the microbial profiling of the qawa...
TRANSCRIPT


THE MICROBIAL PROFILING OF THE QAWA RIVER
USING MOLECULAR METHODS
by
Awei Bainivalu
A thesis presented to the University of the South Pacific as
a partial fulfillment of the requirements for the degree of
MASTER OF SCIENCE
Copyright 2015© by Awei Bainivalu
School of Biological and Chemical Sciences
Faculty of Science Technology and Environment
The University of the South Pacific
Sept, 2015

DECLARATION
Statement by author
I, Awei Bainivalu, declare that this thesis is my own work and that, to the best of my
knowledge, it contains no material previously published, or substantially overlapping with
material submitted for the award of any other degree at any institution, except where due
acknowledgment is made in the text.
Name………………………………………………….
Student ID……………………………………………
Signature……………………………………………...
Date…………………………………………………...
Statement by Supervisor
The research in this thesis was performed under my supervision and to my knowledge is
the sole work of Ms. Awei Bainivalu.
Name………………………………………………….
Designation……………………………………………
Signature……………………………………………...
Date…………………………………………………...

DEDICATION
For my son, Epi and husband, Benjamin Delaimatuku
&
For my parents, Mr. Kelepi and Mrs. Lorna Bolatolu
My joy and rock, I will always be in your debt.

i
ACKNOWLEDGEMENT
“I keep my eyes always on the LORD. With him at my right hand, I will not be shaken”
(Psalms 16: 8)
I would like to thank Prof. Peter Lockhart, Ms. Trish McLenachan, Dr. Patrick Biggs
and Mr. Vinay Narayan for their valuable time, sound advice and assistance while
carrying out my research. Thank you for your support, I will always be grateful.
I am thankful to my brother in law, Epi Snr, for his assistance during my sampling
trips. Also, to my cousin, Unise for your hospitality and making me feel at home when
I am in Labasa for fieldwork.
I am grateful to Mr. Karuna Reddy of USP Research Office, for his time and assistance
with my statistical analysis.
My wonderful friends; Tams, Mike and Kalo, thank you guys for all your help and
always brightening up the room with your jokes and laughter.
I am indeed thankful to the School of Biological and Chemical Science’s technicians:
Dinesh, Mere and Siva for helping me with my laboratory work. A special thanks to
Siva, thank you for the extra effort you always put in for my research.
I would like to show my gratitude to the University of the South Pacific and Fijian
Affairs Board for funding this project. I also would like to thank Allan Wilson Centre
(Massey University), under the guidance of Peter and Trish, for the molecular analysis
of this work.
Finally, last but not the least, to my loving family, thank you for your endless support
and believing in me.
Malo Vakalevu

ii
ABSTRACT
The Qawa River situated in the North Eastern part of Vanua Levu near Labasa plays
a crucial role in the lives of people within the surrounding area. Many small
communities and villages along the Qawa River depend on the river for food, washing,
transportation, recreation and agriculture. South of Labasa, a Fiji Sugar Cooperation
(FSC) mill is situated along the river and discharge from the mill into the river has
been a long standing concern for the residents of Labasa and others living near the
river. Over a twelve month period (in April, August, December and March) dissolved
oxygen (DO) levels and temperature were measured at sites upstream and downstream
from the sugar mill. Water samples were also collected. Estimates of viable counts of
bacteria were made and DNA was extracted. The 16S rRNA V3 and V4 region was
sequenced on an Illumina MiSeq® platform and analyzed using QIIME software. DO
levels were similar between most sites at any one time, but changed dramatically over
the sampling period. They were lowest at the FSC mill site during the crushing season,
but also low at other sites (excluding Namoli) during this period. Total viable counts
for bacteria were greatest at the FSC mill site during the crushing season. QIIME
analysis showed that microbial populations were also similar between sites, but these
changed over time. It is presumed that high nutrient effluent during the crushing
season leads to significant algal and bacterial growth at FSC mill, and that this leads
to a high respiration demand which results in the observed dramatic lowering of DO
levels up and downstream of the FSC mill. DO levels show significant recovery three
months after crushing. However, during the crushing season, organisms of potential
health concern can be identified in the river. A suggestion has been made for future
monitoring of the river.

iii
TABLE OF CONTENTS
PAGES
Acknowledgement i
Abstract ii
Table of Contents iii-v
List of Figures vi-vii
List of Tables viii
Abbreviations ix-xi
Chapter 1: Introduction and Background Literature 1-26
1.1 Rivers and Human Settlement 1-4
1.1.2 Fiji Islands 4-6
1.2 The Qawa River in Vanua Levu Fiji 7-8
1.3 Impact of Human settlement on the health of Qawa River 9-10
1.3.1 Microbes as Natural Decomposers 10-13
1.4 The Sugar Industry in Fiji: its economic importance and
environmental impact on Fijian Rivers including the Qawa River 13-18
1.4.1 Impact of Sugar Mill Effluents from other Sugar Producing Countries 18-19
1.5 Estimating the relative abundance of micro-organisms 19-25
1.5.1 Culture Methods 19-22
1.5.1.1 Viable Cell Counting 22
1.5.2 Molecular Methods 22-25
1.6 Specific Aims of this thesis: Investigation of micro flora 26
of the Qawa River
Chapter 2: Materials and Methods 27-40
2.1 Sampling 27-28
2.2 DO and Temperature measurements 29
2.2.1 Statistical Inferences 29

iv
2.2.1.1 DO: Sampling Site 29
2.2.1.2 DO: Sampling Time 29-30
2.3 Total Viable Count (TVC) 30
2.3.1 Protocol for handling bacteria for TVC 30
2.3.2 Culturing for water samples for TVC 30
2.3.3 Statistical Inferences 31
2.3.3.1 Viable Counts: Sampling Site and Sampling Time 31
2.3.3.2 Correlation test for viable counts (log) and DO: sampling
site vs sampling time 31
2.4 Culturing of indicator species of potential concern from the Qawa River 31
2.4.1 Campylobacter sp. 31-33
2.5 DNA Extraction 33
2.5.1 Protocol for handling bacteria for DNA extraction 33
2.5.2 Isolation of DNA 33
2.5.2.1 Protocol for DNA extraction of bacteria from river water 33-35
2.5.2.2 Protocol for DNA extraction from bacterial cultures 35
2.6 MiSeq® sequencing of bacterial DNA from river water on the NZGL platform 35-38
2.6.1 QIIME bioinformatics pipeline 38-40
Chapter 3: Health of the Qawa River 41-58
Results 41
3.1 Physical Parameters 41
3.1.1 Temperature 41
3.1.2 Dissolved Oxygen (DO) 42
3.1.2.1 DO: Non Parametric Correlation Test
between sampling sites 43-44
3.1.2.2 DO: Non Parametric Correlation Analysis
between sampling times 44
3.2 Microbiological Parameters 45
3.2.1 Total Viable Counts at six sites over the sampling interval 45

v
3.3 Overall Correlation Log Count vs Overall DO 45
3.4 MiSeq® sequencing results 46 3.4.1 Rarefaction analysis 46-47
3.4.2 Microbial profiles upstream vs downstream for
each sampling time 47-49
3.4.3 Taxa Present 49-50
3.5 Culturing of Indicator species and confirmation of identity 50-51
Discussion 52-58
3.6 Qawa River Temperature and DO 51
3.6.1 Temperature 51-52
3.6.2 Dissolved Oxygen 52-53
3.7 Rarefaction Analysis 53-54
3.8 Microbial compositions mirror changes in river DO 54-55
3.9 Microorganisms of concern in the river 55-57
3.9.1 Viable Counts 57-58
Chapter 4: Future Work and Monitoring of Qawa River 59-61
4.1 Comprehensive assessment 59-60
4.1.1 LAMP surveys 60-61
References 62-78
Appendices 79-124

vi
LIST OF FIGURES
PAGES
Figure 1.1: The hydrological cycle. Arrows indicate direction of
water movement 1
Figure 1.2: Main islands, rivers and towns of the Fiji Islands 6
Figure 1.3: Showing the Labasa and Qawa River 7
Figure 1.4: Rainfall classes for the Fiji Islands 9
Figure 1.5: The FSC mill situated on the Qawa River 13
Figure 2.1: Sampling sites along the Qawa River 27
Figure 2.2a: Average monthly water temperature over the year in Vanua Levu 28 Figure 2.2b: Average monthly precipitation over the year in Vanua Levu 29
Figure 2.3: Serial dilution and plating scheme 30
Figure 2.4: Enrichment step for Campylobacter using Bolton broth 33
Figure 2.5: Membrane filtration equipment
Figure 2.6: Secondary structure of the 16S rRNA of bacteria indicating
34
variable V1-V9 regions 37
Figure 2.7: QIIME Bioinformatics pipeline
Figure 3.1: Water temperature (°C) measured at six sites on the Qawa River
40
before, during and following the sugar cane crushing season 41
Figure 3.2: DO levels at six different sampling sites 42
Figure 3.3: Total viable counts for the six sampling sites 45
Figure 3.4: Rarefaction plot for observed species vs time 46
Figure 3.5: Rarefaction plot for phylogenetic diversity vs time 47

vii
Figure 3.6: PCoA plot of microbial profile differences when
color coded for sampling times 48
Figure 3.7: PCoA plot of microbial profile differences when
color coded for location 49
Figure 3.8: Cultured Campylobacter sp. from the Qawa River 51

viii
LIST OF TABLES
PAGES
Table 1.1: Waste produced at different process stages 15
Table 2.1: Sampling locations on the Qawa River 28
Table 3.1: Results of correlation tests for DO values at four sampling
times between sampling sites 43
Table 3.2: Results of correlation test for DO values at the six sites
between sampling times 44
Table 3.3: List of identified taxa from the Qawa River 50
Table 3.4: Confirmation result for Campylobacter sp. 51
Table 3.5: Critical level of DO (mg/L) in river water 54

ix
ABBREVIATIONS
BOD: Biological Oxygen Demand
Bolton Broth: Enrichment broth for Campylobacter sp.
bp: Base Pairs
Bst: Bacillus stearothermophilus
CFU: Colony Forming Unit
COD: Chemical Oxygen Demand
CTAB: Cetyl Trimethyl Ammonium Bromide
DGGE: Denaturing Gradient Gel Electrophoresis
DO: Dissolved Oxygen
DOC: Dissolved Organic Carbon
EDTA: Ethylenediaminetetraacetic acid
FEA: Fiji Electricity Authority
FISH: Fluorescence In situ Hybridization
FSC: Fiji Sugar Co operation
GDP: Gross Domestic Product
GPS: Global Positioning System
LAMP: Loop-mediated Amplification
McCartney Bottles: strong clear glass bottle with wide
mouth and aluminum screw cap with rubber liner
mCCDA: modified Charcoal-Cefoprazone-Deoxycholate
Agar
mRNA: messenger Ribonucleic Acid

x
MiSeq®: desktop sequencer that can produce 2x300
paired-end reads in a single run
NA: Nutrient Agar
NGS: Next Generation Sequencing
NZGL: New Zealand Genomics Limited OTU: Operational Taxonomic Unit
PCoA: Principal Coordinates Analysis
PCR: Polymerase Chain Reaction
ProK: Proteinase K
PVP: Polyvinylpyrrolidone
PWD: Public Works Department
QIIME: Quantitative Insights Into Microbial
Ecology
rpm: Revolution per minute
rRNA: ribosomal Ribonucleic Acid
SDS: Sodium Dodecyl Sulfate
SLB: Sucrose Lysis Buffer
SPSS: Statistical Packages for Social Sciences
SS: Suspended Solids
TDS: Total Dissolved Solids
TE: Tris EDTA Buffer
TVC: Total Viable Counts
USP: University of the South Pacific

xi
V1 toV9: Variable 1 to Variable 9 Region
VBNC: Viable but non culturable
WHO: World Health Organization
YSI Model 85: A handheld multimeter that measures DO, Conductivity, Salinity and Temperature system

1
CHAPTER 1
INTRODUCTION AND BACKGROUND LITERATURE
1.1 Rivers and Human Settlement
Water is the driver of nature and a component of all living things. Without water, life
could not exist. Freshwater accounts for only 2.5 % of the Earth’s surface water and
river water constitutes only 0.0001 % of the total volume of water (Maidment, 1993;
Shiklomanov & Rodda, 2003).
Figure 1.1: The hydrological cycle (Source: http://what-when-how.com/water-
science/hydrologic-cycle-water-science/). Arrows indicate direction of water
movement.
Rivers are terrestrial flowing surface waters that drain distinct watersheds. They
provide a natural water course, usually freshwater, that drains into an ocean or lake,
or other body of water. Rivers form part of the hydrologic cycle which provides a

2
continuous circulation of water between the earth and its atmosphere as shown in
Figure 1.1.
With the sun’s heat, the hydrologic cycle begins with the evaporation of free water to
the atmosphere. Moist air is lifted along with water from evapotranspiration, which is
water transpired from plants and evaporated from the soil. Cold air in the atmosphere
cools the water vapor and causes it to condense into clouds. Water is then returned to
the earth’s surface by precipitation and this either becomes groundwater that can leak
its way into oceans, rivers and streams or alternatively be evaporated from the soil. It
can also become ground surface water, and eventually gather in terrestrial water bodies
and finally into the ocean where the cycle begins again through evaporation (Figure
1.1).
Typically, a river has a geographic origin, known as its headwaters. There are smaller
rivers or streams that merge into the main river; such contributing streams are called
tributaries of the principal river. Thus, rivers are the sum of its tributaries appropriately
termed river systems rather than single rivers. According to Jordaan (2013), rivers can
be of three kinds:
• linear or meandering: rivers which are characterized by one main stream, with lesser
contributing tributaries, following a mildly curving path from source to mouth.
• dendritic: rivers having numerous contributing tributaries. • compound systems: rivers which are meandering and shifting.
These interconnected networks of natural streams, channels and rivers provide
humanity with one of the utmost and valuable natural resources (Arin et al., 2014).
Although rivers and other forms of surface water only account for a small portion of
water available on the planet, they appear large in the human imagination as the result
of their impact on our lives.
Human civilization was born on river banks. The first human civilizations developed
along rivers in Egypt, Mesopotamia, India and China and today many a great city lies

3
along a river. River water is vital for human existence and has been the mainstay for
nearly all human settlements for generations (Liang & Ding, 2004; Nguyen, 2006).
According to Varis et al. (2012) about one-quarter of the world population lives in the
ten largest river basins (from Indus River to Yellow River) of the Monsoon Asia-
Pacific region. Most of these areas have a fast growing population, particularly in
South Asia where the pace of urbanization has extended across a substantial area of
all the basins.
Developments of many major cities, towns and settlements in the world have been
centered with close proximity to rivers (Paul & Meyer, 2001; Kumar et al., 2013).
River water used by communities can be broadly categorized as being for consumptive
and non-consumptive purposes.
Consumptive uses
River waters are used in domestic activities, agriculture, irrigation and industry. In
addition to this, river water carries and transports organisms and important gases and
nutrients to many areas (Erah et al., 2002; Araya et al., 2003). Furthermore, it affects
the global carbon cycle as river waters play an important role in oxidizing, storage and
the release of terrestrial carbon (Downing et al., 2006; Cole et al., 2007). Naiman &
Décamps (1997) documented that in addition to supporting fish and aquatic
ecosystems, rivers sustain vegetation in riparian or floodplain ecosystems. Rivers also
drain water, shaping the features of the Earth as it make their way to the sea (Jordaan,
2013).
Non-consumptive purposes
People use rivers as travel routes, for recreation and for providing power for
hydroelectric plants and the electrical energy we use in our everyday lives. Rivers
also provide an easy means of disposing of waste-water and, in low income situations,
other wastes (Ghosh & McBean, 1998; Kanu & Achi, 2011; Okoko et al., 2012).
Rivers can also be a source of human despair, predominantly during floods when they
break their banks and cause substantial damage to properties and even cause death

4
(Jonkman & Kelman, 2005; Hartvich et al., 2007; Yeo et al., 2007; Lal et al., 2009;
Kumar et al., 2013). Despite its central importance for human existence, 1.2 billion
people worldwide are at risk of not having access to water (UNEP, 2003).
1.1.2 Fiji Islands
In the Fiji Islands, like elsewhere in the Pacific, freshwater resources can be limiting.
In some locations there are large river overflows. In others, there are no surface water
resources and people rely solely on rainwater for their potable and economic needs
(SPC SOPAC, 2012).
As an archipelago of the Pacific, the Fiji Islands comprise approximately 320 islands
of varying sizes. Lying roughly 177 ° E and 178 ° W Longitude and 12 ° and 22 ° S
Latitude, the islands encompass a region of nearly 1.3 million km² with a land area of
18,700 km2, of which only one-third are inhabited (Fiji Department of Energy, 2010).
While all types of oceanic islands are found in the Fiji group, the largest islands (Viti
Levu and Vanua Levu), are of volcanic origin (Gray, 1989) and are described as “high
islands”. Their interiors are mountainous which limits the area of land suitable for
development to mostly river banks and coastal areas (Chandra & Mason, 1998). Fiji
has its most extensive natural water resources and abundance of freshwater on its
larger islands, where annual rainfalls range from 2,000 mm to 6,000 mm fall in the
mountain catchments. This rainfall provides the source of water for Fiji’s diverse river
systems. These sources range from minor mountain streams and steep torrents, to very
large mature rivers in the lowlands. They are meandering between flood plains,
lagoons and oceans deltas generally sheltered by a fringing coral reef. Surface water
is used as the main source of water supply for all cities and major towns on the larger,
high islands of Fiji. This source is used both for drinking as well as for industry (SPC
SOPAC, 2012).
With many permanent rivers and streams, the larger islands are well watered, however
only Viti Levu has rivers of any considerable size. There are about 50 rivers on this
island and these feed three large river systems, the Rewa (130 km), Navua and

5
Sigatoka, which enter the sea along the south coast. The Rewa River has the largest
catchment area of these and covers nearly one third of the island. Occupying the drier
parts of Viti Levu are Fiji’s two most economically important rivers, the Ba and Nadi
Rivers, which have a combined catchment of only 15 % of Viti Levu (Gray, 1989).
For Vanua Levu, the Dreketi River (55 km) is the only river of any considerable size
with respect to 40 rivers present. (Fiji Department of Energy, 2010) (Figure 1.2).

6
Figu
re 1
.2:M
ain
isla
nds,
river
s and
tow
ns o
f the
Fiji
Isla
nds.
(Sou
rce:
McG
ree
et a
l ., 2
010)
.
N.B
: Blu
e ar
row
indi
cate
s are
a
whe
re th
e st
udy
took
pla
ce.

7
1.2 The Qawa River in Vanua Levu Fiji
Vanua Levu has a number of rivers, including the Labasa, the Wailevu and the Qawa
River from the Macuata province. These three form a delta on which the town of
Labasa stands. Situated on the outskirts of Labasa Town, the Qawa and the Labasa
River are connected by an 8 kilometer canal which helps drain the fertile Labasa plains
and adjacent foothills (Gray, 1989).
The Qawa River has a running length of approximately 33.5 km and drains an area of
126 km2 (Yeo, 2001). It is relatively shallow and not navigable by any large vessels.
The use of this river for commercial shipping purposes has declined since the
commissioning of Port Malau (Labasa’s main port) to its current status where the river
is only used as an access for fishing boats and small ferry services to the upper Macuata
coast and its offshore islands.
Figure 1.3: The Labasa and Qawa River (Source: Google Maps, 2013)
Together with the Labasa River, these rivers host one of the five largest stands of
mangroves in Fiji which occur in deltaic formations at the river mouth (Gray, 1989).

8
Mangroves provide breeding and nursery grounds for a wide variety of economically
important fish and crustaceans as well as other aquatic organisms. Mangroves also
help prevent soil erosion by stabilizing sediments with their tangled roots. Furthermore
they maintain water quality and clarity by filtering pollutants, they also play an
important role in the Labasa sewage treatment program (Fiji Department of
Environment, 1999).
Like any other river, the Qawa River plays a crucial role in the lives of people within
the surrounding area. Many small communities and villages that are situated along the
Qawa River depend on the river for food, washing, transportation, recreation and
agricultural purposes. The river is especially used during the period of low/no tap
water supply.
Labasa, in the North Eastern part of Vanua Levu has a population of almost 28,000
according to a recent 2007 census (Fiji Bureau of Statistics, 2013). Labasa is a major
center of Fiji's sugar industry. In contrast to Vanua Levu's windward, southern coast,
it faces the prevailing trade winds and therefore the main mountain ranges which
produce significant rainfall. In general terms, the northern region of Vanua Levu is
dry for six months of the year (Figure 1.4) with an uneven distribution of rainy days.
Rainfall seasonality is more pronounced for the leeward northern side of the high
islands, which only receives 20 % of the annual total in the dry month. This is why
sugarcane, the island’s major crop, thrives in this northern parts of Vanua Levu.
The average annual rainfall of the area around Labasa is between 2,000 to 2,500
mm/yr., with a six month wet season extending between November to April. During
the latter period, 80 % of the annual rainfall is precipitated. There is only slight
temperature variation throughout the year. The monthly average temperature can
range from 24 to 27 degrees Celsius (°C) and have an annual mean of 25.6 °C (Ledua
et al., 1996).

9
1.3 Impact of Human settlement on health of Qawa River
The water from rivers and fresh water lakes plays a significant role in overall
development programs of any country, as a source of water supply for domestic and
industrial purposes, for agriculture, fishery and power generation.
Figure 1.4: Rainfall classes for the Fiji Islands (Source: Atherton et al., 2005)
However, the water resources are also utilized for the disposal of industrial wastes and
sewage, leading to water pollution. Human activities related to urbanization,
industrialization and agriculture have adverse effects on water quality due to various
types of pollutants (Paerl et al., 2003; Mokbel & Yamakanamardi, 2008; Hamuda &
Patkó, 2011; Kipsang et al., 2014). Rivers and their ecosystems are threatened
throughout the world and may be rendered unfit for the use of humans and other biota

10
as a direct result of activities that alter the physical, chemical and biological nature of
the receiving water bodies (Sangodoyin, 1991; Hammer, 1992; Fausch et al., 2002).
1.3.1 Microbes as Natural Decomposers
An important component facilitating ecosystem processes is the bacterial community.
Microbes are natural and vital members of all aquatic communities as they are among
the most vital contributors to the conversion of complex organic compounds and
minerals in the aquatic environment (Fischer et al., 2002; Azam & Worden, 2004;
Handelsman, 2004).
The bacterial biomass is also an important part of natural river systems. Due to low
primary productivity and a high ratio of river bank length to water volume,
allochthonous organic material is commonly the main source of organic matter for the
food web in such environments. Heterotrophic bacteria assimilate this organic matter
with protozoa and metazoa making them key organisms in the base food chain
(Legendre & Rivkin, 2008).
Therefore, not only are these bacteria decomposers as earlier adopted in classical
ecological concepts by Odum (1956), playing a key role in the reduction of organic
matters and the reminerilization (Muylaert et al., 2002) of nutrients; their biomass is
a main food supply to higher trophic levels through the microbial food web in rivers
(Pomeroy, 1980). Azam et al. (1983) and Meyer (1994) called this pathway the
microbial loop, which profoundly increases the productivity of the whole aquatic
system (Lindstrom, 2001).
Bacteria are the most important consumers of the organic carbon entrained in running
waters and thus they play a significant role in the aquatic carbon cycle. They
metabolize detritus arising from the death of higher organisms and organic wastes
from excretions (Azam & Cho, 1987). According to Moran & Hodson (1990),
dissolved organic carbon (DOC) supporting bacterial metabolism originates either
from in situ primary production or from external terrestrial inputs.

11
Like all ecosystems, freshwater ecosystems require energy inputs to sustain the
organisms within, thus consumption of DOC by bacteria is one of the major pathways
of material and energy flow in pelagic food webs (Azam et al., 1983; Meyer, 1994;
Cole, 1999). Degradation of this organic matter contributes to the purification of the
ecosystem and is, therefore, a major process controlling water quality. These bacteria
are responsible for much of the respiration in large rivers, and they may affect the
quantity and quality of matter transported by large rivers to the oceans (Benner et al.,
1995; Castillo et al., 2004).
However, pathogenic microbes may also increase due to pollution which may pose
health risks to humans (Danielopol et al., 2003).
Disposal of wastes into water by humans has been practiced since the earliest
civilizations, mainly because it is cheap and an easy way to rid society’s wastes. Thus
far, water bodies are often used as a dumping ground for wastes such as industrial
effluents, raw sewage, garbage and oil spills (Gafny et al., 2000; Obi et al., 2002;
Singh et al., 2007; Abraham, 2011). Water quality is closely linked to water use and
to the state of economic development and physical health of communities (Manhokwe
et al., 2013).
The deteriorating quality of the receiving waters has substantial implication both in
the immediate situation and over the long term (Ghosh & McBean, 1998). It has been
estimated that nearly 1.5 billion people lack safe drinking water globally, and that at
least 5 million deaths per year can be attributed to waterborne diseases (Onsdorff,
1996).
Most industrial activities involve water intensive processes and consequently
discharge effluent into the water bodies in large amounts. Such processes contaminate
many rivers in Fiji. The Qawa River in particular has been in the spotlight for more
than 20 years due to high pollution content. It is perhaps one of the most polluted rivers
in the South Pacific (Karan, 2010). A number of reports (Lee, 1981; Tamata et al.,

12
1996; Fung & Chand, 1997) have indicated that the Fiji Sugar Cooperation (FSC) mill,
situated along the river, is the main contributor aggravating pollution problems on the
Qawa River (Figure 1.5).
The nature of the Qawa River (slow-moving and meandering) at the discharge point
used by the sugar mill exacerbates the problem of pollution. The depression found in
the river bottom close to the discharge point causes the high density, organic rich
effluent to sink, deoxygenating the river. This inactivity of the river has been
problematic for the local residents particularly during the "dry" season when the river
flow is particularly sluggish (Lee, 1981). Unfortunately, this also corresponds to the
crushing season (June to November) when significant amounts of carbon-rich effluent
were discharged into the river.
.
The status of Qawa River has been a long standing concern for the residents of Labasa.
Its uncharacteristic colour, intolerable stench and depletion of fish resources, death
and loss of marine life, loss of recreational activities and water sports, and loss of
subsistence livelihoods and economic activities based on the river have all been
attributed to the FSC mill which discharges its wastes into the lower reaches of the
river (Tamata et al., 1996).

