pathogenesis of rashes in - microbiology and molecular ...mmbr.asm.org/content/30/4/739.full.pdf ·...
TRANSCRIPT

BACTERIOLOGICAL REVIEWS, Dec., 1966Copyright © 1966 American Society for Microbiology
Vol. 30, No. 4Printed in U.S.A.
Pathogenesis of Rashes in Virus DiseasesC. A. MIMS
Department of Microbiology, John Curtin School of Medical Research, Australian National University,Canberra, Australia
INTRODUCTION..................................................... 739VIRUS ROUTE TO SKIN-PRIMARY LESION ...................................................... 740
Unbroken-Skin..................................................... 740Local Injury..................................................... 740Biting Vectors..................................................... 741
VIRUS ROUTE TO SKIN-SECONDARY LESION..................................................... 741Vascular Localization of Circulating Inert Particles.............................................. 741Vascular Localization and Diapedesis of Leukocytes ............................................ 742Vascular Localization of Viruses..................................................... 743
GENESIS OF THE LESION.......................................... ............................ 744Primarily Vascular Lesions..................................................... 744Hemorrhagic rash as an unusual complication.................................................. 744Mosquito-borne hemorrhagic fevers ..................................................... 744Mite-and tick- borne hemorrhagic fevers..................................................... 745Rickettsial rashes ..................................................... 745
Extravascular Involvement..................................................... ...... 745Role of the ground substance..................................................... 745Dermal response to infection...................................................... 746Involvement ofepidermis ............... ...................................... 746Role of melanocytes..................................................... 747Differences in susceptibility ofepidermal cells................................................. 748Epidermal response to infection..................................................... 749
Part Played by the Immune Response..................................................... 749Delayed type hypersensitivity..................................................... 749Antibodies..................................................... 751Allergic rashes..................................................... 751Direct damage by virus..................................................... 752Regression oflesions..................................................... 752
FACTORS AFFECTING THE PRODUCTION OF RASHES IN VIRUS DISEASES............................... 753DISCUSSION..................................................... 754SUMMARY..................................................... 755LITERATURE CITED..................................................... 756
INTRODUCTIONObservations on the rashes accompanying in-
fectious diseases date back to ancient times, arash being one of the most characteristic andreadily observed signs of disease. Most of theseexanthemata are seen in the course of virus in-fections. In spite of unprecedented expansion inour knowledge of animal virology many questionsregarding the pathogenesis of rashes in virus dis-eases remain unanswered, and little progress hasbeen made since the work of von Pirquet, Coun-cilman, Mallory, Findlay, and others. Forinstance, there is still no explanation for the basicdifference in distribution of the rashes of chicken-pox and smallpox, and it is not known whethermeasles virus, to produce the rash, grows in theskin, and, if it does, in which cells it multiplies.Advances which have taken place in certain
fields of research during the past 5 years seem to
make a fresh approach to the problem of rashesworthwhile. The rapidly advancing field of im-munology is providing a sounder basis for eval-uating the role of cellular and humoral immuneevents in the production of skin lesions, and re-cent work on the inflammatory response is alsogiving important information. The electronmicroscope reveals the ultrastructural backgroundagainst which events in the skin must be con-sidered, and the fluorescent-antibody method, byidentifying infected cells, makes an analysis ofvirus growth in the skin possible at the histo-logical level.Rashes will be discussed from three aspects:
first, the method by which virus arrives in theskin; second, the sites at which virus growthoccurs; and, third, the mechanism by whichlesions are produced. For the most part, rasheswill be discussed as if the virus concerned did
739
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
localize and grow in the skin, although this maynot be so in prodromal rashes, for instance, or inthe rashes seen in certain arbo- and enterovirusinfections. Because their pathogenesis and patho-physiology are better understood than in manyvirus infections, and because very striking rashesare produced, rickettsial diseases will also beconsidered. Some aspects of the pathogenesis ofvirus-induced skin lesions have already been dis-cussed in Platt's excellent short review (108).
VIRUS ROUTE TO SKIN--PRIMARYLESION
For viruses which give rise to skin lesions bylocalizing and growing in the skin, it is convenientto distinguish the lesions initiated by viruses thatpenetrate the skin from the outside from those inwhich virus reaches the skin from the inside afterspreading through the body of the infected host;in other words, to distinguish between primaryand secondary skin lesions. In much of the experi-mental work to be described, the primary lesionsproduced at the site of the injection of virus intothe skin have been studied. Also, some viruses,such as those responsible for warts and papil-lomas, produce a skin lesion only at the site ofinfection and fail to spread through the body. Aseparate discussion of primary lesions, at leastof the early stages in their pathogenesis, is thusnecessary. Although the skin is usually infectedvia injuries or biting vectors, the possibility ofinfection through unbroken skin will also beconsidered. A later section deals with virus in-fection through the skin in relation to the feedingmethods of blood-sucking arthropods, a subjectof some significance in arthropod-transmittedvirus diseases.
Unbroken SkinThe skin is rightly regarded as protecting the
body, both mechanically and chemically, frominvading microorganisms. When bacteria areplaced on normal human skin, they are rapidlyinactivated, the relative roles of acidity, desicca-tion, fatty acids, or other factors probably dif-fering with different bacteria (116). These anti-bacterial factors might also be expected to helpprevent skin infection with viruses, but the onlyreport which could be found on this topic dealtwith influenza virus (101), and this virus wouldnot be expected to infect the skin.
Results of some experiments have indicatedthat virus particles are able to penetrate the skinof normal animals. Keller (68) reported that,when the tail or abdominal skin of normal micewas atraumatically placed in contact with fluidcontaining 1010 plaque-forming units (PFU) of
Bacillus mnegateriumn phage per milliliter, irregularamounts of phage were recoverable from theblood at 0.5 to 2 hr later. The amounts repre-sented an exceedingly small proportion of thephage present at the skin surface. Nevertheless,if these experiments are valid, the phage particlesnot only penetrated normal skin, but they sub-sequently entered dermal blood vessels. However,normal skin probably cannot accurately be de-scribed as unbroken at the microscopic level, and,in any case, under normal circumstances suchlarge amounts of virus are not likely to be locallyavailable at the skin surface. There have beenoccasional reports of infection with viruses orrickettsiae through apparently unbroken skin, butthese infections have occurred under conditionswhere scratching and rubbing of the inoculationsites have often not been controlled (8). Skin canbe broken without visible injury or bleeding, andparticles can thus be placed in contact withliving cells.
Virus infection through truly unbroken skin,therefore, probably never occurs, and small ormoderate amounts of virus are not likely to in-fect unless the skin is scratched or otherwisedamaged. This is the method of human infectionin accidental smallpox vaccination, and in infec-tion with warts, cowpox, milker's nodes, louse-borne typhus rickettsiae, or with the etiologicalagent of cat-scratch disease.
Local InjuryAs discussed above, most instances of infection
through apparently unbroken skin probably in-volve minute local injuries. Deliberate local in-jury will be considered as a method of infection,first, because it is the classical method of vac-cination against smallpox, and, second, becauseit is the method used in much of the experimentalwork on the primary lesion produced by viruses.
In the multiple pressure method of smallpoxvaccination, it seems necessary only to split openthe epidermis and allow virus to encounterepidermal or dermal cells. An increasing amountof evidence suggests that many viruses infect cellsby being phagocytosed (20), often after an initialspecific attachment to receptors on the cell sur-face. Put another way, cells infect themselves.Epidermal cells, for instance, are able to ingestparticles of carbon introduced into the skin of theguinea pigs' foot (107). Roberts (114), using thefluorescent-antibody technique, showed thatepidermal cells were infected in the first cycle ofgrowth after the controlled intradermal inocula-tion of ectromelia virus into the skin of mice.This certainly implies an initial uptake of virusby epidermal cells.
740 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
Inevitably, viruses scratched or injected intothe skin enter dermal lymphatics. This is particu-larly true for intradermal injections, and, insaying that "every intradermal injection is anintralymphatic one," Hudack and McMaster (60)drew attention to the very rich superficial plexusof lymphatics in the skin of man, as well as of themouse (59). When large amounts of ectromeliavirus are injected intradermally and subcutane-ously into mice, virus is detectable in local lymphnodes within a few minutes (Mims, unpublisheddata). The lymphatic system, although extensive,does not deal with all the injected particles in thisway, and the rest have the opportunity to infectdermal cells. If the infecting virus grows in thesecells, its spread in the dermis is almost assured.If it fails to grow, it may be restricted to the initialsite of growth in epidermal or other cells, as isprobably the case in wart or papilloma virus in-fections. Roberts (114), in his immunofluores-cence study of ectromelia virus infection in mouseskin, found that, after scarification of virus intothe dermis, the first infected dermal cells ap-peared to be macrophages, and the focus thenspread through the dermis by means of the in-fection of neighboring cells.
Biting Vectors
When biting vectors transmit virus diseasesmechanically, there being no multiplication ofvirus in the vector, the contaminated mouthpartsintroduce virus into epidermis and dermis, asdiscussed in the preceding sections. This is trueof the transmission of swinepox by the pig louse(123), for instance, and the best studied exampleis the transmission of myxoma virus by mos-quitoes (38).When transmission is said to be biological,
this means that the virus multiplies in the vectorand, after an incubation period, is transmitted bythe saliva during a blood feed. A discussion ofthe process of infection must take into accountthe mechanics of blood feeding. Mosquitoes, inthe act of feeding (49), probe in dermal tissues,emitting "puffs" of saliva as they do so. If theproboscis enters a blood capillary, it is "threaded"along the vessel, and further ejections of salivatake place during the ingestion of blood. Theinfected saliva is thus introduced into dermaltissues, and often directly into the vascular sys-tem. Ticks are exclusively "pool feeders";i.e., they pierce the skin with their cutting mouth-parts, suck up the pool of blood which accumu-lates, and periodically eject saliva into the woundthroughout the feed (72). Virus infection byticks is, therefore, the counterpart of a minuteintradermal injection.
Thus, the mechanically transmitted pox-viruses encounter epidermal and dermal cells,and it is in these cells that they grow. Rickettsia,too, probably grow in these cells, and strikinglocal lesions or eschars are often produced.These viruses and rickettsiae also have the op-portunity to spread systematically, because theyenter dermal lymphatics. The biologically trans-mitted or arboviruses, however, do not give riseto local lesions, in spite of the fact that infectedsaliva is introduced into the dermis. Conceivably,they do not grow in dermal or epidermal cells,and infection takes place only after virus par-ticles have entered the lymphatic or vascular sys-tem.
VIRUS ROUTE TO SKIN-SECONDARY LESIONSecondary skin lesions occur in the course of
generalized virus diseases and will be consideredas due to the localization and growth of virusin the skin. As will be discussed at a later stage,rashes can quite probably arise by allergicmechanisms without the localization and growthof virus in the skin. It is because even less isknown about such rashes that discussion willbe restricted in the first place to lesions producedby the growth of virus in the skin. With rareexceptions, virus reaches the skin via the blood,during a viremic stage of the disease.
Viremias can be classified into those inwhich virus is cell-associated, and those in whichvirus is free in the plasma (91), although viremiassometimes are of mixed character. The essentialprimary event for the production of a skin lesionis for free virus or an infected leukocyte to lo-calize in small blood vessels in the skin, and inmany generalized virus diseases, such as varicella(135), alastrim (83), and myxomatosis (64), avascular lesion is the earliest event in the de-velopment of a skin lesion. To understand thisvascular localization of virus in skin, one mustfirst discuss the behavior of inert virus-sizedparticles when they are introduced into the blood,and also the evidence for the passage of leuko-cytes through the skin blood vessels of normalanimals.
