j. cell sci. printed in great britain - home | journal of cell...
TRANSCRIPT

J. Cell Sci. 23, 85-100 (1977) 85
Printed in Great Britain
GIBBERELLIC ACID-INDUCED GERMINATION
OF SPORES OF ANEMIA PHYLLITIDIS:
AUTORADIOGRAPHIC STUDY OF THE
TIMING AND REGULATION OF NUCLEIC
ACID AND PROTEIN SYNTHESIS IN RELATION
TO CELL MORPHOGENESIS
V. RAGHAVAN
Department of Botany, The Ohio State University, Columbus, Ohio 43210, U.S.A.
SUMMARY
The timing and regulation of nucleic acid and protein synthesis during gibberellic acid-induced germination of spores of the fern, Anemia phyllitidis (L.) Swartz were investigatedusing metabolic inhibitors combined with cytological and autoradiographic techniques. Incu-bation of spores in gibberellic acid for at least 24 h was necessary for their subsequent germina-tion in the absence of the hormone. Based on the action of cycloheximide, it appeared thatcontinuous synthesis of proteins was essential for the hydrolysis of storage reserves, enlarge-ment of the protoplast, division of the spore nucleus and formation of the protonemal cellassociated with germination. Addition of actinomycin D or 5-rluorouracil along with gibberellicacid did not visibly affect germination, although the latter inhibited division of the spore nucleus.Both compounds also inhibited RNA synthesis in germinating spores without any noticeableeffect on DNA or protein synthesis. Cordycepin was most effective in inhibiting germinationwhen it was added during the first 24 h of gibberellic acid action. Spores visibly germinated inthe absence of nuclear division when DNA synthesis was inhibited by hydroxyurea. On theother hand, incubation of spores in an osmoticum such as mannitol led to no discernible signsof germination, although division of the nucleus and formation of the protonemal cell occurredin the absence of cell enlargement. It is tentatively proposed that the dry spore contains a stablemRNA which is activated in the presence of gibberellic acid. This mRNA provides templatesfor the synthesis of proteins necessary for the early events of germination.
INTRODUCTION
Spores of the fern, Anemia phyllitidis germinate in complete darkness in the presenceof low concentrations of gibberellic acid (GA) (Schraudolf, 1962; Weinberg & Voeller,1969). Histochemical and cytological studies have shown that the first noticeable signof spore germination is the hydrolysis of storage protein granules. This is followedby migration of the nucleus from its central location in the spore to one side andlocalized enlargement of the protoplast at the site of the nucleus. The enlarging tipof the protoplast breaks the spore coat (exine) and appears outside as a papillatestructure (protonemal initial). The nucleus undergoes mitosis forming 2 nuclei withthe spindle oriented in the plane of the protonemal initial which is cut off from therest of the spore by a cross-wall to form the protonemal cell. Derivatives of theprotonemal cell form the mature gametophyte or prothallus. A second nuclear division

86 V. Raghavan
in the spore cell in which the cross-wall is oriented at right angles to the first gives riseto the rhizoid cell. The rhizoid cell elongates to form a single-celled rhizoid. Theseordered morphogenetic events occur in spores sown in a medium containing GAduring a germination time course of 3-4 days; in the absence of GA, spores remainessentially dormant and retain the structural features of dry spores (Raghavan, 1976).
Since hydrolysis of protein granules is initiated in spores as early as 12 h afteraddition of GA, it has been deduced that a primary role of the hormone in germinationis in the synthesis of hydrolytic enzymes concerned with the breakdown of storagereserves (Raghavan, 1976). The origin of mRNA which serves as templates for enzymeproteins and the manner in which translation of this body of mRNA is accomplished areof interest, since such data might throw light on the processes involved in cell morpho-genesis, which determine the fate of the progeny of a single-celled spore. With this inmind, the role of transcription and translation processes at different stages of germina-tion of spores of Anemia phyllitidis has been studied. In this paper, the effects ofinhibitors of protein (cycloheximide) and nucleic acid synthesis (actinomycin D,cordycepin, 5-fluorouracil and hydroxyurea) and an osmoticum (mannitol) ongermination and on autoradiographically detectable incorporation of precursors ofmacromolecule synthesis into germinating spores are reported.
MATERIALS AND METHODS
Culture procedures
Spores of Anemia phyllitidis (L.) Swartz supplied by Dr Alice F. Tryon (Harvard University,Cambridge, Mass.) were used in this work. Procedures for aseptic culture of spores have beendescribed previously (Raghavan, 1976) and can be summarized as follows. Spores sterilized inClorox were allowed to imbibe in the dark in 10 ml of a mineral salt medium for4 days at 25 CC.Unless otherwise stated, they were then transferred to a fresh medium containing 100 mg/1.GA with or without the test substance and kept in the dark. All organic compounds weresterilized through Millipore filters and added to the medium. Spores in which the spore coatwas broken and the protonemal initial was visible outside were scored for germination 4 daysafter transfer to GA. Each chemical was tested in at least 2 experiments in which the treatmentswere run in duplicate. At least 200 spores were counted in each case to determine the percentageof germination.
Histological methods
The effect of chemicals on cell morphogenesis during germination was studied by histologicalmethods. For this purpose, spores were fixed in 10 % acrolein overnight in the cold. Proceduresfor dehydration and embedding in glycol methacrylate were as previously reported (Raghavan,1976). Sections cut at 5-7 fim thickness on a rotary microtome using a steel knife were stainedwith azure B or toluidine blue and mounted in Euparal for microscopic examination.
Autoradiography
To study the effect of chemicals on the synthesis of nucleic acids and proteins, fully imbibedspores were transferred to 25 ml of medium containing GA with or without an optimumconcentration of the test substance and incubated in the dark for 60 h. Towards the terminalperiod of a 60-h germination time course, cultures were supplied with [3H]thymidine (5-0 /tCi/ml; sp. act. 250 Ci/mmol), [3H]uridine (50/iCi/ml; sp. act. 25-0 Ci/mmol) or ['Hjleucine(2'O/iCi/ml; sp. act. 460 Ci/mmol), serving, respectively, as precursors for the synthesis ofDNA, RNA and proteins. The period of exposure of spores to [3H]thymidine was 12 h, while

Gibberellic acid-induced spore germination 87
['HJuridine and [3H]leucine were supplied for 4 h each. Spores were collected by washingbriefly in water, followed by 15 min washing in the respective unlabelled precursors (i-o mg/1.),fixed in acrolein and embedded in glycol methacrylate. Sections cut at 5-7 /tm thickness weremounted on slides which were then coated with Kodak NTB3 liquid emulsion and exposed inthe dark for 15 days. The slides were developed for 5 min in chilled 1:1 Kodak D-19 developer,fixed, stained in azure B and mounted in Euparal.