13
Figure 1.5: The FSC mill situated on the Qawa River ©Awei Bainivalu
In addition, surveys conducted at a school and among communities located along the
Qawa River have revealed reports of dizziness, nausea and headache (Karan, 2010).
Health related issues continue to be a major concern especially during the crushing
season of the sugar mill.
1.4 The Sugar Industry in Fiji: its economic importance and environmental impact
on Fijian Rivers including the Qawa River.
The sugar industry has a substantial economic standing in many countries. In Fiji, the
sugar industry is the second largest industry, previously perceived as the backbone of
the Fijian economy, given its contributions to gross domestic product (GDP) and
employment generation. The FSC is government owned and also the largest public
enterprise in the country employing nearly 3,000 people while 200,000 or more
depend on it for their livelihood in rural sugarcane belts of Fiji. The industry still
provides a livelihood for a large part of the population and remains one of the major
earners of foreign exchange. However, despite its significance, the sugar industry also

14
plays a major role in producing a higher amount of water pollution compared to other
industries.
Sugarcane cultivation and production is thought by some to have a damaging impact
on rivers and coral reefs. This includes the Qawa River in Labasa and the Great Sea
Reef of Vanua Levu, which is the third longest barrier reef system in the Southern
Hemisphere. Not only biodiversity of these river systems and coastal areas is
threatened, cultures and livelihoods of communities that rely on these natural
resources are also at risk due to chemicals used on sugar cultivation and wastes from
sugar production (WWF, 2013).
Wastewater with varying levels of pollution load is usually generated at nearly all
stages of sugar production (Table: 1.1). Sugar cane industries are known to be water
intensive industries; meaning that they utilize large amounts of fresh water for the
production process.
The wastewater effluents generated by the sugar cane industries are characterized by
high levels of BOD, Chemical Oxygen Demand (COD), Suspended Solids (SS), pH
and Total Dissolved Solids (TDS) (Ali & Soltan, 1996; Salequzzaman et al., 2008).
Effluents with high BOD when untreated or partially treated can result in aquatic
pollution (Kaur et al., 2010). High BOD means high levels of organic matter will be
available to bacteria as a food source resulting in expansion of bacterial populations
in the water. As oxygen is needed by microbes to successfully decompose the high
levels of organic matter, available Dissolved Oxygen (DO) in the water can be
depleted. On complete exhaustion of the oxygen, the bacteria begin anaerobic
respiration to breakdown (oxidize) organic material in the water. To do this they
reduce sulfates so that the decomposition process can be continued. This reduction
produces a foul smelling Dihydrogensulfide gas (H2S), which in turn can precipitate
iron and other dissolved metals, turning the water black and making it highly toxic for
aquatic life.

15
High COD is a measure of the inorganic and partly organic non-biodegradable content
of the effluents. This is another aspect of concern in untreated wastewater since it has
effects on the receiving water body similar to that of high BOD receiving water bodies
(Akbar & Khwaja, 2006)
Table 1.1: Waste produced at different process stages (Source: Santos, 2008)
Process Stage Main Inputs Wastes and By-Products
Mill House Sugarcane Wastewater containing suspended solids and oil
content Washing from floor cleaning containing sugar Bagasse
Process House Sugar Juice Washing of different components such as
evaporators, juice heater, vacuum pan, clarifiers
etc., generates aggressive effluents with high BOD, COD and TDS concentrations
Boiler House Bagasse and Furnace oil Wastewaters from scrubs
Cooling Pond Water and Chemicals Wastewater
Sugar cane industrial wastewater is also known to contain considerable levels of
suspended solids (SS). Although not considered a human health hazard, suspended
solids can lead to undesirable water quality conditions (Ali & Ahmad, 1993; Baruah
et al., 1993; Singh et al., 1996; Pawar et al., 1998; Thuresson, 2001). Suspended solids
can cloud or reduce light penetration which in turn, has an adverse effect on fish
(clogging gills) and other aquatic life by reducing photosynthesis which ultimately
reduces oxygen available to them in water. Suspended solids may also contain
contaminants, such as nutrients, organic matter, pesticides and heavy metals (Devlin
& McVay, 2001). Cheesman, (2005) mentioned that waste water from some sections
of the sugar mill contains a considerable concentration of suspended solids, which
cause blockage in drainage and ditches. These also increase pollution impacts because
of the slow decomposition of the settled matter.

16
Sugar mill effluents generally are not considered to have high enough levels of total
dissolved solids (TDS) to cause major environmental impact. However, discharge of
wastewater with high TDS level has the potential to have an adverse impact on aquatic
life, making the receiving water unfit for human consumption, domestic use and
irrigation purposes (Singh, 2000).
Sugar cane effluent also increases the color, temperature and pH of aquatic
environments resulting in pollution problems. Colored waters are not only visually
unpleasant, they reduce light penetration. This will result in inefficient photosynthesis
at deeper levels in rivers and consequently a decrease in the dissolved oxygen content
of the water as algal growth is reduced.
Furthermore, sugar mill effluent also has an impact on plant communities in affected
habitats. Increased nutrients will stimulate the growth of some species more than
others, changing the ecological balance (Ali & Soltan, 1996; Arindam, 1999). Borhidi
et al. (1986) and Singh et al. (1998) have reported thick mats of weeds, macrophytes
and submerged aquatic plants that blocked canals as a result of waste water from sugar
mills. Reports also note single fauna and flora species becoming dominant in most
polluted river sites (Ali & Soltan, 1996). Furthermore, while algal growth might
initially lead to an increase in DO, the increasing respiration demands of algal blooms
and bacteria can subsequently cause DO levels to crash, leading to oxygen starvation
for aquatic life. Effluents with high temperatures are also considered responsible for
depleting DO levels in the water as oxygen molecules are less soluble at high
temperature (Baskaran et al., 2009).
Sugar mill effluents generally change the natural pH level of the receiving water body
to some extent, either making it too acidic or alkaline (Robinson, 1990). Excessive
acidity can result in the release of Dihydrogensulfide to the air (Akbar & Khwaja,
2006). Also waste waters have a high salt concentration and are toxic to aquatic life
when discharged into a water course (Kumar & Chopra, 2010). Consequently it has

17
been recommended that wastewaters from sugar mills must be treated properly to
reduce its organic load before disposal into river systems (Akbar & Khwaja, 2006;
Yadav & Pathak, 2012).
In Fiji, sugar cane is processed at four mills namely Lautoka, Rarawai at Ba, Penang
and Labasa. All mills are situated near rivers or a water source. The cooperation has
effluent digestion treatment ponds at the Rarawai, Penang and Labasa mills. Spill
overs from the production processes, as well as pollutants generated from
manufacturing, are directed to these treatment ponds (Fagan et al., 1995), where they
undergo biodegradation. The Lautoka mill has no suitable land to accommodate such
ponds so effluents are discharged at sea. It is supposed that, because the mixing effect
is greater in deep sea than in shallower waters of the foreshore, the impact on marine
life is minimal (Lodhia, 1999).
There is a need to evaluate this assumption and also the FSC sugar mill’s full impact
on the Lautoka marine ecosystem. This is because the discharged effluent is high in
BOD and high in temperature. These features are expected to produce considerable
reduction in dissolved oxygen levels in the receiving water for the six months of the
year when the sugar mill is in operation from June to November (Watling & Chape,
1992).
Tamata & Lloyd (1994) reported high BOD levels resulting from effluents from the
Rarawai mill, located on the banks of Ba River into which effluent is discharged. Both
the Rarawai mill and Labasa mill installed effluent treatment systems consisting of
primary and secondary treatment; however both the Rarawai and Labasa mills showed
high BOD levels indicating treatment systems were not performing to expected
standards. Anderson & Lloyd (1995) mentioned that this could be due to input loading
frequently surpassing design value, leading to anaerobic conditions in ponds. As for
the Ba River, Fagan et al. (1995) reported very low DO at sites close to the discharge
point during the crushing season of Rarawai mill. DO levels in the river downstream
from the mill were also below that necessary to maintain healthy aquatic life (Tamata
& Lloyd, 1994).

18
Reports on water quality in the Qawa River, Labasa during the crushing season have
shown higher water temperature and BOD, and lower DO, than the levels permissible
near the FSC mill discharge outlet, a main site of pollution in this river (Tamata et al.,
1996).The high BOD levels and low DO levels there have been suggested as the cause
of fish kills and loss of aquatic life (Watling, 1985; Fagan et al., 1995). The evident
decrease of mangrove area fringing the river banks has been suggested by Fung &
Chand (1997) as being due to the presence of bagasse (fibrous matter that remains
after sugarcane or sorghum stalks are crushed) on the river side, which kills mangroves
by covering their breathing roots. Water temperature is fairly constant with depth for
the Ba and Qawa River, as the river is relatively shallow. In addition, the uniform
temperature is also attributed to water turbulence generated when large volumes of
water flow during coming tides.
1.4.1 Impact of Sugar Mill Effluents from other Sugar Producing Countries
Reports indicate that sugar mill effluents from sugar producing countries all have
exceedingly high levels of BOD, COD, SS and low levels of DO (Cheesman, 2005;
Carminati, 2008; Usharanji et al., 2010; Saranraj & Stella, 2014). As previously
mentioned, under these conditions oxygen supply in the receiving water bodies is
depleted causing detrimental impact on aquatic life and its ecosystem. The high
organic load in sugar mill wastewaters that leads to water pollution brings about
changes that can overturn the ecological balance of the aquatic system. Akali et al.
(2011) investigated the pollution of the River Nzoia, Kenya from sugar mill effluent
and found that BOD, COD and temperature were higher than the permissible limits of
the World Health Organization (WHO). Similar conclusions were also drawn by
Salequzzaman et al. (2008). These authors have reported that sugar mills in
Bangladesh do not also maintain the WHO standard qualities.
Similar findings have also been made by Akbar & Khwaja (2006), who noted that
wastewater from sugar mills in Pakistan also exceed safe limits. Galindo et al. (2001)
reported the severe organic contamination mainly from sugarcane processing in the
Sali River, Argentina. Lopez-Lopez et al. (2003) determined that high levels of

19
pollution from sugar industry effluents in Mexico may threaten the survival of the
native fish population of the America basin. Corcoran et al. (2010) reported that annual
cleaning of sugar mills in Santa Cruz, Bolivia and the pollution of Danish coastal
waters by sugar factory effluents resulted in the deaths of millions of fish in local rivers
and the occurrence of bacterial pathogens respectively. Furthermore the report also
stated that damages to the Great Barrier Reef, Australia were attributed to waste water
from sugar mills that have altered flow of freshwater into the sea carrying high levels
of nutrients, reducing water quality and impacting on onshore reefs (WWF, 2013).
The impact of the sugar cane industry on the river system has not yet been
systematically studied in Fiji using modern microbiological methods. Such modern
methods involve DNA extraction of bacteria from river water samples, high
throughput sequencing of DNA fragments and assignment of the sequenced DNA to
bacterial species and genera. This approach provides a means for a much more
comprehensive assessment of microbial flora in the river water than has been possible
previously, making this assessment more desirable because the microbial community
is at the foundation of the biogeochemical cycles in aquatic systems (Axmanová et al.,
2006; Judd et al., 2006). Understanding how aquatic microbial populations respond to
FSC activities will bring important insights into understanding ecosystem health and
public health risks (Azam, 1998; Wetzel, 2001).
1.5 Estimating the relative abundance of micro-organisms
1.5.1 Culture Methods
Culturable bacteria refer to bacteria that are viable when sampled from the
environment and will grow on culture media under laboratory conditions.
Previously, detection and analysis of bacteria in the environment were performed
mainly by using culture-based methods (Fry, 2000; Kaeberlein et al., 2002; Zwart et
al., 2002; Rodríguez, 2004; Schleifer, 2004; Neelakanta & Sultana, 2013;).
Handelsman (2004) suggested that precise studies of bacteria in pure culture were
more appealing to microbiologists and therefore, most of the knowledge that fills
modern microbiology textbooks results from organisms isolated in pure culture.

20
Bodour et al. (2003) and Rees et al. (2004) note that the current application of culture
techniques is recognized as having limited scope for studying microbial diversity in
most environments, although culture based techniques have enabled microbiologists
to identify many culturable organisms.
In the past, results obtained by culture-dependent techniques covered only those few
organisms that could be cultivated. Several studies have employed culture-
independent techniques to show that cultivated microorganisms from diverse
environments often may represent very minor components of the microbial
community as a whole. Although efforts have been made to reveal the microbial
ecosystems on the basis of traditional cultivation methods (Spring et al., 2000), it is
now widely recognized that only small proportion of the total cell counts of bacteria
can be cultured under laboratory conditions (Amann et al., 1990; Ward et al., 1990;
Amann et al., 1995; Hugenholtz, 2002; Ling et al., 2015). This situation, according to
Miteva et al. (2004) and Pearce et al. (2003), substantially lessens the understanding
of the actual physiological and metabolic properties of the microbial community
present in the environment. Additionally, microorganisms retrieved using common
culture methods are rarely numerically abundant or functionally significant in the
environment from which they were cultured.
Cultivation based methods are important since the ecological role of prokaryotes in
natural environments can be estimated only when they are successfully cultivated and
characterized (DeLong et al., 1993; Amann et al., 1995; Tamaki et al., 2005).
However, at the same time, conventional cultivation of microorganisms is laborious,
time consuming and most importantly, selective and biased for the growth of specific
microorganisms (Ferguson et al., 1984; Eilers et al., 2000; Ling et al., 2015).
The majority of cells obtained from nature and visualized by microscopy are viable,
but they do not generally form visible colonies on plates (Eilers et al., 2000). The
majority of bacterial community, although viable in their natural environments, do not
grow under laboratory conditions and remain in a viable but non-culturable (VBNC)

21
stage (Lleo et al., 2005; Oliver, 2005). Such VBNC organisms could represent
completely novel groups and may be abundant or very active but remain untapped by
standard culture methods. Injury of cells, availability of nutrients and the presence of
lysogenic bacteriophages among others can be factors affecting non-culturability of
bacteria (Ashbolt et al., 2001).
VBNC bacteria are a major concern in public health risk assessments because many
pathogenic bacteria like Vibrio, Campylobacter and others have been reported to enter
a VBNC state from which they are able to return to the infectious state after passaging
in animal hosts (Yogita, 2005). Many potentially harmful bacteria survive sterilization
treatment and persist in processed food and potable water and in the environment.
Although the discovery of VBNC bacteria has highlighted the significance of growth-
independent microbial detection procedures, it is a fact that standard culture-based
microbiological methods have proven to be sufficiently effective to protect public
health for many decades.
Campylobacter are microaerophilic, very small, curved, thin, gram-negative rods (1.5
to 5 µm), with corkscrew motility. Due to the fastidious nature of these
microorganisms of concern, a pre-enrichment step is a necessity to culture these
bacteria. However, although cultured successfully, the pre-enrichment step in itself
precludes comparative estimates of abundance of these bacteria in the environment, or
in the case of the present study, in the water column. Environmental waters, especially
contaminated waters, are responsible for a number of outbreaks of Campylobacter
infection (Jones & Roworth, 1996; Sopwith et al., 2008). Campylobacter has been
isolated from a variety of environmental water sources, including rivers, lakes, and
ponds and streams (Bolton et al., 1987; Hanninen et al., 1998; Obi et al., 2002).
Lawson et al. (1999) mentioned that infection can occur through ingestion during
recreational water activity or by consumption of contaminated potable water.
Like all culture dependent methods, detection and identification of Campylobacter in
environmental waters is a long process and is labour-intensive. However, the use of

22
conventional culture techniques and the development of new culture media are
encouraged due to the advantages of having pure isolates to undertake physiological
and metabolic studies. In the present work, efforts were made to culture
Campylobacter from the Qawa River.
1.5.1.1 Viable Cell Counting
A viable cell count allows one to detect the number of actively growing and dividing
cells in a sample. The plate count method or spread plate relies on bacteria growing
on agar that become visible to the naked eye and so can be counted. Original samples
are diluted (1:10, 1:100, 1:1000 etc.) in peptone water before incubations so that on
average, counts must be between 30 to 300 colonies of the target bacterium per plate.
Total viable count (TVC) gives a quantitative indication for the presence of
microorganisms in a sample and this is recorded as the number of colony forming units
(CFU) per ml of the sample. In the present work, TVC estimates were made from
water samples taken from the Qawa River before, during and after the crushing season.
1.5.2 Molecular Methods
While culture-based approaches are extremely useful for understanding the
physiological potential of isolated organisms (Dahllöf, 2002; Rodríguez, 2002;
Schleifer, 2004), they do not necessarily provide comprehensive information on the
composition of microbial communities in their natural environment (Moyer et al.,
1994; Berthelet et al., 1996; Borneman & Triplett, 1997; Hugenholtz et al., 1998;
Omar & Ampe, 2000; Xu, 2006). Furthermore, it is also difficult to culture most
bacteria in environmental samples (Kogure et al., 1978; Ward et al., 1990; Amann et
al., 1995; Lleo et al., 2005) because natural populations comprise many species and
therefore evaluating the changes in bacterial community structure is not possible with
culturing techniques (Ward et al., 1990; Amann et al., 1995).
Because of the limitations linked to culture dependent methods, molecular methods
have become of increasing interest to researchers wanting to broaden knowledge of
complex ecosystems (Huws et al., 2007). The understanding of microbial diversity

23
has been greatly enhanced by DNA-based molecular tools. DNA techniques can by-
pass culturing problems by determining the taxonomic diversity of environmental
samples. Using molecular techniques, the phylogenetic and functional diversity of
non-culturable bacteria can be discovered and its distribution documented (Huson et
al., 2011). Furthermore, modern molecular biology offers numerous ways that allow
for the simple exploration of bacterial diversity in natural ecosystems. The culture-
independent methods that this technology provides have exposed an enormous
diversity of uncultured organisms. This finding underlines the potential to identify and
apply non-biased methods for the analysis of bacterial diversity in the environment
(Amann et al., 1995; Hugenholtz, 2002).
Various new DNA-based techniques have been established, which can aid the
identification of single bacterial species in sample material without the cultivation of
the organisms (Ward et al., 1990; Muyzer et al., 1993; Ludwig & Schleifer, 1994;
Amann et al., 1995). Most of the published work carried out to date has been based on
ribosomal sequences, which were used as phylogenetic markers (Woese, 1987).
Ribosomal RNAs (rRNAs) are primordial molecules, participating in protein synthesis
of cells.
According to Rosselló-Mora & Amann (2001), rRNA genes are especially suitable for
microbial diversity and phylogenetic studies due to the following properties:
• ubiquitous distribution
• contain conserved regions between organisms that are phylogenetically distant – this
facilitates their selective amplification with ‘universal’ DNA primers
• not affected by environmental changes
• RNA genes can occur in variable numbers in different organisms, however in many
organisms a process of concerted evolution gives rise to one characteristic sequence
per organism
• almost unaffected by horizontal genetic transference mechanisms

24
The 16S rRNA gene encodes an essential component of ribosomes important in the
translation of messenger RNA (mRNA) for protein synthesis and as indicated above
these molecules have several features that favor their use for molecular systematic
studies. Amplification of 16S rRNA by the polymerase chain reaction (PCR) followed
by sequencing has become widely used for bacterial identification in many kinds of
environmental studies and phylogenetic analyses of 16S rRNA gene have shown itself
to be a valuable technique for exploring bacterial biodiversity (Hiorns et al., 1997;
Glöckner et al., 2000; Pontes et al., 2007). While in some instances 16S rRNA
characterization provides species level information, generally it has been found most
useful for understanding phylogenetic relationships among prokaryotes above the
species level (e.g. Singh et al., 2012).
Pearce et al. (2003) also noted limitation of 16S rRNA gene sequences in their use to
differentiate closely related but ecologically distinct bacteria. In addition Dahllöf
(2002) and Hoffmann & Roggenkamp (2003) have mentioned that amplification bias
with ‘universal primers’ can mislead actual diversity of microbial communities in
environmental samples. This issues has also become evident when sequencing of
different 16S rRNA variable regions, which have sometimes given different
conclusions concerning microbial compositions (Wang et al., 2013). The specific
problem with universal primer pairs for amplification of 16S rRNA gene sequences is
that they may be more or less conserved in some organisms, and so this will lead to
some bacteria being amplified and others not. The relative abundance levels of taxa
can also be misled by such primer bias (Wang et al., 2013). Furthermore, Rosselló-
Mora & Amann (2001) and Jaspers & Overmann (2004) have noted that information
about microbial ecophysiological characteristics (and their genetic determinants)
cannot generally be obtained from 16S rRNA profiling.
Nevertheless, molecular methodologies using 16S rRNA gene sequences have been
helpful for characterizing unculturable bacteria, and are very important for the
improvement and development of culture methods for bacteria. The methodology
which is relatively inexpensive to apply also can provide valuable information on the

25
general similarity and differences of microbial populations. Whilst most recent
publications tend to favour characterization of 16S rRNA gene using Illumina®
sequencing, several publications have used methods such as 16S rRNA gene clone
libraries, fluorescence in situ hybridization (FISH) or denaturing gradient gel
electrophoresis (DGGE) to explore bacterial diversity in waters (e.g. Hiorns et al.,
1997; Hewson et al., 2003; Venter et al., 2004; Lyautey et al., 2005; Gilbride et al.,
2006; Winter et al., 2007; Buesing et al., 2009; Humbert et al., 2009; Ibekwe et al.,
2012; Jones et al., 2013).
The term “metagenomics” has been used to describe this growing field of studies
involving analysis of microbial populations in environmental studies, most typically
based on analyses of 16S rRNA gene (Handelsman, 2004). Next generation
sequencing (NGS) platforms including Roche 454® and Illumina® sequencing systems
have developed widely used commercial protocols for sequencing small segments
(~500 bp regions) of 16S rRNA gene (which are approximately 1,200 to 1,500 bp in
length). DNA amplification and sequencing primers target highly conserved regions
and taxonomic information is then provided from information within the hyper
variable regions (Jeraldo et al., 2011). The primary and secondary structures of the
16S rRNA gene show nine hyper-variable (V1 to V9) regions flanked by relatively
conserved regions. As noted by Lazarevic et al. (2012), most microbial diversity
surveys have been aimed at characterizing a small number of the hyper-variable
regions of the 16S rRNA, such as the V3 and V4 region analyzed in this thesis. These
individual regions, rather than the complete length of the 16S rRNA gene were
typically studied because of the read length limit with NGS sequencing protocols. For
example, with the MiSeq® 16S rRNA protocol used here, a 500 bp PCR product was
initially size fractionated on an agarose gel for sequencing, and the total read length
obtained for each end of sequenced DNA fragment was 250 bp.

26
1.6 Specific Aims of this thesis: Investigation of micro flora of the Qawa River
The work reported in this thesis represents the first systematic investigation of micro
flora in the Qawa River using modern techniques of molecular biology. Over a period
of 12 months, at four sampling intervals (three months before, during, one month after
and three months after the crushing season), measurements of dissolved oxygen (DO)
levels and water temperature were made at six sites on the river, centered in relation
to the location of the FSC mill. At four of these sites the taxonomic composition of
microbial populations was studied and compared using Illumina® 16S rRNA (V3 and
V4) sequencing and QIIME analysis. Based on the findings reported in Chapter 3,
suggestions and recommendations have been made in Chapter 4 for ongoing
monitoring of the Qawa River and other rivers of Fiji that host FSC mills.

27
CHAPTER 2
MATERIALS AND METHODS
2.1 Sampling
Water samples were collected from six sites along the Qawa River (Vanua Levu) as
shown in Figure 2.1 and described in Table 2.1. Two samples were collected in the
upper stream, two samples were taken at the FSC outfall and a further two samples
were collected beyond the outfall towards the river mouth.
Figure 2.1: Sampling sites along the Qawa River (Source: Google Maps, 2013); site
1: “F.E.A”, site 2: “Sawmill”, site 3: “F.S.C outflow”, site 4: “P.W.D”, site 5
“Junction”, site 6: “Namoli”.
Sample collections were carried out before, during and after the crushing season.
Sampling times were three months before crushing (represented as minus3), during
crushing (zero), one month after crushing (plus1) and three months after crushing
Direction of river flow

28
(plus3), making a total of four sampling trips. A total number of 24 water samples
were analyzed in the period of twelve months.
Table 2.1: Sampling locations on the Qawa River
Site GPS Site Description F.E.A
S 16° 25’01.5” E 179° 22’58.0”
· F.EA: downstream; · approx. 2 km from FSC
Sawmill
S 16° 25’30.6” E 179° 23’14.8”
· Sawmill: downstream; · approx. 1 km from FSC
F.S.C
S 16° 25’55.5” E 179° 23’4.84”
· FSC: outfall of sugar mill; · downstream
P.W.D
S 16° 26’28.9” E 179° 24’38.3”
· PWD: upstream; · approx. 2.5 km from FSC
Junction
S 16° 27’00.1” E 179° 25’10.4”
· Junction: upstream; · approx. 4.8 km from FSC
Namoli
S 16° 27’23.0” E 179° 25’31.7”
· Namoli: upstream; · approx. 7 km from FSC
Temperature in Vanua Levu, on average is always high. The warmest month is
December and the wettest month is March. July is the driest month during which the
FSC sugar mill begins its six month crushing season (Figure 2.2a and Figure 2.2b).
Figure 2.2a: Average monthly water temperature over the year in Vanua Levu. (Source: World Weather & Climate Information, 2014)

29
Figure 2.2b: Average monthly precipitation over the year in Vanua Levu. (Source:
World Weather & Climate Information, 2014)
2.2 DO and Temperature measurements
A YSI Model 85 multimeter was used to measure dissolved oxygen (DO) levels and
temperature at the six different sites during sampling. The probe was submerged in the
water, with replicate readings taken and recorded.
2.2.1 Statistical Inferences
2.2.1.1 DO: Sampling Site
DO values from the six sampling sites were treated as independent observations.
Replicates of DO data were graphed using Microsoft Excel and a non-parametric
correlation test was conducted using Kruskal-Wallis on Statistical Packages for Social
Sciences (SPSS) 21 software on data from each sampling site. A combined p value
was then tabulated using the Mann-Whitney test to test the significance (α =0.05) of
differences in DO levels at each of the six sites.
2.2.1.2 DO: Sampling Time
The DO data were collected from six sampling sites for four different time periods,
namely, before crushing (in April: minus3), during crushing (August: zero), one month
after crushing (December: plus1) and three months after crushing (March: plus3).
These were all treated as independent observations. To show the variation from the
four sampling times, DO values were graphed using Microsoft Excel and a non-
parametric correlation test was conducted using Kruskal-Wallis test implemented in

30
SPSS21 software. A combined α value was tabulated using Mann-Whitney test to test
the significance (α =0.05) of differences in DO level at each time period.
2.3 Total Viable Counts (TVC)
2.3.1 Protocol for handling bacteria for TVC
From each of the six sites, five composite samples of 1 L were collected on a straight
transect across the river two to three meters away from the river bank. The water
samples were collected in sterile McCartney bottles and kept in an eskie packed with
ice packs during transportation by air. All samples were kept in a dark and cool place
with temperature maintained at 1 to 4 °C until further analyses. Microbial analyses
were carried out on the water samples within six hours of their arrival at the University
of the South Pacific (USP) Microbiology Laboratory.
2.3.2 Culturing of water samples for TVC
All laboratory aspects of bacteriological analysis were analyzed according to the
Standard Method for the Examination Water and Wastewater (APHA, 2005).
Samples were serially diluted from 10-1 to 10-6 and pour-plated on to Nutrient Agar
(NA) as shown in Figure 2.3.
Figure 2.3: Serial dilution of water samples and plating scheme

31
Three replicates were plated for each dilution. The plates were incubated at 37 °C for
24 to 42 hours and then observed for bacterial growth. Plates that had 30 to 300
colonies were counted and recorded.
2.3.3 Statistical inferences
2.3.3.1 Viable Counts: Sampling Site and Sampling Time
Replicates of viable counts (cfu/ml) data were graphed using Microsoft Excel, to show
changes in absolute counts for the six sampling sites over the 12 month sampling
period.
2.3.3.2 Correlation test for viable counts (log) and DO: sampling site vs
sampling time
A Non Parametric Correlation test was conducted using Kendall’s tau-b (Abdi, 2007)
to investigate differences in overall log count and DO level at the six sites and four
sampling times (significance level (α =0.05)).
2.4 Culturing of indicator species of potential concern from the Qawa River
2.4.1 Campylobacter sp. River water samples (10 ml) from each site were added to 10 ml Bolton broth (Meat
peptone 10 % w/v, 5 % w/v Lactalbumin hydrolysate, 5 % w/v Yeast Extract, 5 mM
NaCl, 1 % w/v Alpha-ketoglutaric acid, 0.5 % w/v Sodium pyruvate, 0.5 % w/v
Sodium metabisulphite, 0.6 % w/v Sodium carbonate, 0.01 % w/v Haemin) tubes in a
duplicate enrichment step culture protocol (Figure 2.4). The culture was incubated at
37 °C for 4 hours and then at 42 °C for an overnight incubation.
The enrichment broth was then plated onto Charcoal Cefoperazone Deoxycholate
(CCD) Modified Agar Base (mCCDA) (10 % w/v Meat extract, 10 % w/v Peptone, 5
mM NaCl, 4 % w/v Bacteriological Charcoal, 3 % w/v Casein hydrolysate, 1 % w/v
Sodium Deoxycholate, 0.25 % w/v Iron (II) sulfate, 0.25 % w/v Sodium pyruvate, 15
% w/v Agar) and incubated at 42 °C under micro-aerophillic conditions using the
CampyPak™ Plus Systems (Fort Richard Laboratories, NZ) for 2 to 5 days.

