peptide arrays for kinome analysis: new opportunities and remaining challenges
TRANSCRIPT

REVIEW
Peptide arrays for kinome analysis: New opportunities
and remaining challenges
Ryan Arsenault1,2, Philip Griebel2,3 and Scott Napper1,2
1 Department of Biochemistry, University of Saskatchewan, Saskatoon, Saskatchewan, Canada2 Vaccine and Infectious Disease Organization, University of Saskatchewan, Saskatoon, Saskatchewan,
Canada3 School of Public Health, University of Saskatchewan, Saskatoon, Saskatchewan, Canada
Received: May 31, 2011
Revised: September 28, 2011
Accepted: October 4, 2011
Phosphorylation is the predominant mechanism of post-translational modification for
regulation of protein function. With central roles in virtually every cellular process, and
strong linkages with many diseases, there is a considerable interest in defining, and ulti-
mately controlling, kinase activities. Investigations of human cellular phosphorylation events,
which includes over 500 different kinases and tens of thousands of phosphorylation targets,
represent a daunting challenge for proteomic researchers and cell biologists alike. As such,
there is a priority to develop tools that enable the evaluation of cellular phosphorylation events
in a high-throughput, and biologically relevant, fashion. Towards this objective, two distinct,
but functionally related, experimental approaches have emerged; phosphoproteome investi-
gations, which focus on the sub-population of proteins which undergo phosphorylation and
kinome analysis, which considers the activities of the kinase enzymes mediating these
phosphorylation events. Within kinome analysis, peptide arrays have demonstrated consid-
erable potential as a cost-effective, high-throughput approach for defining phosphorylation-
mediated signal transduction activity. In particular, a number of recent advances in the
application of peptide arrays for kinome analysis have enabled researchers to tackle
increasingly complex biological problems in a wider range of species. In this review, recent
advances in kinomic analysis utilizing peptides arrays including several of the biological
questions studied by our group, as well as outstanding challenges still facing this technology,
are discussed.
Keywords:
Kinase / Kinome / Peptide array / Phosphoproteome / Phosphorylation /
Protein arrays
1 Background
In the late 1950s, Krebs and Fischer were the first to
describe the role of reversible protein phosphorylation for
the regulation of enzymatic activity [1, 2]. For this pivotal
contribution to science they were awarded the Nobel Prize.
Protein kinases, which catalyze the transfer of the g phos-
phate group from ATP to specific serine, threonine or
tyrosine hydroxyl groups in a target protein substrate, are
now recognized as one of the largest and most important
enzyme classes. Consisting of over 500 members, human
protein kinases are responsible for modifying an estimated
one-third of the human proteome [3, 4] with many members
of the proteome undergoing complex patterns of kinase
modification at multiple sites to generate distinct isoforms
with unique functional characteristics. The presence and
dynamic nature of these phosphorylation isoforms adds a
daunting layer of complexity to characterizing and under-
standing the proteome. While much is unknown of how
Colour Online: See the article online to view Figs. 1, 3–5 in colour.
Abbreviations: Bregs, regulatory B cells; LPS, lipopolysaccharide;
ODN, oligodeoxynucleotide; PP, Peyer’s Patch; TLR, Toll-like
receptor
Correspondence: Dr. Scott Napper, Department of Biochemistry,
University of Saskatchewan, 120 Veterinary Road, University of
Saskatchewan, Saskatoon, Saskatchewan, S7N 5E3 Canada
E-mail: [email protected]
Fax: 11-306-966-7478
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Proteomics 2011, 11, 4595–4609 4595DOI 10.1002/pmic.201100296

these modifications, even within a static infrastructure of
proteins, achieve complex biological responses, there is a
growing appreciation of the importance of kinases in
controlling cellular responses and of the potential for char-
acterizations of global kinase activity (the kinome) to offer
critical insight into biology.
There is a considerable debate as to the most appropriate
level at which to define cell responses. Transcriptional
analysis, based largely on availability and maturity of the
approach, remains the most widely applied technique for
global analysis of cellular responses. However, due to a
multitude of post-transcriptional regulatory events, there are
concerns that descriptions of patterns of gene expression, no
matter how comprehensive, do not accurately describe or
predict cellular phenotypes. Specifically, a major criticism of
genetic approaches is their inability to consider post-
transcription regulatory events such as gene silencing,
mRNA stability, unique translational efficiencies, protein
turnover, sequestration of enzymes away from substrates,
and activation and deactivation of proteins by any number of
post-translational modifications. Intuitively, characteriza-
tions of host responses that occur closer to the functional
phenotype should have greater potential to circumvent these
complicating factors and offer a clearer picture of cellular
response. From this perspective, protein kinases are at the
core of signal transduction with central roles in regulation of
virtually every aspect of cellular behavior. Through their
ability to modulate protein conformation and functional
characteristics, kinases control diverse processes such as
metabolism, transcription, cell cycle progression, cytoskeletal
rearrangement and cell movement, apoptosis, and differ-
entiation. As such, characterizations of host cellular
responses at the level of phosphorylation-mediated signal
transduction have the potential to offer important, and
predictive, insight in cellular mechanisms of phenotypes.
Investigations of cellular response at the level of protein
phosphorylation are also important and appropriate as the
disruption of kinase-mediated signaling cascades are asso-
ciated with a spectrum of diseases including cancer,
inflammation, neurological disorders and diabetes [5].
Indeed within the human genome, over 250 protein kinase
genes map to disease loci [6]. The involvement of kinases in
disease typically results from improper levels of expression/
localization/activity or mutations in the protein sequences
that alter these activities.
The role of kinases in many diseases, as well as their
regulatory role in many central pathways, makes them
logical targets for drug therapy [7]. Fortuitously, the
conserved catalytic cleft of the kinases is highly attractive for
drug design making the kinases highly ‘‘druggable’’ [8].
Kinase inhibitors have also been proposed as a more precise
mechanism for therapeutic intervention than other strate-
gies such as the targeted down-regulation of particular
genes. Not surprisingly, the central role of kinases in many
diseases, cancer in particular, and the potential to treat
complex phenotypes by targeting specific biomolecules,
have prompted drug companies to invest considerable effort
into the development of kinase inhibitors. There are esti-
mates that approximately half of the current Research and
Development budget of the pharmaceutical industry is
focused on kinases and their inhibitors. Kinases are the
most frequently targeted gene class in cancer therapeutics,
and are second only to G protein-coupled receptors across all
therapeutic areas [7, 8]. In addition to the immediate value
of these emerging molecules as therapeutics, these inhibi-
tors also represent a valuable resource with the potential for
utilization in research for hypothesis validation. Given the
magnitude of effort devoted towards their development it is
certain that additional kinase inhibitors, of greater range
and improved specificity, will be developed.
There are a number of licensed, and soon-to-be licensed,
kinase inhibitors that emphasize the potential of these
targets. Gleevac (imatinib), a potent inhibitor of the consti-
tutively active breakpoint cluster region-Abelson murine
leukemia viral oncogene homolog 1 (BCR-ABL) fusion
protein, is approved for the treatment of leukemia and
gastrointestinal stromal tumors [9, 10]. Other protein kinase
inhibitors, such as the epidermal growth factor receptor
(EGFR) inhibitors (Tarceva, Genetech) and getinib (Iressa
AstraZeneca, London UK), have either received FDA
approval or are in the late stage clinical development to treat
different cancers [11, 12]. The potential therapeutic value of
kinase inhibitors is not limited to the treatment of cancers.
For example, ruboxistaurin to treat diabetic retinopathy,
safingol, a protein kinase C inhibitor, for treatment of
atopical dermatitis and fasudil, which has received approval
in Japan, for treatment of cerebral ischemia. Therapeutic
modulation of kinase activity can also have anti-inflamma-
tory and immunosuppressive effects. For example, two
critical immunosuppressive drugs, cyclosporine A and
rapamycin, function through modulation of the phosphor-
ylation status of the cell; cyclosporine A through inhibition
of a phosphatase [13] and rapamycin through inhibition of a
kinase [14]. Other anti-inflammatory drugs that suppress
tumor necrosis factor (TNF)-a and interleukin (IL)-1bexpression also function through kinase inhibition [15].
1.1 Phosphoproteome and kinome analysis
A number of experimental approaches are available for the
analysis of phosphorylation based cellular signaling, these
can be divided into two groups, phosphoproteome and
kinome analysis [16]. The difference is dependent on
whether the consideration is on the protein kinases that
phosphorylate proteins, the kinome, or the targets of these
enzymes, the phosphoproteome. While these types of
analysis are strongly linked, representing the same biologi-
cal phenomena, and are at times considered interchange-
able, the experimental approaches are distinct. We suggest a
strict delineation where phosphoproteome analysis consid-
ers only the proteins containing phosphoryl groups and the
4596 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

kinome refers to the enzymatic activities of the kinase
compliment of a cell, regardless of substrate target. This
distinction between the two should not be exaggerated,
however, as the phosphoproteome represents the net action
of the kinome, as well as that of active phosphatases.
