persistence in massachusetts of the veined white butterfly due to use of the invasive form of cuckoo...
TRANSCRIPT

ORIGINAL PAPER
Persistence in Massachusetts of the veined white butterflydue to use of the invasive form of cuckoo flower
M. V. Herlihy • R. G. Van Driesche •
D. L. Wagner
Received: 2 August 2013 / Accepted: 7 April 2014
� Springer International Publishing Switzerland 2014
Abstract The native pierid butterfly Pieris oleracea
underwent a large range reduction in New England in
the twentieth century, likely due to the introduction the
invasive butterfly Pieris rapae (Lep.: Pieridae) to
North America in 1860, and later the oligophagous
parasitoid Cotesia glomerata (Hymenoptera: Bracon-
idae) in 1884. Thought extirpated from the state by the
1970s, one large dense population of the butterfly was
found in the mid 1980s in a flood plain meadow along
the Housatonic River in Lenox, Berkshire Co., Mas-
sachusetts. We examined how this native pierid was
able to maintain a relatively dense local population by
feeding on a novel, invasive host plant, Cardamine
pratensis (cuckoo flower), in a meadow habitat despite
known parasitoid presence. We approached this ques-
tion in three ways. First, we deployed trap host plants
(cuckoo flower and collards) stocked host larvae (first
and second instars of either P. rapae or P. oleracea) at
the Lenox site and other locations to determine current
rates of C. glomerata attack, for comparison with
historical information. Second, we used olfactometer
experiments to determine if C. glomerata females
could detect the cuckoo flower volatiles released
during P. oleracea larval feeding. Third, we used field-
cage experiments to determine if the plant architecture
found in the flood plain meadow inhibited the ability
of C. glomerata females to locate and parasitize hosts.
Specifically, we asked if overtopping vegetation
prevented or reduced parasitism of P. oleracea larvae
feeding on the covered basal rosettes of C. pratensis,
which is the physical form of host plant for three of the
four butterfly generations at the site.
Keywords Biological control � Non-target
impact � Enemy-free space � Parasitoid
displacement � Pieris oleracea � Pieris rapae �Pieris napi � Cotesia glomerata � Cotesia
rubecula � Cardamine pratensis
Introduction
In the mid-to-late nineteenth century, the native
butterfly Pieris oleracea Harris (formerly Pieris napi
oleracea) began a decline in Massachusetts (Scudder
1889) that culminated in its apparent disappearance
from most of the state, with populations persisting in
parts of Berkshire County in western Massachusetts.
In 1986, a large population was rediscovered in
western Massachusetts (Lenox, Berkshire Co.) by
Roger Pease, an amateur Lepidopterist. It was later
determined that the larvae of this population fed
primarily on an introduced subspecies of cuckoo
M. V. Herlihy (&) � R. G. Van Driesche
Department of Environmental Conservation, University
of Massachusetts, Amherst, MA 01003, USA
e-mail: [email protected]
D. L. Wagner
Department of Ecology and Evolutionary Biology,
University of Connecticut, Storrs, CT 06269, USA
123
Biol Invasions
DOI 10.1007/s10530-014-0698-x

flower (Cardamine pratensis [L.] var. pratensis) in
wet meadows and along edges of rivers (Mike Nelson
unpubl. data). The introduced subspecies of cuckoo
flower is found throughout Massachusetts, while the
native form C. pratensis [L.] var. palustris is state
listed as threatened (USDA and CAPRP 2012). The
introduced plant is a biennial, flowers in spring, and
overwinters as a rosette. Foliage suitable for P.
oleracea larvae is present throughout the growing
season, allowing for four annual generations at Lenox.
Throughout the summer and fall, most of the available
cuckoo flower is overtopped by growth of other
floodplain plants. It is worth noting that although C.
pratensis is a novel host plant for P. oleracea in the
United States, it is a common host for all three sub-
species of P. napi in the United Kingdom (Asher et al.
2001).
Reasons for the original statewide decline of P.
oleracea in Massachusetts are uncertain. Evidence
supports the hypothesis that an introduced biological
control agent, Cotesia glomerata (L.) (Hymenoptera:
Braconidae), likely played an important role (Benson
et al. 2003), although other potentially contributing
factors have been suggested, including the possible
regional loss of a key forest host plant, Cardamine
diphylla [Michx.] A.W. Wood (two-leaf toothwort)
(Keeler et al. 2006). However, the continued abun-
dance of this plant throughout most western Massa-
chusetts did not prevent loss of P. oleracea in that
region (Benson et al. 2003). Historically, the decline
of P. oleracea began after the invasion of the pest
butterfly Pieris rapae (L.) (Lepidoptera: Pieridae) into
North America ca 1860 in Quebec and ca. 1869 in
Massachusetts (Scudder 1889). In response to this
invasion, in 1884 the European parasitoid C. glomer-
ata was introduced into the United States for biolog-
ical control of P. rapae (Clausen 1978), although the
same parasitoid may have co-invaded with P. rapae as
suggested by Scudder (1889). This parasitoid is not
specific to P. rapae, but rather readily attacks a range
of white butterflies (Pierinae) in several genera (Pieris,
Pontia, Tatachila, and Aporia) in different parts of the
world (Herrera Gonzalez 1982). C. glomerata estab-
lished and spread widely across North America and
became the dominant parasitoid of P. rapae in North
America (Van Driesche and Bellows 1988). In labo-
ratory tests, P. oleracea was a physiologically suitable
and preferred host (relative to P. rapae) for C.
glomerata (Van Driesche et al. 2003). Because of
the rarity of P. oleracea in Massachusetts, field attacks
of this host by C. glomerata were not directly
observed. However, attack on artificially deployed
trap host larvae of P. oleracea was observed in both
northern Vermont, where P. oleracea populations
persisted, and in western Massachusetts, where they
largely disappeared (Van Driesche et al. 2004). In
2001 in western Massachusetts, Van Driesche et al.
(2004) found that 100 % of 59 larvae recovered from
450 first or second instars of P. oleracea placed in a
meadow on hedge mustard, Sisymbrium officinale [L.]
Scop., were parasitized by C. glomerata during a
7-day exposure. Of particular relevance is that the
Shelburne, MA (N 42�320 by W 72�380) site used in
Van Driesche et al. (2004) is in the same region of
forested hills only 52 km NE from Lenox, Massachu-
setts, the site of our current study.
Parasitoid pressure from C. glomerata, however,
has not remained stable. Historically C. glomerata was
common and widespread in Massachusetts until
sometime between 2001 and 2007 (Van Driesche
et al. 2004; Van Driesche 2008) during which interval
another introduced parasitoid (Cotesia rubecula [Mar-
shall]) (Hymenoptera: Braconidae) outcompeted it for
dominance of P. rapae hosts (Van Driesche 2008), the
species that previously was the most common host of
C. glomerata in western Massachusetts (Van Driesche
1988). While several introductions of C. rubecula in
North America occurred between 1960 and 1992
(Puttler et al. 1970; Williamson 1971, 1972; Parker
and Pinnell 1972; McDonald and Kok 1992; Wold-
Burkness et al. 2005; Lee and Heimpel 2005), this
species’ presence in New England evidently resulted
from a 1988 release of wasps collected near Beijing,
China (Van Driesche and Nunn 2002). After release,
C. rubecula established and spread rapidly, and by
2007 it accounted for 99 % of the spring (Van
Driesche 2008) and (in 2009) 90 % of the fall
parasitism (Herlihy and Van Driesche 2013) of P.
rapae larvae collected from organic vegetable farms in
western Massachusetts. Because this species has a
narrower host range than C. glomerata (Brodeur et al.