32
Identification of cultured Campylobacter was carried out using:
i. Observation under the light microscope
Smears were prepared of bacterial cultures growing on mCCDA (less than 20 hours
old) on a clean glass slide with a drop of distilled water. Slides were then observed
under oil immersion using an Olympus C65 light microscope.
ii. Gram negative staining
Smears was prepared of bacterial cultures growing on mCCDA (less than 20 hours
old) and heat fixed. The slides were flooded with crystal violet solution for one minute
and rinsed with water. A counter stain, Grams iodine, was added on the slide for
another one minute. This was then washed off by adding 95 % ethanol and
immediately followed by water as a decolourisation step. A final stain, safranin, was
added for a minute and finally rinsed off with water. Slides were observed under oil
immersion using an Olympus C65 light microscope.
iii. Biochemical Tests:
A. Catalase Test
A small amount of culture was transferred to a clean slide with a sterile loop and 3 %
hydrogen peroxide solution. A positive reaction for catalase results from breakdown
of hydrogen peroxide and is evidenced by the evolution of bubbles.
B. Oxidase Test
An oxidase test strip was used to test for oxidation. A toothpick was used to transfer a
small amount of culture and rubbed on to the oxidase strip, without tearing the strip.
Doing this acts to remove the capsules from the surface of the bacteria so the substrate
can interact with the bacteria. A positive oxidized state is deep purple or blue. A
delayed or no colour change was interpreted as a negative result.

33
Figure 2.4: Enrichment step for Campylobacter using Bolton broth
2.5 DNA Extraction
2.5.1 Protocol for handling bacteria for DNA extraction
Water samples of 8 L in volume were used for DNA extraction. These were collected
in sterile 4 X 2 L polyethylene containers and sent to USP Microbiology Laboratory
at upper campus, for water filtration and DNA extraction. Since sample volumes for
DNA extraction were too large to be transported by air, they were sent via sea cargo
and kept in dark cool storage, to keep integrity of the microbes and minimize change
in bacterial populations.
2.5.2 Isolation of DNA
2.5.2.1 Protocol for DNA extraction of bacteria from river water
The protocol for DNA extraction from water samples was adapted from Hewson et al.
(2003) and Humbert et al. (2009) with some modifications. Membrane filters (Figure
2.5) were used to filter 8 L of river water collected from each site. This included a pre
filtration step using 5 µm filter paper. The filtrate was further filtered using 0.22 µm
nitrocellulose filter paper. Pre filtration has been found elsewhere to prevent clogging
on 0.22 µm filters and increase DNA yields (Richard Fong, pers comm).The 0.22 µm
filter was collected and stored for DNA extraction in 5 ml of sucrose lysis buffer (SLB-
400 mM NaCl, 750 mM sucrose, 20 mM EDTA, and 50 mM Tris-HCl (pH 9.0)).

34
The filters were cut into thin strips and placed into sterile 50 ml centrifuge tubes
containing 10 ml Tris-EDTA buffer (TE- 10 mM Tris-HCl pH 8.0, 1 mM EDTA)
buffer. Then 20 to 50 µl of 10 % w/v sodium dodecyl sulfate (SDS) solution was added
and this was incubated at 55 °C for 1 hr. The mixture was then subjected to 2x
freeze/thaw cycles at -70 °C after which 75 µl of Proteinase K (ProK) was added and
the mixture was incubated overnight at 55 °C. After incubation, the mixture was
centrifuged at 14000 rpm again and the top layer was transferred to another new tube,
to which 5 ml of 1:1 phenol/chloroform solution was added.
Figure 2.5: Membrane filtration equipment
This solution was then centrifuged at 14000 rpm and the aqueous phase was
transferred to another new tube to which 50 µl 5 M NaCl and 10 ml of ethanol stored
at 4 °C were added to initiate nucleic acid precipitation. The mixture was incubated at
-20 °C overnight to maximize the yield of precipitate.
The mixture was then centrifuged at 14000 rpm, the supernatant discarded and the
nucleic acid pellet resuspended in 5 ml 80 % v/v ethanol, followed by an incubation
at room temperature for 10 minutes. The mixture was centrifuged at 14000 rpm again,
Membrane filter
Receiving Flask
Filter funnel
Filter holder
Vacuum source

35
the supernatant was discarded and the DNA pellet was air dried. The pellet was
resuspended in 0.05 ml of non-DNA water. The 80 % ethanol washing step was done
twice to help in the removal of any salts from the nucleic acid.
2.5.2.2 Protocol for DNA extraction from bacterial cultures
The DNA extraction protocol used for bacterial cultures was adapted from Wilson
(1997) with minor modifications. After growing Campylobacter on mCCDA, the cells
were cultured in 15 ml of Bolton broth at 37 oC for 4 hours and transferred to 42 oC
for an overnight incubation. The cultures were then centrifuged at 14000 rpm to
produce cell pellets and then pellets were transferred in an Eppendorf tubes for DNA
extraction. DNA extraction was done using a CTAB chloroform/isoamyl alcohol
method as follows:
Cell pellets were resuspended in 0.5 ml of CTAB buffer (100 mM Tris-HCl, 1.4 M
NaCl, 20 mM EDTA, 2 % w/v CTAB, 1 % w/v PVP [mol. weight 360,000], pH 8.0
and 0.4 % w/v 2-mercaptoethanol) and centrifuged at 14000 rpm for 5 minutes. When
centrifugation was complete, the Eppendorf tubes were removed and incubated in a
water bath at 65 oC for 20 minutes.
Following this, the tubes were allowed to cool, then an equal volume of
chloroform/isoamyl alcohol mixture (24:1) was added to cooled tubes and these were
placed on a rotator mixer for another 20 minutes and then centrifuged again at 14000
rpm for 15 minutes. This procedure resulted in an aqueous phase containing nucleic
acids, an interphase containing precipitated protein and a lower solvent phase. The
uppermost aqueous phase was transferred into a new 2 ml sterile tube, to which 0.5 ml
of 5.0 M NaCl and 1 volume of isopropanol were added. The tubes were inverted
several times to mix and incubated at -70 °C for at least one hour. The tubes were then
centrifuged at 14000 rpm for 30 minutes. The supernatant was decanted and the pellet
was washed with 80 % v/v ethanol quickly followed by a 20 second centrifugation
step. The supernatant was discarded and tubes were left in an inverted position at room
temperature. After all traces of ethanol had disappeared, the DNA pellet was
resuspended in 20 µl of sterile PCR grade water and stored at -20 °C.

36
2.6 MiSeq® sequencing of bacterial DNA from river water on the NZGL platform
Extracted DNA was submitted to the NZGL (Massey University) sequencing facility
http://www.nzgenomics.co.nz/. The DNA samples were checked for quality (DNA
degradation) on an Agilent Bioanalyzer
(http://www.genomics.agilent.com/en/Bioanalyzer-System/2100-Bioanalyzer-
Instruments/?cid=AG-PT-106) and for contaminating protein using a Nanodrop
spectrophotometer (http://nanodrop.com/Library/CPMB-1st.pdf).
Samples from the four sites were found suitable (intact and of high quality) for MiSeq®
sequencing. Indexed 16S rDNA V3_V4 Nextera libraries were prepared for each of
these sites and all sampling times. Three libraries were prepared by the Massey
Genome (NZGL)
Service using a standard operating protocol:
http://www.illumina.com/products/nextera_xt_dna_sample_prep_kit.html.
Generating these libraries involved a two-step tailed PCR approach, the first step
amplified the 16S rDNA locus (V3 and V4 region) from each sample and then a second
step added Nextera indexed sequencing adaptors
https://www.illumina.com/content/dam/illumina-
support/documents/documentation/chemistry_documentation/16s/16s-metagenomic-
library-prep-guide-15044223-b.pdf. This protocol allows different samples to be
pooled and sequenced together on an Illumina MiSeq® flow cell or in a single
HiSeq® lane. In our case the Qawa River water libraries were sequenced overnight on
an Illumina MiSeq® sequencer using a 250 bp paired ends sequencing protocol.
The sequencing by NZGL involved first loading the library onto to a MiSeq® flow cell
under conditions where that the library DNA fragments attached to the oligonucleotide
“lawn” of the flow cell. Bridge amplification PCR was then used to generate a cluster
for each attached DNA fragment. Both ends of the cluster fragments (paired end
sequencing) were then sequenced using Illumina’s proprietary reversible terminator
sequencing method which captured images of the florescence signals emitted as the
different dNTPs were incorporated into the synthesized DNA strand complementary

37
to the fragment of DNA used to generate the cluster
http://www.illumina.com/content/dam/illumina-
marketing/documents/products/illumina_sequencing_introduction.pdf.
Figure 2.6: Secondary structure of the 16S rRNA gene of bacteria indicating variable
V1V9 regions, adapted from Yarza et al. (2014).
The V3 and V4 regions of the 16S rRNA gene common to all bacterial species targeted
by the above protocol have been shown in Figure 2.6. Following sequencing, the data

38
generated were QC checked by a NZGL bioinformatician (Mauro Truglio). This
involved checking the quality scores of every base in the fastq file for each sequence
with the software SolexaQA: (solexaqa.sourceforge.net/; Cox et al., 2010) and
separating data with low cumulative quality scores from those with high scores.
The high quality data were then summarized and visualized using the QIIME
(quantitative insights into microbial ecology) analysis pipeline (Caporaso et al., 2010)
on a high performance desktop server with 32 GB RAM by NZGL bioinformatician
Patrick Biggs.
2.6.1 QIIME bioinformatics pipeline
QIIME is an open source software package useful for comparing 16S rDNA sequences
in different samples. QIIME provides for OTU (operational taxonomic unit)
assignment, construction of distances (as a measure of the difference between
microbial sample profiles) and phylogenetic trees from which phylogenetic diversity
(sum of branch lengths separating OTUs for a given sample) can be calculated.
It provides a framework for statistical analysis and visualization of NGS data. Here it
was used to assign 16S rDNA sequence reads to taxa using the RDP classifier and to
compare microbial profiles between sampling times and sites. A resampling analysis
that compared species richness and phylogenetic diversity for different sample sizes
of sequences was also undertaken to determine whether differences in microbial
profiles could be detected for the sample sizes sequenced (100,000 sequenced reads
per sample). From the list of assigned taxa, some organisms of concern for human
health were identified.
An overview of the QIIME bioinformatics pipeline for Roche 454® sequence data has
been given in Caporaso et al. (2010) and is shown in Figure 2.7. The pipeline is very
similar to that used for Illumina® sequence data.
From the outputs produced by the pipeline we have produced a plot of alpha diversity
(within sample diversity) for subsets of sequence reads of increasing number. This is

39
a so called ‘rarefaction plot’ which shows how species number and phylogenetic
diversity estimates change with the sample size of sequenced reads. Species number
and phylogenetic diversity are expected to be highly correlated, but in the case of poor
quality sequence data (as explained above for Solexa QA) the two measures can differ
because sequencing errors can inflate inferred species number greatly but less affect
phylogenetic diversity. The latter is a measure of the sum of branch length differences
in a phylogenetic tree that connects the observed ‘species’. PCoA plots have also been
used here to visualize the difference in microbial profiles estimated for different
samples. Previously such plots have proven useful for demonstrating that microbial
composition (inferred from illumina® sequence data) differ between environmental
isolates (Caporaso et al., 2012).

40
Figure 2.7: QIIME Bioinformatics pipeline, from (Caporaso et al., 2010)

41
CHAPTER 3
HEALTH OF THE QAWA RIVER
RESULTS
3.1 Physical Parameters
3.1.1 Temperature
River water temperature varied throughout the sampling seasons on the Qawa River.
There was an overall increase of 3 to 4 degrees celsius in the three months leading up
to the crushing season.
Figure 3.1: Water temperature (°C) measured at six sites on the Qawa River before,
during and following the sugar cane crushing season.
Temperature peaked during the crushing season (sampling time zero) and then fell
over three to four months following the period of crushing (Figure 3.1). Temperature
remained relatively constant between sites at each sampling time.
0
5
10
15
20
25
30
35
minus3 zero plus1 plus3 Sampling Times
Temperature ( ((° C)
FEA saw mill FSC PWD Junction Namoli

42
3.1.2 Dissolved Oxygen (DO)
Figure 3.2 shows changing levels of dissolved oxygen (DO) at six sites on the Qawa
River before (minus3), during (zero) and after the crushing season (plus1 & plus3).
DO values measured at the FSC outflow were lowest during the crushing season. Over
the time interval measured they were highest one to three months following the
crushing season.
Figure 3.2: DO levels at six different sampling sites (minus3 = three months prior to
crushing; zero = crushing season; plus1 = one month after crushing; plus3= three
months after crushing).
DO values ranged from 0.2 mg/L during crushing to approximately 8.0 mg/L three
months after crushing. Unlike the other sites sampled, Namoli, the site most upstream
of the FSC outflow, maintained high values of DO across the sampling period. A non-
parametric correlation test was used to further examine the difference between DO
levels at this site and at other sites.
0 1 2 3 4 5 6 7 8 9
10
FEA Saw mill FSC PWD Junction Namoli Sampling Sites
Dissolved Oxygen (mg/L)
minus3 zero plus1 plus3

43
3.1.2.1 DO: Non Parametric Correlation Test between sampling sites
As similar trends in DO were observed between six sampled sites across the sampling
interval, a pairwise correlation analysis was made of DO values between sampling
sites. The null hypothesis being that the trend for change in DO values with respect to
time was similar (correlated) between sites.
Table 3.1: Results of correlation test for DO values at four sampling times between
sampling sites.
Compared Sampling
Sites (DO) Significance level
α = 0.05
FEA Vs saw mill 1.00
Vs FSC 0.91
Vs PWD 0.29
Vs Junction 0.40
Vs Namoli 0.01*
Saw mill Vs FSC 0.67
Vs PWD 0.52
Vs Junction 0.40
Vs Namoli 0.01*
FSC Vs PWD 0.29
Vs Junction 0.22
Vs Namoli 0.05*
PWD Vs Junction 0.67
Vs Namoli 0.09
Junction Vs Namoli 0.09
NB. *: significant
At α< 0.05 the null hypothesis (that the pattern of changing DO values across the
sampling period was similar between sites) could not be rejected for most of the
pairwise site comparisons. The exception concerned Namoli. DO values always
remained high at this site across the sampling period. The sites closest to Namoli

44
(PWD and Junction) displayed a similar trend to FEA, saw mill and FSC sites in
respect of changing DO values over time. However, DO values at these two sites
(PWD and Junction) were only significantly different from Namoli at α<= 0.09.
It was concluded from this analysis that low DO values attributed to the FSC outflow
(see later results and discussion regarding this point) were a feature most characteristic
of sites downstream (FEA) of the FSC outflow site and at upstream sites closest to the
FSC outflow site (FSC and PWD).
3.1.2.2 DO: Non Parametric Correlation analysis between sampling times To
evaluate the extent to which DO values changed over time, the pattern of DO values
at six study sites was compared between sampling times. At α<0.05 the pattern of DO
values was significantly different at different sampling times for most pairwise
comparisons. Only between one and three months (plus1 month Vs plus3 months)
after crushing were the patterns of DO values not significantly different at α<0.05.
This finding is consistent with the inference of different microbial compositions found
in the river at different sampling times.
Table 3.2: Results of correlation test for DO values at six sites between sampling times
Comparison of sampling times (DO) significance level
α = 0.05
minus3 months Vs crushing (zero) 0.02*
Vs plus1 month 0*
Vs plus3 months 0*
crushing (zero) Vs plus1 month 0*
Vs plus3 months 0*
plus1 month Vs plus3 months 0.07
NB. *: significant

45
3.2 Microbiological Parameters
3.2.1 Total Viable Counts at six sites over the sampling interval
Total viable counts (duplicate estimates) were made for six sites at four sampling
times. The lowest number of viable counts was recorded from FEA site (values were
similar three months before and three months after the crushing season).
Figure 3.3: Total viable counts from the six sampled sites (minus3 = three months
before crushing; zero = crushing; plus1 = one month following crushing; plus3 = three
months following crushing).
The highest counts were recorded from the FSC site (the FSC outflow) during the
crushing season. At this time there was an increase in the total number of viable counts
at this site by x 103. Figure 3.3 indicates that the massive increase in microbial load
on the river was restricted to the FSC site.
3.3 Overall Correlation of Log Count vs Overall DO
A comparison was made between DO values and log transformed total viable counts
to identify any correlation between these two measurements. A nonparametric
correlation test using Kendall’s coefficient of concordance was 0, indicating no
detectable correlation between the log counts and DO.
0.0E+00 4.0E+02 8.0E+02 1.2E+03 1.6E+03 2.0E+03 2.4E+03 2.8E+03 3.2E+03 3.6E+03 4.0E+03
FEA Saw mill FSC PWD Junction Namoli Sampling Sites
Total Viable Counts (cfu/ml)
minus3 zero plus1 plus3

46
3.4 MiSeq® Sequencing Results
3.4.1. Rarefaction analysis
Figure 3.4. shows the results of jackknife resampling (sampling without replacement)
of sequence reads and calculation of phylogenetic diversity and also the number of
observed species for the subsets of sequenced reads. The graph shows that differences
in the phylogenetic diversity of the samples collected at different times became evident
when as few as 10,000 reads had been analyzed. Most phylogenetic and species
diversity is seen prior to crushing and least diversity three months after crushing. As
the rarefaction curves have not plateaued at 100,000 reads sequenced per sample, it
suggests that further sequencing of the 16S rDNA in the collected samples would
likely uncover further diversity of organisms.
Figure 3.4 Rarefaction plot: observed species vs time (red: minus3; blue: plus1; green:
crushing; brown: plus3)

47
Figure 3.5 Rarefaction plot: phylogenetic diversity vs time.
3.4.2. Microbial profiles upstream vs downstream for each sample time
A PCoA plot was calculated for the sequenced reads using QIIME software. Figure
3.5 represents the similarity between microbial profiles for individual samples
(100,000 reads each sample). Samples are color coded for sampling time. The plot
shows that water samples collected at the same time from different river sites have
similar microbial compositions. Figure 3.6 shows the samples color coded by location.
Samples from the same location, but collected at different times have different
microbial profiles.

48
Figure 3.6 PCoA plot of microbial profile differences when color coded for sampling
times
When color coded for sampling times (red: minus3; green: crushing; blue: plus1;
brown: plus3) the PCoA plots shows a clustering of individual samples from different
locations (Figure 3.6). That is, the microbial composition at the four sites is similar at
any one sampling time. Most variation between the profiles at different sites occurs a
month after crushing.
The profiles of samples taken from the same locations at different times do not cluster
together (Figure 3.7). That is, profiles for water samples taken from the same site are
dissimilar to each other at different sampling times.

49
Figure 3.7 PCoA plot of microbial profile differences when color coded for locations
3.4.3 Taxa present
Results from the 16S rDNA sequencing and QIIME analysis generated a list of taxa
from the Qawa River of potential interest for future environmental monitoring of the
river. A list of selected bacteria is shown in Table 3.3. A complete list and output from
QIIME has been provided as an appendix.

50
Table 3.3: list of selected identified taxa from the Qawa River
No. Taxa Identified Comments
1 Rhabdochlamydia sp. � Mostly present at one month after crushing
2 Synechococcus sp. � Present before and during crushing
3 Phenylobacterium sp. � Present at one month and three months after crushing
4 Rhodospirillaceae � Predominant at one month after crushing
5 Vibrio sp. � Predominant during crushing
6 Campylobacter sp. � Predominant during crushing
7 Pseudomonas sp. � Present before and one month after crushing
8 Aeromona sp. � Present at one month after crushing
9 Legionellales � Present at one month and three months after crushing
10 Enterobacteriales � Mostly present before crushing
11 Flavobacterium sp. � Mostly present during crushing
12 Arcobacter sp. � Mostly present during crushing
3.5 Culturing of Indicator species and confirmation of identity
Campylobacter sp. was successfully cultured from all sampling sites on the Qawa
River. Cultures grown on mCCDA agar showed a characteristic whitish colour and
raised colonies. Confirmation tests and results are provided in Table 3.4.

51
Figure 3.8: Cultured Campylobacter sp. from Qawa River on mCCDA agar
Table 3.4: Confirmation result for Campylobacter sp. Test Result
i. Gram Staining � Pink stain
� Gram negative
ii. Observation under the light microscope
� S-shaped or curved morphology
iii.
Biochemical Tests:
a. Catalyze Test
b. Oxidase Test
� �
Formation of bubbles on colonies Positive catalyze test
� Blue color on test strip
� Positive oxidase test
Campylobacter er rrrrsp. .

52
DISCUSSION
3.6 Qawa River Temperature and DO
Temperature and DO, together with other several important physical parameters of
water determines water quality.
3.6.1 Temperature
Temperature is fairly constant at the Qawa River (Figure 3.1) .The uniform
temperature observed is attributed to the large volume of water flow distributed across
a relatively shallow river. During the crushing season, temperature was marginally
higher than at the other sites during the second sampling period which corresponded
to the crushing season of FSC (Figure 3.1). The water that is drawn from the river for
cooling at the sugar mill is returned warmer than receiving water so this might account
for the higher temperature at this site. In the non-crushing season, the water
temperature at the FSC outfall does not vary much from adjacent sites. It was noted
that the temperature from all sites was relatively similar due to climatic conditions
during the time at which temperature was recorded (NASA, 2005). Conditions typical
of the dry season occur, especially in this geographic region of the Fijian islands. With
much less rain received during this period and slower river flows, warmer river waters
are expected than are typically seen during the wet season.
The lowest temperature recorded was from the most upstream site Namoli which could
have been attributed to the shading effect of riparian vegetation. Temperature can
become an important factor as it determines the solubility of DO and other gases in
the water bodies. Thus it is also noteworthy that Namoli displayed DO values different
from the other sites (Figure 3.2).
3.6.2 Dissolved Oxygen (DO)
DO is one of the most important parameters in water quality assessment. Although
there was no significant difference in DO for the majority of the sites sampled at any
particular sampling time, there was a significant difference in DO recorded at the most
upstream site (Namoli) when compared with the two most downstream sites (FEA and
Sawmill). There was also a marginal difference in DO at the FSC outfall compared to

53
the Namoli site. DO was highest and temperature lowest at the Namoli site, consistent
with the expectation that solubility of oxygen is high when water temperature is low.
Water turbulence and the cooling effect caused by the riparian vegetation at the
Namoli site may also contribute to the higher DO values recorded at Namoli. Although
no formal study was concocted, it was observed that fish and other aquatic organisms
seemed to thrive upstream at the four times of water sampling.
DO values changed significantly between sampling times (Table 3.2); before
(minus3), during (zero), and after crushing (plus1 and plus3 months). Lowest levels of
DO at the Qawa River correspond to the sugar mills crushing season. This presumably
reflects the negative impact of the sugar mill on the Qawa River DO. During crushing,
there is an influx of organic load in the water bodies which is received from the effluent
discharge from the sugar mill. Bacteria and algae are expected to utilize these organic
compounds as a food source. Whilst initial algal growth will help to increase DO (as
a byproduct of photosynthesis) the respiratory resulting from population growth load
(which will absorb DO from the water column) is expected to cause DO in the water
to fall markedly.
Furthermore, material which enters the river does not get properly flushed out to
because of the gentle gradient of the river and its silty bottom. Thus both climate and
river features were likely to have contributed to the low DO levels observed during
the crushing season, which was below a level required to sustain life in the Qawa
River. DO level ranged from <0.5 mg/L to 1 mg/L at the FSC outfall and sampling
points taken downstream within the vicinity of FSC during the crushing season (Figure
3.2). This DO value was well below the recommended DO level for healthy aquatic
life (Table 3.5) (APHA, 2005). The significance of this impact is perhaps also
evidenced by observations of dead fish observed floating on some parts of the river
closer to the FSC site during the crushing season (Tamata et al., 1996; Fung & Chand,
1997). Similar observations have also been reported elsewhere. For example, Ali &
Soltan (1996) documented fish mortalities in Bangladesh as a result of discharge of
sugar mill effluents.

54
Table 3.5: critical levels of DO mg/L in river water (Source: APHA, 2005)
DO mg/L Impact on aquatic organisms
0-2 Not enough oxygen to support life
2-4 Only a few fish and aquatic insects can survive
4-7 Good for many aquatic animals
7-11 Very good for most stream fish
3.7 Rarefaction Analysis
The results described from MiSeq® analyses were calculated for 100,000 reads per
sample. As the rarefaction plots indicate that the species number and phylogenetic
diversity in the samples have not plateaued, it is possible that further sampling will
identify additional taxa and further phylogenetic diversity. Comparisons were made
of 100,000 reads per sample and the clustering of profiles in Figure 3.5 indicates that
this number of reads was sufficient to detect similar microbial profiles at different
sampling times and different microbial profiles at the same sites at different times.
Note that the absolute level of phylogenetic diversity measured in the analysis will
also be determined by the phylogenetic resolution of the molecular marker analyzed,
in this case the V3 and V4 region of the 16S rDNA molecule. It is known for example
that random shotgun sequencing (sequences of random DNA reads) provides higher
levels of phylogenetic resolution than 16S rDNA sequencing (Singh et al., 2012;
Poretsky et al., 2014) and thus rarefaction curves and total measures of species
richness/phylogenetic diversity might look different for random shotgun sequencing.
An interesting point that is noteworthy is that the rarefaction plot gives an indication
of relative phylogenetic diversity in the river at different sampling times. Even though
total viable counts increased dramatically during the crushing season this did not
dramatically increase overall phylogenetic diversity, rather while the absolute
numbers of bacteria increased at the FSC outflow site, the microbial composition
remained similar throughout the river at the four sites tested. This observation is
perhaps consistent with previous reports that increased nutrients from organic loads
can stimulate the growth of some species but not others (Borhidi et al.,1986; Ali &

55
Soltan,1996; Singh et al., 1998; Arindam 1999). In this situation growth has the
potential to reduce phylogenetic diversity.
3.8 Microbial compositions mirror changes on river DO
There was a very weak to negligible correlation between the log viable counts and DO.
DO values tended to be similar between sites at any one time, while the massive
increase in bacterial growth during the crushing season was only observed at the FSC
mill site. This finding is interesting and it would be worth further investigating the
relationship between changes in microbial populations and bacterial abundance at sites
adjacent to the FSC mill. DO values at different sites (the exception being Namoli) at
the same sampling time are similar. Mirroring this, was the finding that microbial
compositions are also similar at different sites at the same sampling times (clustered
points, Figure 3.6). Thus there appears to be a close relationship between DO levels
and microbial composition in the river. If it is assumed that changing river DO levels
are largely driven by a biotic response to effluent from the FSC mill, then it is clear
that the FSC mill has a very significant impact on the ecology of the river.
A river ecosystem is strongly affected by human activities. The pollutants discharged
into a river from human activities may destroy the ecosystem of the river (Qadir et.
al., 2013).
Microorganisms in rivers play key roles in degrading the pollutants and therefore in
preventing the river ecosystem from being destroyed (Kenzaka et al., 2001; Salman
et. al, 2013). The influx of organic matter most especially when the sugar mill is in
operation, can alter the natural purification system of the receiving water body. This
creates opportunities for invasive plants and organisms, including bacteria, to outgrow
those indigenous to the area. This drastically degrades the water body as the capacity
to for a given water body to perform its “cleanup” has been compromised. As a result,
accumulation of pollutants will occur which further degrades the aquatic ecosystem.
Vakabua (1991) noted that during the sugarcane crushing season, the presence of oil
in the Ba River prompted the growth of oil utilizing bacteria including Pseudomonas.