Phosphoproteome analysis seeks to define the sub-
population of the proteome with respect to the identities and
points of phosphorylation. Phosphoproteome analysis
shares the same challenges that are associated with standard
proteomic analysis but is complicated experimentally by: the
relative scarcity of phosphoproteome members, the dynamic
nature of protein phosphorylation and the proteome size
overwhelming, or suppressing, the ability to detect the
phosphoproteome [17–19]. The small concentration of
phosphoproteins relative to the entire proteome is a signif-
icant challenge in phosphoproteome analysis. While a large
fraction of the proteome undergoes phosphorylation, many
of the proteins involved in signal transduction are expressed
at very low levels. Thus, the proteins of most interest when
studying cellular signaling are those that are the most
difficult to isolate. This is exacerbated by the fact that these
proteins, in addition to being found in low abundance, are
often phosphorylated at sub-stoichiometric levels. This
means that a small fraction of a given signaling protein is
phosphorylated at any one time; only 1–2% of the total
individual protein compliment of a cell is found in the
phosphorylated form [16–19]. When considering all these
experimental limitations, a promising alternative for char-
acterization of cellular phosphorylation is to focus instead
on the kinome. Investigations of enzymatic activities offer
greater potential for targeted, and perhaps mostly impor-
tantly, sensitive, analysis. Specifically, the well-defined and
highly conserved chemistry of enzymatic phosphorylation
permits rapid characterization of kinase activity, provided an
appropriate substrate is available.
1.2 Kinome Analysis through peptide arrays
A central obstacle for global kinome analysis is the nature of
the substrates to be employed. While proteins are the
physiological substrates for the kinases they are problematic
to mass produce and relatively unstable on array format. An
alternative is to use peptides that represent sequences
surrounding a site of phosphorylation. Many protein kina-
ses recognize phosphoacceptor sites determined by residues
surrounding the phosphorylated amino acid, as opposed to
higher order secondary or tertiary structures. Specifically,
the target specificity of many kinases is a function of the
residues in the 14 and �4 flanking positions of the phos-
phoacceptor site [20]. Synthetic peptides modeled on the site
of phosphorylation have been shown to be appropriate
substrates with Vmax and Km values approaching that of the
intact protein [21]. Relative to the complete protein, peptides
are easily synthesized, inexpensive, highly stable and
amenable to array technology [22]. Construction of arrays
representing hundreds to thousands of immobilized
peptides allows the profiling of cellular signaling activities
by determining the activities of hundreds of kinases in a
single experiment.
The basic premise of kinome analysis through peptide
arrays is that peptides representing the phosphorylation
target sites of proteins are synthesized and spotted onto an
array surface [23]. Detailed review of the commonly
employed methodologies for peptide synthesis and array
spotting are available elsewhere [16]. Following production
of the array a sample containing active kinase, or kinases,
such as a cellular lysate, is applied for the array and these
enzymes phosphorylate their respective target sequences
using ATP as the phosphate source. This phosphorylation
event is visualized using one of a number of methods
including phosphorylation-specific antibodies, radioactivity
or phospho-specific stains (Fig. 1). Quantification of the
extent of phosphorylation of a given peptide by lysates
representing different experimental conditions (control
versus for treatment) enable evaluation of relative kinase
activity. It is also possible to infer the extent of cellular
phosphorylation of the protein that is represented by the
peptide.
A large number of peptide arrays for kinome analysis are
commercially available. These arrays range in size from
dozens to thousands of peptides, representing defined
phosphorylation sites. A partial listing of the peptide arrays
that are currently available for kinome analysis is presented
in (Table 1).
2 Success stories
While still an emerging technology, the literature contains
numerous examples of the successful application of peptide
arrays for kinome analysis. The following studies carried out
by our group represent characterizations of cellular
responses to stimuli of a range of biological complexity,
from characterizing responses to defined, singular ligands
to describe changes in phosphorylation-mediated signal
transduction during disease states. These studies were
carried out in bovine and in most cases are related to the
study of immunological signaling. The use of bovine as a
model is important for a number of reasons, the commercial
importance of bovine cannot be overstated and any under-
standing that aids in the health of the bovine population is
extremely important to this industry. The ability to do
infection studies and to collect adequate amounts of mate-
rial, for examples cells and tissue, is key for peptide array
studies where large numbers of cells may be needed.
Immunological studies involving specific immune stimu-
lating ligands are well suited for peptide array work as they
tend to work through specific receptors and activate defined
signaling pathways. These pathways can then be picked out
of a complex signaling network more easily than studies that
have broad cellular effects.
Proteomics 2011, 11, 4595–4609 4597
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

2.1 Validating and expanding known signaling
pathways to defined ligands
Much of the early confidence in the ability of peptide arrays
to offer accurate insight into cell signaling events stemmed
from investigations with isolated cells in response to a
defined ligand. In particular, validation of the kinome array
technology was first performed by analyzing cellular
responses induced by a specific ligand for which signal-
ing events had been characterized through independent
Figure 1. Methods of detecting phosphorylated peptides: (A) Kinases specific for their target peptide sequence phoshorylate the appro-
priate residue on the array. The g phosphate group of ATP which is the phosphate donor for the kinase is radioactive and its emissions can
be visualized and quantified. (B) Direct detection of phospho-peptides by using a labelled antibody specific for the phosphorylated form of
the peptide. (C) Indirect detection of the phospho-peptide by first using a phospho-specific antibody which is then visualized by with a
labeled anti-immunoglobulin antibody. Indirect labeling can enhance sensitivity by amplifying the signal from the phosphorylation event.
(D) A labeled phosphoamino acid chelator is added after the kinase reaction. Surface plasma resonance or MALDI-TOF MS can then be
used for detection. (E) Fluorescent phosphospecific stain which only binds phosphorylated peptide can be used to directly detect the
phosphorylated residues present on the array following completion of the kinase reaction. Adapted from Jalal et al. (2007) [16].
Table 1. Commercially available peptide arrays
Name Technology Company Number ofsubstrates
Visualizationmethod
PepScan Spotted peptide array PepScanTherapeutics
10 000–100 000 Variable
PepStar Custom spotted peptide array JPT 100–1000 VariablePamChip Immobilized peptide on porous surface Pamgene 50–150 FluorescencePeptide array Unbound 96-well array ChemPep 96 RadiolabelCelluSpots Custom spotted peptide array cellulose
membraneIntavis AG 1–384 Variable
Reverse-phase proteinmicroarray
Sample blotting peptide array TheranosticsHealth
100s Fluorescence
4598 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

methodologies. In one such example our group conducted
an investigation of signaling in bovine monocytes following
stimulation with ligands for different members of the same
receptor family. The Toll-like receptors (TLRs) are a family
of pathogen recognition receptors that alert the host to the
presence of microbial challenge [24]. Various microbial
associated ligands activate specific TLRs. For example,
lipopolysaccharide (LPS) activates TLR4, whereas microbial
DNA (CpGs) activates TLR9. Outside their academic inter-
est, the responses induced by each of these ligands is also of
considerable medical important with LPS representing a
causative agent of sepsis and CpG’s are promising immu-
nomodulatory molecules.
While activation of distinct members of the TLR family
has been reported to activate a conserved, and well defined,
signaling response, there are also specialized responses
associated with activation of individual family members that
are less well defined. Within this investigation, while
confirming the activation of the characterized TLR pathway,
unique, phosphorylation events which were ligand-specific
and not previously associated to TLR activity were defined
and validated (Fig. 2). For example, array data predicted
Tyr40 phosphorylation of Etk in response to LPS stimula-
tion, but not CpG stimulation. This specific response was
confirmed through the use of a phosphorylation-specific
antibody. Additionally, based on patterns of phosphorylation
of peptides corresponding to the p40phox and p47phox
proteins the arrays predicted activation of the oxidative burst
by CpG, but not LPS. The phenotypic consequences of these
apparent differences in signaling were confirmed through
functional assays. This highlights the ability of kinome
arrays to describe complex phenotypes that are multifaceted
in their regulation through phosphorylation [25].
Kinomic characterization with peptide arrays of human
monocyte responses to activation with LPS have also been
characterized by other labs with a greater emphasis on the
temporal changes that occur following exposure to a ligand
[26]. The results of this study were in close agreement with
our study and confirm the interspecies conservation of TLR
signaling pathway. These results provided novel insight into
the complexity and mechanisms of TLR signaling and
highlight the utility and power of the kinome arrays.
2.2 Understanding signaling responses induced by
phenotypically conserved, but mechanistically
distinct, ligands
Peptide arrays have been successfully applied to elucidate
signaling events induced by individual ligands, which may
share common signaling events despite interacting with
distinct receptors. The cytokines IL-32 and IL-17 are emer-
ging as critical players in the pathophysiology of chronic
inflammation and autoimmune disease. It is speculated that
the molecular mechanisms governing IL-32- and IL-17-
mediated cellular responses are differentially dependent on
the TNF pathway. Peptide-array-based kinome analysis was
used to demonstrate that a peptide target corresponding to
TNF-receptor-1 (TNF-R1) was phosphorylated in the
presence of cytokine IL-32 but not IL-17. This critical finding
was then independently validated through the use of a
monoclonal antibody to specifically block TNF-R1 and
Figure 2. Conserved and
distinct TLR4 and TLR9 signal-
ing in bovine monocytes:
Signaling events indicated by
peptide array kinome analysis
in bovine monocytes following
treatment with LPS or CpG.
