pharmacology, biochemistry and behavior · was measured 30s later and then at 5-min intervals for...
TRANSCRIPT

Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
Contents lists available at SciVerse ScienceDirect
Pharmacology, Biochemistry and Behavior
j ourna l homepage: www.e lsev ie r .com/ locate /pharmbiochembeh
Antinociceptive effect of stimulating the zona incerta with glutamate in rats
Ariane Petronilho, Gláucia M. Reis, Quintino M. Dias, Rafael S. Fais, Wiliam A. Prado ⁎Department of Pharmacology, Faculty of Medicine of Ribeirão Preto, University of São Paulo, Ribeirão Preto, SP 14049-900, Brazil
⁎ Corresponding author at: Department of PharmacRibeirão Preto, University of São Paulo, Av. Bandeira14049-900, Brazil. Tel.: +55 16 36023038; fax: +55 16
E-mail address: [email protected] (W.A. Prado
0091-3057 © 2012 Elsevier Inc.doi:10.1016/j.pbb.2012.01.022
Open access under the Else
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 June 2011Received in revised form 23 January 2012Accepted 25 January 2012Available online 1 February 2012
Keywords:Zona incertaPain controlDorsolateral funiculusAntinociceptionGlutamate
The zona incerta (ZI) is a subthalamic nucleus connected to several structures, some of them known to be in-volved with antinociception. The ZI itself may be involved with both antinociception and nociception. The anti-nociceptive effects of stimulating the ZIwith glutamate using the rat tail-flick test and a ratmodel of incision painwere examined. The effects of intraperitoneal antagonists of acetylcholine, noradrenaline, serotonin, dopamine,or opioids on glutamate-induced antinociception from the ZI in the tail-flick test were also evaluated. The injec-tion of glutamate (7 μg/0.25 μl) into the ZI increased tail-flick latency and inhibited post-incision pain, but did notchange the animal performance in a Rota-rod test. The injection of glutamate into sites near the ZI was noneffective. The glutamate-induced antinociception from the ZI did not occur in animals with bilateral lesion ofthe dorsolateral funiculus, or in rats treated intraperitoneally with naloxone (1 and 2 m/kg), methysergide (1 and2 m/kg) or phenoxybenzamine (2 m/kg), but remained unchanged in rats treated with atropine, mecamylamine,or haloperidol (all given at doses of 1 and 2 m/kg). We conclude that the antinociceptive effect evoked from theZI is not due to a reduced motor performance, is likely to result from the activation of a pain-inhibitory mechanismthat descends to the spinal cord via the dorsolateral funiculus, and involves at least opioid, serotonergic andα-adrenergic mechanisms. This profile resembles the reported effects of these antagonists on the antinoci-ception caused by stimulating the periaqueductal gray or the pedunculopontine tegmental nucleus.
© 2012 Elsevier Inc. Open access under the Elsevier OA license.
1. Introduction
The zona incerta (ZI) is a subthalamic nucleus located ventrolateralto the medial lemniscus and dorsomedial to the substantia nigra(Ricardo, 1981). Almost all major ascending and descending fiber tractscross the ZI and target numerous brain centers from the thalamus to thespinal cord (Mitrofanis, 2005). The ZI seems to be involved with differ-ent brain functions, such as intake of liquids and food (Gonzalez-Limaet al., 1993), locomotion (Mogenson et al., 1985), and sexual behavior(Edwards and Isaacs, 1991). The ZImay also act as a diencephalic centerthat initiates and controls responses to a somatic or visceral stimulusand influences arousal and attention as well (Mitrofanis, 2005).
There are evidences for the influence of the ZI on nociception andantinociception: some neurons in the ZI respond to peripheral noxiousstimulation (Yen et al., 1989; Okada et al., 2002), and their metabolicactivity rises significantly during both phases of the response to intra-plantar formalin (Porro et al., 2003); electrical stimulation of the ZIproduces antinociception in the rat tail-flick test, but strong aversionwas observed during the stimulation period (Prado and Roberts, 1985).
The ZI interconnects with various brainstem nuclei, including theperiaqueductal gray, pedunculopontine tegmental nucleus (PPTg)
ology, Faculty of Medicine ofntes 3900, Ribeirão Preto, SP36332301.).
vier OA license.
(Ricardo, 1981; Roger and Cadusseau, 1985; Shammah-Lagnado et al.,1985; Beitz, 1989; Heise and Mitrofanis, 2004), the nucleus raphemagnus (NRM) (Erzumulu and Killackey, 1980; Ricardo, 1981; Senbaet al., 1981; Peschanski, 1984; Desbois and Villanueva, 2001), giganto-cellularis nucleus (Giα) (Watanabe and Kawana, 1982; Carlton et al.,1983) and the anterior pretectal nucleus (APtN) (Foster et al., 1989;Terenzi et al., 1995; Giber et al., 2008), structures from which electricalstimulation evokes antinociception (Rees and Roberts, 1993; Millan,1999).
Stimulation-produced analgesia (SPA) from several brain structuresinvolves the activation of descending pathways that act to inhibit nox-ious inputs in the spinal dorsal horn (Millan, 1999). SPA from theNRM is mediated by serotonergic and opioid mechanisms (Fields andBasbaum, 1978), while SPA from Giα is mediated by neurons that pro-ject to and activate spinally projecting noradrenergic neurons located inthe A7 cell group (Clark and Proudfit, 1991). Endogenous opioids, sero-tonin and noradrenaline are involved in mechanisms activated duringSPA from PAG (Millan, 2002), whereas SPA from the APtN is mediatedby opioid and α-adrenergic mechanisms (Rees et al., 1987).
The present study reassessed the effects of stimulating the ZI in therat tail-flick test. Intrinsic neurons and many crossing nerve fibers havebeen described in the ZI (Masri et al., 2009), and thus stimulation ofthe ZI was performed using local injection of glutamate, which isknown to activate cells, but not nerve fibers (Fries and Zieglgansberger,1974). The effects of stimulating the ZI against post-incision pain inrats were assessed as well. The effects of intraperitoneal antagonists

361A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
on glutamate-induced antinociception from the ZI were also evaluat-ed to determine whether antagonists of acetylcholine, noradrena-line, serotonin, dopamine, or opioids are effective, as they areknown to be against SPA from PAG, NRM, Giα, or APtN.