56
3.9 Microorganisms of concern/indicator in the river
Selected identified taxa from the Qawa River included:
• Rhabdochlamydia sp.: Mostly present at one month after crushing. These are
'chlamydia-like' organisms; they have a range of hosts from bivalves to fishes and
also been suggested as emerging pathogens for humans and animals (Thomas et
al., 2006). They live in a variety of aquatic habitats including waste water plants.
• Synechococcus sp.: Present before and during crushing and thrives particularly
well on coastal plumes of major rivers, enriched with nutrients such as nitrate and
phosphate, which are active ingredients of fertilizers used by cane farmers. They
are also capable of extensive growth, resulting in bloom events that can cause
significant threat to human and animal health (Carmicheal, 1992).
• Phenylobacterium sp.: Present at one month and three months after crushing and
grow optimally only on artificial compounds like chloridazon, an active
ingredient of a herbicide used for control of broadleaf weeds in sugar beet
(Eberspaher & Lingens, 2006).
• Rhodospirillaceae: Predominant at one month after crushing. Often found in
anaerobic aquatic environments, such as stagnant water and eutrophic ponds,
although they are able to survive in air (Allaby, 1998).
• Vibrio sp.: Predominant during crushing. They are facultative anaerobes that can
cause water borne illness (such as cholera) and foodborne infections usually
associated with eating undercooked seafood (Singh et al., 2012).
• Campylobacter sp.: Predominant during crushing, are pathogenic and can infect
humans and other animals. At least a dozen species of Campylobacter have been
implicated in human disease. They are now recognized as one of the main causes
of bacterial foodborne disease in many developed countries (Biggs et al., 2011;
Gras et al., 2012).
• Pseudomonas sp.: Present before and one month after crushing. They are
opportunistic human pathogens known to cause eye, nose and throat infections to
swimmers. Studies have also revealed some drug resistant strains (Nonaka et al.,
2010).

57
• Aeromonas sp.: Present at one month after crushing. They are facultative
anaerobic, mostly are associated with food and water pathogens. Agents of wide
spectra of diseases in man and animals (Ghenghesh et al., 2007).
• Legionellales: Present at one month and three months after crushing. They are
Gramnegative bacteria and include notable pathogens. Intracellular parasites
infecting invertebrates, animals and man (Garrity et al., 2005).
Enterobacteriales: Mostly present before crushing. These pathogens are known
as “enteric bacteria” including members such as Salmonella, Escherichia coli,
Yersinia, Klebsiella and Shigella; live in the intestine of animals. They are
responsible for a variety of human illnesses (World of Microbiology and
Immunology, 2003). The 16S rRNA data was not phylogenetically informative
in respect to identifying the genera and species of the Enterobacteria in the
samples.
• Flavobacterium sp.: Predominant during crushing. They are found in soil and fresh
water environments. Several species are known to cause disease in fresh water fish
(Mudarris et al., 1994).
• Arcobacter sp.: Predominant during crushing. It show an unusually wide range of
habitats and some species can be human and animal pathogens (Lehner et al.,
2005).
3.9.1 Viable Counts
Wastewater discharges provide bacteria and organic matter to the recipient systems.
Allochthonous bacteria and most enteric and pathogenic microorganisms are released
directly or through wastewaters to water bodies. Bacterial populations shift (Xu, 2006;
Ibekwe et al., 2012) in response to environmental changes, such as nutrient
enrichment.
As expected, the highest viable counts were recorded at the FSC site and the lowest
counts from the FEA site (furthest downstream), when samples from various sites were
compared. As suggested previously, the high absolute counts found during crushing
season are most likely attributable to the outflow of the sugar mill effluent into the

58
river. The low viable counts recorded furthest downstream are possibly due to the
mixing of seawater which will support a different diversity of microflora.
Because of their species diversity and ability to rapidly respond to their changing
environment, bacteria are potentially useful indicators of water quality (Lemke et. al.,
1997; Lear et al., 2013). Results show that during the crushing season, the bacterial
load in the river is further intensified as compared to the non-crushing season (Figure
3.3). The high viable counts correlate with expected high BOD and low measured DO
for this site which will accompany organic pollution during the crushing of sugarcane.
Agricultural activities have been well documented as sources of bacterial pollution in
rivers (Gunkel et. al., 2007; Akali et al., 2011; Salman et. al., 2013).
In addition, Fung & Chand (1997) reported high levels of total coliform and fecal
coliforms in the Qawa River which was more than the recommended level according
to WHO standards (WHO, 1993). The presence of Enterobacteria (Table 3.3) supports
this finding, further indicating that the Qawa River had heavy contamination of sewage
waste. The presence of these organisms might be attributed to the use of mangroves
along the Qawa and Labasa River as sinks for sewage treatment programs (Gray,
1989). This was evidenced perhaps from the observation that sampling sites furthest
from FSC still recorded high viable counts, which might be attributed to domestic and
municipal wastes which could be a secondary source of bacterial pollution in the Qawa
River.
The recovery of Campylobacter from river waters that could be cultured in the present
study is a further point of concern and should be considered for assessing public health
risks associated with the river and its different uses by people in the region.
Suggestions for future monitoring of the river have been made in the final chapter of
this thesis.

59
CHAPTER 4
Future Work and Monitoring of Qawa River
Whilst the 16S rRNA V3 and V4 amplicon sequencing provides for general
conclusions that can be drawn from the changing microbial composition of the Qawa
River, it does not explicitly provide information on health risks associated with the
sugar mill operation. This might be better evaluated using more comprehensive
random shotgun sequencing and also by initiating a routine survey for specific
organisms of concern using rapid and relatively inexpensive diagnostic DNA tests.
These suggestions are discussed in turn.
4.1 Comprehensive assessment
Recent published work (Poretsky et al., 2014) has highlighted the strengths and
weaknesses of 16S rRNA sequencing. Their conclusions are also evident in findings
of the present work. That is while the 16S rRNA sequencing showed clear patterns in
differences between microbial populations, in general it was unable to provide
resolution of species. For example, during crushing high levels of Vibrio were found
in the river immediately upstream from the FSC outflow site however the exact species
could not be identified. This finding is in contrast with a recent study of marine waters
near Suva using random shotgun sequencing that could more precisely identify the
Vibrio species present (Singh et al. 2012). The samples already collected for 16S
rRNA sequencing would also be suitable for shotgun sequencing and so further
information relevant to public health assessment could be made by analyzing these
samples.
A similar analysis could also be made of water samples collected from other rivers in
Viti Levu hosting sugar mills during the crushing season. One aspect of the technology
that has rapidly advanced in the last year is the computational speed required to match
DNA sequence reads to organisms in electronic databases. New computational
methods of assigning reads to sequences are reportedly 20,000x faster than when the
human genome was first sequenced (http://ab.inf.uni-tuebingen.de/software/diamond/)

60
and this means that it is possible to obtain comprehensive results on microbial profiles
in a timely fashion. However, even more rapid and potentially useful (because of its
relative cost) would be future surveys based on using the Loop Mediated
Amplification (LAMP) DNA amplification technique which is currently seeing
widespread uptake in clinical settings. Of particular recent interest is its potential for
quantitative analyses, which can provide more information for decision making than
do presence-absence tests (Temple et al., 2013). Companies such as Diagenetix in
Hawaii are now producing relative low costs LAMP devices ($ 6,000 FD) that can be
used to make rapid quantitative assessments for the presence for specific pathogens
(each test costs ~ $1 FD).
4.1.1 LAMP surveys
Loop Mediated Amplification is an emerging tool in clinical diagnostics (e.g. Zeng et
al., 2014) and environmental monitoring (e.g. Karanis et al., 2007). It involves a DNA
amplification reaction which takes place at a constant temperature. Thus it does not
require use of an expensive PCR machine. The enzyme, typically Bst, functions at 65 oC and the chemical reagents are produced dried down. The enzyme system are stable
under tropical conditions. Where genome data is available for pathogens of concern
(e.g. Vibrio cholera, Campylobater jejuni), primers can be designed that will
specifically amplify DNA from these organisms. Thus this technology provides, in a
10 minutes to 1 hr diagnostic test, a means of detecting the presence and amount of a
pathogen in any water or food sample.
For many of the organisms listed in section 3.9 genome data is available that can be
used to design LAMP primers (GENBANK: ncbi.nlm.nih.gov) for testing water
samples in Fijian rivers. Strain specific primers could also be developed for cultures
obtained from Fijian environmental isolates such as reported here for Campylobacter.
Using such methodology, many environmental samples as well as samples from
aquatic life (e.g. fish and mollusks) could be examined at different times of the year
to best evaluate specific public health concerns surrounding use of river water as well
as food safety.

61
Specifically, such monitoring could be undertaken to gain a better understanding on
the impact of marine life by activities of the Lautoka FSC mill. As mentioned in the
introduction, there is no treatment at this site, rather effluent from the mill is
discharged offshore. Further, the effectiveness of treatments at the Rarawai, Penang
and Labasa mills could also be assessed and monitored. Doing so would facilitate
assessment of public health risks associated with the water sources at these sites at
different times of the year. Using LAMP monitoring, both food and water could be
studied.

62
REFERENCES
Abdi, H. (2007). Abdi, H. (2007). Kendall rank correlation. In N. Salkind (Ed.), Encyclopedia of measurement and statistics. (pp. 509-511). Thousand Oaks, CA: Sage Publications, Inc.
Abraham, W. R. (2011). Megacities as sources for pathogenic bacteria in rivers and their fate downstream. International Journal of Microbiology, 2011, 1-13 doi:10.1155/2011/798292.
Akali, N., Nyongesa, N., Neyole, E., & Miima, J. B. (2011). Effluent Discharge by Mumias Sugar Company in Kenya: An Empirical Investigation of the Pollution of River Nzoia. Sacha Journal of Environmental Studies, 1, 1–30.
Akbar, N. M., & Khwaja, M. A. (2006). Study on Effluents from Selected Sugar Mills in Pakistan: Potential Environmental, Health and Economic Consequences of an Excessive Pollution Load. Sustainable Development Policy Institute (SDPI), (pp 144), Pakistan.
Ali, A. D., & Ahmad, M. S. (1993). Environmental Chemistry of groundwater in parts of Gandak basin, India. Indian Journal of Environmental Hydrology, 1, 31–36.
Ali, M. M., & Soltan, M. E. (1996). The impact of three industrial effluents on submerged aquatic plants in the River Nile, Egypt. Hydrobiologia, 340, 77–83.
Allaby, M. (1998). Rhodospirillaceae. Retrieved January 4, 2015 from http://www.encyclopedia.com/doc/107-Rhodospirillaceae.html
Amann, R. I., Binder, B. J., Olson, R. J., Chisholm, S. W., Devereux, R., & Stahl, D. A. (1990). Combination of 16S rRNA targeted oligonucleotide probes with flowcytometry for analyzing mixed microbial populations. Applied and Environmental Microbiology, 56, 1919–1925.
Amann, R. I., Ludwig, W., & Schleifer, K. H. (1995). Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiological Reviews, 59, 143–169.
Anderson, E., & Lloyd, B. (1995). Monitoring Performance of Effluent Treatment Systems at Fiji Sugar Corporation Rarawai Mill and Labasa Mill (pp. 1-23). IAS Technical Report No. 94, University of the South Pacific, Suva.
APHA. (2005). Standard methods for the examination of water and wastewater (21st ed.). American Public Health Association.
Araya, R., Tani, K., Takagi, T., Yamaguchi, N., & Nasu, M. (2003). Bacterial activity and community composition in stream water and biofilm from an urban river

63
determined by fluorescent in situ hybridization and DGGE analysis. FEMS Microbiology Ecology, 43, 111–119.
Arin, H. B., Dung, C. V, & Zitta, W. S. (2014). Groundwater supply in Jos-East Local Government Area of Plateau state, Nigeria. Journal of Environmental Science and Water Resources, 3, 1–6.
Arindam, A. (1999). Influence of carbonaceous sugar mill Effluent on phytosociology (herbaceous). Advances in Plant Science, 12, 159–162.
Ashbolt, N. J., Grabow, W., & Snozzi, M. (2001). Indicators of microbial water quality. In L. Fewtrell & J. Bartram (Eds.), Water Quality: Guidelines, Standards and Health (pp. 1–28). London, UK: IWA Publishing.
Atherton, J., Olson, D., Farley, L., & Qauqau, I. (2005). Fiji Watersheds at Risk: Watershed Assessment for Healthy Reefs and Fisheries. Wildlife Conservation Society-South Pacific (pp. 1-37), Suva.
Axmanová, S., Koutný, J., Cupalová, J., & Rulík, M. (2006). Bacterial growth and community composition in fractions of dissolved organic carbon of different molar mass from interstitial water. Folia Microbiologica, 51, 439–444.
Azam, F. (1998). Microbial control of oceanic carbon flux: the plot thickens. Science, 280, 694–696.
Azam, F., & Cho, B. (1987). Bacterial utilization of organic matter in the sea. In M., Fletcher, J., Jones, & T. R.,Gray (Eds.), Ecology of microbial communities (pp. 261–281). Cambridge: Cambridge University Press.
Azam, F., Fenchel, T., Field, J. G., Gray, J., Meyer-Reil, L., & Thingstad, F. (1983). The ecological role of water-column microbes in the sea. Marine Ecology Progress Series, 10, 257–263.
Azam, F., & Worden, A. (2004). Microbes, molecules and marine ecosystems. Science, 303(16), 22–24.
Baruah, A., Sharma, R. W., & Boralo, G. C. (1993). Impact of sugar mill and distillery effluents on Water Quality of River Gelabil, Assam. Indian Journal of Environmental Health, 25, 288–293.
Baskaran, L., Sundaramoorthy, P., Chidambaram, A. L. A., & Sankar, G. K. (2009). Amelioration of sugar mill effluent polluted soil and its effect of green gram (Vigna radiata). Botany Research International, 2, 107–114.
Benner, R., Opsah, S., Chin-Leo, G., Richey, R. E., & Forsberg, R. (1995). Bacterial carbon metabolism in the Amazon river system. Limnology and Oceanography, 40, 1262–1270.

64
Berthelet, M., Whyte, L. G., & Greer, C. (1996). Rapid, direct extraction of DNA from soils for PCR analysis using polyvinylpolypyrrolidone spin columns. FEMS Microbiology Letters, 138, 17–22.
Biggs, P. J., Fearnhead, P., Hotter, G., Mohan, V., Collins-Emerson, J., Besser, T. E., & French, N. P. (2011). Whole-Genome Comparison of Two Campylobacter jejuni Isolates of the Same Sequence Type Reveals Multiple Loci of Different Ancestral Lineage. PloS One, 6(11). e27121. doi:10.1371/journal.pone.0027121
Bodour, A., Wang, J., Brusseau, M., & Maier, R. (2003). Temporal change in culturable phenanthrene degraders in response to long-term exposure to phenanthrene in a soil column system. Environmental Microbiology, 5, 888–895.
Bolton, F. J., Coates, D., Hutchinson, D. N., & Godfree, A. F. (1987). A study of thermophilic campylobacters in a river system. Journal of Applied Bacteriology, 62, 167–176.
Borhidi, A., Slabarria-Fernandez, D., & Voros, L. (1986). Hydrobotanical studies in the framework of Cuban Zapata Project. Acta Botanica Hungarica, 32, 119–133.
Borneman, J., & Triplett, E. W. (1997). Molecular microbial diversity in soils from Eastern Amazonia: evidence for unusual microorganisms and microbial population shifts associated with deforestation. Applied and Environmental Microbiology, 63, 2647–2653.
Buesing, N., Filippini, M., Bürgmann, H., & Gessner, M. O. (2009). Microbial communities in contrasting freshwater marsh microhabitats. FEMS Microbiology Ecology, 69, 84–97.
Caporaso, J. G., Lauber, C. L., Walters, W. A, Berg-Lyons, D., Lozupone, C. A, Turnbaugh, P. J., & Knight, R. (2010). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proceedings of the National Academy of Sciences of the United States of America, 108, 4516–4522.
Caporaso, J. G., Lauber, C. L., Walters, W. A, Berg-Lyons, D., Huntley, J., Fierer, N., & Knight, R. (2012). Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The ISME Journal, 6, 1621–1624.
Carmicheal, W. W. (1992). Cyanobacteria secondary metabolites. Journal of Applied Bacteriology, 72, 445–459.
Carminati, A. V. (2008). An Assessment of the Effects of Sugarmill Activities on the
Ecological Integrity of the Mvoti and Amatikulu Rivers. University of Johhanesburg.

65
Castillo, M. M., Tallan, T. D., Sinsabaugh, R. L., & Kling, G. (2004). Seasonal and interannual variation of bacterial production in lowland rivers of the Orinoco basin. Freshwater Biology, 49, 1400–1414.
Chandra, R., & Mason, K. (1998). An Atlas of Fiji. Department of Geography, The University of the South Pacific, Suva.
Cheesman, O. D. (2005). Environmental Impacts of Sugar Production. United Kingdom: CABI Publishing.
Cole, J. J. (1999). Aquatic microbiology for ecosystem scientists: new and recycled paradigms in ecological microbiology. Ecology, 2, 215–225.
Cole, J. J., Prairie, Y. T., Caraco, N. F., McDowell, W. H., Tranvik, L. J., Striegl, R. G., & Melack, J. (2007). Plumbing the Global Carbon Cycle: Integrating Inland Waters into the Terrestrial Carbon Budget. Ecosystems, 10, 171–184.
Corcoran, E., Nellemann, C., Baker, E., Bos, R., Osborn, D., & Savelli, H. (2010). Sick Water? The Central Role of Wastewater Management in Sustainable Development. UNEP (pp. 1-85) Norway.
Cox, M. P., Peterson, D. A., & Biggs, P. J. (2010). SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinfomatics, 11, 485–491.
Dahllöf, I. (2002). Molecular community analysis of microbial diversity. Current Opinion in Biotechnology, 13, 213–217.
Danielopol, D., Griebler, C., Gunatilaka, A., & Notenboom, J. (2003). Present state and future prospects for groundwater ecosystems. Environmental Conservation, 30, 1–27.
DeLong, E. F., Franks, D. G., & Alldredge, A. L. (1993). Phylogenetic diversity of aggregate-attached vs. free-living marine bacterial assemblages. Limnology and Oceanography, 28, 924–934.
Devlin, D. L., & McVay, K. A. (2001). Suspended Solids: A Water Quality Concern for Kansas. Kansas State University, USA. Retrieved February 12, 2014 from http://www.oznet.ksu.edu.
Downing, J. A., Prairie, Y. T., Cole, J. J., Duarte, C. M., Tranvik, L. J., Striegl, R. G., & Middelburg, J. (2006). The global abundance and size distribution of lakes, ponds and impoundments. Limnology and Oceanography, 51, 2388–2397.
Eberspaher, J., & Lingens, F. (2006). The Genus Phenylobacterium. The Prokaryotes, 5, 250–256.

66
Eilers, H., Pernthaler, J., Glöckner, F. O., & Amann, R. (2000). Culturability and in situ abundance of pelagic bacteria from the North Sea. Applied and Environmental Microbiology, 66, 3044–3051.
Erah, O., Akujieze, N., & Oteze, E. (2002). The quality of ground water in Benin City, A baseline study on organic chemicals and microbial contaminants of health importance in boreholes and open wells. Tropical Journal of Pharmaceuticals Research, 1, 75–82.
Fagan, L., Cakausese, N., & Anderson, E. (1995). Water Quality in Ba River and Estuary: Environmental Effects of Multiple Resource Use (pp 1-47). IAS Environmental Report No 95, University of the South Pacific, Suva.
Fausch, K. D., Torgersen, C. E., Baxter, C. V, & Li, H. W. (2002). Landscapes to Riverscapes: Bridging the Gap between Research and Conservation of Stream Fishes. BioScience 52, 483–497.
Ferguson, R. L., Buckley, E. N., & Palumbo, A. V. (1984). Response of marine bacterioplankton to differential filtration and confinement. Applied and Environmental Microbiology, 47, 49–55.
Fiji Bureau of Statistics. (2013). 2007 Census of Population and Housing. Retrieved January 4, 2015 from http://www.statsfiji.gov.fj.
Fiji Department of Energy. (2010). Fiji: Renewable Energy Report. Suva.
Fiji Department of Environment. (1999). Fiji's National Biodiversity Strategic and Action Plan. Suva.
Fischer, H., Sachse, A., Steinberg, C. E. W., & Pusch, M. (2002). Differential retention and utilization of dissolved organic carbon by bacteria in river sediments. Limnology and Oceanography, 47, 1702–1711.
Fry, J. (2000). Bacterial diversity and “unculturables.” Micobiology Today, 27, 186–
188.
Fung, C., & Chand, K. (1997). Status of Water Quality in the Qawa River before the Sugarcane Crushing Season of 1996 (pp. 1-19 ). IAS Environmental Studies Report No. 83, University of the South Pacific, Suva.
Gafny, S., Goren, M., & Gasith, A. (2000). Habitat condition and fish assemblage structure in a coastal mediterranean stream (Yarqon, Israel) receiving domestic effluent. Hydrobiologia, 422/423, 319–330.
Galindo, M. C., Vece, M. B., Perondi, M. E., Monserrat-Araoz, M., Garcia, G., del Hidalgo, M., & Blesa, M. A. (2001). Chemical behaviour of the Sali River, Province of Tucuman, Argentina. Environmental Geology, 40, 847–852.

67
Garrity, G. M., Bell, J. A., & Lilburn, T. (2005). Legionellales ord. nov. In Bergery’s Manual of Systematic Bacteriology (pp. 210–247). Springer, USA.
Ghenghesh, K. S., Ahmed, S. F., El-khalek, R. A., Al-gendy, A., & Klena, J. (2007). Aeromonas -Associated Infections in Developing Countries. Journal of Infection in Developing Countries, 2, 81–98.
Ghosh, N., & McBean, E. A. (1998). Water Quality Modeling of the Kali River, India. Water, Air, and Soil Pollution, 102, 91–103.
Gilbride, K. A, Lee, D. Y., & Beaudette, L. A. (2006). Molecular techniques in wastewater: Understanding microbial communities, detecting pathogens, and real time process control. Journal of Microbiological Methods, 66, 1–20.
Glöckner, F. O., Zaichikov, E., Belkova, N., Denissova, L., Pernthaler, J., Pernthaler, A., & Amann, R. (2000). Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributed phylogenetic clusters including an abundant group of actinobacteria. Applied and Environmental Microbiology, 66, 5053–5065.
Google Maps. (2013). Qawa River, Labasa, Vanualevu. Retrieved March 14, 2013 from https://www.google.com.fj/maps/@-16.4519612,179.3583162,6105m/data=!3m1!1e3!5m1!1e2?hl=en-FJ
Gras, L. M., Smid, J. H., Wagenaar, J. A., Boer, A. G. De, Havelaar, A. H., Friesema, I. H. M., French, N. P., Busani, L., & Pelt, W. Van. (2012). Risk Factors for Campylobacteriosis of Chicken, Ruminant and Environmental Origin: A Combined Case- Control and Source Attribution Analysis. Plos One, 7, e42599. doi:10.1371/journal.pone.0042599.
Gray, A. J. (1989). A Directory of Wetlands in Ocenia. Wetlands International, Suva. (pp 1-41) Fiji.
Gunkel, G., Kosmol, J., Sobral, M., & Rohn, H. (2007). Sugar Cane Industry as a Source of Water Pollution – Case Study on the Situation in Ipojuca River, Pernambuco, Brazil. Water, Air and Soil Pollution, 180, 261–269.
Hammer, D. A. (1992). Creating Freshwater Wetlands, Lewis Publishers. (pp 1-480) USA.
Hamuda, H., & Patkó, I. (2011). Variations in Water Quality of Danube River at Budapest City. Obuda University e-Bulletin, 13–30.
Handelsman, J. (2004). Metagenomics: Application of Genomics to Uncultured Microorganisms. Microbiology and Molecular Biology Reviews, 68, 669–685.

68
Hanninen, M. L., Niskanen, M., & Korhonen, L. (1998). Water as a reservoir for Campylobacter jejuni infection in cows studied by serotyping and pulsed-field gel electrophoresis (PFGE). Zentralblatt Für Veterinaer- Medizin Reihe B, 45, 37–42.
Hartvich, F., Langhammer, J., & Vilimek, V. (2007). The 2002 flood consequences in the catchment of Otava River, Czech Republic. Water Resources, 34, 14–26.
Hewson, I., Vargo, G. A., & Fuhrman, J. A. (2003). Bacterial diversity in shallow oligotrophic marine benthos and overlying waters: effects of virus infection, containment, and nutrient enrichment. Microbial Ecology, 46, 322–336.
Hiorns, W. D., Methé, B. A., Nierzwicki-Bauer, S. A., & Zehr, J. P. (1997). Bacterial diversity in Adirondack mountain lakes as revealed by 16S rRNA gene sequences. Applied and Environmental Microbiology, 63, 2957–2960.
Hoffmann, H., & Roggenkamp, A. (2003). Population genetics of the Nomen species Enterobacter cloacae. Applied and Environmental Microbiology, 69, 5306–5318.
Hugenholtz, P. (2002). Exploring prokaryotic diversity in the genomic era. Genome Biology, 3, reviews0003.1–0003.8
Hugenholtz, P., Goebbel, B. M., & Pace, N. R. (1998). Impact of culture- independent studies on the emerging phylogenetic view of bacterial diversity. Journal of Bacteriology, 180, 4765–4774.
Humbert, J. F., Dorigo, U., Cecchi, P., Le Berre, B., Debroas, D., & Bouvy, M. (2009). Comparison of the structure and composition of bacterial communities from temperate and tropical freshwater ecosystems. Environmental Microbiology, 11, 2339–2350.
Huson, D. H., Mitra, S., Ruscheweyh, H., Weber, N., & Schuster, S. C. (2011). Integrative analysis of environmental sequences using MEGAN4. Genome Research, 21, 1552–1560.
Huws, S. A., Edwards, J. E., Kim, E. J., & Scollan, N. D. (2007). Specificity and sensitivity of eubacterial primers utilized for molecular profiling of bacteria within complex microbial ecosystems. Journal of Microbiological Methods, 70, 565–569.
Ibekwe, A. M., Leddy, M. B., Bold, R. M., & Graves, A. K. (2012). Bacterial community composition in low-flowing river water with different sources of pollutants. FEMS Microbiology Ecology, 79, 155–166.
Jaspers, E., & Overmann, J. (2004). Ecological significance of micro- diversity: identical 16S rRNA gene sequences can be found in bacteria with highly

69
divergent genomes and ecophysiologies. Applied and Environmental Microbiology, 70, 4831–4839.
Jeraldo, P., Chia, N., & Goldenfeld, N. (2011). On the suitability of short reads of 16S rRNA for phylogeny-based analyses in environmental surveys. Environmental Microbiology, 13, 3000–3009.
Jones, A. C., Liao, T. S. V., Najar, F. Z., Roe, B. A., Hambright, K. D., & Caron, D. A., (2013). Seasonality and disturbance: annual pattern and response of the bacterial and microbial eukaryotic assemblages in a freshwater ecosystem. Environmental Microbiology, 15, 2557–2572.
Jones, I. G., & Roworth, M. (1996). An outbreak of Escherichia coli O157 and campylobacteriosis associated with contamination of a drinking water supply. Public Health, 110, 277–282.
Jonkman, S., & Kelman, I. (2005). An analysis of the causes and consequences of flood disaster deaths. Disasters, 29, 75–97.
Jordaan, J. M. (2013). The Uses of River Water and Impacts. In Encyclopedia of Life Support Systems (Vol. III).
Judd, K. E., Crump, B. C., & Kling, G. W. (2006). Variation in dissolved organic matter controls bacterial production and community composition. Ecology, 87(8), 2068– 2079.
Kaeberlein, T., Lewis, K., & Epstein, S. S. (2002). Isolating “uncultivable”
microorganisms in pure culture in a simulated natural environment. Science, 296, 1127–1129.
Kanu, I., & Achi, O. (2011). Industrial Effluents and Their Impact on Water Quality of Receiving Rivers in Nigeria. Journal of Applied Technology in Environmental Sanitation, 1, 75–86.
Karan, M. (2010, June 11). Man from Qawa River. The Fiji Times. Suva.
Kaur, A., Vats, S., Rekhi, S., Bhardwaj, A., Goel, J., Ranjeet, S., & Gaur, K. K. (2010). Physico-chemical analysis of the industrial effluents and their impact on the soil microflora. International Conference on Ecological Informatics and Ecosystem Conservation 2, 595–599.
Karanis, P., Thekisoe, O., Kiouptsi, K., Ongerth, J., Igarashi, I., & Inoue, N. (2007). Development and Preliminary Evaluation of a Loop-Mediated Isothermal Amplification Procedure for Sensitive Detection of Cryptosporidium Oocysts in Fecal and Water Samples. Applied and Environmental Microbiology, 73, 5660– 5662.