Vascular Localization of Circulating InertParticles
In certain virus diseases with rashes, such asthose produced by entero- and arboviruses,virus is free in the plasma. Foreign particles,viral or nonviral, are cleared from the blood byreticuloendothelial cells, particularly those liningthe liver and spleen sinusoids, and larger par-ticles tend to be removed more rapidly thansmaller ones (91). Large particles, indeed, would
741VOL. 30, 1966
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
be cleared by reticuloendothelial cells beforethere was much opportunity for localizationelsewhere, unless capillary reactivity was al-tered. Small particles, on the other hand, with alonger cirulatory half life, could more easilylocalize in small skin blood vessels.Do particles introduced into the blood, in
fact, localize in skin vessels? Convincing evidencefor localization is usually obtained with the elec-tron microscope, and particles in blood vesselwalls are likely to be seen in sections only whenthey are very numerous. Thus, in most experi-ments, inflammatory changes have been inducedin vessels, so that particles would localize inthem on a grand scale. After histamine or seroto-nin treatment of rat scrotal tissues, for instance,intravenously injected colloidal mercuric sulfidewas detectable between the endothelial cells ofcapillaries and postcapillary venules (84). Par-ticles appeared to be held up by the basementmembrane but later were seen in the cytoplasmof pericapillary phagocytic cells. It was noted thatan occasional particle was seen in vessel wallsin untreated animals. Alksne (1) made similarobservations on dermal capillaries in the skinof the mouse, and he also saw occasional par-ticles in capillary walls in untreated mice, in areaswhere the hair had been clipped just before themercuric sulfide injection. One weakness in suchevidence is that very large numbers of particleswere suddenly introduced into the blood, whereasin a virus infection there would be a more gradualentry of particles into the blood. Localizationin skin or other blood vessels might then be lesscommon. There is also the theoretical possibilitythat the large injections of colloidal particlescould lead to local histamine-mediated changes inblood vessels, as in the anaphylactoid response.Apart from these observations, there is very
little direct information about the vascular local-ization of circulating virus-sized particles in theskin of normal animals. However, it can reason-ably be assumed that minute injuries or infectionsoccur regularly in the apparently normal skin ofanimals. Small-scale inflammatory changes mightthen take place, and, however transient thesechanges, circulating virus particles could localizein small blood vessels. There are certain otherphysiological circumstances under which par-ticles localize in skin vessels. Capillaries whichare regenerating after injury have been shownto have temporary gaps between endothelialcells, and deficiencies in the basement membrane,so that circulating inert particles localize in, andleak from, these vessels (119). During hairgrowth cycles there are changes in the capillarynetworks supplying hair follicles, both in man(34), and in animals such as the mouse (142).
The actively growing capillaries around follicleswhich are resuming activity might then differfrom normal capillaries in their reaction to cir-culating particles. Finally, the microcirculatorychanges in the skin which are known to takeplace after changes in environmental temperaturemight affect the localization of blood-borneparticles, and perhaps even the vasodilation ofreactive hyperemia could act in the same way.There is also some important indirect evidence
indicating that circulating particles might lo-calize in the skin vessels of normal animals.Grotte (52) injected different molecular speciesof dextran intravenously into dogs, and deter-mined the concentration in lymph from the leg.Dextran of molecular weight 300,000 was foundin the lymph, and Grotte concluded that thesemolecules, of effective diffusion diameter 225 A,leaked through capillary walls to enter lympha-tics. Courtice and Garlick (19) estimated concen-trations of different lipoproteins in the serumand in the lymph from the foot of hypercholes-terolemic rabbits. The particle size of the lipo-proteins was determined by electron microscopy,and the authors concluded that smaller particlespassed through capillary walls more rapidlythan larger ones. Even particles 600 to 700 A indiameter were readily recoverable in lymph.Although the rabbits were anesthetized andtheir legs were immersed in warm water to pro-mote lymph flow, these experiments could meanthat particles the size of small viruses leak throughcapillary walls under normal circumstances.Such events might not be common enough to bedetected in sections by electron microscopy. Thesite of leakage in the foot is not known, but is islikely to be in the skin.
Vascular Localization and Diapedesis ofLeukocytes
In the viremias of myxomatosis, mousepox,measles, rinderpest, and distemper, for instance,virus is associated with leukocytes (91). Platelets,lymphocytes, monocytes, or polymorphs maybe infected. Healthy cells which bear virus par-ticles, or are at an early stage of infection withleukocyte-pathogenic viruses, would be expectedto move through small blood vessels like normalleukocytes, and could thus carry infection throughto extravascular tissues. The ultrastructuraldetails of this diapedesis have now been carefuUystudied (86). Platelets would not be expected topass through blood vessel walls, but the adherenceof platelets to vascular endothelium, an earlyresponse to mild damage or inflammation, couldenable platelet-borne viruses to infect vascularendothelium.
742 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
If, as discussed earlier, small-scale inflamma-tory events occur in apparently normal skin,these may lead to leukocyte localization anddiapedesis. There is direct evidence that leuko-cytes leave blood vessels in the skin in normalanimals. Yoffey and Drinker (160) reported thatsmall numbers of leukocytes were present inafferent lymph from the limbs of normal catsand dogs. In these experiments, the animals wereanesthetized and the limbs were massaged topromote lymph flow. In more physiologicalstudies, Hall and Morris (60) cannulated afferentlymphatics in the legs of sheep, and found thatafferent lymph contained up to 3,000 leukocytesper mm3, about 80% of these being maturelymphocytes. The sheep were maintained inpens and the cannulas remained in position formany weeks. It must be assumed from theseexperiments that leukocytes leave blood vesselsin the skin and perhaps the muscles of normalanimals. This is also in accord with currentbeliefs about the wanderings and recirculationof lymphocytes.
It is now necessary to see to what extent theseconsiderations about free particles and leukocytesapply to the vascular localization of circulatingviruses.
Vascular Localization of Viruses
As discussed above, there is some experimentalprecedent for the localization of free or cell-associated viruses in skin blood vessels. Thereare numerous testimonies to the localization ofvirus lesions in provoked skin areas, and aninflammatory response, or an increased vascular-ity, can usually be invoked as the cause of thislocalization. Much of this evidence was reviewed,in relation to the "stickiness" and permeabilityof capillary endothelium, by Findlay (39). Ex-perimentally, it has been usual to record the de-velopment of lesions in treated skin areas afterintravenous injection of large doses of virus.For instance, fowlpox lesions localize at the sitewhere feathers have been pulled out (13), andvaccinia lesions where the skin of rabbits hasbeen plucked, shaved, scarified, treated withhistamine, or even where vasodilation has beenproduced by sympathetic nerve section (15, 16,39, 140).Some viruses localize in the unprovoked skin
of apparently normal animals after intravenousinjection. When large doses of neurovacciniawere injected intravenously into rabbits, lesionswere seen 4 days later on the untreated skin of thelips and nose, and also on the tongue and on thegeneral body surface (26, 76). Downie (27)injected cowpox virus intravenously into normal
rabbits and recorded skin lesions on the lips,eyelids, ears, and tongue. Platt (109) injectedherpes simplex virus intravenously into normalmale guinea pigs and observed a blotchy rashon the perineum 2 to 4 days later. When cowpoxvirus is injected intravenously in large doses intomice, nearly all of the virus is taken up by reticu-loendothelial cells, but it also localizes and growsin the skin of the tail, ears, snout, anogenitalregions, paws, and tongue (93). Fluorescent-antibody studies have revealed that the injectedvirus localizes in small dermal blood vesselsand then grows in dermal and epidermal cells.The skin becomes bright red on the 3rd or 4thday, so that a confluent rash is formed on thehairless areas. It is not known whether free orcell-associated virus localizes in the skin in theseinstances, but free particles would have to localizein spite of rapid clearance from the blood byreticuloendothelial cells.
In the secondary skin lesions which localizein irritated areas in the course of exanthematousvirus diseases, infected leukocytes may be con-cerned rather than free particles, although freevirus may also be present in the blood. Varicellacan localize under strips of adhesive tape ornapkins, and in areas affected by acne (112), orsecondary syphilis (132). The rash in measlesmay be more profuse in "mottled" skin areas(77), at the site of intradermally injected tuber-culin (118), or in other irritated areas (137).Ricketts (111) drew attention to the localizationof smallpox lesions in irritated skin areas, andFenner (37) localized mousepox skin lesions inshaved areas. Some of the experimental findingson the localization of foot-and-mouth diseasevirus may also involve local vascular changes(108), but it seems probable that genuine dif-ferences in the reactivity of the epidermis aresometimes equally important. Also, the abilityof an already localized virus to infect dermaland epidermal tissues and produce a macro-scopic lesion may well depend on nonvascularregional differences in the skin.
In summary, skin lesions may appear in irri-tated skin areas, but lesions also arise in appar-ently unprovoked skin, both experimentally andin the course of exanthematous virus diseases.Either each lesion arises in a subclinically in-flamed area, or there is localization in normalblood vessels. Most of the evidence concernsviruses which are associated with leukocytesduring their viremic stage. The possibility thatviruses in the plasma, whether exanthematous ornot, localize in skin vessels, will be discussed morefully later.
VOL. 30, 1966 743
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
GENESIS OF THE LESION
The essential first act in the genesis of theskin lesion, the localization of virus in bloodvessel walls, has been discussed. The skin lesionmay then develop as a primarily vascular one or,alternatively, as a result of the spread of infectioninto other tissues in the skin (see Fig. 1). Broadlyspeaking, a lasting local dilation of subpapillarydermal blood vessels produces a macule. If thereis also edema and an infiltration of cells into thearea, the macule becomes a papule. There maybe secondary changes in the epidermis leadingto desquamation or changes in pigmentation,but a primary involvement of the epidermisusually results in vesiculation, ulceration, andscabbing.
Primary Vascular Lesions
Viruses growing in cells of the blood vesselwall could damage these cells directly, and thusinitiate an inflammatory response. There isdirect evidence that vascular endothelium is in-fected in certain virus diseases, because specificinclusion bodies (73, 124, 135), or viral antigen(18, 57, 165) have been observed in the endo-thelial cells of small blood vessels. At a laterstage, antigen-antibody reactions could contributeto the pathological changes (see below) Theremight be edema, vasodilation, and cellular infil-tration, giving rise to a visible skin lesion, oreven thrombosis and hemorrhage if damage tothe vessel was severe.
In some instances, the infection may not pro-ceed far beyond the blood vessel wall. Indeed,the infectious process would be automaticallyrestricted to the vessel wall if there were no in-fected emigrating leukocytes, and if at the sametime neither dermal cells nor perivascular phago-cytic cells could be infected.
Primarily vascular lesions are seen in a numberof different virus diseases. For instance, there is aprimarily vascular lesion in equine viral arteritis,leading to medial necrosis of smaller arteries (66).In African swine fever (wart-hog disease) andhog cholera, distinctive changes in the dermalblood vessels provide the basis for the observedskin lesions (89, 124), and the skin lesions insheeppox have been attributed largely to theeffects of vascular thrombosis (110).Hemorrhagic rash as an unusual complication.
Hemorrhagic manifestations, including petechialrashes, are seen occasionally in a number of virusdiseases, including varicella (87), variola, andmeasles (61). Almost nothing is known of thepathogenesis of these hemorrhagic rashes, exceptthat an altered host reactivity must be important.The very rare instances of measles complicated
by purpura appear to be thrombocytopenic inorigin. Hemorrhagic chickenpox is seen par-ticularly in tropical countries, where differencesin nutrition and the presence of parasites andother infections may play a part. It may be rele-vant that hemorrhage can be induced aroundestablished vaccinia lesions in the skin of rabbitsby the intravenous injection of endotoxin (50,71).
Mosquito-borne hemorrhagic Je'vers. Hemor-rhagic manifestations and petechial rashes areseen more regularly in certain arthropod-trans-mitted virus diseases. These were classified byGajdusek (45), who distinguished betweenthose transmitted by mites or ticks and thosetransmitted by mosquitoes.The mosquito-borne fevers with hemorrhagic
manifestations include dengue and yellowfever, and in many outbreaks of these two dis-eases hemorrhages from small blood vesselshave been reported. In hemorrhagic yellow feverit is difficult to assess the importance of primaryvascular damage, because the liver lesions leadto deficiencies in prothrombin and other coagu-lation factors (90, 133), and this contributes tothe production of hemorrhages. Skin hemor-rhages are occasionally seen. In hemorrhagicdengue, there are also coagulation defects andoften thrombocytopenia (98), but here vasculardamage is of primary importance. The outbreaksof hemorrhagic dengue in Thailand have beencarefully studied from a physiopathologicalstandpoint (54), and it is evident that there is aprimary injury to small blood vessels. This leadsto increased capillary fragility, as indicated by apositive tourniquet test (46), and a leakage ofplasma from the blood, with hemoconcentrationand possible shock. Hemorrhages are seen invarious parts of the body, and there is often apetechial rash.Maculopapular rashes have also been reported
in dengue outbreaks. Sabin (117) infected volun-teers with dengue virus, and biopsies of the rashshowed an endothelial swelling of small bloodvessels, perivascular edema, and a mononuclearinfiltration. It is clear that in dengue small bloodvessels in the skin are damaged, and this maygive rise to a maculopapular, or even petechialrash, the tendency to hemorrhage perhaps beingincreased when there is a coagulation defect orthrombocytopenia. What is not clear, is whetherthe vessel damage is due to a direct destructiveeffect of the virus. Sabin could not inhibit therash locally with intradermally injected immuneserum, and, curiously enough, the rash sparedthe site of initial infection in the skin, whetherthis had been by needle or by mosquito bite.
