phd thesis jdg (druk1) - department
TRANSCRIPT



Thesis for obtaining the degree of Doctor in Veterinary Sciences
(Ph.D.) Ghent University, 2013
Detection and mechanisms
of macrocyclic lactone resistance
in the bovine nematode
Cooperia oncophora
Jessie De Graef
Promotors
Prof. Dr. P. Geldhof and Prof. Dr. E. Claerebout
Laboratory of Parasitology Department of Virology, Parasitology and Immunology
Faculty of Veterinary Medicine, Ghent University Salisburylaan 133, B-9820 Merelbeke


1
TABLE OF CONTENTS
LIST OF FIGURES 4
LIST OF TABLES 5
LIST OF ABBREVIATIONS 6
CHAPTER 1: Anthelmintic resistance in cattle nematodes – A review
1.1 Introduction to anthelmintic resistance 9
1.2 Detecting anthelmintic resistance 12 1.2.1 Controlled efficacy test 12
1.2.2 Faecal egg count reduction test 13
1.2.3 In vitro assays 14
1.2.4 Molecular detection techniques 15
1.3 Anthelmintic resistance on Belgian cattle farms 15
1.4 Factors affecting the development of anthelmintic resistance 16 1.4.1 Parasite genetics and biology 17
1.4.2 Refugia and management factors 17
1.4.3 Sub-therapeutic drug levels 18
1.5 Macrocyclic lactones 18 1.5.1 Pharmacokinetics 19
1.5.2 Effects on nematodes and mode of action 20
1.5.3 Structure and localization of the receptor 21
1.6 Mechanisms of macrocyclic lactone resistance 23
1.6.1 Glutamate-gated chloride channels 23
1.6.2 P-glycoproteins 24
1.6.3 Other candidate genes 26
1.7 Conclusion 28
OBJECTIVES 29
CHAPTER 2: Assessing resistance against macrocyclic lactones in gastro-intestinal nematodes
in cattle using the faecal egg count reduction test and the controlled efficacy test
2.1 Introduction 35
2.2 Materials and methods 35
2.2.1 Nematode isolates 35
2.2.2 Experimental design 36
2.2.3 Parasitological techniques 36
2.2.4 Determination of efficacy 37
2.3 Results 37
2.3.1 Ostertagia ostertagi 37
2.3.2 Cooperia oncophora 37
2.4 Discussion 41

2
CHAPTER 3: Screening of the Cooperia oncophora transcriptome database for candidate
genes involved in macrocyclic lactone resistance
3.1 Introduction 45
3.2 Materials and methods 45 3.2.1 Parasite material and RNA extraction 45
3.2.2 ‘Next-generation’ sequencing and processing of the reads 46
3.2.3 Identification of candidate resistance genes 46
3.2.4 Degenerate PCR approach and full-length amplification 47
3.2.5 Reverse transcriptase PCR 47
3.3 Results 48
3.3.1 Transcript reconstruction 48
3.3.2 Identification of GluCl subunit genes 48
3.3.3 Identification of ABC transporter genes 51
3.4 Discussion 51
CHAPTER 4: Gene expression and mutation analysis of glutamate-gated chloride channels in
resistant Cooperia oncophora isolates following in vivo exposure to macrocyclic lactones
4.1 Introduction 57
4.2 Materials and methods 58 4.2.1 Parasite material 58
4.2.2 RNA extraction and cDNA synthesis 58
4.2.3 Quantitative real-time PCR 58
4.2.4 Mutation analysis of the full-length Con-avr-14B and Con-glc-6 sequences 59
4.3 Results 60 4.3.1 Analysis of constitutive and inducible transcriptional changes of GluCl subunit genes 60
4.3.2 Mutation analysis of Con-avr-14B 64
4.3.3 Mutation analysis of Con-glc-6 64
4.4 Discussion 68
CHAPTER 5: Gene expression analysis of ABC transporters in a resistant Cooperia oncophora
isolate following in vivo and in vitro exposure to macrocyclic lactones
5.1 Introduction 73
5.2 Materials and methods 74 5.2.1 Parasite material 74
5.2.2 RNA extraction and cDNA synthesis 74
5.2.3 Quantitative real-time PCR 75
5.3 Results 75 5.3.1 Analysis of constitutive transcriptional changes of ABC transporter genes 75
5.3.2 Analysis of inducible transcriptional changes after in vivo exposure of adult worms 77
5.3.3 Analysis of inducible transcriptional changes after in vitro exposure of third stage larvae 78
5.4 Discussion 79

3
CHAPTER 6: General discussion
6.1 Introduction 83
6.2 Is the FECRT sti l l useful in the field? 83
6.3 Molecular background of macrocyclic lactone resistance 85 6.3.1 The role of glutamate-gated chloride channels in macrocyclic lactone resistance 85
6.3.2 The role of ABC transporters in macrocyclic lactone resistance 87
6.4 Prospects for molecular methods to detect macrocyclic lactone resistance in the
field 88
6.5 Delaying macrocyclic lactone resistance 90
6.6 Conclusion 91
SUMMARY/SAMENVATTING 93
APPENDIX A 104
APPENDIX B 106
REFERENCES 108
ACKNOWLEDGEMENTS/DANKWOORD 125

4
LIST OF FIGURES
FIGURE 1.1: The main effects of macrocyclic lactones on nematodes: paralysis of the pharynx,
somatic muscles and uterus.
FIGURE 1.2: Schematic representation of a glutamate-gated chloride channel.
FIGURE 1.3: The role of P-glycoprotein (PGP) in macrocyclic lactone (ML) efflux from the cell
and a model for transmembrane topology of PGP.
FIGURE 3.1: Phylogenetic analysis of identified glutamate-gated chloride channel subunits in
Cooperia oncophora, Ostertagia ostertagi, Haemonchus contortus and Caenorhabditis elegans.
FIGURE 4.1: Fold changes in constitutive and inducible mRNA transcript levels of glc-2, glc-3,
glc-4, glc-6, avr-14A and avr-14B in Cooperia oncophora adult worms.
FIGURE 4.2: Sequence alignment of the predicted protein sequences of Cooperia oncophora
avr-14B and glc-6 genes with orthologous sequences from Caenorhabditis elegans,
Haemonchus contortus and Ostertagia ostertagi.
FIGURE 4.3: Isoform frequencies of predicted full-length protein sequences of Con-AVR-14B
and Con-GLC-6.
FIGURE 4.4: Summary of all amino acid substitutions in the predicted full-length protein
sequences of Con-AVR-14B and Con-GLC-6 and their frequencies per Cooperia oncophora
isolate investigated.
FIGURE 5.1: Fold changes in constitutive mRNA transcript levels of ABC transporter genes in
Cooperia oncophora eggs, L3 and adult worms.
FIGURE 5.2: Fold changes in inducible mRNA transcript levels of ABC transporter genes in
Cooperia oncophora adult worms after in vivo exposure to macrocyclic lactones.
FIGURE 5.3: Fold changes in inducible mRNA transcript levels of ABC transporter genes in
Cooperia oncophora L3 after in vitro exposure to ivermectin.

5
LIST OF TABLES
TABLE 1.1: Introduction of anthelmintic drugs onto the market and the development of
resistance to the drug.
TABLE 1.2: Prevalence of anthelmintic resistance in bovine nematodes.
TABLE 2.1: Arithmetic means and range of the Ostertagia ostertagi faecal egg counts and
post-mortem total worm counts.
TABLE 2.2: Arithmetic means and range of the Cooperia oncophora faecal egg counts and
post-mortem total worm counts.
TABLE 2.3: Comparison of the faecal egg count reduction calculation methods
TABLE 3.1: Summary of assembly and annotation information from the Cooperia oncophora
transcriptome database.
TABLE 3.2: Sequence sizes, accession numbers and transcription patterns throughout the life
cycle of Cooperia oncophora of the 6 so far identified glutamate-gated chloride channel
subunits.
TABLE 3.3: Sequence sizes, accession numbers, blast analyses and reverse-transcriptase PCR
results of the 15 partially identified Cooperia oncophora ABC transporter genes to determine
their correct annotation and to show their transcription pattern throughout the life cycle of C.
oncophora.

6
LIST OF ABBREVIATIONS
AA amino acids
ABC ATP-binding cassette
ABZ albendazole
ATP adenosine triphosphate
Bp base pair
BZs benzimidazoles
cDNA complementary deoxyribonucleic acid
CEGs core eukaryotic genomes
CET controlled efficacy test
DNA deoxyribonucleic acid
EC50 effective concentration to have half-maximal effect
e.g. exempli gratia
EPG eggs per gram faeces
EST expressed sequence tag
F female
FBZ fenbendazole
FEC faecal egg count
FECR faecal egg count reduction
FECRT faecal egg count reduction test
GABA γ-aminobutyric acid
GluCl glutamate-gated chloride channel
GST glutathione S-transferase
HAF half transporter
I/Ts imidothiazoles/tetrahydropyrimidines
i.e. id est
IVM ivermectin
L1, L2, L3, L4 first, second, third, fourth stage larvae
LDA larval development assay
LEV levamisole
LFA larval feeding assay
LMIA larval midration inhibition assay
NBD nucleotide-binding domain
M male
MLs macrocyclic lactones
MMT micro-motility meter test
mRNA messenger ribonucleic acid
MRP multidrug resistant protein
MOX moxidectin
NADP nicotinamide adenine dinucleotide phosphate
PCR polymerase chain reaction
PGE parasitic gastroenteritis
PGP P-glycoprotein
TST targeted selective treatment
WAAVP world association for the advancement of veterinary parasitology
xL3 ex-sheathed third stage larvae

CHAPTER 1 Anthelmintic resistance in cattle nematodes - A review

8
Based on: De Graef J, Claerebout E and Geldhof P. Anthelmintic resistance in cattle
nematodes. Vlaams Diergeneeskundig Tijdschrift. 2013 (submitted).

CHAPTER 1: Anthelmintic resistance in cattle nematodes
9
1.1 Introduction to anthelmintic resistance
Worldwide, infections with parasitic nematodes restrict welfare and productivity of livestock.
Control of these parasites relies heavily on the administration of anthelmintic drugs. Between
1960 and 1990s, the pharmaceutical industry made major progress in developing deworming
compounds with excellent broad-spectrum activity and safety [1]. This led to the discovery of
three major drug classes available for ruminants, each with distinct modes of action: the
benzimidazoles (BZs), the imidazothiazoles and tetrahydropyrimidines (I/Ts) and the
macrocyclic lactones (MLs). Relatively short after their introduction onto the market, all
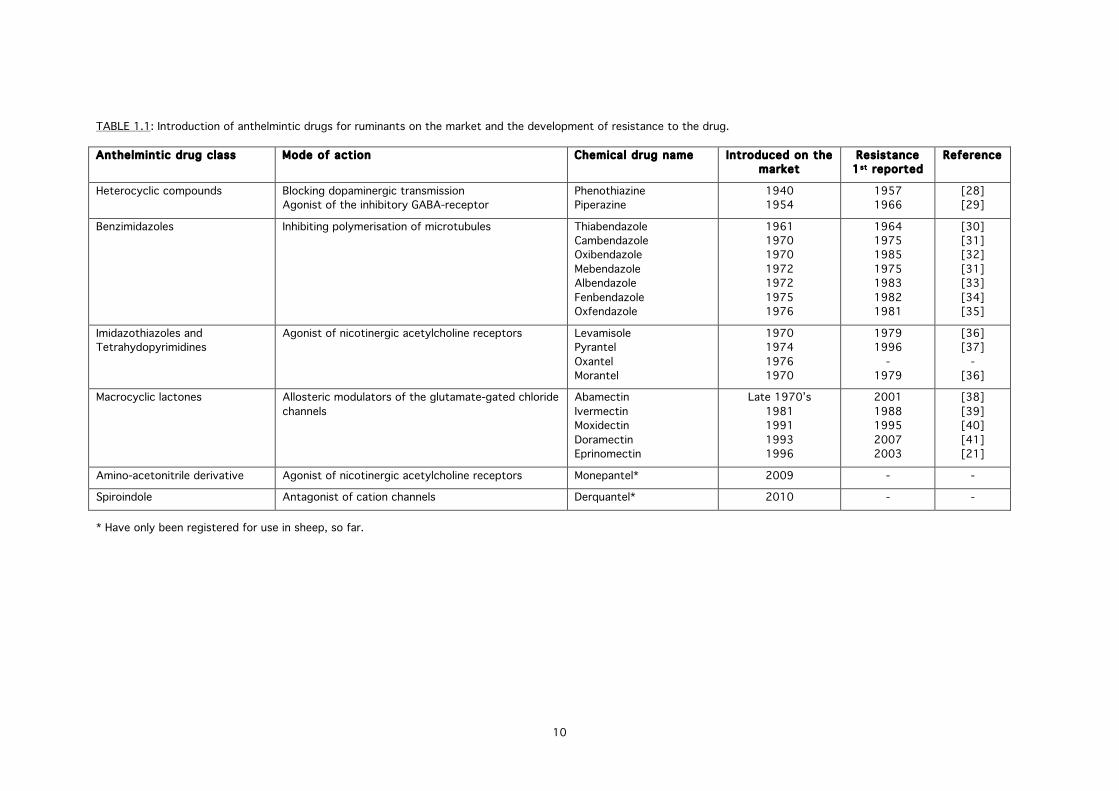
anthelmintic drug classes have become the victim of their own success (TABLE 1.1). The more
intensively parasites are being controlled with drugs, the more likely resistance develops [2].
Anthelmintic resistance occurs when parasites, usually eliminated by a specific dose, suddenly
survive the treatment. Since resistance is inherited, the surviving worms will pass their
resistance alleles to their progeny [3]. Today, the problem of anthelmintic resistance is by far
the most severe in small ruminants. Multidrug-resistance is documented to numerous gastro-
intestinal nematodes of sheep and goats (e.g. Haemonchus contortus, Teladorsagia
circumcincta, Trichostrongylus spp. and Cooperia spp.) [4-9]. In South Africa, New Zealand,
Australia and the UK, multidrug-resistance even has forced farmers to stop sheep and goat
farming [10-12].
Compared to small ruminants, relatively few field surveys have been performed to investigate
the prevalence of anthelmintic resistance in cattle parasites. Therefore, the number of cases of
cattle nematodes resistant to anthelmintic drugs might be considerably underestimated.
Resistance against I/Ts or BZs is reported in most of the major gastro-intestinal nematodes of
cattle (e.g. Cooperia spp., Haemonchus placei, Ostertagia ostertagi and Trichostrongylus spp.).
The prevalence of ML-resistance in cattle nematodes, especially Cooperia spp., is increasing in
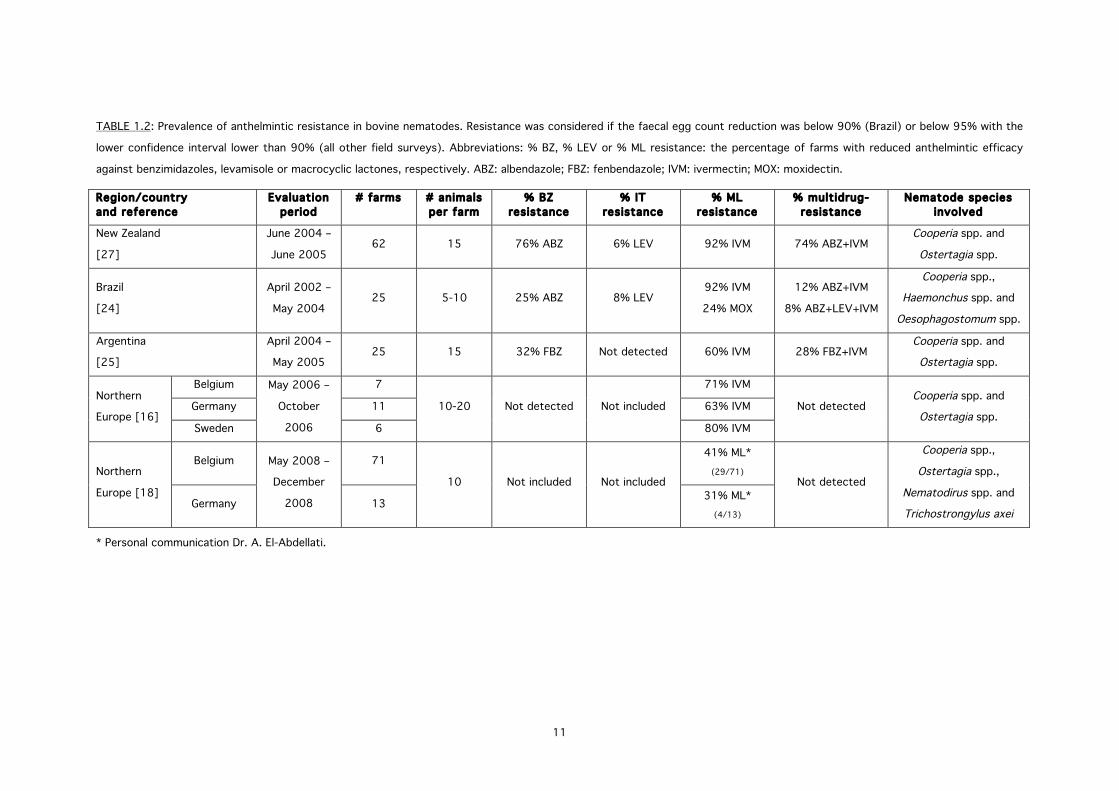
New Zealand, Argentina, Brazil, the USA and Northern Europe, including Belgium [13-27].
TABLE 1.2 summarizes the results of the few field surveys that were conducted in order to
assign the extent of anthelmintic resistance in bovine nematodes. Most alarmingly are the
reports of multidrug-resistance against both MLs and BZs, with a prevalence reaching 74%,
12% and 28% for New Zealand, Brazil and Argentina, respectively [24, 25, 27]. The main
species found after the treatment failure was C. oncophora.
In these 5 surveys, anthelmintic resistance was determined based on the faecal egg count
reduction test (see section 1.2.2) in combination with larval group cultures of the treated
animals and un-treated (if included) animals. Unfortunately, only a low number of farms per
region were included, which makes it difficult to estimate the precise anthelmintic resistance
prevalence on a regional/national scale. The reason why only a few farms participate is
because of the stringent selection criteria and the discouraging workload.
10
TABLE 1.1: Introduction of anthelmintic drugs for ruminants on the market and the development of resistance to the drug.
Anthelmintic drug class Mode of action Chemical drug name Introduced on the market
Resistance 1st reported
Reference
Heterocyclic compounds Blocking dopaminergic transmission Agonist of the inhibitory GABA-receptor
Phenothiazine Piperazine
1940 1954
1957 1966
[28] [29]
Benzimidazoles Inhibiting polymerisation of microtubules Thiabendazole Cambendazole Oxibendazole Mebendazole Albendazole Fenbendazole Oxfendazole
1961 1970 1970 1972 1972 1975 1976
1964 1975 1985 1975 1983 1982 1981
[30] [31] [32] [31] [33] [34] [35]
Imidazothiazoles and Tetrahydopyrimidines
Agonist of nicotinergic acetylcholine receptors Levamisole Pyrantel Oxantel Morantel
1970 1974 1976 1970
1979 1996
- 1979
[36] [37]
- [36]
Macrocyclic lactones Allosteric modulators of the glutamate-gated chloride channels
Abamectin Ivermectin Moxidectin Doramectin Eprinomectin
Late 1970’s 1981 1991 1993 1996
2001 1988 1995 2007 2003
[38] [39] [40] [41] [21]
Amino-acetonitrile derivative Agonist of nicotinergic acetylcholine receptors Monepantel* 2009 - -
Spiroindole Antagonist of cation channels Derquantel* 2010 - -
* Have only been registered for use in sheep, so far.
11
TABLE 1.2: Prevalence of anthelmintic resistance in bovine nematodes. Resistance was considered if the faecal egg count reduction was below 90% (Brazil) or below 95% with the
lower confidence interval lower than 90% (all other field surveys). Abbreviations: % BZ, % LEV or % ML resistance: the percentage of farms with reduced anthelmintic efficacy
against benzimidazoles, levamisole or macrocyclic lactones, respectively. ABZ: albendazole; FBZ: fenbendazole; IVM: ivermectin; MOX: moxidectin.
Region/country and reference
Evaluation period
# farms # animals per farm
% BZ resistance
% IT resistance
% ML resistance
% multidrug-resistance
Nematode species involved
New Zealand
[27]
June 2004 –
June 2005 62 15 76% ABZ 6% LEV 92% IVM 74% ABZ+IVM
Cooperia spp. and
Ostertagia spp.
Brazil
[24]
April 2002 –
May 2004 25 5-10 25% ABZ 8% LEV
92% IVM
24% MOX
12% ABZ+IVM
8% ABZ+LEV+IVM
Cooperia spp.,
Haemonchus spp. and
Oesophagostomum spp.
Argentina
[25]
April 2004 –
May 2005 25 15 32% FBZ Not detected 60% IVM 28% FBZ+IVM
Cooperia spp. and
Ostertagia spp.
Northern
Europe [16]
Belgium May 2006 –
October
2006
7
10-20 Not detected Not included
71% IVM
Not detected Cooperia spp. and
Ostertagia spp. Germany 11 63% IVM
Sweden 6 80% IVM
Northern
Europe [18]
Belgium May 2008 –
December
2008
71
10 Not included Not included
41% ML* (29/71)
Not detected
Cooperia spp.,
Ostertagia spp.,
Nematodirus spp. and
Trichostrongylus axei Germany 13 31% ML*
(4/13)
* Personal communication Dr. A. El-Abdellati.