70
Kenzaka, T., Yamaguchi, N., Prapagdee, B., Mikami, E., & Nasu, M. (2001). Bacterial Community Composition and Activity in Urban Rivers in Thailand and Malaysia. Journal of Health Science, 47, 353–361.
Kipsang, R., Muia, R., Makindi, A. W., & Omondi, D. O. (2014). Changes in the densities of faecal and organic matter contaminants from upstream to downstream along Nyangores River of Mara catchment, Kenya. Journal of Environmental Science and Water Resources, 3, 15–25.
Kogure, K., Shimizu, U., & Taga, N. (1978). A tentative direct microscopic method for counting living marine bacteria. Canadian Journal of Microbiology, 25, 415–
420.
Kumar, V., & Chopra, A. K. (2010). Influence of sugar mill effluent on physicochemical characteristics of soil at Haridwar (Uttarakhand), India. Journal of Applied and Natural Science, 2, 269–279.
Kumar, V., Rouquette, J. R., & Lerner, D. N. (2013). Integrated modelling for Sustainability Appraisal of urban river corridors: going beyond compartmentalised thinking. Water Research, 47, 7221–7234.
Lal, P. N., Rita, R., & Khatri, N. (2009). Economic Costs of the 2009 Floods in the Fiji Sugar Belt and Policy Implications. (pp. 1-52) IUCN, Switzerland.
Lawson, A. J., Logan, J. M., O’Neill, J., Desai, M., & Stanley, J. (1999). Large-scale survey of Campylobacter species in human gastroenteritis by PCR and PCR– enzyme-linked immunosorbent assay. Journal of Clinical Microbiology, 37, 3860– 3864
Lazarevic, V., Whiteson, K., Gaia, N., Gizard, Y., Hernandez, D., Farinelli, L., Østerås, M., Francois, P., & Schrenzel, J. (2012). Analysis of the salivary microbiome using culture-independent techniques. Journal of Clinical Bioinformatics, 2, 1-4.
Lear, G., Washington, V., Neale, M., Case, B., Buckley, H., & Lewis, G. (2013). The biogeography of stream bacteria. Global Ecology and Biogeography, 22, 544–
554.
Ledua, E., Matoto, S., Sesewa, A., Siassi, Y., Raj, D., & Korovulavula, J. (1996). Management of Coastal Fisheries in Macuata and Bua Provinces, Vanua Levu, Fiji. Suva
Lee, G. (1981). Report on a preliminary investigation into the alleged pollution of the Qawa River. University of the South Pacific, Suva.
Legendre, L., & Rivkin, R. B. (2008). Planktonic food webs: Microbial hub approach. Marine Ecology Progress Series, 365, 289–309.

71
Lehner, A., Tasara, T., & Stephan, R. (2005). Relevant aspects of Arcobacter spp. as potential foodborne pathogen. International Journal of Food Microbiology, 102, 127–135.
Lemke, M., Brown, B., & Leff, L. (1997). The Response of Three Bacterial Populations to Pollution in a Stream. Microbial Ecology, 34, 224–231.
Liang, G., & Ding, S. (2004). Impacts of human activity and natural change on the wetland landscape pattern along the Yellow River in Henan Province. Journal of Geographical Sciences, 3, 339–348.
Lindstrom, E. S. (2001). Investigating influential factors on Bacterioplankton community composition: results from a field study of five mesotrophic lakes. Microbial Ecology, 42, 598–605.
Ling, L. L., Schneider, T., Peoples, A. J., Spoering, A. L., Engels, I., Conlon, B. P., & Lewis, K. (2015). A new antibiotic kills pathogens without detectable resistance. Nature, 517, 455–459.
Lleo, M. M., Bonato, B., Tafi, M. C., Signoretto, C., Pruzzo, C., & Canepari, P. (2005). Molecular vs culture methods for the detection of bacterial faecal indicators in groundwater for human use. Letters in Applied Microbiology, 40, 289–94.
Lodhia, S. K. (1999). Accountants and Environmental Accounting: An initial and exploratory study into contemporary practices in Fiji. University of the South Pacific.
Lopez-Lopez, E., Favari, L., Martinez-Taboche, L., Madrigal, M., & Soto, C. (2003). Hazard Assessment of a mixture of pollutants from a sugar industry of three fish species of Western Mexico by the responses of enzymes and Lipid peroxidation. Bulletin of Environmental and Contamination and Toxicology, 70, 739–745.
Ludwig, W., & Schleifer, K. H. (1994). Bacterial phylogeny based on 16S and 23S rRNA sequence analysis. FEMS Microbiology Reviews, 15, 155–173.
Lyautey, E., Lacoste, B., Ten-Hage, L., Rols, J. L., & Garabetian, F. (2005). Analysis of bacterial diversity in river biofilms using 16S rDNA PCR-DGGE: methodological settings and fingerprints interpretation. Water Research, 39, 380–388.
Maidment, D. (1993). Hydrological Cycle. In D. Maidment (Ed.), Handbook of Hydrology. New York: McGraw-Hill, USA.
Manhokwe, S., Matiashe, I., & Jombo, T. (2013). An analysis of the water quality of groundwater sources in selected high density areas in Gweru urban, Zimbabwe. Journal of Environmental Science and Water Resources, 2, 302–309.

72
McGree, S., Yeo, S. W., & Devi, S. (2010). Flooding in the Fiji Islands between 1840 and 2009, (pp 1- 69) Fiji.
Miteva, V. I., Scheridant, P. P., & Brenchley, J. E. (2004). Phylogenetic and physiological diversity of microorganisms isolated from a deep Greenland glacier ice core. Applied and Environmental Microbiology, 70, 202–213.
Mokbel, W. A., & Yamakanamardi, S. M. (2008). Temporal variations in the abundance of heterotrophic bacteria in ground water according to land use patterns in Mysore district, India. African Journal of Environmental Science and Technology 2, 59–67.
Moran, M. A., & Hodson, R. E. (1990). Bacterial production on humic and nonhumic of dissolved organic carbon. Limnology and Oceanography, 35, 1744–1756.
Moyer, C. L., Dobbs, F. C., & Karl, D. M. (1994). Estimation of diversity and community structure through restriction fragment length polymorphism distribution analysis of bacterial 16S rRNA genes from a microbial mat at an active, hydrothermal vent system, Loihi Seamount, Hawaii. Applied and Environmental Microbiology, 60, 871–879.
Mudarris, M., Austin, B., Segers, P., Vacanneyt, M., Hoste, B., & Bernardet, J. F. (1994). Flavobacterium scophthalmum sp. nov., a Pathogen of Turbot. International Journal of Systematic Bacteriology, 44, 447–453.
Muylaert, K., Van der Gucht, K., Vloemans, N., De Meester, L., Gillis, M., & Vyverman, W. (2002). Relationship between bacterial community composition and bottom up versus top down variables in four eutrophic shallow lakes. Applied and Environmental Microbiology, 68, 4740–4750.
Muyzer, G., de Waal, E. C., & Uitterlinden, A. G. (1993). Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and Environmental Microbiology, 59, 695–700.
Naiman, R. J., & Décamps, H. (1997). The ecology of interfaces: Riparian zones. Annual Review of Ecology and Systematics, 28, 621–658.
NASA. (2005). What’s the Difference Between Weather and Climate? Retrieved
August 24, 2015 from http://www.nasa.gov/mission_pages.
Neelakanta, G., & Sultana, H. (2013). The Use of Metagenomic Approaches to Analyze Changes in Microbial Communities. Microbiology Insights, 6, 37–48.
Nguyen, J. Van. (2006). Bacterial Abundance & Community Composition of the Water Sediment System in the Venice Lagoon during the Sediment Transportation Project (pp. 1-60). University of California, USA.

73
Nonaka, L., Inubushi, A., Shinomiya, H., Murase, M., & Suzuki, S. (2010). Differences of genetic diversity and antibiotics susceptibility of Pseudomonas aeruginosa isolated from hospital, river and coastal seawater. Environmental Microbiology Reports, 2, 465–472.
Obi, C. L., Potgieter, N., Bessong, P. O., & Matsaung, G. (2002). Assessment of the microbial quality of river water sources in rural Venda communities in South Africa. Water SA, 28, 287–292.
Odum, H. T. (1956). Efficiencies, size of organisms, and community structure. Ecology, 37, 592–597.
Okoko, A. A., Muia, A. W., Moturi, W. N., & Oyake, M. (2012). Levels of E.coli contamination of River Awach and household water in western Kenya. Journal of Environmental Science and Water Resources, 1, 120–126.
Oliver, J. (2005). The viable but nonculturable state in bacteria. Journal of Microbiology, 43, 93–100.
Omar, N., & Ampe, F. (2000). Microbial community dynamics during production of the Mexican fermented maize dough pozol. Applied and Environmental Microbiology, 66, 3664–3673.
Onsdorff, K. A. (1996). Pollution; Water Environment. Law Practice Journal, 4, 14–
18.
Paerl, H. W., Dyble, J., Moisander, P. H., Noble, R. T., Piehler, M. F., Pinckney, J. L., & Valdes, L. M. (2003). Microbial indicators of aquatic ecosystem change: current applications to eutrophication studies. FEMS Microbiology Ecology, 46, 233–246.
Paul, M., & Meyer, J. (2001). Streams in the urban landscape. Annual Review of Ecology and Systematics, 32, 333–365.
Pawar, N. J., Pondhe, G. M., & Patil, F. (1998). Groundwater pollution due to sugarmill effluent at Sonai, Maharashtra, India. Environmental Geology, 34, 151–158.
Pearce, D. A., van der Gast, C. J., Lawley, B., & Ellis-Evans, J. C. (2003). Bacterioplankton community diversity in a maritime Antarctic lake, determined by culture-dependent and culture-independent techniques. FEMS Microbiology Ecology, 45, 59–70.
Pomeroy, L. R. (1980). Microbial role in aquatic food webs. In R. R. Colwell & J. Foster (Eds.), Aquatic Microbial Ecology (pp. 85–109). University of Maryland SeaGrant, College Park, USA.

74
Pontes, D. S., Lima-Bittencourt, C. I., Chartone-Souza, E., & Amaral Nascimento, A. M. (2007). Molecular approaches: advantages and artifacts in assessing bacterial diversity. Journal of Industrial Microbiology & Biotechnology, 34, 463–473.
Poretsky, R., Rodriguez, L. M., Luo, C., Tsementzi, D., & Konstantinidis, K. T. (2014). Strengths and Limitations of 16S rRNA Gene Amplicon Sequencing in Revealing Temporal Microbial Community Dynamics. PloS One, 9, 1–12.
Qadir, A., Malik, R. N., Feroze, A., Jamil, N., & Mukhtar, K. (2013). Spatiotemporal distribution of contaminants in Nullah Palkhu -highly polluted stream of Pakistan. Journal of Environmental Science and Water Resources, 2, 342–353.
Rees, G. N., Baldwin, D. S., Watson, G. O., Perryman, S., & Nielsen, D. L. (2004). Ordination and significance testing of microbial community composition derived from terminal restriction fragment length polymorphisms: application of multivariate statistics. Antonie van Leeuwenhoek, 86, 339–47.
Robinson, A. (1990). pH and the Environment. United States: Geology Department, University of California, Davis. Retrieved January 4, 2015 from https://www.geology.ucdavis.edu/~pcp/Reading/pH.pdf
Rodríguez, V. F. (2002). Approaches to prokaryotic biodiversity: a population genetics perspective. Environmental Microbiology, 4, 628–633.
Rodríguez, V. F. (2004). Environmental genomics, the big picture? FEMS
Microbiology Letters, 231, 153–158.
Rosselló-Mora, R., & Amann, R. (2001). The species concept for prokaryotes. FEMS Microbiology Reviews, 25, 39–67.
Salequzzaman, M., Islam, S. M. T., Tasnuva, A., Kashem, M. A., & Masud, M. (2008). Environmental Impact of Sugar Industry- A Case Study on Kushtia Sugar Mills in Bangladesh. Journal of Innovation and Development Strategy, 2, 31–35.
Salman, J. M., Al-azawey, A. S. N., & Hassan, F. M. (2013). Study of Bacterial Indicators in Water and Sediments from Al- Hilla River, Iraq. Hydrology Current Research, S13: 001. doi:10.4172/2157-7587.
Sangodoyin, A. (1991). Groundwater and Surface Water Pollution by Open Refuse Dump in Ibadan, Nigeria. Journal of Discovery and Innovations, 3, 24–31.
Santos, C. (2008). Study on Cleaner production: Opportunities for the Sugar Industry in Belize (pp. 1-94). National Central University, Taiwan.
Saranraj, P., & Stella, D. (2014). Impact of Sugar Mill Effluent to Environment and Bioremediation: A Review. World Applied Sciences Journal, 30, 299–316.

75
Schleifer, K. (2004). Microbial diversity: facts, problems and prospects. System of Applied Microbiology, 27, 3–9.
Shiklomanov, I., & Rodda, J. (2003). Worlds water resources at the beginning of the 21st century (pp. 1-25).Cambridge University Press, UK.
Singh, R., Narayan, V., McLenachan, P., Winkworth, R. C., Mitra, S., Lockhart, P. J., & Rao, D. (2012). Detection and diversity of pathogenic Vibrio from Fiji. Environmental Microbiology Reports, 4, 403–411.
Singh, A., Ghosh, S., & Pankaj, S. (2007). Water quality management of a stretch of river Yamuna: An interactive fuzzy multi-objective approach. Water Resources Management, 21, 515–523.
Singh, R. (2000). Pollution in Abu drainage- a preliminary report. Advances in Polymer Sciences, 13, 43–45.
Singh, H., Mishra, J. P., & Mahaver, L. R. (1998). Observation of certain characteristics of water in relation to pollution of river Ramganga. Journal of Environmental Biology, 19, 353–356.
Singh, B. B., Srivastava, G. K., & Gupta, R. P. R. (1996). Effect of Industrial Effluents on groundwater quality at Sardarnagar, Gorakhpur. Journal of Living World, 3, 61–64.
Sopwith, W., Birtles, A., Matthews, M., Fox, A., Gee, S., Painter, M., & Bolton, E. (2008). Identification of potential environmentally adapted Campylobacter jejuni strain, United Kingdom. Emerging Infectious Diseases, 14, 1769–1773.
SPC SOPAC. (2012). Catalogue of Rivers for the Pacific Islands (pp. 1-169). Applied Geoscience and Technology Division, Suva.
Spring, S., Schlze, R., Overmann, J., & Schleifer, K. H. (2000). Identification and characterization of ecologically significant prokaryotes in the sediment of freshwater lakes: molecular and cultivation studies. FEMS Microbiology Reviews, 24, 573–590.
Tamaki, H., Sekiguchi, Y., Hanada, S., Nakamura, K., Nomura, N., Matsumura, M., & Kamagata, Y. (2005). Comparative Analysis of Bacterial Diversity in Freshwater Sediment of a Shallow Eutrophic Lake by Molecular and Improved Cultivation Based Techniques. Applied and Environmental Microbiology, 71, 2162–2169.
Tamata, B., & Lloyd, B. (1994). Environmental Assessment of the Effect of the FSC Rarawai Sugar Mill on the Ba River (p. 37). IAS Environmental Studies Report No. 73, University of the South Pacific, Suva.

76
Tamata, B. R., Kubuabola, S. L., & Fung, C. (1996). Status of Water Quality in the Qawa River during the Sugarcane Crushing Season of 1995 (p. 22). IAS Environmental Studies Report No. 82, University of the South Pacific, Suva.
Temple, T. N., du Toit, L. J., Derie, M. L., & Johnson, K. B. (2013). Quantitative Molecular Detection of Xanthomonas hortorum pv. carotae in Carrot Seed Before and After Hot-Water Treatment. Plant Disease Journal, 97, 1585–1592.
Thomas, V., Casson, N., & Greub, G. (2006). Criblamydia sequanensis, a new intracellular Chlamydiales isolated from Seine River water using amoebal coculture. Environmental Microbiology, 8, 2125–2135.
Thuresson, S. (2001). Treatment of Distillery Effluent in Vegetated Surface Flow Wetlands in Muhoroni, Kenya. Swedish University of Agricultural Sciences, Uppsala (pp. 1-45) Sweden.
UNEP. (2003). The Environment in the News. Nairobi, Kenya. Retrieved February 6, 2014 from www.unep.org/cpi/briefs/BRIEF12march.doc
Usharanji, K., Umarani, K., Ayyasami, P. M., Shanthi, K., & Lakshmanaperumalsamu, P. (2010). Physico-Chemical and Bacteriological Characteristics of Noyyal River and Ground Water Quality of Perur, India. Journal of Applied Sciences and Environmental Management, 14, 29–35.
Vakabua, J. N. (1991). Studies on Ba River to Ascertain/Deny the Destruction of
Marine Resources by Sugar mill spillage (p. 19). Department of Fisheries Technical Report No 91, Suva.
Varis, O., Kummu, M., & Salmivaara, A. (2012). Ten major rivers in monsoon AsiaPacific: An assessment of vulnerability. Applied Geography, 32, 441–454.
Venter, J. C., Remington, K., Heidelberg, J. F., Halpern, A. L., Rusch, D., Eisen, J. A., & Smith, H. O. (2004). Environmental genome shotgun sequencing of the Sargasso Sea. Science, 304, 66–74.
Wang, J., McLenachan, P., Biggs, P. J., Winder, L. H., Schoenfeld, B. I. K., Narayan, V. V., & Lockhart, P. J. (2013). Environmental bio-monitoring with high-throughput sequencing. Briefings in Bioinformatics, 14, 575–588.
Ward, D. M., Weller, R., & Bateson, M. (1990). 16S rRNA sequences reveal numerous uncultured microorganisms in a natural community. Nature, 345, 63–65.
Watling, D. (1985). A Mangrove Management Plan for Fiji, Phase I. Government Press, (pp. 1-67) Suva, Fiji.
Watling, D., & Chape, S. (1992). Environment: Fiji. The National State of the Environment Report (pp. 1-154). IUCN, Fiji.

77
Wetzel, R. G. (2001). Bacterioplankton. In R. G. Wetzel (Ed.), Limnology. Lake and river ecosystems (3rd ed., pp. 489–525). New York: Academic Press.
WHO. (1993). Guidelines for drinking water quality. Geneva, Switzerland. Retrieved August 12, 2014 from http://www.who.int/water_sanitation_health/dwq/gdwq2v1/en/
Wilson, K. (1997). Preparation of Genomic DNA from Bacteria. In Current Protocols in Molecular Biology (pp. 1–5). Townsville, Australia: John Wiley & Sons, Inc.
Winter, C., Hein, T., Kavka, G., Mach, R. L., & Farnleitner, A. H. (2007). Longitudinal changes in the bacterial community composition of the Danube River: a whole-river approach. Applied and Environmental Microbiology, 73, 421–431.
World of Microbiology and Immunology. (2003). Enterobacteriaceae. Retrieved January 4, 2015 from http://www.encyclopedia.com/doc/IG2-3409800194.html
World Weather & Climate Information. (2014). Average Weather and Climate in Fiji. Retrieved October 12, 2014 from https://weather-and-climate.com/average-monthly-Rainfall-Temperature-Sunshine-in-Fiji
WWF. (2013). Sustaining the Sweetness of the Sugar Industry. Suva. January 4, 2015 from http://www.wwfpacific.org.fj
Xu, J. (2006). Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances. Molecular Ecology, 15, 1713–1731.
Yadav, J., & Pathak, R. K. (2012). Analysis and Physico-Chemical Parameters of Sarvar Devla Sugar Mill Studies of Effluents. Current World Environment, 7, 313–315.
Yarza, P., Yilmaz, P., Pruesse, E., Glöckner, F. O., Ludwig, W., Schleifer, K. H., & Rosselló-Móra, R. (2014). Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nature Reviews Microbiology, 12, 635–645.
Yeo, S. (2001). A Review of Flooding in Macuata Province, Fiji Islands. SOPAC (pp. 1154) Suva.
Yeo, S., Blong, R. J., & McAneney, K. J. (2007). Flooding in Fiji: findings from a 100year historical series. Hydrological Sciences Journal, 52(5), 1004–1015.
Yogita, N. S. (2005). Viable but non-culturable bacteria: their impact on public health. Current Science, 89, 1650-1651.

78
Zeng, Y., Zhang, X., Nie, K., Ding, X., Ring, B. Z., Xu, L., & Ma, X. (2014). Rapid quantitative detection of Human immunodeficiency virus type 1 by a reverse transcription-loop-mediated isothermal amplification assay. Gene, 541, 123–128.
Zwart, G., Crump, B. C., Agterveld, M. P. K., Hagen, F., & Han, S. (2002). Typical freshwater bacteria: an analysis of available 16S rRNA gene sequences from plankton of lakes and rivers. Aquatic Microbial Ecology, 28, 141–155.
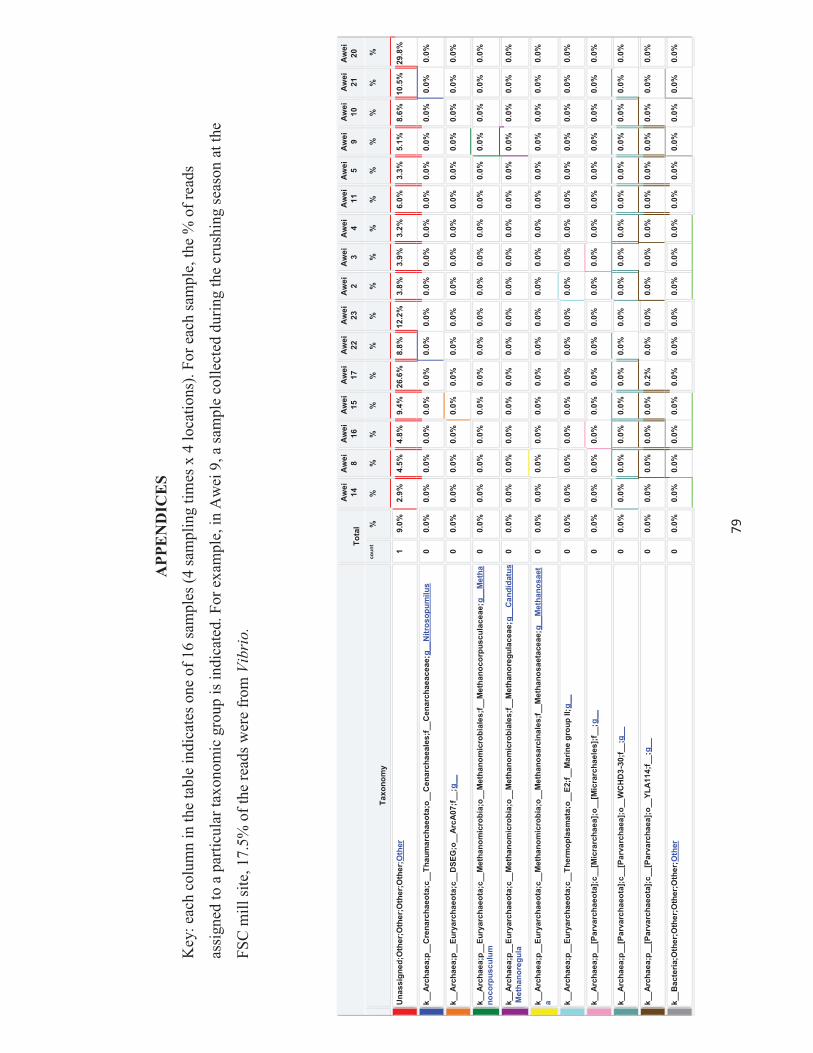
79
APP
EN
DIC
ES
Key
: eac
h co
lum
n in
the
tabl
e in
dica
tes o
ne o
f 16
sam
ples
(4 sa
mpl
ing
times
x 4
loca
tions
). Fo
r eac
h sa
mpl
e, th
e %
of r
eads
assi
gned
to a
par
ticul
ar ta
xono
mic
gro
up is
indi
cate
d. F
or e
xam
ple,
in A
wei
9, a
sam
ple
colle
cted
dur
ing
the
crus
hing
seas
on a
t the
FSC
mill
site
, 17.
5% o
f the
read
s wer
e fr
om V
ibri
o.
Tota
l A
wei
14
A
wei
8
Aw
ei
16
Aw
ei
15
Aw
ei
17
Aw
ei
22
Aw
ei
23
Aw
ei
2
Aw
ei
3
Aw
ei
4
Aw
ei
11
Aw
ei
5
Aw
ei
9
Aw
ei
10
Aw
ei
21
Aw
ei
20
Taxo
nom
y
coun
t %
%
%
%
%
%
%
%
%
%
%
%
%
%
%
%
%
Una
ssig
ned;
Oth
er;O
ther
;Oth
er;O
ther
;Oth
er
1
9.0%
2.
9%
4.5%
4.
8%
9.4%
26.
6%
8.8%
12.
2%
3.8%
3.
9%
3.2%
6.
0%
3.3%
5.
1%
8.6%
10.
5% 2
9.8%
k__A
rcha
ea;p
__C
rena
rcha
eota
;c__
Thau
mar
chae
ota;
o__C
enar
chae
ales
;f__C
enar
chae
acea
e;g_
_Nitr
osop
umilu
s
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__Eu
ryar
chae
ota;
c__D
SEG
;o__
Arc
A07
;f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__Eu
ryar
chae
ota;
c__M
etha
nom
icro
bia;
o__M
etha
nom
icro
bial
es;f_
_Met
hano
corp
uscu
lace
ae;g
__M
etha
no
corp
uscu
lum
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__Eu
ryar
chae
ota;
c__M
etha
nom
icro
bia;
o__M
etha
nom
icro
bial
es;f_
_Met
hano
regu
lace
ae;g
__C
andi
datu
s M
etha
nore
gula
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__Eu
ryar
chae
ota;
c__M
etha
nom
icro
bia;
o__M
etha
nosa
rcin
ales
;f__M
etha
nosa
etac
eae;
g__M
etha
nosa
et
a
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__Eu
ryar
chae
ota;
c__T
herm
opla
smat
a;o_
_E2;
f__M
arin
e gr
oup
II;g_
_
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__[P
arva
rcha
eota
];c__
[Mic
rarc
haea
];o__
[Mic
rarc
hael
es];f
__;g
__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__[P
arva
rcha
eota
];c__
[Par
varc
haea
];o__
WC
HD
3-30
;f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__A
rcha
ea;p
__[P
arva
rcha
eota
];c__
[Par
varc
haea
];o__
YLA
114;
f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
2%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;O
ther
;Oth
er;O
ther
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%