Canonical signaling pathways
as well as novel signaling
events were observed. Adapted
from Arsenault et al. (2009) [25].
Proteomics 2011, 11, 4595–4609 4599
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

suppress IL-32-induced downstream responses. This
approach provided independent confirmation that IL-32-
mediated activity was dependent on TNF-R1. In contrast,
and consistent with the array results, blocking TNF-R1 did
not impact IL-17-induced downstream responses. This study
provided critical insight into the molecular mechanism of
the differential dependence of IL-32 and IL-17 on the TNF-
pathway and is likely to be of immediate therapeutic
importance.
The same investigation of IL-32- and IL-17-induced
signaling responses also identified p300 (transcriptional
co-activator) and death-associated protein kinase-1 (DAPK-1)
as common protein phosphorylation targets for IL-32 and IL-
17. Phosphorylation of p300 and DAPK-1 upon cytokine
stimulation was confirmed by immunoblots. The presence of
these common targets was further supported by additional
results demonstrating similar transcriptional responses, the
direct activation of nuclear factor k-light-chain-enhancer of
activated B cells (NF-kB) and the induction of chemokine
production by both IL-32 and IL-17. Furthermore, knock
down of p300 and DAPK-1 altered downstream responses
induced by IL-32 and IL-17, and impacted certain cellular
responses induced by TNF-a and IL-1b. These findings
allowed the authors to hypothesize that p300 and DAPK-1
represent nodes where the inflammatory networks of IL-32
and IL-17 overlap, and that p300 and DAPK-1 impact both
TNF-dependent and -independent processes. Therefore p300
and DAPK-1 may be viable therapeutic targets for chronic
inflammatory diseases [27]. Importantly, in this example, use
of peptide arrays greatly accelerated discovery in contrast to
using other techniques to investigate hypothesized targets
one at a time.
2.3 Understanding phenotypic variability
The power of kinome analysis, and in particular utilizing
peptide arrays, is well exemplified by efforts to elucidate the
cellular mechanisms of phenotypic differences between
closely related cells. The challenge of these scenarios is that
the cells may appear to present a highly conserved infra-
structure of biomolecules but subtle differences in signal
transduction through this network may dictate unique
functional phenotypes. Intestinal Peyer’s patch (PP) regu-
latory cluster of differentiation (CD)211 B cells (Bregs)
suppress TLR9-induced innate immune responses.
However, it is unknown if TLR9 activation is similarly
regulated in PP Bregs. We investigated the TLR9 agonist,
CpG oligodeoxynucleotides (ODN) induced response in the
PP Bregs [28]. It was observed that PP CD211 Bregs had high
expression of TLR9 mRNA, but would not proliferate upon
stimulation with CpG ODN. Furthermore, while CD211 B
cells from blood secreted IgM or IL-12 following CpG ODN
stimulation, PP CD211 Bregs did not. We hypothesized that
PP CD211 Bregs unresponsiveness resulted from a lack of
ability of CpG to activate TLR9 signaling in this type of cell.
This hypothesis was confirmed when kinome analysis
showed dynamic patterns of phosphorylation in adapter
proteins of TLR such as interleukin-1 receptor-associated
kinase (IRAK)1, mitogen-activated protein kinase kinase
kinase 7 (TAK1), I-k-B kinase (IKK) and NF-B-p65 in blood
CD211 B cells stimulated with CpG, indicating an activa-
tion of the TLR9 pathway. In contrast, CpG ODN stimula-
tion of PP CD211 Bregs displayed phosphorylation events in
TLR9 adapter proteins indicative of TLR9 pathway inacti-
vation. This lack of apparent TLR9-signaling events for
peptides representing proteins which are early inter-
mediates in the TLR-signaling pathway indicated that
signaling is blocked close to the receptor (Fig. 3). These
Figure 3. TLR9 signaling differences in related cell types: There
are numerous phosphorylation differences between PP CD211
Bregs and blood CD211 B cells. The spots indicate the proteins
and residues which were differentially phosphorylated following
stimulation with CpG ODN. CD211 B cells from blood displayed
strong TLR9 responses which CD211 Bregs displayed no TLR9
response. Adapted from Booth et al. (2010) [28].
4600 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

observations point toward a novel means by which TLR-
expressing cells control TLR responses in cells with regu-
latory functions. Our observations suggest a novel
mechanism by which TLR responses are regulated in TLR-
expressing cells with regulatory functions [28].
2.4 Cancer profiling
Cancer is the prototypical kinase disease and for many
researchers the study of kinases is intimately associated
with the study of cancer. It is very often a disregulation
of a subset or even a single protein kinase that is the cause
of cancer. In fact, kinases and their regulators are
commonly mutated oncogenes and tumor suppressors [29].
Kinase targeting is by any measure a massive portion of
current cancer research. Thus, it is logical that cancer
has had an outsized impact on the field of kinome
analysis and the development of peptide arrays. The power
that peptide arrays have is their ability to screen multiple
(up to 1000 or more) kinase target sites in one experiment.
In the drug discovery field, this is an important considera-
tion and can significantly increase the rate of research. In
addition, the kinome profiling of various tumors and
cancerous cells provides important biological information
on how cancers from individual patients compare
with respect to what they have in common and what defines
their individual biology. Important work in this field has
been carried out on pediatric brain tumor profiling
[30], leukemia profiling [31], chondrosarcoma [32], colon
cancer [33].
The screening of therapeutic agents that inhibit key
phosphorylated signaling molecules is a major focus of
cancer research. However, a critical consideration is that the
animal model or cell culture-based model is accurately
representing the true biology of target cancer cells in the
host. The combination of species-specific kinome arrays and
the Bovine Leukemia Virus (BLV) ovine model system,
allowed analysis of changes in kinase activity that occur
between host and culture in transformed B cells [34]. The
results indicated that the phosphorylation patterns were
significantly altered when cancer cells were removed from
host and passaged for use in tissue culture. These changes
were in pathways that define transformation in this cell type,
which could possibly alter results if one was using such a
host-derived cell line for therapeutic screens. This observa-
tion suggests that the external environment of the cells has a
profound effect on biologically important cell signaling
events. This analysis, using high-throughput kinome tech-
niques, was able to identify key phosphorylation events that
define cancer progression in B cells. In addition, to deter-
mine the alteration of critical signaling events between
primary cancer cells in vivo and cultured cells in vitro. Loss
or gain of key signaling pathways when cells are cultured
has a significant impact on the translation of knowledge
from the bench to the clinic [34].
3 New opportunities for peptide arraykinome analysis
3.1 Species-specific peptide arrays
Until recently the peptide arrays that have been created for
kinome analysis were based on phosphorylation events
characterized from the homologous species. The vast
majority of characterized phosphorylation events are for
human and mouse with only limited information available
for other species. This creates a barrier for creation of arrays
for species where the phosphoproteome has yet to be
defined. However, as the specific sites for protein phos-
phorylation, and their subsequent biological consequences,
are often conserved we previously hypothesized that it would
be possible to predict the sequence contexts of phosphor-
ylation events in proteins of other species based on genomic
information.
To test this hypothesis, our lab interrogated Phosphosite
(www.phosphosite.org) and Phosphobase (phospho.
elm.eu.org) which are publically available online databases
that hold information on manually curated and literature-
based serine, threonine and tyrosine phosphorylation sites.
The information contained within these sites is predomi-
nantly for phosphorylation events from human and mouse.
Search results produce peptide sequences, usually about 15
amino acids in length, which correspond to characterized
phosphorylation sites. Information about the recognizing
kinase, as well as links that describe the biological function
of the specific modification, is listed. The amount of
sequence conservation between bovine and human kinase
phosphorylation target recognition sites was investigated by
searching nearly a thousand peptides of 15 amino acids in
length, representing human phosphorylation sites, against
the NCBI-NR protein database by Blastp program in order to
generate orthologous bovine peptides. The results indicated
that approximately half of the bovine sequences matched
100% to those of human. The majority of the remaining half
had limited sequence differences, usually amounting to only
1 or 2 amino acids. An annotation comparison between the
query and hit sequences was used to confirm that both
referred back to the same protein identity [16]. These results
show that phosphorylation sites are not fully conserved
between human and bovine and these differences would
limit the utility of using human arrays to analyze the bovine
kinome by this method. Interestingly, other labs have
utilized human-based peptides arrays for species as evolu-
tionarily distant as Arabidopsis [35]. This, in our opinion,
speaks more highly of the enthusiasm to apply peptide
kinome to species of interest, and the lack of species-
appropriate tools, than to the value or appropriateness of
attempting to transcend species barriers with generic
peptide arrays.
For our first generation bovine-specific peptide array, 300
peptides were selected from the initial list of 880 peptides.
Target peptides represented proteins involved in a spectrum
Proteomics 2011, 11, 4595–4609 4601
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
of biological events but with emphasis on proteins, path-
ways and processes associated with innate immunity.