2. Materials and methods
2.1. Animals and surgery
Experiments were carried out using male Wistar rats (150–160 g),provided by the Animal House of the University of São Paulo (USP;Campus of Ribeirão Preto). These animals were housed in groups of fiveper cage, and given free access to food and water throughout the experi-ments. The room temperature was controlled (22±1 °C), and a light–dark cycle (07:00–19:00 h lights on) was maintained. The experimentswere approved by the Commission of Ethics in Animal Research, Facultyof Medicine of Ribeirão Preto, University of São Paulo (Process number012/2006). The guidelines of the Committee for Research and EthicalIssues of IASP (Zimmermann, 1983) were followed throughout theexperiments. Each rat was used on only one occasion. Each animal wasanesthetizedwith tribromoethanol (250 mg/kg, i.p.) and a 12-mm lengthof a 23-gauge stainless steel guide cannula was stereotaxically implantedinto the skull until its tip lays 3 mmabove the right ZI. The coordinates (inmm) were AP=+4.1(from the interaural line), L=−2.3 (from thesagittal suture), andH=−3.3 (from the skull surface). The guide cannulawas fixed to the skull with two steel screws and dental cement, and keptpatent with a sterile obturator until the time of drug administration. Agroup of rats taken for the evaluation of incision pain was contralateralor bilaterally implanted with guide cannulae in the ZI. After receivingpenicillin (50 mg/kg, i.m.), the animal was allowed to recover for atleast 1 week before the start of the experiments.
2.2. Microinjection procedures
Drug or saline was microinjected into the ZI using a glass needle(70–80 μm) protected by a system of telescoping steel tubes as de-scribed elsewhere (Azami et al., 1980). The assembly was insertedinto the guide cannula immediately before microinjection and the nee-dle advanced to protrude 3 mmbeyond the guide cannula tip. The injec-tion of 2% lignocaine (0.25 μl) into the dorsal or ventral portions of theanterior pretectal nucleus significantly increased the incision pain inrats, but the effects from the different parts of the nucleus did not differ(Villarreal et al., 2004). Therefore, the volume of solution was certainlydiffusing throughout the entire nucleus. The area of the ZI (about3 mm2) is a bit larger than the area of the anterior pretectal nucleus(about 2 mm2). By this reason, the volume of 0.25 μl was chosen foreach injection into the ZI. This volume was delivered at a constantrate over a period of 2 min, and the needle was removed 20 s after thecompletion of this procedure.
2.3. Tail-flick test
Each animalwas lightly restrained by handwith the tail laid across awire coil at room temperature (23±2 °C). The coil temperature wasthen raised by the passage of an electric current and the tail-flick latency(TFL) was measured. Heat was applied to a portion of the ventral sur-face of the tail between 4 and 6 cm from the tip. Each trial was terminat-ed after 6 s to minimize the probability of skin damage. TFL wasmeasured at 5-min intervals until a stable baseline was obtained overthree consecutive trials. Only rats showing stable baseline TFL after upto 6 trials were used in each experiment. The animals were then treatedwith intraperitoneal saline (control) or antagonist, and the TFL wasmeasured again within 30 s and then at 5-min interval for up to 3 hafter phenoxybenzamine or 15 min after the other antagonists. Gluta-mate or saline (0.25 μl) was then injected into the ZI, and the TFL wasmeasured within 30 s and then at 5-min intervals for up to 15 min.
2.4. Model of incision pain
Rats were placed in an elevated clear plastic cagewith a nylonmeshbottom, which allowed easy access to the paw plantar surface. Beforeeach test, the animals remained in the cage for 15 min to allow behav-ioral acclimation. The baseline threshold tomechanical punctate stimu-lation was measured with an electronic von Frey apparatus (IITCElectronic Equipments, CA, U.S.A.), consisting of a hand-held probeunit to which a rigid plastic tip (tip area=0.44 mm2) was connected.The plastic tip was applied with increasing force in an upward directionagainst sites near the heel, 1–2 mmadjacent to themedial border of thewound and to similar sites in the non-incised hind paw. The movementof the probe was interrupted when a withdrawal of the stimulated pawoccurred. During this procedure the applied force was continuouslyrecorded by a main unit connected to the probe. A single trial consistedof 3 applications of the tip, once every 5 s in each hind paw. Themean ofthree readings was taken as the mechanical threshold for a particulartiming. The animal was then anesthetized with isoflurane (in oxygenflow; 2% for induction and 0.5% for maintenance) via a loose-fitting,cone-shaped mask, and a 1-cm longitudinal incision was made with asurgical blade through the skin and fascia of the plantar aspect of theright hind paw, starting 0.5 cm from the proximal edge of the heel, asdescribed elsewhere (Brennan et al., 1996). The plantaris muscle wasleft intact during the procedure. The skin was then sutured with two5-0 nylon stitches. The thresholds for both hind paws of each rat weremeasured 2 h after the incision. A solution (0.25 μl) containing salineor glutamate was then injected into the ZI contralateral to the incisedpaw, and the mechanical threshold for both hind paws of each animalwas measured 30 s later and then at 5-min intervals for up to 25 min.
2.5. Rota-rod
TheRota-rod (UgoBasile, Comerio, Italy) consisted of a 7-cmdiameterdrum rotating at 25 rpm, and was used to assess motor coordination ofrats implanted 1 week earlier with a guide cannula. Each rat was placedon the drum for up to 2 min and the time of permanence was recordedon 2 occasions. An initial trial was conducted in naive rats, and the timespent by each rat on the drum was taken as baseline in the test.Twenty-four hours later, the time spent by each animal on the drumwas revaluated 30 s after the injection of saline or glutamate into the ZI.
2.6. Dorsolateral funiculus lesion
Rats were initially implanted with the guide cannula directed to theZI as described above. Two days later, they were anesthetized with tri-bromoethanol (250 mg/kg, i.p.), and a lesion of the dorsolateral funicu-lus (DLF) was made in 8 of them, bilaterally at the T1–T3 level under asurgical microscope. After laminectomy, the lesionwasmade by cuttinga portion of the dorsolateral quadrant of the spinal cord with a sharpknife. The overlying muscles and skin were sutured, and the animalwas allowed to recover for at least 5 days before the experiments. Theother rats were submitted to similar procedures, except for the cord le-sion, and served as sham-lesion rats. DLF-lesion rats were then injectedwith saline (4 animals) or glutamate (4–5 animals) in the ZI. Sham-lesion rats were also injected with saline (4–5 animals) or glutamate(4–5 animals) in the ZI.