CHAPTER 1: Anthelmintic resistance in cattle nematodes
12
On the other hand, some differences may be observed between the experimental designs: e.g.
the detection limit of the faecal egg counts, the selection of the farms, the initial faecal egg
counts before treatment, the route of administration of the anthelmintic and the calculation
method to determine the efficacies. These methodological considerations will have a
considerable impact on the study result and make it difficult to compare and extrapolate the
observations [18].
In this chapter, a summary is given of the detection methods to evaluate the efficacy of MLs
against ruminant nematodes. Subsequently, the most recent findings about the resistance
status of nematode species in cattle in Belgium are described. Then, factors affecting the
development of anthelmintic resistance will be discussed and finally, the current knowledge of
molecular mechanisms at the base of ML-resistance will be summarized.
1.2 Detecting anthelmintic resistance
Frequent drug treatments increase the selection pressure on the resistance alleles that
appeared in the parasite population. At a certain point, the anthelmintic drug is no longer
useful in protecting the host against parasite infections and a change to another drug, with a
different mode of action, is necessary. It is of great importance to detect anthelmintic
resistance as early as possible, whilst the frequency of resistance alleles in the parasite
population is still low. In this way, the onset of anthelmintic resistance could be delayed and
the efficacy of the currently used anthelmintic drugs could be maintained for longer [42].
The World Association for the Advancement of Veterinary Parasitology (WAAVP) provided
guidelines on the detection of anthelmintic resistance [43-46]. However, in cattle, it is still
non-standardized and therefore it remains difficult to assign the correct resistance status and
to compare data among different surveys. The most accepted methods are two in vivo
methods: the faecal egg count reduction test (FECRT) and the controlled efficacy test (CET).
Although the CET is the most reliable method, the FECRT is more commonly used [43, 44]. In
the following paragraphs the strengths and drawbacks of the available diagnostic tests for ML-
resistance are discussed.
1.2.1 Controlled efficacy test
This in vivo test is suitable for all types of anthelmintic drugs and is the gold standard for
evaluating their efficacy. The CET, or slaughter trial, requires the infected host to be sacrificed.
Therefore, this test is rarely used for diagnosing resistance in the field, but ideal for dose-
confirmation studies or when confirmation of resistance is required. The percentage efficacy is
determined by comparing the means of surviving parasites in groups of treated and untreated
animals after an artificial or natural infection [44, 46, 47]. Resistance is confirmed when the
reduction in worm counts is <90%, or more than 1000 worms survived the treatment [47].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
13
1.2.2 Faecal egg count reduction test
This in vivo procedure is currently the most practical method for field diagnosis of resistance,
against any anthelmintic drug. Based on the microscopic detection of nematode eggs in faecal
samples of the infected host before and after treatment, the reduction in faecal egg counts
(FECs) is calculated. Standards for the FECRT only exist for sheep at the moment. An accurate
determination of resistance is more difficult in infected cattle than in small ruminants, since
the FECs tend to be lower [48, 49]. A population of worms is declared to be resistant if the
percentage reduction is <95% and the lower 95% confidence interval is <90%, resistance is
suspected if only 1 of the 2 criteria is met [43].
The major limitation of the FECRT is its lack of sensitivity [42]. The modified McMaster
technique, with a detection limit of 50 eggs per gram (EPG), often fails to detect low numbers
of eggs. As a consequence, an early diagnosis of resistance is impeded [50]. If pre-treatment
egg counts are <150 EPG, a more sensitive counting method is recommended. Recently, the
commercial FECPAK counting system was introduced, which has a sensitivity of up to 10 EPG
to test for nematode egg counts in cattle (www.fecpak.com). The FLOTAC technique, with a
detection limit of 1-2 EPG, reaches the required sensitivity, but loses on practicality [51]. The
Moredun Technique can also be used with a sensitivity up to 1 EPG [52]. Another drawback of
the FECRT is that it is not species-specific, unless morphological or molecular analysis of the
parasite material is conducted. In a mixed infection it is difficult to differentiate microscopically
the eggs between nematode species [53]. In order to calculate species-specific drug
efficacies, it is suggested to conduct larval cultures of pre- and post-treatment samples, from
which third stage larvae can be harvested and differentiated. A third disadvantage is that the
FECRT is not user friendly (labour intensive). Therefore, its use as a monitoring tool is limited.
A possible option would be the use of pooled samples.
The interpretation of the FECRT is affected by a complex interplay of various factors, including
the detection limit of the FEC method, the number of animals per treatment group and the
level of excretion and aggregation of the FECs [54]. Besides, the correlation between egg
counts and worm numbers is not always so clear, especially not in cattle [44, 55, 56]. Due to
the temporary sterilising effect of BZs and MLs, faecal samples should be collected 8-10 or 14-
17 days after treatment with BZs or MLs, respectively [44]. The variability in the FECR data
can also be attributed to the calculation methods (i.e. geometric means of FECs appear to
overestimate the efficacy compared to arithmetic FEC means) and multiple formulas that are
available (i.e. formulas can include/exclude untreated control groups or can be based on
individual FECs instead of group mean FECs) [43, 46, 47, 57-59].
The outcome of the FECRT is also prone to confounding factors, which also apply to the CET.
To reduce the likelihood of false positive results (reduced anthelmintic efficacy without true
anthelmintic resistance), some requirements should be taken into account. Weighing the

CHAPTER 1: Anthelmintic resistance in cattle nematodes
14
animals is essential to avoid a sub-optimal treatment dosage. The pharmacokinetics of the
drug vary according to the route of administration (bolus, topical, oral or injectable),
formulation, body condition, age and physiological status. All of these factors contribute to
differences in (persistent) activity of the anthelmintic and may result in a lower drug efficacy if
the product is eliminated from the body of the host too fast [60-62].
1.2.3 In vitro assays
In vitro assays have the advantages of low cost and having no inter-host variation, as well as
the opportunity for replication and standardization [63]. ML-resistance can be detected by the
following in vitro tests: � Larval migration inhibition assay (LMIA); � Micro-motility meter test
(MMT); � Larval development assay (LDA) and � Larval feeding assay (LFA). Recently, the
first three of these in vitro tests have been adapted and evaluated for the detection of
resistance in gastro-intestinal nematodes of cattle [64].
Migration and motility tests are based on the drug-induced paralysis of the body musculature
of trichostrongyloid nematodes. In the LMIA, ex-sheathed third stage larvae (L3) are incubated
in serial dilutions of anthelmintic for 24 hours, and subsequently transferred onto a sieve for a
further 24 hours. Resistant L3 will be able to migrate through the sieve, while susceptible L3
remain on the mesh. Subsequently, the percentage migrated L3 is calculated. In the MMT,
movements of L3 or adult worms, incubated in anthelmintic dilutions, will fractionate light rays,
which are measured with a photo-detector. The numerical representation of this signal is
termed the motility index. Active worms give higher indices than paralyzed worms [64, 65]. L3
are a non-feeding stage, can easily be collected, maintained and transported and hence, the
use of L3 is advantageous.
The LDA measures the potency of the anthelmintic as inhibitor of the development. In case of
the LDA, trichostrongyloid eggs are incubated for 6-8 days in a growth medium, with
Escherichia coli and yeast as a food source and with the anthelmintic under test. Subsequently,
the percentage developed L3 is calculated. Fresh eggs are the most crucial factor for
successful performance of the LDA [64, 66]. A commercial LDA (Drenchrite®) has been
developed for the detection of BZ- and levamisole (LEV)-resistance in sheep and goat
nematodes [67]. ML-resistance can also be diagnosed with the LFA in which first stage larvae
(L1) are cultured with fluorescein-5-isothiocyanate-labelled E. coli and serial dilutions of the
anthelmintic. Under a fluorescence microscope the ratio of fed and unfed larvae at each drug
concentration is determined [68].
The results of in vitro tests are interpreted using EC50 values, describing the concentration at
which a drug is half-maximal effective (50% of the parasites is killed). As by definition,
resistant isolates will have higher EC50 values compared to susceptible isolates. The biggest
challenge for all diagnostic bioassays remains the establishment of reference EC50 values.

CHAPTER 1: Anthelmintic resistance in cattle nematodes
15
Therefore, the accuracy, sensitivity, repeatability and reproducibility for different isolates and
species (mixed infections) in different laboratories still require optimization. Additionally,
validation against in vivo data is still required, since the pharmacology of the drug in the host-
parasite system is lost in in vitro assays [63].
1.2.4 Molecular detection techniques
Theoretically, molecular tests are capable of detecting resistance alleles when the frequency of
these alleles is still very low. Therefore, a genetic test for resistance requires the knowledge of
the molecular basis of resistance. The identification of mutations in target genes or the
detection of alterations in the expression of genes could lead to the development of probes,
respectively for pyrosequencing or real-time PCR. These techniques would enable the
determination of susceptible or resistant populations [69-71]. So far, molecular markers for
detecting and measuring anthelmintic-resistance only exist for BZs in sheep nematodes.
Therefore, the WAAVP strongly encourages further investigation of the genetic mechanisms of
resistance, especially in bovine nematodes. Despite their expensive equipment, molecular
techniques could be more sensitive and less time-consuming than current in vivo and in vitro
detection methods. As for in vivo and in vitro tests, the challenge still remains the correct
identification of resistance in mixed parasite infections. Furthermore, tests based on the
detection of one single mutation to diagnose resistance, will make an underestimation, if
resistance may have resulted from more than one underlying mechanism [44, 69, 72, 73].
1.3 Anthelmintic resistance on Belgian cattle farms
The predominant nematode species infecting cattle in the temperate, European climate are O.
ostertagi and C. oncophora with 100% prevalence on pastures grazed by cattle [74]. In
Belgium, 72% of the farms use MLs to control parasite infections, of which 27% specifically
use ivermectin (IVM) [75]. Cooperia spp. are considered to be the dose-limiting species for
MLs, this means that the recommended dose is determined based on the efficacy against
these species [76].
The first report of a reduced IVM efficacy on Belgian cattle farms dates from 2006 [16]. At
that time, 7 farms were investigated and on all farms reduced efficacies were observed 21
days after IVM treatment, with FEC reductions ranging from 58-95%. After a revisit, the
reduced IVM efficacy could only be confirmed on 1 farm, with a FEC reduction of 54% on day
21 post-treatment. On all farms, only C. oncophora was recovered from the larval cultures. On
one particular farm, the evolution of IVM-resistance was further monitored during 4
consecutive years and showed a rapid increase of the resistance level in C. oncophora. After
IVM treatment, reductions in FECs of 73%, 40% and 0% were recorded, respectively in 2006,
2007 and 2008. One year later, side-resistance against moxidectin (MOX) was also determined

CHAPTER 1: Anthelmintic resistance in cattle nematodes
16
(FECR of 83%), despite the fact that MOX had never been used on this farm before. This might
suggest that the use of any type of MLs is inappropriate, once IVM-resistance has been
detected. On the other hand, fenbendazole (FEN), belonging to the BZ drug class, was still fully
(100%) effective on this farm [19].
Later, a new survey on a larger number of farms was conducted in order to make a better
estimation on the prevalence of anthelmintic resistance in Belgium and Germany (TABLE 1.2).
Of 88 farms included in this study, 84 farms used MLs. A FECR <95% was observed on 33 out
of the 84 farms (39%). Cooperia spp. were the most prevalent parasites after treatment.
However, using a Monte-Carlo simulation analysis, to correct for the used McMaster technique
with a detection limit of 50 EPG, reduced efficacies could only be confirmed on 25% of the
farms. Moreover, when four farms were revisited, only on 1 farm resistance against IVM could
be confirmed. These results showed that a reduced efficacy, observed with the FECRT, is not
only caused by anthelmintic resistance, but that the detection limit of the FEC technique used
and the (in)correct administration of the anthelmintic drugs are confounding factors of major
importance [18].
So far, emerging ML-resistance has only been reported for C. oncophora and not for the more
pathogenic O. ostertagi on Belgian cattle farms. Since C. oncophora is the dose-limiting species
for MLs, it is expected for resistance to appear first in this species. Moreover, Cooperia spp.
are predominantly parasites of younger cattle, as immunity to Cooperia spp. tends to develop
earlier than to O. ostertagi, for example. Consequently, anthelmintic programs tailored to treat
first year animals are likely to preferentially select for anthelmintic resistance in Cooperia spp.
[49, 77]. On cattle farms in Sweden and Germany, ML-resistance has been suspected in O.
ostertagi, so possibly, the existing levels of resistant O. ostertagi in Belgium are still below the
detection threshold [16].
1.4 Factors affecting the development of anthelmintic resistance
The development rate of anthelmintic resistance appears to be slow at first, but once a certain
level of resistance genes is established, the following treatments result in an exponential
increase of these resistance genes to a level where treatment failure occurs [2, 3]. Once
resistance is present in a parasite population, there is no evidence for reversion or loss of
resistance, although some (temporarily) effect of counter selection by another anthelmintic
drug cannot be dismissed [78, 79]. In parasites of sheep, the dynamics of selection for
anthelmintic resistance are well studied [80] and some predisposing factors are likely to be
similar in the nematode parasites of cattle [49]. These factors act either independently or in
an additive fashion and can be associated with the parasite species, the infected host, the
drug treatment, on-farm control management or the environment.

CHAPTER 1: Anthelmintic resistance in cattle nematodes
17
1.4.1 Parasite genetics and biology
Parasites in a population do not respond uniformly to treatment, this is due to their genetic
diversity. The high genetic diversity is linked to the huge population size and high reproduction
rate of parasites [81]. It is presumed that resistance alleles already exist within the parasite
population, prior to the first introduction with a drug [82]. But also, an alternative hypothesis
exists, suggesting there are multiple origins of resistance by spontaneous and recurrent
mutations [83]. Although the genetics of resistance are still poorly understood, resistance will
develop more quickly if only one gene is involved in resistance compared to the involvement of
multiple genes. Resistance will also develop faster if genes for resistance are dominant, rather
than recessive, because then, both heterozygote and homozygote worms will survive the
treatment and contribute to the next generation [84-86]. Furthermore, some parasites have
biological characteristics that favour resistance alleles to build up faster in the population, such
as their direct life cycles (no intermediate host), a short generation time and high fecundity. It
is assumed that, if resistant parasites would have an enhanced fitness or resistance is linked to
other fitness genes, then the spread of resistance in the population will also increase. Fitness
includes all those properties that enable more worms to complete their life cycle such as the
egg-laying rate, persistence of worms in the host (a reduced hypobiosis shortens their life
cycle), survival on the pasture, the ability to migrate on herbage and their infectivity when
ingested [69]. It has also been suggested that IVM-resistant C. oncophora in cattle became
more pathogenic than susceptible worms [15, 82].
1.4.2 Refugia and management factors
The larvae on pasture, the percentage of animals left untreated and the arrested larval stages
not affected by treatment of the host comprise the parasites in refugia. The proportion of
parasites in refugia should be high, but stay optimal, in order to dilute the resistance genes in
the pool of susceptible genes, and hence delaying the development of anthelmintic resistance.
Together, the parasites in refugia, the frequency of anthelmintic treatment and the extent of
under-dosing are mainly responsible for inducing anthelmintic resistance [87]. To decrease the
selection pressure, it is of major importance that treatment and pasture management are
fulfilled in ways that maintain refugia. Anthelmintic treatments should progress according to a
strategic plan, where frequency, time of treatment and selective treatment of first year or
infected animals are tightly followed. Short-interval treatments that approach the pre-patent
period for the parasite reduce the opportunities for susceptible worms to reproduce and
diminish the parasites in refugia. On farms with an intensive breeding and/or grazing program,
calves are given multiple treatments and are grazed away from the adults, hence, pasture
contamination derives from worms surviving more frequent treatments, this creates a very
high selection pressure for anthelmintic resistance to develop [88]. Therefore, it is encouraged
to implement an alternate grazing system, where calves are allowed to graze on pastures used

CHAPTER 1: Anthelmintic resistance in cattle nematodes
18
the previous year by older animals [69]. It should also be avoided to treat animals and then,
moving them immediately to a clean pasture. If doing so, the contamination of the new pasture
will only be attributed to a subpopulation that is resistant to the treatment. In this light,
farmers should be aware that summer drought is a variable factor that clears out the free-
living stages on pasture [88]. Additionally, bought-in animals should be effectively quarantine
drenched before they are placed on dirty pasture, to dilute out the progeny of any survivors of
the quarantine treatment [89].
1.4.3 Sub-therapeutic drug levels
To ensure that treatments are fully efficacious it is important to first weigh a cohort of the
heaviest animals – if not all animals – and hence the anthelmintic drug can be given at the
correct therapeutic dose level [90]. Sub-therapeutic concentrations will allow more worms to
survive the treatment and increase the development rate of resistance. A reduced
bioavailability of the drug has been associated with the route of administration and the type of
animal. Especially the inconsistent performance of topical (pour-on) applications has been
questioned as a predisposing factor for resistance [91]. Also, an enhanced drug metabolism
for some types of animals or breeds (such as described for goats and Belgian Blue cattle [61,
81]) can contribute to selection for resistance. The selection pressure on anthelmintic
resistance to develop is also affected by the pharmacokinetics of the drug. With the use of
persistent (long-acting) or slow release drugs, the drug concentrations tail off slowly towards
the end of their elimination phase as a result from an extended half-life. This effect will have
the same influence as under-dosing animals and therefore it is preferable to use short-acting
drugs [18, 49, 62, 82, 92].
1.5 Macrocyclic lactones
The avermectins (ivermectin, abamectin, doramectin and eprinomectin) and milbemycins
(moxidectin) are 2 subclasses within the ML family. Both chemical groups are produced
through fermentation by the soil-dwelling actinomycetes from the genus Streptomyces and
have a broad spectrum of anti-parasitic properties. All MLs share a 16-membered macrocyclic
backbone, but structurally differ from each other by the presence of substituents and/or
double carbon bonds. Naturally produced metabolites are milbemycin A3, A4, D and avermectin
A1a, A1b, A2a, A2b, B1a, B1b, B2a, B2b. IVM (22,23-dihydro-avermectin B1) is a semi-synthetic
derivate of a mixture of avermectin B1a and B1b. MLs are administered principally to control
gastro-intestinal nematode parasites, but also to assist in the control of several ecto-parasites
including ticks, mites and lice. However, MLs are ineffective against flatworms and tapeworms
[81, 93].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
19
1.5.1 Pharmacokinetics
MLs are characterized by their high lipophilicity, which results in a wide distribution and a good
absorption of the compounds and enables them to concentrate particularly in adipose tissue.
The slow release of the drug from these lipid reservoirs prolongs the residence of the drug in
the bloodstream. The concentration of drug residues is highest in liver and fat and lowest in
the brain [81]. Also, there is a good correlation between the IVM plasma concentration and the
IVM concentration measured in the parasite’s target tissue, such as mucosal tissue, skin and
lung. MLs are poorly metabolized and the major clearance pathway is through biliary secretion.
Consequently, faecal excretion accounts for 90% of the dose administered, with less than 2%
of the dose excreted in urine. Following subcutaneous dosing, ivermectin levels in cattle faeces
peak within 3-5 days. Moxidectin residues are excreted more slowly and peak excretion
(>58%) in cattle faeces persist for more than 28 days. The higher the lipophilicity of the MLs,
the higher the concentration secreted in milk. For example, moxidectin pour-on and
eprinomectin have no milk withholding time and can be used in lactating cattle [81, 94].
The exceptional potency of ML compounds makes it possible to use very low doses. MLs are
administered to cattle as oral drench, topical or injectable formulations. The delivery routes
and formulation strongly affect the systemic availability, as well as the availability in the
gastrointestinal tract, where several endoparasites are found. The greatest bioavailability is
achieved with injection then by the oral route and followed by topical administration (pour-on).
Parenteral administration (injectable and topical) delays absorption compared to the oral route.
Absorption is faster and results in a higher peak plasma concentration in an aqueous vehicle
compared to an oil-based vehicle. Concerning topical formulations, their systemic bioavailability
is low and does not exceed 15% of that for subcutaneous injection in cattle [62, 81, 95].
However, the animal’s licking and grooming behavior can affect the bioavailability of the drug.
Pour-on administration of IVM resulted in a lower systemic availability when licking was
prevented, but with licking, a substantial amount of topically applied IVM could also access the
systemic circulation via oral consumption [91, 96, 97]. Orally administered MLs associate with
particulate digesta in the rumen that delay the rate of passage of the drug down the gastro-
intestinal tract. Although oral formulations achieve lower blood-level concentrations compared
to injectable formulations, they reach considerable concentrations at the gastrointestinal level.
Therefore, it is not surprising that oral formulations of MLs obtain the highest efficacy against
gastrointestinal nematodes [98-100]. The anthelmintic action depends on the ability of active
drug to reach its specific receptor within the target species Subsequently, drug entry and
accumulation in the parasite are important issues to achieve an optimal clinical efficacy.
Passive transcuticular drug transfer is the predominantly entry mechanism and depends on the
lipophilicity of the anthelmintic [101].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
20
The systemic availability of a drug can also be affected by several host-related matters,
including the animal species, breed differences, metabolism, body composition, diet intake and
body condition. For example, reducing the feed intake prior to an oral or injectable treatment
will prolong drug absorption or will modify the exchange pattern between plasma and fat
reservoirs, resulting in a greater plasma availability and increasing anthelmintic efficacy. The
lower fat content of Belgian Blue carcasses enables a subcutaneously injected drug to reach
the systemic circulation directly, whereas in Holstein calves, the drug will accumulate in the
subcutaneous fat reservoirs. As a result, a significantly higher area under the plasma
concentration-time curve and a higher maximal plasma concentration is recorded in the Belgian
Blue breed [61, 81].
1.5.2 Effects on nematodes and mode of action
MLs can paralyse the pharynx, the somatic muscles and/or the uterus of parasites. Inhibition of
the pharyngeal pumping leads to worm death, due to starvation. From a recent study with the
sheep nematode H. contortus it appears that the somatic musculature is a more important
target site for abamectin and most likely for ML drugs in general [102]. For susceptible worms,
the EC50 values were 10-fold higher in the feeding assay compared to the EC50 values from the
motility assay. However, paralysis of the body-wall muscles was only restricted to the mid-
body of the worm. As a result of their reduced mobility, the removal of parasitic worms from
the gastro-intestinal tract of the host will be facilitated. Further, by inhibiting the uterine
muscles in female worms, the release of eggs already present in the uterus and/or the
production of new eggs can be suppressed by ML treatment. In adult filarial nematodes (e.g.
Onchocerca volvulus), suppression of the new microfilariae production is the most important
effect of MLs. It is important to realize that each of these major effects may differ between
species and developmental stages as they might have different sensitivities towards the
anthelmintic. Moreover, the ability of the ML to reach a particular site of action (or multiple
sites), can explain different anthelmintic effects [103-106].
FIGURE 1.1: The main effects of macrocyclic lactones on nematodes: paralysis of the pharynx, somatic
muscles and uterine muscles. From [107].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
21
MLs exert their paralysing effect by binding the glutamate-gated chloride channels (GluCls).
The irreversible activation of these receptors by an ML, evokes an irreversible increase in
chloride ion uptake, this induces the hyperpolarization of nerve cell membranes and leads to
the paralysis of the parasite. MLs can interact with a wide variety of ligand-gated anion and
cation channels, though the GluCls are thought to be the main targets. This was recognized
after an experiment in which total mRNA, isolated from Caenorhabditis elegans, was expressed
in Xenopus laevis oocytes. Subsequently, using the micro-electrode voltage clamp technique, it
was demonstrated that avermectin-sensitive currents were sensitive to glutamate and that
avermectin potentiated the effects of glutamate. MLs have the ability to kill parasites without
affecting the mammalian host, probably because the GluCl receptors are uniquely found in
invertebrates, and because MLs do not cross (exceptions in breeds of mice and dogs) the
blood-brain barrier to reach putative receptors in the central nervous system of the host
(FIGURE 1.1) [106, 108-110].
1.5.3 Structure and localization of the receptor
The GluCl receptors belong to the superfamily of Cys-loop ligand-gated ion channels. Within
the Cys-loop family, 2 broad categories can be distinguished: the cation-selective or excitatory
members (e.g. nicotinic acetylcholine and serotonin receptors) and the anion-selective or
inhibitory members (e.g. γ-aminobutyric acid, glycine, histamine and the GluCl receptors). As
for all members of this superfamily, the native GluCl receptor is presumed to be a pentamer,
made up of 5 subunits that are arranged together around a central pore. The individual subunit
sequences are characterized by: a long N-terminal extracellular domain, 4 membrane-spanning
domains, a long intracellular loop between the third and fourth membrane-spanning domain and
a short extracellular C-terminal. The N-terminal fragments of all subunits constitute the ligand-
binding site and the second membrane-spanning domain is the main contributor of the central
pore. The ‘Cys-loop’ term comes from the conservative loop of 13 residues between 2
cysteine residues in the extracellular N-terminal domain (FIGURE 1.2) [106, 110-112].
The first GluCl subunits to be cloned were the GluClα1 and GluClβ subunits of C. elegans,
encoded by Cel-glc-1 and Cel-glc-2, respectively. In the meantime, 4 more genes have been
identified that encode GluCl α-type subunits in C. elegans: glc-3, glc-4, avr-14 and avr-15. At
least 2 of these genes, Cel-avr-14 and Cel-avr-15, are alternatively spliced to yield 2 different
subunits (AVR-14A, AVR-14B, AVR-15A and AVR-15B). The properties of the different C.
elegans GluCl subunits have been examined by expression in X. laevis oocytes. Homomeric
channels of GluClα1 (GLC-1), GLC-3, AVR-14B, AVR-15A and AVR-15B were all sensitive to
IVM. Concerning the AVR-14A subunit, it remained unclear if the unresponsiveness to either
IVM or glutamate was due to a lack of sensitivity or to the inability to associate into
homomeric channels. GluClβ (GLC-2) homomers were activated by glutamate, but insensitive to
IVM. Interestingly, the co-expression of GluClα1/GluClβ subunits, AVR-14B/GluClβ subunits,