Specific peptides were selected based on biological signifi-
cance and degree of sequence conservation surrounding the
phospho-acceptor. These criteria were predicated on the
assumption that sequence conservation may reflect func-
tional conservation. Notably, many proteins undergo phos-
phorylation at multiple sites to control discrete aspects of
protein function. Therefore, numerous proteins on the array
were represented by multiple peptides, representing distinct
phosphorylation events, to allow more detailed insight into
cellular responses under different treatment conditions.
This has proven a powerful feature of the arrays in providing
a comprehensive assessment of how critical proteins are
regulated by complex and dynamic patterns of phosphor-
ylation at multiple sites. Each peptide was printed in tripli-
cate on the array to provide a measure of technical
reproducibility within each experiment (Fig. 4).
Our first generation arrays, and many of the arrays
reported in the literature, employ identical blocks that are
printed in triplicate. There may be an advantage, however, to
randomly place peptides within replicate blocks to minimize
possible effects resulting from the relative placement of a
peptide within the grid. Peptide synthesis and printing is
performed by a commercial partner (JPT Technologies) and
utilization of the arrays in the lab does not depend on any
specialized equipment that would not be present in labs
conducting transcriptional array investigations.
We have employed a similar bioinformatics approach to
develop a porcine peptide array. A similar degree of conser-
vation was observed when using human phosphoproteome
databases to identify putative porcine phosphorylation sites as
was encountered for the bovine array. Generation of arrays
for species for more evolutionarily distant organisms does
become problematic. For example, peptide sequences corre-
sponding to thousands of characterized phosphorylation
events were investigated using the same protocol to establish
the degree of conservation across several mammalian species
as well as select plants and insects. For organisms such as
plants and insects it is much more problematic to predict
phosphorylation sites based on these linear bioinformatic
approaches (Table 2). For these organisms, greater effort to
take into account unique biology associated with these
species as well as degrees of kinase conservation, their
consensus sequences and phosphorylation targets will also
assist in creating arrays for these species.
Databases are emerging describing the kinase comple-
ment for economically important plant species such as rice
(phylomics.ucdavis.edu/kinase/). Given the economic
importance of these organisms, in particular plants, and the
potential for kinome analysis to offer functional insight into
phenotypes and provide biomarkers for selective breeding,
these efforts are certainly justified. Interestingly, a number
of groups have reported on peptide array kinome analysis of
plants utilizing peptide arrays corresponding to human
phosphorylation sequences.
3.1.1 Non-traditional species
A tremendous advantage of kinome analysis for systems
biology investigations is that kinases are highly conserved
across species. This facilitates extrapolation of research find-
ings to other species and allows researchers to test inhibitors
of human kinases in animal models of disease. The only, but
often overlooked, caveat is the requirement for appropriatemodels of disease. The recent characterization of the mouse
kinome has been heralded for its ability to ‘‘enhance the
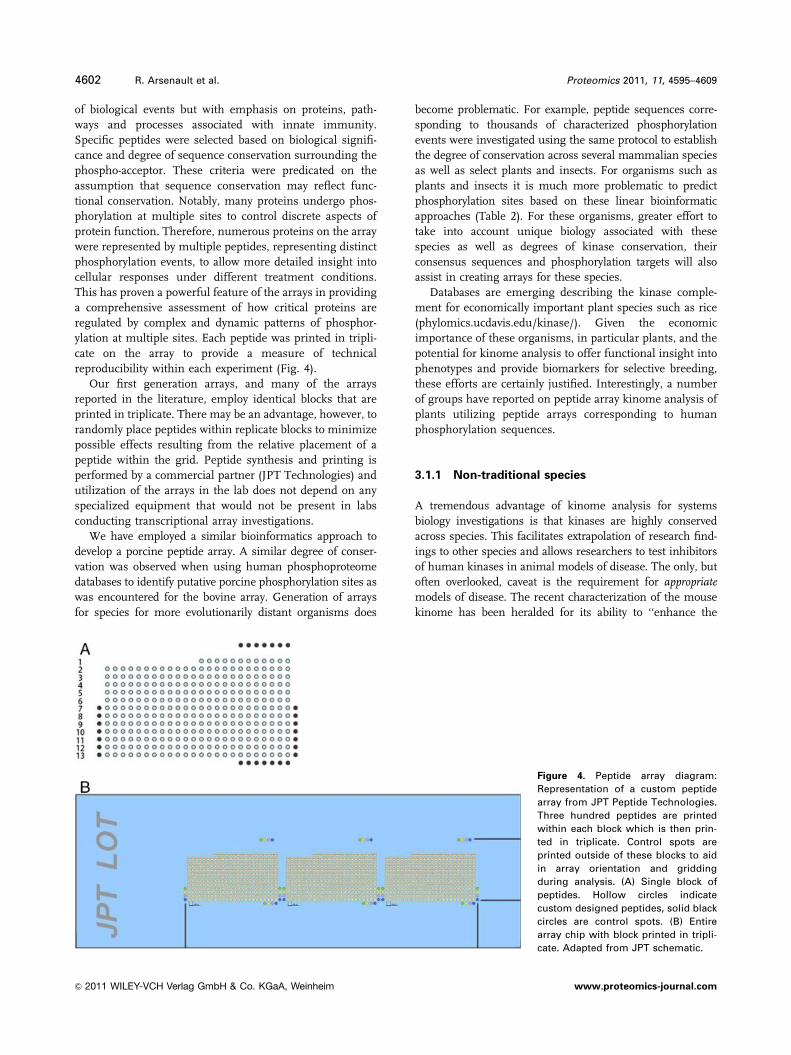
Figure 4. Peptide array diagram:
Representation of a custom peptide
array from JPT Peptide Technologies.
Three hundred peptides are printed
within each block which is then prin-
ted in triplicate. Control spots are
printed outside of these blocks to aid
in array orientation and gridding
during analysis. (A) Single block of
peptides. Hollow circles indicate
custom designed peptides, solid black
circles are control spots. (B) Entire
array chip with block printed in tripli-
cate. Adapted from JPT schematic.
4602 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
exploration of the roles of all kinases in mouse models of
human diseases’’ [36]. While mice offer unprecedented
opportunity for generation of mutants to study kinase func-
tion and consequences of kinase ablation, the value of the
mouse as a model for human disease are far less definitive.
The availability, cost and ease of handling inbred,
genetically defined mouse strains have made this the species
of choice for most animal experimental work. Unfortunately,
these selection criteria do not ensure accurate disease models
and efficient knowledge translation. An ever increasing
number of infection studies, with a variety of different
stimuli, pathogens and treatments, have demonstrated
divergent biological mechanisms, responses and distinct
pathologies when comparing mice and humans [37]. This
has prompted many researchers to invest greater resources
into the development of animal models, which more accu-
rately reproduce relevant human physiology and pathology.
Furthermore, the use of animals in biomedical research
ethically requires selection of species that match the human
disease as closely as possible to minimize the number of
animals required and maximize the potential benefit.
There are a number of factors that limit the capacity of
mouse models to accurately reflect human responses. First,
from our specific interest in infection and immunity, when
vaccinating or challenging an outbred population it has been
frequently observed that there is a binomial distribution
in immune responses referred to as ‘‘low’’ and ‘‘high’’
responders [38]. This reflects a genetic component that
influences the magnitude of immune responses within
individual animals. This individual variation is an essential
consideration when developing therapeutics for humans
and is best represented by outbred species. Second, effective
animal models must share the same physiological char-
acteristics as humans, or at least reflect them as closely as
possible. For example, the skin physiology of the pig is very
similar to humans that render the pig a good model for
studying intracutaneous or topical delivery of compounds.
3.1.2 Interspecies comparative analysis
Efficient translation of research from animal models to
clinical application will require more detailed knowledge
regarding conservation of cell signaling responses in model
species. For comparative kinomic investigations, it is
possible to design arrays reflecting specific biology or
signaling pathways for all species of interest. This can be
achieved through the selection of species-specific peptides
from orthologous proteins when sufficient proteomic
information is available for all species. Alternatively, it is
possible to design arrays using peptides that are absolutely
conserved in sequence for all species of interest. These
arrays could then be used to interrogate samples of interest
from each species. This approach would minimize potential
technical problems associated with the use of different
arrays but it may be necessary to then validate that
conserved peptides are selected from orthologous proteins.
The use of outbred species for kinome analysis also raises
a number of challenges due to genetic and environmental
differences among individual animals. Both these factors
may significantly influence kinase activity before and after
experimental manipulation of isolated cells or the host.
Therefore, experimental design and subsequent data analysis
must accommodate these potential sources of data variation.
Variation in kinase activity among individual animals has
presented a challenge when analyzing data for conserved
responses to a specific pathogen or stimulus. This variation
in kinase activity may also provide an opportunity to further
understand the biological basis for specific differences in
host phenotype or responses. We have initiated kinome
studies in animal populations with defined differences in
their susceptibility to pathogen infection. It is too early in
these studies to determine whether kinome analysis will
provide specific biomarkers of disease resistance and
susceptibility or provide significant insight into the biological
mechanisms mediating these specific phenotypes.