2.7. Histology
At the end of the experiments, the animal was deeply anesthetizedwith intraperitoneal sodium thiopental and perfused through theheart with 4% paraformaldehyde in 0.1 M phosphate buffered saline.Fast green (1%, 0.25 μl) was injected through the guide cannula tolabel the site of intracerebral injection. The brain was removed andthe dye spot localized from 50-μm serial coronal sections stainedwith neutral red, and identified on diagrams from the atlas of

362 A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
Paxinos and Watson (1986). The thoracic spinal cord of rats submit-ted to DLF lesion was removed and the extent of the lesions wasalso identified on diagrams from the same atlas. The ZI is anatomicallydivided into the rostral, dorsal, ventral and caudal sectors (Mitrofanis,2005), but no attempt was made to group the rats according to eachsector.
2.8. Drugs
The following drugs were used: monosodium L-glutamate, atropinesulfate, mecamylamine, methysergide maleate, naloxone hydrochloride,phenoxybenzamine, and haloperidol and were purchased from Sigma(St. Louis, MO, USA). The antagonists were all prepared and diluted insaline, and their doses referred to the salt. The doses of glutamate usedhere (3.5 and 7.0 μg, corresponding to 23 and 47 nmol, respectively)were within the range (30–60 nmol) proposed elsewhere (Satoh et al.,1983; Jensen and Yaksh, 1989).
2.9. Data analysis
Tail-flick latencies (in seconds) are reported as means±SEM.Comparisons between control (rats treated in ZI and intraperitoneallywith saline) and test groups were made by multivariate analysis ofvariance (MANOVA) with repeated measures to compare the groupsover all times. The factors analyzed were treatments, time and treat-ment×time interaction. In the case of treatment×time interaction,one-way analysis of variance followed by Duncan's test was performedfor each time. The analysis was performed using the statistical softwarepackage SPSS/PC+, version 17.0.
The changes in the animal's performance in the Rota-rod werecompared using ANOVA followed by the Tukey's test. The level of signif-icance was set at Pb0.05 in all cases.
3. Results
3.1. Antinociception evoked by glutamate from ZI
The groups of rats used in each experiment were not significantlydifferent regarding baseline TFL. Rats injected with saline (0.25 μl;n=8) into the ZI had TFL that did not differ significantly from base-line throughout the observation period. A strong increase in TFL wasobtained soon after the injection of glutamate (7 μg/0.25 μl; n=8)into the ZI, the effect remaining significantly above control for atleast 5 min (Fig. 1A). A higher dose of glutamate (14 μg/0.25 μl;n=7) did not induce an additional effect, while a lower dose(3.5 μg/0.25 μl; n=8) induced a non-significant increase in TFL. Thecurves in Fig. 1A were significantly different regarding treatment(F3,27=43.62; Pb0.0001), time (F6,162=116.49; Pb0.0001), andtreatment×time interaction (F18,162=27.59; Pb0.0001). During orafter injection of glutamate, no animal showed gross disturbance ofmotor coordination or exhibited signs of aversion. The animalswalked normally and responded to non-noxious stimulation through-out the period of observation. The location of the microinjection sitesis shown in Fig. 1B. Some rats were purposely injected with glutamate(7 μg/0.25 μl) at sites near the ZI, and the results from sites in thesame region were pooled and compared to that obtained from ratsstimulated with glutamate in the ZI. A strong increase in TFL was con-firmed for the injection into the ZI (n=6), but not at sites neighbor-ing the ZI (Fig. 1C). The location of these sites is shown in Fig. 1D andincludes the ventral posteromedial (n=9) and ventral posterolateral(n=3) thalamic nuclei, and areas lateral (n=3) or ventral (n=9) tothe ZI.
The time course of the effects produced by the injection of sa-line or glutamate in the ZI on the post-incision pain is shown inFig. 1E. The groups did not differ significantly regarding the me-chanical thresholds for both hind paws measured immediately
before the incision (baseline). Two hours after the incision, a re-duction in the mechanical threshold of the incised paw was ob-served, thus characterizing the post incision pain. In contrast, nochange was observed in the non-incised hind paw as comparedto baseline. The post incision pain was abolished for at least10 min after the injection of glutamate (n=5) into the ZI contra-lateral to the incised paw, but remained unchanged after the in-jection of saline (n=5) into the same region. The mechanicalthreshold of the non incised paw was not changed significantlyafter the injection of saline (n=5) or glutamate (n=5) into theZI. The curves in Fig. 1E were significantly different regardingtreatment (F3,20=146.36; Pb0.0001) and time (F10,200=105,45;Pb0.0001), and had significant treatment×time interaction(F30,200=53.74; Pb0.0001). The location of the injection sites inthe ZI is shown in Fig. 1F. The mechanical threshold of bothhind paws of non incised rats was not changed significantlyafter the injection of saline (n=5) or glutamate (n=5) bilateralinto the ZI (Fig. 1G). The curves in Fig. 1G were not significantlydifferent regarding treatment (F3,18=0; P=0.89) and time(F6,108=1.06; P=0.53), nor had significant treatment×time in-teraction (F18,108=4.08; P=0.3). The location of the injectionsites in the ZI is shown in Fig. 1H. Representative microscopeimage with a dye spot restrict to the ZI is shown in Fig. 2.
3.2. Effect of dorsolateral funiculus lesion on the antinociception evokedby glutamate from ZI
The injection of saline (0.25 μl) into the ZI of DLF-lesion (n=5)or sham DLF-lesion (n=4) rats did not produce significant changesin TFL regarding baseline values. A strong increase in TFL followingthe injection of glutamate (7 μg/0.25 μl) into the ZI was confirmedin sham (n=5) DLF-lesion, but not in real (n=6) DLF-lesion rats(Fig. 3A). The curves in Fig. 3A were significantly different regardingtreatment (F(3,16)=9.43; Pb0.001), time (F(6,96)=8.38; Pb0.001)and treatment versus time interaction (F(18,96)=5.61; Pb0.001).The extent of the DLF lesion is shown in Fig. 3B to demonstratethat the tissue damage was restricted to the DLF and the adjacentdorsal horn. The location of the injection sites in the ZI at which glu-tamate induced antinociceptive effect is shown in Fig. 3C.