80
k__B
acte
ria;p
__;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
C1;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
C1;
c__H
DB
W-W
B69
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
AT-
s2-5
7;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
ia-5
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
ia-6
;o__
BPC
015;
f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
ia-6
;o__
CC
U21
;f__;
g__
0
0.0%
0.
0%
0.0%
0.
1%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
2%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
ia-6
;o__
iii1-
15;f_
_;g_
_
0
0.1%
0.
0%
0.0%
0.
3%
0.0%
0.
0%
0.1%
0.
6%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
2%
0.1%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
ia-6
;o__
iii1-
15;f_
_RB
40;g
__
0
0.5%
0.
0%
0.0%
0.
8%
0.3%
0.
2%
2.4%
0.
3%
0.0%
0.
0%
0.0%
0.
1%
0.0%
0.
0%
0.0%
4.
1%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
ia-6
;o__
iii1-
15;f_
_mb2
424;
g__
0
0.2%
0.
0%
0.0%
1.
5%
0.4%
0.
8%
0.3%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Aci
doba
cter
iia;o
__A
cido
bact
eria
les;
f__A
cido
bact
eria
ceae
;g__
Edap
hoba
cter
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Hol
opha
gae;
o__H
olop
haga
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Solib
acte
res;
o__S
olib
acte
rale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Solib
acte
res;
o__S
olib
acte
rale
s;f_
_PA
UC
26f;
g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.3%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
1%
0.0%
k__B
acte
ria;p
__A
cido
bact
eria
;c__
Solib
acte
res;
o__S
olib
acte
rale
s;f_
_Sol
ibac
tera
ceae
;g__
0
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
0.
0%
0.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__So
libac
tere
s;o_
_Sol
ibac
tera
les;
f__S
olib
acte
race
ae;g
__C
andi
datu
s So
libac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__So
libac
tere
s;o_
_Sol
ibac
tera
les;
f__[
Bry
obac
tera
ceae
];g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__Sv
a072
5;o_
_Sva
0725
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__TM
1;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__[C
hlor
acid
obac
teria
];o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

81
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__[C
hlor
acid
obac
teria
];o__
PK29
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__[C
hlor
acid
obac
teria
];o__
RB
41;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Aci
doba
cter
ia;c
__[C
hlor
acid
obac
teria
];o__
RB
41;f_
_Elli
n607
5;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__;
g__
0
0.2%
0.
5%
0.0%
0.8
% 0
.3%
0.0
% 0
.0%
0.0
% 0
.5%
0.4
% 0
.4%
0.1
% 0
.5%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__C
111;
g__
0
1.5%
0.
0%
0.7%
0.1
% 0
.1%
0.4
% 0
.1%
0.1
% 3
.6%
2.8
% 3
.6%
5.3
% 3
.5%
0.7
% 3
.1%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__E
B10
17;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__I
amia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__J
dFB
GB
act;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__M
icro
thrix
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__O
CS1
55;g
__
0
0.0%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__T
K06
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__Z
A34
09c;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__k
oll1
3;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
cidi
mic
robi
ia;o
__A
cidi
mic
robi
ales
;f__w
b1_P
06;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__;
g__
0
2.2%
0.
5%
2.4%
3.1
% 0
.5%
0.1
% 0
.1%
1.1
% 2
.2%
2.1
% 4
.3%
9.1
% 1
.4%
3.3
% 4
.2%
0.1
% 0
.8%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__A
CK
-M1;
g__
0
0.4%
0.
1%
0.0%
5.7
% 0
.7%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__A
ctin
osyn
nem
atac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__B
revi
bact
eria
ceae
;g__
Bre
viba
cter
ium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.4%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__C
ellu
lom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__C
ellu
lom
onad
acea
e;g_
_Dem
equi
na
0
0.1%
0.
0%
0.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.9
% 0
.1%
0.0
% 0
.0%

82
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__C
oryn
ebac
teria
ceae
;g__
Cor
yneb
acte
rium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__D
erm
abac
tera
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__D
erm
abac
tera
ceae
;g__
Bra
chyb
acte
rium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__D
erm
acoc
cace
ae;g
__D
erm
acoc
cus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__D
ietz
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__D
ietz
iace
ae;g
__D
ietz
ia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__F
rank
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__F
rank
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__G
ordo
niac
eae;
g__G
ordo
nia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__I
ntra
spor
angi
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__I
ntra
spor
angi
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;Oth
er
0
0.1%
0.
0%
0.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.1%
0.1
% 0
.3%
0.1
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
0
2.7%
0.
0%
8.4%
0.0
% 0
.0%
0.4
% 0
.0%
0.1
% 2
.2%
3.6
% 2
.8%
9.2
% 2
.5%
7.7
% 4
.7%
0.0
% 0
.7%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Agr
ococ
cus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Can
dida
tus
Aqu
ilu
na
0
2.8%
0.
1%
3.7%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 7
.6%
8.4
% 8
.6%
4.7
% 4
.6%
3.2
% 3
.3%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Can
dida
tus
Rho
do
luna
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Cla
viba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Leuc
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Mic
roba
cter
ium
0
0.1%
0.
0%
0.3%
0.0
% 0
.0%
0.2
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.0%
0.4
% 0
.3%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Myc
etoc
ola
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Pseu
docl
avib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

83
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Rat
hayi
bact
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
bact
eria
ceae
;g__
Salin
ibac
teriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
cocc
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
cocc
acea
e;g_
_Art
hrob
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
cocc
acea
e;g_
_Mic
roco
ccus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
cocc
acea
e;g_
_Nes
tere
nkon
ia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
icro
cocc
acea
e;g_
_Rot
hia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__M
ycob
acte
riace
ae;g
__M
ycob
acte
rium
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.6%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
akam
urel
lace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
ocar
diac
eae;
g__N
ocar
dia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
ocar
diac
eae;
g__R
hodo
cocc
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
ocar
dioi
dace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
ocar
dioi
dace
ae;g
__A
erom
icro
bium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
ocar
dioi
dace
ae;g
__N
ocar
dioi
des
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__N
ocar
dioi
dace
ae;g
__Pr
opio
nici
mon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__P
ropi
onib
acte
riace
ae;g
__Pr
opio
niba
cter
i um
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__P
seud
onoc
ardi
acea
e;g_
_Pra
user
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__P
seud
onoc
ardi
acea
e;g_
_Pse
udon
ocar
dia 0
0.
0%
0.0%
0.
0% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__S
trep
tom
ycet
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__S
trep
tom
ycet
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__S
trep
tom
ycet
acea
e;g_
_Str
epto
myc
es
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Act
inom
ycet
ales
;f__T
suka
mur
ella
ceae
;g__
Tsuk
amur
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

84
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__A
ctin
obac
teria
;o__
Bifi
doba
cter
iale
s;f_
_Bifi
doba
cter
iace
ae;g
__B
ifido
bact
eriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__C
orio
bact
eriia
;o__
Cor
ioba
cter
iale
s;f_
_Cor
ioba
cter
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__M
B-A
2-10
8;o_
_031
9-7L
14;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__O
PB41
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Gai
ella
les;
f__;
g__
0
0.1%
0.
3%
0.0%
0.2
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.1%
0.9
% 0
.1%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Gai
ella
les;
f__A
K1A
B1_
02E;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Gai
ella
les;
f__G
aiel
lace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Sol
irubr
obac
tera
les;
Oth
er;O
ther
0
0.3%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.5%
0.2
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
1.7
% 1
.4%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Sol
irubr
obac
tera
les;
f__;
g__
0
0.6%
0.
0%
0.0%
0.2
% 0
.3%
0.8
% 0
.1%
0.3
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
7.7
% 0
.3%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Sol
irubr
obac
tera
les;
f__C
onex
ibac
tera
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Sol
irubr
obac
tera
les;
f__C
onex
ibac
tera
ceae
;g__
Con
exib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Sol
irubr
obac
tera
les;
f__P
atul
ibac
tera
ceae
;g__
0
0.2%
0.
0%
0.0%
1.6
% 1
.5%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Act
inob
acte
ria;c
__Th
erm
oleo
phili
a;o_
_Sol
irubr
obac
tera
les;
f__S
oliru
brob
acte
race
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Anc
K6;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
0319
-6E2
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
Arm
atim
onad
ia;o
__A
rmat
imon
adal
es;f_
_Arm
atim
onad
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
Arm
atim
onad
ia;o
__A
rmat
imon
adal
es;f_
_Arm
atim
onad
acea
e;g_
_Arm
atim
ona
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
Cht
hono
mon
adet
es;o
__SJ
A-2
2;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
[Fim
briim
onad
ia];o
__[F
imbr
iimon
adal
es];f
__;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
[Fim
briim
onad
ia];o
__[F
imbr
iimon
adal
es];f
__[F
imbr
iimon
adac
eae]
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Arm
atim
onad
etes
;c__
[Fim
briim
onad
ia];o
__[F
imbr
iimon
adal
es];f
__[F
imbr
iimon
adac
eae]
;g__
Fim
brii
mon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

85
k_
_Bac
teria
;p__
BH
I80-
139;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
BR
C1;
c__N
PL-U
PA2;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.0%
k_
_Bac
teria
;p__
BR
C1;
c__P
RR
-11;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.6
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;O
ther
;Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
ME4
3;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__;
g__
0
0.3%
0.
0%
1.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
3.6
% 0
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__B
A00
8;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__B
acte
roid
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__B
acte
roid
acea
e;g_
_Bac
tero
ides
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__G
ZKB
119;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__M
L635
J-40
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__M
arin
ilabi
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__P
orph
yrom
onad
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__P
orph
yrom
onad
acea
e;g_
_Pal
udib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__P
orph
yrom
onad
acea
e;g_
_Par
abac
tero
ides
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__P
orph
yrom
onad
acea
e;g_
_Por
phyr
omon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__P
revo
tella
ceae
;g__
Prev
otel
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__R
iken
ella
ceae
;g__
Blv
ii28
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__S
B-1
;g__
0
0.1%
0.
0%
0.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__V
C21
_Bac
22;g
__
0
0.1%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.5%
0.0
% 0
.0%

86
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__B
acte
roid
ia;o
__B
acte
roid
ales
;f__[
Para
prev
otel
lace
ae];
g__[
Prev
otel
la]
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyc
loba
cter
iace
ae;g
__
0
0.2%
2.
0%
0.0%
0.0
% 1
.2%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyc
loba
cter
iace
ae;g
__A
lgor
ipha
gus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__
0
0.4%
0.
0%
0.0%
1.3
% 0
.5%
0.0
% 0
.8%
0.5
% 0
.2%
0.4
% 1
.4%
0.0
% 1
.0%
0.0
% 0
.1%
0.1
% 0
.3%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__D
yado
bact
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__Em
ticic
ia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__Fl
ecto
baci
llus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__Le
adbe
ttere
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__M
icro
scill
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Cyt
opha
gace
ae;g
__Po
ntib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
g__
0
0.1%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.4%
0.1
% 0
.1%
0.1
% 0
.0%
0.2
% 0
.1%
0.0
% 0
.1%
0.2
% 0
.2%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
g__F
lexi
bact
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
g__F
lexi
thrix
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
g__F
ulvi
virg
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
g__R
eich
enba
chie
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_Fla
mm
eovi
rgac
eae;
g__R
osei
virg
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__C
ytop
hagi
a;o_
_Cyt
opha
gale
s;f_
_[A
moe
boph
ilace
ae];
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__;
g__
0
0.3%
0.
3%
0.1%
0.1
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.1%
0.2
% 0
.1%
0.6
% 0
.2%
0.1
% 0
.5%
1.4
% 0
.1%

87
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__C
ryom
orph
acea
e;O
ther
0
0.0%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__C
ryom
orph
acea
e;g_
_
0
1.4%
10
.9%
2.
4% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
2.3
% 0
.4%
2.1
% 0
.4%
1.6
% 1
.7%
0.4
% 0
.0%
0.0
%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__C
ryom
orph
acea
e;g_
_Cro
cini
tom
ix
0
0.2%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
1.4
% 0
.0%
0.0
% 1
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__C
ryom
orph
acea
e;g_
_Flu
viic
ola
0
0.8%
4.
0%
0.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.6%
0.8
% 2
.7%
0.1
% 4
.5%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__C
ryom
orph
acea
e;g_
_Ow
enw
eeks
ia
0
0.0%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;Oth
er
0
0.1%
0.
4%
0.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.1%
0.2
% 0
.0%
0.1
% 0
.0%
0.1
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
0
1.3%
8.
4%
1.7%
0.0
% 0
.0%
0.0
% 0
.2%
0.1
% 1
.5%
1.7
% 1
.4%
2.2
% 1
.0%
2.0
% 1
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Aeq
uoriv
ita
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Cap
nocy
toph
aga
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Flav
obac
teriu
m
0
0.3%
0.
3%
0.1%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.9%
1.7
% 0
.8%
0.0
% 0
.8%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Gill
isia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Gra
mel
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Leeu
wen
hoek
iella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Mar
ibac
ter
0
0.0%
0.
3%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Mes
onia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Mur
icau
da
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
1.6
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Myr
oide
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Pola
ribac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Rob
igin
itale
a
0
0.1%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.6%
0.0
% 0
.4%
0.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Sale
gent
ibac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Sedi
min
icol
a
0
0.3%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
3.2
% 0
.1%
0.0
% 1
.3%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Suffl
avib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

88
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Tena
ciba
culu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__F
lavo
bact
eria
ceae
;g__
Win
ogra
dsky
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__[
Wee
ksel
lace
ae];
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__[
Wee
ksel
lace
ae];
g__C
hrys
eoba
cter
ium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__[
Wee
ksel
lace
ae];
g__C
loac
ibac
teriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Fl
avob
acte
riia;
o__F
lavo
bact
eria
les;
f__[
Wee
ksel
lace
ae];
g__W
aute
rsie
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
Oth
er;O
ther
0
0.0%
0.
1%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__;
g__
0
1.0%
0.
2%
0.1%
3.1
% 1
.0%
0.0
% 1
.7%
0.0
% 0
.4%
0.2
% 2
.1%
0.1
% 1
.2%
0.1
% 0
.4%
4.6
% 0
.3%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__N
S11-
12;g
__
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.5%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
2.1
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__S
phin
goba
cter
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.1
% 0
.0%
0.2
% 0
.1%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__S
phin
goba
cter
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__S
phin
goba
cter
iace
ae;g
__Pa
rape
dob
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__S
phin
goba
cter
iace
ae;g
__Pe
doba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__Sp
hing
obac
terii
a;o_
_Sph
ingo
bact
eria
les;
f__S
phin
goba
cter
iace
ae;g
__Sp
hing
oba
cter
ium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[R
hodo
ther
mi];
o__[
Rho
doth
erm
ales
];f__
Rho
doth
erm
acea
e;g_
_
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
1.1
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[R
hodo
ther
mi];
o__[
Rho
doth
erm
ales
];f__
[Bal
neol
acea
e];O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[R
hodo
ther
mi];
o__[
Rho
doth
erm
ales
];f__
[Bal
neol
acea
e];g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[R
hodo
ther
mi];
o__[
Rho
doth
erm
ales
];f__
[Bal
neol
acea
e];g
__B
alne
ola
0
0.1%
0.
0%
0.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
0.1
% 0
.4%
0.0
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.5%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__;
g__
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__C
hitin
opha
gace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

89
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__C
hitin
opha
gace
ae;g
__
0
0.1%
0.
0%
0.0%
1.3
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.3%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__C
hitin
opha
gace
ae;g
__C
hitin
opha
ga
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.5
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__C
hitin
opha
gace
ae;g
__Se
dim
inib
acte
rium
0
0.1%
0.
0%
0.0%
1.4
% 0
.2%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__S
apro
spira
ceae
;g__
0
0.1%
0.
0%
0.4%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 1
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__S
apro
spira
ceae
;g__
Hal
isco
men
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__S
apro
spira
ceae
;g__
Lew
inel
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Bac
tero
idet
es;c
__[S
apro
spira
e];o
__[S
apro
spira
les]
;f__S
apro
spira
ceae
;g__
Sapr
ospi
ra
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cal
dith
rix;c
__C
aldi
thrix
ae;o
__C
aldi
thrix
ales
;f__B
A05
9;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cal
dith
rix;c
__C
aldi
thrix
ae;o
__C
aldi
thrix
ales
;f__C
aldi
thrix
acea
e;g_
_LC
P-26
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_;g_
_
0
0.7%
0.
3%
0.0%
0.6
% 0
.5%
1.7
% 0
.7%
0.1
% 0
.1%
0.2
% 0
.3%
0.0
% 0
.3%
0.0
% 0
.0%
0.0
% 7
.2%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Chl
amyd
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Chl
amyd
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Chl
amyd
iace
ae;g
__C
hlam
ydia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Crib
lam
ydia
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Crib
lam
ydia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Crib
lam
ydia
ceae
;g__
Estr
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Par
achl
amyd
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Par
achl
amyd
iace
ae;g
__
0
0.1%
0.
0%
0.0%
0.0
% 0
.1%
1.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Par
achl
amyd
iace
ae;g
__C
andi
datu
s Pr
otoc
hlam
ydi
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Par
achl
amyd
iace
ae;g
__Pa
rach
lam
ydia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

90
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Rha
bdoc
hlam
ydia
ceae
;g__
Can
dida
tus
Rha
bdoc
hla
myd
ia
0
1.1%
0.
1%
0.0%
0.3
% 1
.9%
10.
9% 0
.2%
0.0
% 0
.1%
0.1
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.9
% 2
.1%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Sim
kani
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Sim
kani
acea
e;g_
_Sim
kani
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Wad
dlia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
amyd
iae;
c__C
hlam
ydiia
;o__
Chl
amyd
iale
s;f_
_Wad
dlia
ceae
;g__
Wad
dlia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
BSV
26;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
BSV
26;o
__A
89;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
BSV
26;o
__C
20;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
BSV
26;o
__VC
38;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.5
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
Chl
orob
ia;o
__C
hlor
obia
les;
f__C
hlor
obia
ceae
;g__
Chl
orob
acul
um
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
Igna
viba
cter
ia;o
__Ig
navi
bact
eria
les;
f__I
gnav
ibac
teria
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.2%
k_
_Bac
teria
;p__
Chl
orob
i;c__
Igna
viba
cter
ia;o
__Ig
navi
bact
eria
les;
f__[
Mel
iorib
acte
race
ae];
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
Igna
viba
cter
ia;o
__Ig
navi
bact
eria
les;
f__l
heB
3-7;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orob
i;c__
OPB
56;o
__;f_
_;g_
_
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 1
.7%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.6
% 0
.1%
k_
_Bac
teria
;p__
Chl
orob
i;c__
SJA
-28;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_Ana
erol
inea
les;
f__A
naer
olin
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_CFB
-26;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_Cal
dilin
eale
s;f_
_Cal
dilin
eace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_Cal
dilin
eale
s;f_
_Cal
dilin
eace
ae;g
__C
aldi
linea
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

91
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_GC
A00
4;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_H39
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_S02
08;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_SB
R10
31;f_
_A4b
;g__
0
0.1%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.7%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_SB
R10
31;f_
_SJA
-101
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_SH
A-2
0;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_SJA
-15;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ana
erol
inea
e;o_
_env
OPS
12;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Chl
orof
lexi
;o__
AK
IW78
1;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Chl
orof
lexi
;o__
[Ros
eifle
xale
s];f_
_;g_
_
0
1.5%
3.
5%
0.0%
12.
6% 7
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Deh
aloc
occo
idet
es;o
__D
ehal
ococ
coid
ales
;f__D
ehal
ococ
coid
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Deh
aloc
occo
idet
es;o
__G
IF9;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ellin
6529
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Kte
dono
bact
eria
;o__
Ther
mog
emm
atis
pora
les;
f__T
herm
ogem
mat
ispo
race
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
MVP
-21;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
S085
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
SAR
202;
o__;
f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.5
% 0
.5%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
SHA
-26;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
TK10
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
TK10
;o__
AK
YG88
5;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
TK17
;o__
;f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.7%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.1%

92
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
TK17
;o__
mle
1-48
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
k_
_Bac
teria
;p__
Chl
orof
lexi
;c__
Ther
mom
icro
bia;
o__J
G30
-KF-
CM
45;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__4C
0d-2
;o__
MLE
1-12
;f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.1%
0.1
% 0
.3%
1.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__4C
0d-2
;o__
SM1D
11;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__4C
0d-2
;o__
SM2F
09;f_
_;g_
_
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 1
.6%
0.0
% 0
.2%
0.3
% 0
.4%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__4C
0d-2
;o__
YS2;
f__;
g__
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__C
hlor
opla
st;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__C
hlor
opla
st;o
__C
hlor
ophy
ta;f_
_;g_
_
0
0.1%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.6%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__C
hlor
opla
st;o
__C
rypt
ophy
ta;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__C
hlor
opla
st;o
__Eu
glen
ozoa
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__C
hlor
opla
st;o
__St
ram
enop
iles;
f__;
g__
0
0.2%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 2
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__C
hlor
opla
st;o
__St
rept
ophy
ta;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__M
L635
J-21
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__O
scill
ator
ioph
ycid
eae;
o__O
scill
ator
iale
s;f_
_Pho
rmid
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__O
scill
ator
ioph
ycid
eae;
o__O
scill
ator
iale
s;f_
_Pho
rmid
iace
ae;g
__O
scill
ator
ia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__O
scill
ator
ioph
ycid
eae;
o__O
scill
ator
iale
s;f_
_Pho
rmid
iace
ae;g
__Ph
orm
idiu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__Sy
nech
ococ
coph
ycid
eae;
Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__Sy
nech
ococ
coph
ycid
eae;
o__S
ynec
hoco
ccal
es;f_
_Syn
echo
cocc
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__Sy
nech
ococ
coph
ycid
eae;
o__S
ynec
hoco
ccal
es;f_
_Syn
echo
cocc
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Cya
noba
cter
ia;c
__Sy
nech
ococ
coph
ycid
eae;
o__S
ynec
hoco
ccal
es;f_
_Syn
echo
cocc
acea
e;g_
_Syn
ec
hoco
ccus
0
1.0%
0.
0%
4.6%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.1%
1.7
% 0
.1%
5.0
% 4
.5%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Def
errib
acte
res;
c__D
efer
ribac
tere
s;o_
_Def
errib
acte
rale
s;f_
_Def
errib
acte
race
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

93
k_
_Bac
teria
;p__
Def
errib
acte
res;
c__D
efer
ribac
tere
s;o_
_Def
errib
acte
rale
s;f_
_Def
errib
acte
race
ae;g
__Fl
exis
tipes
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Def
errib
acte
res;
c__D
efer
ribac
tere
s;o_
_Def
errib
acte
rale
s;f_
_V1B
07b9
3;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Elus
imic
robi
a;c_
_Elu
sim
icro
bia;
o__E
lusi
mic
robi
ales
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Elus
imic
robi
a;c_
_Elu
sim
icro
bia;
o__F
AC
88;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Elus
imic
robi
a;c_
_Elu
sim
icro
bia;
o__I
Ib;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Elus
imic
robi
a;c_
_Elu
sim
icro
bia;
o__M
VP-8
8;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Elus
imic
robi
a;c_
_End
omic
robi
a;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
FBP;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
FCPU
426;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fibr
obac
tere
s;c_
_Fib
roba
cter
ia;o
__Fi
brob
acte
rale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fibr
obac
tere
s;c_
_Fib
roba
cter
ia;o
__Fi
brob
acte
rale
s;f_
_Fib
roba
cter
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Alic
yclo
baci
llace
ae;g
__A
licyc
loba
cillu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__A
noxy
baci
llus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__B
acill
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.6
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__G
eoba
cillu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__M
arin
ococ
cus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__O
cean
obac
illus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Bac
illac
eae;
g__T
hala
ssob
acill
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Pae
niba
cilla
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

94
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Pae
niba
cilla
ceae
;g__
Paen
ibac
illus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Pla
noco
ccac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Pla
noco
ccac
eae;
g__L
ysin
ibac
illus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Pla
noco
ccac
eae;
g__P
lano
cocc
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Sta
phyl
ococ
cace
ae;g
__Je
otga
licoc
cus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_Sta
phyl
ococ
cace
ae;g
__St
aphy
loco
ccus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_The
rmoa
ctin
omyc
etac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_[Ex
iguo
bact
erac
eae]
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Bac
illal
es;f_
_[Ex
iguo
bact
erac
eae]
;g__
Exig
uoba
cter
ium
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Gem
ella
les;
f__G
emel
lace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Aer
ococ
cace
ae;g
__A
eroc
occu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Aer
ococ
cace
ae;g
__Fa
ckla
mia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Ent
eroc
occa
ceae
;g__
Ente
roco
ccus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Lac
toba
cilla
ceae
;g__
Lact
obac
illus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Leu
cono
stoc
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Leu
cono
stoc
acea
e;g_
_Leu
cono
stoc
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__B
acill
i;o__
Lact
obac
illal
es;f_
_Str
epto
cocc
acea
e;g_
_Str
epto
cocc
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Chr
iste
nsen
ella
ceae
;g__
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

95
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;g
__A
lkal
iphi
lus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;g
__C
lost
ridiis
alib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;g
__C
lost
ridiu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;g
__Th
erm
oana
erob
acte
rium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Clo
strid
iace
ae;g
__Ti
ndal
lia_A
noxy
natr
onum
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Gra
cilib
acte
race
ae;g
__G
raci
libac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Lac
hnos
pira
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Lac
hnos
pira
ceae
;g__
0
0.1%
0.
0%
1.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
1.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Lac
hnos
pira
ceae
;g__
Cop
roco
ccus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Lac
hnos
pira
ceae
;g__
Epul
opis
cium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Pep
toco
ccac
eae;
g__D
ehal
obac
ter_
Synt
roph
obot
ulus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Pep
toco
ccac
eae;
g__D
esul
fosp
oros
inus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Pep
toco
ccac
eae;
g__D
esul
foto
mac
ulum
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Rum
inoc
occa
ceae
;g__
0
0.0%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Rum
inoc
occa
ceae
;g__
Osc
illos
pira
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Syn
trop
hom
onad
acea
e;g_
_Syn
trop
hom
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Vei
llone
llace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Vei
llone
llace
ae;g
__M
egas
phae
ra
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Vei
llone
llace
ae;g
__Ve
illon
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_Vei
llone
llace
ae;g
__va
dinH
B04
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[A
cida
min
obac
tera
ceae
];O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