CHAPTER 1: Anthelmintic resistance in cattle nematodes
22
AVR-15A/GluClβ subunits and AVR-15B/GluClβ subunits resulted in the formation of
heteromeric channels that were sensitive to IVM [106, 113-118]. Different channels might
have different affinities for IVM or MOX, although for both drugs the same binding sites have
been found, there appears to be a difference in response to the 2 drugs in terms of larval
development rate, pharyngeal pumping and mobility [119]. Reporter gene experiments in C.
elegans demonstrated that both GluClβ and AVR-15 subunits were expressed in pharyngeal
muscle cells and furthermore, AVR-15 subunits were dispersed over the motor nervous system
[114, 120]. Gene expression of Cel-avr-14B was detected in extra-pharyngeal neurones in the
head, sensory neurons and ventral cord motor neurones [121, 122]. Cel-glc-1 appears in the
extra-pharyngeal neurones as well [115]. The expression sites of Cel-glc-3 and Cel-glc-4 are
still unknown. Anyway, the observed inhibition of pharyngeal pumping and mobility in C.
elegans is supported by the presence of IVM-sensitive GluCl subunits on the pharyngeal muscle
and motor neurones, which innervate somatic muscles.
Most research on parasitic GluCl receptors has been done on H. contortus. So far, 6 genes
encoding 7 GluCl subunits have been identified in H. contortus. No orthologous of Cel-glc-1 and
Cel-avr-15 have been identified in H. contortus. But, vice versa, Hco-glc-5 and Hco-glc-6
appear to encode for parasite-specific GluCl subunits [106, 123, 124]. In conclusion, GluCl
receptors can have different sensitivities, different sites of expression and different effects on
nematodes due to several possible subunit combinations. Furthermore, the GluCl subunits are
not completely conserved between nematode species. This all together, hampers the
understanding of the action mechanisms of MLs and has important implications for the
development of resistance.
FIGURE 1.2: Schematic representation of a glutamate-gated chloride channel. The macrocyclic lactone (ML)
compound binds to the receptor channel and causes an influx of chloride ions resulting in an irreversible
hyperpolarisation. Adapted from [125].
Chloride ions!
ML!
Outside cell!
Inside cell!
Cell membrane! ML! ML! ML!

CHAPTER 1: Anthelmintic resistance in cattle nematodes
23
1.6 Mechanisms of macrocyclic lactone resistance
A thorough understanding of the mechanisms by which a parasitic nematode develops
anthelmintic resistance is indispensable for the development of a more sensitive and reliable
molecular detection technique. With such a test, an early detection of anthelmintic resistance
is aimed, in order to restrict and circumvent the problem. Parasites can use specific
mechanisms, which involve the drug target, and/or unspecific mechanisms, which alter the
drug concentration, to become resistant. Examples of strategies are: � A change in the
molecular target, due to e.g. a mutation, making the binding site unrecognizable for the drug;
� Up- or down-regulation of target genes to overcome or avert drug action; � Enhanced
removal of the drug from the target site by a change in detoxification efflux pumps such as P-
glycoproteins; and � An alteration in the metabolism of the drug, e.g. through cytochrome
P450, that inactivates the drug or impedes its activation [82, 126]. Theoretically, drugs
belonging to the same anthelmintic family, share their mode of action, and therefore possibly
give rise to the same specific mechanism of resistance. However, if this side-resistance is
achieved by an unspecific strategy, drugs with the same mode of action will not automatically
give rise to the same level of resistance, because of the unspecific and diversified contribution.
Cross-resistance, between drugs that have different modes of action, will be achieved through
an unspecific, receptor-independent mechanism of resistance [127, 128]. Genes involved in
specific (GluCl receptors) and unspecific (P-glycoproteins) ML-resistance mechanisms are
addressed in the following paragraphs.
1.6.1 Glutamate-gated chloride channels
Molecular genetic work on C. elegans has demonstrated that it requires the simultaneous
mutation of at least 3 genes (Cel-glc-1, Cel-avr-14 and Cel-avr-15) encoding GluCl α-type
subunits, before high-level (4000-fold) resistance to ivermectin is achieved. In contrast, double
mutants in any two of these genes led to only modest resistance (7- to 10–fold) and null
mutations of the genes individually did not confer to resistance [115]. More recently, a
naturally occurring deletion (only 4 amino acids) in the ligand-binding domain of Cel-glc-1 has
been identified, which confers resistance to avermectins [129]. However, it is likely that many
of the resistant C. elegans mutants have dysfunctions, which would be lethal in parasitic
nematodes, and thus be irrelevant to resistance in parasitic nematodes. One gene, avr-14 in
particular, seems to be widely conserved in different parasitic nematode species of ruminants,
such as C. oncophora, O. ostertagi, H. contortus and T. circumcincta.
The genetic variability of Con-avr-14B and Con-glc-2 (encoding for an α- and β-type GluCl
subunit) was compared between an IVM-susceptible and IVM-resistant C. oncophora isolate
from the UK, using single-strand conformation polymorphism. Con-avr-14B could be associated
with IVM-resistance, while no difference in allele frequencies was observed for Con-glc-2 [130].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
24
By cloning the full-length Con-avr-14B and Con-glc-2 sequences from IVM-susceptible and IVM-
resistant C. oncophora, 3 mutations in Con-avr-14B (E114G, V235A and L256F) were
identified in the N-terminal extracellular domain of IVM-resistant worms. After expression in X.
laevis oocytes, whole-cell current recordings demonstrated that only the L256F mutation in
Con-avr-14B accounted for a loss (2- to 3-fold) in sensitivity to glutamate, IVM and MOX
[131]. Recently, the L256F polymorphism in the avr-14B gene was investigated in Belgian C.
oncophora and O. ostertagi isolates. Unfortunately, the L256F mutation appeared to be absent
in IVM-resistant isolates of both parasites. Still, a loss in allelic diversity of the Con-avr-14B
gene could be observed in the IVM-resistant isolate, compared to the susceptible isolate.
Additionally, transcription levels of avr-14B were significantly lower in male and female worms
of the IVM-resistant C. oncophora and O. ostertagi isolates relative to the susceptible ones
[132]. Similar results were observed for avr-14B in T. circumcincta. Pyrosequencing analysis
also failed to detect the presence of the L256F mutation in multiple-resistant T. circumcincta
isolates, although there was a significant change in allele frequencies following IVM exposure
[133].
In H. contortus, avr14B was expressed in X. laevis oocytes, carrying the candidate mutations
(E114G, V235A and L256F). As with the C. oncophora strain from the UK, electrophysiological
recordings showed that only the L256F mutation in Hco-avr-14B caused a significant loss (3-
to 6.5-fold) of sensitivity for glutamate and IVM [134]. Whether this mutation is present or
not, it remains a candidate polymorphism associated with IVM-resistance. Other genes,
encoding GluCl subunits in H. contortus, which have been associated with IVM-resistance, are
Hco-glc-3, Hco-glc-5 and Hco-glc-6. Their resistance-associated changes were either
demonstrated in mRNA transcription profiles, allele frequencies or reporter gene assays [116,
123, 135-141].
1.6.2 P-glycoproteins
One of the receptor-independent mechanisms of ML-resistance includes an increased efflux of
the drug by P-glycoproteins (PGPs). PGPs are large integrated membrane proteins belonging to
the superfamily of ATP-binding cassette (ABC) transporters, which carry diverse substrates
from the inside to the outside of cells. The ABC transporter family plays an important role in
the absorption, distribution, metabolism and elimination of xenobiotic compounds, both inside
the host and inside the parasite. Other members of the ABC transporter family include the
Half-transporters (HAFs) and the Multidrug resistant proteins (MRPs). PGPs are composed of 2
homologous halves, each half contains six membrane-spanning α-helices and a cytoplasmic
nucleotide-binding domain (NBD), which includes an ATP-binding site. A linker region separates
both halves. The membrane-spanning regions form the membrane channel that acts as ligand-
binding site. The NBDs are the motor domains of ABC-transporters and contain the highly
conserved Walker A, Walker B and ABC signature (LSGGQ) motif (FIGURE 1.3) [127, 142-144].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
25
The observation of PGPs being involved in the efflux of IVM was first discovered in mammals. In
a pgp-knockout mouse line and PGP-deficient Collie dogs, an enhanced sensitivity to IVM was
demonstrated, resulting in extreme neurotoxicity followed by death. Thus, PGPs in the blood
brain barrier offer protection against accumulation of toxic drug concentrations reaching the
central nervous system. Furthermore IVM and to a lesser extent MOX have been proven to be
excellent substrates for PGPs in mammals [145-148]. In this light, research on similar defence
mechanisms in parasites was started. So far in C. elegans 15 pgp genes, 9 haf genes and 8
mrp genes have been identified. In parasitic nematode species of ruminants, the number of
identified ABC transporter genes is still expanding. Until now, 9 pgp genes, 1 haf gene and 2
mrp genes have been described in H. contortus and recently, 11 partial pgp sequences were
identified in T. circumcincta [140, 144, 149-151].
FIGURE 1.3: The role of P-glycoprotein (PGP) in macrocyclic lactone (ML) efflux from the cell and a model
for transmembrane topology of PGP. (A) PGP actively pumps out the MLs against a concentration gradient.
(B) When overexpressed, such as in case of ML-resistance, PGP inhibits the ML compounds from reaching
their site of action and limits their efficacy. (C) With the use of PGP-inhibitors, PGP function is blocked and
the ML compounds can accumulate in the cell. Adapted from [144].
Transmembrane domains!Nucleotide binding domains!Active efflux!Macrocyclic lactones!Efflux inhibitor!
Key:!
A! B! C!
Transmembrane domains 1-6! Transmembrane domains 7-12!
N! C! ATP site! ATP site!
Nucleotide binding domain 1! Nucleotide binding domain 2!
"
#$!#$!#$! #$!#$! #$!
#$!#$! #$!
#$!

CHAPTER 1: Anthelmintic resistance in cattle nematodes
26
P-glycoprotein A, encoded by pgp-2, in H. contortus was the first ABC transporter found to be
associated with ML-resistance in parasitic nematodes. A higher mRNA transcription and
changes in allelic diversity were observed for Hco-pgp-2 in IVM-selected worms compared to
susceptible worms [152, 153]. More recently, again Hco-pgp-2 and Hco-pgp-9 showed an
increased expression level in a triple-resistant H. contortus isolate, relative to the susceptible
isolate [140]. The mRNA transcription level of pgp-9 was also up-regulated in a triple-resistant
T. circumcincta isolate, in comparison with a susceptible isolate [149]. In addition to the
changes in constitutive gene transcription, also induced changes have been observed after
exposure to MLs. An inducible overexpression of Hco-pgp-A(2), -B, -C, -D and –E was shown in
an IVM-resistant isolate following in vivo exposure to IVM. MOX exposure in vivo induced
overexpression in only Hco-pgp-C and –E. [154]. In vitro exposure of IVM-resistant and MOX-
resistant C. elegans isolates, either generated by receptor knockdown or through step-wise
exposure to non-lethal doses of IVM, was associated with an inducible overexpression of pgp-1,
pgp-2, pgp-4, pgp-12, pgp-14, mrp-1, mrp-2, mrp-3, mrp-4, mrp-5, mrp-6, mrp-7, mrp-8, haf-
1, haf-2 and haf-3 [155-157]. Furthermore, the in vivo administration of ABC transporter-
inhibitors (e.g. verapamil, ketoconazole, loperamide,…) in combination with an anthelmintic
drug seems to improve the pharmacokinetics and efficacy of the drugs in different infected
hosts [158-164]. PGP-inhibitors have also demonstrated to restore the susceptibility of
resistant H. contortus, T. circumcincta and C. elegans isolates to MLs in in vitro assays [145,
156, 165]. Reversal of resistance by such interfering agents, confirm the involvement of ABC
transporters in (multi)-drug-resistant worms (FIGURE 1.3).
1.6.3 Other candidate genes
Increasing evidence suggests that resistance is often the result of changes in genes other than
the immediate drug targets, which are the GluCl receptors for MLs. For example, in H.
contortus, the lgc-37 gene (previously known as HG1) encodes for a subunit of a GABA-gated
chloride channel and has been shown to be under selection of MLs. Significant differences in
allele frequencies were detected between unselected and IVM- and MOX-selected H. contortus
strains. Subsequently, an unselected and IVM-selected allele, differing in 4 amino acids, were
expressed in X. laevis oocytes. Electrophysiological recordings demonstrated that IVM
increased the GABA response in cells transfected with the susceptible allele and IVM
attenuated the GABA response in cell transfected with the resistant allele. A substitution of
K169R was found to reduce the sensitivity to GABA and MOX [166-168]. Another
neurotransmitter receptor that recently has been linked to ML-resistance is a dopamine-gated
anion channel, encoded by the H. contortus ggr-3 gene. Hco-ggr-3 is significantly down-
regulated in ML-selected strains of H. contortus. Moreover, a single nucleotide polymorphism in
the 3’ un-translated region appears to be associated with ML-selection [169].

CHAPTER 1: Anthelmintic resistance in cattle nematodes
27
An intriguing further observation is that IVM-resistant H.contortus worms have defects in the
amphids (sensory organs in the nematode head that contain the sensory neurones). Since
GluCl receptors are expressed in amphid and extra-pharyngeal neurones [122], derangements
in the microtubules of these neurones might prevent the drug from accessing its target sites
[170]. As a consequence, repeated use of MLs might predispose parasitic nematodes to BZ-
resistance, as the main target for BZs is β-tubulin [171-173]. Further, the amphid and extra-
pharyngeal neurones are connected to the pharyngeal muscle cells via linking neurones, in
which the gap junction annexins convey the IVM-induced hyperpolarisation to the pharynx. In
the absence of annexins (encoded by unc-7 and unc-9 in C. elegans), IVM-toxicity is restricted
to those cells expressing the targeted GluCl receptors, the hyperpolarisation signal is not
transferred and the worm survives the treatment. It has also been suggested that genes
mediating the cuticle permeability might be involved in ML-resistance. The amphid dye filing
(dyf) genes, such as osm-1 in C. elegans, might act additively to regulate IVM-uptake. When
Cel-osm-1 is mutated, a reduced effect of IVM is observed, probably due to a lower uptake of
the drug [115].
Alternatively, the drug might be subjected to detoxification through the cell’s natural
antioxidant defence enzymes, including glutathione-thioredoxin systems or cytochrome P450
enzymes. Generally, unique enzymes metabolize xenobiotics into more polar metabolites that
are easier to excrete. In H. contortus worms resistant to BZ treatment, an increased
glutathione S-transferase (GST) activity was reported compared to susceptible worms. Also,
when an inhibitor of glutathion synthesis was administered in combination with a BZ compound,
an increased sensitivity of the resistant H. contortus isolate was detected to this drug [126,
174-177]. Furthermore, IVM-resistance in H. contortus was recently dedicated to an increased
expression of several thioredoxins, which are essential for free radical scavenging [178].
However, further research is required to determine the role of xenobiotic metabolizing
enzymes in the development of drug-resistance.

CHAPTER 1: Anthelmintic resistance in cattle nematodes
28
1.7 Conclusion
After every treatment, resistance alleles will be enriched in the parasite population. But the
speed with which anthelmintic resistance develops is still unknown. Due to the irreversibility of
resistance it is of major importance to prolong the efficacy of the currently used drugs and to
intervene with the spread of resistance alleles. This is only possible if the recommended
treatment strategies in combination with advised pasture management are correctly applied
and if anthelmintic resistance can be detected accurately and in a very early stage. Lately,
some reports have emerged on resistance against MLs in the cattle nematodes C. oncophora
and O. ostertagi, using the FECRT. However, the FECRT is labour intensive, time-consuming,
not species-specific and detects resistance only when it is too late. A more sensitive,
molecular test is urgently needed. However, such a molecular test requires a better
understanding of the action mechanisms of the drugs and the genetic basis of ML-resistance.
Additionally, new insights in the molecular mechanisms of resistance could lead to the
identification of novel candidate drug-receptors. New anthelmintic drugs, with a novel mode of
action, could combat resistance against the currently available drugs.

OBJECTIVES

30

OBJECTIVES
31
OBJECTIVES
In Belgium, Cooperia oncophora is the most common cattle parasite in which resistance,
especially against MLs, occurs. In the field, the most commonly used method for diagnosing
ML-resistance is the insensitive FECRT. As described in CHAPTER 1, the FECRT has to be
employed with caution to avoid under-estimation of anthelmintic resistance. Therefore the
first objective of this PhD thesis was to evaluate the accuracy of the FECRT to assess
resistance against IVM and MOX in a C. oncophora field isolate, through comparison with the
reduction in worm burden after treatment (CET). Additionally, the required parasite material
for further molecular analyses was collected during this trial (CHAPTER 2).
The spread of resistance-alleles could be delayed with a sensitive molecular detection
technique for ML-resistance. But, a molecular test requires a genetic marker for resistance.
Molecular changes in GluCl receptors and ABC transporters are likely to play an important role
in the molecular mechanisms of ML-resistance. Therefore, the second objective was to
determine the C. oncophora transcriptome and identify members of the GluCl receptor and
ABC transporter families (CHAPTER 3).
At last, the third objective was to investigate the molecular changes in the identified GluCl
subunit and ABC transporter genes of C. oncophora between IVM-susceptible and IVM-resistant
isolates (CHAPTER 4 and CHAPTER 5).
In CHAPTER 6, it will be discussed whether the FECRT is still useful in the field, how this thesis
improved our knowledge on the molecular mechanisms of ML-resistance and how the results
can contribute in the development of a more sensitive detection technique for ML-resistance in
the field.