The injection of saline (0.25 μl) bilateral into the ZI of DLF-lesion (n=4) or sham DLF-lesion(n=5) rats also did not producesignificant changes in mechanical threshold of incised paw regard-ing pre incision values. The post incision pain was abolished for atleast 10 min after the injection of glutamate (7 μg/0.25 μl) bilateralinto the ZI of sham (n=5) DLF-lesion, but remained unchangedafter similar procedure in real (n=5) DLF-lesion rats (Fig. 4A).In this experiment, the mechanical threshold of non incisedpaws was also increased after glutamate (not shown in figures).The extent of the DLF lesion is shown in Fig. 4B and the locationof the injection sites in the ZI at which glutamate induced antino-ciceptive effect is shown in Fig. 4C.
3.3. Effects of injecting glutamate into the ZI on the Rota-rodperformance
Two groups of 5 rats each were used in this experiment andthe results are shown in Fig. 5A. The groups did not differ signif-icantly with regard to baseline performance on the Rota-rod.Rats injected with saline (0.25 μl) had performances that did notdiffer significantly from baseline. Similar results were obtainedwith rats injected with glutamate (7 μg/0.25 μl). The data in Figs. 5Awere significantly different (ANOVA: F3,23=2.99; P=0.71). Thelocation of the injection sites in the ZI for this experiment is shownin Fig. 5B.

Fig. 1. Time course of the changes in tail-flick latency (A and C) or incision pain (E) produced by the injection (arrow) of saline (0.25 μl) or glutamate into the ZI (A and E) or at siteswithin or near the ZI (C). A surgical incision was performed in the plantar aspect of the right paw. The effects of the injection of saline or glutamate bilateral into the ZI on the me-chanical threshold of both hind paws of non incised rats are shown in G. The doses of glutamate are given in A as μg/0.25 μl. In C, E and G, glutamate was used at a dose of 7 μg/0.25 μl. In E, glutamate was injected into the ZI contralateral to the incised paw. Points are means±S.E.M. of 7–8 animals in A, 3–9 animals in C, and 5 animals in E and G. Pointsare different from saline-treated rats (*) or from all the other groups (#), Pb0.05. The location of injection sites for the experiments shown in A, C, E and G is shown in B, D, F and H,respectively, on diagrams taken from Paxinos andWatson (1986). Abbreviations: VPM= ventral posteromedial thalamic nucleus; VPL= ventral posterolateral thalamic nucleus; ZI= zona incerta; RP = right paw; LP = left paw.
363A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
3.4. Effect of intraperitoneal non-selective antagonists on the glutamate-induced antinociception from ZI
Rats pretreated intraperitoneally with saline, methysergide, nal-oxone, phenoxybenzamine, atropine, mecamylamine, or haloperidol
were injected with saline (0.25 μl) or glutamate (7 μg/0.25 μl) intothe ZI. Glutamate or saline was injected into the ZI 3 h after phenox-ybenzamine and 15 min after the other antagonists. No change in TFLwas produced by the injection of saline into the ZI of rats treated in-traperitoneally with saline (0.1 ml/kg; n=7), compared to baseline
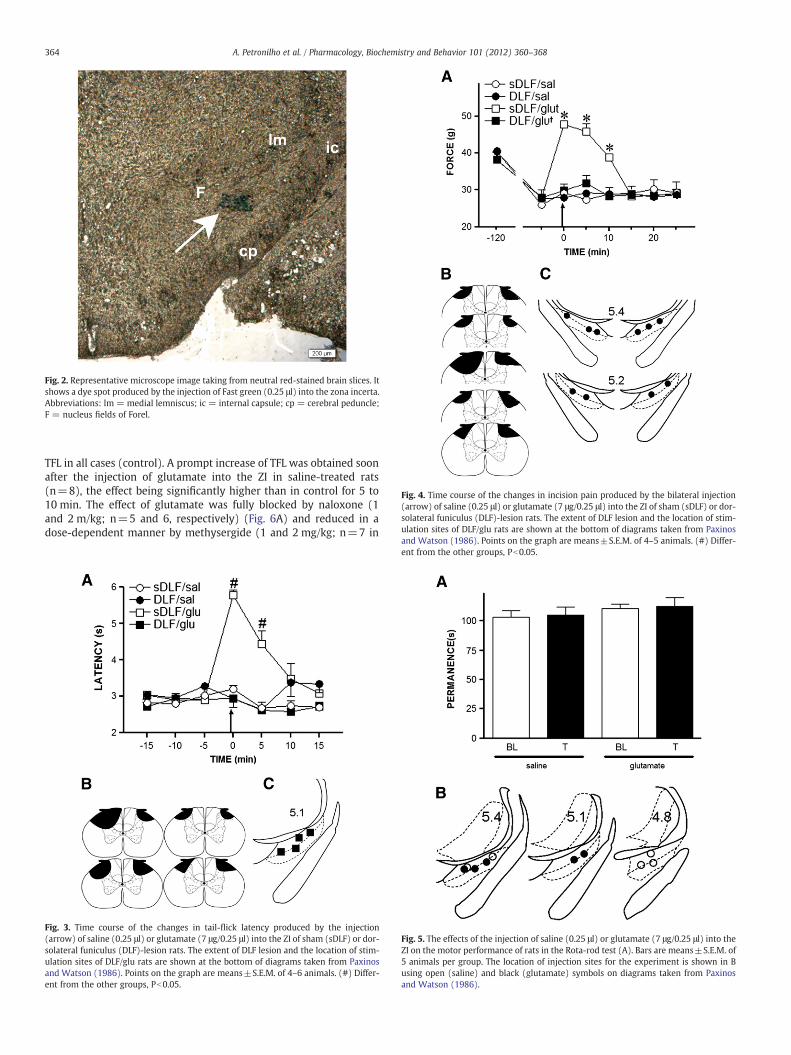
Fig. 2. Representative microscope image taking from neutral red-stained brain slices. Itshows a dye spot produced by the injection of Fast green (0.25 μl) into the zona incerta.Abbreviations: lm = medial lemniscus; ic = internal capsule; cp = cerebral peduncle;F = nucleus fields of Forel.