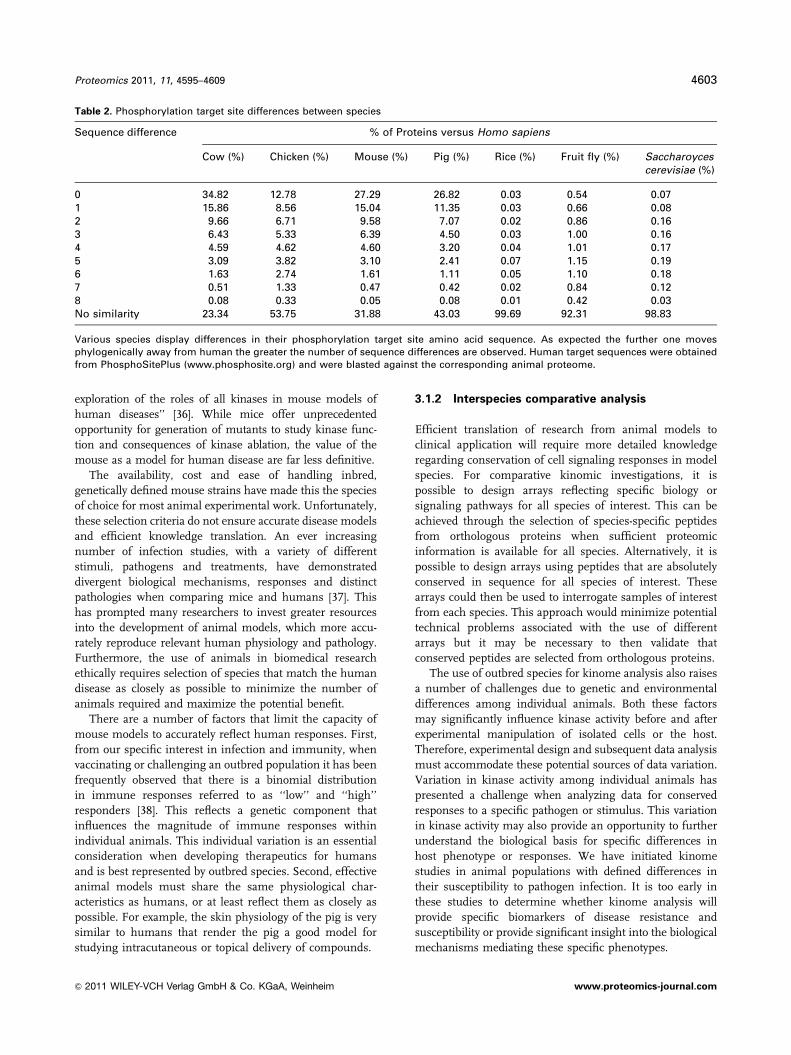
Table 2. Phosphorylation target site differences between species
Sequence difference % of Proteins versus Homo sapiens
Cow (%) Chicken (%) Mouse (%) Pig (%) Rice (%) Fruit fly (%) Saccharoycescerevisiae (%)
0 34.82 12.78 27.29 26.82 0.03 0.54 0.071 15.86 8.56 15.04 11.35 0.03 0.66 0.082 9.66 6.71 9.58 7.07 0.02 0.86 0.163 6.43 5.33 6.39 4.50 0.03 1.00 0.164 4.59 4.62 4.60 3.20 0.04 1.01 0.175 3.09 3.82 3.10 2.41 0.07 1.15 0.196 1.63 2.74 1.61 1.11 0.05 1.10 0.187 0.51 1.33 0.47 0.42 0.02 0.84 0.128 0.08 0.33 0.05 0.08 0.01 0.42 0.03No similarity 23.34 53.75 31.88 43.03 99.69 92.31 98.83
Various species display differences in their phosphorylation target site amino acid sequence. As expected the further one movesphylogenically away from human the greater the number of sequence differences are observed. Human target sequences were obtainedfrom PhosphoSitePlus (www.phosphosite.org) and were blasted against the corresponding animal proteome.
Proteomics 2011, 11, 4595–4609 4603
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Transcriptional analysis has been used to compare host
responses to a variety of pathogens and identify pathogen-
specific immune evasion strategies [39]. Validation of sustained
transcriptional responses following infection is difficult and
recent evidence that pathogens produce miRNA [40] further
complicates the interpretation of host transcriptional responses.
We have used kinome arrays biased for innate immune
responses to compare cellular responses to a variety of patho-
gens. These analyses provide the opportunity to not only identify
specific innate immune responses that have been disrupted, but
also identify the specific adapter proteins that have been targeted
by the pathogen. This level of knowledge may be critical when
comparing clinical isolates of pathogen that vary in virulence or
designing attenuated pathogens for use in vaccines.
3.2 Process-specific peptide arrays
An additional advantage of the peptide arrays is that
peptides can be selected to enable focussed investigation of a
particular priority cellular process. Such wilful construction
offers the researcher richer information about signaling
events associated with a particular biological response such
as apoptosis, cell division or, in the case of our bovine array,
innate immunity. Our approach to date has been to devote a
considerable portion (half to two thirds) of the peptides on
the array to central, but generic, signaling pathways. This
enables novel discovery of how a stimulus may impact
cellular responses in a manner which may not be immedi-
ately intuitive. The remaining peptides represent focussed,
hypothesis-driven selections which enable the researcher to
better interrogate specific cellular responses which are of
priority. As the design and production of customized arrays
becomes increasingly routine it is easy to envision kinome
investigations through peptide arrays following an iterative
process of array design with increasing focus and repre-
sentation of particular processes within an investigation.
3.3 Non-radioactive protocols
Many of the initial peptide array kinome protocols utilized32P-g-ATP for detection and quantification of peptide phos-
phorylation. While effective such approaches are disadvan-
taged in their cost, sensitivity and safety. More recent
manuscripts describe the use of phospho-specific fluor-
escent stains that are compatible with the arrays, less costly
and suitable for use with currently available microarray data
scanners. In our hands the use of these stains, in particular
Diamond ProQ phosphostain, offers greater sensitivity and
reproducibility than radioactive protocols [25].
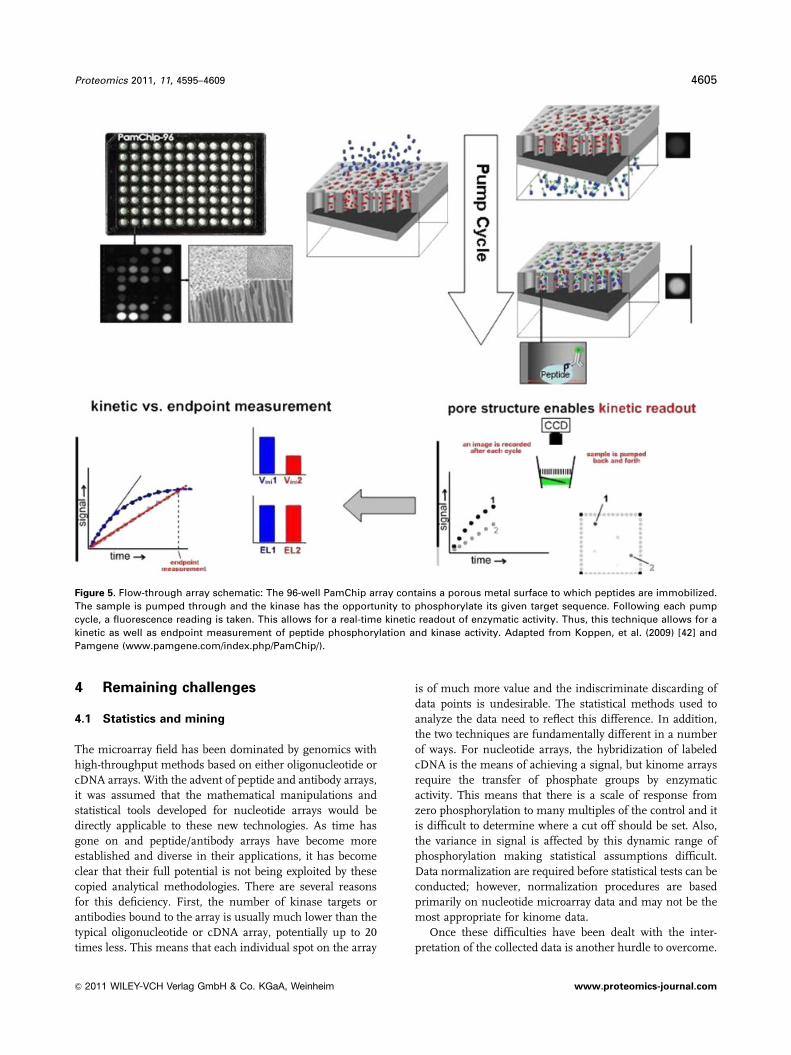
3.4 Flow-through 3D arrays
A variation of the peptide array for kinome analysis discussed
above is the flow-through 3D array [41]. This technology is
epitomized by the PamChip system developed by Pamgene of
the Netherlands. Peptides that correspond to specific kinase
target sites are covalently immobilized on an activated alumi-
num oxide surface. The unique nature of this array is the
surfaces on which the peptides are immobilized are a series of
pores through which the analyte solution can be pumped. As
the solution is pumped back and forth through the pores any
kinase contained within will phoshorylate its corresponding
target peptide. This is visualized using phosphorylation-specific
antibodies bound to a fluorophore. The pumping rate and
number of cycles can be varied and a fluorescence reading is
taken after every pump cycle. Following each pumping cycle
more peptides will be phosphorylated resulting in greater
fluorescent signal. This allows the user to take kinetic as well as
end point measurements of kinase enzyme activity (Fig. 5) [42].