744 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
It seems unlikely that circulating toxins cause theblood vessel damage, but mechanisms involvingthe immune response are easier to visualize(see below). Dengue virus growing in capillaryendothelium may do very little direct damage,but, if antigen is present there at the time thatfree antibody appears in the serum, there wouldbe an antigen-antibody reaction with pathologi-cal consequences in the vessel walls. Little isknown about- the pathogenesis of hemorrhagicdengue, and the hemorrhagic manifestations arenot usually seen in occidental patients, but thepossibility of a hyperergic host response is promi-nent in much of the current thinking on the sub-ject (54).
Mite- and tick-borne hemorrhagic fevers. Someof the hemorrhagic mite- and tick-borne fevershave also been investigated from the physio-pathological point of view. Here, too, damageto small blood vessels is one of the primarypathological changes, resulting in capillary leak-age, increased capillary fragility, and hemor-rhages (74). Patients may have lasting flushes ormild rashes, as well as petechial rashes (33).Greisman (51) made direct observations on smallblood vessels in the nail-fold bed, and describedprolonged dilatation and hemorrhage in patientswith Korean hemorrhagic fever. As is the casewith mosquito-borne hemorrhagic fevers, weare ignorant about the relative importance ofdirect viral damage, toxins, and immune reactionin the pathogenesis of the vascular lesions.
Rickettsial rashes. Rickettsia can be observeddirectly in vascular endothelium in the lesionsof Rocky Mountain spotted fever (143), typhus,and scrub typhus (144, 145), and very strikingrashes are seen in these diseases. When rickettsiaeare injected intravenously in large doses to miceor rats, they produce widespread capillary endo-thelial damage, and animals die in shock as aresult of the leakage of plasma through capi-laries (138). Death occurs within hours, and thereare no gross pathological changes in vessels.Infectious particles, not toxins, are responsiblefor the effect. Rickettsiae, clearly, have a pri-mary pathological action on vascular endothe-lium. Equally clearly, when patients with rick-ettsiae in blood vessel walls produce circulatingantibodies, antigen-antibody reactions will tendto damage these vessel walls. This could accountfor the vascular lesions, for there need be noclassical Arthus reaction. Allen and Spitz (2)suggested that the hyperergic state was importantin the pathogenesis of the vascular lesion inscrub typhus and other rickettsial diseases. Intra-epidermal vesicles are seen in the skin lesions ofone rickettsial disease, and these are sufficientlylike chickenpox (25, 62) to have led to the dis-
tinctive name of rickettsialpox. But it is notknown whether rickettsiae grow in the epidermisto produce these epidermal lesions, which arealso described in the early development of thescrub typhus eschar (2).
Thus, in most rickettsial diseases, the rashappears to be produced by rickettsiae growing inblood vessel walls, and then reacting with cir-culating antibodies and perhaps with immunecells. The evidence does not warrant conclusionsabout the relative importance of direct damage byrickettsiae, and damage mediated by the im-mune response.
Extravascular InvolvementThe infectious process may spread to extra-
vascular tissues, either after the infection ofperivascular cells and release of virus into sur-rounding tissues, or by way of migrating leuko-cytes. Unfortunately, although the possibilitiescan be discussed in this way, it is not often pos-sible to say exactly what happens in any par-ticular virus disease. In any case, if dermal fibro-blasts or macrophages become infected, aspread of the infection through the dermis ispossible. At this stage, the quality of the groundsubstance of the dermal connective tissue mightplay an important role.
Role of the ground substance. Do viruses spreadextracellularly in the skin? The ground substanceis viscous and normally offers considerableresistance to the passage of foreign material(81). However, when very large numbers ofcolloidal gold particles were injected subcutane-ously into mice, electron microscopic examina-tion showed that at least some particles spreadsignificantly through the dermis (47). But stand-ard injection techniques, in which comparativelylarge volumes are introduced under pressure,are grossly unphysiological compared with thenormal process of infection through skin, andeven more so compared with the evolution ofhematogenous lesions in the skin. The changesin the ground substance that occur under somecircumstances increase the difficulty with whichinert particles spread in the dermis. At the sametime, smaller skin lesions are produced by cer-tain viruses, and with greater difficulty (127).During pregnancy, for instance, rabbits developsmaller skin lesions at the site of inoculation ofmyxoma virus (126). There might be increasedopportunity for the spread of free particles inedematous tissues, through which dyes are knownto move more rapidly (67, 80). Hyaluronidase,the spreading factor, has been shown to havepronounced effects on the movement of par-ticles within the skin, and on the production ofskin lesions by poxviruses (31). Although the
745VOL. 30, 1966
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
physiological role of hyaluronidase in tissueslike the skin is still unsettled, the fact that it hassuch effects when injected locally shows that theconsistency of the ground substance can be im-portant in the development of virus skin lesions.
Thus, the ability of free particles to spreadthrough dermal connective tissue, although prob-ably lirnited, can be influenced by certain physio-logical changes. To what extent would this beimportant during the genesis of virus skin lesions?It would seem that virus particles released locallyand in small numbers are likely to be taken upat an early stage by phagocytic cells. Later on inthe evolution of a lesion, if there was edema, or alarger scale release of virus particles, there mightbe an opportunity for significant spread of freeparticles. But the spread of virus extracellularlyis probably unimportant compared with spreadby the movement of infected cells or by the infec-tion of successive cells, either of which could inturn be influenced by changes in the groundsubstance.Dermal response to infection. In the dermis,
vasodilation, edema, and cellular infiltrationcombine to produce pathological changes whichare not often very characteristic for differentviruses. The type of cellular infiltration may de-pend on the relative importance of nonspecificinflammatory and delayed type hypersensitivityresponses. There are also likely to be changes inthe quality of the ground substance.Many features of the dermal response are
discussed elsewhere, but the characteristic lesionscaused by myxoma virus and the Yaba poxviruswill be mentioned here. In rabbits infected withmyxoma virus, the skin lesions contain largeamounts of muci-carmine staining extracellularfluid. This mucinous or myxomatous fluid appearsto be produced by fibroblasts, some of whichtransform into myxoma cells (64). The closelyrelated fibroma virus, while it may lead to theproduction of a similar material in newbornrabbits (32), induces the formation of a moresolidly cellular tumor in the dermis of adults.Fibroblasts are known to be capable of formingmucin, and they do so regularly in young em-bryos, as well as in certain rare mucinous tumors.The fact that a virus infection may specificallyinduce this change in the secretory activity offibroblasts is of some interest, but such aspects ofthe infection have not been investigated experi-mentally. The Yaba poxvirus is mentioned be-cause it induces an unusual response in the der-mal histiocytes of monkeys. These cells, whichare infected, multiply to produce "histiocyto-mas," and, if India ink is injected with the virus,the dermal nodules are seen to consist of ink-laden cells (125).
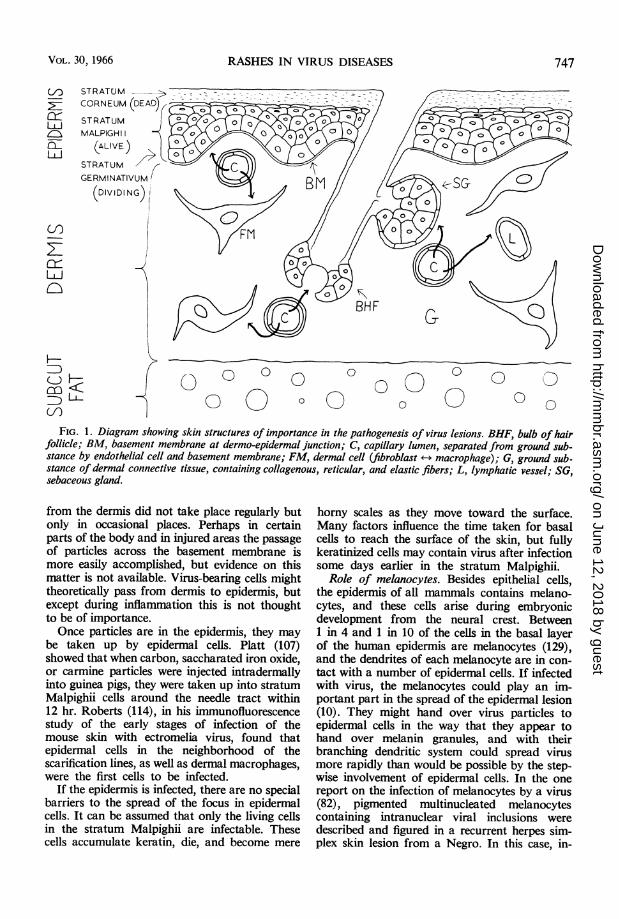
Involvement of epidermis. Once infection hasspread into dermal connective tissue, the surfaceepidermis, hair follicles, and sebaceous glandsmay in turn be infected (Fig. 1). Although theentire body surface is covered with a sheet of epi-dermis, in nearly all mammals most of the epider-mis is in fact below the body surface, for it isembedded in the dermis in the form of hairfollicles and glands. Hair follicles and sebaceousglands, indeed, are selectively affected in certainvirus diseases, the severe scarring followingattacks of variola major, for instance, being aresult of the destruction of deep-seated sebaceousglands (12).A submicroscopic (350 A) membrane sheet
separates basal epidermal cells from the dermis(121). This membrane is thinner and differentin appearance from basement membranes else-where, and, although metabolites must passthrough it to nourish the epidermis, perhaps byway of the vesicles described by Odland (100),it might be expected to act as a barrier to thespread of free particles into the epidermis.Platt (107) found that iron dextran introducedbelow the epidermis of guinea pigs did not passthrough into epidermal cells, except in injuredareas and in certain parts of the body. The lattercomprised the tip of the tongue, ears, feet, belly,and prepuce, and the possibility that in the nor-mal animal minute undetectable traumata werealways occurring at these sites was discussed.He drew attention to differences in the phagocy-tic activity of epidermal cells, but pointed outthat differences in the integrity of the basementmembrane, and thus in the passage of particlesacross it, may have played a part. Selby's mate-rial (121) included skin from the footpad of rats,but there have been no studies of regional dif-ferences in completeness of the membraneat the dermo-epidermal junction. Gerard andTyler (47) found that when colloidal gold wasinjected subcutaneously into mice some of theparticles spread into the dermis, and some cameto lie adjacent to the basement membrane. Butthey generally failed to pass into the epidermis,except for occasional particles which were seenbetween epidermal cells.
It can be concluded that free particles in thedermis, although capable of passing through thebasement membrane and thus encountering epi-dermal cells, do so uncommonly. Thus, in Rob-erts' (114) careful immunofluorescence study ofthe infection of the mouse skin with ectromeliavirus, the primary dermal lesion was shown tospread laterally through the dermis and occasion-ally to reinfect the epidermis to produce "islandfoci" which were separate from the primary epi-dermal focus. The reinfection of the epidermis
746 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from
RASHES IN VIRUS DISEASES
()
C 00 0 0O0 o0
o () (
FIG. 1. Diagram showing skin structures of importance in the pathogenesis of virus lesions. BHF, bulb of hairfollicle; BM, basement membrane at dermo-epidermal junction; C, capillary lumen, separated from ground sub-stance by endothelial cell and basement membrane; FM, dermal cell (fibroblast +-+ macrophage); G, ground sub-
stance of dermal connective tissue, containing collagenous, reticular, and elastic fibers; L, lymphatic vessel; SG,sebaceous gland.
from the dermis did not take place regularly butonly in occasional places. Perhaps in certainparts of the body and in injured areas the passageof particles across the basement membrane ismore easily accomplished, but evidence on thismatter is not available. Virus-bearing cells mighttheoretically pass from dermis to epidermis, butexcept during inflammation this is not thoughtto be of importance.Once particles are in the epidermis, they may
be taken up by epidermal cells. Platt (107)showed that when carbon, saccharated iron oxide,or carmine particles were injected intradermallyinto guinea pigs, they were taken up into stratumMalpighii cells around the needle tract within12 hr. Roberts (114), in his immunofluorescencestudy of the early stages of infection of themouse skin with ectromelia virus, found thatepidermal cells in the neighborhood of thescarification lines, as well as dermal macrophages,were the first cells to be infected.
If the epidermis is infected, there are no specialbarriers to the spread of the focus in epidermalcells. It can be assumed that only the living cellsin the stratum Malpighii are infectable. Thesecells accumulate keratin, die, and become mere
horny scales as they move toward the surface.Many factors influence the time taken for basalcells to reach the surface of the skin, but fullykeratinized cells may contain virus after infectionsome days earlier in the stratum Malpighii.