32

CHAPTER 2 Assessing resistance against macrocyclic lactones in gastro-
intestinal nematodes in cattle using the faecal egg count
reduction test and the controlled efficacy test

34
Based on: De Graef J, Sarre C, Mills BJ, Mahabir S, Casaert S, De Wilde N, Van Weyenberg M,
Geldhof P, Marchiondo A, Vercruysse J, Meeus P and Claerebout E. Assessing resistance against
macrocyclic lactones in gastro-intestinal nematodes in cattle using the faecal egg count
reduction test and the controlled efficacy test. Veterinary Parasitology. 2012 (189[2-4]:
378-382).

CHAPTER 2: Faecal egg count reduction test versus controlled efficacy test
35
2.1 Introduction
In cattle, in the majority of the anthelmintic resistance cases, resistance against the
macrocyclic lactones (MLs) has been reported. MLs are divided into two groups: avermectins
(ivermectin, abamectin, doramectin and eprinomectin) and milbemycins (moxidectin). Both
chemical groups are different in structure, but share a common mode of action [93]. Isolates
of the dose-limiting species Cooperia oncophora are mostly implicated in ML-resistance in
cattle, but cases of ML-resistant Ostertagia ostertagi are also emerging in temperate climate
regions [17, 25, 27].
In field conditions, the detection of anthelmintic resistance is usually based on the faecal egg
count reduction test (FECRT). However, the major limitation of this technique is its lack of
sensitivity, in addition, the reliability of the FECRT is strongly affected by the number of
animals per treatment group and the level of excretion and aggregation of the faecal egg
counts (FECs) [54]. Further, the correlation between FECs and worm numbers is not always so
clear, especially not in cattle, since the number of egg counts only reflects to the female worm
population and some species show a strong density-dependence in egg production [44, 55,
56, 179]. Moreover, ML treatment causes a (temporary) sterilising effect on the uterine
muscles of the parasite, which can result in an under-estimation of the ML-resistance problem
[44]. In addition, a reduced efficacy observed by FECRT is not always caused by anthelmintic
resistance, but can be due to confounding factors such as a sub-optimal treatment dosage
[18].
Due to these important drawbacks of the FECRT, the main objective of this study was to
evaluate the accuracy of the FECRT to assess the resistance status of C. oncophora and O.
ostertagi in cattle, using a controlled efficacy test as a reference. For this purpose, the
efficacy of ivermectin and moxidectin was evaluated in an IVM-resistant C. oncophora field
isolate [19] and an IVM-resistant laboratory isolate of O. ostertagi [90]. Further, possible side-
resistance against MOX was investigated in both IVM-resistant nematode isolates. In addition,
parasite material (eggs, third stage larvae and adult worms of C. oncophora) was recovered
before and/or after treatment of the animals for later molecular investigations (CHAPTER 4
and CHAPTER 5).
2.2 Materials and methods
2.2.1 Nematode isolates
An anthelmintic susceptible O. ostertagi population was previously selected for IVM-resistance
by repeatedly exposing the population to sub-therapeutic and therapeutic levels of IVM over
10 generations. In each selection round, a group of calves was infected with the progeny of
the previous IVM-selected O. ostertagi population. In the last selection round a therapeutic IVM

CHAPTER 2: Faecal egg count reduction test versus controlled efficacy test
36
dose (0.2 mg/kg bodyweight) only reduced the faecal egg counts by 57% and 65% on days 7
and 14 after treatment, respectively [90]. An IVM-resistant C. oncophora isolate was collected
from a Belgian farm in 2008. The presence of IVM-resistant C. oncophora worms on this farm
was first detected in 2006 [16]. During the following years, the FECR on day 21 post-
treatment decreased from 73% in 2006, over 40% in 2007, to 0% in 2008 [19]. Both isolates
are maintained in the laboratory by regular passages, without treatment, through helminth-free
calves. Infective third stage larvae (L3) were harvested from coprocultures of 14 days at
25°C, followed by Baermannisation [180].
2.2.2 Experimental design
Thirty male, 10-month old Holstein Friesian calves, individually weighing 177 kg – 258 kg, were
used in this trial. Animals were free of gastrointestinal helminth infections, as confirmed by
faecal egg counts. During the trial, animals were kept indoors to prevent infection with
parasitic nematodes and were individually tethered. The calves were fed hay and commercial
pellets, and given ad libitum access to water. Each calf was orally infected with 25,000 L3 the
IVM-resistant O. ostertagi isolate and 25,000 L3 of IVM-resistant C. oncophora, 28 days prior
to treatment (Day -28). On day -4, faecal samples were examined to confirm that all calves
had positive egg counts. The animals were randomly assigned to 3 treatment groups (n =
10/group). At day 0, the calves received a single dose of their assigned product, administered
as subcutaneous injections at the dose rate of 0.2 mg/kg bodyweight. The animals in group 1
received MOX (Cydectin® 1%, Pfizer), while those in group 2 received IVM (Ivomec® 1%, Merial)
and the calves in group 3 received a placebo treatment (0.9% sodium chloride). Fourteen and
15 days after treatment the animals were euthanized and nematode burdens were determined.
2.2.3 Parasitological techniques
Faecal samples were collected from the rectum of the animals on days -4, 0, 7 and 14 to
determine the FEC using a modified McMaster technique with a sensitivity of 50 eggs per gram
faeces. Coprocultures were made of the individual faecal samples taken at days 0, 7 and 14
post-treatment. Third stage larvae were collected by the Baermann technique and 100 larvae
were differentiated from each sample. O. ostertagi and C. oncophora egg counts were
estimated by multiplying the individual total egg count with the % Ostertagia spp. or the %
Cooperia spp. obtained from the faecal cultures.
The calves were randomly designated for necropsy on either day 14 or 15 post-treatment. The
abomasa and small intestines were recovered and processed according to the techniques
described by [46]. Two percent of the abomasal washings, the abomasal digests and the small
intestinal washings were analyzed to determine the total worm burdens (adults and inhibited
larval stages). Furthermore, 10 female C. oncophora worms were collected randomly per
animal and mounted on microscope slides for in utero egg counts.

CHAPTER 2: Faecal egg count reduction test versus controlled efficacy test
37
2.2.4 Determination of efficacy
The efficacy of the test compounds IVM and MOX was determined by calculating the faecal egg
count reduction through the formula proposed by the WAAVP guidelines [43]: %FECR = 100 x
(1 – T/C), where T and C are the arithmetic means of the FECs, respectively in the treated and
control group. Anthelmintic resistance was confirmed by a FECR lower than 95% and a 95%
confidence limit less than 90%. In the controlled slaughter test the efficacy of anthelmintic
treatment against O. ostertagi and C. oncophora was calculated using the same formula, where
T and C were defined by the arithmetic mean worm counts, respectively in the treated and
control group [46]. In addition, for worm counts the geometric means were also calculated and
used to determine efficacy, as would have been the case for the studies conducted to obtain
the original label claims [46]. The 95% confidence intervals for the reductions were calculated
using the RESO software as recommended by [43]. Treatments were compared with Dunn’s
multiple procedure in the Kruskal-Wallis test. If the overall treatment effect was statistically
significant (P<0.05), paired comparisons were made based on rank sums in the Mann-Whitney
test (GraphPad Prism® software, version 5.0c).
2.3 Results
Individual data for FECs on days 0, 7 and 14 post-treatment, % C. oncophora and O.ostertagi
larvae from coprocultures on days 0, 7 and 14 post-treatment, C. oncophora and O. ostertagi
worm burdens and mean in utero egg counts in 10 surviving female C. oncophora worms per
animal can be found in APPENDIX A.
2.3.1 Ostertagia ostertagi
The arithmetic means of the O. ostertagi egg and worm counts are presented in TABLE 2.1.
Prior to treatment, O. ostertagi egg counts were similar in the three treatment groups, with
means ranging from 213 to 245 EPG. On days 7 and 14 after IVM treatment, egg counts of O.
ostertagi were significantly lower in the treated animals compared to the control group
(P<0.05), with a reduction of 73% and 68% on days 7 and 14, respectively. O. ostertagi total
worm counts were reduced by 84% (geometric mean 89%) after IVM treatment and were
significantly lower in the IVM group than in the controls (P<0.05). MOX treatment resulted in
>99% reduction of O. ostertagi egg counts and worm counts (geometric mean >99%).
2.3.2 Cooperia oncophora
The arithmetic means of the C. oncophora egg and worm counts are presented in TABLE 2.2.
On day 0, mean C. oncophora egg counts varied, from 512 EPG to 755 EPG (P>0.05) between
treatment groups. On day 7 and 14 post-treatment, mean C. oncophora egg counts in the IVM
group were not significantly lower compared to the control group, with a reduction of 19% and
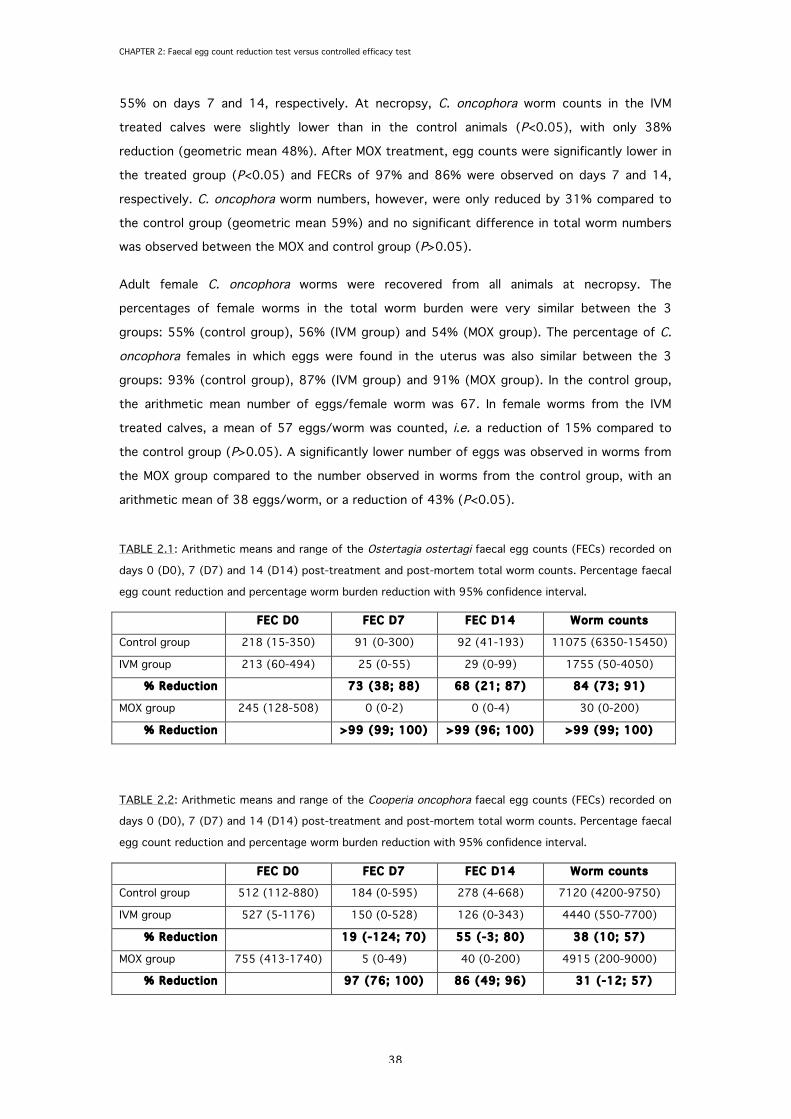
CHAPTER 2: Faecal egg count reduction test versus controlled efficacy test
38
55% on days 7 and 14, respectively. At necropsy, C. oncophora worm counts in the IVM
treated calves were slightly lower than in the control animals (P<0.05), with only 38%
reduction (geometric mean 48%). After MOX treatment, egg counts were significantly lower in
the treated group (P<0.05) and FECRs of 97% and 86% were observed on days 7 and 14,
respectively. C. oncophora worm numbers, however, were only reduced by 31% compared to
the control group (geometric mean 59%) and no significant difference in total worm numbers
was observed between the MOX and control group (P>0.05).
Adult female C. oncophora worms were recovered from all animals at necropsy. The
percentages of female worms in the total worm burden were very similar between the 3
groups: 55% (control group), 56% (IVM group) and 54% (MOX group). The percentage of C.
oncophora females in which eggs were found in the uterus was also similar between the 3
groups: 93% (control group), 87% (IVM group) and 91% (MOX group). In the control group,
the arithmetic mean number of eggs/female worm was 67. In female worms from the IVM
treated calves, a mean of 57 eggs/worm was counted, i.e. a reduction of 15% compared to
the control group (P>0.05). A significantly lower number of eggs was observed in worms from
the MOX group compared to the number observed in worms from the control group, with an
arithmetic mean of 38 eggs/worm, or a reduction of 43% (P<0.05).
TABLE 2.1: Arithmetic means and range of the Ostertagia ostertagi faecal egg counts (FECs) recorded on
days 0 (D0), 7 (D7) and 14 (D14) post-treatment and post-mortem total worm counts. Percentage faecal
egg count reduction and percentage worm burden reduction with 95% confidence interval.
FEC D0 FEC D7 FEC D14 Worm counts
Control group 218 (15-350) 91 (0-300) 92 (41-193) 11075 (6350-15450)
IVM group 213 (60-494) 25 (0-55) 29 (0-99) 1755 (50-4050)
% Reduction 73 (38; 88) 68 (21; 87) 84 (73; 91)
MOX group 245 (128-508) 0 (0-2) 0 (0-4) 30 (0-200)
% Reduction >99 (99; 100) >99 (96; 100) >99 (99; 100)
TABLE 2.2: Arithmetic means and range of the Cooperia oncophora faecal egg counts (FECs) recorded on
days 0 (D0), 7 (D7) and 14 (D14) post-treatment and post-mortem total worm counts. Percentage faecal
egg count reduction and percentage worm burden reduction with 95% confidence interval.
FEC D0 FEC D7 FEC D14 Worm counts
Control group 512 (112-880) 184 (0-595) 278 (4-668) 7120 (4200-9750)
IVM group 527 (5-1176) 150 (0-528) 126 (0-343) 4440 (550-7700)
% Reduction 19 (-124; 70) 55 (-3; 80) 38 (10; 57)
MOX group 755 (413-1740) 5 (0-49) 40 (0-200) 4915 (200-9000)
% Reduction 97 (76; 100) 86 (49; 96) 31 (-12; 57)
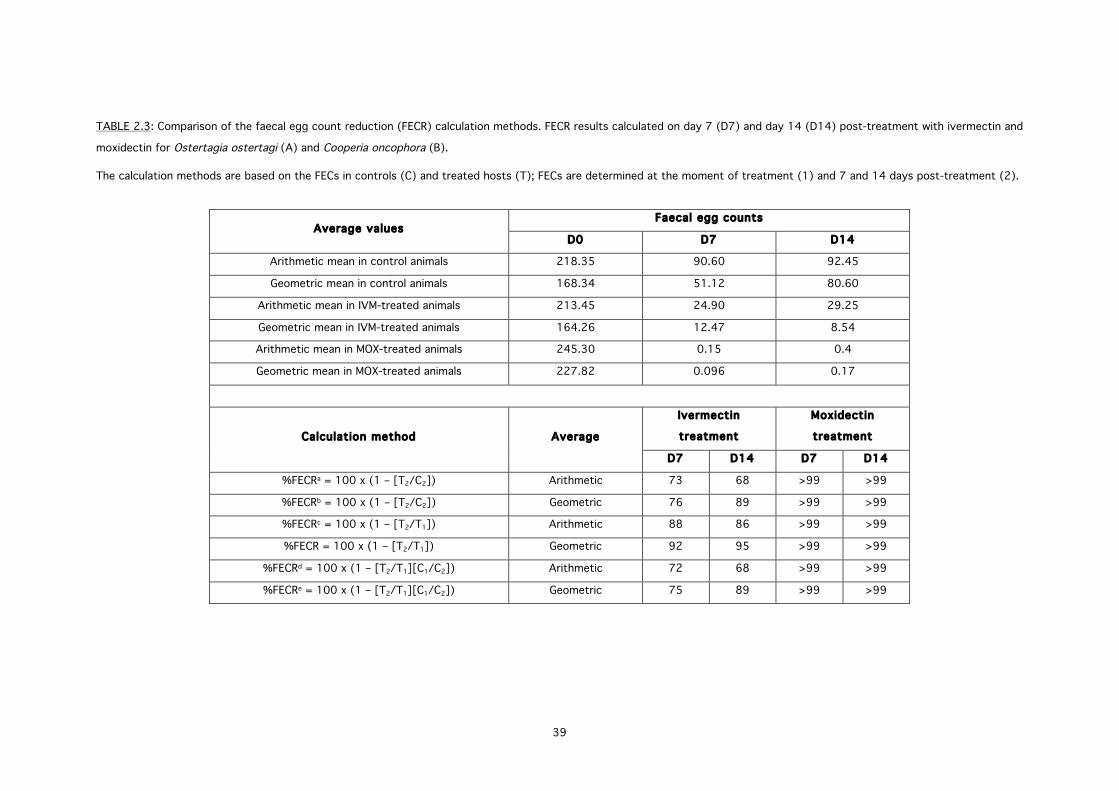
39
TABLE 2.3: Comparison of the faecal egg count reduction (FECR) calculation methods. FECR results calculated on day 7 (D7) and day 14 (D14) post-treatment with ivermectin and
moxidectin for Ostertagia ostertagi (A) and Cooperia oncophora (B).
The calculation methods are based on the FECs in controls (C) and treated hosts (T); FECs are determined at the moment of treatment (1) and 7 and 14 days post-treatment (2).
Average values Faecal egg counts
D0 D7 D14
Arithmetic mean in control animals 218.35 90.60 92.45
Geometric mean in control animals 168.34 51.12 80.60
Arithmetic mean in IVM-treated animals 213.45 24.90 29.25
Geometric mean in IVM-treated animals 164.26 12.47 8.54
Arithmetic mean in MOX-treated animals 245.30 0.15 0.4
Geometric mean in MOX-treated animals 227.82 0.096 0.17
Calculation method Average
Ivermectin
treatment
Moxidectin
treatment
D7 D14 D7 D14
%FECRa = 100 x (1 – [T2/C2]) Arithmetic 73 68 >99 >99
%FECRb = 100 x (1 – [T2/C2]) Geometric 76 89 >99 >99
%FECRc = 100 x (1 – [T2/T1]) Arithmetic 88 86 >99 >99
%FECR = 100 x (1 – [T2/T1]) Geometric 92 95 >99 >99
%FECRd = 100 x (1 – [T2/T1][C1/C2]) Arithmetic 72 68 >99 >99
%FECRe = 100 x (1 – [T2/T1][C1/C2]) Geometric 75 89 >99 >99

40
Average values Faecal egg counts
D0 D7 D14
Arithmetic mean in control animals 511.65 184.40 277.55
Geometric mean in control animals 436.29 69.73 156.99
Arithmetic mean in IVM-treated animals 526.55 150.10 125.75
Geometric mean in IVM-treated animals 325.06 40.98 34.69
Arithmetic mean in MOX-treated animals 754.70 4.85 39.60
Geometric mean in MOX-treated animals 695.40 0.48 3.26
Calculation method Average
Ivermectin
treatment
Moxidectin
treatment
D7 D14 D7 D14
%FECRa = 100 x (1 – [T2/C2]) Arithmetic 19 55 97 86
%FECR = 100 x (1 – [T2/C2]) Geometric 41 78 >99 98
%FECRb = 100 x (1 – [T2/T1]) Arithmetic 71 76 >99 95
%FECR = 100 x (1 – [T2/T1]) Geometric 87 89 >99 >99
%FECRc = 100 x (1 – [T2/T1][C1/C2]) Arithmetic 21 56 98 90
%FECRd = 100 x (1 – [T2/T1][C1/C2]) Geometric 21 70 >99 99
a Formula proposed by Coles et al. [43] b Formula proposed by Kochapakdee et al. [181] c Formula proposed by Dash et al. [58] d Formula proposed by Presidente [47]

CHAPTER 2: Faecal egg count reduction test versus controlled efficacy test
41
2.4 Discussion
Based on the FECRT, the O. ostertagi and C. oncophora isolates used in this trial, were
previously declared to be IVM-resistant [19, 90]. The FECRT results in the present experiment
supported the presence of IVM-resistance in both isolates with efficacies ranging from 73%
(95% CI: 38; 88) to 68% (95% CI: 21; 87) for O. ostertagi and 19% (95% CI: -124; 70) to
55% (95% CI: -3; 80) for C. oncophora, respectively at 7 and 14 days after IVM treatment. As
larval development and recovery of the larvae from the coprocultures might differ between
both species [182], a bias in the species-specific FEC cannot be excluded. However, the IVM-
resistant status of both isolates was also corroborated by the controlled efficacy test, with
reductions in worm burden of 84% (95% CI: 73; 91) for O. ostertagi and only 38% (95% CI:
10; 57) for C. oncophora, after IVM treatment. The obtained data suggest, at least with these
strains and in young animals with little acquired immunity, a good correlation between the
FECRT and the controlled efficacy test for assessing IVM efficacy in IVM-resistant cattle
parasites.
MOX showed a very high efficacy against the IVM-resistant O. ostertagi isolate, with 100%
reduction in FEC and worm counts. The FECRT suggested borderline resistance against MOX in
the IVM-resistant C. oncophora, with FECRs varying from 97% (95% CI: 76; 100) to 86%
(95% CI: 49; 96), respectively at 7 and 14 days after MOX treatment. However, failure of MOX
treatment was demonstrated more clearly by the controlled efficacy test. C. oncophora worm
counts in the MOX group were only 31% (95% CI: -12; 57) lower than in the control group,
indicating that most worms from the IVM-resistant C. oncophora isolate also survived the MOX
treatment. Side-resistance against MOX in this IVM-resistant C. oncophora isolate was already
suspected in 2008, when MOX treatment on this farm reduced FEC by only 83% [19], despite
the fact that MOX had never been used on this farm before. It has been proposed that IVM and
MOX share a common action mechanism, but the ligand-gated chloride channels they target
are diverse within the parasite and there are subtle differences in how these compounds
interact with the target [119, 183]. In addition, drug transporter P-glycoprotein may be
involved in ML-resistance, and IVM and MOX might have different affinity for these P-
glycoproteins [127, 145, 163]. These subtle differences may explain why MOX was still
effective against the IVM-resistant O. ostertagi isolate. The fact that side-resistance against
MOX was only observed in the IVM-resistant C. oncophora isolate, and not in the IVM-resistant
O. ostertagi, could also be due to a higher level of ML-resistance in the C. oncophora isolate. C.
oncophora is the dose limiting species for IVM [81], and consequently, IVM-resistance develops
more rapidly in this parasite. On the other hand, one should take into account that the IVM-
resistant O. ostertagi isolate was selected in the lab and is not necessarily representative for
IVM resistant field isolates.

