the samc conditions, as the other groups, which have found a difference between A S patients and healthy controls?
Weiss: We used the same conditions and we exchanged D N A probes. so they could confirm our data. Their pattern was identical to ours.
P a n a y i : You have looked at the structure of the region of the B27 gene. Is there any possibility that regulatory genes could be present in this region and explain the difference between AS patients and healthy controls?
For example, do we know if up-regulation of class I antigens on B27 cells from AS patients or controls occurs diffcrently following Stimulation with interferon or 'tumour necrosis factor* (TNF)?
Weiss: We compared the B27 expression with TNF-cv and TNF-ß expression by Northern blot analysis between AS patients and healthy individuals, and we did not find any difference.
However, we did not stimulate the cells to see if there was any differential expression, but these experiments are now in progress.
W i l l i a m s : You showed some sequence data where there was homology with B27.1. but not with B27.2 and B27.3. Could this homology be due to chance?
Weiss: I do a lot of comparisons and always find homologies, both in human and mouse sytems. So I am not sure, that just because one finds a homology, it has to mean something. In the case of the short peptide sequence, if you propose that the six amino acids fragment is the cross-reactive epitope. that can't be the case, because the HLA-B27 Subtypes share a sequence which consists of five amino acids. Furthermore this same five amino acids sequence is also found in other H L A - B antigens such as B7 and B8.