Role of melanocytes. Besides epithelial cells,the epidermis of all mammals contains melano-cytes, and these cells arise during embryonicdevelopment from the neural crest. Between1 in 4 and 1 in 10 of the cells in the basal layerof the human epidermis are melanocytes (129),and the dendrites of each melanocyte are in con-tact with a number of epidermal cells. If infectedwith virus, the melanocytes could play an im-portant part in the spread of the epidermal lesion(10). They might hand over virus particles toepidermal cells in the way that they appear tohand over melanin granules, and with theirbranching dendritic system could spread virusmore rapidly than would be possible by the step-wise involvement of epidermal cells. In the onereport on the infection of melanocytes by a virus(82), pigmented multinucleated melanocytescontaining intranuclear viral inclusions weredescribed and figured in a recurrent herpes sim-plex skin lesion from a Negro. In this case, in-
(./) S-
2E C(
LL
wn
s-G
(I)
zu-JCL
VOL. 30, 1966 747
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
clusion bodies were "confined almost entirelyto the pigment carrying cells," and this raisesthe interesting possibility that melanocytes playan important part in recurrent herpes and per-haps even in zoster. The cornea, a site for re-current herpes, also appears to contain melano-cytes (Langerhans' cells) which impart a slightpigmentation to the cornea in some animals.The only experimental evidence concerning therole of melanocytes in the development of virusskin lesions comes from the work of Roberts(114). He examined fluorescent antibody-stained epidermal strips from developing ectro-melia skin lesions in mice, and could find noevidence that melanocytes facilitated the spreadof infection. He was unable to determine whetherthe melanocytes were, in fact, infected.
Melanocytes, therefore, because there are manyof them in the skin of all mammals, could theo-retically play an important part in the develop-ment of epidermal lesions. In one instance wheretheir role was investigated, they did not appearto be important, but information is scarce, andusually it is not even possible to say whether theyare infected by viruses growing in the skin.
Differences in susceptibility of epidermal cells.Although there are no special barriers to thespread of infection in the epidermis, there is thepossibility that the epidermal cells themselves,in certain areas or under certain circumstances,are more susceptible to infection. Thus, moreactively mitotic epidermal regions, epidermalcells round hair follicles during the active stagesof hair growth, or epidermal cells in exposedregions like the mouth, paws, prepuce, or udders,might be more readily infected. Where lesionsappear in certain areas of skin in generalizedinfections, a difference in the ability of circulatingvirus to localize in skin blood vessels must alsobe considered. It is difficult, as Platt (108) haspointed out, to decide whether there are alsolocal differences in the susceptibility of epidermalcells. For instance, it might be simply the vascu-lar localization of virus which leads to the initia-tion of smallpox lesions in sebaceous glands andhair follicles, just as vascular localization ac-counts for the appearance of vaccinial lesionsin the shaved skin areas of intravenously injectedrabbits. Genuine differences in the behavior ofepidermal cells are therefore more readily re-vealed when primary lesions are studied.
Platt (106) showed that when foot-and-mouthvirus was injected into the skin of guinea pigsthere were marked regional differences in sus-ceptibility. The metatarsal pads were highlysusceptible, the ear less so, and the trunk almostinsusceptible. Skin from metatarsal pads, how-
ever, when grafted onto the chest, was much lesssusceptible to directly inoculated virus. Thechange in susceptibility was not regularly relatedto alterations in mitotic activity in the graftedskin, and changes in skin temperature were notthought to be important. Platt suggested thatthere was a change in the susceptibility of theepithelial cells themselves.Kidd and Parsons (70) reported that local in-
jections of Shope papilloma virus producedlesions in the skin, but not in the mucous mem-brane inside the mouth or urogenital tract ofdomestic rabbits. In this case, lesions are pro-duced by the proliferative response of epidermalcells, and, although the findings are of consider-able interest, they do not lead to any firm con-clusions about the development of the infectiousfocus in epidermal cells. Friedewald (42) foundthat, when the skin of rabbits was made hyper-plastic by treatment with certain chemicals,it became more susceptible to intradermallyinjected or scarified papilloma virus. Whiteley(140) showed that the same virus, inoculated byscarification into the skin of rabbits, producedmuch larger tumors in skin with actively multi-plying epidermis, 5 and 10 days after plucking.Here again, it is only possible to say that tumorsare more readily induced, and once establishedgrow faster, in thick, actively mitotic epidermis.In a similar experiment with vaccinia virus, therewas no great difference between the lesions pro-duced in active and in quiescent skin (140).
Finally, the growth of viruses in the skin canalmost certainly be directly affected by tempera-ture. There are various observations suggestingthat this occurs, but where, in generalized virusinfections, skin lesions are reduced in cooledextremities, the results may be attributable topoor vascular localization in cool skin. Moredirect evidence is available from the work ofRoberts (115). He injected ectromelia virus intothe footpads of mice kept in the cold and at nor-mal temperatures, and compared virus growthcurves in the inoculated feet. From the start,less virus was produced in the feet of mice keptin the cold. When virus was injected into thehairy and therefore warmer skin of the back,there was no difference in skin titers at the siteof injection. Although the results might have beendue to changes in the susceptibility of cells as aresult of reduced blood flow, the probable ex-planation is that in the feet of mice in the coldthe cells, particularly those in the epidermis,were at a suboptimal temperature for virus growth(115). In an experiment with an attenuatedstrain of myxoma virus, Roberts (personalcommunication) made parallel titrations intra-
748 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
dermally on the freshly clipped and on the nor-mal furred sides of a rabbit's back. End points,as judged by the appearance of a lesion, were100-fold higher on the clipped side. This recallsthe work of Thompson (131), who showedthat rabbits in the warm room, with skin tem-peratures several degrees higher than controlrabbits, developed very much smaller tumorsafter the intradermal inijection of fibroma virus.Rabbits in the warm room were also less likelyto develop skin lesions when injected intrader-mally with myxoma virus (102). The most ac-ceptable explanation for the above observationsis that myxoma and fibroma viruses grow betterat lower temperatures (88), although, strictlyspeaking, the difference could be more in skinreactivity than in virus growth.Epidermal response to infection. A few general
features of the pathogenesis of the epidermallesion will be mentioned. Infected epidermal cells,like other cells infected with cytopathic viruses,tend to swell and become vacuolated, and theymay degenerate. The first signs of infection areseen in the living cells of the stratum Malpighii(see Fig. 1). Affected cells often swell, and maydevelop intranuclear or intracytoplasmic in-clusions, depending on the virus. Cells may be-come vacuolated and necrotic, with nucleardisruption, and, when a focus of such cells hasdeveloped, a hole is produced in the epidermis.In response to the cell damage, inflammatoryfluid inevitably appears, and the hole becomes avesicle (variola, vaccinia, foot-and-mouth dis-ease). Sometimes intercellular edema is pro-nounced, and a fluid-filled cavity appears as thecells are parted or degenerate; vesicles may arisein this way without substantial cell necrosis.Since vesicles arise primarily in the stratumMalpighii, there is a roof of stratum corneumand often of granulosum or prickle cells of thestratum Malpighii. The floor may consist ofbasal epidermal (stratum germinativum) cellsor sometimes, when these are destroyed, thedermis itself. Vesicles may become pustular asleukocytes accumulate in them. In some instances,intraepidermal vesicles are small and do notunite, and a macroscopic vesicle is never seen,as in measles (85), extremely mild smallpox, ormalignant fulminating smallpox. Inevitably,vesicles are striking features in thick or thickenedepidermis, where a good roof is available. Forinstance, human smallpox lesions in the palmsand soles, where the stratum corneum is par-ticularly thick, may not disappear until after alllesions elsewhere in the body have healed. Onthe other hand, in the mouse, where the epider-mis is thin and the stratum Malpighii is onlyabout two cells deep, visible vesicles do not
easily develop, and papules give rise directly toulcers (37). In mucous membranes, although thedevelopment of lesions is basically the same asin the skin, the roof of vesicles, being sodden,breaks down early to produce an ulcer (herpessimplex); one result is that infectious virus isliberated at an earlier stage than from the skin.
In lesions caused by many viruses, epidermalcells proliferate, so that the epidermis becomessomewhat thicker (sheep-pox, smallpox, cow-pox). In some cases, the infected cells perhapsrespond by proliferating (fowlpox), but in othercases it seems likely that nearby but uninfectedepidermal cells react in this way, as indicated bythe work of Roberts (124) on ectromelia skinlesions in mice. The proliferating cells at the edgeof enlarging lesions are themselves eventuallyinfected and may become necrotic. Sometimesthe proliferative response in the epidermis isgross, so that tumors are produced. For instance,in the rabbit papilloma (132) or the human wart(66), cells in the stratum germinativum are initi-ally infected and respond by mitotic activity.The viruses causing these lesions perhaps have aslow growth cycle in epidermal cells, and virusparticles are not common until the cells arebecoming imperfectly keratinized as they ascendto the epidermal surface, presumably a few daysafter their infection. These atypically keratinizedcells come to form most of the dry horny papil-loma or wart.
Part Played by the Immune Response
In discussing immune responses, it is customaryto distinguish between the part played by cir-culating antibodies and that played by an al-tered cellular response or delayed type of hyper-sensitivity. This distinction may prove to be lessmeaningful in the last analysis, but it is never-theless a valid one, and the immune response willbe dealt with under these headings. The possiblerole of virus allergens in rashes must also be con-sidered. Immune damage will then be contrastedwith direct damage by viruses, and, finally, thefactors responsible for the regression of virusrashes will be discussed.Delayed type hypersensitivity. A hypersensi-
tivity reaction to viral materials is sometimesdemonstrable in animals recovered from virusinfections. As usually tested for by the intrader-mal injection of viral antigens, this consists of aslowly evolving cellular infiltration of the skin.This allergic or delayed type of hypersensitivityreaction can be elicited after infections with anumber of nonviral microorganisms, includingtuberculosis, brucellosis, glanders, leprosy, andlymphogranuloma inguinale, all of which are
VOL. 30, 1966 749
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
intracellular parasites involving especially macro-phages. It is also seen after certain poxvirusinfections [variola, vaccinia, ectromelia (37)]and mumps (35). All that need be said about ithere, and indeed almost all that can be saidabout it, is that it is a specific cellular response toan antigen.
Fifty years ago, von Pirquet (136) suggestedthat an altered host reactivity to virus materialplayed an important role in the development ofvaccinia, and perhaps also measles, Germanmeasles, and chickenpox skin lesions. Von Pir-quet studied the skin responses of unvaccinatedpatients to daily inoculations of vaccinia virusat different skin sites. He found that the evolu-tion of individual lesions proceeded independ-ently until 8 to 11 days, and then at the same timearound each lesion an inflammatory zone de-veloped. From these and similar studies on vac-cinia virus, and from related observations intuberculosis, serum-sickness, and other condi-tions, he concluded that the exanthem is producedas a result of the changed reactivity of the host.He suggested that circulating antibodies reactwith the virus in the skin to produce a toxic sub-stance which gives rise to the inflammatoryevents in the lesion. Pincus and Flick (104)confirmed and extended von Pirquet's observa-tion on vaccinial skin lesions. With the viruspreparations used by these workers, an allergicresponse could be detected as early as the 4thday after inoculation, and their evidence indi-cated that the subsequent evolution of the lesion,including vesicle formation, was attributable tothe allergic response.For many years, von Pirquet's idea remained
interesting but ill-defined. Ledingham (73) foundthat the intradermal injection of India ink locallyinhibited the reaction of allergy to vaccinia virusin rabbits, and that it also prevented the develop-ment of the local lesion produced by primary in-fection with vaccinia virus. The effect was sharplyconfined to the India ink injection site, and atypical vaccinial lesion once developed just out-side the edge of the ink mass. These findings wereconfirmed by Widelock (141), who also showedthat the ink injection did not affect virus multi-plication in the skin. It would be interesting toknow whether a smallpox eruption fails to in-volve tattoo marks. Presumably, the effect in-volves a generally changed reactivity of bloodvessels, because there is also no inflammatoryreaction to pneumococci or oil of citronella inthe ink injection site (4). The experiments, there-fore, do no more than indicate that for the de-velopment of any of these responses there mustbe an extravasation of fluid and cells into theskin.