Fig. 4. Time course of the changes in incision pain produced by the bilateral injection(arrow) of saline (0.25 μl) or glutamate (7 μg/0.25 μl) into the ZI of sham (sDLF) or dor-solateral funiculus (DLF)-lesion rats. The extent of DLF lesion and the location of stim-ulation sites of DLF/glu rats are shown at the bottom of diagrams taken from Paxinos
364 A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
TFL in all cases (control). A prompt increase of TFL was obtained soonafter the injection of glutamate into the ZI in saline-treated rats(n=8), the effect being significantly higher than in control for 5 to10 min. The effect of glutamate was fully blocked by naloxone (1and 2 m/kg; n=5 and 6, respectively) (Fig. 6A) and reduced in adose-dependent manner by methysergide (1 and 2 mg/kg; n=7 in
Fig. 3. Time course of the changes in tail-flick latency produced by the injection(arrow) of saline (0.25 μl) or glutamate (7 μg/0.25 μl) into the ZI of sham (sDLF) or dor-solateral funiculus (DLF)-lesion rats. The extent of DLF lesion and the location of stim-ulation sites of DLF/glu rats are shown at the bottom of diagrams taken from Paxinosand Watson (1986). Points on the graph are means±S.E.M. of 4–6 animals. (#) Differ-ent from the other groups, Pb0.05.
and Watson (1986). Points on the graph are means±S.E.M. of 4–5 animals. (#) Differ-ent from the other groups, Pb0.05.
Fig. 5. The effects of the injection of saline (0.25 μl) or glutamate (7 μg/0.25 μl) into theZI on the motor performance of rats in the Rota-rod test (A). Bars are means±S.E.M. of5 animals per group. The location of injection sites for the experiment is shown in Busing open (saline) and black (glutamate) symbols on diagrams taken from Paxinosand Watson (1986).
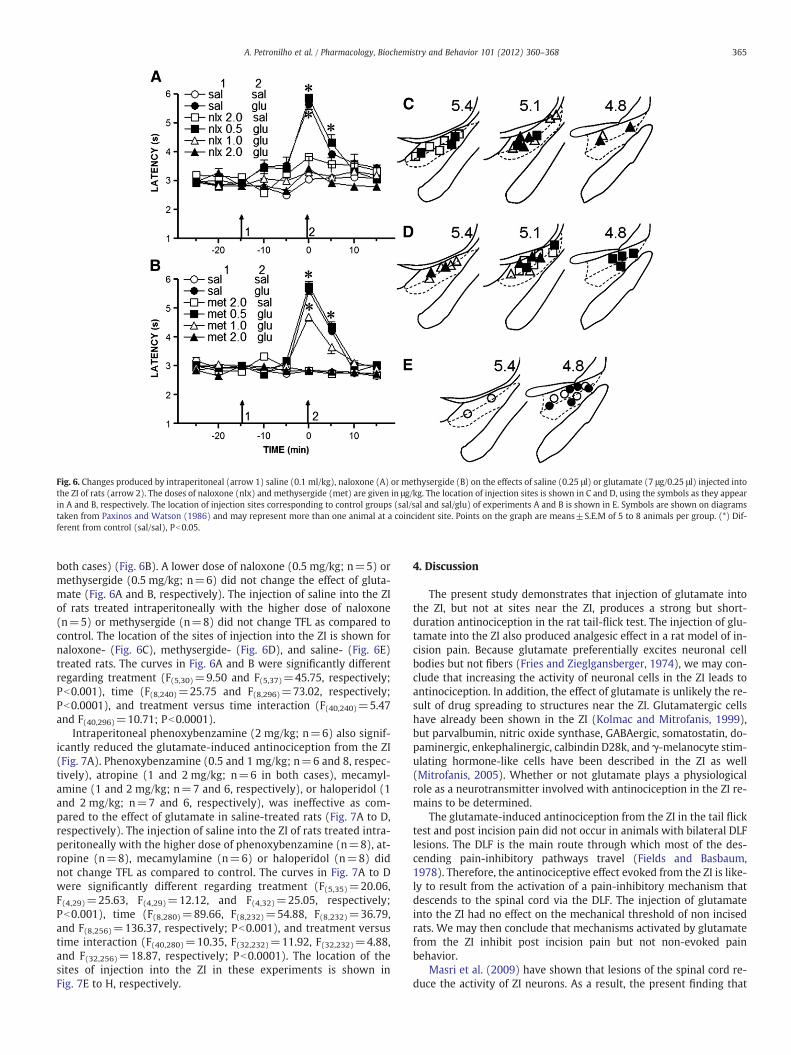
Fig. 6. Changes produced by intraperitoneal (arrow 1) saline (0.1 ml/kg), naloxone (A) or methysergide (B) on the effects of saline (0.25 μl) or glutamate (7 μg/0.25 μl) injected intothe ZI of rats (arrow 2). The doses of naloxone (nlx) and methysergide (met) are given in μg/kg. The location of injection sites is shown in C and D, using the symbols as they appearin A and B, respectively. The location of injection sites corresponding to control groups (sal/sal and sal/glu) of experiments A and B is shown in E. Symbols are shown on diagramstaken from Paxinos and Watson (1986) and may represent more than one animal at a coincident site. Points on the graph are means±S.E.M of 5 to 8 animals per group. (*) Dif-ferent from control (sal/sal), Pb0.05.
365A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
both cases) (Fig. 6B). A lower dose of naloxone (0.5 mg/kg; n=5) ormethysergide (0.5 mg/kg; n=6) did not change the effect of gluta-mate (Fig. 6A and B, respectively). The injection of saline into the ZIof rats treated intraperitoneally with the higher dose of naloxone(n=5) or methysergide (n=8) did not change TFL as compared tocontrol. The location of the sites of injection into the ZI is shown fornaloxone- (Fig. 6C), methysergide- (Fig. 6D), and saline- (Fig. 6E)treated rats. The curves in Fig. 6A and B were significantly differentregarding treatment (F(5,30)=9.50 and F(5,37)=45.75, respectively;Pb0.001), time (F(8,240)=25.75 and F(8,296)=73.02, respectively;Pb0.0001), and treatment versus time interaction (F(40,240)=5.47and F(40,296)=10.71; Pb0.0001).