1998; Van Driesche et al. 2003), it has rarely been
observed attacking species other than P. rapae in the
field (van Driesche et al. 2003, 2004). With this
background, we can construct a likely timeline for
both the presence of the P. oleracea population at
Lenox, MA, site (noted to occur there since at least
1986) and the region-wide displacement of C.
M. V. Herlihy et al.
123

glomerata by C. rubecula in western Massachusetts
(no earlier than 2001), leaving 1986–2001 as a period
when population increase of P. oleracea in Lenox on
cuckoo flower had to have occurred in the presence of
C. glomerata as a relatively abundant parasitoid,
which is a key point for understanding the history of
parasitoid-butterfly interactions at the study site.
The objectives of our current study were to describe
the current levels of parasitoid pressure from C.
glomerata at the Lenox site using trap–host experi-
ments and to determine what aspects of use of cuckoo
flower may have allowed for parasitoid escape by P.
oleracea historically (at least 1986–2001, likely
longer) when both C. glomerata and P. oleracea were
known to both be present at the site.
To estimate the current level of C. glomerata
pressure at the Lenox site, we conducted trap–host
exposures for 3 years (2008–10) in the wet meadow
and riverside areas where this P. oleracea population
still exists. Broadly, we found that at our study
location, as in the rest of western Massachusetts, C.
glomerata is now uncommon and there is currently
little-to-no parasitism of P. oleracea.
To meet our second objective and understand how
P. oleracea achieved population growth at the site pre-
2001, when C. glomerata was likely still common, we
used a combination of olfactometer and field-cage
experiments to determine what aspect of this Lenox
meadow habitat provided protection from parasitism.
We investigated three mechanisms that might have
created at least partially enemy-free space (Jeffries
and Lawton 1984): (1) that C. glomerata was unable to
detect or respond to volatiles from cuckoo flower
foliage infested by P. oleracea, plausible since
European cuckoo flower has a different glucosinolate
profile than either American cuckoo flower or brassica
crops like cabbage, collards, etc. (Agerbirk et al. 2010;
Agerbirk and Chew unpubl. data); (2) that the more
complex leaf architecture of the pinnate leaves of C.
pratensis may have lowered wasp foraging efficiency
as compared to the simpler leaves of crops such as
collards and cabbage (but not kale); or (3) that the
position of cuckoo flower rosettes, growing beneath
taller vegetation in summer and fall, may have reduced
the ability of C. glomerata to find infested cuckoo
flower under overtopping meadow plants (commonly
ca 0.6 m tall) as found by Sato and Ohsaki (1987) in
Japan, working with C. glomerata and the closely
related Pieris napi japonica Shirozu.
More broadly, there are at least two other hypotheses
about factors that might have affected population growth
of P. oleracea at the Lenox site that were not considered in
this study. First among these is the invasion of the region
by garlic mustard (Alliaria petiolata [M. Bieb.]) (Keeler
et al. 2006). While this species acts as a toxic egg trap for P.
oleracea (Keeler and Chew 2008), it is an abundant host to
which the butterfly may be adapting (Courant et al. 1994;
Keeler and Chew 2008; Chew et al. 2012). However, now
and in the critical past period (1986–2001) under study, the
overwhelming majority of garlic mustard plants were toxic
to most P. oleracea larvae. Garlic mustard is also found in
many areas in Massachusetts where P. oleracea popula-
tions did not persist. Therefore we discard this factor as
unimportant to the issue under study.
Second, the size of the resource patch formed by the
cuckoo flower population at the Lenox site is both large
and available for a very long period each year. This
allows P. oleracea butterflies to have four generations
per year at the site, which raises the butterfly’s effective
annual population rate of increase and helps it support
mortality from parasitism without a decrease in density.
We view this factor as likely important, but not one
readily tested experimentally. Consequently here we
focused on C. glomerata response to cuckoo flower
volatiles, the potential effect of leaf architecture, and the
effect of overtopping vegetation in reducing parasitism
and allowing for butterfly population growth.
Although much ecological research on exotic plants
has been focused on negative impacts on native species
through competition, exotic plants have other ecological
effects, including providing enemy free space to native
lepidopterans. In California, 82 butterflies (40 % of the
state’s species) have been shown to use exotic host
plants as food resources and to gain enemy free space
(Graves and Shapiro 2003; Shapiro 2002). We propose
that, despite parasitoid pressure due to C. glomerata and
range reduction of the host plant C. diphylla due to
invasion of garlic mustard, P. oleracea was able to
persist in Massachusetts by exploiting the exotic form of
C. pratensis as a host plant.
Materials and methods
Trap host experiments
Trap host studies were carried out in various sections
of the Lenox, Massachusetts site (Fig. 1) in the
Persistence in Massachusetts of the veined white butterfly
123

floodplain of the Housatonic River in 2008, 2009 and
2010 to detect parasitism by Cotesia using first and
second instar larvae of Pieris spp. butterflies. Similar
host exposures were also made in some additional
locations in 2009 and 2010 as positive controls of the
trap host technique. The floodplain sites were all on
property of the MA Department of Fish and Game and
were mowed yearly. Trap host larvae deployed in
early 2008 were placed in the field on naturally
occurring cuckoo flower plants. Starting with the third
experiment in 2008 and thereafter, trap host larvae
were placed, in the laboratory, on potted plants, which
were transported to field sites and placed out for
exposure to parasitoids. Exposures based on potted
plants were done either on previously dug field cuckoo
flower rosettes (a 100 cm2 area of plants), green-
house–grown collards (var. Georgia, 25 cm tall, 5–6
leaves), or field-dug broccoli transplants (25 cm tall,
5–6 leaves). After deployment in the field for the
desired number of days (3–4 in most cases, but up to
12 days in some experiments, see Table 1 for details),
remaining larvae were recovered and either reared on
host plants to the pupal stage (2008, some experiments
of 2009) or dissected (some experiments of 2009, all of
2010), either process being suitable for detection of
parasitism by Cotesia and both being procedures our
laboratory has carried out many other times. Details of
all nine trap host experiments (three in 2008, four in
2009, and two in 2010) are given in Table 1.
2008 experiments
In 2008, only one location (Lenox, MA, Fig. 1) was
used to experimentally expose larvae of P. oleracea to
measure field parasitism rates. The Lenox, Massachu-
setts site is located near the intersection of New Lenox
Rd. and the Housatonic River in Berkshire Co.
(42�230N; 73�140W). Two floodplain meadows at this
location were used to expose trap hosts in 2008 (A and
B, Fig. 1) in a series of two experiments (Table 1).