96
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[A
cida
min
obac
tera
ceae
];g_
_Aci
dam
inob
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[A
cida
min
obac
tera
ceae
];g_
_Fus
ibac
ter
0
0.1%
0.
0%
0.5%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.4
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[A
cida
min
obac
tera
ceae
];g_
_WH
1-8
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[M
ogib
acte
riace
ae];
g__A
naer
ovor
ax
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[Ti
ssie
rella
ceae
];g_
_Ana
eroc
occu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Clo
strid
iale
s;f_
_[Ti
ssie
rella
ceae
];g_
_Fin
egol
dia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__C
lost
ridia
;o__
Ther
moa
naer
obac
tera
les;
f__C
aldi
cellu
losi
rupt
orac
eae;
g__C
aldi
cellu
lo
siru
ptor
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__Er
ysip
elot
richi
;o__
Erys
ipel
otric
hale
s;f_
_Ery
sipe
lotr
icha
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__Er
ysip
elot
richi
;o__
Erys
ipel
otric
hale
s;f_
_Ery
sipe
lotr
icha
ceae
;g__
PSB
-M-3
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Firm
icut
es;c
__Er
ysip
elot
richi
;o__
Erys
ipel
otric
hale
s;f_
_Ery
sipe
lotr
icha
ceae
;g__
RFN
20
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fuso
bact
eria
;c__
Fuso
bact
eriia
;o__
Fuso
bact
eria
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fuso
bact
eria
;c__
Fuso
bact
eriia
;o__
Fuso
bact
eria
les;
f__F
usob
acte
riace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fuso
bact
eria
;c__
Fuso
bact
eriia
;o__
Fuso
bact
eria
les;
f__F
usob
acte
riace
ae;g
__C
etob
acte
rium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fuso
bact
eria
;c__
Fuso
bact
eriia
;o__
Fuso
bact
eria
les;
f__F
usob
acte
riace
ae;g
__Fu
soba
cter
ium
0
0.0%
0.
0%
0.4%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Fuso
bact
eria
;c__
Fuso
bact
eriia
;o__
Fuso
bact
eria
les;
f__F
usob
acte
riace
ae;g
__u1
14
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
GN
02;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
GN
02;c
__3B
R-5
F;o_
_;f_
_;g_
_
0
1.4%
0.
0%
0.8%
0.2
% 0
.0%
0.2
% 0
.0%
4.0
% 0
.0%
0.0
% 0
.0%
7.5
% 0
.0%
3.1
% 6
.8%
0.2
% 0
.0%
k_
_Bac
teria
;p__
GN
02;c
__B
B34
;o__
;f__;
g__
0
0.3%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
5.2
% 0
.1%
k_
_Bac
teria
;p__
GN
02;c
__B
D1-
5;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
GN
02;c
__G
KS2
-174
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.2
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
GN
02;c
__G
N07
;o__
;f__;
g__
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.8%
1.9
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
GN
02;c
__IIB
17;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

97
k_
_Bac
teria
;p__
GN
02;c
__SB
YW_2
184;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
GN
04;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
GN
04;c
__G
N15
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
-1;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
-2;o
__;f_
_;g_
_
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.6%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
3.1
% 0
.1%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
atim
onad
etes
;Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
atim
onad
etes
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
atim
onad
etes
;o__
C11
4;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
atim
onad
etes
;o__
Gem
mat
imon
adal
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
atim
onad
etes
;o__
Gem
mat
imon
adal
es;f_
_Gem
mat
imon
adac
eae;
g__
Gem
mat
imon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__G
emm
atim
onad
etes
;o__
KD
8-87
;f__;
g__
0
0.0%
0.
0%
0.0%
0.2
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Gem
mat
imon
adet
es;c
__JL
-ETN
P-Z3
9;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
H-1
78;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Kaz
an-3
B-2
8;c_
_;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Lent
isph
aera
e;c_
_[Le
ntis
phae
ria];O
ther
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Lent
isph
aera
e;c_
_[Le
ntis
phae
ria];o
__Le
ntis
phae
rale
s;f_
_Arc
tic95
B-1
0;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Lent
isph
aera
e;c_
_[Le
ntis
phae
ria];o
__Le
ntis
phae
rale
s;f_
_Len
tisph
aera
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Lent
isph
aera
e;c_
_[Le
ntis
phae
ria];o
__Vi
ctiv
alla
les;
f__V
ictiv
alla
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Lent
isph
aera
e;c_
_[Le
ntis
phae
ria];o
__Z2
0;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Lent
isph
aera
e;c_
_[Le
ntis
phae
ria];o
__Z2
0;f_
_R4-
45B
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
NK
B19
;c__
;o__
;f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.5
% 0
.0%

98
k_
_Bac
teria
;p__
NK
B19
;c__
SHA
B59
0;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
NK
B19
;c__
TSB
W08
;o__
;f__;
g__
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
NK
B19
;c__
noFP
_H4;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Nitr
ospi
rae;
c__N
itros
pira
;o__
Nitr
ospi
rale
s;f_
_Nitr
ospi
race
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Nitr
ospi
rae;
c__N
itros
pira
;o__
Nitr
ospi
rale
s;f_
_Nitr
ospi
race
ae;g
__N
itros
pira
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Nitr
ospi
rae;
c__N
itros
pira
;o__
Nitr
ospi
rale
s;f_
_[Th
erm
odes
ulfo
vibr
iona
ceae
];g_
_BD
2-6
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Nitr
ospi
rae;
c__N
itros
pira
;o__
Nitr
ospi
rale
s;f_
_[Th
erm
odes
ulfo
vibr
iona
ceae
];g_
_LC
P-6
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OC
31;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OD
1;c_
_;o_
_;f_
_;g_
_
0
0.1%
0.
1%
0.0%
0.2
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.3%
k_
_Bac
teria
;p__
OD
1;c_
_AB
Y1;o
__;f_
_;g_
_
0
0.9%
0.
3%
1.2%
0.3
% 0
.5%
2.4
% 0
.6%
3.1
% 0
.1%
0.2
% 0
.1%
2.2
% 0
.1%
1.5
% 1
.4%
0.8
% 0
.3%
k_
_Bac
teria
;p__
OD
1;c_
_Mb-
NB
09;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OD
1;c_
_SM
2F11
;o__
;f__;
g__
0
0.1%
0.
1%
0.0%
0.1
% 0
.2%
1.5
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OD
1;c_
_ZB
2;o_
_;f_
_;g_
_
0
2.8%
0.
6%
0.3%
3.3
% 3
.5%
6.0
% 4
.1%
7.3
% 0
.3%
0.8
% 0
.8%
0.3
% 1
.5%
0.3
% 0
.5%
8.4
% 7
.1%
k_
_Bac
teria
;p__
OP1
1;c_
_;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_OP1
1-1;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_OP1
1-2;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_OP1
1-2;
o__W
CH
B1-
07;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_OP1
1-3;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_OP1
1-4;
o__;
f__;
g__
0
0.2%
0.
0%
0.0%
0.0
% 0
.4%
2.8
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_WC
HB
1-64
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_WC
HB
1-64
;o__
K2-
4-19
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP1
1;c_
_WC
HB
1-64
;o__
d153
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.1%
0.2
% 0
.0%

99
k_
_Bac
teria
;p__
OP3
;c__
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP3
;c__
BD
4-9;
o__;
f__;
g__
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP3
;c__
PBS-
25;o
__;f_
_;g_
_
0
0.0%
0.
1%
0.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP3
;c__
koll1
1;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP3
;c__
koll1
1;o_
_GIF
10;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
OP3
;c__
koll1
1;o_
_GIF
10;f_
_kpj
58rc
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
PAU
C34
f;c__
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__02
8H05
-P-B
N-P
5;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__B
D7-
11;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__C
6;o_
_MVS
-107
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__C
6;o_
_d11
3;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__O
M19
0;o_
_CL5
00-1
5;f_
_;g_
_
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__O
M19
0;o_
_agg
27;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__A
KA
U35
64;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__C
CM
11a;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__M
SBL9
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__P
hyci
spha
eral
es;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.3
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__P
hyci
spha
eral
es;f_
_Phy
cisp
haer
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__S
-70;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__W
D21
01;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

100
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Ph
ycis
phae
rae;
o__m
le1-
8;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
a3;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
a4;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
B97
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Gem
mat
ales
;f__G
emm
atac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Gem
mat
ales
;f__G
emm
atac
eae;
g__G
emm
ata
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Gem
mat
ales
;f__I
sosp
haer
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Pire
llula
les;
f__P
irellu
lace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Pire
llula
les;
f__P
irellu
lace
ae;g
__
0
1.1%
0.
1%
0.1%
0.1
% 0
.1%
0.9
% 0
.1%
0.0
% 2
.1%
2.7
% 0
.5%
8.7
% 0
.7%
0.1
% 1
.6%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Pire
llula
les;
f__P
irellu
lace
ae;g
__A
17
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Pire
llula
les;
f__P
irellu
lace
ae;g
__Pi
rellu
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__Pl
anct
omyc
etia
;o__
Plan
ctom
ycet
ales
;f__P
lanc
tom
ycet
acea
e;g_
_Pla
ncto
myc
es
0
0.2%
0.
1%
0.0%
0.1
% 0
.1%
0.1
% 0
.3%
0.9
% 0
.1%
0.1
% 0
.6%
0.0
% 0
.4%
0.0
% 0
.0%
0.4
% 0
.1%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__va
dinH
A49
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__va
dinH
A49
;o__
DH
61;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__va
dinH
A49
;o__
PHO
S-H
E93;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__va
dinH
A49
;o__
PeH
g47;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Plan
ctom
ycet
es;c
__va
dinH
A49
;o__
p04_
C01
;f__;
g__
0
0.0%
0.
0%
0.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;O
ther
;Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;O
ther
;Oth
er;O
ther
0
0.2%
0.
0%
0.1%
0.0
% 0
.0%
0.1
% 0
.4%
1.5
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.3
% 0
.1%
0.2
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__;f_
_;g_
_
1
3.5%
0.
0%
0.1%
0.7
% 0
.8%
3.7
% 6
.3%
7.9
% 6
.7%
6.7
% 6
.3%
0.2
% 7
.6%
0.1
% 0
.0%
4.7
% 4
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__B
D7-
3;f_
_;g_
_
0
0.5%
0.
5%
0.2%
2.2
% 0
.5%
1.0
% 0
.5%
0.5
% 0
.2%
0.1
% 0
.2%
0.2
% 0
.2%
0.2
% 0
.4%
0.6
% 0
.4%
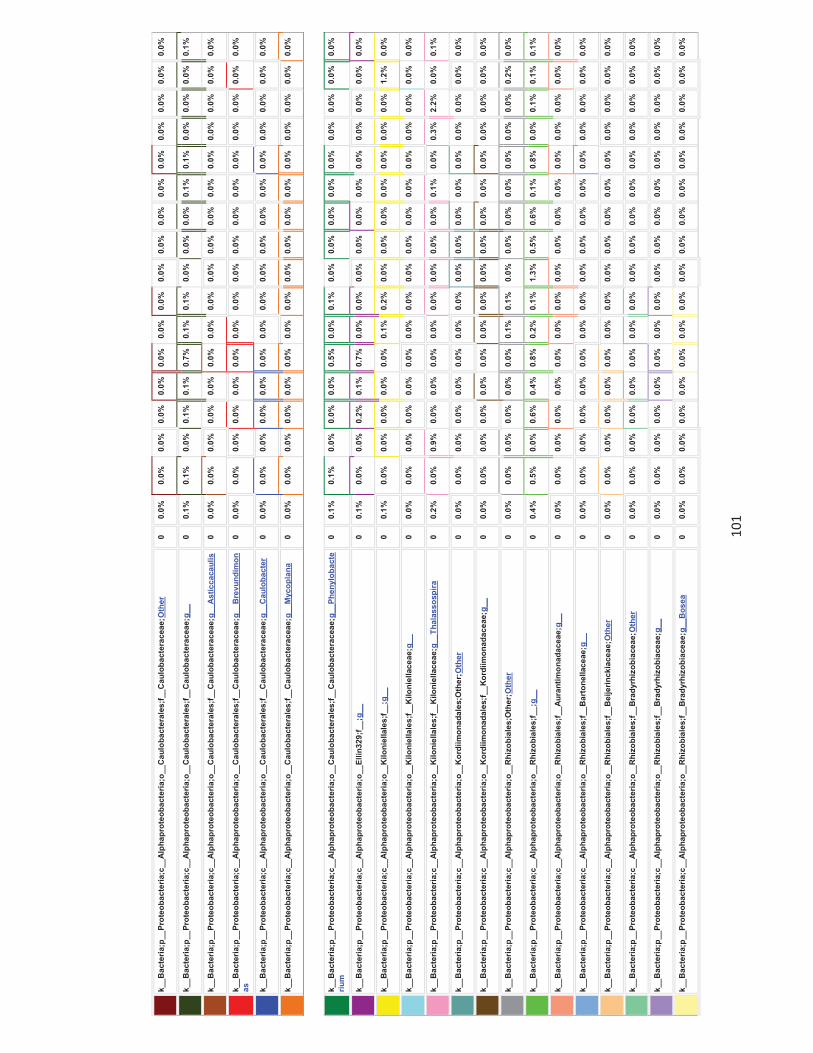
101
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;g_
_
0
0.1%
0.
1%
0.0%
0.1
% 0
.1%
0.7
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;g_
_Ast
icca
caul
is
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;g_
_Bre
vund
imon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;g_
_Cau
loba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;g_
_Myc
opla
na
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__C
aulo
bact
eral
es;f_
_Cau
loba
cter
acea
e;g_
_Phe
nylo
bact
e riu
m
0
0.1%
0.
1%
0.0%
0.0
% 0
.0%
0.5
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__El
lin32
9;f_
_;g_
_
0
0.1%
0.
0%
0.0%
0.2
% 0
.1%
0.7
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__K
iloni
ella
les;
f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
1.2
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__K
iloni
ella
les;
f__K
iloni
ella
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__K
iloni
ella
les;
f__K
iloni
ella
ceae
;g__
Thal
asso
spira
0
0.2%
0.
0%
0.9%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.3
% 2
.2%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__K
ordi
imon
adal
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__K
ordi
imon
adal
es;f_
_Kor
diim
onad
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_;g_
_
0
0.4%
0.
5%
0.0%
0.6
% 0
.4%
0.8
% 0
.2%
0.1
% 1
.3%
0.5
% 0
.6%
0.1
% 0
.8%
0.0
% 0
.1%
0.1
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Aur
antim
onad
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bar
tone
llace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bei
jerin
ckia
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bra
dyrh
izob
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bra
dyrh
izob
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bra
dyrh
izob
iace
ae;g
__B
osea
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

102
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bra
dyrh
izob
iace
ae;g
__B
rady
rhiz
obiu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bru
cella
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Bru
cella
ceae
;g__
Och
roba
ctru
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Coh
aesi
bact
erac
eae;
g__C
ohae
siba
cter
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.5
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__D
evos
ia
0
0.1%
0.
1%
0.0%
0.2
% 0
.1%
0.2
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.0%
0.5
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__H
ypho
mic
robi
u m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__P
arvi
bacu
lum
0
0.9%
0.
0%
0.0%
0.0
% 0
.1%
0.7
% 5
.8%
2.0
% 0
.1%
0.1
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
3.3
% 2
.4%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__P
edom
icro
bium
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__R
hodo
bium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Hyp
hom
icro
biac
eae;
g__R
hodo
plan
es
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.5
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Met
hylo
bact
eria
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Met
hylo
bact
eria
ceae
;g__
Met
hylo
bact
eri
um
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Met
hylo
cyst
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Met
hylo
cyst
acea
e;g_
_Met
hylo
pila
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Met
hylo
cyst
acea
e;g_
_Met
hylo
sinu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Met
hylo
cyst
acea
e;g_
_Ple
omor
phom
ona
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;O
ther
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 1
.7%
0.9
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.3%
0.6
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;g
__
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 1
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.1%
1.4
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;g
__A
min
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;g
__C
hela
tivor
ans
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

103
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;g
__M
esor
hizo
bium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;g
__N
itrat
iredu
ctor
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Phy
lloba
cter
iace
ae;g
__Ph
yllo
bact
eriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Rhi
zobi
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Rhi
zobi
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Rhi
zobi
acea
e;g_
_Agr
obac
teriu
m
0
0.1%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.7%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Rhi
zobi
acea
e;g_
_Kai
stia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Rho
dobi
acea
e;g_
_Afif
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hizo
bial
es;f_
_Xan
thob
acte
race
ae;g
__Xa
ntho
bact
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Hyp
hom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Hyp
hom
onad
acea
e;g_
_
0
0.8%
1.
5%
0.1%
0.7
% 0
.4%
0.1
% 4
.9%
2.8
% 0
.0%
0.3
% 0
.2%
0.1
% 0
.3%
0.0
% 0
.1%
0.3
% 0
.3%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Hyp
hom
onad
acea
e;g_
_Hirs
chia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Hyp
hom
onad
acea
e;g_
_Hyp
hom
on
as
0
0.2%
0.
1%
0.1%
0.0
% 0
.0%
0.0
% 0
.4%
0.3
% 0
.0%
0.2
% 0
.1%
0.7
% 0
.1%
0.0
% 0
.6%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Hyp
hom
onad
acea
e;g_
_Mar
icau
lis
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
1.6
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Hyp
hom
onad
acea
e;g_
_Oce
anic
auli
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;O
ther
0
0.2%
0.
1%
1.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.2
% 0
.2%
0.1
% 0
.1%
0.2
% 0
.8%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_
1
3.3%
0.
6%
20.4
%
0.0%
0.1
% 0
.0%
0.1
% 0
.4%
2.5
% 2
.1%
2.9
% 6
.4%
2.2
% 6
.2%
9.1
% 0
.1%
0.2
%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Ana
eros
por
a
1
3.2%
2.
3%
11.3
%
1.2%
4.8
% 0
.0%
0.0
% 2
.6%
4.0
% 4
.5%
7.8
% 0
.1%
6.6
% 4
.2%
1.2
% 0
.0%
0.0
%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Lok
tane
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Mar
ivita
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%

104
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Oct
adec
aba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Par
acoc
cus
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Pha
eoba
cte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Rho
doba
cte
r 0
0.9%
10
.4%
0.
0% 0
.2%
2.0
% 0
.6%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
0.3
% 0
.1%
0.1
% 0
.0%
0.0
%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Rho
dovu
lu
m
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Tha
lass
obiu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
bact
eral
es;f_
_Rho
doba
cter
acea
e;g_
_Tro
pici
bact
er
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Ace
toba
cter
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Ace
toba
cter
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Ace
toba
cter
acea
e;g_
_Ros
eom
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
1
3.5%
0.
7%
0.2%
2.9
% 2
.0%
7.1
% 4
.4%
13.
9%
4.0%
1.1
% 2
.3%
1.3
% 2
.3%
0.3
% 1
.3%
5.3
% 7
.5%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Azo
spiri
llum
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Inqu
ilinu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Mag
neto
spiri
l lu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Nis
aea
0
0.1%
0.
0%
0.6%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.5
% 0
.0%
0.2
% 0
.8%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Nov
ispi
rillu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Ole
omon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Phae
ospi
rillu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

105
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Rho
dosp
irillu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Rho
dovi
brio
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
hodo
spiri
llale
s;f_
_Rho
dosp
irilla
ceae
;g__
Sker
man
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__;
g__
0
0.2%
0.
2%
0.1%
0.5
% 0
.3%
0.0
% 0
.1%
0.1
% 0
.3%
0.3
% 0
.3%
0.3
% 0
.3%
0.1
% 0
.3%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__A
EGEA
N_1
12;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__A
napl
asm
atac
eae;
g__N
eoric
ketts
ia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__H
olos
pora
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__P
elag
ibac
tera
ceae
;g__
0
0.3%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
1.8
% 1
.3%
0.8
% 0
.3%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 1
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__R
icke
ttsia
ceae
;g__
0
0.4%
0.
2%
0.2%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
1.4
% 0
.1%
0.5
% 4
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__R
icke
ttsia
ceae
;g__
Orie
ntia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__R
icke
ttsia
les;
f__m
itoch
ondr
ia;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_;g_
_
0
0.0%
0.
3%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Ery
thro
bact
erac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Ery
thro
bact
erac
eae;
g__
0
0.2%
0.
9%
0.0%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.6%
0.1
% 0
.5%
0.4
% 0
.4%
0.0
% 0
.0%
0.0
% 0
.5%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Ery
thro
bact
erac
eae;
g__C
itrom
ic
robi
um
0
0.1%
0.
0%
0.0%
0.2
% 0
.1%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Ery
thro
bact
erac
eae;
g__E
ryth
rob
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Ery
thro
bact
erac
eae;
g__L
utib
act
eriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
g__
0
0.1%
0.
2%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.2%
0.0
% 0
.2%
0.0
% 0
.0%
0.3
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
g__K
aist
ob
acte
r 0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.0%

106
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
g__N
ovos
p hi
ngob
ium
0
0.8%
6.
0%
0.0%
0.6
% 4
.9%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.3%
0.2
% 1
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
g__S
phin
g ob
ium
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
1.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
g__S
phin
g om
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__A
lpha
prot
eoba
cter
ia;o
__Sp
hing
omon
adal
es;f_
_Sph
ingo
mon
adac
eae;
g__S
phin
g op
yxis
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;Oth
er;O
ther
;Oth
er
0
0.1%
0.
1%
0.0%
0.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
;f__;
g__
0
1.4%
0.
9%
0.0%
1.6
% 1
.7%
3.5
% 0
.0%
0.0
% 3
.9%
3.7
% 3
.3%
0.0
% 3
.3%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
ASS
O-1
3;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__;
g__
0
0.1%
0.
1%
0.0%
0.7
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__A
lcal
igen
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__A
lcal
igen
acea
e;g_
_
0
0.4%
0.
0%
0.7%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
2.2
% 0
.0%
0.5
% 2
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__A
lcal
igen
acea
e;g_
_Ach
rom
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__A
lcal
igen
acea
e;g_
_Tet
rath
ioba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__B
urkh
olde
riace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__B
urkh
olde
riace
ae;g
__B
urkh
olde
ria
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__B
urkh
olde
riace
ae;g
__Sa
linis
pora
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;O
ther
0
1.3%
5.
2%
0.0%
3.6
% 4
.1%
0.1
% 0
.0%
0.0
% 0
.7%
2.9
% 1
.9%
0.1
% 2
.4%
0.1
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_
0
1.9%
5.
8%
0.3%
2.7
% 4
.9%
0.2
% 0
.0%
0.0
% 1
.9%
2.8
% 3
.4%
0.7
% 5
.3%
0.2
% 1
.7%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Aci
dovo
rax
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Com
amon
as
0
0.3%
0.
2%
0.0%
1.4
% 2
.7%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Del
ftia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

107
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Gie
sber
geria
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Hyd
roge
noph
a ga
0
0.1%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
0.3
% 0
.4%
0.0
% 0
.5%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Hyl
emon
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Lim
noba
cter
0
0.2%
0.
8%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
0.5
% 0
.6%
0.9
% 0
.3%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Lim
noha
bita
ns
0
0.3%
2.
8%
0.0%
0.5
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.3%
0.0
% 0
.4%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Met
hylib
ium
0
0.0%
0.
5%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Pol
arom
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_RS6
2
0
0.4%
2.
7%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.3%
1.1
% 0
.7%
0.0
% 1
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Ram
libac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Ros
eate
les
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Rub
riviv
ax
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Sim
plic
ispi
ra
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__C
omam
onad
acea
e;g_
_Tep
idim
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__O
xalo
bact
erac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__O
xalo
bact
erac
eae;
g__
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__O
xalo
bact
erac
eae;
g__C
upria
vidu
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__O
xalo
bact
erac
eae;
g__J
anth
inob
acte
ri um
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__O
xalo
bact
erac
eae;
g__P
olyn
ucle
obac
t er
0
0.0%
0.
1%
0.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Bur
khol
deria
les;
f__O
xalo
bact
erac
eae;
g__R
alst
onia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Ellin
6067
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Gal
lione
llale
s;f_
_Gal
lione
llace
ae;g
__G
allio
nella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
IS-4
4;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

108
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
MN
D1;
f__;
g__
0
0.1%
0.
0%
0.0%
0.3
% 0
.1%
0.7
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
MW
H-U
niP1
;f__;
g__
0
0.5%
1.
2%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
0.8
% 1
.2%
1.8
% 1
.7%
0.1
% 0
.7%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Met
hylo
phila
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Met
hylo
phila
les;
f__M
ethy
loph
ilace
ae;O
ther
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Met
hylo
phila
les;
f__M
ethy
loph
ilace
ae;g
__
0
2.0%
1.
0%
0.0%
4.3
% 2
.6%
0.1
% 0
.0%
0.0
% 6
.5%
5.8
% 5
.9%
0.1
% 5
.8%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Met
hylo
phila
les;
f__M
ethy
loph
ilace
ae;g
__M
ethy
lote
nera
0
0.0%
0.
4%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Nei
sser
iale
s;f_
_Nei
sser
iace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Nei
sser
iale
s;f_
_Nei
sser
iace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Nei
sser
iale
s;f_
_Nei
sser
iace
ae;g
__A
quita
lea
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Nei
sser
iale
s;f_
_Nei
sser
iace
ae;g
__C
hrom
obac
teriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Nei
sser
iale
s;f_
_Nei
sser
iace
ae;g
__Vo
gese
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Nitr
osom
onad
ales
;f__N
itros
omon
adac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Proc
abac
teria
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Proc
abac
teria
les;
f__P
roca
bact
eria
ceae
;g__
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__
0
0.1%
0.
4%
0.0%
0.2
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.2
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__A
zoar
cus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__C
39
0
0.0%
0.
1%
0.0%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__C
andi
datu
s A
ccu
mul
ibac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__D
echl
orom
onas
0
0.0%
0.
1%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__M
ethy
love
rsat
ilis
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
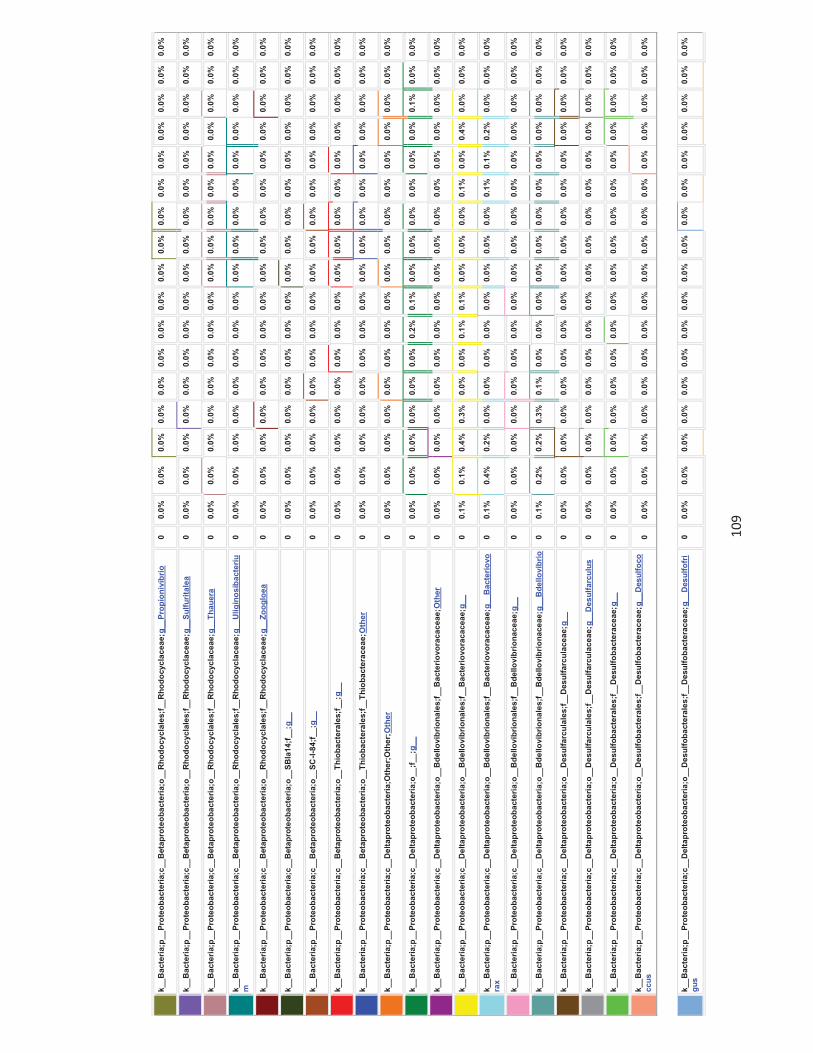
109
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__P
ropi
oniv
ibrio
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__S
ulfu
rital
ea
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__T
haue
ra
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__U
ligin
osib
acte
riu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Rho
docy
clal
es;f_
_Rho
docy
clac
eae;
g__Z
oogl
oea
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
SBla
14;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
SC-I-
84;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Thio
bact
eral
es;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__B
etap
rote
obac
teria
;o__
Thio
bact
eral
es;f_
_Thi
obac
tera
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;O
ther
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__B
dello
vibr
iona
les;
f__B
acte
riovo
raca
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__B
dello
vibr
iona
les;
f__B
acte
riovo
raca
ceae
;g__
0
0.1%
0.
1%
0.4%
0.3
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.4
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__B
dello
vibr
iona
les;
f__B
acte
riovo
raca
ceae
;g__
Bac
terio
vo
rax
0
0.1%
0.
4%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
0.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__B
dello
vibr
iona
les;
f__B
dello
vibr
iona
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__B
dello
vibr
iona
les;
f__B
dello
vibr
iona
ceae
;g__
Bde
llovi
brio
0
0.1%
0.
2%
0.2%
0.3
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
farc
ulal
es;f_
_Des
ulfa
rcul
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
farc
ulal
es;f_
_Des
ulfa
rcul
acea
e;g_
_Des
ulfa
rcul
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
foba
cter
ales
;f__D
esul
foba
cter
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
foba
cter
ales
;f__D
esul
foba
cter
acea
e;g_
_Des
ulfo
co
ccus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
foba
cter
ales
;f__D
esul
foba
cter
acea
e;g_
_Des
ulfo
fri
gus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