In his far-reaching survey of the pathogenesisof virus diseases, Burnet (14) drew attention tovon Pirquet's work and gave a more up-to-dateinterpretation of the measles exanthem in termsof von Pirquet's hypothesis.A few years ago, the subject of the role of al-
lergy and delayed type hypersensitivity in thedeveopment of viral skin lesions was reopenedby the observations on patients with agamma-globulinemia, and by a number of interestinglaboratory experiments with vaccinia virus. Pin-cus and Flick (103) found that, when an anti-mononuclear cell serum was injected intra-dermally into guinea pigs, the lesions producedby vaccinia virus injected into the same sitefailed to evolve in the normal way. An erythema-tous area developed in the first few days, butneither papules nor vesicles appeared. Serumhemagglutination inhibiting antibody titers ap-peared as in control animals, indicating that theguinea pigs had indeed been infected. The de-layed type hypersensitivity response to dinitro-chlorobenzene could be inhibited in the sameway, and it was concluded that this response wasessential for the normal evolution of the vac-cinial skin lesion. Impressive confirmatoryevidence was obtained when rabbits were madeimmunologically tolerant to vaccinia, and theninjected intradermally with virus (41). In con-trast to control animals, skin reactions were eithercompletely absent or else abortive, with slightredness and scabbing but without vesicle forma-tion. These rabbits failed to produce antibody tovaccinia virus, and many died with generalizedvisceral infection. Virus can be assumed to havegrown in the skin, because typical vaccinial in-clusion bodies were seen in epidermal cells inthe lesions, just as in Widelock's (141) India inkexperiments, where epidermal cell swelling andhyperplasia occurred as usual, in spite of the factthat typical lesions failed to appear. There isstrong evidence, therefore, that in vaccinial skininfections much of the dermal swelling andedema, and even vesicle formation in the epider-mis, are mediated by a delayed type hypersensi-tivity response. Vesicles are also produced inthe delayed type hypersensitivity response ofcontact dermatitis, and sometimes when a tuber-culin patch is applied to the skin of sensitizedsubjects.There are a few observations suggesting that
host responses play an essential part in the de-velopment of the measles rash. Hecht's giantcell pneumonia appears to be an atypical responseto measles virus infection, and in three patientsfrom whose lungs virus was isolated postmortemthere had been neither Koplik spots nor rash(36). Each patient had an underlying chronic
750 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
disease, and it seems possible that the absenceof the rash was associated with a failure in hostreactivity to virus growth.
Antibodies. During the normal evolution of aprimary vaccination lesion in man, an erythema-tous area appears round the central zone after 7or more days, depending on the amount of virusinoculated. This coincides with the appearance
in the blood of circulating antibodies, and canbe attributed to an antigen-antibody reactionin the lesion. If viral antigens are present inblood vessel walls and circulating precipitatingantibody appears, then antigen-antibody com-plexes may be precipitated in vessel walls andan Arthus-type reaction is generated with primarydamage to these vessels. Antigen-antibody reac-tions may also occur extravascularly and lead toan increased capillary permeability, as seen inthe passive cutaneous anaphylaxis response. Inretrospect, the "allergy" referred to by von
Pirquet in his observations on primary vaccinialskin lesions probably consists of an antigen-antibody mediated response. Similar observationson the evolution of myxoma virus skin lesionsin rabbits have been made (Marshall, personalcommunication), and clear evidence about therole of circulating antibodies was obtained inthe following experiment (Mims and Roberts,unpublished data). A rabbit was given 40 intra-dermal inoculations with 104 LD50 of a virulentstrain of myxoma virus, and on day 3, when thelesions were slightly raised reddish papules, 40ml of myxoma-immune rabbit serum were in-jected intravenously. Within 1 hr, the skin lesionsdeveloped a deep red zone at their edges, sothat the lesion came to resemble those oftenseen with attenuated virus strains after 8 or moredays. This is taken as good evidence that anti-body plays a part, at least in the later stages inthe development of skin lesions.Although antibodies may be responsible for
the erythematous zone around established lesions,there is evidence that antibodies do not other-wise play a part in the evolution of the vaccinialskin lesion. X-irradiation, which suppressedcirculating antibody formation but not the de-velopment of delayed type hypersensitivity,failed to affect the development of vaccinia skinlesions (41, 43). Again, agammaglobulinemicpatients who are unable to produce antibodies,or at least have greatly reduced ability to do so(5), nevertheless generally develop typical skinlesions on primary vaccination and give immuneresponses to revaccination (48).Thus in the case of the best-studied viral skin
lesion, that produced by vaccinia virus, it seemsthat a delayed type hypersensitivity responseplays an important part in the early evolution of
the lesion. Although, as discussed earlier, theswelling and vacuolation of cells and the forma-tion of microvesicles may be a purely cellularresponse to infection, there is evidence thatvesicle production in the gross sense is mediatedby the delayed type hypersensitivity response.Later, the appearance of circulating antibodymay give the lesion an added inflammatorycharacter. However, it may often prove difficultto distinguish between the parts played by theseparately characterizable components of theimmune response. Skin-sensitizing antibodieswould be of possible importance in viral skinlesions, but nothing is known about these anti-bodies in virus diseases.
Allergic rashes. Certain clinically observedallergic rashes should be mentioned. They arefairly well characterized, but usually have nocounterpart in experimental animals, and theirpathogenesis is not understood. They maynevertheless be important in understanding thepathogenesis of viral rashes. These allergic rashesare seen in a number of human diseases and skinsensitivity can often be demonstrated, but theallergen has clearly not been locally introducedinto the skin at the site of the rash. For instance,urticarial erythematous or vesicular rashes areseen in serum sickness, in food and other aller-gies, in helminth infestations, in drug eruptions,and in the trichophytids of fungus infections. Theagent responsible for the eruption must beassumed to be blood-borne. Presumably antigensor allergens enter the circulation, localize in skinblood vessels, and an immunological reactionwhich occurs at some stage gives rise to the rash.The rash may be restricted in distribution, evento a localized skin area in the fixed drug erup-tions; it may appear in showers, and may tendto be symmetrical. Whatever the pathogenesis ofthese rashes, or the factors responsible for theirlocalization, their possible occurrence in virusdisease must be considered. The fleeting pro-dromal rashes sometimes seen in chickenpox,smallpox, or measles might well be rashes ofthis sort. The definitive rashes in certain entero-and arbovirus infections (Table 1) may alsocome into this category. Even in the case ofmeasles, there is no good evidence that infectiousvirus rather than viral allergen localizes in theskin to produce the rash, although Debre,Bonnett, and Broca (22) showed that intrader-mally injected immune serum locally inhibitedthe development of the measles rash, and theclosely related distemper virus has been shownby fluorescent-antibody staining to grow in theskin of intranasally infected ferrets (79).
Soluble viral antigens can certainly be liberatedinto the blood in generalized virus infections.
751VOL. 30, 1966
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
Rivers and Ward (113) demonstrated nonin-fectious precipitinogen in the serum of rabbitsinfected with myxoma virus. Using the precipitinreaction, Hughes (63) demonstrated solubleantigens in the serum of monkeys infected withyellow fever virus, and, more interestingly, theexperiments of Davis (21) indicated that theantigens in such sera can induce anaphylaxis inpassively sensitized animals.
In summary, some of the rashes seen in virusdiseases may well be allergic in nature, withcirculating materials localizing in the skin togive rise to immediate or delayed reactions.Such rashes have usually been described inman, and, although comparable skin reactionsare recorded in animals, for example, in fungusinfections in the guinea pig (56, 128), they mayprove to be of significance in man rather thananimals.
Direct damage by viruses. In spite of the factthat the immune response may be important inthe genesis of skin lesions produced by certainpoxviruses and perhaps by measles virus, therole of direct cell damage by viruses should notbe underestimated. For instance, viruses growingin blood vessels may damage cells to produceendothelial swelling, extravasation of fluid andleukocytes, or even hemorrhage and infarctionwith consequent anoxic changes in tissues. Atany stage, the products of cell destruction maythemselves generate nonspecific inflammatoryresponses. Thus, in spite of the fact that theimmune response cannot be important in thelesions produced by vaccinia virus on the chorio-allantoic membrane of chick embryos, necrosis,vacuolation, edema, and hemorrhage are never-theless prominent features. The role of the directdestructive action of viruses on skin is perhapsmore clearly separable from that of secondaryreactions in the primary lesions in foot-and-mouth disease. In Platt's (105) experiments onguinea pigs, where epidermal necrosis withpapules and even small vesicles were visible asearly as 24 to 48 hr after infection, it would seemunlikely that the immune response could havemade a notable contribution.The possible role of direct damage by toxins
may be mentioned. In the Schick test for exam-ple, Corynebacterium diphtheriae toxin injectedinto the skin of nonimmune subjects gives macu-lopapular lesions. In scarlet fever, circulatingstreptococcal toxins localize in the skin to pro-duce erythematous lesions, but in this casehypersensitivity to the toxin perhaps plays apart. Soluble viral toxins that are active inanimals have not been demonstrated but theyremain a possibility.
Finally, it should be pointed out that, even if
viruses can sometimes produce lesions in theskin without the help of the immune response,this cannot occur if infected cells show no cyto-pathic changes. Thus, in carrier mice congeni-tally infected with LCM virus, a state of immunetolerance permits the growth of virus in cells andtissues throughout the body (92). This virus,moreover, is completely noncytopathic for in-fected mouse cells, and carriers remain well inspite of the fact that all tissues are heavily in-fected. Foci of infection in the skin are seen byfluorescent-antibody staining, involving dermaland epidermal cells, but infected skin areas arecompletely healthy and show no pathologicalchanges.
Regression of lesions. The part played by anti-body and delayed type hypersensitivity in theregression of virus skin lesions merits brief dis-cussion. Antibodies may not be important,because vaccinial skin lesions regress normallyin most agammaglobulinemic patients, and alsoin X-irradiated animals which fail to producedetectable antibodies (43). The fact that pas-sively administered antibody may cause regressionof lesions, and that in immune animals antibodyprevents the formation of lesions, does not meanthat antibody is responsible for the regressionof lesions under normal circumstances. There isalso evidence that delayed type hypersensitivitydoes not contribute to the regression of primaryvaccinial skin lesions (134). If this is true, atleast for vaccinial lesions, it may be asked what,in fact, limits the growth of the primary lesionand prevents its spreading and covering a largepart of the body surface. There is good reasonto believe that interferon, which is present invaccinia skin lesions (97), may be of primaryimportance (44). It has been suggested thatinterferon is concerned in the recovery fromprimary virus infection, and antibody is involvedin the resistance to reinfection (6). Unfortunately,there are few studies that permit clear-cut con-clusions. For instance, in vaccinia gangrenosum,a rare complication of smallpox vaccination,there is indeed a progressive enlargement of theprimary lesion. Although antibodies are usuallynot demonstrable, and most cases heal after theadministration of immune globulins, this is notalways so, and in two instances the advance ofthe lesion was finally halted by the local injectionof immune leukocytes (69, 99). In most cases,interferon production was not investigated. It isdifficult to draw clear conclusions, because thesepatients may have had different and multipledeficiencies. Although lesions usually regressedwhen antibody was given, this does not meanthat antibodies normally perform this function,and there may have been an accompanying
752 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
failure in interferon production, or in otherresistance factors.
FACTORS AFFECTING THE OCCURRENCE OF RASHESIN VIRUS DISEASES
Rashes are seen in only a small proportion ofall generalized virus diseases, and it is of someinterest to attempt to account for the presenceor absence of a rash in different virus infections(see Table 1). Why do so many viruses, in spiteof the fact that they circulate in the blood, failto produce rashes? Unfortunately our basicignorance about rashes permits no more than afew general observations and speculations.
First, the following classification of the exan-thematous virus diseases can be made. Thereare two sets of viruses which grow in the dermisand epidermis and which, therefore, producelocal lesions when injected into the skin. Thefirst set comprises the poxviruses, herpes simplex-varicella, and probably the measles-distemper-
TABLE 1. Occurrenice of cutaneous eruptions ingeneralized virus diseases (75, 139)
Present' Absent or not recorded
Coxsackie Ag, Ai,,, Coxsackie (most)(B,, B3, B5)
Echo 9, 16, (4, 6, 18) Echo- (others)Vesicular exanthem EMCFoot-and-mouth dis- Poliovirusesease
(Reoviruses 2) Reoviruses (most)Arboviruses: group A Arboviruses: group A
(Chikungunya, (most), group BO'nyong-nyong), (most), group Cgroup B (West Nile, and miscellaneousDengue)
Measles-distemper- Fowl plague, mumps,rinderpest Newcastle disease
virusPox viruses CytomegalovirusesInfectious mononu- Infectious hepatitis
cleosisRubella RabiesVesicular stomatitisVaricella-zoster-
herpes simplex(Adenoviruses 1, 2, 3, Adenoviruses (most)
7)African horse sick-
nessPanleucopenia of catsPolyoma, simian virus
40, K virusAvian leucosis - sar-coma complex
And many other virusdiseases of animals
a Those in parentheses are less well established.
rinderpest group. These viruses are all large,are associated with leukocytes when in the blood,and can be thought of as being carried out ofdermal blood vessels by leukocytes, to infectdermal and epidermal cells. The second setincludes the foot-and-mouth disease-vesicularexanthem group, and probably coxsackie Ag andAi,, (3). These viruses are all very small, nonebeing larger than 30 , in diameter. They areprobably free in the plasma of infected animals,and they probably all grow in dermal or epi-dermal cells. Particles of this magnitude areperhaps capable of leaking through small skinblood vessels in normal animals (19, 52), andlesions can then be produced if they grow indermal or epidermal cells.