When 5-fluorouracil was the inhibitor, ["HJadenosine (s-o/tCi/ml; sp. act. 17-0 Ci/mmol)was used as the precursor to monitor RNA synthesis during an exposure period of 4 h. Since[sH]adenosine is incorporated into both DNA and RNA, sections were washed in DNase(o-i mg/ml in 0-003 M MgSO4, pH 6#S, 4 h at 30 °C) before coating them with the emulsion.Although the pattern of autoradiographically detectable synthesis of macromolecules remainedessentially unchanged by incubating spores for shorter periods of time in higher concentrationsof the isotope [for example 2 h in PH]thymidine (10 /tCi/ml), 1 h in [*H]uridine, fHJadenosine(10/iCi/ml) or [3H]leucine (5/iCi/ml)], in the interest of economy and autoradiographicresolution, we have used throughout this work longer periods of incubation of spores in lowerconcentrations of isotopes.
Chemicals
GA (75 % K salt) and hydroxyurea used in this work were purchased from NutritionalBiochemicals Corp., Cleveland, Ohio. Radioisotopes as specified above were obtained fromSchwarz/Mann, Orangeburg, New York. Actinomycin D, cycloheximide and 5-fluorouracilwere generously supplied by Merck, Sharpe & Dohme (Rahway, New Jersey), The UpjohnCompany (Kalamazoo, Michigan) and Hoffman-La Roche (Basle, Switzerland), respectively.All other chemicals were of reagent grade.
RESULTS
When fully imbibed spores were cultured in a medium containing GA and examined4 days later, maximum germination obtained in the sample was about 70%. By thistime, the germinated spore formed a 1- or 2-celled protonemal filament and a moder-ately long rhizoid (Fig. 1). Incubation of the spores in the same medium for anadditional 2 days in the dark led to no further increase in the percentage of germinationalthough formation of a few more cells in the protonemal filament and further elonga-tion of the rhizoid occurred. To determine the minimum period of exposure to GAfor germination, fully imbibed spores were treated with the hormone for varying periodsof time, washed thoroughly in distilled water and transferred to the basal medium(without GA). Percentages of germination were determined 4 days after the beginningof GA application. As seen from the results in Table i, about 30% of spores germi-nated after contact with GA for 12 h and maximum germination was obtained after24 h in GA. Further contact of spores with GA did not lead to an increase in thepercentage of germination. From these results it appears that the germination-promoting effects of GA are accomplished in the first 24 h of treatment of spores withthe hormone. However, apart from an incipient hydrolysis of storage reserves,sections of spores examined after 24 h in GA did not reveal any other signs of hormoneaction.
Requirement for protein synthesis
It was stated earlier that hydrolysis of storage reserves was initiated in spores asearly as 12 h after GA application. The requirement for protein synthesis for
V. Raghavan
8
Pgh< 1

Gibberellic acid-induced spore germination 89
GA-induced hydrolysis of storage reserves during germination was investigated usingcycloheximide. This drug which prevents the initiation and formation of nascentpolypeptides on ribosomes, is considered to be a general inhibitor of protein synthesis(Lin, Mosteller & Hardesty, 1966). Continuous incubation of fully imbibed spores in
Table 1. Effect of pretreatment of spores with GAfor varying periods oftime on their subsequent germination in the basal medium
Pret rea tment in G A , h . . .o 1 2 4 8 12 18 24 36 48 60 72 96 (continuous)
% germination . . .o o o 6 12 28 57 69 65 71 67 62 62
GA and cycloheximide (0-5 mg/1.) inhibited germination in the entire population ofspores when examined 4 days later. In sections, spores treated with the drug exhibitednone of the usual symptoms of germination including hydrolysis of storage proteingranules and were thus very similar in structure to the dry spores (Fig. 2). The effect
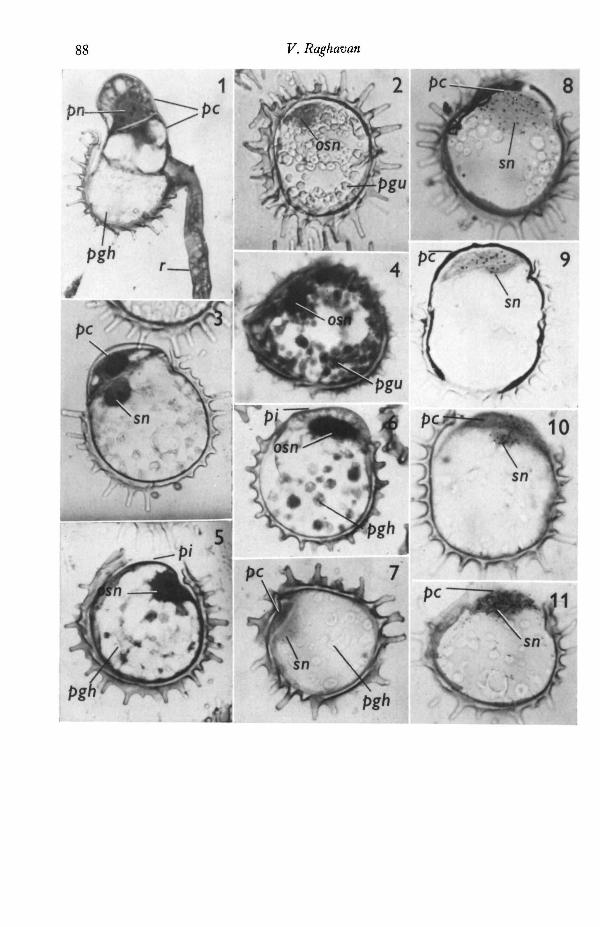
Figs. 1-7. Effect of inhibitors of synthesis of nucleic acids and protein, and ofmannitol on cell morphogenesis during germination. Fully imbibed spores were allowedto germinate in medium containing GA with or without the test substance for 4 days.All x 950. osn, original spore nucleus before division; pc, protonemal cell(s); pgh,storage protein granule in the process of hydrolysis; pgu, unhydrolysed proteingranule; pi, protonemal initial; pn, nucleus of the protonemal cell; r, rhizoid; sn, sporenucleus after division.