CHAPTER 2: Faecal egg count reduction test versus controlled efficacy test
42
The discrepancy in the present study between the FECs and the worm counts for C. oncophora
in the MOX group can be explained by the reduced fecundity after MOX treatment and possibly
a reduced egg excretion. Apparently, MOX decreased the number of eggs in female worms that
survived the treatment, which was not the case in the IVM group. A significant difference in
the presence of eggs inside the uterus of Cooperia spp. after MOX treatment compared to
untreated controls was also demonstrated by [184]. From these results it appears that the
FECRT is not a reliable assay to detect MOX resistance, as some cases of MOX resistance in C.
oncophora may be overlooked, due to suppressed fecundity in MOX-treated worms. Although
generally a good correlation between the FECRT and the controlled efficacy test is observed in
isolates that are solidly resistant or susceptible to an anti-parasitic drug [184-186], poor
correlations between both tests have been reported in cases where anthelmintic resistance
was suspected [187]. This may have important consequences for anthelmintic resistance
surveys in the field, as the FECRT is the only practical test available at this moment. Several
studies in which a high efficacy of MOX treatment was reported in IVM-resistant nematodes
[188-190], were based on the FECRT only, and may have underestimated the prevalence of
MOX resistance. The FECRT has been criticised before, because of its low sensitivity and other
confounding factors, e.g. under-dosing [18]. In this case, however, improving the detection
limit (<50 eggs per gram) of the FECRT would probably not overcome the problem. According
to the WAAVP guidelines [43], faecal samples should be collected 14-17 days post-treatment
with MOX, but these results suggest that probably a longer interval is required, before the
efficacy of MOX can be evaluated by the FECRT.
Further, the results of this study demonstrate that the outcome of the FECRT can be
dependent on the mathematical technique used to analyse the data (TABLE 2.3). The
efficacies obtained with the FECRT using the methods of Coles et al. and Dash et al. were
closest to the results of the CET [43, 58]. These methods propose the use of arithmetic
means and include the values of the untreated control group. Geometric means were likely to
overestimate efficacy, owing to all positive pre-treatment counts, but some zero post-
treatment counts [59]. In the case of MOX, only the methods of Coles et al. and Dash et al.
declared (a low level of) resistance in the C. oncophora isolate 14 days post-treatment,
despite the low correlation with the CET.
In conclusion, IVM-resistance was confirmed by the FECRT and a controlled efficacy test in a
field isolate of C. oncophora and a lab isolate of O. ostertagi. While the IVM-resistant O.
ostertagi isolate was still susceptible to MOX treatment, side-resistance against MOX was
present in the IVM-resistant C. oncophora isolate. MOX-resistance in C. oncophora was not
irrefutably detected by the FECRT, due to suppression of the egg production in the resistant
female worms. To accurately determine MOX-resistance, the sensitivity of the FECRT is inferior
to the controlled efficacy test.
CHAPTER 3 Screening of the Cooperia oncophora transcriptome
database for candidate genes involved
in macrocyclic lactone resistance

44
Based on: Heizer E, Zarlenga DS, Gasser RB, De Graef J*, Geldhof P and Mitreva M.
Transcriptome analyses reveal protein and domain families that delineate stage-related
development in the economically important parasitic nematodes, Ostertagia ostertagi and
Cooperia oncophora. BMC Genomics. 2012 (submitted).
* Contribution: Providing total RNA from the life cycle stages of C. oncophora and O. ostertagi.

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
45
3.1 Introduction
Molecular changes in drug target genes or genes that are suspected of being involved in drug
efflux or drug metabolism are likely to play an important role in the underlying genetic
mechanisms of anthelmintic resistance [154, 166]. In order to develop a sensitive molecular
tool for diagnosing macrocyclic lactone (ML)-resistance in cattle nematodes, the search for the
genuine resistance markers continues. Thinking of a candidate gene approach, a detailed
exploration of the transcriptome of the cattle nematode Cooperia oncophora would elucidate
the extent of the glutamate-gated chloride (GluCl) channel receptor and ABC transporter
families in this parasite. However, currently no transcriptomic or genomic data is available at all
for C. oncophora.
The present study [191] has generated extensive information on the transcriptome of C.
oncophora for all developmental stages, representing established gene expression patterns in
the entire life cycle. In the next paragraphs, the results of the transcript assemblies for C.
oncophora will only be cited briefly, whereas most attention will be paid to the in-depth
analysis of the identified candidate ML-resistance genes, i.e. those encoding for GluCl channel
subunits and ABC transporters, from the C. oncophora transcriptome dataset.
3.2 Materials and methods
3.2.1 Parasite material and RNA extraction
The anthelmintic-susceptible C. oncophora population used in this study has never been
exposed to drug treatments and is maintained in the Laboratory for Veterinary Parasitology at
the Ghent University by regular passages through helminth-free calves [132]. Eggs were
purified from the faeces of artificially infected calves by a centrifugation-flotation method and
finally collected on a 38 μm sieve. To collect first-stage larvae (L1), eggs were incubated for
16 h at 28°C in deionized water. The hatched L1 were purified by Baermannisation. Second-
and third- stage larvae (L2 and L3) were collected by culturing the faeces at 25°C for 72 h
and 14 days, respectively, followed by Baermannisation. Ex-sheathed L3 (xL3) were obtained
by adding 0.5% of sodium hypochlorite (in distilled water) for 15 min [64]. Fourth-stage larvae
(L4) were obtained from animals euthanized 8 days post-infection, hereby, intestinal contents
and washings were poured over a 116 μm sieve, L4 were retained and then placed on a
Baermann apparatus. Similarly, adult worms were recovered live at necropsy at 21 days post-
infection and microscopically further partitioned into male (M) and female (F) worms. Total
RNA was prepared by grinding the parasite samples (pellets of at least 100 adults, 1000 larvae
and 50000 eggs) on ice in 0.2 ml glass homogenizers (Wheaton), followed by the Trizol®
(Gibco Invitrogen) method according to the manufacturer’s recommendations. Residual
genomic DNA was removed by DNase I treatment (Roche). The RNA integrity was verified with

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
46
the Experion™ RNA StdSens starter kit (Bio-Rad) and the RNA concentration was determined
using a Nanodrop® ND-1000 spectrophotometer (NanoDrop Technologies).
3.2.2 ‘Next-generation’ sequencing and processing of the reads
Library construction was based on the SMART cDNA library construction (Clontech
Laboratories). Then, the cDNA library fragments were immobilized onto DNA capture beads,
emulsified and subjected to PCR in order to amplify the DNA template. Further, the emulsion
was chemically broken and the DNA library beads were recovered. Subsequently, DNA library
beads were loaded onto a PicoTiterPlate device and sequenced on the Genome Sequencer 454
Titanium instrument using the GS FLX titanium Sequencing kit (Roche 454) [191]. The
assembly of the sequencing reads was carried out using the Newbler assembler v2.5
runMapping software and for the clustering of the reads the cd-hit-est was used at 99%
identity [192]. Next, the reads were mapped to the PHRAP assembly (http://phrap.org) for
expression profiling. Utilizing prot4est, assembled contigs and isotigs were translated [193].
Predicted polypeptides were compared to the core eukaryote genes (CEGs) to estimate the
completeness of the C. oncophora transcriptome. Then, predicted polypeptides were further
analysed with InterProScan, using tags to search for InterPro domains, GO terms and Pfam
domains [194]. Finally, functional classification was carried out on the contigs, based on the
homology with the KEGG (Kyoto Encyclopaedia of Genes and Genomes) database. Assembled
contigs and isotigs are now available for acquisition and searching at http://nematode.net
[195].
3.2.3 Identification of candidate resistance genes
The data-mining portal NemaBLAST (available at http://nematode.net) was used to blast
protein query sequences of C. elegans and H. contortus genes, encoding subunits of the GluCl
receptor and members of the ABC transporter family, versus EST contigs and genes of C.
oncophora. Partial sequence information of the matching C. oncophora GluCl receptor subunits
and ABC transporters was requested from the NemaGene portal. Subsequently, gene-specific
primers were designed using the online Primer3 software (http://frodo.we.mit.edu/primer3).
cDNA was synthesized from 1 μg total RNA from C. oncophora adults by random priming using
the iScript cDNA synthesis kit (Bio-Rad) according to the manufacturer’s recommendations.
PCR amplifications were carried out in a 25 μl reaction volume: containing1 μl of adult cDNA
template, 1 μM of each primer, 0.4 mM of each dNTP, 1.4 mM MgCl2, 1 unit GoTaq® DNA
polymerase (Promega), 5 μl of 5x GoTaq® Buffer (Promega) and ultra pure water. All reactions
were run as follows: 2 min at 95°C, followed by 35 cycles of 95°C for 30 s, 60°C for 30 s and
72°C for 30 s, followed by a final extension at 72°C for 10 min and then hold on 10°C. PCR
products were visualized on a 1.5% agarose gel and stained with 0.5 μg/ml ethidium bromide.
Bands were excised and purified with the Geneclean kit® (MPBio). Purified PCR products were

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
47
cloned using the pGEM®-T easy vector (Promega) and Escherichia coli DH5α competent cells
(Stratagene) according to the manufacturer’s protocols. Plasmid products were sequenced
bidirectional with SP6 and T7 vector primers. The sequences were analysed with DNASTAR
software (Lasergene version 8). To assign the C. oncophora candidate resistance genes with
the correct nomenclature, the partial sequences were subjected to BLASTx analysis against
the Caenorhabditis taxid and BLASTp analysis, respectively on NCBI
(http://blast.ncbi.nlm.nih.gov/Blast.cig) and NEMBASE4 (http://nematodes.org) [196].
3.2.4 Degenerate PCR approach and full-length amplification
The full-length or partial cDNA sequences of Con-pgp-2, Con-pgp-3, Con-pgp-12 and Con-pgp-
16 were obtained as described in [197], using degenerated primers followed by RACE-PCR.
Con-pgp-9 was partially isolated by a standard PCR using degenerated primers based on
sequence homology between C. elegans pgp-9 (GenBank ID: NM_075086) and T. circumcincta
pgp-9 (provided by Dr. P. Skuce) sequences. Furthermore, the full-length sequences of the
identified GluCl subunit genes (i.e. Con-glc-3, Con-glc-4 and Con-glc-6) were generated. The 5’
end of Con-glc-3 was isolated by a standard PCR in 2 steps. In the first step degenerated
primers based on sequence homology between C. elegans glc-3 (GenBank ID: NM_072040),
Cylicocyclus nassatus glc-3 (GenBank ID: AY727925) and H. contortus glc-3 (GenBank ID:
JF298242) sequences were used and isolated a 880 bp fragment. From this fragment an
antisense gene-specific primer was designed (Glc3R5’) to amplify the last part of the 5’ un-
translated region in combination with the nematode spliced leader sequence (SL1). The 5’
ends of Con-glc-4 and Con-glc-6 were isolated in only 1 step, using the SL1 primer in
combination with the antisense gene-specific primers Glc4R5’ and Glc6R5’, respectively.
Cloning and sequencing of all fragments was performed as described above. The sequences of
all primer pairs used are summarized in APPENDIX B.
3.2.5 Reverse transcriptase PCR
To investigate the transcription pattern of the individual GluCl subunit and ABC transporter
genes throughout the life cycle of C. oncophora, reverse transcription PCRs (SuperScript™
One-Step RT-PCR with Platinum® Taq, Invitrogen) were carried out. PCR mixtures had final
concentrations of 200 ng RNA template, 0.8 μM of both forward and reverse primer, 1 unit of
RT/Platinum® Taq Mix, in a reaction buffer containing 0.2 mM of each dNTP and 1.2 mM MgCl2.
PCR conditions were set as follows: 30 min at 50°C, 2 min at 94°C, followed by 40 cycles of
denaturing (15 s at 94°C), annealing (30 s at 60°C) and elongation (12 s at 72°C), followed by
a final elongation step at 72°C for 10 min, after which the PCR mixtures were kept at 10°C.
Glyceraldehyde 3-phosphate dehydrogenase (Con-gapdh) was included as an internal standard.

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
48
3.3 Results
3.3.1 Transcript reconstruction
Sequencing of the transcriptome of C. oncophora resulted in 9,603,581 reads. After screening
for possible host contamination 8,743,854 parasite-derived reads were left. Putative protein
translations resulted in 29,900 predicted polypeptides, covering an estimated 81% of the
complete transcriptome of C. oncophora. In C. oncophora, 6,406 polypeptides (21.4%) were
found in all stages, and less than 1% of the peptides were expressed in a single stage. On
average, 35% of the peptides in any stage were constitutively transcribed and the majority of
these peptides were predicted to be involved in genetic information processing, as
transcription and translation. Comparison of the stage-specific expression of polypeptides
within species revealed that the majority of polypeptides, expressed in each stage, were not
differentially expressed. C. oncophora females exhibit the highest percentage of up-regulated
polypeptides, whereas L3 with sheath show the highest percentage of down-regulated
transcripts. InterProScan analysis of the free-living (egg, L1, L2, L3) and parasitic (xL3, L4, M,
F) stages revealed that some domains and associated functions are abundant in both groups,
while others are uniquely to a single stage or group [191]. In TABLE 3.1 the assembly and
annotation information is summarized.
3.3.2 Identification of GluCl subunit genes
By screening the C. oncophora transcriptome database for genes that encode possible GluCl
subunits, 4 genes were identified. Based on the BLASTx results on NCBI, the four C. oncophora
sequences were identified as homologues of glc-3, glc-4 and avr-14 and a parasite-specific
gene glc-6. Out of these 4 genes, only Con-avr-14 had been described before [130-132]. The
full-length cDNA sequences of Con-glc-3, Con-glc-4 and Con-glc-6 were generated using the
nematode spliced leader sequence SL1 and encode for predicted proteins of 492 amino acids
(AA), 503 AA and 444 AA, respectively. The TMHMM server version 2.0
(http://cbs.dtu.dk/services/TMHMM-2.0) was used to predict the membrane protein topology
and confirmed the characteristics of Cys-loop ligand-gated ion channels for Con-glc-3, Con-glc-
4 and Con-glc-6. A phylogenetic tree of the identified C. oncophora GluCl subunits with
homologues from Caenorhabditis elegans, Haemonchus contortus and Ostertagia ostertagi is
presented in FIGURE 3.1. Reverse transcriptase PCR of all GluCl subunits in C. oncophora
showed that all these genes were constitutively expressed throughout the life cycle of C.
oncophora, except for a lower transcription in the eggs of Con-glc-4, Con-glc-6 and the Con-
avr-14 splice variants (TABLE 3.2).
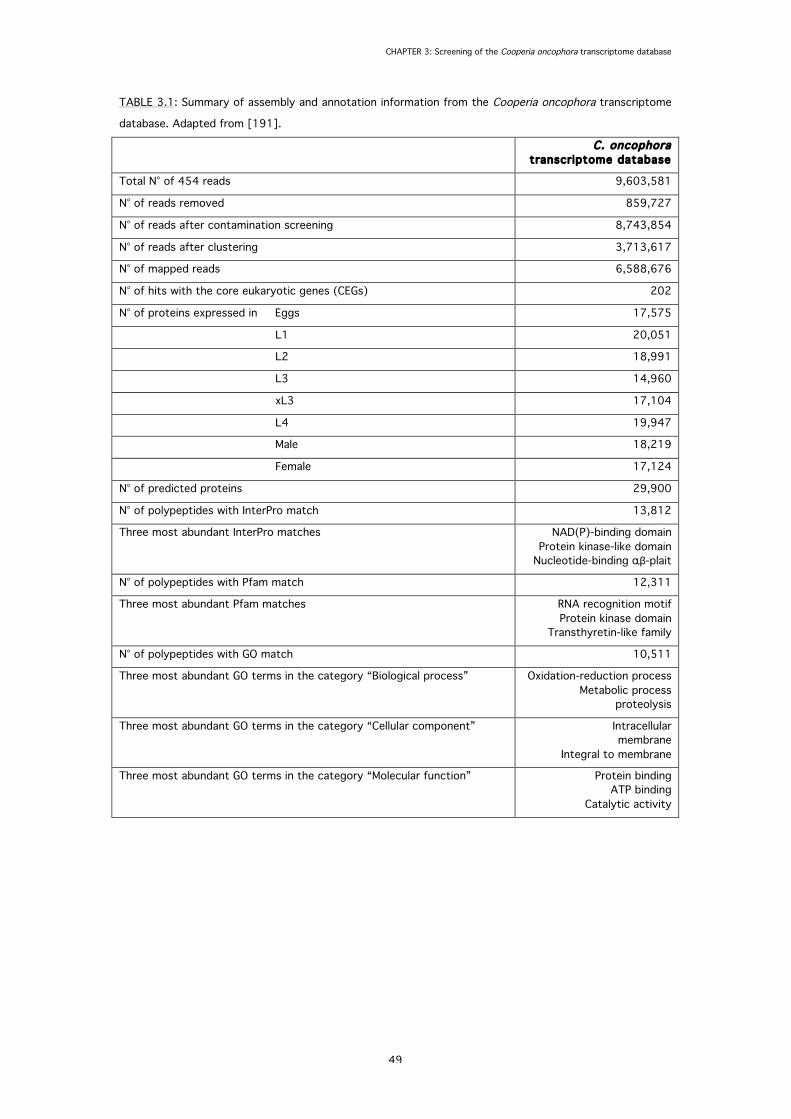
CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
49
TABLE 3.1: Summary of assembly and annotation information from the Cooperia oncophora transcriptome
database. Adapted from [191].
C. oncophora transcriptome database
Total N° of 454 reads 9,603,581
N° of reads removed 859,727
N° of reads after contamination screening 8,743,854
N° of reads after clustering 3,713,617
N° of mapped reads 6,588,676
N° of hits with the core eukaryotic genes (CEGs) 202
N° of proteins expressed in Eggs 17,575
L1 20,051
L2 18,991
L3 14,960
xL3 17,104
L4 19,947
Male 18,219
Female 17,124
N° of predicted proteins 29,900
N° of polypeptides with InterPro match 13,812
Three most abundant InterPro matches NAD(P)-binding domain Protein kinase-like domain
Nucleotide-binding αβ-plait
N° of polypeptides with Pfam match 12,311
Three most abundant Pfam matches RNA recognition motif Protein kinase domain
Transthyretin-like family
N° of polypeptides with GO match 10,511
Three most abundant GO terms in the category “Biological process” Oxidation-reduction process Metabolic process
proteolysis
Three most abundant GO terms in the category “Cellular component” Intracellular membrane
Integral to membrane
Three most abundant GO terms in the category “Molecular function” Protein binding ATP binding
Catalytic activity