Cuckoo flower was abundant and in bloom in both
sites during the first experiments in 2008 in May. The
larvae used as trap hosts were all P. oleracea in 2008,
reared from eggs obtained from field-caught P.
oleracea adults from spring and summer flights at
the Lenox, Massachusetts site (under a MA Depart-
ment of Fish and Game, Natural Heritage Program
permit). In 2008, three separate trap–host exposures of
P. oleraceae were made (Exps. 1, 2, and 3), the first
two of which were done by placing larvae on field-
Fig. 1 Aerial photograph of the Lenox study site (42�230N; 73�140W), showing locations A, B, and C where trap host larvae were
exposed to detect Cotesia parasitism, as well as area D, an organic vegetable farm where trap host exposures were also made
M. V. Herlihy et al.
123
grown cuckoo flower plants and the third with larvae
on potted collards (an artificial but suitable host plant,
highly attractive to Cotesia parasitoids). Details of
each trap host exposure are summarized in Table 1.
2009 experiments
In 2009, four experiments were run, spanning a longer
period (May 24–October 2), with work at three
Table 1 History of trap host exposures (Pieris spp.) to detect Cotesia parasitism at the Lenox, Massachusetts, site where P. oleracea
has increased, as well as several sites used as positive controls (in bold), 2008–2010
Year Exp # Dates Place Plant Host #L/Pl Total# trap
hosts/#pla% Par Rb
(par/n)
% Par Gc
(par/n)
2008 1 May 17–29 Lenox area B CK Fd (in situ) Pieris oleracea 1L 97/91 n/a 0 (0/17)
2 May 23–29 Lenox area B CK F (in situ) Pieris oleracea 5–8L 35/6 n/a 0 (0/19)
3 July 18–24 Lenox areas A&B collards (potted) Pieris oleracea 5L 40e/8 n/a n/a
2009 1a May 24–June 2 Lenox areas A&B collards (potted,) P. rapae 5L 84/17 12.5 (1/12) 0 (0/12)
1b May 31–June 3 Lenox area B CK F (potted) Pieris oleracea 5L 120/24 n/a 0 (0/83f)
1c May 31–June 3 N’hamptonfComm. Garden CK F (potted) Pieris oleracea 5L 80/16 n/a 0 (0/34)
2a July 14–17 Lenox areas A&B CK F (potted) Pieris oleracea 4L 41/10 n/a 0 (0/4)i
2b July 14–17 Lenox areas A&B CK F (potted) Pieris rapae 4L 48/12 0 (0/9) 0 (0/9)i
3 Aug 13–16 Lenox area C CK F (potted) Pieris oleracea 3–9L 147/30 n/a 0 (0/75)
4a Sept 24–Oct 2g Lenox area C Broccoli (potted) Pieris oleracea 5L/11 55 n/a 0 (0/12)
4b Sept 24–Oct 2g Lenox area C Broccoli (potted) Pieris rapae 5L/11 55 0 (0/12) 0 (0/12)
4c Sept 24–Oct 2g Hadley farm area Broccoli (potted) Pieris oleracea 5L/10 50 n/a 22 (5/23)
4d Sept 24–Oct 2g Hadley farm area Broccoli (potted) Pieris rapae 5L/10 50j 53 (10/19) 11 (2/19)
2010 1 June 23–28 URI North Kingston Collards (potted) Pieris rapae 8L 32/8 0 (0/13) 0 (0/13)
Collards (potted) Pieris oleracea 8L 32/8 n/a 0 (0/7)
CK-F (potted) Pieris rapae 8L 32/8 0 (0/9) 0 (0/9)
CK-F (potted) Pieris oleracea 8L 32/8 n/a 0 (15)
June 23–28 Lenox area C (meadow) Collards (potted) Pieris rapae 8L 32/4 0 (0/20) 0 (0/20)
Collards (potted) Pieris oleracea 8L 32/4 n/a 0 (0/24)
CK-F (potted) Pieris rapae 8L 32/4 0 (0/7) 0 (0/7)
CK-F (potted) Pieris oleracea 8L 32/4 n/a 0 (0/22)
June 23–28 Lenox area D (organic farm) Collards (potted) Pieris rapae 8L 32/4 0 (0/20) 0 (0/20)
Collards (potted) Pieris oleracea 8L 32/4 n/a 0 (0/23)
CK-F (potted) Pieris rapae 8L 32/4 0 (0/17) 0 (0/17)
CK-F (potted) Pieris oleracea 8L 32/4 n/a 0 (0/16)
2 July 24–July 28 Lenox area D (organic farm) Collards (potted) Pieris rapae 6L 60/10 46 (13/28) 0 (0/28
Collards (potted) Pieris oleracea 6L 60/10 n/a 10 (3/31)
CK-F (potted) Pieris oleracea 6L 60/10 n/a 0 (0/23)
Lenox area C (meadow) Collards (potted) Pieris rapae 6L 60/10 0 (0/38) 0 (0/38)
Collards (potted) Pieris oleracea 6L 60/10 n/a 0 (0/32)
CK-F (potted) Pieris oleracea 6L 60/10 n/a 0 (0/36)
a The number of trap host larvae exposed and the number of plants on which they were dispersed
b R = Cotesia rubecula. This is a specific parasitoid of Pieris rapae and so its recovery is expected only when P. rapae this the trap host
c G = Cotesia glomerata. This parasitoid could be recovered from either P. rapae or P. oleracea
d CK F = cuckoo flower
e All lost to flood
f One of the 83 recovered larvae was parasitized by an unidentified nematode
g Hard frost occurred during this experiment, affecting quality of trap host plants
h Northampton Community Garden and Hadley farm area were two distantly located sites used as positive controls to demonstrate ability of method to detect C.
glomerata parasitism in areas where C. glomerata was present
i Most larvae lost due to heavy damage of plants by snails
j Two additional larvae of the 19 recovered were parasitized by unidentified tachinids
Persistence in Massachusetts of the veined white butterfly
123

locations: (1) New Lenox (areas A and B as in 2008
plus a new area, C); (2) the Northampton Community
Gardens, Northampton, Massachusetts (Prince Street,
42�180N; 72�390W); and (3) an organic vegetable farm
in Hadley, Massachusetts (42�200N; 72�360W). The
latter two locations were intended as positive controls
to demonstrate the efficacy of the trap host method.
Area C at the New Lenox site was a large wet meadow
(ca 2 ha) (Fig. 1) with a water table close to surface
such that small variations in the surface produced wet
or dry areas. Wet areas contained abundant cuckoo
flower, which by summer were overtopped with other
wetland plants. Butterflies were detected by us in this
meadow for the first time in 2009, but clearly were
present there earlier.
In 2009, in addition to P. oleracea, we also exposed
similar-sized larvae of P. rapae, a primary host of the
Cotesia parasitoids under study. For experiments, both
species of butterflies were collected in the wild (P.
oleracea from Lenox, MA and P. rapae from Hadley,
MA) and held over host plants outdoors for oviposition
(collards for P. rapae and cuckoo flower for P.
oleraceae). Leaf fragments with eggs were excised
and placed on moist filter paper in petri dishes to track
hatch. Hatching larvae were allowed several hours to
feed and then were held under chill (ca. 10 �C) until
deployed in field experiments. Details of the four
experiments are in Table 1.