Other exanthematous viruses, including certainarboviruses and echoviruses, rubella, and infec-tious mononucleosis, can be thought of as failingto progress beyond the stage of vascular orperivascular infection. They are, therefore, un-likely to produce local lesions when injected intothe skin. They may even produce lesions by wayof allergic responses rather than by the growthof virus in the skin.Thus, any circulating virus, free or cell-asso-
ciated, which localizes in a skin blood vessel,has the opportunity to infect the vessel wall andgive rise to a vascular lesion. The infectious focuscould spread to involve dermal and epidermalcells as well, if they were susceptible. Even ifvirus fails to grow in blood vessels, leukocytesmight carry it through, or, if small, it mightleak through to extravascular tissues, where itwould again have an opportunity to produce alesion by growing in dermal or epidermal cells.
If localization in vessel walls takes placeaccording to the size of virus particles, themechanisms which localize exanthematous viruseswould also localize viruses which never give riseto skin lesions. Thus, if the rash inirecho 1 orWest Nile infections is a result of the localizationof virus in the skin, it can be asked why thereare no rashes in poliovirus or yellow fever virusinfections. In these last two diseases, circulatingvirus is known to localize in small blood vesselsin the brain (polio) and liver (yellow fever) toproduce lesions. Circulating virus is probablyfree in the plasma in each of these four infections,and it would seem unlikely that there is anythingcharacteristic of echo 16 or West Nile virusparticles which makes them localize in the skin,or about polio and yellow fever virus particleswhich makes them localize in the brain andliver. More probably, in each case virus particleslocalize in the same way according to uptake byreticuloendothelial cells and lodgement in capil-laries and venules as discussed earlier. But West
753VOL. 30, 1966
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
Nile virus, for instance, although lodged inKupffer cells, cannot initiate infection in theliver, just as poliovirus in small blood vessels inthe skin cannot give rise to a lesion. This doesnot exclude a possible affinity of certain virusesfor vascular endothelium in general, or for theendothelium in certain vascular beds. CoxsackieA virus might localize specifically in stripedmuscle blood vessels, or mumps virus in theblood vessels of certain glands. This explanationof the facts must have at least a limited validity.
In summary, the production of a rash byleukocyte-associated viruses may depend ontheir ability to grow in dermal and epidermalcells. Circulating free virus particles perhapslocalize nonspecifically in skin blood vesselsaccording to their size, and only those whichgrow in vessel walls or pass through vessel wallsto grow in extravascular tissues produce rashes.Obviously, there are likely to be major differencesin the ability of different viruses to grow invessel walls and in dermal and epidermal cells.With so little evidence, the above speculationsmay prove to be unfounded; indeed, all that can
be done with complete safety is to invoke the"dermotropism" of certain viruses, just as
Levaditi did 40 years ago.
DISCUSSION
In this survey of the pathogenesis of virusrashes, attention has been drawn to generalprinciples governing the development of skinlesions. Certain aspects of the subject have notbeen dealt with very thoroughly. For instance,virus-induced tumors of the skin have receivedno more than brief mention, and on the subjectof the zoster rash, there seemed little to add toDownie's (28) survey. The subject of rashesneeded reviewing, if only to restate the old prob-lems in modern terms, to bring together relevantinformation, and, in doing so, perhaps to have a
clearer insight into the gaps in our understandingof the behavior of the skin in virus diseases.Without a doubt, a full understanding of thepathogenesis of rashes awaits advances in our
basic knowledge of the skin and its responses.Generally, skin lesions produced by viruses
which grow in the skin when injected locallyhave been well studied, compared with thosethat produce rashes but no local lesion. Thus,experiments with poxviruses are often discussedat length, whereas remarks about arbo- orenterovirus rashes are largely speculatory. It isno accident that nearly all observations andmany of the experiments concern man. It mightbe expected that the skin of man, so closelyobserved by patient and physician for centuries,
would for this reason alone yield more informa-tion. Even the most minute blemishes are clearlyvisible on human skin because it is naked, andthey are more clearly visible on white than onblack skin. But there is another importantreason why the human skin figures so promi-nently in any discussion on rashes. Human skinis unique. In taking the place of fur, it still hasto be both an instrument of isolation from, andof communication with, the outside world. Oneimportant consequence is that the skin of manmust be thicker for mechanical protection, andthis involves differences in structure and cellrenewal rates, as well as a change in activity forthe oily skin glands. Also, because the sensoryfunction of hairs was largely lost, a reviseddistribution of skin sense organs was necessary.Finally, and most important, the skin became athermoregulatory organ of great significance. Toa greater extent than most mammals, man con-trols the rate of radiant heat loss from the skinby controlling the flow of blood through theskin. This has led to a very elaborate network ofsuperficial blood vessels, together with a delicatebut equally complex system of nervous andhumoral control. Finally, in a poorly understoodgeneral way human skin tends to reflect thestate of the central nervous system. This is notonly true for more obviously psychogenic re-sponses like blushing (every blush is a fleetingrash), but it is also true for the influence of themind on the skin, as illustrated in any textbookof psychosomatic medicine. A Mantoux reactioncan be inhibited by direct suggestion underhypnosis, presumably by an effect on the vascu-lar contribution to the response (11).
Thus, the nakedness of man has given his skinimportant protective, sensory, and thermoregula-tory adaptations which, taken together, makehuman skin unique. Interestingly, the skin of thedomestic pig has points of resemblance to thatof man, but there are at the same time funda-mental differences, both in vasculature and inskin glands (95). Human skin, in constant closetouch with the outside world, is both sensitiveand reactive, and it has been graphically describedas a "turbulent tissue" (94). It is this turbulentreactivity, perhaps, which often makes humanskin respond more vigorously than the hairedskin of animals to virus infections, as well as toother disease agents. Skin lesions are of course afeature of many virus diseases of animals, butthese lesions tend to be on exposed hairless areaswhere the skin has the human properties ofthickness, sensitivity, and vascular reactivity.Hence, although rashes may involve the generalbody surface of animals, it is udders, scrotums,ears, prepuces, teats, noses; and paws that are
754 Mims
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
more regular sites for lesions. For instance, theclosely related 'diseases of measles, distemper,and rinderpest can be compared. In rinderpest,areas of red moist skin, perhaps representing aconfluent eruption, with occasional vesiculation,are sometimes seen on the udder, scrotum, andinside of the thighs. In distemper, the exanthemis said to be seen on the abdomen and inneraspect of the thighs in 50% of cases, but therehas been some uncertainty as to whether it is infact due to the virus (23, 29). Yet, in humanmeasles one of the most florid and characteristicrashes known occurs, involving the general bodysurface. Even in susceptible monkeys, the samevirus produces skin lesions sparingly and irregu-larly.Mucous membranes are also a site for lesions,
and are important in shedding infectious virus,but human mucosae do not differ in any impor-tant way from those of most other mammals.Accordingly, enanthems are equally a feature inanimal and human virus diseases.The mucosal lining of the alimentary canal,
like the skin, is an epithelial structure, and,because it is composed of naked living cells, it isin some ways even more intimately exposed tothe external world. The localization of circulatingviruses in intestinal epithelium would presumablytake place according to the same general princi-ples as those discussed for the skin. The intestinalepithelium is probably often infected during virusdiseases, but there may be no signs of infectionuntil the appearance of gross symptoms likediarrhea, which is seen, for instance, in distemperin dogs (30), in cattle infected with rinderpestvirus (120), and even in measles (96). Little isknown about the development or incidence ofthese enteric lesions. Doubtless the thinness ofthe epithelial surface, the high rate of cell shed-ding, the ubiquity of other microorganisms, andthe mucus-lined, fluid surroundings, would givesuch lesions their special character.The above comparisons of lesion sites in man
and in animals highlight our ignorance of thefactors governing the distribution of skin lesionsin different virus diseases. This lack of under-standing can be illustrated when we ask whyfoot-and-mouth disease virus, even when itinfects man, produces lesions on the feet, hands,and mouth (40), or why the agent of Fort Braggfever gives skin lesions almost restricted to thepretibial region (130). Attention is therebydrawn to the importance of local differences inthe skin and hairs, sebaceous glands, vascularity,dermo-epidermal junctions, or epidermal thick-ness, which may vary widely in different regions(104). Any of these differences could lead todifferences in the localization of virus, the growth
of virus, or the production of lesions. For in-stance, Lewis and Harner (77) charted in a child'sleg the local variations in vascular activity whichare the basis for skin mottling, and they wereable to show that a measles rash involved theareas where blood vessels were more dilated.The work of Lewis on the vascular responses
of human skin (78) illustrates forcibly the powerand importance of clinical observation. Importantfacts and clear clues to principles may emergefrom such observations, as has been seen in therecent work on agammaglobulinemia (48). Clini-cal observations have already provided manyclues whose significance we have so far failed tounderstand. For instance, it is known thattuberculosis is activated by measles, that tuber-culin-positive patients become tuberculin-negativefor 1 or 2 weeks after a measles eruption (17),and that a measles rash appears early and se-verely at the site of a recent positive tuberculinskin test (118). In both diseases, macrophagesplay an important part and giant cells areformed.May not important principles lie hidden iobservations such as these? Again, it has bednreported that when lepers are vaccinated there isa generalized activation of leprous skin lesions,and the vaccinia skin lesions themselves areparticularly severe (24, 55). Also, why is it thatthe pyrexias of smallpox, tuberculosis, primaryatypical pneumonia, and brucellosis fail toactivate latent herpes simplex and give coldsores (9)? The clinician, as well as the biochemist,pathologist and immunologist, will assuredlyremain of fundamental importance in advancingour understanding of rashes.
SUMMARYThe subject of the pathogenesis of rashes in
virus diseases has been surveyed and discussed.Although this is an old and at present unfashion-able subject, recent advances in immunology,experimental pathology, and the pathogenesis ofvirus diseases seemed to justify a collectingtogether of information and a restatement ofproblems in modern terms.A preliminary section deals with the initiation
of infection in the skin, including the mechanismsof infection by biting arthropods.The localization of circulating viruses in skin
blood vessels is discussed at some length. Circu-lating inert particles appear to localize in bloodvessels in the apparently normal skin of animals,and leukocytes pass through vessel walls intotissues during the normal course of their wander-ings. The localization of free and leukocyte-associated viruses in skin blood vessels is surveyedin the light of these findings. The characteristicdistribution of skin lesions in different virus
VOL. 30,.1966 755
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

MIMS
exanthemata remains a mystery, although thereis some understanding of the mode of action oflocalizing factors.The spread of the infectious process after the
initial localization of virus in dermal bloodvessels is discussed. In some infections, thelesion remains restricted to the vicinity of theblood vessel, and the pathogenesis of the hemor-rhagic fevers is surveyed. The part played bythe ground substance in the spread of infectionthrough the dermis is examined. Infection ofthe epidermis is discussed, the possible role ofmelanocytes is considered, and the pathologicalresponse in the !pidermis is briefly referred to.A large section is devoted to the role of the
immune response in the pathogenesis of skinlesions. Recent work appears to support some ofthe suggestions made many years ago, but inmost viral rashes it is still not possible to assessthe relative importance of direct cell damage byviruses, nonspecific inflammatory changes, viraltoxins, and the immune or allergic response.
In a speculative attempt to account for theabsence of rashes in many generalized virus infec-tions, those infections with rashes are brieflyclassified and the pathogenesis of the rashes issurveyed.
In the discussion differences between the oc-currence of rashes in man and in animals iscommented on in relation to the uniqueness ofhuman skin. The production of lesions in mucousmembranes and in the epithelium of the ali-mentary canal is referred to. Finally, our basicignorance about rashes is reaffirmed, and, sincerashes are seen in man rather than in animals,attention is drawn to the continued importanceof clinical observations.
LITERATURE CITED1. ALKSNE, J. E. 1959. The passage of colloidal
particles across the dermal capillary wall un-der the influence of histamine. Quart. J.Exptl. Physiol. 44:51-66.
2. ALLEN, A. C., AND S. SPITZ. 1945. A comparativestudy of the pathology of scrub typhus (tsu-tsugamushi disease) and other rickettsialdiseases. Am. J. Pathol. 21:603-645.