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
50
FIGURE 3.1: Phylogenetic analysis of identified glutamate-gated chloride channel subunits in Cooperia
oncophora, Ostertagia ostertagi, Haemonchus contortus and Caenorhabditis elegans.
TABLE 3.2: Sequence sizes, accession numbers and transcription patterns throughout the life cycle of
Cooperia oncophora of the 6 so far identified glutamate-gated chloride channel subunits.
Transcript name
Isogroup from transcriptome
database
Full-length cDNA (bp)
Accession number Transcription throughout C. oncophora’s l ife cycle
E L1 L2 L3 L4 ♂ ♀
Con-glc-2 - 1296 AY372757
Con-glc-3 07564 1476 HF545673
Con-glc-4 15318 1509 HF545674
Con-glc-6 03101 1332 HF545675
Con-avr-14A - 1266 EU006790
Con-avr-14B Con-gapdh
17167
CONTROL
1314
204
AY372756
FR690828
0.1
Cel-glc-2
Con-glc-2
Hco-glc-2
Cel-glc-4
Con-glc-4Hco-glc-4
Cel-glc-1
Cel-avr-15ACel-avr-15B
Cel-glc-3
Con-glc-3
Hco-glc-3
Hco-glc-5
Cel-avr-14A
Oos-avr-14ACon-avr-14AHco-avr-14A
Cel-avr-14BHco-avr-14BCon-avr-14B
Oos-avr-14B
Con-glc-6Hco-glc-6
Oos-glc-6
0.1
Cel-glc-2
Con-glc-2
Hco-glc-2
Cel-glc-4
Con-glc-4Hco-glc-4
Cel-glc-1
Cel-avr-15ACel-avr-15B
Cel-glc-3
Con-glc-3
Hco-glc-3
Hco-glc-5
Cel-avr-14A
Oos-avr-14ACon-avr-14AHco-avr-14A
Cel-avr-14BHco-avr-14BCon-avr-14B
Oos-avr-14B
Con-glc-6Hco-glc-6
Oos-glc-6
!"
!"
!"
!"
!"!"
!"
!"
Con-AVR-14B Hco-AVR-14B Oos-AVR-14B
Hco-GLC-6
Oos-GLC-6
Con-GLC-6
Hco-GLC-2
Con-GLC-2
Cel-GLC-2
Cel-AVR-14B
Cel-AVR-14A
Hco-AVR-14A Con-AVR-14A
Oos-AVR-14A
Hco-GLC-5
Hco-GLC-3
Con-GLC-3
Cel-GLC-3
Cel-GLC-1
Cel-AVR-15B Cel-AVR-15A
Cel-GLC-4
Hco-GLC-4 Con-GLC-4
1000
1000
1000
1000
1000 740
997
867
957 817
1000
1000
1000
986
1000
630 986
1000 592
1000 753

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
51
3.3.3 Identification of ABC transporter genes
Analysis of the C. oncophora transcriptome dataset resulted in the identification of 12 partial
sequences encoding ABC transporters ranging in size from 129 bp to 1248 bp (TABLE 3.3).
Based on the best BLASTp results on the NEMBASE4 server against the C. elegans protein
database, the C. oncophora sequences were subsequently assigned the putative correct gene
name. Four partial P-glycoprotein sequences were identified with highest homology to pgp-1,
pgp-2, pgp-3 and pgp-11, respectively. Furthermore, five partial half transporters were
identified (i.e. haf-2, haf-3, haf-4, haf-7 and haf-9) and three partial sequences encoding MRPs
(i.e. mrp-1, mrp-4 and mrp-7). An additional three C. oncophora pgp sequences were identified
by a degenerate PCR approach with homology to i.e. C. elegans pgp-9, pgp-12 and pgp-16.
Reverse transcriptase PCR showed that all ABC transporter genes identified were constitutively
transcribed throughout the life cycle of C. oncophora, except for only a very low transcript
level of Con-pgp-1 and Con-pgp-16 and no expression of Con-pgp-9 in eggs.
3.4 Discussion
Sequencing of all transcripts in the eggs, L1, L2, L3, L4, male and female worms of C.
oncophora provided an important resource for exploring the extent of candidate gene families,
proven to be involved in ML-resistance [123, 127, 130-132, 135, 140, 149, 152, 154-156].
Four GluCl subunit genes, 7 pgp genes, 5 haf genes and 3 mrp genes were identified in this
study, either by mining the C.oncophora transcriptome dataset or by a degenerate PCR
approach. Twelve out of the 15 ABC transporter genes and all 4 GluCl subunit genes were
identified in the transcriptome database, indicating that these are likely to be the most highly
transcribed members of their gene families in C. oncophora under normal conditions.
Transcripts of Con-pgp-9, Con-pgp-12 and Con-pgp-16, on the other hand, were only identified
by PCR, suggesting a lower constitutive transcript level.
It is important to note that since most of the identified ABC transporter genes were partial,
the currently assigned gene names may still change once more sequence information becomes
available. In contrast to the situation in mammals, nematodes have a much more diverse
repertoire of ABC transporter genes [127, 198]. The reason for this diversity is unknown, but
it is suggested that they might be essential in protecting different neurones in the body of the
nematodes from a broad spectrum of toxins [154] and may also be a defence mechanism for
their changing environmental conditions i.e. free-living to parasitic stage. This wide repertoire
of ABC transporters in nematodes also suggests that many transporters have overlapping
substrate-specificities. Therefore, it is likely that a combination of several ABC transporters is
required for resistance to MLs.
Most of the ABC transporter genes investigated were constitutively transcribed throughout
the complete life cycle of C. oncophora, suggesting that they have a basic function in the

CHAPTER 3: Screening of the Cooperia oncophora transcriptome database
52
metabolism of these worms. Similarly, Con-glc-2 and Con-glc-3 appeared to be constitutively
transcribed throughout the complete life cycle of C. oncophora as well, while the other GluCl
subunit genes investigated (i.e. Con-glc-4, Con-glc-6 and Con-avr-14), showed a lower
transcription in the egg stage. This could be explained by the fact that these genes are mainly
expressed in the parasite’s nervous system, which is still under development in the egg stage
[106]. Since MLs are not ovicidal, it was already suggested that the gene(s) of interest are not
expressed in parasite eggs [81]. Similarly, the level of expression of the α-subunit encoded by
H. contortus glc-5 was found to be highest in adult worms, then in L3 and at lowest in the egg
stage [137].
MLs act on GluCl channels, uniquely found in invertebrates, which are presumed to be made up
of 5 subunits surrounding a central pore. So far, the number of identified GluCl subunits in C.
oncophora, H. contortus and C. elegans is already higher than 5. This indicates that nematodes
contain multiple forms of these ML-receptor channels, which may differ in their sensitivity to
the current drugs. α-subunits of GluCl channels appear to be sensitive to IVM and in this study,
2 new C. oncophora genes (i.e. glc-3 and glc-6) encoding for α-subunits were identified. Also
Con-glc-4 was newly identified and encodes for the more divergent, but conserved γ-subunit
[110, 113]. The Con-avr-14 transcript identified from the transcriptome database encoded for
the IVM-sensitive B splice variant [115, 131, 134]. The alternative splicing pattern of avr-14 is
conserved across species as C. elegans, H. contortus, C. oncophora and O. ostertagi and it is
suggested the avr-14B transcript may have arisen as a result of partial gene duplication of the
avr-14A transcript [120]. No orthologues of the IVM-sensitive Cel-glc-1 and Cel-avr-15 genes
have been identified in the C. oncophora transcriptome dataset. Interestingly, a glc gene could
be identified with highest homology to the IVM-sensitive H. contortus glc-6, which appears to
be absent in the free-living nematode C. elegans. If GLC-6 is a parasite-specific GluCl subunit
that is required in the formation of fully functional ML-receptor channels, then, glc-6 could be
the ultimate candidate resistance gene in parasitic worms [106, 123, 199].
The transcriptomic data produced by Heizer et al. [191] revealed that many differences in the
most prevalent InterPro signatures were between the free-living and parasitic stages of C.
oncophora. In the parasitic stages, polypeptides containing domains that traditionally function
in the degradation of proteins dominated, while polypeptides involved in growth and
development are more prominent in the free-living stages. These differences may be linked to
host adaptation and therefore parasitism. Further in-depth explorations can help to provide
novel methods to control infections and alternative drug targets. Additionally, this approach
can provide resources for expression microarrays for both ML-resistant (exposed to drugs/not
exposed to drugs) and ML-susceptible C. oncophora worms, and hence help to identify markers
of ML-resistance. In the longer term, comparative analyses of the biology, evolution and
adaptation to parasitism in C. oncophora will be extremely useful for annotating C. oncophora’s
upcoming genome [191].
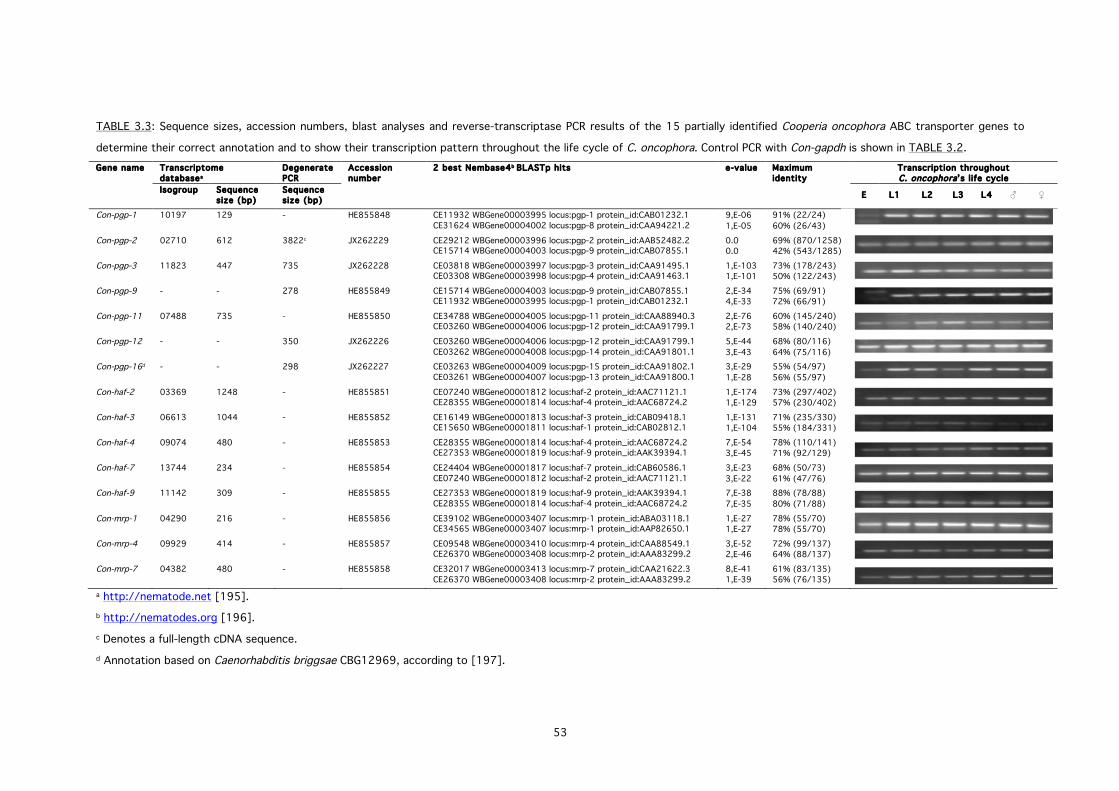
53
TABLE 3.3: Sequence sizes, accession numbers, blast analyses and reverse-transcriptase PCR results of the 15 partially identified Cooperia oncophora ABC transporter genes to
determine their correct annotation and to show their transcription pattern throughout the life cycle of C. oncophora. Control PCR with Con-gapdh is shown in TABLE 3.2. Gene name Transcriptome
databasea Degenerate PCR
Accession number
2 best Nembase4b BLASTp hits e-value Maximum identity
Transcription throughout C. oncophora’s l ife cycle
Isogroup Sequence size (bp)
Sequence size (bp) E L1 L2 L3 L4 ♂ ♀
Con-pgp-1 10197 129 - HE855848 CE11932 WBGene00003995 locus:pgp-1 protein_id:CAB01232.1 9,E-06 91% (22/24) CE31624 WBGene00004002 locus:pgp-8 protein_id:CAA94221.2 1,E-05 60% (26/43)
Con-pgp-2 02710 612 3822c JX262229 CE29212 WBGene00003996 locus:pgp-2 protein_id:AAB52482.2 0.0 69% (870/1258) CE15714 WBGene00004003 locus:pgp-9 protein_id:CAB07855.1 0.0 42% (543/1285)
Con-pgp-3 11823 447 735 JX262228 CE03818 WBGene00003997 locus:pgp-3 protein_id:CAA91495.1 1,E-103 73% (178/243) CE03308 WBGene00003998 locus:pgp-4 protein_id:CAA91463.1 1,E-101 50% (122/243)
Con-pgp-9 - - 278 HE855849 CE15714 WBGene00004003 locus:pgp-9 protein_id:CAB07855.1 2,E-34 75% (69/91) CE11932 WBGene00003995 locus:pgp-1 protein_id:CAB01232.1 4,E-33 72% (66/91)
Con-pgp-11 07488 735 - HE855850 CE34788 WBGene00004005 locus:pgp-11 protein_id:CAA88940.3 2,E-76 60% (145/240) CE03260 WBGene00004006 locus:pgp-12 protein_id:CAA91799.1 2,E-73 58% (140/240)
Con-pgp-12 - - 350 JX262226 CE03260 WBGene00004006 locus:pgp-12 protein_id:CAA91799.1 5,E-44 68% (80/116) CE03262 WBGene00004008 locus:pgp-14 protein_id:CAA91801.1 3,E-43 64% (75/116)
Con-pgp-16d - - 298 JX262227 CE03263 WBGene00004009 locus:pgp-15 protein_id:CAA91802.1 3,E-29 55% (54/97) CE03261 WBGene00004007 locus:pgp-13 protein_id:CAA91800.1 1,E-28 56% (55/97)
Con-haf-2 03369 1248 - HE855851 CE07240 WBGene00001812 locus:haf-2 protein_id:AAC71121.1 1,E-174 73% (297/402) CE28355 WBGene00001814 locus:haf-4 protein_id:AAC68724.2 1,E-129 57% (230/402)
Con-haf-3 06613 1044 - HE855852 CE16149 WBGene00001813 locus:haf-3 protein_id:CAB09418.1 1,E-131 71% (235/330) CE15650 WBGene00001811 locus:haf-1 protein_id:CAB02812.1 1,E-104 55% (184/331)
Con-haf-4 09074 480 - HE855853 CE28355 WBGene00001814 locus:haf-4 protein_id:AAC68724.2 7,E-54 78% (110/141) CE27353 WBGene00001819 locus:haf-9 protein_id:AAK39394.1 3,E-45 71% (92/129)
Con-haf-7 13744 234 - HE855854 CE24404 WBGene00001817 locus:haf-7 protein_id:CAB60586.1 3,E-23 68% (50/73) CE07240 WBGene00001812 locus:haf-2 protein_id:AAC71121.1 3,E-22 61% (47/76)
Con-haf-9 11142 309 - HE855855 CE27353 WBGene00001819 locus:haf-9 protein_id:AAK39394.1 7,E-38 88% (78/88) CE28355 WBGene00001814 locus:haf-4 protein_id:AAC68724.2 7,E-35 80% (71/88)
Con-mrp-1 04290 216 - HE855856 CE39102 WBGene00003407 locus:mrp-1 protein_id:ABA03118.1 1,E-27 78% (55/70) CE34565 WBGene00003407 locus:mrp-1 protein_id:AAP82650.1 1,E-27 78% (55/70)
Con-mrp-4 09929 414 - HE855857 CE09548 WBGene00003410 locus:mrp-4 protein_id:CAA88549.1 3,E-52 72% (99/137) CE26370 WBGene00003408 locus:mrp-2 protein_id:AAA83299.2 2,E-46 64% (88/137)
Con-mrp-7 04382 480 - HE855858 CE32017 WBGene00003413 locus:mrp-7 protein_id:CAA21622.3 8,E-41 61% (83/135) CE26370 WBGene00003408 locus:mrp-2 protein_id:AAA83299.2 1,E-39 56% (76/135)
a http://nematode.net [195]. b http://nematodes.org [196]. c Denotes a full-length cDNA sequence. d Annotation based on Caenorhabditis briggsae CBG12969, according to [197].

54

CHAPTER 4 Gene expression and mutation analysis of glutamate-gated
chloride channels in resistance Cooperia oncophora isolates
following in vivo exposure to macrocyclic lactones

56
Based on: De Graef J, Claerebout E, Vercruysse J, Wolstenholme A, Mitreva M and Geldhof P.
Gene expression and mutation analysis of the parasite-specific glutamate-gated chloride
channel GLC-6 in resistant Cooperia oncophora isolates following in vivo exposure to
macrocyclic lactones. International Journal for Parasitology: Drug and Drug Resistance. 2013
(submitted).

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
57
4.1 Introduction
Macrocyclic lactones (MLs), such as the avermectin ivermectin (IVM) and the milbemycin
moxidectin (MOX), paralyse the pharyngeal, somatic and/or uterine muscles of susceptible
parasites by binding to the invertebrate glutamate-gated chloride (GluCl) channels. The GluCl
channels are members of the Cys-loop ligand-gated ion channel family and are formed by the
assembly of 5 subunits [110, 113] (CHAPTER 1).
The composition of the GluCl gene family in nematodes varies substantially between species. In
the free-living nematode Caenorhabditis elegans, 6 GluCl subunit genes have been identified
and well characterized. Cel-glc-1, Cel-glc-3, Cel-avr-14 and Cel-avr-15 encode for GluCl α-type
subunits, Cel-glc-2 encodes for a β-subunit, whereas Cel-glc-4 encodes for a rather distant γ-
type of subunit [106]. Only avr-14, which can be alternatively spliced to create 2 subunits,
has a clear homologue in parasitic nematode species such as Haemonchus contortus, H. placei,
Cooperia oncophora, Ostertagia ostertagi, Teladorsagia circumcincta, Brugia malayi, Trichinella
spiralis and Dirofilaria immitis [121, 130, 132, 133, 200-202]. On the other hand, 2 parasite-
specific GluCl subunits, GLC-5 and GLC-6, have been characterized in H. contortus [123, 137].
From the C. oncophora transcriptome database, 3 GluCl subunit genes were newly identified,
i.e. glc-3, glc-4 and the parasite-specific glc-6 (CHAPTER 3).
In C. elegans, simultaneous mutation of glc-1, avr-14 and avr-15 confers a very high level of
IVM-resistance (4000-fold) [115]. Further, it appears that only the GluCl α-type subunits are
IVM-gated, such as C. elegans GLC-1, GLC-3, AVR-14B, AVR-15A and AVR-15B and H.
contortus GLC-5 and GLC-6 [113-117, 123, 141]. Therefore, these GluCl subunit genes are of
major interest for investigation of resistance-associated changes. In C. oncophora, 3 amino
acid (AA) substitutions, E114G, V235A and L256F, were identified in the AVR-14B subunit of
an IVM-resistant isolate from the United Kingdom. Electrophysiological recordings
demonstrated that only the L256F mutation accounted for a loss (2- to 3-fold) in sensitivity
to glutamate, IVM and MOX [131]. However, studies investigating the presence of the L256F
mutation in several resistant isolates of C. oncophora, O. ostertagi, H. contortus and T.
circumcincta were all negative [132, 133, 140]. Nevertheless, it was suggested that avr-14B
was under selection of IVM in Belgian C. oncophora isolates, as a loss in allelic diversity and a
decrease in transcription levels was observed in IVM-resistant worms [132].
The aim of the present study was to investigate the possible involvement of all identified GluCl
channel subunit genes in C. oncophora (see CHAPTER 3) in the mechanism of ML-resistance.
Firstly, by analysing constitutive and inducible changes in gene transcription levels between a
susceptible and ML-resistant field isolates and secondly, by investigating AA substitutions
between susceptible, IVM and/or MOX pressurised and unpressurised C. oncophora populations.