2010 experiments
In 2010, the main experimental area was again area C
at the New Lenox site (the large wet meadow).
Experiments intended as positive controls were run in
two additional locations: (1) a cabbage plot on farm
land of the University of Rhode Island in N. Kingston,
Rhode Island (41�290N; 71�320W), selected because it
was the last site in the region known to us from
previous surveys where C. glomerata still dominated
and (2) River Valley Farm (area D in Fig. 1), an
organic vegetable farm close to our main research site
area C in New Lenox.
Pieris oleracea and P. rapae butterflies larvae used
in experiments in 2010 were reared from butterflies
sourced as in previous years and reared in a similar
manner. Collard plants used in experiments were
grown outdoors from Hart’s brand seed in 10 cm
square pots in garden soil, while cuckoo flower plants
were again dug from the New Lenox site. Plants
bearing trap host larvae were elevated ca 45 cm above
the soil on wooden stands (a pole topped with a plastic
container (20 cm w 9 20 cm l 9 8 cm h). The pole
was treated with Tanglefoot as a barrier to snails, a
pest discovered to be abundant at the site, and the
container in which the pot sat was filled with water to
sustain the plant.
Two experiments were run in 2010, each with four
treatments (two plants [collards and cuckoo flower]
crossed with larvae of two Pieris species (P. rapae and
P. oleracea). Exp. #1 (June 23–June 28) was run at
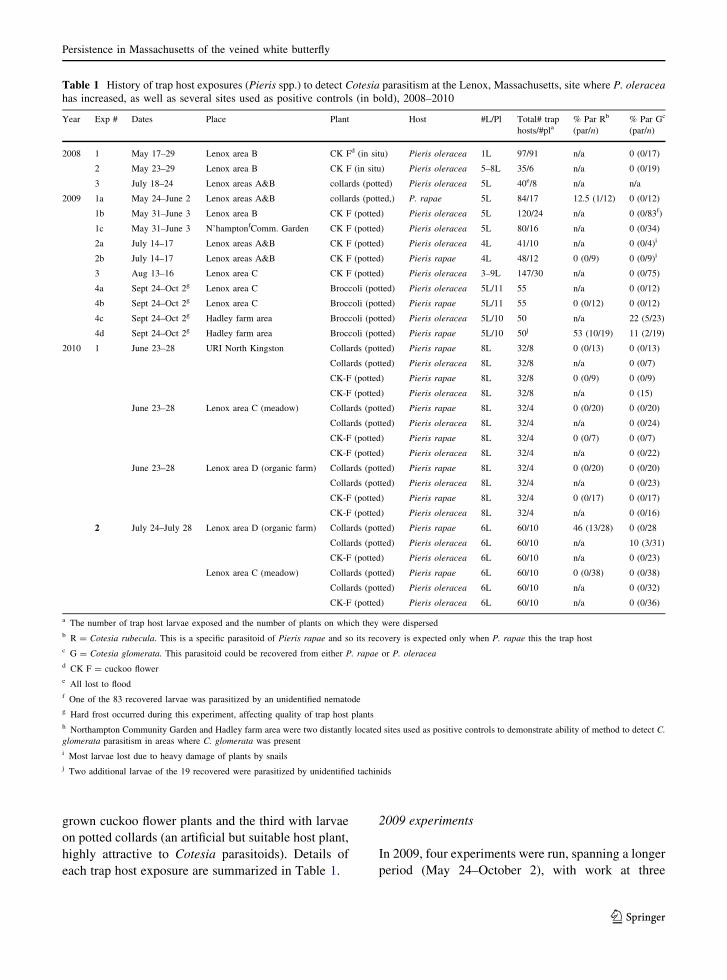
three locations (North Kingston RI and areas C and D
at Lenox) during the second flight of the P. oleracea
meadow population at the Lenox site (Fig. 2). At the
North Kingston Site, each treatment was replicated
eight times, with eight trap–host larvae per plant,
placing the 32 plants in stands as described above, in
an existing field of cabbage (free of insecticide
treatment). At the Lenox, MA site, 16 plants (four of
each treatment; each plant with eight larvae) were
placed in areas C (meadow) and D (River Valley farm)
(Fig. 1). In area C, plants were arranged in two
transects in the meadow and vegetation was cleared
from 60 cm circles around pots to increase visibility of
exposed larvae. At area D (the farm), plants were
arranged in a single row between a row of cultivated
kale plants and the forest edge, spacing pots about
3–4 m apart. Exp. #2 (July 24–28) was again run at
Lenox (in Areas C and D) during the third flight of the
meadow P. oleracea population (Fig. 2). The same
protocol was used as in Exp. 1 of 2010. Details of
protocol for both experiments are summarized in
Table 1.
Sources of plants and insects for olfactometer
and field-cage experiments
Cuckoo flower (C. pratensis) plants used in our
olfactometer experiments were dug at the study site
in Lenox, MA (42�2303700N; 73�1403300W) and placed
in 10 cm-dia pots prefilled with Pro-Mix BX mycorise
growing mix. Plants were kept in a greenhouse and
watered every 2 days until used for experimentation.
Collards (Brassica oleracea L. var. Blue max hybrid)
for experiments were purchased as 4-week-old plants
from Harvest Farm in Whately, MA. They were
transplanted into 10 cm-dia pots prefilled with Pro-
Mix BX mycorise growing mix. The plants were kept
in a greenhouse and watered every 2 days until used.
M. V. Herlihy et al.
123
Cotesia glomerata wasps used in experiments were
obtained from our laboratory colony, originally started
with cocoons collected in Massachusetts and supple-
mented with material from Virginia and Illinois
(because it became impossible to find adequate
numbers in Massachusetts). C. glomerata cocoon
masses were held at about 3 �C until needed for
experiments. Cocoons were then put into cages with
honey and sugar solution (‘‘Instant Hummingbird
Nectar,’’ Perky Pet Woodstream Corp. Lititz, PA), at
20 �C, natural light, and 50–65 % R. H. and left for
adult emergence. Once adults had emerged, wasps
were continually supplied with honey and a sugar
solution, but were not exposed to host larvae or host
plants. Naıve female wasps, exposed to males for
several days for mating, were used when they were
3–5 days old. After use in experiments, females were
returned to the rearing colony, where they were placed
into a separate rearing cage and exposed to first and
second instars of P. rapae or P. oleracea on fresh
collard leaves for oviposition. Parasitized P. rapae and
P. napi larvae were reared on collard leaves to produce
C. glomerata cocoon masses.
Pieris oleracea larvae used in our experiments were
obtained from our laboratory colony, initiated in 2010
with adult female butterflies collected at the Lenox,
Massachusetts site and supplemented by additional
butterflies from the Lenox, Massachusetts population
over a 3 year period (2010–2012) (under permit from
the State Natural Heritage Program). Butterflies were
reared through several generations in the laboratory in
the course of the summer of 2012 for olfactometer and
overtopping vegetation cage experiments. Pupae were
stored at about 3 �C until needed and were then moved
to Bug dorm cages (61 cm 9 61 cm 9 61 cm) (Bio-
Quip Products, Rancho Dominguez, CA, USA), where
emerging adults were provided with sugar solution
(‘‘Instant Hummingbird Nectar,’’ Perky Pet Wood-
stream Corp. Lititz, PA) and potted cuckoo flower
plants dug from Lenox, Massachusetts. Eggs laid on
plants were allowed to hatch and larvae to develop on
the cuckoo flower plants until larvae were used.