3. ALsop, J., T. H. FLEwET1, AND J. R. FOSTER.1960. Hand-foot-and-mouth disease in Bir-mingham in 1959. Brit. Med. J. 2:1708-1711.
4. AVERY, R. C., AND R. H. RIGDON. 1939. Failureof an acute inflammatory process to extendinto area previously injected with Indian ink.Proc. Soc. Exptl. Biol. Med. 42:67-69.
5. BARON, S., J. P. NASON, R. M. FRIEDMAN, G. M.OWEN, H. BLEVY, AND E. V. BARNETT. 1962.Antibody production by hypogammaglobu-linaemic patients. J. Immunol. 88:443-449.
6. BARON, S. 1963. Mechanism of recovery fromviral infection. Advan. Virus Res. 10:39-64.
7. BARTON, J. C., AND R. G. GREEN. 1943. Histo-logic adaptation of the virus of fox encepha-litis. Am. J. Hyg. 37:21-36.
8. BAUER, J. H., AND N. P. HUDSON. 1928. Passageof the virus of yellow fever through the skin.Am. J. Trop. Med. 8:371-378.
9. BENNETT, 1. L., AND A. NICASTRI. 1960. Fever asa mechanism of resistance. Bacteriol. Rev.24:16-34.
10. BILLINGHAM, R. E. 1948. Dendritic cells. J. Anat.82:93-108.
11. BLACK, S., J. H. HUMPHREY, AND J. S. F. NIVEN.1963. Inhibition of Mantoux reaction bydirect suggestion under hypnosis. Brit. Med.J. 1:1649-1652.
12. BRAs, G. 1952. Observations on the formationof smallpox scars. A.M.A. Arch. Pathol.54:149-156.
13. BURNET, E. 1906. L'epithelioma contagieux desoiseux. Ann. Inst. Pasteur 22:742-765.
14. BURNET, F. M. 1955. Principles of animalvirology, chapter 9. Academic Press, Inc.,New York.
15. CALMETTE, A., AND C. GUERIN. 1901. Recherchessur la vaccine experimentale. Ann. Inst.Pasteur 15:161-168.
16. CAMus, L. 1917. Influence de la vasodilatationsur la localisation des pustules vaccinalesspontanees. Bull. Acad. Med. 77(series 3):111.
17. CHRISTENSEN, P. E., H. SCHMIDT, H. 0. BANG,V. ANDERSON, B. JORDAL, AND 0. JENSEN.1953. An epidemic of measles in southernGreenland, 1951. 11. Measles and tubercu-losis. Acta Med. Scand. 144:450-454.
18. COFFIN, D. L., A. M. COONS, AND V. J. CABASSO.1953. A histological study of infectious caninehepatitis by means of fluorescent antibody. J.Exptl. Med. 98:13-20.
19. COURTICE, F. C., AND D. G. GARLICK. 1962.The permeablllty of the capillary wall to thedifferent plasma lipoprotems of the hyper-cholerolaemic rabbit in relation to their size.Quart. J. Exptl. Physiol. 47:221-227.
20. DALES, S. 1965. Penetration of animal virusesinto cells. Progr. Med. Virol. 7:1-43.
21. DAVIS, G. E. 1931. Complement fixation inyellow fever in monkey and in man. Am. J.Hyg. 13:79-128.
22. DEBRE, R., H. BONNET, AND R. BROCA. 1923.Sur l'inhibition locale de l'eruption morbil-leuse par l'injection preable de serum de con-valescent. Compt. Rend. Soc. Biol. 89:70-72.
23. DE MONDBREUN, W. A. 1937. The histopathol-ogy of natural and experimental canine dis-temper. Am. J. Pathol. 13:187-212.
24. DENNEY, 0. E., AND R. HOPKINS. 1922. Specificleprous reactions and abnormal vaccinia in-duced in lepers by smallpox vaccination. Pub-lic Health Rept. U.S. 37:3141-3149.
25. DOLGOPOL, V. B. 1948. Histologic changes inrickettsialpox. Am. J. Pathol. 24:119-126.
26. DOUGLAS, S. R., W. SMITH, AND L. R. W. PRICE.1929. Generalized vaccinia in rabbits with
756 BACTERIOL. REV.
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
especial reference to lesions in the internalorgans. J. Pathol. Bacteriol. 32:99-120.
27. DOWNIE, A. W. 1939. A study of the lesionsproduced experimentally by cowpox virus. J.Pathol. Bacteriol. 48:361-379.
28. DOWNIE, A. W. 1959. Chickenpox and zoster.Brit. Med. Bull. 15:197-200.
29. DUNKIN, G. W. 1926. Vesico-pustules in dis-temper. Vet. Rec. 6:193-197.
30. DUNKIN, G. W., AND P. P. LAIDLAW. 1926.Studies in dog distemper. II. Experimentaldistemper in the dog. J. Comp. Pathol. Therap.39:213-221.
31. DURAN-REYNALS, F. 1942. Tissue permeabilityand the spreading factors in infection. Bac-teriol. Rev. 6:197-252.
32. DURAN-REYNALS, F. 1945. Immunological fac-tors that influence the neoplastic effects of therabbit fibroma virus. Cancer Res. 5:25-39.
33. EARLE, D. P. 1954. Symposium on epidemichaemorrhagic fever. Am. J. Med. 16:617-8.
34. ELLIS, R. A., AND G. MORETTI. 1959. Vascularpatterns associated with catagen hair folliclesin the human scalp. Ann. N.Y. Acad. Sci.83:448-457.
35. ENDERS, J. F., S. COHEN, AND L. W. KANE. 1945.Immunity in mumps. II. The development ofcomplement fixing antibody and dermal hy-persensitivity in human beings followingmumps. J. Exptl. Med. 81:119-135.
36. ENDERS, J. F., K. MCCARTHY, A. MITus, ANDW. J. CHEATHAM. 1959. Isolation of measlesvirus at autopsy in cases of giant cell phe-nomena without rash. N. Engl. J. Med.261 :875-881.
37. FENNER, F. 1948. The clinical features andpathogenesis of mouse pox (infectious ectro-melia of mice). J. Pathol. Bacteriol. 60:529-552.
38. FENNER, F., AND F. N. RATCLIFFE. 1965. Myxo-matosis. Cambridge Univ. Press, London.
39. FINDLAY, G. M. 1928. Histamine and infection.J. Pathol. Bacteriol. 31:633-644.
40. FLAUM, A. 1939. Foot-and-mouth disease inman. Acta Pathol. Microbiol. Scand. 16:197-213.
41. FLICK, J. A., AND W. B. PINCUS. 1963. Inhibitionof the lesions of primary vaccinia and of de-layed hypersensitivity through immunologicaltolerance in rabbits. J. Exptl. Med. 117:633-646.
42. FRIEDEWALD, W. F. 1942. Cell state as affectingsusceptibility to a virus. J. Exptl. Med. 75:197-219.
43. FRIEDMAN, R. M., AND S. BARON. 1961. Therole of antibody in recovery from infectionwith vaccinia virus. J. Immunol. 87:379-382.
44. FRIEDMAN, R. M., S. BARON, C. E BUCKLER, ANDR. L. STEINMULLER. 1962. The role of antibody,delayed hypersensitivity and interferon pro-duction in recovery from primary infectionwith vaccinia virus. J. Exptl. Med. 116:347-356
45. GAJDUSEK, C. 1962. Virus haemorrhagic fever.J. Pediat. 60: 841-857.
46. GEORGOPOULOS, M. 1928. Die Profung des Zu-standes der peripherem Gefasse als Methodezur fruhzeitigen und sicheren diagnose desDenguefiebers. Muench. Med. Wochchr. 75:1793-4.
47. GERARD, A. G., AND H. Y. TYLER. 1964. Cen-trifugal migration and transportation of sub-dermally injected colloidal gold. J. Invest.Dermatol. 42:305-306.
48. GOOD, R. A., R. A. BRIDGES, AND R. M. CON-DIE. 1960. Host-parasite relationships inpatients with dysproteinaemias. Bacteriol.Rev. 24:115-132.
49. GORDON, R. M., AND W. H. R. LUMSDEN. 1939.A study of the behaviour of the mouthparts ofmosquitoes when taking up blood from livingtissue, together with some observations on theingestion of microfilariae. Ann. Trop. Med.Parasitol. 33:259-278.
50. GRATIA, A., AND R. LINZ. 1931. Phenomene deSchwartzman et infection vaccinale. Compt.Rend. Soc. Biol. 108:238-240.
51. GREISMAN, S. E. 1957. Capillary observations inpatients with haemorrhagic fever and otherinfectious illnesses. J. Clin. Invest. 36:1688-1700.
52. GROTTE, G. 1956. Passage of dextran moleculesacross the blood-lymph barrier. Acta Chir.Scand. Suppl. 211:1-84.
53. HALL, J. G., AND B. MORRIS. 1963. The lymph-borne cells of the immune response. Quart. J.Exptl. Physiol. 48:235-247.
54. HALSTEAD, S. B. 1965. Dengue and haemorrhagicfevers of south-east Asia. Yale J. Biol. Med.37:434-451.
55. HASSELTINE, H. E. 1923. The effect of vacciniaon leprosy. Public Health Rept. U.S. 38:1-10.
56. HENRICI, A. T. 1939. Experimental trichophytidin guinea pigs. Proc. Soc. Exptl. Biol. Med.41 :349-353.
57. HOOK, E. W., C. N. LUTrRELL, K. SLATEN, ANDR. R. WAGNER. 1962. Hemorrhagic encephal-opathy in chicken embryos infected withinfluenza virus. IV. Endothelial localizationof viral antigens determined by immunofluores-cence. Am. J. Pathol. 41:593-599.
58. HOWATSON, A. F. 1962. Viruses connected withtumours and warts. Brit. Med. Bull. 18:193-198.
59. HUDACK, S., AND P. D. MCMASTER. 1932. Thepermeability of the wall of the lymphaticcapillary. J. Exptl. Med. 56:223-238.
60. HUDACK, S., AND P. D. MCMASTER. 1933. Thelymphatic participation in human cutaneousphenomena. J. Exptl. Med. 57:751-774.
61. HUDSON, J. B., L. WEINSTEIN, AND F. W. CHANG.1956. Thrombocytopenic purpura in measles.J. Pediat. 48:48-56.
62. HUEBNER, R. J., P. STAMPS, AND C. ARMSTRONG.1946. Rickettsialpox a newly describedrickettsial disease. 1. Isolation of the etio-logical agent. Public Health Rept. U.S.61:1605-1614.
VOL. 30, 1966 757
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
63. HUGHES, T. P. 1933. A precipitin reaction inyellow fever. J. Immunol. 25:275-294.
64. HURST, E. W. 1937. Myxoma and the Shopefibroma. I. The histology of myxoma. Brit. J.Exptl. Pathol. 18:1-15.
65. JOHNSON, R. T. 1965. Virus invasion of the cen-tral nervous system. Am. J. Pathol. 46:929-940.
66. JONFS, T. C., E. R. DOLL, AND J. T. BRYANS.1957. The lesions of equine viral arteritis.Cornell Vet. 47:52-68.
67. JUHLIN, L. 1956. The role of oedema in theclearance of Evans blue and haemoglobinfrom inflammatory foci. Acta Pathol. Micro-biol. Scand. 38:385-402.
68. KELLER, R. 1958. Passage of bacteriophagethrough intact skin of mice. Science 128:718-719.
69. KEMPE, C. H. 1960. Studies on smallpox andcomplications of smallpox vaccination. Pedi-atrics 26:176-189.
70. KIDD, J. G., AND R. J. PARSONS. 1936-37. Tissue.affinity of Shope papilloma virus. Proc. SocExptl. Biol. Med. 35:438-441.
71. KOPLIK, L. H. 1935. Shwartzman phenomenonin vaccine virus lesions. Am. J. Pathol. 11:842-843.
72. LAVOIPIERRE, M. M. J., AND R. F. RIEK. 1955.Observations on the feeding habits of Argasidticks and on the effect of their bites on labora-tory animals, together with a note on theproduction of coxal fluid by several of thespecies studied. Ann.Trop. Med. Parasitol.49:96-113.
73. LEDINGHAM, J. C. G. 1927. The role of the reticu-loendothelial system of the cutis in experimen-tal vaccinia and other infections: experimentswith Indian ink. Brit. J. Exptl. Pathol. 8:12-25.
74. LEEDHAM, C. L. 1953. Epidemic hemorrhagicfever: A summarization. Ann. Internal Med.38:106-112.
75. LERNER, A. M., J. 0. KLEIN, J. D. CHERRY,AND M. FINLAND. 1963. New viral exanthems.New Engl. J. Med. 269:678-685, 736-740.