110
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
foba
cter
ales
;f__D
esul
fobu
lbac
eae;
g__
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
foba
cter
ales
;f__D
esul
fobu
lbac
eae;
g__D
esul
fobu
lb
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
foba
cter
ales
;f__D
esul
fobu
lbac
eae;
g__D
esul
foca
ps
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
fovi
brio
nale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
fovi
brio
nale
s;f_
_Des
ulfo
halo
biac
eae;
g__D
esul
fove
rm
icul
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
fovi
brio
nale
s;f_
_Des
ulfo
mic
robi
acea
e;g_
_Des
ulfo
m
icro
bium
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
fovi
brio
nale
s;f_
_Des
ulfo
vibr
iona
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
fovi
brio
nale
s;f_
_Des
ulfo
vibr
iona
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
fovi
brio
nale
s;f_
_Des
ulfo
vibr
iona
ceae
;g__
Des
ulfo
vi
brio
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;f_
_Des
ulfu
rom
onad
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;f_
_Geo
bact
erac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;f_
_Geo
bact
erac
eae;
g__G
eoba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;f_
_Pel
obac
tera
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;f_
_Pel
obac
tera
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__D
esul
furo
mon
adal
es;f_
_Pel
obac
tera
ceae
;g__
Pelo
bact
er
0
0.1%
0.
0%
0.5%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__FA
C87
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__G
MD
14H
09;f_
_;g_
_
0
0.0%
0.
6%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
IZ46
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.4%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__0
319-
6G20
;g__
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__C
ysto
bact
erin
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

111
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__H
alia
ngia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__M
yxoc
occa
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__M
yxoc
occa
ceae
;g__
Ana
erom
yxob
acte
r
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__N
anno
cyst
acea
e;g_
_Nan
nocy
stis
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__N
anno
cyst
acea
e;g_
_Ple
sioc
ystis
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__O
M27
;g__
0
0.0%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__M
yxoc
occa
les;
f__P
olya
ngia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__N
B1-
j;f__
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__N
B1-
j;f__
JTB
38;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__N
KB
15;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__PB
19;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sp
iroba
cilla
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.2
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sv
a085
3;f_
_;g_
_
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 1
.1%
0.0
% 0
.5%
0.0
% 0
.4%
0.0
% 0
.3%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sv
a085
3;f_
_JTB
36;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sv
a085
3;f_
_S25
_123
8;g_
_
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
0.0
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.5%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_Syn
trop
hace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_Syn
trop
hace
ae;g
__D
esul
fom
oni
le
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_Syn
trop
hace
ae;g
__Sy
ntro
phus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_Syn
trop
hoba
cter
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_Syn
trop
hoba
cter
acea
e;g_
_Syn
tr
opho
bact
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

112
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__Sy
ntro
phob
acte
rale
s;f_
_Syn
trop
horh
abda
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__D
elta
prot
eoba
cter
ia;o
__[E
ntot
heon
ella
les]
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__C
ampy
loba
cter
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__C
ampy
loba
cter
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__C
ampy
loba
cter
acea
e;g_
_Arc
ob
acte
r 0
0.9%
0.
0%
5.7%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
6.6
% 2
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__C
ampy
loba
cter
acea
e;g_
_Sul
fur
ospi
rillu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__H
elic
obac
tera
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__H
elic
obac
tera
ceae
;g__
Hel
icob
a ct
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__H
elic
obac
tera
ceae
;g__
Sulfu
ric
urvu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ep
silo
npro
teob
acte
ria;o
__C
ampy
loba
cter
ales
;f__H
elic
obac
tera
ceae
;g__
Sulfu
rim
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;O
ther
;Oth
er;O
ther
0
0.1%
0.
0%
0.0%
0.2
% 0
.0%
0.2
% 0
.0%
0.0
% 1
.0%
0.3
% 0
.1%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__34
P16;
f__;
g__
0
0.3%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.6%
1.3
% 0
.7%
0.5
% 1
.0%
0.0
% 0
.1%
0.4
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
erom
onad
ales
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
erom
onad
ales
;f__A
erom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
erom
onad
ales
;f__A
erom
onad
acea
e;g_
_
0
0.1%
0.
5%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.3%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
erom
onad
ales
;f__A
erom
onad
acea
e;g_
_Oce
anim
ona
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
erom
onad
ales
;f__A
erom
onad
acea
e;g_
_Tol
umon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
erom
onad
ales
;f__S
ucci
nivi
brio
nace
ae;g
__Su
ccin
ivi
brio
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

113
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;O
ther
;Oth
er
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
1.0
% 0
.1%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_;g_
_
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.7%
0.8
% 1
.0%
0.0
% 0
.8%
0.0
% 0
.0%
0.3
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_125
ds10
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_211
ds20
;g__
0
0.2%
0.
1%
0.0%
0.4
% 0
.3%
0.0
% 0
.3%
0.3
% 0
.1%
0.1
% 0
.4%
0.5
% 0
.8%
0.0
% 0
.0%
0.1
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_
0
0.2%
1.
6%
0.5%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.1
% 0
.2%
0.3
% 0
.2%
0.1
% 0
.2%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Aga
rivor
a ns
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Alte
rom
on
as
0
0.1%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.4%
0.0
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_BD
2-13
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Can
dida
tu
s En
dobu
gula
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Cel
lvib
rio
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Gla
ciec
ola
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.2%
0.0
% 0
.5%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_HB
2-32
-21 0
0.
1%
0.1%
0.
0% 0
.0%
0.0
% 0
.0%
0.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.7
%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_HTC
C22
07
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Mar
inim
ic
robi
um
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Mar
inob
ac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_Mic
robu
lb
ifer
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Alte
rom
onad
acea
e;g_
_ZD
0117
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Col
wel
liace
ae;g
__Th
alas
som
ona
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Fer
rimon
adac
eae;
g__F
errim
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_HTC
C21
88;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

114
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_HTC
C21
88;g
__
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_HTC
C21
88;g
__H
TCC
0
2.5%
0.
4%
0.0%
0.0
% 0
.1%
0.0
% 1
5.4%
8.7
% 0
.5%
0.2
% 0
.3%
0.7
% 0
.3%
0.0
% 0
.0%
11.
9% 1
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Idi
omar
inac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Idi
omar
inac
eae;
g__I
diom
arin
a
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 1
.0%
0.6
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 1
.4%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Idi
omar
inac
eae;
g__P
seud
idio
mar
in
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_J11
5;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.6
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_J11
5;g_
_
0
0.0%
0.
2%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_J11
5;g_
_Spo
ngiib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_OM
60;g
__
0
0.1%
0.
2%
0.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.2
% 0
.3%
0.0
% 0
.2%
0.1
% 0
.3%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Psy
chro
mon
adac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_Psy
chro
mon
adac
eae;
g__P
sych
ro
mon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_She
wan
ella
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_She
wan
ella
ceae
;g__
Shew
anel
la
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.7
% 0
.2%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_[C
hrom
atia
ceae
];O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_[C
hrom
atia
ceae
];g_
_
0
0.2%
0.
1%
0.0%
0.2
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.5
% 0
.4%
0.4
% 0
.8%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__A
ltero
mon
adal
es;f_
_[C
hrom
atia
ceae
];g_
_Rhe
inhe
imer
a
0
0.3%
0.
8%
0.0%
2.5
% 0
.9%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.5%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__C
ardi
obac
teria
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__C
hrom
atia
les;
Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__C
hrom
atia
les;
f__;
g__
0
0.6%
0.
8%
0.4%
0.7
% 0
.1%
0.3
% 0
.0%
0.1
% 2
.6%
1.2
% 0
.8%
0.1
% 0
.7%
0.8
% 0
.2%
0.1
% 0
.3%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__C
hrom
atia
les;
f__C
hrom
atia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__C
hrom
atia
les;
f__E
ctot
hior
hodo
spira
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

115
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__C
hrom
atia
les;
f__E
ctot
hior
hodo
spira
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
0
0.6%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.3%
3.0
% 2
.0%
0.1
% 3
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Citr
obac
t er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Edw
ards
i el
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Erw
inia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Esch
eric
hi
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Kle
bsie
ll a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Salm
onel
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__En
tero
bact
eria
les;
f__E
nter
obac
teria
ceae
;g__
Serr
atia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__H
TCC
2188
;f__;
g__
0
0.5%
0.
0%
0.8%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 1
.5%
0.5
% 1
.1%
0.1
% 1
.7%
0.4
% 0
.8%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__H
TCC
2188
;f__H
TCC
2089
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__;
g__
0
0.1%
0.
0%
0.0%
0.2
% 0
.1%
0.3
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__C
oxie
llace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__C
oxie
llace
ae;g
__
0
0.4%
0.
3%
0.0%
0.3
% 0
.3%
1.0
% 0
.4%
3.1
% 0
.1%
0.1
% 0
.2%
0.2
% 0
.2%
0.0
% 0
.3%
0.1
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__C
oxie
llace
ae;g
__A
quic
ella
0
0.1%
0.
0%
0.0%
0.6
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__C
oxie
llace
ae;g
__C
oxie
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__C
oxie
llace
ae;g
__R
icke
ttsie
lla
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__E
ndoe
ctei
nasc
idia
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__F
ranc
isel
lace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__F
ranc
isel
lace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

116
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__F
ranc
isel
lace
ae;g
__Fr
anci
sella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__L
egio
nella
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__L
egio
nella
ceae
;g__
0
0.1%
0.
1%
0.1%
0.2
% 0
.6%
0.7
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.2
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__L
egio
nella
ceae
;g__
Legi
onel
la
0
0.0%
0.
0%
0.0%
0.1
% 0
.1%
0.5
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Le
gion
ella
les;
f__L
egio
nella
ceae
;g__
Tatlo
ckia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__M
ethy
loco
ccal
es;f_
_;g_
_
0
0.3%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 5
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__M
ethy
loco
ccal
es;f_
_Cre
notr
icha
ceae
;g__
Cre
noth
rix
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__M
ethy
loco
ccal
es;f_
_Met
hylo
cocc
acea
e;g_
_Met
hylo
cal
dum
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__M
ethy
loco
ccal
es;f_
_Met
hylo
cocc
acea
e;g_
_Met
hylo
m
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__;
g__
0
1.1%
0.
0%
2.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 4
.8%
1.5
% 0
.6%
2.8
% 0
.8%
0.9
% 4
.7%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__A
lcan
ivor
acac
eae;
g__A
lcan
ivor
a x
0
1.6%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 5
.6%
2.8
% 0
.1%
0.9
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
2.1
% 1
3.6%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
ahel
lace
ae;g
__H
ahel
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
alom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
alom
onad
acea
e;g_
_
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 1
.1%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.3
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
alom
onad
acea
e;g_
_Can
dida
tus
Por
tiera
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
alom
onad
acea
e;g_
_Chr
omoh
al
obac
ter
0
0.2%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 2
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
alom
onad
acea
e;g_
_Hae
rere
hal
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__H
alom
onad
acea
e;g_
_Hal
omon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;g__
0
0.8%
0.
0%
2.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 3
.4%
0.8
% 0
.7%
0.1
% 1
.1%
2.9
% 1
.1%
0.0
% 0
.0%

117
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;g__
Mar
inob
a ct
eriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;g__
Mar
inom
on
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.5
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;g__
Nep
tuni
ib
acte
r 0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;g__
Nep
tuno
m
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
cean
ospi
rilla
ceae
;g__
Ole
ibac
te
r 0
0.4%
0.
0%
1.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.4
% 0
.1%
0.5
% 0
.1%
1.2
% 2
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
leip
hila
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__O
leip
hila
ceae
;g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
1.2
% 0
.2%
0.4
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__S
UP0
5;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__S
acch
aros
piril
lace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__S
acch
aros
piril
lace
ae;g
__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.4
% 0
.1%
0.1
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__O
cean
ospi
rilla
les;
f__S
acch
aros
piril
lace
ae;g
__R
eine
ke
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__PY
R10
d3;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Pa
steu
rella
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;g
__A
cine
toba
cte
r 0
0.0%
0.
1%
0.1%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;g
__A
lkan
indi
ges
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;g
__En
hydr
obac
t er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;g
__Pe
rluci
diba
c a
0
0.3%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%
1.1
% 1
.8%
0.0
% 1
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Mor
axel
lace
ae;g
__Ps
ychr
obac
t er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

118
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Pse
udom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Pse
udom
onad
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Ps
eudo
mon
adal
es;f_
_Pse
udom
onad
acea
e;g_
_Pse
ud
omon
as
0
0.4%
0.
1%
0.0%
1.5
% 1
.4%
0.2
% 0
.0%
0.0
% 0
.1%
1.4
% 0
.8%
0.2
% 0
.9%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Sa
linis
phae
rale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Sa
linis
phae
rale
s;f_
_Sal
inis
phae
race
ae;g
__Sa
linis
pha
era
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
ioha
lorh
abda
les;
f__;
g__
0
0.1%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.0%
0.3
% 0
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__P
isci
ricke
ttsia
ceae
;g__
0
0.1%
0.
0%
0.0%
0.1
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.2%
0.0
% 0
.0%
0.3
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__P
isci
ricke
ttsia
ceae
;g__
Met
hylo
phag
a
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__P
isci
ricke
ttsia
ceae
;g__
Pisc
irick
etts
ia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__P
isci
ricke
ttsia
ceae
;g__
Thio
mic
rosp
ir a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__T
hiot
richa
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Th
iotr
icha
les;
f__T
hiot
richa
ceae
;g__
Thio
thrix
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Pse
udoa
ltero
mon
adac
eae;
Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Pse
udoa
ltero
mon
adac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.4
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Pse
udoa
ltero
mon
adac
eae;
g__P
seud
oa
ltero
mon
as
0
0.6%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 1
.2%
6.4
% 0
.7%
0.2
% 0
.6%
0.0
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Vib
riona
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Vib
riona
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Vib
riona
ceae
;g__
Phot
obac
teriu
m
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

119
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Vi
brio
nale
s;f_
_Vib
riona
ceae
;g__
Vibr
io
0
1.6%
0.
0%
3.9%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 2
.7%
0.9
% 0
.1%
0.0
% 0
.1%
17.
5%
0.5%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Sin
obac
tera
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Sin
obac
tera
ceae
;g__
0
0.7%
1.
4%
0.0%
1.5
% 0
.3%
0.1
% 2
.7%
1.9
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.3
% 2
.2%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Sin
obac
tera
ceae
;g__
Hyd
roca
rb
onip
haga
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Sin
obac
tera
ceae
;g__
Nev
skia
0
0.1%
0.
9%
0.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Sin
obac
tera
ceae
;g__
Ster
oido
ba
cter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_
0
0.1%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.3%
0.4
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Dok
don
ella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Lut
eiba
ct
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Lut
eim
o na
s
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Lys
oba
cter
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Pse
udo
xant
hom
onas
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Rho
dan
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Ste
notr
op
hom
onas
0
0.1%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
1.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_The
rmo
mon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__Xa
ntho
mon
adal
es;f_
_Xan
thom
onad
acea
e;g_
_Xan
tho
mon
as
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__[M
arin
icel
lale
s];f_
_[M
arin
icel
lace
ae];
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__G
amm
apro
teob
acte
ria;o
__[M
arin
icel
lale
s];f_
_[M
arin
icel
lace
ae];
g__M
arin
icel
la
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__TA
18;o
__C
V90;
f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.2%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.4%

120
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__TA
18;o
__PH
OS-
HD
29;f_
_;g_
_
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.5
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
k_
_Bac
teria
;p__
Prot
eoba
cter
ia;c
__Ze
tapr
oteo
bact
eria
;o__
Mar
ipro
fund
ales
;f__M
arip
rofu
ndac
eae;
g__M
arip
rofu
ndus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
SAR
406;
c__A
B16
;o__
;f__;
g__
0
0.1%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.4
% 0
.6%
0.0
% 0
.0%
k_
_Bac
teria
;p__
SAR
406;
c__A
B16
;o__
Arc
tic96
B-7
;f__A
7140
17;g
__ZA
3312
c
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
SAR
406;
c__A
B16
;o__
SSW
63A
u;f_
_SH
AS4
60;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
SBR
1093
;c__
VHS-
B5-
50;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
SR1;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__G
N05
;o__
LF03
0;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__G
N05
;o__
SBYZ
_608
0;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__M
VP-1
5;o_
_PL-
11B
10;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
phae
roch
aeta
les;
f__S
phae
roch
aeta
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
phae
roch
aeta
les;
f__S
phae
roch
aeta
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
phae
roch
aeta
les;
f__S
phae
roch
aeta
ceae
;g__
Spha
eroc
haet
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
piro
chae
tale
s;f_
_Spi
roch
aeta
ceae
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
piro
chae
tale
s;f_
_Spi
roch
aeta
ceae
;g__
0
0.0%
0.
0%
0.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
piro
chae
tale
s;f_
_Spi
roch
aeta
ceae
;g__
Spiro
chae
ta
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__S
piro
chae
tes;
o__S
piro
chae
tale
s;f_
_Spi
roch
aeta
ceae
;g__
Trep
onem
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__[
Lept
ospi
rae]
;o__
[Lep
tosp
irale
s];f_
_Lep
tosp
irace
ae;g
__Le
pton
ema
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Spiro
chae
tes;
c__[
Lept
ospi
rae]
;o__
[Lep
tosp
irale
s];f_
_Lep
tosp
irace
ae;g
__Tu
rner
iella
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Syne
rgis
tete
s;c_
_Syn
ergi
stia
;o__
Syne
rgis
tale
s;f_
_Det
hios
ulfo
vibr
iona
ceae
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM6;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

121
k_
_Bac
teria
;p__
TM6;
c__S
BR
H58
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM6;
c__S
JA-4
;Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM6;
c__S
JA-4
;o__
;f__;
g__
0
0.2%
0.
0%
0.0%
0.2
% 0
.0%
0.2
% 0
.2%
0.0
% 0
.1%
0.1
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
1.2
% 0
.2%
k_
_Bac
teria
;p__
TM6;
c__S
JA-4
;o__
S119
8;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM6;
c__S
JA-4
;o__
YJF2
-48;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
Oth
er;O
ther
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__M
JK10
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__S
C3;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
1;o_
_;f_
_;g_
_
0
0.7%
0.
0%
0.0%
2.8
% 2
.3%
6.1
% 0
.0%
0.0
% 0
.1%
0.1
% 0
.1%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;O
ther
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_;f_
_;g_
_
0
1.2%
0.
0%
0.1%
0.1
% 1
8.5%
0.
0% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_Blg
i18;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_CW
040;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_CW
040;
f__F
16;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_EW
055;
f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.2%
1.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_I02
5;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
TM7;
c__T
M7-
3;o_
_I02
5;f_
_Rs-
045;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Tene
ricut
es;c
__M
ollic
utes
;o__
;f__;
g__
0
0.2%
0.
0%
2.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
1.7
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Tene
ricut
es;c
__M
ollic
utes
;o__
Ach
olep
lasm
atal
es;f_
_Ach
olep
lasm
atac
eae;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Tene
ricut
es;c
__M
ollic
utes
;o__
Ach
olep
lasm
atal
es;f_
_Ach
olep
lasm
atac
eae;
g__A
chol
epla
sma
0
0.2%
0.
0%
0.6%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.4
% 0
.1%
0.2
% 0
.1%
0.5
% 1
.2%
0.0
% 0
.0%

122
k_
_Bac
teria
;p__
Tene
ricut
es;c
__M
ollic
utes
;o__
Ento
mop
lasm
atal
es;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Tene
ricut
es;c
__M
ollic
utes
;o__
RF3
9;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Tene
ricut
es;c
__R
F3;o
__M
L615
J-28
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Ther
mot
ogae
;c__
Ther
mot
ogae
;o__
Ther
mot
ogal
es;f_
_The
rmot
ogac
eae;
g__K
osm
otog
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
0.1
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__O
pitu
tale
s;f_
_Opi
tuta
ceae
;g__
0
0.1%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 1
.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__O
pitu
tale
s;f_
_Opi
tuta
ceae
;g__
Opi
tutu
s
0
1.1%
2.
4%
0.0%
5.4
% 1
.5%
0.3
% 0
.0%
0.1
% 0
.6%
1.5
% 2
.4%
0.2
% 2
.8%
0.0
% 0
.0%
0.4
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__P
unic
eico
ccal
es;f_
_Pun
icei
cocc
acea
e;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__P
unic
eico
ccal
es;f_
_Pun
icei
cocc
acea
e;g_
_Cor
alio
mar
garit
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__P
unic
eico
ccal
es;f_
_Pun
icei
cocc
acea
e;g_
_MB
11C
04
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__[
Cer
asic
occa
les]
;f__[
Cer
asic
occa
ceae
];g_
_
0
0.1%
0.
0%
0.0%
1.1
% 0
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__O
pitu
tae;
o__[
Pela
gico
ccal
es];f
__[P
elag
icoc
cace
ae];
g__P
elag
icoc
cus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
co-5
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
co-5
;o__
LD1-
PB3;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
co-5
;o__
MSB
L3;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
co-5
;o__
R76
-B12
8;f_
_;g_
_
0
0.1%
0.
0%
0.3%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
1.2
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
co-5
;o__
SS1-
B-0
3-39
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
co-5
;o__
WC
HB
1-41
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_
0
0.3%
0.
0%
0.0%
0.0
% 0
.2%
3.5
% 0
.3%
0.2
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.3%

123
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_Akk
erm
an
sia
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_Hal
ofer
ul
a
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_Lut
eoli
bact
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_MSB
L3
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_Per
sici
r ha
bdus
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.6%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_Pro
sthe
co
bact
er
0
0.1%
0.
1%
0.0%
0.0
% 0
.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
1.4
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__V
erru
com
icro
biae
;o__
Verr
ucom
icro
bial
es;f_
_Ver
ruco
mic
robi
acea
e;g_
_Ver
ruc
omic
robi
um
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Met
hyla
cidi
phila
e];o
__M
ethy
laci
diph
ilale
s;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Met
hyla
cidi
phila
e];o
__M
ethy
laci
diph
ilale
s;f_
_LD
19;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Met
hyla
cidi
phila
e];o
__S
-BQ
2-57
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];Oth
er;O
ther
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
;f__;
g__
0
0.1%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.3%
0.0
% 0
.3%
0.0
% 0
.2%
0.0
% 0
.3%
0.0
% 0
.5%
0.5
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;Oth
er;O
ther
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__;
g__
0
0.1%
0.
0%
0.0%
0.3
% 0
.1%
0.0
% 0
.2%
0.3
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.1%
0.0
% 0
.2%
0.1
% 0
.1%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__E
llin5
15;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__E
llin5
17;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__R
4-41
B;g
__
0
0.5%
2.
1%
0.0%
4.0
% 1
.2%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__[
Pedo
spha
erac
eae]
;Oth
er
0
1.0%
0.
0%
0.0%
0.0
% 0
.8%
0.0
% 9
.9%
2.1
% 0
.0%
0.0
% 0
.0%
2.1
% 0
.0%
0.0
% 0
.0%
1.2
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__[
Pedo
spha
erac
eae]
;g__
Pedo
spha
era
0
0.0%
0.
0%
0.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Pedo
spha
erae
];o__
[Ped
osph
aera
les]
;f__a
uto6
7_4W
;g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Spar
toba
cter
ia];o
__[C
htho
niob
acte
rale
s];f_
_[C
htho
niob
acte
race
ae];
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%

124
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Spar
toba
cter
ia];o
__[C
htho
niob
acte
rale
s];f_
_[C
htho
niob
acte
race
ae];
g__C
andi
da
tus
Xiph
inem
atob
acte
r 0
0.2%
0.
1%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.8%
1.1
% 0
.1%
0.0
% 0
.7%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Spar
toba
cter
ia];o
__[C
htho
niob
acte
rale
s];f_
_[C
htho
niob
acte
race
ae];
g__C
htho
ni
obac
ter
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Spar
toba
cter
ia];o
__[C
htho
niob
acte
rale
s];f_
_[C
htho
niob
acte
race
ae];
g__h
eter
o C
45_4
W
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
Verr
ucom
icro
bia;
c__[
Spar
toba
cter
ia];o
__[C
htho
niob
acte
rale
s];f_
_zEL
20;g
__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WPS
-2;c
__;o
__;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS2
;c__
SHA
-109
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS3
;c__
PRR
-12;
o__G
N03
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS3
;c__
PRR
-12;
o__G
N03
;f__K
SB4;
g__
0
0.0%
0.
0%
0.1%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.4%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS3
;c__
PRR
-12;
o__S
edim
ent-1
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS3
;c__
PRR
-12;
o__S
edim
ent-1
;f__P
RR
-10;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS5
;c__
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS6
;c__
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS6
;c__
B14
2;o_
_;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.0%
k_
_Bac
teria
;p__
WS6
;c__
SC72
;o__
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS6
;c__
SC72
;o__
A-2
AF;
f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS6
;c__
SC72
;o__
MA
T-C
R-H
2-G
03;f_
_;g_
_
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WS6
;c__
SC72
;o__
WC
HB
1-15
;f__;
g__
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
WW
E1;c
__[C
loac
amon
ae];o
__[C
loac
amon
ales
];f__
[Clo
acam
onac
eae]
;Oth
er
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
ZB3;
c__B
S119
;o__
;f__;
g__
0
0.1%
0.
0%
0.1%
0.3
% 0
.6%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.1
% 0
.2%
0.0
% 0
.0%
k_
_Bac
teria
;p__
[The
rmi];
c__D
eino
cocc
i;o__
Dei
noco
ccal
es;f_
_Dei
noco
ccac
eae;
g__D
eino
cocc
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
k_
_Bac
teria
;p__
[The
rmi];
c__D
eino
cocc
i;o__
Ther
mal
es;f_
_The
rmac
eae;
g__M
eiot
herm
us
0
0.0%
0.
0%
0.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%
0.0
% 0
.0%