Excess larvae were returned to the P. oleracea colony
where they were transferred from cuckoo flower to
collard leaves, both being highly suitable host plants.
Olfactometer construction
A standard four-arm olfactometer was used for
experiments. A central chamber (55 mm in diame-
ter 9 100 mm in height) was connected to four glass
arms (50 mm in diameter 9 100 mm long), which
were each connected to bait chambers (100 mm in
diameter 9 200 mm long) containing an odor source.
Wire screens were placed between the openings of
each of the odor source chambers and the arm leading
to the central chamber, allowing air flow, but
preventing wasps from entering the odor source
chamber. The odor source chambers were connected
to a pressure control valve (allowing for air flow rate
through each arm to be equalized), and air was drawn
through a deionized water bubbler to control the
humidity and an activated charcoal filter to remove
unwanted volatiles. The central chamber was linked to
Fig. 2 Observed counts of
P. oleracea butterflies in
flight at the Lenox,
Massachusetts, meadow site
during the 2010 season
Persistence in Massachusetts of the veined white butterfly
123

a vacuum pump via 10 cm-dia plastic tubing. The
pump was set at 400 ml min-1, to pull air evenly
through all four arms, without affecting the wasps’
ability to fly through the arms. Light brown foam
boards (43 cm h) were placed on all four sides of the
olfactometer, which was placed on a laboratory table,
to prevent directional bias in lighting. Fluorescent
lights were positioned above the center of the olfac-
tometer to balance the light environment.
Experimental design for olfactometer tests
As a preliminary control to ensure that the physical set
up of the olfactometer was unbiased, tests were run
with four empty odor-source chambers in which a
single, 3- to 5-day-old, mated female wasp was
released into the central chamber, and whose position
was recorded after 30 min. Also, as a positive control
to ensure the wasps being tested were physiologically
prepared to respond to odors, in an additional exper-
iment, P. oleracea-infested collards (a host plant–
species combination known to be attractive to C.
glomerata) were tested against empty control arms,
with the two controls being placed 180� opposite each
other.
For our experiments, the test protocol was similar to
that of Karimzadeh et al. (2012). During each exper-
imental run, one naıve, 3–5-day-old, mated female C.
glomerata wasp was released into the central chamber.
Two arms (180� apart) contained the same bait (plant
or plant–host complex) and the other two were
unbaited controls, giving the wasp a choice between
either two plants (or two plant host complexes) and
two controls. We tested (1) uninfested collards vs
uninfested cuckoo flower and (2) P. oleracea-infested
collards versus P. oleracea-infested cuckoo flower.
For tests with uninfested plants, collard leaves were
cut at the end of the petiole nearest the leaf and
weighed; cuckoo flower leaves were then cut along the
petiole to match the weight of the collard leaves. This
was done because cuckoo flower leaves are pinnate
and thus easier than collards to downsize to a desired
weight without cutting the actual leaf blade (which
would increase the release of volatiles attractive to
parasitoids). Uninfested leaves were placed in test
chambers 10 min before each experiment. For exper-
imental runs with infested leaves, 10 second instar P.
oleracea were placed on each collard leaf, or group of
cuckoo flower leaves. All leaves were weighed and
foliar weight standardized between treatments. Foli-
age with larvae was placed in the odor source
chambers and larvae were allowed to feed for 1 h in
each experimental run to allow time for plants to
respond to feeding by releasing both constituent and
induced compounds. Tests ran until wasps made a
decision and entered an arm. If wasps were unrespon-
sive, a run was terminated after 30 min. Wasps
entering and remaining in an olfactometer arm beyond
a line 4 cm into the arm were counted as having made
a decision and those remaining in the central chamber
were considered unresponsive. Wasps that reversed
direction within an arm were counted as unresponsive.
The position of odor sources was rotated 90� after each
run (=the trial of one wasp) by disconnecting the odor
source chambers and reconnecting them to a new arm.
After each group of replicates run on a given day (ca
15–20 per day), the odor source chambers were
removed and washed in an odorless detergent and
rinsed with water and allowed to air-dry overnight. In
total, there was a minimum of 30 replicates per
treatment (each being the trial of one wasp).
Overtopping vegetation field cage experiment
To determine if overtopping by other vegetation
affected the rate of host detection by C. glomerata,
field experiments were run on the University of
Massachusetts Amherst campus in a meadow using
open-bottom, cube cages (0.6 m3) with black mesh
fabric (EQUINOX No-See-Um Netting Eastern
Mountain Sports) sides. In each run (one replicate)
of the experiment, we placed four such cages at the
field site. In total, there were eight replicates, run from
June to August 2012, each replicate on a different day.
Each cage was a single replicate of one of four
treatments being tested: (1) collards and clipped native
vegetation, (2) collards and unclipped vegetation, (3)
cuckoo flower and clipped native vegetation, and (4)
cuckoo flower and unclipped vegetation. Test plants of
each species were 10 cm tall. Clipped vegetation was
cut with scissors within 2–5 cm of the soil. Unclipped
vegetation was ca 35 cm tall. There were two test
plants per cage and five first instars of P. oleracea
were placed on each plant (=10 larvae per cage). Then
two 3 to 5-day-old C. glomerata females, which had
been given the opportunity to mate but had no previous
exposure to C. pratensis or B. oleracea volatiles, were
released into each cage. The larvae were exposed to
M. V. Herlihy et al.
123

parasitoids for 24 h. The test was then terminated and
the larvae removed, refrigerated to arrest develop-
ment, and then dissected within 48 h to detect
parasitoid oviposition.
Statistical analyses
All statistical analyses were conducted using the
program R (version 2.13.1). Binomial exact tests were
used to compare female wasp choices in the olfac-
tometer. Cage experiment percent parasitism data
were arcsine transformed in order to meet the
assumption of normality and then analyzed with
ANOVA, followed by a Tukey HSD test.
Results
Trap host recoveries and parasitism, 2008–2010
Over the 3 years of field deployment of P. oleracea
larvae as trap hosts, 903 larvae were deployed (172,
363, and 368 in 2008, 2009, and 2010, respectively)
and 417 (46 %) were recovered after field exposure
(Table 1). Rates of recovery of trap host larvae
improved (21, 48, and 56 % in 2008, 2009, and
2010) as problems from flooding and herbivory by
snails were resolved by use of elevated stands
protected with Tanglefoot). Of the 417 P. oleracea
larvae recovered after field exposure, only 3 (0.7 %)
were parasitized by C. glomerata and none by C.
rubecula. The only detection at the Lenox site of C.
glomerata parasitism occurred at area D, an organic
vegetable farm (Fig 1) close to area C, the meadow
where the largest P. oleracea numbers occurred. P.
rapae butterflies occurred at all of the Lenox research
areas (A, B, C, D) but were most common at the farm
(area D).