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
58
4.2 Materials and methods
4.2.1 Parasite material
In total 3 anthelmintic-susceptible and 2 IVM-resistant C. oncophora isolates have been
investigated in this study. The anthelmintic-susceptible isolates have never been exposed to
drug treatments and originate from Ghent (Belgium) (CoIVSus), Moredun (UK) (CoIVSusM) and
Weybridge (UK) (CoIVSusW). The IVM-resistant C. oncophora isolates (CoIVR08 and CoIVR09)
were collected from 2 different Belgian cattle farms, respectively in 2008 and 2009 [19,
132]. The controlled efficacy test revealed 38% and 31% reduction in worm burden,
respectively for IVM and MOX for the CoIVR08 isolate (CHAPTER 2). For the CoIVR09 isolate,
the efficacy of an IVM treatment was 0%, as determined by FECRT [132]. Adult worms were
collected as described in CHAPTER 3. In vivo exposed resistant adult worms were recovered
live at necropsy, 14 days post-treatment with IVM or MOX (0.2 mg/kg bodyweight) during the
infection trial described in CHAPTER 2. Considering the theoretical assumption that target
tissue concentrations above 1 ng/g represent the minimal drug level required for optimal anti-
parasitic activity, IVM and MOX would still be present at sufficiently high levels 14 days post
subcutaneous injection. Both, IVM and MOX concentrations remain >1 ng/g in gastrointestinal
mucosal tissue for 18 days post-treatment [94, 203]. Pools of worms (n = 100) were stored
at -80°C until required.
4.2.2 RNA extraction and cDNA synthesis
Total RNA samples were extracted by grinding the parasites in 0.2 ml glass homogenizers
(Wheaton) on ice, followed by a Trizol® extraction (Invitrogen). Residual genomic DNA was
removed by DNase I treatment (Roche). The RNA quality was verified with the Experion™ RNA
StdSens Starter kit (Bio-Rad) and the RNA concentration was determined using a Nanodrop®
ND-1000 spectrophotometer (NanoDrop Technologies). cDNA was synthesized from 1 μg total
RNA by random priming using the iScript cDNA synthesis kit (Bio-Rad) according to the
manufacturer’s recommendations.
4.2.3 Quantitative real-time PCR
Quantitative real-time PCRs were performed on the GluCl encoding genes identified in CHAPTER
3 to compare their constitutive (without drug exposure) and inducible (after exposure to IVM
or MOX) transcriptional changes between CoIVSus, CoIVR08 and CoIVR09 adult worms. Three
independent RNA extractions were performed as biological replicates. Total RNA was converted
to cDNA and diluted 1/5 or used undiluted for Con-glc-2 and Con-avr-14B. Real-time PCR
reactions were prepared with the SYBR Green Master Mix (Applied Biosystems) using 6.4 μl
H2O, 0.8 μl of each amplification primer (10 μM) and 2 μl of cDNA to give a 20 μl reaction
volume. All amplification runs were performed on a StepOnePlus Real-Time PCR System
(Applied Biosystems), under the following conditions: 95°C for 20 sec, followed by 40-50

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
59
cycles of 95°C for 5 sec, optimal annealing temperature (APPENDIX B) for 20 sec and an
extension of 72°C for 15 sec. A melting curve analysis was performed at the end of the
reaction to ensure specificity of the primers. Each run also included a five-point dilution series
of pooled cDNA and a non-template control. Technical replicates of each sample were
performed in duplicate within the same run. For each transcript, the mean Ct value of the
replicates was calculated and then corrected for the run efficiency. Subsequently, Ct values
were transformed in relative quantities (Q) using the delta Ct method: Q = E (min Ct – sample Ct).
Where E is the amplification efficiency and min Ct is the lowest Ct value. The relative quantities
were then normalized with the normalisation factor, obtained by the geNorm software for
reference genes Con-gapdh and C. oncophora β-tubulin (Con-tubb) [204, 205]. Transcript
levels were statistically analysed using an independent-samples t-test (SPSS Statistics 19).
Constitutive changes in transcription levels were regarded as being significant if P<0.05 in
comparison with the expression level in unexposed susceptible adults. Inducible changes in
transcription levels were regarded as being significant if P<0.05 in comparison with the
expression level in unexposed resistant adults.
4.2.4 Mutation analysis of the full-length Con-avr-14B and Con-glc-6 sequences
The Superscript™ One-Step Reverse Transcriptase for Long Templates kit (Invitrogen) was used
to amplify the full-length C. oncophora glc-6 (HF545675) and avr-14B (AY372756) cDNA
sequences from RNA of pooled adult worms of the following isolates: CoIVSus, CoIVSusM,
CoIVSusW, CoIVR08, CoIVR08 exposed to IVM, CoIVR08 exposed to MOX, CoIVR09 and
CoIVR09 exposed to IVM. PCR mixtures typically consisted of 250-500 ng RNA, 1 μM of both
forward and reverse primers (APPENDIX B), 0.25 μl of both RT/Platinum® Taq High Fidelity Mix
and Platinum® Taq DNA polymerase, 6.25 μl of the 2x Reaction Mix and DEPC water upon a
total volume of 12.5 μl. The following cycling conditions were established: one cycle of 30 min
at 50°C and 2 min at 94°C were performed in the cDNA synthesis and pre-denaturation step,
followed by 35 cycles of denaturing (15 s at 94°C), annealing (30 s at 62°C) and elongation
(90 s at 72°C). Afterwards a final extension step at 72°C for 5 min was included and then kept
at 10°C.
Amplified products were purified from the 1.5% agarose gels with the Geneclean kit® (MPBio),
ligated in the pGEM®-T easy vector (Promega) and transformed in E. coli DH5α competent cells
(Stratagene). Plasmid products were sequenced in 2 bidirectional reactions: one with SP6 and
T7 vector primers and the other one using a set of internal primers (Fi and Ri) (APPENDIX B), in
order to cover the complete full-length sequences of Con-glc-6 and Con-avr-14B in 4
overlapping fragments. At least 20 clones per isolate were sequenced to obtain the most
abundant full-length sequences of Con-glc-6 and Con-avr-14B per isolate. Sequences were
analysed with DNASTAR software (Lasergene version 8). Only mutations that were present in

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
60
at least 2 sequenced clones were considered in the analyses, to reduce the probability of
identifying sequencing artefacts.
4.3 Results
4.3.1 Analysis of constitutive and inducible transcriptional changes of GluCl subunit genes
Constitutive and inducible expression levels of the identified C. oncophora GluCl subunit genes
were compared between susceptible worms (pooled males and females), IVM-resistant
CoIVR08 worms (unexposed or in vivo exposed to IVM and MOX) and IVM-resistant CoIVR09
worms (unexposed or in vivo exposed to IVM). The real-time PCR results are shown in FIGURE
4.1. For Con-glc-2, constitutive expression levels were significantly increased in both IVM-
resistant isolates, respectively 2.9 and 4.7-fold in CoIVR08 and CoIVR09 worms. In vivo
exposure to IVM or MOX induced a decrease of Con-glc-2 in CoIVR08 and CoIVR09 compared
to both unexposed resistant isolates, but still the net result showed higher transcript levels in
both CoIVR08 (1.7 and 2.2-fold) and CoIVR09 (3.3-fold) relative to unexposed susceptible
worms (FIGURE 4.1). For Con-glc-3, the constitutive expression level was 2.1-fold higher in
resistant CoIVR08 worms compared to susceptible worms (P<0.01). IVM exposure induced a
2.8-fold decrease of Con-glc-3 mRNA transcripts in CoIVR08 worms relative to unexposed
resistant worms (P<0.01). No constitutive transcriptional differences were observed for Con-
glc-4, but a significant decrease (2.2 –fold) was induced in the CoIVR08 adults after exposure
to IVM. A significant constitutive decrease (1.8-fold) was observed for Con-glc-6 in resistant
CoIVR09 worms compared to the CoIVSus isolate, which was maintained after in vivo IVM
exposure of CoIVR09. Furthermore, in vivo exposure to IVM resulted in a significant down-
regulation of Con-glc-6 transcript levels (2.7-fold) in the CoIVR08 isolate compared to the
unexposed CoIVR08 adult worms. The expression profiles of Con-avr-14A and Con-avr-14B are
comparable in the CoIVR08 isolate. The constitutive transcript level of Con-avr-14B was 1.6-
fold higher in the CoIVR08 isolate compared to susceptible worms (P<0.05). Exposure of
CoIVR08 worms to MOX maintained these significant up-regulations in both Con-avr-14A and
Con-avr-14B, whereas IVM exposure induced a 2.4-fold decrease of Con-avr-14B mRNA
transcripts in CoIVR08 worms relative to unexposed resistant worms (P<0.01).

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
61
FIGURE 4.1: Fold changes in constitutive and inducible mRNA transcript levels of glc-2, glc-3, glc-4, glc-6,
avr-14A and avr-14B in Cooperia oncophora adult worms. The transcript levels in unexposed susceptible
worms have been set at 1 and the transcript levels ± SD in unexposed CoIVR08, in vivo exposed CoIVR08
(to IVM and MOX), unexposed CoIVR09 and in vivo exposed CoIVR09 (to IVM) were expressed relative to
this (A). The transcript levels in unexposed resistant worms have been set at 1 and the transcript levels ±
SD in in vivo exposed CoIVR08 (to IVM and MOX, and in vivo exposed CoIVR09 (to IVM) were expressed
relative to this (B) Changes with P<0.05 were regarded as being significant (* P<0.05, ** P<0.01).
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
Fold
cha
nge
in tr
ansc
ript l
evel
s
CoIVSusCoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVSusCoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVSusCoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVSusCoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVSusCoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVSusCoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
Con-glc-2
Con-avr-14A
Con-glc-3
Con-glc-4 Con-glc-6
Con-avr-14B
Fold
cha
nge
in tr
ansc
ript l
evel
s Fo
ld c
hang
e in
tran
scrip
t lev
els
**
**
**
*
* ***
** *
** *
A

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
62
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
Fold
cha
nge
in tr
ansc
ript l
evel
s
CoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
Con-glc-2
Con-avr-14A
Con-glc-3
Con-glc-4 Con-glc-6
Con-avr-14B
Fold
cha
nge
in tr
ansc
ript l
evel
s Fo
ld c
hang
e in
tran
scrip
t lev
els
CoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
CoIVR08 CoIVR08exposedto IVM
CoIVR08exposedto MOX
CoIVR09 CoIVR09exposed to IVM
***
** **
**
B

63
FIGURE 4.2: Sequence alignment of the predicted protein sequences of Cooperia oncophora avr-14B (AY372756) and glc-6 (HF545675) with orthologous sequences from
Haemonchus contortus (Y14234 and ABV68895), Ostertagia ostertagi (CBX19419 and HF545676) and Caenorhabditis elegans (CE26421). Conserved residues are shaded in
grey. The cysteine residues belonging to the Cys-loop are in red, the predicted signal peptide is in bold and the boxes correspond with the 4 predicted membrane-spanning
domains. Residues presumably involved in hydrogen-bonding and van der Waals interactions with ivermectin are in blue and green, respectively.
Con-AVR-14B MRTSVPLATR IGPILALICI VITIISTVEG KRKLKEQEII QRILNNYDWR VRPRGLNASW PDTGGPVLVT VNIYLRSISK IDDVNMEYSA QFTFREEWVD ARLAYGRFED –ESTEVPPFV 119 Hco-AVR-14B MRNSVPLATR IGPMLALICT VSTIMSAVEA KRKLKEQEII QRILNNYDWR VRPRGLNASW PDTGGPVLVT VNIYLRSISK IDDVNMEYSA HFTFREEWVD ARLAYGRFED –ESTEVPPFV 119 Oos-AVR-14B MRTSVPLAAR IGPIVALICT ISTMICSVEG KRKLKEQEII QRILNNYDWR VRPRGLNASW PDTGGPVLVT VNIYLRSISK IDDVNMEYSA QFTFREEWVD ARLAYGRFED –ESTEVPPFV 119 Cel-AVR-14B ---------M WHYRLTTILL IISIIHSIRA KRKLKEQEII QRILKDYDWR VRPRGMNATW PDTGGPVLVT VNIYLRSISK IDDVNMEYSA QFTFREEWTD QRLAYERYEE SGDTEVPPFV 111
Con-AVR-14B VLATSENADQ SQQIWMPDTF FQNEKEARRH LIDKPNVLIR IHKDGSILYS VRLSLVLSCP MSLEFYPLDR QNCLIDLASY AYTTQDIKYE WKEQNPVQQK DGLRQSLPSF ELQDVVTKYC 239 Hco-AVR-14B VLATSENADQ SQQIWMPDTF FQNEKEARRH LIDKPNVLIR IHKDGSILYS VRLSLVLSCP MSLEFYPLDR QNCLIDLASY AYTTQDIKYE WKEQNPVQQK DGLRQSLPSF ELQDVVTKYC 239 Oos-AVR-14B VLATSENADQ SQQIWMPDTF FQNEKEARRH LIDKPNVLIR IHKDGSILYS VRLSLVLSCP MSLEFYPLDR QNCLIDLASY AYTTQDIKYE WKEQNPVQQK DGLRQSLPSF ELQDVVTKYC 239 Cel-AVR-14B VLATSENADQ SQQIWMPDTF FQNEKEARRH LIDKPNVLIR IHKNGQILYS VRLSLVLSCP MSLEFYPLDR QNCLIDLASY AYTTQDIKYE WKEKKPIQQK DGLRQSLPSF ELQDVVTDYC 231
Con-AVR-14B TSKTNTGEYS CARVKLLLRR EYSYYLIQLY IPCIMLVVVS WVSFWLDKDA VPARVSLGVT TLLTMTTQAS GINSKLPPVS YIKAVDVWIG VCLAFIFGAL LEYAVVNYYG RKEFLRKEKK 359 Hco-AVR-14B TSKTNTGEYS CARVKLLLRR EYSYYLIQLY IPCIMLLVVS WVSFWLDKDA VPARVSLGVT TLLTMTTQAS GINSKLPPVS YIKAVDVWIG VCLAFIFGAL LEYAVVNYYG RKEFLRKEKK 359 Oos-AVR-14B TSKTNTGEYS CARVKLLLRR EYSYYLIQLY IPCIMLVVVS WVSFWLDKDA VPARVSLGVT TLLTMTTQAS GINSKLPPVS YIKAVDVWIG VCLAFIFGAL LEYAVVNYYG RKEFLRKEKK 359 Cel-AVR-14B TSLTNTGEYS CARVVLRLRR EYSYYLIQLY IPCIMLVVVS WVSFWLDKDA VPARVSLGVT TLLTMTTQAS GINSKLPPVS YIKAVDVWIG VCLAFIFGAL LEYAVVNYYG RKEFLRKEKK 351
Con-AVR-14B KKTRLDDCVC PSERPALRLD LSTFRRRGWT PLN-RLLDVL GRNADLSRRV DLMSRITFPT LFTVFLVFYY SVYVKQ-SNL E 438 Hco-AVR-14B KKTRLDDCVC PSERPALRLD LSNYRRRGWT PLN-RLLDML GRNADLSRRV DLMSRITFPS LFTAFLVFYY SVYVKQ-SNL D 438 Oos-AVR-14B KKTRLDDCVC PSERPALRLD LSTFRRRGWA PLN-RLLDVL GRNADLSRRV DLMSRITFPS LFTAFLVFYY AAYVRQQSNL E 439 Cel-AVR-14B KKTRIDDCVC PSDRPPLRLD LSAYRSVKRL PIIKRISEIL STNIDISRRV DLMSRLTFPL TFFSFLIFYY VAYVKQ-SRD - 430
Con-GLC-6 MRSAF-ELII VFGSLSTILT SDVDAQVTST NSSTKMKPEE IMDVFISKSY DRRIRPPNRD SDGKNGPVLV SVNAYIRSMS NIDFVRMQYG VQVTFRQFWH DPRLAYEQMF PGVSVPKFII 119 Hco-GLC-6 MRITLMELVL VLGSMSPTLQ SDAASQTLSM SSSRNITVGE IMNVFINSSY DRRIRPPNRD SKGVNGPVMV KVNAYIRSIS NIDFVRMQYN LQVTFRQLWQ DSRLAYQNSF PNDKVPKFII 120 Oos-GLC-6 MRSTFAELVL IFGAFSSKFM SDAATQVAPT SDTPKIQAGE IMNVFINSSY DKRIRPPNRD STGKNGSVTV NVNAYIRSMS NIDFVRMQYN LQVTFRQFWS DPRLAYENLY PRKKFPKFII 120
Con-GLC-6 ITEKNLIWTP DTFFLNEKQA HRHEIDKLNL MIRIYANGSV MSSERLSFTF SCPMYLQKYP MDEQNCDMLL ASYAFTTDDI VYRWDEQNPI QYHAQLNTSL PNFSLQAART GECTSTTTTG 239 Hco-GLC-6 ITEKDLIWTP DTFFLNEKQA HRHEIDKLNL LLRIYSNGSV MYSERLSLTL SCPMYLHKYP MDEQYCQMLL ASYAFTTDDI VYQWEEQNPI QYHVLLNTSL PNFLLNAAET GECTSSTTTG 240 Oos-GLC-6 ITEKDLIWIP DTFFLNEKEA HRHEIDKLNL LMRIYANGSV MYSERLSLTL SCPMYLHKYP MDEQKCRLLL ASYAFTTDDI VYRWEEQNPI QYHVQLNTSL PNFSLASAEI GDCTSSTTTG 240
Con-GLC-6 EYSCLKTMFT LKRMFRFYLA QIYLPSTLLV VVSWVSFWLE RTAVPARVTL GVTTLLTMTT QAAAINNSLP PVSYIKAVDV WIGVCLAFIF AAVLEFAIVS YCASLRHGPC PTHEKYHDVV 359 Hco-GLC-6 EYSCLKVMFT MKRMFRFYLA QIYLPSTLLV VVSWVSFWLD RTAVPARVTL GVTTLLTMTT QAAAINNSLP PVSYIKAVDV WIGVCLAFIF AAVLEFAVLS YCASLMH--- -VHEKCQKVA 356 Oos-GLC-6 EYSCIQTMFT MKRMFRFYVA QIYLPSTLLV VVSWVSFWLE RTAVPARVTL GVTTLLTMTT QAAAINNSLP PVSYIKAVDV WIGTCLAFIF AAVVEFAIVS YCAAPKHGPS –HHKECLCAA 359
Con-GLC-6 REAQEKTKPQ EKPNRSDSWG TEKELESKLL KGSEGEETTK GKLSLWQRWK AGADPPKVID LKSRIMFPVF FIMFNIFYWT WYSFL 444 Hco-GLC-6 KEAHNENKSP MRGRQK--RD DNKHPDLKPL QPSDTDRPSK RGLSFWQRWK VGADPPKMID LKSRIIFPLF FIVFNVTYWT LYSFL 439 Oos-GLC-6 REATDKDRSS DEVNRKVVGA EAKNTEMEPL QASNVTRPHK GGFSFWRRWK EGADPPKVID LKSRIMFPVF FIVFNLVYWT WYSFL 444

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
64
4.3.2 Mutation analysis of Con-avr-14B
The predicted protein sequence of Con-avr-14B is well conserved and shows 97%, 97% and
85% identity, respectively with orthologous sequences from H. contortus, O. ostertagi and C.
elegans. AA heterogeneity can be found between species in the signal peptide sequence and
the internal loop between transmembrane domain (TM) 3 and TM4 (FIGURE 4.2). To
investigate the presence of mutations associated with ML-resistance, at least 20 clones of the
full-length cDNA sequences, encoding Con-AVR-14B, were obtained from 3 susceptible isolates
(CoIVSus, CoIVSusM and CoIVSusW), from 2 IVM-resistant Belgian field isolates (CoIVR08 and
CoIVR09) and from 3 IVM-resistant isolates after in vivo exposure to MLs (CoIVR08 + IVM,
CoIVR08 + MOX and CoIVR09 + IVM). A total of 180 protein sequences were compared and
resulted in the identification of 29 isoforms of Con-AVR-14B. In the CoIVSusM isolate, the
original sequence (AY372756) published by [130] was the only isoform identified (isoform 1).
Isoform 1 was not present in the CoIVSus and CoIVR09 isolates. Respectively 4, 2, 5, 5, 11,
1and 7 isoforms were identified in the CoIVSus, CoIVSusW, CoIVR08, CoIVR08 + IVM, CoIVR08
+ MOX, CoIVR09 and CoIVR09 + IVM isolates (FIGURE 4.3A). The AA substitutions and their
frequencies per isolate are shown in (FIGURE 4.4A). Over all isolates 23 AA substitutions were
identified which were present in at least 2 sequenced clones. All mutations are isolate-specific,
apart from I14V, which appears in both the CoIVR08 + MOX and CoIVR09 + IVM worms. Two
mutations, I24T/N and V326A, were enriched after IVM and/or MOX exposure of the CoIVR08
worms. Eight mutations were newly introduced after exposure of the CoIVR08 or CoIVR09
isolate. Most mutations appeared in the N-terminal extracellular domain (48%) and the
intracellular loop between TM3 and TM4 (26%).
4.3.3 Mutation analysis of Con-glc-6
The predicted protein sequence of Con-glc-6 is less conserved between species compared to
Con-avr-14B and shows only 74% identity with orthologous sequences from H. contortus and
O. ostertagi. Most AA heterogeneity was also found in the signal peptide sequence and the
internal loop between TM3 and TM4 (FIGURE 4.2). Similar to the mutation analysis of Con-AVR-
14B, at least 20 clones of the full-length cDNA sequences, encoding GLC-6, were analysed in
the same 8 isolates. A total of 175 protein sequences were compared and resulted in the
identification of 101 isoforms of Con-GLC-6. All isoforms were isolate specific. Respectively 7,
13, 9, 10, 14, 21, 14 and 13 isoforms were identified in the CoIVSus, CoIVSusM, CoIVSusW,
CoIVR08, CoIVR08 + IVM, CoIVR08 + MOX, CoIVR09 and CoIVR09 + IVM isolates (FIGURE
4.3B). The AA substitutions and their frequencies per isolate are shown in (FIGURE 4.4B). Over
all isolates 96 AA substitutions were identified which were present in at least 2/175 Con-GLC-
6 protein sequences. Forty-four mutations were isolate-specific, whereas 52 changes could be
found over 2 or more isolates. However, grouping the isolates by types susceptible (Sus),
resistant (Res) or resistant exposed (Res+) revealed that 22 out of these 52 mutations were
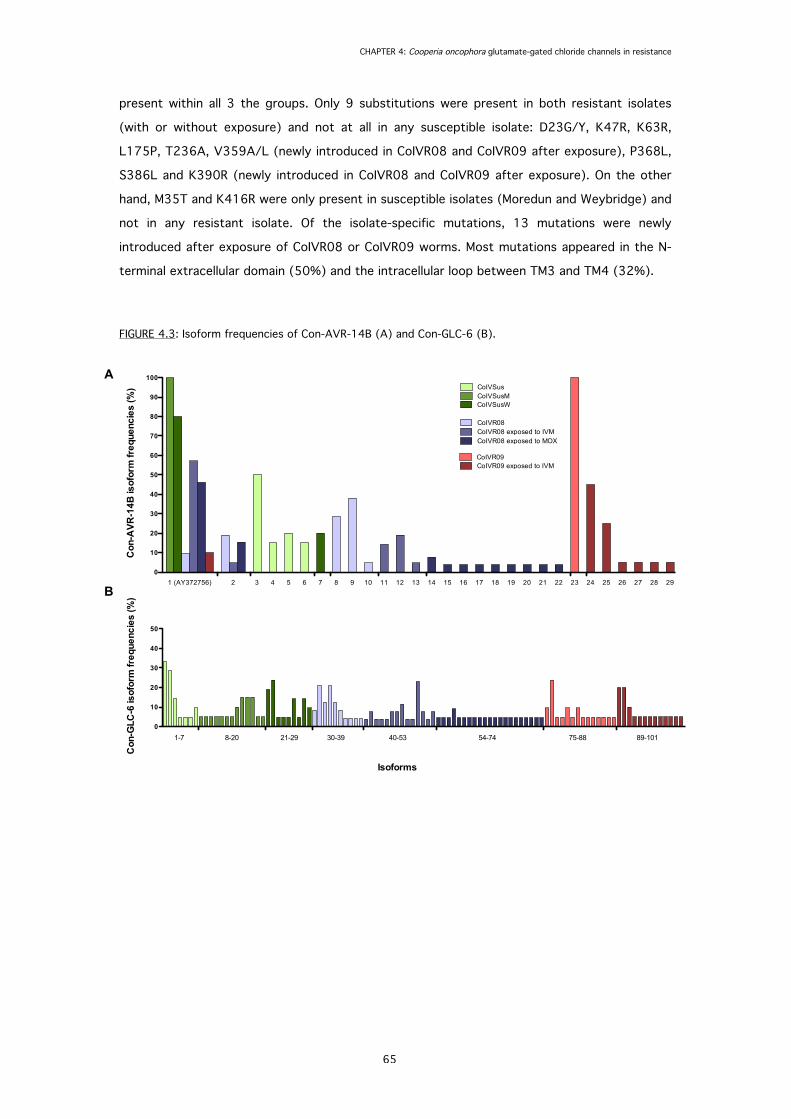
CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
65
present within all 3 the groups. Only 9 substitutions were present in both resistant isolates
(with or without exposure) and not at all in any susceptible isolate: D23G/Y, K47R, K63R,
L175P, T236A, V359A/L (newly introduced in CoIVR08 and CoIVR09 after exposure), P368L,
S386L and K390R (newly introduced in CoIVR08 and CoIVR09 after exposure). On the other
hand, M35T and K416R were only present in susceptible isolates (Moredun and Weybridge) and
not in any resistant isolate. Of the isolate-specific mutations, 13 mutations were newly
introduced after exposure of CoIVR08 or CoIVR09 worms. Most mutations appeared in the N-
terminal extracellular domain (50%) and the intracellular loop between TM3 and TM4 (32%).
FIGURE 4.3: Isoform frequencies of Con-AVR-14B (A) and Con-GLC-6 (B).
1 (AY372756) 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 290
10
20
30
40
50
60
70
80
90
100
CoIVSusMCoIVSus
CoIVSusW
CoIVR08CoIVR08 exposed to IVMCoIVR08 exposed to MOX
CoIVR09 exposed to IVMCoIVR09
0
10
20
30
40
50
Con
-AV
R-1
4B is
ofor
m fr
eque
ncie
s (%
)C
on-G
LC-6
isof
orm
freq
uenc
ies
(%)
Isoforms
A
B
1-7 8-20 21-29 30-39 40-53 54-74 75-88 89-101