This technique has been applied for a number of important
studies including kinome profiling of pediatric brain tumors
[30], development of protein kinase C (PKC) inhibitors [43] and
leukemia treatment target identification [31].
3.5 Bead-based solution-phase heterogeneous
kinase assays
Solution-phase peptide-based kinase assays are an alter-
native to the arrays based on immobilized peptide on a solid
array surface discussed above. The technique is based on the
covalent linking of peptide substrates onto beads via an
acrylamide linker [44]. Each individual bead will contain a
number of identical peptide substrates linked to its surface.
A mixture of beads containing different peptide substrates
are combined within a single well of a 96-well plate. To these
wells is added cell lysates that contain active protein kinases.
These kinases will then phosphorylate their respective target
peptides. The reaction is then stopped and synthetically
phosphorylated internal standards are added to beads.
Fluorescent antibodies against phospho-peptides are added
to the wells to bind to the phosphorylated peptide. Sylvester
and Kron [44] used Luminex beads and the Luminex flow
system to identify both the beads, thus the peptide substrate,
and the antibody, thus the peptides phosphorylation state,
through a measurement of two channel fluorescence. The
fluorescent of the bead indicated peptide identification and
the fluorescence of the phospho-specific antibody indicated
phosphorylation of the peptide substrate by the active kinase
in the sample. The ability to screen multiple peptides within
a single well of a 96-well plate allows various conditions,
concentrations or treatments to be assayed within a single
experiment. The main advantage of this technique over
solid-supported peptide substrates is that the kinases have
greater access to their target peptides within solution [45].
With a uniform distribution within solution, beads linked to
peptide substrates have a higher reaction rate with kinases
than solid-phase arrays. In addition, beads provide a larger
surface area for reactions than a two-dimensional array
surface and a high loading capacity of peptide on the beads.
4604 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
4 Remaining challenges
4.1 Statistics and mining
The microarray field has been dominated by genomics with
high-throughput methods based on either oligonucleotide or
cDNA arrays. With the advent of peptide and antibody arrays,
it was assumed that the mathematical manipulations and
statistical tools developed for nucleotide arrays would be
directly applicable to these new technologies. As time has
gone on and peptide/antibody arrays have become more
established and diverse in their applications, it has become
clear that their full potential is not being exploited by these
copied analytical methodologies. There are several reasons
for this deficiency. First, the number of kinase targets or
antibodies bound to the array is usually much lower than the
typical oligonucleotide or cDNA array, potentially up to 20
times less. This means that each individual spot on the array
is of much more value and the indiscriminate discarding of
data points is undesirable. The statistical methods used to
analyze the data need to reflect this difference. In addition,
the two techniques are fundamentally different in a number
of ways. For nucleotide arrays, the hybridization of labeled
cDNA is the means of achieving a signal, but kinome arrays
require the transfer of phosphate groups by enzymatic
activity. This means that there is a scale of response from
zero phosphorylation to many multiples of the control and it
is difficult to determine where a cut off should be set. Also,
the variance in signal is affected by this dynamic range of
phosphorylation making statistical assumptions difficult.
Data normalization are required before statistical tests can be
conducted; however, normalization procedures are based
primarily on nucleotide microarray data and may not be the
most appropriate for kinome data.
Once these difficulties have been dealt with the inter-
pretation of the collected data is another hurdle to overcome.
Figure 5. Flow-through array schematic: The 96-well PamChip array contains a porous metal surface to which peptides are immobilized.
The sample is pumped through and the kinase has the opportunity to phosphorylate its given target sequence. Following each pump
cycle, a fluorescence reading is taken. This allows for a real-time kinetic readout of enzymatic activity. Thus, this technique allows for a
kinetic as well as endpoint measurement of peptide phosphorylation and kinase activity. Adapted from Koppen, et al. (2009) [42] and
Pamgene (www.pamgene.com/index.php/PamChip/).
Proteomics 2011, 11, 4595–4609 4605
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

With any high-throughput method there will be a wealth of
data generated and one must decide how to mine it for
significant and biologically relevant results. This has been
historically difficult with kinomic array data and many
papers have focused on identifying data which confirm
previous low-throughput methods and then focusing on one
or two novel phosphorylation events that can be confirmed
with Western blots or bioassays. While it is important that
array data agree with the previous reports the main goal of
this method is to discover novel biology and in many cases
uncover targets for therapy. A more systematic data mining
approach is needed to fully explore the value of kinome data.
4.2 Kinase specificity/efficiency
A great unknown that all kinomic researchers are faced with
is the specificity with which an individual kinase recognizes
a spotted target sequence and how efficiently it phosphor-
ylates that sequence. While enzymatic work has been done
showing kinases phosphorylate target peptides with
comparable mechanics to native protein [46, 47], the speci-
ficity and efficiency of each individual kinase or each target
sequence is unknown. This can greatly affect the data
interpretation if one considers the magnitude of phosphor-
ylation on the array as biologically significant. Indeed as
Sikkema et al. [30] wrote in their kinome profiling study,
‘‘ydetermining the sensitivity of a substrate for specific
kinases will prove to be of tremendous value in future
application of the peptide array.’’ They go on to summarize
the largest drawback of the current state of peptide array
kinome research. ‘‘Therefore, generating hypotheses is the
limit of what is possible at the current developmental stage
of peptide microarray technology.’’ The peptide arrays in
their current state simply point us in directions of further
study using other, more established, methods. This is
further discussed in the following section.
4.3 A systematic approach needed
A new approach must be taken in the analysis of data
generated by the kinomic peptide arrays. Instead of simply
being a means of generating hypotheses for future work, the
peptide arrays can be developed into a tool to provide a
better picture of what is going on in the cell under a given
condition. This is currently not possible and will require
tools that synthesize all of the data from the array and
organize it into functional groupings or networks. This
‘‘value-added’’ approach to kinomic peptide array data will
allow researchers to not only simply find a single interesting
phosphorylation event to study further or give a list of
differential phosphorylation events between two treatments/
diseases, but provide biological relevant data. Examples may
include which cell signaling pathways are affected by a given
treatment and at what point is it diverging from control,
what cell function is affected by a disease and how a specific
function is linked to other core functions, such as metabo-
lism, cell cycle and apoptosis.
In addition to the previously mentioned additional
capacities, in the future integration of kinome data with
other high-throughput methods will be needed. As Parikh
and Peppelenbosch [48] wrote in their review of kinome
profiling and cancer ‘‘ythe future lies in moving beyond
the idea of an individual ‘‘omics’’ approach and studying a
biological system in its entirety by combining data from all
different omics technologies, so that disease can be better
modeled and thoroughly understood compared with a single
pathway approach.’’ The synthesis of genomics, tran-
scriptomics, proteomics and kinomics to aid in under-
standing is the important development still to come to move
peptide array kinomics into its own rather than being a
hypothesis generating technique.
4.4 Validation of kinome data
The philosophy of our lab is that the information emerging
from the arrays should be utilized to suggest signal beha-
viors and their associated phenotypes, which can then be
validated through independent experimental approaches.
Such validation could come in the form of targeted phos-
phoproteome investigations with phosphorylation-specific
antibodies. Alternatively, where species-appropriate phos-
phorylation-specific antibodies are not available, it is
necessary to experimentally investigate the phenotypic
events that are suggested by the kinome data such as
apoptosis, oxidative burst and pro/anti-inflammatory
responses. The practice of validating kinome results through
independent approaches also helps to determine the
number of biological, and most importantly technical,
replicates that are appropriate. The number of replicates
should be sufficient to ensure that a reliable and statistically
significant picture of the signal emerges.
4.5 Biological importance
The protocols employed for peptide-array-based kinome
analysis introduce biological artefacts which compromise the
value of the emerging data. In particular, disruption of cells
to release cellular kinases results in an inherent loss of
cellular structure and organization that is undoubtedly
biologically important. Loss of this organization within the
context of a cellular lysate likely impacts the activity of
kinases through the loss of regulating interactions. Similarly,
the peptide arrays may present a kinase the opportunity to
phosphorylate a substrate which it would not normally
encounter in the cell due to compartmentalization or the
absence of co-incidental substrate and kinase expression.