Fig. 1. Control, medium containing only GA. The spore has germinated to form a2-celled protonemal filament and an elongate rhizoid.
Fig. 2. Cycloheximide (0-5 mg/1.). The spore has not germinated.
Fig. 3. Actinomycin D (200 mg/1.). The spore has germinated to form the protonemalcell.
Fig. 4. Cordycepin (8-0 mg/1.). The spore has not germinated.
Fig. 5. s-Fluorouracil (50-0 mg/1.).
Fig. 6. Hydroxyurea (120 mg/1.). In this and the previous figure, spores have germi-nated to form the protonemal initial, although nuclear division has not occurred.
Fig. 7. Mannitol (o-6 M). The spore has not visibly germinated, but mitosis of thespore nucleus followed by cytokinesis has occurred to form a protonemal cell withinthe confines of the exine.Figs. 8-11. Autoradiographs showing the incorporation of radioactive precursors ofnucleic acids and protein into 60-h-old spores germinating in medium containingGA. In all cases, nuclear division has occurred and spores have germinated to form theprotonemal cell.
Fig. 8. [3H]thymidine. Silver grains are associated with the original spore nucleus,nucleus of the protonemal cell and cytoplasm of the latter.
Fig. 9. ['H]uridine.Fig. 10. [3H]adenosine. In this and the previous figure, silver grains are associated
with the nucleoli and the rest of the nuclear material.
Fig. 11. [*H]leucine. The cytoplasm of the protonemal cell and the nuclei of thespore cell and protonemal cell are covered with silver grains.

90 V. Raghavan
of the drug was transient since spores resumed normal germination when they weretransferred to a medium lacking cycloheximide. Therefore, it seems that cycloheximide-sensitive synthesis of proteins is essential for the hydrolysis of storage reserves and forthe subsequent initiation of the protonemal cell and rhizoid. As shown in Figs. 12-14in the presence of cycloheximide, spores did not incorporate labelled precursors intoDNA, RNA or proteins. For comparison, the corresponding autoradiographs ofspores germinated for the same length of time in the absence of the inhibitor are shownin Figs. 8, 9, 11. As described in an earlier paper (Raghavan, 1976), in 60-h-oldgerminating spores, pHJthymidine was incorporated into the spore nucleus, nucleusof the protonemal cell and the immediately surrounding cytoplasm of the latter,while [3H]uridine incorporation appeared in the nucleolus and the rest of the nuclearmaterial of both cells. Incorporation of [3H]thymidine into nucleus and cytoplasm isreadily removed by DNase digestion. This as well as the fact that cytoplasmic incor-poration of pHJthymidine is restricted to a short period of the germination timecourse [48-72 h after addition of GA; see Raghavan (1976)] suggests the existence ofa genuine cytoplasmic DNA synthesis during germination of spores of Anemiaphyllitidis. This also rules out the possibility that thymidine is demethylated touridine and incorporated into RNA during the long incubation period employed in thepresent study. [3H]Leucine incorporation, indicative of protein synthesis, or redistri-bution of newly synthesized protein, was mainly confined to the cytoplasm of theprotonemal cell and to some extent to the nuclei of the spore cell and protonemal cell.It is not established with certainty whether in 60-h-old germinating spores studiedhere the appearance of pH]leucine label in the nucleus is due to the synthesis ofproteins in that organelle or due to the transport of proteins synthesized in thecytoplasm to the nucleus. However, as shown in figs. 42 and 43 of an earlier study(Raghavan, 1976), during 36-48 h after addition of GA, before the basophilic cyto-plasm surrounding the nucleus is established in the germinating spore, [3H]leucine isexclusively incorporated into the nucleus. It should also be noted that no silver grains,suggestive of incorporation of isotopes, were seen associated with the great bulk of thespore cytoplasm containing reserve storage granules in various stages of hydrolysis.
Another experiment was performed to determine the time after addition of GA whenproteins essential for the breakdown of storage reserves were formed. In this experiment,spores were supplied with 0-5 mg/1. cycloheximide at different times after transfer toGA. Samples were fixed for histological studies at the time of addition of the inhibitorand at the end of the experimental period (4 days after addition of GA). Addition ofcycloheximide to spores after 6 h in GA completely prevented the breakdown ofstorage reserves, while introduction of the drug at later times permitted their pro-gressive hydrolysis. Thus it seems that there is no restricted period in the germinationtime course when proteins necessary for the hydrolysis of storage reserves are formedand that the production of hydrolases continues throughout the period of germination.This experiment also showed that cycloheximide given any time up to 12 h followingGA application inhibited initiation of the protonema in virtually all spores, suggestingthat proteins essential for protonemal initiation are synthesized subsequent to theaddition of cycloheximide. Protonemal initiation occurred normally if cycloheximide

Gibberellic acid-induced spore germination 91
was added between 24-36 h of GA treatment, although nuclear division delimiting theprotonemal cell was inhibited by the drug. Still later additions of the drug (36-48 hafter GA) permitted the occurrence of the first nuclear division while elongation of theprotonemal cell was arrested. Evidently, proteins for protonemal initiation, nucleardivision and cell elongation are synthesized in the same temporal sequence as theappearance of these morphogenetic landmarks in the germinating spore. Comparisonsof sections of spores collected at different times before and after application ofcycloheximide showed only slight differences in the degree of hydrolysis of storagereserves and in cell morphogenesis indicating that, following its administration to themedium, the drug penetrated the spores relatively rapidly.