Intraperitoneal phenoxybenzamine (2 mg/kg; n=6) also signif-icantly reduced the glutamate-induced antinociception from the ZI(Fig. 7A). Phenoxybenzamine (0.5 and 1 mg/kg; n=6 and 8, respec-tively), atropine (1 and 2 mg/kg; n=6 in both cases), mecamyl-amine (1 and 2 mg/kg; n=7 and 6, respectively), or haloperidol (1and 2 mg/kg; n=7 and 6, respectively), was ineffective as com-pared to the effect of glutamate in saline-treated rats (Fig. 7A to D,respectively). The injection of saline into the ZI of rats treated intra-peritoneally with the higher dose of phenoxybenzamine (n=8), at-ropine (n=8), mecamylamine (n=6) or haloperidol (n=8) didnot change TFL as compared to control. The curves in Fig. 7A to Dwere significantly different regarding treatment (F(5,35)=20.06,F(4,29)=25.63, F(4,29)=12.12, and F(4,32)=25.05, respectively;Pb0.001), time (F(8,280)=89.66, F(8,232)=54.88, F(8,232)=36.79,and F(8,256)=136.37, respectively; Pb0.001), and treatment versustime interaction (F(40,280)=10.35, F(32,232)=11.92, F(32,232)=4.88,and F(32,256)=18.87, respectively; Pb0.0001). The location of thesites of injection into the ZI in these experiments is shown inFig. 7E to H, respectively.
4. Discussion
The present study demonstrates that injection of glutamate intothe ZI, but not at sites near the ZI, produces a strong but short-duration antinociception in the rat tail-flick test. The injection of glu-tamate into the ZI also produced analgesic effect in a rat model of in-cision pain. Because glutamate preferentially excites neuronal cellbodies but not fibers (Fries and Zieglgansberger, 1974), we may con-clude that increasing the activity of neuronal cells in the ZI leads toantinociception. In addition, the effect of glutamate is unlikely the re-sult of drug spreading to structures near the ZI. Glutamatergic cellshave already been shown in the ZI (Kolmac and Mitrofanis, 1999),but parvalbumin, nitric oxide synthase, GABAergic, somatostatin, do-paminergic, enkephalinergic, calbindin D28k, and γ-melanocyte stim-ulating hormone-like cells have been described in the ZI as well(Mitrofanis, 2005). Whether or not glutamate plays a physiologicalrole as a neurotransmitter involved with antinociception in the ZI re-mains to be determined.
The glutamate-induced antinociception from the ZI in the tail flicktest and post incision pain did not occur in animals with bilateral DLFlesions. The DLF is the main route through which most of the des-cending pain-inhibitory pathways travel (Fields and Basbaum,1978). Therefore, the antinociceptive effect evoked from the ZI is like-ly to result from the activation of a pain-inhibitory mechanism thatdescends to the spinal cord via the DLF. The injection of glutamateinto the ZI had no effect on the mechanical threshold of non incisedrats. We may then conclude that mechanisms activated by glutamatefrom the ZI inhibit post incision pain but not non-evoked painbehavior.
Masri et al. (2009) have shown that lesions of the spinal cord re-duce the activity of ZI neurons. As a result, the present finding that

Fig. 7. Changes produced by intraperitoneal (arrow 1) saline (0.1 ml/kg), phenoxybenzamine (A), atropine (B), mecamylamine (C) or haloperidol (D) on the effects of saline(0.25 μl) or glutamate (7 μg/0.25 μl) injected into the ZI of rats (arrow 2). The doses of phenoxybenzamine (pbz), atropine (atr), mecamylamine (mec), and haloperidol (hal)are given in μg/kg. The location of injection sites for the experiments shown in A to D is shown in E to H, respectively, on diagrams taken from Paxinos and Watson (1986), andmay represent more than one animal at a coincident site. The location of injection sites corresponding to groups sal/sal and sal/glu in B is the same as shown in Fig. 3E. Pointsare means±S.E.M of 4 to 8 animals per group. (*) Different from control (sal/sal); (#) different from all the other groups, Pb0.05.
366 A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
ZI-evoked analgesia is prevented by spinal lesion might reflect thesuppressed activity of ZI. However, the extent of the DLF lesion usedin the present study was limited to the superficial dorsal horn,while the anterolateral quadrant of the spinal cord and the ventraland ventrolateral funiculi were targeted in Masri's experiment. There-fore, the suppressed activity of the ZI consequent to spinal lesion is un-likely to be the reason for the lack of analgesic effect of stimulating theZI in DLF-lesion animals.
The ZI plays a role in locomotor activity and this role is supported byanatomical evidences of ZI connections with motor-related centers,such as the red nucleus, the pontine nuclei and several hypothalamic nu-clei, the cervical and lumbosacral regions of the spinal cord (Mitrofanis,2005). We found here that the injection of glutamate into the ZI wasnot followed by changes of the animal performance in a Roda-rod test,which is used to assess motor changes (Dunham and Miya, 1957). Wemay then conclude that the antinociceptive effect shown here is unlikelyto result from a motor side effect of stimulating the ZI with glutamate.
Compared to control, the glutamate-induced antinociception fromthe ZI was prevented by intraperitoneal naloxone, methysergideor phenoxybenzamine (non-selective opioid, serotonergic and α-
adrenergic antagonist, respectively). In contrast, intraperitonealatropine, mecamylamine, or haloperidol (non-selective muscarin-ic, nicotinic, and dopaminergic antagonists, respectively) was inef-fective against glutamate-induced analgesia from the ZI eventhough used at doses equal to or higher than those shown to beeffective against chemically or electrically induced antinociception(Sahley and Berntson, 1979; Drago et al., 1984; Rees et al., 1987).Therefore, at least, serotonergic, α-adrenergic, and endogenousopioid mechanisms but not cholinergic or dopaminergic mecha-nisms seem to be involved in the modulation of the glutamate-induced antinociception from the ZI. However, we cannot makeconclusions about the particular site of action for each of the an-tagonists while using systemic antagonists.
Autoradiographic experiments showed connections from the ZI tothe thalamus, hypothalamus, APtN, superior colliculus, PAG, cunei-form nucleus, red nucleus, PPTg, NRM and Giα (Watanabe andKawana, 1982; Carlton et al., 1983). Glutamatergic projections fromthe ZI to PAG (Beitz, 1989) and PPTg (Heise and Mitrofanis, 2004)have already been demonstrated. The ZI also projects to the PAGand APtN in rats (Foster et al., 1989). The PAG (see Millan, 1999),

367A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
PPTg (Katayama et al., 1984; Iwamoto, 1989; Iwamoto and Marion,1993) and APtN (see Rees and Roberts, 1993) are structures fromwhich electrical stimulation evokes antinociception. ZI efferentsterminating within the spinal gray matter, particularly the ventralhorn, have already been demonstrated (Ricardo, 1981; Watanabeand Kawana, 1982; Peschanski et al., 1983; Roger and Cadusseau,1985; Romanowski et al., 1985; Shammah-Lagnado et al., 1985;Shiosaka et al., 1985; Shaw and Mitrofanis, 2001, 2002).