Trap host exposures of P. rapae larvae at the Lenox
site were also made because we knew this was a highly
attractive host for both C. glomerata and C. rubecula
(and thus a good probe for both parasitoids). In total
435 P. rapae larvae were placed on trap host plants
(2008–2010) and 163 were recovered. Of these, none
were parasitized by C. glomerata, while 14 (9 %) were
attacked by C. rubecula. Again, all parasitism detected
for larvae of this butterfly was in area D, the organic
vegetable farm described above.
While our trap host method has been used previ-
ously on an extensive basis to detect Cotesia attack on
Pieris species (Benson et al. 2003; Van Driesche et al.
2004) as part of this study, we set out trap hosts in three
areas where we believed C. glomerata populations
might still remain despite its largely having been
suppressed in the region by competition from C.
rubecula (Van Driesche 2008; Herlihy and Van
Driesche 2013). Of these sites (Northampton, MA;
Hadley, MA; and N. Kingston, Rhode Island), Cotesia
parasitism was detected only at the Hadley site (a farm
field planted to organic cole crops). At that site, we
observed 22 % (5/23) parasitism by C. glomerata of P.
oleracea larvae. At the same site and dates, P. rapae
trap host larvae suffered 53 % (10/19) parasitism by C.
rubecula and 11 % by C. glomerata. As a positive
control, these data showed that the trap host system
functioned and was able to detect Cotesia parasitism.
It also demonstrated that P. oleracea is an attractive
host, at least on collards, for C. glomerata.
Response of C. glomerata to host and plant
volatiles
As a preliminary assessment to detect potential
directional bias in the olfactometer, wasp response
(n = 32) to four unbaited arms was assessed and no
significant difference among arms was detected
(X2 = 0.25, df = 3, P = 0.969) (Fig. 3). As a posi-
tive control to demonstrate that the test parasitoids
were capable of responding as expected to a known
high quality host/plant combination, female wasps
were given the choice of collard foliage infested with
P. oleracea larvae versus empty (unbaited) arms, and
in this test 23 of 25 parasitoids chose infested collards
over empty arms (P = 0.56 in two-tailed binomial
exact test with significance set at 0.05), a highly
significant difference.
In the first experiment, C. glomerata females were
offered uninfested collards vs uninfested cuckoo
flower plants and both were found to be equally
attractive; of 105 responsive females, 59 chose
infested collards and 46 chose infested cuckoo flower,
with no significant difference in a two-tailed binomial
exact test at the 0.05 level (P = 0. 24) (Fig. 3). In the
final test, C. glomerata females were offered a choice
of P. oleracea-infested cuckoo flower plants versus P.
oleracea-infested collards and of 58 responsive
Persistence in Massachusetts of the veined white butterfly
123
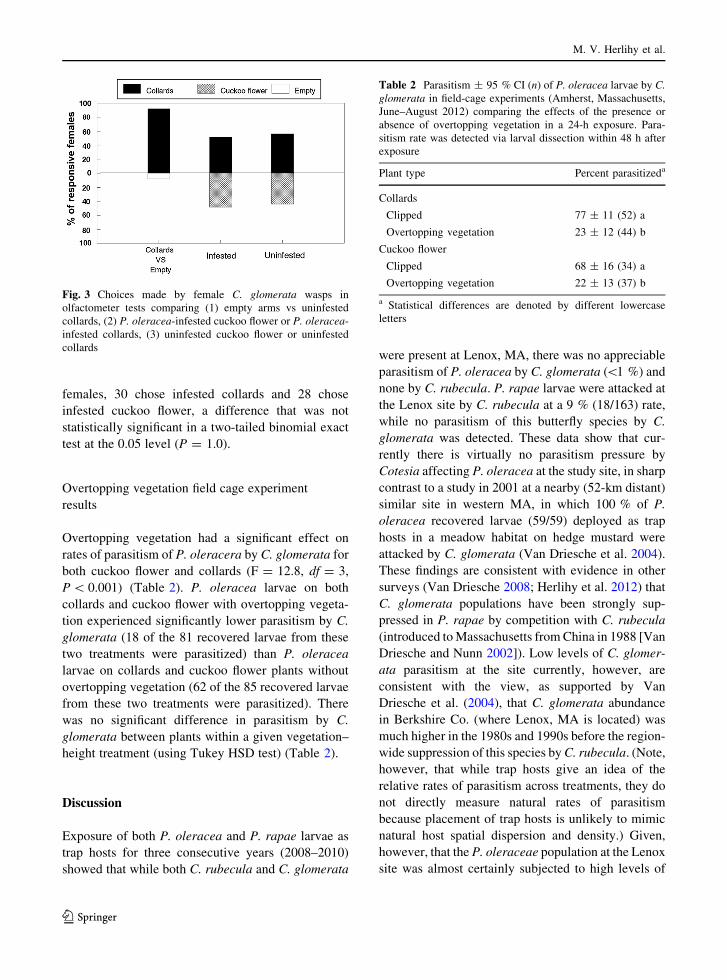
females, 30 chose infested collards and 28 chose
infested cuckoo flower, a difference that was not
statistically significant in a two-tailed binomial exact
test at the 0.05 level (P = 1.0).
Overtopping vegetation field cage experiment
results
Overtopping vegetation had a significant effect on
rates of parasitism of P. oleracera by C. glomerata for
both cuckoo flower and collards (F = 12.8, df = 3,
P \ 0.001) (Table 2). P. oleracea larvae on both
collards and cuckoo flower with overtopping vegeta-
tion experienced significantly lower parasitism by C.
glomerata (18 of the 81 recovered larvae from these
two treatments were parasitized) than P. oleracea
larvae on collards and cuckoo flower plants without
overtopping vegetation (62 of the 85 recovered larvae
from these two treatments were parasitized). There
was no significant difference in parasitism by C.
glomerata between plants within a given vegetation–
height treatment (using Tukey HSD test) (Table 2).
Discussion
Exposure of both P. oleracea and P. rapae larvae as
trap hosts for three consecutive years (2008–2010)
showed that while both C. rubecula and C. glomerata
were present at Lenox, MA, there was no appreciable
parasitism of P. oleracea by C. glomerata (\1 %) and
none by C. rubecula. P. rapae larvae were attacked at
the Lenox site by C. rubecula at a 9 % (18/163) rate,
while no parasitism of this butterfly species by C.
glomerata was detected. These data show that cur-
rently there is virtually no parasitism pressure by
Cotesia affecting P. oleracea at the study site, in sharp
contrast to a study in 2001 at a nearby (52-km distant)
similar site in western MA, in which 100 % of P.
oleracea recovered larvae (59/59) deployed as trap
hosts in a meadow habitat on hedge mustard were
attacked by C. glomerata (Van Driesche et al. 2004).