The specificity of kinases depends not only on their ability
to recognize and modify a protein target, but also on their
4606 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

spatial and temporal access to these targets. The intracellular
environment is highly complex and organized with the
particular and dynamic location of various biomolecules,
including kinases and their substrates, providing an impor-
tant level of control for biological functions. As such the
translocation of a cellular kinase from the cytosol to more
specialized locations can serve to regulate activity. For
example, b-adrenergic receptor kinase translocates from the
cytosol to the cell membrane to phosphorylate and desensi-
tize G-protein-coupled receptors [49–51], while mitogen-
activated protein kinase (MAPK) translocates from the cyto-
sol to the nucleus to regulate gene transcription [52]. This
situation can become even more complex for multipurpose
kinases that regulate distinct and functionally unrelated
processes within in the same cell. For example, PKCdregulates both depolarization events, apoptosis or
cell growth through either translocation to the cell
membrane to inhibit inward recitifying K1 channels or
translocation to the mitochondria to activate ATP-dependent
K1 channels or translocation to the nucleus for regulation of
gene transcription respectively [53–55]. Depending upon the
specific needs of a cell these distinct patterns of translocation
and associated responses can be achieved simultaneously.
Targeted regulation of kinase activity through compart-
mentalization is not limited to dynamic patterns of kinase
localization, but is also achieved through the discrete
formation of factors that regulate kinase activity. For
example, formation of cyclic adenosine monophosphate
(cAMP), a potent activator of protein kinase A, has been
shown to occur within discreet microdomains in cardiac
myocytes in response to b-adrenergic stimulation [49]. This
in turn would be anticipated to result in equally compart-
mentalized activation of protein kinase A (PKA), which was
suggested by the authors of the paper to represent a
mechanism for compartmentalized activation of PKA in
specific sub-cellular locations [56].
The loss of cellular organization likely represents a
greater challenge for kinome rather than phosphoproteome
investigations as kinome analysis depends on measure-
ments of enzymatic activity which are taken post-lysis,
whereas phosphoproteome analysis characterizes events
which occur prior to cell lysis.
5 Concluding remarks
A reliable methodology for characterizing host signal
transduction activity offers tremendous promise to under-
stand both cellular physiology and pathophysiology. This is
not a trivial task giving the complexity and dynamic nature
of these responses. Given the current demonstrated poten-
tial for utilizing the kinases to understand biology as well as
providing targets for therapeutic intervention, these efforts
are well justified. As we continue to realize the potential of
kinase inhibitors tools such as the peptide arrays for kinome
analysis may ultimately lead to better application of a wide
variety of therapies to enhance individual patient medicine
and minimize the risk of adverse reactions.
While much of this initial excitement has been within the
context of cancer the potential to define the cellular
mechanisms of disease through kinome analysis and the
opportunity to treat these diseases with kinases inhibitors is
being appreciated for an increasingly wide spectrum of
diseases. We anticipate that in the future kinome analysis
will have increasingly important roles in the study of
host–pathogen interactions. The interaction between a
particular pathogen and its target cell are often complex and
multifaceted presenting a considerable challenge for any
experimental approach. The effort required to understand
these complex interactions are justified, in particular for
pathogens that establish chronic infections, because
understanding the mechanisms used by a pathogen to
subvert the host response provides critical targets for ther-
apeutic intervention. In particular, many pathogens have
been shown to target host phosphorylation-mediated signal
transduction pathways. Therefore, understanding host
responses at the level of the kinome is highly appropriate.
Interestingly, the pathogenic mechanisms of several patho-
gens have been shown to involve the production eukaryotic-
like kinase effector molecules, which are translocated into
the host cell for direct subversion of host processes [57].
These eukaryotic-like kinases have been identified as highly
attractive therapeutic targets [58].
A large number of practical and philosophical questions
and challenges remain to be addressed for peptide-array
kinome analysis in the coming years. We anticipate that the
development of customized software packages for the analy-
sis and interpretation of kinome data will represent a turning
point in the utility of this approach. That is researchers can
place greater confidence in the biological meaning of
kinomics data with less requirement for development of
specialized expertise for understanding the data there will be
a greater uptake of the technology into non-specialized labs
wishing to embark in this sub-discipline of proteomics.
There also remains a key philosophical question about
the arrays that holds considerable practical importance. Do
they represent kinome arrays or phosphoproteome arrays?
Or both? As the arrays depend upon the activities of the
cellular kinases to phosphorylate the peptide targets this
would seem consistent with a kinome array. However, as the
extent of phosphorylation of the peptide is used as a
surrogate marker to predict the extent of phosphorylation of
the corresponding protein within the cell this is more
consistent with phosphoproteome analysis. This is not a
trivial distinction. For example, in each scenario, one might
consider the number of peptides that would be required for
a comprehensive array. From the perspective of a kinome
array just over 500 peptides, if they represent substrates of
ideally specificity of the corresponding kinases, would offer
a global perspective on the relative activities of each kinase.
From a phosphoproteome perspective, the array would
require peptides to represent each cellular phosphorylation
Proteomics 2011, 11, 4595–4609 4607
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

event, tens of thousands in number, to obtain a compre-
hensive understanding of phosphorylation-mediated signal
transduction events within the cell. There would obviously
be considerable differences as well in how this data would
be handled, interpreted and validated. Future studies will be
required to further define kinase specificity before the
optimal balance between peptide array design and data
interpretation can be achieved.
The authors acknowledge the financial contributions of theCanada Research Chairs program (Neonatal Mucosal Immu-nology) and the Natural Sciences and Engineering ResearchCouncil.
The authors have declared no conflict of interest.
6 References
[1] Krebs, E. G., Fischer, E. H., Phosphorylase activity of skeletal
muscle extracts. J. Biol. Chem. 1955, 216, 113–120.
[2] Fischer, E. H., Krebs, E. G., Conversion of phosphorylase b
to phosphorylase a in muscle extracts. J. Biol. Chem. 1955,
216, 121–132.
[3] Manning, G., Whyte, D. B., Martinez, R., Hunter, T., Sudar-
sanam, S., The protein kinase compliment of the human
genome. Science 2002, 298, 1912–1934.
[4] Hunter, T., Protein kinases and phosphatases: the yin and
yang of protein phsophorylation and signaling. Cell 1995,
80, 225–236.
[5] Manning, G., Caenepeel, S., Encyclopedia of protein kinases
in human diseases, Catalog Reference Manual, Cell
Signaling Technology, Beverly, MA, 2005.
[6] Knuutila, S., Bjorkqvist, A. M., Autio, K., Tarkkaqnen, M.
et al., DNA copy number amplifications in human
neoplasms: review of comparative genomic hybridization
studies. Am. J. Pathol. 1998, 152, 1107–1123.
[7] Cohen, P., Protein kinases–the major drug targets of the
twenty-first century? Nat. Rev. Drug Discov. 2002, 1, 309–315.
[8] Hopkins, A. L., Groom, C. R., The druggable genome. Nat.
Rev. Drug Discov. 2002, 1, 727–730.
[9] Druker, B., Tamura, S., Buchdunger, E., Ohno, S. et al.,
Effects of a selective inhibitor of the Abl tyrosine kinase on
the growth of Bcr-Abl positive cells. Nat. Med. 1996, 2,
561–566.
[10] Druker, B., Talpaz, M., Resta, D., Peng, B. et al., Efficacy and
safety of a specific inhibitor of the CR-ABL tyrosine kinase in
chronic myeloid leukemia. N. Engl. J. Med. 2001, 2,
561–566.
[11] Noble, M., Endicott, J., Johnson, L., Protein kinase inhibi-
tors: insights into drug design from structure. Science 2004,
303, 1800–1805.
[12] Eglen, R. M., Reisine, T., The current status of drug
discovery against the human kinome. Assay Drug Dev.
Tech. 2009, 7, 22–43.
[13] Liu, J., Farmer, J. D., Lane, W. S., Friedman, J. et al.,
Calcineurin is a common target of cyclophilin–cyclosporin A
and FKBP–FK506 complexes. Cell 199, 166, 807–815.
[14] Heitman, J., Movva, N. R., Hall, M. N., Targets for cell cycle
arrest by the immunosuppressant rapamycin in yeast.
Science 1991, 253, 905–909.
[15] Lee, J. C., Laydon, J. T., McDonnell, P. C., Gallagher, T. F.
et al., A protein kinase involved in the regulation of
inflammatory cytokine biosynthesis. Nature 1994, 372,
739–746..
[16] Jalal, S., Kindrachuk, J., Napper, S., Phosphoproteome and
kinome analysis: unique perspectives on the same problem.
Curr. Anal. Chem. 2007, 3, 1–15.
[17] Mann, M., Ong, S. E., Gronborg, M., Stern, H. et al., Trends
Biotechnol. 2002, 20, 261.
[18] Kalume, D. E., Molina, H. and Pandey, A., Tackling the
phosphoproteome: tools and strategies. Curr. Opin. Chem.
Biol. 2004, 7, 64–69.
[19] Gorg, A., Weiss, W., Dunn, M. J., Current two-dimensional
electrophoresis technology for proteomics. Proteomics
2004, 4, 3665–3685.
[20] Kreegipuu, A., Blom, N., Brunak, S., Jarv, J., Statistical
analysis of protein kinase specificity determinants. FEBS
Lett. 1998, 430, 45–50.
[21] Zhu, H., Klemic, J. F., Chang, S., Bertone, P. et al., Analysis
of yeast protein kinases using protein chips. Nature Genet.
2000, 26, 283–289.
[22] Ouyang, Z., Takats, Z., Blake, T. A., Gologan, B. et al.,
Preparing protein microarrays by soft-landing of mass-
selected ions. Science 2003, 301, 1351–1354.