The stimulation-produced antinociception from the PAG activatesopioid and monoamine systems in the brain because naloxone andmonoamine antagonists potently reduce the effects of PAG stimulation(Basbaum and Fields, 1978).
The SPA from the NRM is mediated by serotonergic and endogenousopioid but not noradrenergic mechanisms (Fields and Basbaum, 1978;Azami et al., 1982), while noradrenaline but not serotonin mediatesthe effect from the parabrachial nucleus and locus coeruleus (seeMillan, 2002). Giα neurons that project to and activate spinally project-ing noradrenergic neurons located in the A7 cell group are involved inthe mediation of the SPA from the Giα (Clark and Proudfit, 1991).
The stimulation-induced antinociception from the PPTg is blockedby systemic naloxone, methysergide, phenoxybenzamine and atropine,but not by propranolol, haloperidol or mecamylamine (Dias et al.,2009). The PPTg participates as a relay station in a descending pathwayfrom the APtN (Terenzi et al., 1992), a brain structure fromwhich elec-trical stimulation produces a strong and long-lasting antinociception,which is mediated by opioid, α-adrenergic and muscarinic cholinergicmechanisms (Rees et al., 1987).
The injection of glutamate into the ZI inhibits the rat tail-flick reflexby mechanisms that resemble those activated by PAG or PPTg stimula-tion. Similarities include the effectiveness of opioid, serotonergic andα-adrenoceptor antagonists. Differing from the effect from the ZI,the SPA from the PPTg was sensitive to systemic atropine. The ZIwas stimulated here with glutamate which certainly activates thebrain structure in a manner more restricted than that provided byelectrical stimulation, as utilized in the aforementioned studies.
5. Conclusions
The injection of glutamate into the rat ZI produces a strong butshort-duration antinociception that does not occur in DLF-lesion ratsor in rats pretreated intraperitoneally with naloxone, methysergide orphenoxybenzamine. Therefore, the effect of the ZI is likely to resultfrom the activation of a pain-inhibitory pathway that descends to thespinal cord via the DLF and involves at least opioid, serotonergic andα-adrenergic mechanisms.
Acknowledgments
Supported by FAPESP. A.P., R.S.F., G.M.R. and Q.M.D. were the recipi-ents of FAPESP fellowships. The authors greatly appreciate the technicalassistance of M.A. Carvalho and P.R. Castania. Dr. A. Leyva helpedwith English editing of the manuscript.
References
Azami J, LlewelynMB, RobertsMHT. An extra-fine assembly for intracerebralmicroinjection.J Physiol 1980;305:18P–9P.
Azami J, Llewelyn MB, Roberts MHT. The contribution of nucleus reticularis paragiganto-cellularis and nucleus raphe magnus to the analgesia produced by systemicallyadministered morphine, investigated with the microinjection technique. Pain1982;12:229–46.
Basbaum AI, Fields HL. Endogenous pain control mechanisms: review and hypothesis.Ann Neurol 1978;4:451–62.
Beitz AJ. Possible origin of glutamatergic projections to the midbrain periaqueductalgrey and deep layer of the superior colliculus of the rat. Brain Res 1989;23:25–35.
Brennan TJ, Vandermeulen EP, Gebhart GF. Characterization of a rat model of incisionalpain. Pain 1996;64:493–501.
Carlton SM, Leichnetz GR, Young EG, Mayer DJ. Supramedullary afferents of the nucleusraphe magnus in the rat: a study using the transcannula HRP gel and autoradio-graphic techniques. J Comp Neurol 1983;214:43–58.
Clark FM, Proudfit HK. Projections of neurons in the ventromedial medulla to pontinecatecholamine cell groups involved in the modulation of nociception. Brain Res1991;540:105–15.
Desbois C, Villanueva L. The organization of lateral ventromedial thalamic connectionsin the rat: a link for the distribution of nociceptive signals to widespread corticalregions. Neuroscience 2001;102:885–98.
Dias QM, Crespilho SF, Silveira JWS, Prado WA. Muscarinic and α1-adrenergic mechanismscontribute to the spinal mediation of stimulation-induced antinociception from thepedunculopontine tegmental nucleus in the rat. Pharmacol Biochem Behav 2009;92:488–94.
Drago F, Caccamo G, Continella G, Scapagnini U. Amphetamine-induced analgesia doesnot involve brain opioids. Eur J Pharmacol 1984;101:267–9.
Dunham NW, Miya TS. A note on a simple apparatus for detecting neurological deficitin rats and mice. J Am Pharm Assoc 1957;46:208–9.
Edwards DA, Isaacs S. The zona incerta lesions: effects on copulation, partner preferenceand other socio-sexual behaviours. Behav Brain Res 1991;44:145–50.
Erzumulu RS, Killackey HP. Diencephalic projections of the subnucleus interpolaris ofthe brainstem trigeminal complex in the rat. Neuroscience 1980;5:1891–901.
Fields JL, Basbaum AI. Brainstem control of spinal pain-transmission neurons. Annu RevPhysiol 1978;40:217–48.
Foster GA, Sizer AR, Rees H, Roberts MHT. Afferent projections to the rostral anteriorpretectal nucleus of the rat: a possible role in the processing of noxious stimuli.Neuroscience 1989;29:685–94.
Fries W, Zieglgansberger W. A method to discriminate axonal from cell body activityand to analyse ‘silent cell’. Exp Brain Res 1974;21:441–5.
Giber K, Slézia A, Bokor H, Bodor AL, Ludányi A, Katona I, et al. Heterogeneous outputpathways link the anterior pretectal nucleus with the zona incerta and the thalamusin rat. J Comp Neurol 2008;506:122–40.
Gonzalez-Lima F, Helmstetter FJ, Agudo J. Functional mapping of the rat brain duringdrinking behavior: a fluorodeoxyglucose study. Physiol Behav 1993;54:605–12.