Requirement for RNA synthesis
The possible origin of mRNA which codes for proteins for the hydrolysis ofstorage reserves and for cell morphogenesis during GA-induced germination wasinvestigated by the use of actinomycin D. The primary target of this compound isassumed to be mRNA, the synthesis of which is inhibited by binding DNA and thusblocking RNA polymerase (Reich, Franklin, Shatkin & Tatum, 1962), although incertain systems low concentrations of the drug may selectively inhibit rRNA synthesiswith relatively little effect on the synthesis of mRNA (Wilson & Hoagland, 1967;Sugano,Suda, Kawada & Sugano, 1971; Woodcock & Mansbridge, 1971). The rationalefor our experiment is that if a morphological or physiological process of germinationoccurs normally in the presence of actinomycin D, it is possible that the synthesis ofmRNA necessary for initiating this process has taken place before the drug was applied.When fully imbibed spores were incubated in GA and actinomycin D (ioomg/1.),they germinated to the same extent as in the control. A still higher concentration of thedrug (200 mg/1.) also did not appreciably inhibit germination of spores. Histologicalexamination of germinating spores revealed that hydrolysis of storage reserves,initiation of the protonema and the first nuclear division delimiting the protonemalcell were insensitive to high levels of actinomycin D, although as seen in Fig. 3,elongation of the protonemal cell was inhibited by the drug. In spores germinating inthe presence of 200 mg/1. actinomycin D, the nucleus occasionally underwent asecond division to form the rhizoid cell. These results taken in conjunction with thewidely accepted role of actinomycin D as an inhibitor of mRNA synthesis make itreasonable to speculate that proteins of early germination stages are encoded froma stable mRNA that is present in the dry spore in an inactive form. Presumably, thismRNA is not transcribed until the spore begins to germinate in the presence of GA.However, synthesis of actinomycin D-sensitive new mRNA appears necessary for theelongation of newly cut off protonemal cell.
The autoradiographic localization of the synthesis of nucleic acids and proteins inspores germinating in a medium containing actinomycin D as shown in Figs. 15-17supports this interpretation. In the presence of the drug, 60-h-old germinating sporesdid not incorporate any pHJuridine into RNA. In contrast, spores treated withactinomycin D showed normal accumulation of silver grains over the nuclei andcytoplasm when pHJleucine was used as the precursor, suggesting that protein

92 V. Raghavan
synthesis was unimpaired even when RNA synthesis was inhibited. The potentialability of the spore nucleus to divide in the presence of the drug was associated with amoderately intense incorporation of pffjthymidine. Clearly these data establish theexistence of active DNA- and protein-synthesizing machinery in spores germinatingin the presence of sufficient actinomycin D to stop RNA synthesis detectable byautoradiography.
Cordycepin effects
Since the above results suggested the existence of stored mRNA in dry, dormantspores, it was of interest to determine how mRNA is kept in an inactive form untilspores begin to germinate in the presence of GA. One obvious possibility is that theprocessing of mRNA, which involves post-transcriptional addition of adenylic acid
Table 2. Effect of addition of cordycepin (8-0 mgjl.) at different timesafter GA on germination
Time, h, after GA, ataddition of cordycepin
% germination...0
...0
2
0
4 i0 i
5 12
I 2 0
18
46
24
65
36
59
48
65
Control (no cordycepin)68
residues to the 3'-0H end of RNA molecules, is delayed until spores are subjected toGA action. This possibility was examined by testing the effects of cordycepin(3'-deoxyadenosine) on cell morphogenesis during germination and on the incorpora-tion of radioactive precursors of nucleic acid and protein synthesis. Cordycepin is ananalogue of adenosine, and is believed to affect post-transcriptional polyadenylationof mRNA (Darnell, Philipson, Wall & Adesnik, 1971). It was found that if cordycepin(8-o mg/1. or higher) and GA were added together to fully imbibed spores, there wasno germination at the end of 4 days. Sections of spores revealed almost no signs of GAaction beyond mild hydrolysis of storage protein granules (Fig. 4). If the concentrationof cordycepin used was 5-0 mg/1. a small percentage of spores germinated to formbarely visible protonemal initials; in sections, the first nuclear division appeared to beinhibited in such spores. At progressively lower concentrations of the inhibitor, anincreasing number of spores escaped from inhibition and germinated normally.Inhibition of germination induced by concentrations of cordycepin as high as 20-0 mg/1.was fully reversible upon transfer of spores to a medium lacking the inhibitor. Thisshows that the observed action of the inhibitor was not due to toxicity, and that thetreatment had no residual deleterious effects. It was also found that spores sown ina medium containing cordycepin did not incorporate radioactive precursors of DNA,RNA and protein. If cordycepin acts by interfering with the post-transcriptionalpolyadenylation of mRNA, these results suggest that processing of mRNA entailingpolyadenylation occurs upon incubation of spores in a medium containingGA and that perhaps cordycepin inhibits germination by inhibiting GA-inducedpolyadenylation.
In another experiment, cordycepin (8-o mg/1.) was administered to spores at

Gibberellic acid-induced spore germination 93
different times following the addition of GA and the percentages of germinationdetermined 4 days from the time of transfer to GA (Table 2). It was found that if thedrug was added any time up to 4 h after GA, no germination occurred. About 20%of spores escaped from inhibition if cordycepin was added up to 12 h after transfer toGA and cordycepin added 24 h or more after GA had no inhibitory effect on germina-tion. We have seen earlier that pretreatment of spores in GA for 24 h allows then-subsequent germination in the absence of the hormone. This minimum period ofhormone action required to trigger germination lies well within the period of sensiti-vity of spores to cordycepin when presumably GA-induced polyadenylation of mRNAis taking place. Moreover, the percentages of spore germination at different intervalsgiven in Table 2 correlate well with similar data (Table 1) obtained after pretreatmentof spores in GA for different periods.