[23] Houseman, B. T., Huh, J. H., Kron, S. J., Mrksich, M.,
Peptide chips for the quantitative evaluation of protein
kinase activity. Nature Biotech. 2002, 20, 270–274.
[24] Kindrachuk, L., Potter, J., Wilson, H., Griebel, P., Napper, S.,
Activation and regulation of toll-like receptor 9: CpGs and
beyond. Mini. Rev. Med. Chem. 2008, 8, 590–600.
[25] Arsenault, R. J., Jalal, S., Babiuk, L. A., Potter, A. et al.,
Kinome analysis of toll-like receptor signaling in bovine
monocytes. J. Recept. Signal. Transduct. Res. 2009, 29,
299–311.
[26] Diks, S. H., Kok, K., O’Toole, T., Hommes, D. W. et al.,
Kinome profiling for studying lipopolysaccharide signal
transduction in human peripheral blood mononuclear cells.
J. Biol. Chem. 2004, 279, 49206–49213.
[27] Turner-Brannen, E., Choi, K. G., Arsenault, R., El-Gagalwy,
H. et al., Inflammatory cytokines IL-32 and IL-17 have
common signaling intermediates despite differential
dependence on TNF-receptor 1. J. Immunol. 2011, 186,
7127–7135.
[28] Booth, J. S., Arsenault, R., Napper, S., Griebel, P. J. et al.,
TLR9 signaling failure renders Peyer’s patch regulatory B
cells unresponsive to stimulation with CpG oligodeoxy-
nucleotides. J. Innate Immun. 2010, 5, 483–494.
[29] Knight, Z. A., Lin, H., Shokat, K. M., Targeting the cancer
kinome through polypharmacology. Nat. Rev. Cancer. 2010,
10, 130–137.
4608 R. Arsenault et al. Proteomics 2011, 11, 4595–4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

[30] Sikkema, A. H., Diks, S. H., den Dunnen, W. F. A., ter Elst, A.
et al., Kinome profiling in pediatric brain tumors as a new
approach for target discovery. Cancer Res. 2009, 69,
5987–5995.
[31] ter Elst, A., Diks, S. H., Kampen, K. R., Hoogerbrugge, P. M.
et al., Identification of new possible targets for leukemia
treatment by kinase activity profiling. Leukemia Lymphoma
2011, 52, 122–130.
[32] Schrage, Y. M., Briaire-de Bruijn, I. H., de Miranda, N. F. C.
C., van Oosterwijk, J. et al., Kinome profiling of chon-
drosarcoma reveals SRC-pathway activity and dasatinib as
option for treatment. Cancer Res. 2009, 69, 6216–6222.
[33] Tuynman, J. B., Vermeulen, L., Boon, E. M., Kemper, K.
et al., Cyclooxygenase-2 inhibition inhibits c-Met kinase
activity and Wnt activity in colon cancer. Cancer Res. 2008,
68(4),1213–1220.
[34] Van den Broke, A., Arsenault, R., Cleuter, Y., Dehouck, C.,
Kinome profiling of bovine leukemia virus-indiced ovine
leukemia: an approach for identifying altered signaling
pathways and drugable targets in cancer. JAIDS 2011, 56,
98.
[35] Ritsema, T., Joore, J., van Workum, W., Pieterse, C. J.M.,
Kinome profiling of Arabidopsis using arrays of kinase
consensus substrates. Plant Methods 2007, 3, 3.
[36] Caenepeel, S., Charydczak, G., Sudarsanam, S., Hunter, T.,
Manning, G., The mouse kinome: discovery and compara-
tive genetics of all mouse protein kinases. Proc. Natl. Acad.
Sci. USA 2004, 32, 11707-11712.
[37] Gerdts, V., van den Hurk, S., Griebel, P., Babiuk, L., Use of
animal models in the development of human vaccines.
Future Med. 2007, 2, 667–675.
[38] Wilkie, B. N., Mallard, B. A., Genetic effects on vaccination.
Adv. Vet. Med. 1999, 41, 39–51.
[39] Wilson, K. T., Crabtree, J. E., Immunology of Helicobacter
pylori: insights into the failure of the immune response and
perspectives on vaccine studies. Gastroenterology 2007,
133, 288–308.
[40] He, S., Yang, Z., Skogerbo, G., Ren, F. et al., The properties
and functions of virus encoded microRNA, siRNA, and other
small noncoding RNAs. CRC Crit. Rev. Microbiol. 2008, 34,
175–188.
[41] Hilhorst, R., Houkes, L., van den Berg, A., Ruijtenbeek, R.,
Peptide microarrays for detailed, high-throughput substrate
identification, kinetic characterization, and inhibition
studies on protein kinase A. Anal. Biochem. 2009, 387,
150–161.
[42] Koppen, A., Houtman, R., Pijnenburg, D., Jeninga, E. H.
et al., Nuclear receptor coregulator interaction profiling
identifies TRIP3 as a novel peroxisome proliferator-
activated receptor y cofactor. Mol. Cell. Proteomics 2009, 8,
2212–2226.
[43] Poot, A. J., van Ameijde, J., Slijper, M., van den Berg, A.
et al., Development of selective bisubstrate-based inhibitors
against protein kinase C (PKC) isozymes by using dynamic
peptide microarrays. ChemBioChem 2009, 10, 2042–2051.
[44] Sylvester, J. E., Kron, S. J., A bead-based activity screen for
small-molecule inhibitors of signal transduction in chronic
myelogenous leukemia cells. Mol. Cancer Ther. 2010, 9,
1469–1481.
[45] Wu, D., Sylvester, J. E., Parker, L. L., Zhou, G., Kron, S. J.,
Peptide reporters of kinase activity in whole cell lysates.
Biopolymers 2010, 94, 475–486.
[46] Zetterqvist, O., Ragnarsson, U., Humble, E., Berglund, L.,
Engstrom, L., The minimum substrate of cyclic AMP-
stimulated protein kinase, as studied by synthetic peptides
representing the phosphorylatable site of pyruvate kinase
(type L) of rat liver. Biochem. Biophys. Res. Commun. 1976,
70, 696–703.
[47] Kemp, B. E., Graves, D. J., Benjamini, E., Krebs, E. G., Role
of multiple basic residues in determining the substrate
specificity of cyclic AMP-dependent protein kinase. J. Biol.
Chem. 1977, 252, 4888–4894,
[48] Parikh, K., Peppelenbosch, M. P., Kinome profiling of clin-
ical cancer specimens. Cancer Res. 2010, 70, 2575.
[49] Benovic, J. L., Strasser, R. H., Caron, M. G., Lefkowitz, R. J.,
Beta-adrenergic receptor kinase: identification of a novel
protein kinase that phosphorylates the agonist-occupied
form of the receptor. Proc. Natl. Acad. Sci. USA 1986, 83,
2797–2801.
[50] Strasser, R. H., Benovic, J. L., Caron, M. G., Lefkowitz, R. J.,
Beta-agonist and prostaglandin EI-induced translocation of
the beta-adrenergic receptor kinase: evidence that the
kinase may act on multiple adenylate cyclase-coupled
receptors. Proc. Natl. Acad. Sci. USA 1986, 83, 6362–6366.
[51] Benovic, J. L., Mayor, F., Somers, R. L., Caron, M. G.,
Leftowitz, R. J., Light-dependent phosphorylation of
rhodopsin by beta-adrenergic receptor kinase. Nature 1986,
321, 869–872.
[52] Pouyssegur, J., Lenormand, P., Fidelity and spatio-temporal
control in MAP kinase (ERKs) signalling. Eur. J. Biochem.
2003, 270, 3291–3299.
[53] Perillan, P. R., Chen, M., Potts, E. A., Simard, J. M., Trans-
forming growth factor beta1 regulates Kir2.3 inward recti-
fier K1channels via phospholipase C and protein kinase C
gamma in reactive astrocytes from adult rat brain. J. Biol.
Chem. 2001, 227, 1974–1980.
[54] Majumder, P. K., Mishra, N. C., Sun, X., Bharti, A. et al.,
Targeting of protein kinase C gamma to mitochondria in the
oxidative stress response. Cell Growth Differ. 2001, 12,
465–470.
[55] O’Flaherty, J. T., Chadwell, B. A., Kearns, M. W., Sergeant,
S., Daniel, L. W., Protein kinases C translocation responses
to low concentrations of arachidonic acid. J. Biol. Chem.
2001, 276, 24743–24750.
[56] Skalhegg, B. S., Tasken, K., Specificity in the cAMP/PKA
signalling pathway. Differential expression, regulation, and
subcellular localization of subunits of PKA. Front Biosci.
2000, 5, d679–d693.
[57] Alber, T., Signaling mechanisms of the Mycobacterium
tuberculosis receptor Ser/Thr protein kinases. Curr. Opin.
Struct. Biol. 2009, 6, 650–657.
[58] Schreiber, M., Res, I., Matter, A., Protein kinases as
antibacterial targets. Curr. Opin. Cell Biol. 2009, 2,
325–330.
Proteomics 2011, 11, 4595–4609 4609
& 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com