Heise CE, Mitrofanis J. Evidence for a glutamatergic projection from the zona incerta tothe basal ganglia in rats. J Comp Neurol 2004;468:482–95.
Iwamoto ET. Antinociception after nicotine administration into the mesopontine tegmen-tum of rats: evidence for muscarinic actions. J Pharmacol Exp Ther 1989;251:412–21.
Iwamoto ET, Marion L. Adrenergic, serotonergic and cholinergic components of nicotinicantinociception in rats. J Pharmacol Exp Ther 1993;265:777–89.
Jensen TS, Yaksh TL. Comparison of the antinociceptive effect of morphine and glutamateat coincidental sites in the periaqueductal gray and medial medulla in rats. Brain Res1989;476:1–9.
Katayama Y,Watkins LR, Becker DP, Hayes RL. Non-opiate analgesia induced by carbacholmicroinjection into the pontine parabrachial region of the cat. Brain Res 1984;296:263–83.
Kolmac CI, Mitrofanis J. Distribution of various neurochemicals within zona incerta:an immunocytochemical and histochemical study. Anat Embryol 1999;199:265–80.
Masri R, Quiton RL, Lucas JM, Murray PD, Thompson SM, Keller A. Zona incerta: a role incentral pain. J Neurophysiol 2009;102:181–91.
Millan MJ. The induction of pain: an integrative review. Prog Neurobiol 1999;57:1-164.Millan MJ. Descending control of pain. Prog Neurobiol 2002;66:355–474.Mitrofanis J. Some certainty for the “zone of uncertainty”? exploring the function of the
zona incerta. Neuroscience 2005;130:1-15.Mogenson GJ, Swanson LW,WuM. Evidence that projections from substantia innominata
to the zona incerta and mesencephalic locomotor region contribute to locomotoractivity. Brain Res 1985;334:65–76.
Okada N, Matsumoto N, Kitada Y. Responses of diencephalic nociceptive neurons toorofacial stimuli and effects of internal capsule stimulation in the rat. Arch OralBiol 2002;47:815–29.
Paxinos G, Watson C. The rat brain in stereotaxic coordinates. New York: AcademicPress; 1986.
Peschanski M. Trigeminal afferents to the diencephalon in the rat. Neuroscience1984;12:465–87.
Peschanski M, Manyth PW, Besson JM. Spinal afferents to the ventrobasal thalamiccomplex in rat: an anatomical study using wheat-germ-agglutinin conjugated tohorseradish peroxidase. Brain Res 1983;278:240–4.
Porro CA, Cavazzuti M, Lui F, Giuliani D, Pellegrini M, Baraldi P. Independent timecourses of supraspinal nociceptive activity and spinally mediated behavior duringtonic pain. Pain 2003;104:291–301.
Prado W, Roberts MHT. An assessment of the antinociceptive and aversive effects ofstimulating identified sites in the rat brain. Brain Res 1985;340:219–28.
Rees H, Roberts MHT. The anterior pretectal nucleus: a proposed role in sensory proces-sing. Pain 1993;53:121–35.
Rees H, PradoWA, Rawlings S, Roberts MHT. The effects of intraperitoneal administrationof antagonists and development of morphine tolerance on the antinociceptioninduced by stimulation of the anterior pretectal nucleus of the rat. Br J Pharmacol1987;92:769–79.
Ricardo JA. Efferent connections of the subthalamic region in the rat. II. The zona incerta.Brain Res 1981;214:43–60.
Roger M, Cadusseau J. Afferents to the zona incerta in the rat: a combined retrogradeand anterograde study. J Comp Neurol 1985;241:480–92.
Romanowski CAJ, Mitchell IJ, Crossman AR. The organization of the efferent projectionsof the zona incerta. J Anat 1985;143:75–95.
Sahley TL, Berntson GG. Antinociceptive effects of central and systemic administrationsof nicotine in the rat. Psychopharmacology (Berl) 1979;65:279–83.

368 A. Petronilho et al. / Pharmacology, Biochemistry and Behavior 101 (2012) 360–368
Satoh M, Oku R, Akaike A. Analgesia produced by microinjection of L-glutamate into therostral ventromedial bulbar nuclei of the rat and its inhibition by intrathecal alpha-adrenergic blocking agents. Brain Res 1983;261:361–4.
Senba E, Takagi H, Shiosaka S, Sakanaka M, Inagaki SN, Takatsuki K, et al. On the afferentprojections from some meso-diencephalic nuclei to n. raphe magnus of the rat. BrainRes 1981;211:387–92.
Shammah-Lagnado SJ, Negrão N, Ricardo JA. Afferent connections of the zona incerta: ahorseradish peroxidase study in the rat. Neuroscience 1985;15:109–34.
Shaw VE, Mitrofanis J. Lamination of spinal cells projecting to the zona incerta of rats.J Neurocytol 2001;30:695–704.
Shaw VE, Mitrofanis J. Anatomical evidence for somatotopic maps in zona incerta of rat.Anat Embryol 2002;206:119–30.
Shiosaka S, Kawai Y, Shibasaki T, TohyamaM. The descendingαMSHergic (αmelanocyte-stimulating hormone-ergic) projections from the zona incerta and lateral hypothalam-ic area to the inferior colliculus and spinal cord in the rat. Brain Res 1985;338:371–5.
Terenzi MG, Rees H, Roberts MHT. The pontine parabrachial region mediates some ofthe descending inhibitory effects of stimulating the anterior pretectal nucleus. BrainRes 1992;594:205–14.
Terenzi MG, Zagon A, Roberts MHT. Efferent connections from the anterior pretectalnucleus to the diencephalon and mesencephalon in the rat. Brain Res 1995;701:183–91.
Villarreal CF, Kina VAV, Prado WA. Antinociception induced by stimulating the anteriorpretectal nucleus in two models of pain in rats. Clin Exp Pharmacol Physiol 2004;31:608–13.
Watanabe K, Kawana E. The cells of origin of incertofugal projections to the tectum,thalamus, tegmentum, and spinal cord in rat: a study using autoradiographic andhorseradish peroxidase methods. Neuroscience 1982;10:2389–406.
Yen CT, Fu TC, Chen RC. Distribution of thalamic nociceptive neurons activated fromthe tail of the rat. Brain Res 1989;498:118–22.
ZimmermannM. Ethical guidelines for investigations of experimental pain in consciousanimals. Pain 1983;16:109–10.