5-Fluorouracil effects
The experiments with actinomycin D suggest that certain early morphogeneticevents of germination proceed normally in spores when the synthesis of putativemRNA is inhibited. This does not tell us anything about the involvement of othertypes of RNA in cell morphogenesis during germination. We considered the possi-bility that at some stage of germination spores might be making rRNA and tRNA;this was tested by studying the effect of 5-fluorouracil on germination. There isevidence in the literature that 5-fluorouracil, an analogue of uracil, can be incor-porated into all molecular species of RNA (Heidelberger, 1965), although some workon plant systems has indicated that it can preferentially block the synthesis of rRNAand tRNA when given at certain concentrations (Gressel & Galun, 1966; Key, 1966).It was found that addition of 5-fluorouracil at concentrations ranging from o-i to 50-0mg/1. did not affect the germination of spores which formed readily discernibleprotonemal initials. Here again, cytological observations of sections of spores areimportant. Whereas hydrolysis of storage reserves and enlargement of the protoplastproceeded normally in the presence of 5-fluorouracil (50-0 mg/1.), division of thespore nucleus was inhibited (Fig. 5). In very many spores, the nucleus failed tomigrate to the site from which the protonemal initial arose, or it migrated to a sitedifferent from the origin of the latter. In all cases, however, the nucleus appearedconspicuously swollen and highly granular with 1 or 2 prominent nucleoli. Sinceelongation of the protonemal initial was also inhibited in the presence of 5-fluorouracil,germinated spores appeared as abnormal uninucleate structures. From these results,it seems that a fraction of RNA which is apparently synthesized in the presence ofactinomycin D is sensitive to 5-fluorouracil and that the synthesis of 5-fluorouracil-sensitive RNA is necessary for initiating mitotic division delimiting the protonemal cell.Spores germinating in a medium containing 5-fluorouracil did not synthesize RNAas studied by incorporation of pHJadenosine (Figs. 10, 19), although incorporationof [3H]leucine, indicative of protein synthesis was nearly as heavy as in the control(Fig. 20). Moderate incorporation of [sH]thymidine into the spore nucleus was alsonoted, indicating that the block to division occurred in the post-synthetic period ofthe mitotic cycle (Fig. 18).
7 CEL 23
V. Raghavan

Gibber elite acid-induced spore germination 95
Requirement for DNA synthesis
During normal germination of the spore, enlargement of the protoplast is generallyfollowed by mitosis and cytokinesis. Our previous experiments have shown that thethe spore nucleus incorporates [3H]thymidine preparatory to mitosis indicating thatthe latter involves stimulation of DNA synthesis. To determine whether germinationcan proceed in the absence of DNA synthesis, effects of hydroxyurea on germinationwere studied. Several workers (Young & Hodas, 1964; Yarbro, Kennedy & Barnum,1965; Rosenkranz, Garro, Levy & Carr, 1966) have shown that this compound is anefficient inhibitor of DNA synthesis with little effect on RNA and protein synthesis.If hydroxyurea was added in a range of concentrations along with GA, spores germi-nated to the same extent as in a medium lacking the drug. Hydrolysis of storagereserves and enlargement of the protoplast into the protonemal initial occurrednormally even in spores incubated in a medium containing 120-0 mg/1. hydroxyurea,
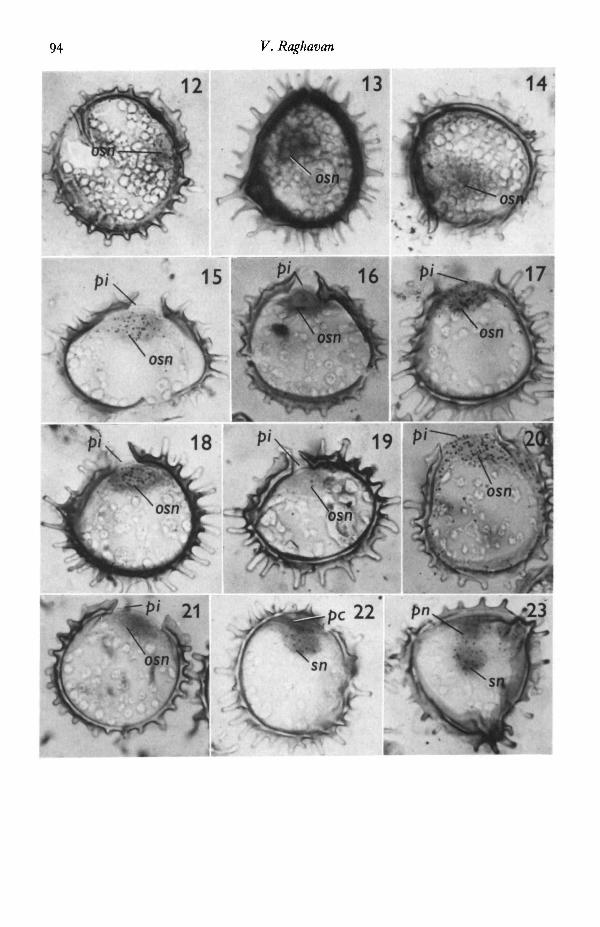
Figs. 12-23. Autoradiographs showing the effect of cycloheximide (05 mg/1.), actino-mycinD (200 mg/1.), 5-fluorouracil (50-0 mg/1.), hydroxyurea (120 mg/1.) and mannitol(o-6 M) on the incorporation of radioactive precursors of nucleic acids and proteininto 60-h-old germinating spores, x 950.
Fig. 12. PHjthymidine, cycloheximide.Fig. 13. [3H]uridine, cycloheximide.Fig. 14. [3H]leucine, cycloheximide. Spores shown in these figures have not germi-
nated. A few silver grains are seen over the nucleus in Fig. 12 but none are seen inFigs. 13, 14. The nuclei in the latter 2 figures appear black due to the uptake of stain.
Fig. 15. [sH]thymidine, actinomycin D. The spore has germinated to form the proto-nemal initial. Silver grains are seen mostly over the original spore nucleus and to someextent over the surrounding cytoplasm.
Fig. 16. PHJuridine, actinomycin D. The spore has germinated to form the proto-nemal initial. No incorporation of the isotope is seen.
Fig. 17. PH]leucine, actinomycin D. The spore has germinated to form the proto-nemal initial. Silver grains are seen over the original spore nucleus and the cytoplasmof the protonemal initial.
Fig. 18. [*H]thymidine, 5-fluorouracil. The spore has germinated to form the proto-nemal initial. Silver grains are mostly associated with the original spore nucleus.