These findings are consistent with evidence in other
surveys (Van Driesche 2008; Herlihy et al. 2012) that
C. glomerata populations have been strongly sup-
pressed in P. rapae by competition with C. rubecula
(introduced to Massachusetts from China in 1988 [Van
Driesche and Nunn 2002]). Low levels of C. glomer-
ata parasitism at the site currently, however, are
consistent with the view, as supported by Van
Driesche et al. (2004), that C. glomerata abundance
in Berkshire Co. (where Lenox, MA is located) was
much higher in the 1980s and 1990s before the region-
wide suppression of this species by C. rubecula. (Note,
however, that while trap hosts give an idea of the
relative rates of parasitism across treatments, they do
not directly measure natural rates of parasitism
because placement of trap hosts is unlikely to mimic
natural host spatial dispersion and density.) Given,
however, that the P. oleraceae population at the Lenox
site was almost certainly subjected to high levels of
Fig. 3 Choices made by female C. glomerata wasps in
olfactometer tests comparing (1) empty arms vs uninfested
collards, (2) P. oleracea-infested cuckoo flower or P. oleracea-
infested collards, (3) uninfested cuckoo flower or uninfested
collards
Table 2 Parasitism ± 95 % CI (n) of P. oleracea larvae by C.
glomerata in field-cage experiments (Amherst, Massachusetts,
June–August 2012) comparing the effects of the presence or
absence of overtopping vegetation in a 24-h exposure. Para-
sitism rate was detected via larval dissection within 48 h after
exposure
Plant type Percent parasitizeda
Collards
Clipped 77 ± 11 (52) a
Overtopping vegetation 23 ± 12 (44) b
Cuckoo flower
Clipped 68 ± 16 (34) a
Overtopping vegetation 22 ± 13 (37) b
a Statistical differences are denoted by different lowercase
letters
M. V. Herlihy et al.
123

parasitism for several decades or more in the past, it is
of interest to know how the butterfly population could
have evaded parasitism or compensated for it when
feeding on the invasive cuckoo flower stands at the
study site.
It is known that C. glomerata can detect hosts from
long distances via olfactory signals from the plant–
host complex, using volatiles released by the plant
(Steinberg et al. 1993). C. glomerata can also detect
densities of hosts on a host plant and the age of the
plant, allowing it to find and exploit plants with the
highest number of host larvae on younger host plants
(Mattiacci and Dicke 1995; Geervliet et al. 1998). The
goal of one of our experiments was to determine
whether these olfactory signals from the plant–host
complex were being exploited by C. glomerata on C.
pratensis plants. Some plants seem to be exploited by
herbivorous insects because they offer some degree of
enemy-free space because their parasitoids do not
detect or recognize volatiles from such plants when
they are infested. For example, the Japanese subspe-
cies Pieris napi japonica is thought to use a nutrition-
ally inferior host plant (Arabis sp.) in order to avoid
parasitism by C. glomerata (Ohsaki and Sato 1990).
However, in our case, we found that C. glomerata did
not distinguish between volatiles from C. pratensis
and B. oleracea, either from uninfested plants or ones
infested by P. oleracea (Table 2), suggesting that in
the field, C. glomerata should be able detect volatiles
produced when P. oleracea larvae feed on C.
pratensis.
Other factors able to modify parasitism risk in the
field, apart from plant chemistry, are features affecting
plant apparency and foliar complexity (Sato and
Ohsaki 1987; Meiners and Obermaier 2004; Oberma-
ier et al. 2008). In our field cage experiment, we found
that in cages without overtopping vegetation, C.
glomerata wasps were able to easily locate, and
parasitize P. oleracea larvae on C. pratensis, with no
difference in attack rates from that for larvae on
collards (B. oleracea), showing that the more struc-
turally complex pinnate foliage of C. pratensis did not
reduce the ability of the wasp to locate and parasitize
larvae. However, we did find that overtopping vege-
tation greatly decreased parasitism of P. oleracea, on
both C. pratensis and B. oleracea, reducing parasitism
to about one-third of our controls without overtopping
vegetation–which suggests that overtopping vegeta-
tion provided protection from C. glomerata
parasitism. Whether this was from physical complex-
ity slowing or impeding wasp movement or from
effects of vegetation on odor-plume detection or wasp
movement toward odors is unknown.
In summary, the adoption of a novel host, i.e.,
cuckoo flower, by P. oleracea at our Lenox, Massa-
chusetts site evidently conferred some degree of
enemy-free space from C. glomerata by virtue of the
host plant rosettes being obscured most of the year by
taller vegetation, despite the odor signature of infested
cuckoo flower being highly attractive to the parasitoid.
Also, cuckoo flower was an extremely abundant
resource that was available for the whole growing
season, allowing four generations of the butterfly per
year. Multiple generations and an abundant host plant
likely enhanced the population growth rate and hence
density of the butterfly, which in turn may have
allowed the population to tolerate significant levels of
parasitism.
Acknowledgments The authors thank Richard Casagrande of
the University of Rhode Island, George Boettner and Stephen
Donahue of the University of Massachusetts Amherst, Patrick
DeFlorio of Yankee Glassblower, Jian Duan and Craig Oppel of
USDA ARS Newark, DE, Lisa Dachinger of River Valley Farm,
Lenox, Massachusetts, and Emmet Van Driesche. For help with
the rearing of insects and conduct of sentinel larval studies at
Lenox in 2008 and 2009, we thank Alex Meleg, Sabina Perkins,
April Rodd, and Ryan Wagner. This material is based upon work
supported by the National Institute of food and Agriculture, U.S.
Department of Agriculture, the Massachusetts Agricultural
Experiment Station and the Department of Plant, Soil and
Insect Sciences under Project number MAS00957.