76. LEVADITI, C., AND S. NICOLAU. 1923. Ectoder-moses neurotropes. Etudes sur la vaccine.Ann. Inst. Pasteur 37:1-106.
77. LEWIS, T., AND I. HARMER. 1926. Observationsupon the rupture of minute vessels in the skinand upon the distribution of cutaneous haem-orrhages and other skin eruptions. Heart13:337-351.
78. LEwIS, T. 1927. The blood vessels of the humanskin and their responses. Shaw & Sons,London.
79. LIu, C., AND D. L. COFFIN. 1957. Studies oncanine distemper infection by means of fluo-rescein-labeled antibody. I. The pathogenesis,pathology and diagnosis of the disease inexperimentally infected ferrets. Virology 3:115-131.
80. MCMASTER, P. D., AND R. J. PARSONS. 1950.The movement of substances and the state of
the fluid in the intradermal tissue. Ann. N.Y.Acad. Sci. 52:992-1000.
81. MCMASTER, P. D. 1941. An inquiry into thestructural conditions affecting fluid transportin the interstitial tissue of the skin. J. Exptl.Med. 74:9-28.
82. MCNAIR-SCOTr, T. F., H. BLANK, L. L. CORIELL,AND H. CROUSE. 1950. Pathology and patho-genesis of the cutaneous lesions of variola,vaccinia, herpes simplex, herpes zoster andvaricella, p. 74-98. In J. G. Kidd [ed.], Thepathogenesis and pathology of virus deseases.Columbia Univ. Press, New York.
83. MACCULLUM, W. G., AND L. M. MOODY. 1921.Alastrim in Jamaica. Am. J. Hyg. 1:388-408.
84. MAJNO, G., AND G. E. PALADE. 1961. Studies oninflammation. I. The effect of histamine andserotonin on vascular permeability: An elec-tron microscopic study. J. Biophys. Biochem.Cytol. 11:571-605.
85. MALLORY, F. B., AND E. M. MEDLAR. 1919. Theskin lesions in measles. J. Med. Res. 41: 327-348.
86. MARCHESI, V. T., AND H. W. FLOREY. 1960.Electron micrographic observations on theemigration of leucocytes. Quart J. Exptl. Phys-iol. 45: 343-348.
87. MARETIC, Z., AND M. P. M. COORAY. 1963.Comparisons between chicken pox in a tropi-cal and a European country. J. Trop. Med.Hyg. 66:311-315.
88. MARSHALL, I. D. 1959. The influence of ambienttemperature on the course of myxomatosis inrabbits. J. Hyg. 57:484-497.
89. MAURER, F. D., R. A. GRIESEMER, AND T. C.JorN.s. 1958. The pathology of African swinefever A comparison with hog cholera. Am. J.Vet. Res. 19:517-539.
90. MIMs, C. A. 1956. The coagulation defect inRift Valley fever and yellow fever virus infec-tions. Ann. Trop. Med. Parasitol. 50:147-149.
91. MiMs, C. A. 1964. Aspects of the pathogenesisof virus diseases. Bacteriol. Rev. 28:30-71.
92. MIMs, C. A. 1966. Immunofluorescent study ofthe carrier state and mechanism of verticaltransmission in lymphocytic choriomeningitisvirus infection in mice. J. Pathol. Bacteriol.89:395-402.
93. MiMs, C. A. 1967. The response of mice to theintravenous injection of cowpox virus. Brit. J.Exptl, Pathol., in press.
94. MONTAGNA, W. 1962. The structure and functionof skin. 2nd ed. Academic Press, Inc., NewYork.
95. MONTAGNA, W., AND J. S. YUN. 1964. The skniof the domestic pig. J. Invest. Dermatol. 43:11-21.
96. MORLEY, D. C. 1962. Measles in Nigeria. Am. J.Diseases Children 103:230-233.
97. NAGANO, Y., AND Y. KOJIMA. 1958. Inhibitionde l'infection vaccinale par un facteur liquidedans le tissu infecte par le virus homologue.Compt. Rend. Soc. Biol. 152:1627-1629.
758 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

RASHES IN VIRUS DISEASES
98. NELSON, E. R. 1960. Hemorrhagic fever in chil-dren in Thailand. J. Pediat. 56:101-108.
99. O'CONNELL, C. J., D. T. KARZON, A. L. BAR-RON, M. E. PLAUT, AND V. M. ALI. 1964.Progressive vaccinia with normal antibodies.Ann. Internal Med. 60:282-289.
100. ODLAND, G. F. 1958. The fine structure of theinterrelationship of cells in the human epider-mis. J. Biophys. Biochem. Cytol. 4:529-538.
101. PARKER, E. R., AND W. J. MAcNEAL. 1944.Persistence of influenza virus on human hand.J. Lab. Clin. Med. 29:121-126.
102. PARKER, R. F., AND R. L. THOMPSON. 1942. Theeffect of external temperature on the courseof infectious myxomatosis of rabbits. J.Exptl. Med. 75:567-573.
103. PINCUS, W. B., AND J. A. FLICK. 1963. Inhibitionof the primary vaccinial lesion and of delayedhypersensitivity by an antimononuclear cellserum. J. Infect. Diseases 113:15-21.
104. PINCUS, W. B., AND J. A. FLICK. 1963. The roleof hypersensitivity in the pathogenesis of vac-cinia virus infection in humans. J. Pediat.62:57-62.
105. PLATT, H. 1958. Observations on the pathologyof experimental foot-and-mouth disease in theadult guinea pig. J. Pathol. Bacteriol. 76:119-131.
106. PLATT, H. 1960. The localization of lesions inexperimental foot-and-mouth disease. Brit. J.Exptl. Pathol. 41:150-159.
107. PLATT, H. 1963. The engulfment of particulateand colloidal materials by epidermal cells. J.Pathol. Bacteriol. 86:113-122.
108. PLATT, H. 1963. The susceptibility of the skinand epidermoid mucous membranes to virusinfection in man and other animals. Guy'sHosp. Rept. 112:479-497.
109. PLATT, H. 1964. The local and generalized formsof experimental herpes simplex infection inguinea pigs. Brit. J. Exptl. Pathol. 45:300-308.
110. PLOWRIGHT, W., W. G. MACLEOD, AND R. D.FERRIS. 1959. The pathogenesis of sheep poxin the skin of sheep. J. Comp. Pathol. Therap.69:400-413.
111. RICKETTS, T. F. 1908. The diagnosis of smallpox.Cassell & Co. Ltd., London.
112. RIVERS, T. M., AND W. S. TILLETT. 1923. Studieson varicella. J. Exptl. Med. 38:673-690.
113. RIVERS, T. M., AND S. M. WARD. 1937. Infec-tious myxomatosis of rabbits. J. Exptl. Med.66:1-13.
114. ROBETS, J. A. 1962. Histopathogenesis of mousepox. II. Cutaneous infection. Brit. J. Exptl.Pathol. 43:462-468.
115. ROBERTS, J. A. 1964. Enhancement of the viru-lence of attenuated ectromelia virus in micemaintained in a cold environment. AustralianJ. Exptl. Biol. Med. Sci. 42:657-666.
116. ROTHMAN, S., AND A. L. LORINCZ. 1963. Defencemechanisms of the skin. Ann. Rev. Med. 14:215-242.
117. SABIN, A. B. 1952. Research on dengue during
World War ll. Am. J. Trop. Med. Hyg.1:30-50.
118. SC1HICK, B. 1910. Scheinbaves Aufflammenabgelaufener Tuberkulinreaktionen Wahrendder Eruption von Masern Fruhzeitige Erup-tion von Masern-efflorescenzen au entzund-lich gereizten Hautparteilen. Monatsschr.Kinderheilk. 9(orig ):137-140.
119. SCHOEFL, G. L. 1964. Electron microscopic ob-servations on the regeneration of blood vesselsafter injury. Ann. N.Y. Acad. Sci. 116:789-802.
120. SCOTT. G R. 1964. Rinderpest. Advan. Vet. Sci.9:113-224.
121. SELBY, C. C. 1955. An electron microscope studyof the epidermis of mammalian skin in thinsections. I. Dermo-epidermal junction andbasal cell layer. J. Biophys. Biochem. Cytol.1:429-443.
122. SHOPE, R. E. 1933. Infectious papillomatosis ofrabbits. J. Exptl. Med. 58:607-624.
123. SHOPE, R. E. 1940. Swine pox. Arch. Ges.Virusforsch. 1:457-467.
124. SIEFRIED, O., AND C. B. CAIN. 1932. Histologicalstudies on hog cholera. II. Lesions of thevascular system. J. Exptl. Med. 56:345-349.
125. SPROUL, E. E., R. S. METZGAR, AND J. T. GRACE.1963. The pathogenesis of Yaba virus-inducedhistiocytomas in primates. Cancer Res. 23:671-675.
126. SPRUNT, D. H. 1932. Infectious myxomatosis(Sanarelli) in pregnant rabbits. J. Exptl. Med.56:601-608.
127. SPRUNT, D. H. 1950. The ground substance ininfection. Ann. N.Y. Acad. Sci. 52:1052-1060.
128. SULZBERGER, M. B. 1950. Immunologic changesbrought about by fungi and fungous products.Ann. N.Y. Acad. Sci. 50:767-772.
129. SZABO, G. 1959. Quantitative histological inves-tigations on the melanocyte system of thehuman epidermis, p. 99-125. In M. Gordonfed.], Pigment cell biology. Academic Press,Inc., New York.
130. TATLOCK, H. 1947. Studies on a virus from apatient with Fort Bragg Fever (Pretibialfever). J. Clin. Invest. 26:287-297.
131. THOMPSON, R. L. 1938. The influence of temper-ature upon proliferation of infectious fibromaand infectious myxoma viruses in vivo. J. In-fect Diseases 62:307-312.
132. TILLETT, W. S., AND T. M. RIVERS. 1924.Atypical distribution of varicella lesions depen-dant upon a coexistent syphilitic infection.Johns Hopkins Hosp. Bull. 35:137-140.
133. TREJOS, A., AND A. ROMERO. 1954. Prothrombinlevels in yellow fever. Rev. Biol. Trop. 2:69-73.
134. TURK, J. L., A. C. ALLISON, AND M. N. OXMAN.1962. Delayed hypersensitivity in relation tovaccination and multiplication of vacciniavirus in the guinea pig. Lancet 1:405-407.
VOL. 30, 1966 759
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from

BACTERIOL. REV.
135. TYZZER, E. E. 1906. The histology of the skinlesions in varicella. J. Med. Res. 14:361-392.
136. VON PIRQUET, C. E. 1911. Allergy. Arch InternalMed. 7:259-288, 383-436.
137. VON PJRQUET, C. E. 1913. Das Bild der Masernauf der ausseren Haut. Z. Kinderheilk.6(orig.) :1-226.
138. WATrENBERG, L. W., B. L. ELISBERG, C. L.WISSEMAN, AND J. E. SMADEL. 1955. Studieson rickettsial toxins. 11. Altered vascularphysiology in rickettsial toxaemia of mice. J.Immunol. 74:147-157.
139. WENNER, H. A., AND T. YONG LON. 1963. Virusdiseases associated with cutaneous eruptions.Progr. Med. Virol. 5:219-294.
140. WHITELY, H. J. 1956. The effect of the hair-growth cycle on the development and distribu-tion of virus-induced lesions in the skin of therabbit. J. Pathol. Bacteriol. 72:1-13.
141. WIDELOCK, D. 1938. Studies on vaccinia virus:determiination of the correlative factors be-tween the rate of the multiplication of the
virus in the skin and the histological changesproduced. J. Infect. Diseases 62:27-35.
142. WOLBACH, B. S. 1951. The hair cycle of themouse and its importance in the study of se-
quences of experimental carcinogenesis. Ann.N.Y. Acad. Sci. 53:517-536.
143. WOLBACH, S. B. 1919. Studies on Rocky Moun-tain spotted fever. J. Med. Res. 41:1-197.
144. WOLBACH, S. B. 1948. The pathology of therickettsial diseases of man, p. 118. In Therickettsial diseases of man. American Associa-for the Advancement of Science, Washington,D.C.
145. WOLBACH, S. B., J. L. TODD, AND F. W. PAL-FREY. 1922. The etiology and pathology oftyphus. Harvard Univ. Press, Cambridge,Mass.
146. YOFFEY, J. M., AND C. K. DRINKER. 1939. Thecell content of peripheral lymph and its bear-ing on the problem of the circulation of thelymphocyte. Anat. Rec. 73:417-429.
760 MIMS
on June 12, 2018 by guesthttp://m
mbr.asm
.org/D
ownloaded from