Fig. 19. PHJadenosine, 5-nuorouracil. The exine is ruptured; the protonemal initialis barely visible. No incorporation of the isotope is seen.
Fig. 20. f/HJleucine, 5-fluorouracil. The spore has germinated to form the proto-nemal inital. Silver grains are seen over the original spore nucleus and the surroun-ding cytoplasm.
Fig. 21. [3H]thymidine, hydroxyurea. The spore has germinated to form the pro-tonemal initial. No incorporation of the isotope is seen. The nucleus is dark due to theuptake of stain.
Fig. 22. [3H]uridine, mannitol. Spore has not germinated, although mitosis of thespore nucleus, followed by cytokinesis, has resulted in formation of the protonemalcell within the confines of the exine. Labelling is mostly into the nuclear material.
Fig. 23. PHJleucine, mannitol. The spore has not germinated; the original sporenucleus has undergone mitosis without cytokinesis within the confines of the exine toform the nucleus of the potential protonemal cell. Silver grains are seen over both nuclei.
7-2

96 V. Raghavan
but division of the spore nucleus was inhibited (Fig. 6). Nuclei of spores germinatingin the presence of hydroxyurea appeared highly condensed and did not synthesizeDNA in spite of the availability of pHJthymidine in the medium (Fig. 21). AlthoughRNA synthesis was somewhat low in spores germinating in the presence of hydroxy-urea, incorporation of pHJleucine, however, appeared normal. Similar results werealso obtained with 5-fluorodeoxyuridine (20-0 mg/1.), another well known inhibitor ofDNA synthesis, but complete inhibition of incorporation of [3H]thymidine into thespore nucleus was not secured by this drug. The data suggest that the early morpho-genetic events of germination are independent of DNA synthesis in the spore nucleusand exclude the possiblity of GA action at the level of DNA synthesis in inducinggermination.
Role of cell enlargement
The first visible sign of spore germination is the breakage of exine by enlargementof the protoplast followed by division of the spore nucleus. In order to verify whethernuclear division in the normal sequence of germination is dependent upon the enlarge-ment of the protoplast, spores were allowed to germinate in medium supplementedwith GA and mannitol (o-6 M). Although there were no visually apparent signs ofgermination when examined at 4 days, sections of spores fixed in acrolein plus mannitoland processed in the usual way showed that the protoplast had divided into a proto-nemal cell within the confines of the spore coat even in the absence of any enlargement(Fig. 7). In addition, other GA-induced signs of germination such as hydrolysis ofstorage reserves, and migration of the nucleus had also occurred. These changes wereaccompanied by the synthesis of DNA, RNA and proteins as evidenced by theincorporation of radioactive precursors into germinating spores, but the levels ofradioactivity were lower than in controls (Figs. 22, 23). The latter is probably due toreduced uptake of precursors rather than to reduction in the effective sites of theirincorporation. It is clear that although the osmoticum prevents the protoplast fromovercoming the restraining force of the spore coat, nonetheless, cell morphogenesisduring germination is unaffected.
DISCUSSION
This study attempts to characterize the relationship between nucleic acid- andprotein-synthetic activities during early stages of GA-induced germination of spores ofA. phyllitidis and to present a speculative interpretation of the mechanism of hormoneaction in spore germination. The initial stages of germination studied are the enlarge-ment of the protoplast and its division to form the protonemal cell and the extensivehydrolysis of the storage protein granules that precedes cell morphogenesis.
The fact that spores germinate when DNA synthesis is inhibited, or divide whenenlargement of the protoplast is prevented, makes it unlikely that GA promotesgermination by acting primarily on the nucleus or on the cell wall. For this reason, andprincipally because the first detectable sign of GA action in germination is thehydrolysis of storage protein granules, considerable attention was given to this pheno-

Gibberellic acid-induced spore germination 97
menon in the present analysis. It was found that the breakdown of storage reserveswas a critical first step in the visible manifestation of GA action, since in its absencegermination did not occur. On the basis of the observed inhibition of hydrolysis ofstorage reserves by cycloheximide, one is led to conclude that this initial response ofspores to GA entails protein synthesis and that most of this protein synthesis is ofenzymes for hydrolysis of storage reserves. A host of other enzymes such as thoseinvolved in nucleic acid and cell wall biosynthesis are probably synthesized alsoduring the early hours of GA action, since addition of cycloheximide along with GAcompletely inhibits DNA and RNA synthesis and enlargement of the spore proto-plast associated with germination. Of course, there is the possibility demonstrated insome plants (Bennett, Smithers & Ward, 1964; Ross, 1968) that inhibition of nucleicacid synthesis by cycloheximide is a secondary effect of interference with proteinsynthesis. The reversible inhibition of GA-induced germination of Anemia spores bycycloheximide observed in this work is similar to that noted in the dark-germinatingspores of Pteridium aquilinum (Raghavan, 1970, 1971) and in the light-requiringspores of Onoclea sensibilis (Towill & Ikuma, 1975).
The finding that spores germinate normally to the stage of formation of a welldefined protonemal cell in the presence of high concentrations of actinomycin D is animportant lead to determine the origin of mRNA which presumably codes for thefirst proteins of germination. Assuming that actinomycin D operates in this system as aninhibitor of mRNA synthesis (Reich et al. 1962), this observation is suggestive of thepresence in the spore of a relatively stable mRNA which provides templates for thesynthesis of proteins for initiation of the germination process. This possibility takesinto consideration the fact that there is little reduction in autoradiographically detec-table protein synthesis in the spore during the period of action of actinomycin D. Inthe spores of Pteridium aquilinum, actinomycin D effects similar to those reported here,in conjunction with sedimentation data of labelled RNA, were used as evidence forthe existence of stable RNA species to code for proteins for the initiation of therhizoid and protonema (Raghavan, 1970, 1971).