References
Agerbirk N, Olsen CE, Chew FS, Orgaard M (2010) Variable
glucosinolate profiles of Cardamine pratensis (Brassi-
caceae) with equal chromosome numbers. J Agric Food
Chem 58:4693–4700
Asher J, Warren M, Fox R, Harding P, Jeffcoate G, Jeffcoate S
(2001) Millennium atlas of butterflies in Britain and Ire-
land. Oxford University Press, New York
Benson J, van Driesche RG, Pasquale A, Elkinton J (2003)
Introduced braconid parasitoids and range reduction of a
native butterfly in New England. Biol Cont 28:197–213
Biever KD (1992) Distribution and occurrence of Cotesia
rubecula (Hymenoptera: Braconidae), a parasite of Arto-
geia rapae in Washington and Oregon. J Econ Entomol
85:739–742
Brodeur J, Geervliet JBF, VET, LEM (1998) Effects of Pieris
host species on life history parameters in a solitary spe-
cialist and gregarious generalist parasitoid (Cotesia spe-
cies). Entomol Exp Appl 86:145–152
Persistence in Massachusetts of the veined white butterfly
123

Chew FS, van Driesche RG, Casagrande RA (2012) A native
butterfly confronts exotic plants and parasitoids. Mass
Butterflies 39:2–6
Clausen CP (1978) Introduced parasites and predators of
arthropod pests and weeds: a world review. USDA Agri-
culture Handbook 480, Washington
Corrigan JE (1982) Cotesia (Apanteles) rubecula [Hymenop-
tera: Braconidae] recovered in Ottawa, Ontario ten years
after its release. Proc Entomol Soc Ontario 113:71
Courant AV, Holbrook AE, van der Reijden ED, Chew FS
(1994) Native pierine butterfly (Pieridae) adapting to nat-
uralized crucifer? J Lepid Soc 48:168–170
Geervliet JBF, Ariens S, Dicke M, Vet LEM (1998) Long-distance
assessment of patch profitability through volatile infochem-
icals by the parasitoids Cotesia glomerata and C. rubecula
(Hymenoptera: Braconidae). Biol Cont 11:113–121
Graves SD, Shapiro AM (2003) Exotics as hosts of the Cali-
fornia butterfly fauna. Biol Conserv 110:413–433
Herlihy MV, van Driesche RG (2013) Effect of Cotesia rube-
cula (Hymenoptera: Braconidae) on survival of larval
cohorts of Pieris rapae (Lepidoptera: Pieridae) on collards:
evaluation of an introduced biological control sgent. Fla
Entomol 96:360–369
Herlihy MV, van Driesche RG, Abney MR, Brodeur J, Bryant
AB, Casagrande RA, Delaney DA, Elkner TE, Fleischer
SL, Groves RL, Gruner DS, Harmon JP, Heimpel GE,
Hemady K, Kuhar TP, Maund CM, Olmstead RD, Seaman
AM, Skinner M, Weinzierl R, Yeargan KV, Szendrei Z
(2012) Occurrence of Cotesia rubecula (Hymenoptera:
Braconidae) and its displacement of Cotesia glomerata
(Hymenoptera: Braconidae) in eastern North America. Fla
Entomol 95:458–464
Herrera Gonzalez J (1982) La vida silvestre: >Se extinguen las
mariposas en Chile? Santiago. Academia Superior de
Ciencias Pedagogicas de Santiago, Chile
Jeffries MJ, Lawton JH (1984) Enemy free space and the structure
of ecological communities. Biol J Linn Soc 23:269–286
Karimzadeh J, Hardie J, Wright DJ (2012) Plant resistance
affects the olfactory response and parasitism success of
Cotesia vestalis. J Insect Behav 25, doi 10.1007/s10905-
012-9331-y
Keeler MS, Chew FS (2008) Escaping an evolutionary trap:
preference and performance of a native insect on an exotic
invasive host. Oceologia 156:559–568
Keeler MS, Chew FS, Goodale BC, Reed JM (2006) Modeling
the impacts of two exotic invasive species on a native
butterfly: top-down vs. bottom-up effects. J Animal Ecol
75:777–788
Lee JC, Heimpel GE (2005) Impact of flowering buckwheat on
lepidopteran cabbage pests and their parasitoids at two
spatial scales. Biol Control 34:290–301
Mattiacci L, Dicke M (1995) Host-age discrimination during
host location by Cotesia glomerata, a larval parasitoid of
Pieris brassicae. Entomol Exp Appl 76:37–48
McDonald RC, Kok LT (1992) Colonization and hyperparasit-
ism of Cotesia rubecula (Hymenoptera: Braconidae), a
newly introduced parasite of Pieris rapae, in Virginia.
Entomophaga 37:223–228
Meiners T, Obermaier E (2004) Hide and seek on two spatial
scales—vegetation structure affects herbivore oviposition
and egg parasitism. Basic Appl Ecol 5:87–94
Obermaier E, Heisswolf A, Poethke HJ, Randlkofer B, Meiners
T (2008) Plant architecture and vegetation structure: two
ways for insect herbivores to escape parasitism. Eur J
Entomol 105:233–240
Ohsaki N, Sato Y (1990) Avoidance mechanisms of three Pieris
butterfly species against the parasitoid wasp Apantales
glomeratus. Ecol Entomol 15:169–176
Parker FD, Pinnell RE (1972) Further studies of the biological
control of Pieris rapae using supplemental host and para-
site releases. Environ Entomol 1:150–157
Puttler B, Parker FD, Pinnell RE, Thewke SE (1970) Introduction
of Apanteles rubecula Marshall and other parasites of Pieris
rapae in British Columbia. J Econ Entomol 63:304–305
Sato Y, Ohsaki N (1987) Host-habitat location by Apanteles
glomeratus and effect of food-plant exposure on host-
parasitism. Ecol Entomol 12:291–297
Scudder SH (1889) The butterflies of the Eastern United States
and Canada. Vol. 1. Pub. by author, Cambridge
Shapiro AM (2002) The California urban butterfly fauna is
dependent on alien plants. Divers Distrib 8:31–40
Steinberg S, Dicke M, Vet LEM (1993) Relative Importance of
infochemicals from first and second trophic level in long-
range host location by the larval parasitoid Cotesia glom-
erata. J Chem Ecol 19:47–59
USDA, CAPRP (2012) http://plants.usda.gov/java/
profile?symbol=CAPRP
van Driesche RG (1988) Survivorship patterns of larvae of
Pieris rapae (L.) (Lepidoptera: Pieridae) in Massachusetts
kale, with special reference to mortality due to Apanteles
glomeratus L. (Hymenoptera: Braconidae). Bull Entomol
Res 78:397–405
van Driesche RG (2008) Biological control of Pieris rapae in
New England: Host suppression and displacement of Co-
tesia glomerata by Cotesia rubecula (Hymenoptera: Bra-
conidae). Fla Entomol 91:22–25
van Driesche RG, Bellows TS (1988) Host and parasitoid
recruitment for quantifying losses from parasitism, with
reference to Pieris rapae and Cotesia glomerata. Ecol
Entomol 13:215–222
van Driesche RG, Nunn C (2002) Establishment of a Chinese
strain of Cotesia rubecula (Hymenoptera: Braconidae) in
the northeastern United States. Fla Entomol 85:386–388
van Driesche RG, Nunn C, Kreke N, Goldstein B, Benson J
(2003) Laboratory and field host preferences of introduced
Cotesia spp. parasitoids (Hymenoptera: Braconidae)
between native and invasive Pieris butterflies. Biol Control
28:214–221
van Driesche RG, Nunn C, Pasqual A (2004) Life history pat-
tern, host plants, and habitat determinants of population
survival of Pieris napi oleracea interacting with an intro-
duced braconid parasitoid. Biol Control 29:278–287
Williamson GD (1971) Insect liberation in Canada. Parasites and
predators 1970. Ag Canada (Liberation Bulletin) No. 34
Williamson GD (1972) Insect liberation in Canada. Parasites and
predators 1971. Ag Canada (Liberation Bulletin) No. 35
Wold-Burkness SJ, Hutchison WD, Lee JC, Hines RL, Bolin
PC, Heimpel GE (2005) A long-term survey of parasitoid
species composition and parasitism of Trichoplusia ni
(Lepidoptera: Noctuidae), Plutella xylostella (Lepidoptera
: Plutellidae), and Pieris rapae (Lepidoptera: Pieridae) in
Minnesota cabbage. J Entomol Sci 40:211–221
M. V. Herlihy et al.
123