Spores of Anemia germinate in the presence of high concentrations of 5-fluorouracil,although germination is not accompanied by division of the protoplast to form theprotonemal cell. This contrasts markedly with cell morphogenesis observed in sporesgerminating in the presence of actinomycin D. Autoradiography of pHJthymidineincorporation has shown that the block to cell division in spores treated with 5-fluorouracil specifically occurs in the post-synthetic period of the cell division cycle.Taken together the evidence lends considerable support to the view that cytokinesisin the dividing spore cell is dependent upon the synthesis of 5-fluorouracil-sensitiveRNA. Since 5-fluorouracil is known to inhibit the synthesis of rRNA and tRNA incertain plants (Gressel & Galun, 1966; Key, 1966), it may be that the same species ofRNA are also involved in the translation process for cytokinesis in germinatingspores. The contrasting effects of actinomycin D and 5-fluorouracil on cell morpho-genesis and the high concentrations of the former used in this study might rule outthe possibility of actinomycin D acting in our system as an inhibitor of rRNAsynthesis (Wilson & Hoagland, 1967; Sugano et al. 1971; Woodcock & Mansbridge,

98 V. Raghavan
1971). In terms of the extent to which spore germination can proceed using preformedmRNA, it seems that all phases of germination studied here excluding nucleardivision, but including DNA synthesis, may be attributed to the action of actinomycinD-insensitive mRNA present in the dry spore, and activated at the time of imbibitionin the presence of GA.
If the above reasoning is correct in all essential details, the critical role of GA inactivating stored mRNA is of primary significance in spore germination. It is nowestablished that most mRNA of plant (Higgins, Mercer & Goodwin, 1973; Manahan,App & Still, 1973; Van de Wall, 1973), animal and viral origin (Darnell et al. 1971;Edmonds, Vaughan & Nakazato, 1971; Lee, Mendecki & Brawerman, 1971; Adesnik,Salditt, Thomas & Darnell, 1972) contain poly-A sequences. Recently, it has beenshown that addition of GA promotes the synthesis of poly-A-containing RNA relativeto other species of RNA in isolated barley aleurone layers (Ho & Varner, 1974;Jacobsen & Zwar, 1974). It seems not unreasonable that in Anemia spores, processingof mRNA involving polyadenylation is delayed until they are incubated in the presenceof GA, which induces the synthesis of poly-A RNA. The observation that cordycepin,an inhibitor of polyadenylation, inhibits GA-induced germination is consistent withthis notion. It is also significant that the period of transcriptional control by cordycepinis short, comprising the first 24 h following GA addition, which incidentally is theminimum period of exposure of spores to the hormone required to induce germina-tion. Recently, Walbot, Capdevila & Dure (1974) have shown that germination ofcotton embryo and formation of the enzyme carboxypeptidase C in the germinatingembryo are inhibited by cordycepin, but not by high concentrations of actinomycin D.The results have been interpreted to suggest that formation of proteins for early germina-tion are encoded from mRNA stored in the dehydrated embryo and that polyadeny-lation is the control mechanism that activates this mRNA to provide templates forthe first proteins of germination. The actions of actinomycin D and cordycepin thusappear to be similar in the natural germination of the dormant seed embryo and theGA-induced germination of the dormant fern spore.
In arriving at this hypothesis on the mode of action of GA in the germination ofspores of A. phyllitidis, we have eliminated 2 other possibilities of cordycepin effect.One is that cordycepin does not enter the spores after 24 h, thereby allowing thehormone to exercise its germination-promoting function even in the presence of thedrug. In a more recent experiment, this possibility was tested by determining auto-radiographically the incorporation of [3H]uridine and [3H]leucine into sporesat 60 h with cordycepin added at 24 h following GA treatment. It was found thatcompared to the control, there was virtually a complete absence of incorporation of[3H]uridine into the nucleolus and nuclear material of cordycepin-treated spores,while the incorporation of [3H]leucine was unaffected. These results rule out thepossibility of non-entry of cordycepin at a certain time scale to germination as a factorin the release of spores from inhibition by the drug.
Inhibition of protein synthesis in cordycepin-treated spores raises the possibilitythat the drug, known to be a moderate inhibitor of protein synthesis in other systemsalso (Craig, 1973), might inhibit the process during the first 24 h of germination with

Gibberellic acid-induced spore germination 99
little effect thereafter. Since the proposed hypothesis predicts that proteins for earlygermination are translation products of pre-existing mRNA, cordycepin inhibition ofprotein synthesis is consistent with the notion that any treatment which blocks proces-sing of pre-existing mRNA will also block protein synthesis.
Because of limits of information provided by autoradiographic techniques tomeasure nucleic acid and protein synthesis and the effects of various inhibitors on them,the conclusions reached here are admittedly tentative. Moreover, inferences fromexperiments using inhibitors have to be treated with caution because of lack ofinformation on their effect on the uptake of radioactive precursors by spores, and onthe unknown effects of inhibitors in addition to those assumed to be the major effects.To verify further the postulation that spores of A. phyllitidis contain a preformedmRNA and that GA-induced polyadenylation is the regulatory step controlling theprocessing of this mRNA during germination, it is necessary to show: {a) that nomolecular species of RNA having messenger properties are synthesized during earlystages of GA-induced germination of spores; (b) that formation of RNA containingpoly-A segment is enhanced during GA-induced germination of spores; and (c) thatcordycepin inhibits the formation of poly-A. The single-celled nature of fern spores,uncomplicated by the presence of storage tissues such as endosperm or perisperm andthe presence of a relatively simple and well defined differentiation pattern duringgermination, make these investigations worthwhile to serve as a model system forGA-induced germination of seeds.
Thanks are due to Dr Alice F. Tryon for supplying spores of Anemia phyllitidis used in thiswork.
REFERENCES
ADESNIK, M., SALDITT, M., THOMAS, W. & DARNELL, J. E. (1972). Evidence that all messengerRNA molecules (except histone messenger RNA) contain poly(A) sequences and that thepoly(A) has a nuclear function. J. molec. Biol. 71, 21-30.
BENNETT, L. L. JR., SMITHERS, D. & WARD, C. T. (1964). Inhibition of DNA synthesis inmammalian cells by actidione. Biochim. biophys. Acta 87, 60—69.
CRAIG, N. (1973). The effects of inhibitors of RNA and DNA synthesis on protein synthesisand polysome levels in mouse L-cells. J. cell. Physiol. 82, 133-150.
DARNELL, J. E., PHILIPSON, L., WALL, R. & ADESNIK, M. (1971). Polyadenylic acid sequences :role in conversion of nuclear RNA into messenger RNA. Science, N.Y. 174, 507-510.
EDMONDS, M., VAUGHAN, M. H., JR. & NAKAZATO, H. (1971). Polyadenylic acid sequences in theheterogeneous nuclear RNA and rapidly-labeled polyribosomal RNA of HeLa cells: Possibleevidence for a precursor relationship. Proc. natn. Acad. Sci. U.S.A. 68, 1336-1340.
GRESSEL, J. & GALUN, E. (1966). Effect of 5-fluorouracil on the soluble RNA of Trichoderma.Biochem. biophys. Res. Commun. 24, 162-168.
HEIDELBERBER, C. (1965). Fluorinated pyrimidines. Progr. Nucleic Acid Res. molec. Biol. 4, 2-50.HIGGINS, T. J. V., MERCER, J. F. B. & GOODWIN, P. B. (1973). Poly(A) sequences in plant
polysomal RNA. Nature, New Biol. 246, 68-70.Ho, D. T. & VARNER, J. E. (1974). Hormonal control of messenger ribonucleic acid metabolism
in barley aleurone layers. Proc. natn. Acad. Sci. U.S.A. 71, 4783-4786.JACOBSEN, J. V. & ZWAR, J. A. (1974). Gibberellic acid causes increased synthesis of RNA
which contains poly(A) in barley aleurone tissues. Proc. natn. Acad. Sci. £7.5.^4.71,3290-3293.KEY, J. L. (1966). Effect of purine and pyrimidine analogues on growth and RNA metabolism
in the soybean hypocotyl - the selective action of 5-fluorouracil. PI. Physiol., Lancaster 41,1257-1264.

ioo V. Raghavan
LEE, S. Y., MENDECKI, J. & BRAWERMAN, G. (1971). A polynucleotide segment rich in adenylicacid in the rapidly-labeled polyribosomal RNA component of mouse sarcoma 180 ascitescells. Proc. natn. Acad. Sci. U.S.A. 68, 1331-1335.
L I N , S.-Y., MOSTELLER, R. D. & HARDESTY, B. (1966). The mechanism of sodium fluoride andcycloheximide inhibition of hemoglobin biosynthesis in the cell-free rericulocyte system.J. molec. Biol. 21, 51-69.
MANAHAN, C. O., APP , A. A. & STILL, C. C. (1973). The presence of polyadenylate sequencein the ribonucleic acid of a higher plant. Biochem. biophys. Res. Commun. 53, 588-595.
RAGHAVAN, V. (1970). Germination of bracken fern spores. Regulation of protein and RNAsynthesis during initiation and growth of the rhizoid. Expl Cell Res. 63, 341-352.
RAGHAVAN, V. (1971). Synthesis of protein and RNA for initiation and growth of the protonemaduring germination of bracken fern spore. Expl Cell Res. 65, 401-407.
RAGHAVAN, V. (1976). Gibberellic acid-induced germination of spores of Anemia phyllitidis:Nucleic acid and protein synthesis during germination. Am. J. Bot. 63, 960—972.
REICH, E., FRANKLIN, R. M., SHATKIN, A. J. & TATUM, E. L. (1962). Action of actinomycin Don animal cells and viruses. Proc. natn. Acad. Sci. U.S.A. 48, 1238-1245.
ROSENKRANZ, H. S., GARKO, A. J., LEVY, J. A. & CARR, H. S. (1966). Studies with hydroxyureaI. The reversible inhibition of bacterial DNA synthesis and the effect of hydroxyurea on thebactericidal action of streptomycin. Biochim. biophys. Ada 114, 501-515.
Ross, C. W. (1968). Influence for cycloheximide (actidione) upon pyrimidine nucleotidemetabolism and RNA synthesis in cocklebur leaf discs. Biochim. biophys. Acta 166, 40-47.
SCHRAUDOLF, H. (1962). Die Wirkung von Phytohormonen aus Keimung und Entwicklung vonProthallien. I. Ausl&sung der Antheridienbildung und Dunkelkeimung bei Schizaeaceendurch Gibberellinsaure. Biol. Zbl. 8i , 731-740.
SUGANO, H., SUDA, S., KAWADA, T. & SUGANO, I. (1971). Characterization of rapidly labeled40-S ribonucleoprotein particles in rat liver cytoplasm. Biochim. biophys. Acta 238, 139-149.
TOWILL, L. & IKUMA, H. (1975). Photocontrol of the germination of Onoclea spores III .Analysis of germination processes by means of cycloheximide. PI. Physiol., Lancaster 55,803-808.
VAN DE WALLE, C. (1973). Polyadenylic sequences in plant RNA. FEBS Letters, Amsterdam34, 31-34-
WALBOT, V., CAPDEVILA, A. & DURE, L. S., III . (1974). Action of 3'd adenosine (cordycepin) and3'd cytidine on the translation of the stored mRNA of cotton cotyledons. Biochem. biophys.Res. Commun. 60, 103-110
WEINBERG, E.S. & VOELLER, B.R. (1969). Induction of fern spore germination. Proc. natn. Acad.Sci. U.S.A. 64, 835-842.
WILSON, S. H. & HOAGLAND, M. B. (1967). Physiology of rat-liver polysomes. The stability ofmessenger ribonucleic acid and ribosomes. Biochem. J. 103, 556-566.
WOODCOCK, D. M. & MANSBRIDGE, J. N. (1971). Rapidly labeled ribonucleoprotein particles inrat liver cytoplasm and their relevance to the transport of messenger RNA. Biochim. biophys.Acta 240, 218-232.
YARBRO, J. W., KENNEDY, B. J. & BARNUM, C. P. (1965). Hydroxyurea inhibition of DNAsynthesis in ascites tumor. Proc. natn. Acad. Sci. U.S.A. 53, 1033-1035.
YOUNG, C. W. & HODAS, S. (1964). Hydroxyurea: Inhibitory effect on DNA metabolism.Science, N.Y. 146, 1172-1174.
(Received 25 June 1976)