66
FIGURE 4.4: Amino acid substitutions in Con-AVR-14B (A) and Con-GLC-6 (B) and their frequencies per isolate. The positions where substitutions were found are underlined. The
boxes indicate the 4 predicted membrane-spanning domains. Con-AVR-14B (AY372756) M R T S V P L A T R I G P I L A L I C I V I T I I S T V E G K R K L K E Q E I I Q R I L N N Y D W R V R P R G L N A S W P D T G G P V L V T V N I Y L R S I S K I D D V N M E Y S A 90
CoIVSus Ghent - - - - T(15%)CoIVSus Moredun - - - - -CoIVSus Weybridge - - - - -CoIVR08 - T(19%) V(29%) - -CoIVR09 - - - - -CoIVR08 + IVM - T(19%) - P(14%) -CoIVR08 + MOX V(8%) T/N(31%) - - -CoIVR09 + IVM V(5%) - - - -
Con-AVR-14B (AY372756) Q F T F R E E W V D A R L A Y G R F E D E S T E V P P F V V L A T S E N A D Q S Q Q I W M P D T F F Q N E K E A R R H L I D K P N V L I R I H K D G S I L Y S V R L S L V L S C P M 180
CoIVSus Ghent - - P(15%) P(15%)CoIVSus Moredun - - - -CoIVSus Weybridge - - - -CoIVR08 E(38%) del(29%) - -CoIVR09 - - - -CoIVR08 + IVM - - - -CoIVR08 + MOX - - - -CoIVR09 + IVM - - - -
Con-AVR-14B (AY372756) S L E F Y P L D R Q N C L I D L A S Y A Y T T Q D I K Y E W K E Q N P V Q Q K D G L R Q S L P S F E L Q D V V T K Y C T S K T N T G E Y S C A R V K L L L R R E Y S Y Y L I Q L Y I 270
CoIVSus Ghent - - - -CoIVSus Moredun - - - -CoIVSus Weybridge - - - -CoIVR08 - R(29%) - -CoIVR09 P(100%) - - -CoIVR08 + IVM - - - A(14%)CoIVR08 + MOX - - - -CoIVR09 + IVM - - S(10%) -
Con-AVR-14B (AY372756) P C I M L V V V S W V S F W L D K D A V P A R V S I G V T T L L T M T T Q A S G I N S K L P P V S Y I K A V D V W I G V C L A F I F G A L L E Y A V V N Y Y G R K E F L R K E K K K 360
CoIVSus Ghent R(50%) A(70%) - - -CoIVSus Moredun - - - - -CoIVSus Weybridge - - - - L(20%)CoIVR08 - - A(5%) - -CoIVR09 - - - - -CoIVR08 + IVM - - A(14%) - -CoIVR08 + MOX - - - - -CoIVR09 + IVM - - - Stop(25%) -
Con-AVR-14B (AY372756) K T R L D D C V C P S E R P A L R L D L S T F R R R G W T P L N R L L D V L G R N A D L S R R V D L M S R I T F P T L F T V F L V F Y Y S V Y V K Q S N L ECoIVSus Ghent - R(15%) - - -CoIVSus Moredun - - - - -CoIVSus Weybridge - - - - -CoIVR08 - - - - -CoIVR09 - - - - -CoIVR08 + IVM G(19%) - P(19%) Q(14%)-CoIVR08 + MOX - - - - N(8%)CoIVR09 + IVM - - - - -
438

67
Con-GLC-6 (Consensus) M R S A F E L I I V F G S L S T I L T S D V D A Q V T S T N S S T K M K P E E I M D V F I S K S Y D R R I R P P N R D S D G K N G 65CoIVSus Ghent F(19%) F/T(48%) - - - I(5%) I(29%) P(14%)A(14%) R(21%) - - - - - - - - - -CoIVSus Moredun F(75%) T(5%) - - - - - - - - T(5%) N(15%) - T(15%) - - - - N(10%) -CoIVSus Weybridge F(86%) - - - - - - - - - T(14%) - G(24%) - - T(24%) - N(10%) - -CoIVR08 F/L(46%) T(25%) - - G(33%) - - - - - - - - - - - - - - -CoIVR09 F(57%) F(5%) R/G(19%) - G(10%) A(5%) M(5%) - M(5%) A(5%) - - - - R(5%) - - - N(10%) R(5%)CoIVR08 + IVM F(54%) T(15%) - N(23%) - - - - - - - - D(4%) T(4%) R(23%) - - - - R(8%)CoIVR08 + MOX F(36%) F/T(14%) - N/G(18%)G(5%) - I(5%) - A(5%) R(5%) - - - - R(5%) - - N(5%) I(5%) -CoIVR09 + IVM F(20%) T/F(45%) - - Y(10%) A(10%) I(10%) - - R(10%) - - - - - - Q(20%) - N(20%) -
Con-GLC-6 (Consensus) P V L V S V N A Y I R S M S N I D F V R M Q Y G V Q V T F R Q F W H D P R L A Y E Q M F P G V S V P K F I I I T E K N L I W T P D 130CoIVSus Ghent H(10%) - T(29%) - - S(14%) - - - I(38%) A(38%) -CoIVSus Moredun - A(5%) T(25%) - - - - - R(15%) A(5%) - -CoIVSus Weybridge - - T(71%) - S(5%) - S(10%) W(19%) - - - T(10%)CoIVR08 - - T(13%) - - S(8%) - - - - - -CoIVR09 - I(10%) T(33%) S/D(14%) - S(24%) - - - - - -CoIVR08 + IVM - - T(54%) - K(8%) S(12%) - - - - - -CoIVR08 + MOX - - T(41%) - - S(18%) - - - - - -CoIVR09 + IVM - - T(20%) - - S(30%) - - - I(5%) A(5%) -
Con-GLC-6 (Consensus) T F F L N E K Q A H R H E I D K L N L M I R I Y A N G S V M S S E R L S F T F S C P M Y L Q K Y P M D E Q N C D M L L A S Y A F T 195CoIVSus Ghent - - - P(29%) - - H(43%) -CoIVSus Moredun - - G(10%) - - - - -CoIVSus Weybridge - - - - - - - -CoIVR08 - T(21%) - - - - H(21%) G(13%)CoIVR09 - - - - - P(5%) - -CoIVR08 + IVM - V(12%) - - L(8%) - - -CoIVR08 + MOX T(9%) - - - - P(5%) H(5%) -CoIVR09 + IVM - - - - - - H(5%) -
Con-GLC-6 (Consensus) T D D I V Y R W D E Q N P I Q Y H A L L N T S L P N F S L Q A A R T G E C T S T T T T G E Y S C L K T M F T L K R M F R F Y L A Q 260CoIVSus Ghent - - - - H(14%) K(33%) - - - - Y(33%) -CoIVSus Moredun - - - - - Q(20%) - A(15%) - - Y(10%) -CoIVSus Weybridge A(19%) - - - - Q(48%) - - G(19%) - - -CoIVR08 - V(8%) - - - Q(21%) A(21%) - - I(8%) - -CoIVR09 - - - V(10%) - Q(14%) A(5%) - - - - C(10%)CoIVR08 + IVM I(4%) - E(23%) - - Q(46%) - - - - Y(8%) -CoIVR08 + MOX - - G(5%) - - Q/K(18%) - - - - Y(5%) -CoIVR09 + IVM - - - - - Q(40%) - - - - - -
Con-GLC-6 (Consensus) I Y L P S T L L V V V S W V S F W L E R T A V P A R V T L G V T T L L T M T T Q A A A I N N S L P P V S Y I K A V D V W I G V C L 325CoIVSus Ghent - - - - - -CoIVSus Moredun - - - - G(15%) C(10%)CoIVSus Weybridge - S(19%) G(14%) - G(24%) H(5%)CoIVR08 - - - - - -CoIVR09 - - - - - -CoIVR08 + IVM - - - - - H(8%)CoIVR08 + MOX T(9%) - - - G(5%) -CoIVR09 + IVM - - - A(10%) - -
Con-GLC-6 (Consensus) A F I F A A V L E F A I V S Y C A S L R H G P C P T H E K Y H D V V R E A Q E K T K P Q E K P N R S D S W G T E K E L E S K L L K 390CoIVSus Ghent V(14%) - A(10%) H(43%) Q(33%) A(48%) - V(10%) S(10%) - - - - E(10%) Q(19%) - - - - - -CoIVSus Moredun - - A(20%) H(10%) - A(30%) - - - - - - L(10%) - Q(35%) - R(5%) - R(5%) - -CoIVSus Weybridge - - I(14%) - - A(62%) - - - E(24%) - - L(24%) - Q(43%) - - - - - -CoIVR08 - - - H(38%) - A(13%) - - R(21%) - T(21%) L(13%) L(13%)E(8%) Q(8%) - - L(8%) R(8%) P(8%) -CoIVR09 - - - H(52%) - A(14%) - - - - - L(24%) - - Q(5%) - R(24%) L(14%) - - -CoIVR08 + IVM - T(4%) - H(35%) L(4%) - - S/T/V(35%) R(12%) - - - - - Q(12%) S(23%) R(23%) L(8%)R(23%) - R(4%)CoIVR08 + MOX - T(9%) S(5%) H(36%) L(5%) A/F(14%) A(9%) - - - - L(5%) - - Q(23%) - R(9%) - - - -CoIVR09 + IVM - - - H(25%) - A(30%) L(20%) - - - - - - - Q(30%) - R(15%) L(10%) - - R(5%)
Con-GLC-6 (Consensus) G S E G E E T T K G K L S L W Q R W K A G A D P P K V I D L K S R I M F P V F F I M F N I F Y W T W Y S F LCoIVSus Ghent A(10%) - - A(10%) T(5%) - - - - - - - A(10%) C(10%) - L(10%) -CoIVSus Moredun - P(15%) - - T(30%) - R(10%) - P(5%) - C(10%) L(10%) - L(55%) - - -CoIVSus Weybridge - - - - T(14%) - R(43%) - P(5%) - - L(24%) - L(14%) - - -CoIVR08 A/R(13%) - V(8%) A(8%) N/T/R(46%) - - - - R(8%) - L(8%) - L/C(54%) - L(8%) -CoIVR09 - - - - T/S(57%) - - - - - - L(19%) - L(19%) - - -CoIVR08 + IVM - - - - T(12%) - - - P(23%) - - - - L(62%) - T(8%) S(12%)CoIVR08 + MOX - - - I(5%) T(18%) - - - - - - L(14%) - L(36%) - - -CoIVR09 + IVM del(5%) - - - S(20%) - N(20%) - - - L(5%) G(5%) L(40%) T(10%) - -
444

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
68
4.4 Discussion
One way in which parasite populations can develop resistance is by using specific mechanisms,
which involve the drug target, i.e. GluCl channels for MLs. For example, a mutation in the
receptor could make the binding site unrecognizable for the drug, or an altered expression
pattern of target genes could overcome or avert the drug action [110]. In this study, the
GluCl subunit genes glc-2, glc-3, glc-4, glc-6, avr-14A and avr-14B, identified in C. oncophora,
were investigated for their possible involvement in ML-resistance by gene expression analysis,
where the main focus was on similar trends in both ML-resistant isolates. Additionally, Con-glc-
6 and Con-avr-14B were further investigated for resistance-associated AA substitutions.
Quantitative real-time PCRs were performed to compare constitutive (without drug exposure)
and inducible (after exposure to IVM and MOX) changes in gene transcription levels. The IVM-
resistant C. oncophora isolates used in this study (CoIVR08 and CoIVR09) have been
characterized before in terms of the Con-avr-14 gene [132]. Con-avr-14 encodes for the
splice-variants AVR-14A and AVR-14B, which are likely to be under the same transcriptional
regulation. While El-Abdellati et al. (2011) observed significantly lower levels of both Con-avr-
14A and Con-avr-14B in L3, adult males and adult females of the resistant isolates [132],
here, no significant down-regulations could be observed in unexposed resistant worms. The
observed changes in transcript levels are rather small, have low statistically significance and
could therefore change in a rapid and unpredictable manner. The presented Con-avr-14
transcription patterns are more in line with the constitutive up-regulations of avr-14B found in
L3 of multiple-resistant T. circumcincta isolates. The authors supposed that the up-regulation
of avr-14B in these isolates was influenced by the co-selection with other anthelmintic drug
classes [133]. Regarding Con-glc-2, a constitutive up-regulation in IVM-resistant isolates
compared to susceptible worms was observed, which is consistent with the observations of
[132]. Although glc-2 encodes for the IVM-insensitive β-subunit, these subunits do have the
ability to co-assemble and form glutamate- and IVM-gated heteromeric channels, what makes
them still potential contributors in the ML-resistance mechanism, even if it is only to
compensate [113, 118, 130, 131]. Curiously, in vivo exposure to the MLs did not further
increase the Con-glc-2 levels in CoIVR08 or CoIVR09, but induced small down-regulations
compared to unexposed adults of both isolates. Further, in vivo IVM exposure of both CoIVR08
and CoIVR09 worms resulted in significantly lower Con-glc-6 levels compared to unexposed
CoIVSus adult worms, which might raise ML-resistance. So far, GLC-6 has only been identified
in parasitic nematode species, i.e. C. oncophora and H. contortus. Moreover, expression of an
Hco-GLC-6 subunit in a highly resistant triple mutant C. elegans strain caused the transgenic
worms to become IVM sensitive, suggesting that this subunit is IVM-gated [123].
The presence of mutations associated with ML-resistance was investigated by comparing the
full-length protein sequences of AVR-14B and GLC-6 between 3 regionally different susceptible

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
69
C. oncophora isolates, 2 IVM-resistant Belgian field isolates (CoIVR08 and CoIVR09) and both
resistant isolates after in vivo exposure to IVM or MOX. At first sight, it was noticed that Con-
GLC-6 was more polymorphic between and within species than Con-AVR-14B. This could
suggest that Con-glc-6 is under more evolutionary pressure compared to Con-avr-14B.
Consequently, the sequence variations in glc-6 could underlie natural variations in sensitivity to
anthelmintic drugs, as observed for glc-1 in C. elegans [129]. To be resistance-associated a
mutation needs to be enriched in the resistant populations compared to susceptible
populations and the frequency of the mutation should further increase following exposure to
the drug [124]. However, most of the identified mutations were isolate-specific
polymorphisms. Exposure of the isolates to MLs yielded mutations that were different from the
unexposed isolates, even exposure to IVM or MOX of the same isolates generated different
mutations. Since only 20 clones were sequenced per isolate, the identified mutations are
believed to be the most abundant ones for these genes in the investigated C. oncophora
populations. Further, compared to the findings of [132], after only one single passage of
CoIVSus and CoIVR08 through helminth-free calves, the identified AA substitutions in Con-
AVR-14B were all new, apart from I24T/N. This could suggest a continuous genetic evolution
of these GluCl subunit genes in the worm populations and may actually reflect the
heterogeneity of the investigated parasite populations. Besides, mutations had to be present
in at least 2 sequenced clones, to reduce the probability of identifying sequencing artefacts.
Intriguingly, the I24T/N mutation is located in the signal peptide of Con-AVR-14B. Signal
peptides are essential in cellular trafficking of membrane and secretory proteins, resulting in
the correct translocation across or integration into membranes [206-208]. Moreover, signal
peptide sequences typically have a hydrophobic character and by substitution of the polar
Isoleucine residue, at position 24, by an apolar Threonine or Asparagine residue, the correct
integration of the GluCl receptor could be affected, leading to impaired drug binding and
probably to ML-resistance.
Most mutations were found in the N-terminal extracellular domain and the intracellular loop
between TM3 and TM4, for both Con-AVR-14B and Con-GLC-6. The N-terminal extracellular
domain consists of 10 β-sheets where the cysteine loop and the ligand-binding site are located.
Mutations in this region may affect channel gating – the process whereby agonist-induced
conformational changes in the ligand-binding domain are converted to channel opening and
closure [209]. Regarding Con-AVR-14B, the isolate-specific mutations in the N-terminal domain
at positions 79, 162 and 173 have been identified before in T. circumcincta isolates [133],
but it is unknown if they confer loss in IVM-sensitivity. Similar to the results of [132], the
L256F polymorphism, which has previously been accounted for losses in sensitivity to IVM in
resistant C. oncophora worms [131], could not be determined in any of the unexposed or
exposed C. oncophora isolates investigated. Surprisingly, in this study the V235A mutation
could be identified in CoIVR08 worms exposed to IVM. Previously, the V235A mutation has

CHAPTER 4: Cooperia oncophora glutamate-gated chloride channels in resistance
70
been associated with an IVM-resistant Con-AVR-14B subunit [131], but when expressing a
Con-AVR-14B isoform with this mutation in Xenopus oocytes, responses to glutamate and IVM
were similar to those of the susceptible Con-AVR-14B subunit. The large intracellular TM3-TM4
loop has the highest inter-subunit variability and carries motifs for phosphorylation and
ubiquitination. Possibly, this domain regulates receptor assembly, trafficking, anchoring and
gating of the receptor ion-channel and therefore, changes in the TM3-TM4 loop could affect
the sensitivity to anthelmintics and mediate resistance [210-212].
Recently, the crystal structure of IVM bound to a homopentameric (five GLC-1 subunits) C.
elegans GluCl channel has been published [111]. IVM binds on the periphery of the
transmembrane domains and stabilizes an open-pore conformation. Its site occupies TM3 on
the principle subunit, TM1 on the complementary subunit, the pore-lining TM2 and the TM2-
TM3 loop [111, 213]. Several AA substitutions in the IVM binding site of GluCl channel
subunits have been shown to render ML-resistance in nematodes and insects, but none of
them were observed in the investigated C. oncophora populations [213-218]. In this study,
only 2 isolate-specific mutations were identified in the assumed IVM binding site of Con-AVR-
14B, including the presence of a premature stop codon at the beginning of TM3. For Con-GLC-
6, 7 AA substitutions were determined in the described IVM binding site, but they were either
isolate-specific or present in both susceptible and resistant isolates.
Whether the transcriptional changes of Con-glc-2 and Con-glc-6 are potential resistance
mechanisms warrants further investigation. The identification of multiple isolate-specific
mutations suggests that there is a continuous evolution in the GluCl subunit genes and
therefore support the hypothesis that resistance can arise by recurrent spontaneous
mutations at different geographical locations during the period of selection [83]. Although the
high level of ML-resistance in CoIVR08 and CoIVR09 cannot be explained by any of the
mutations neither in Con-glc-6 nor Con-avr-14B, it cannot be excluded that some mutations
could disrupt the GluCl channel functionality and confer resistance.

CHAPTER 5 Gene expression analysis of ABC transporters in a resistant
Cooperia oncophora isolate following in vivo and in vitro
exposure to macrocyclic lactones

72
Based on: De Graef J, Demeler J, Skuce P, Mitreva M, von Samson-Himmelstjerna G,
Vercruysse J, Claerebout E and Geldhof P. Gene expression analysis of ABC transporters in a
resistant Cooperia oncophora isolate following in vivo and in vitro exposure to macrocyclic
lactones. Parasitology, 2013 (2:1-10).

CHAPTER 5: Cooperia oncophora ABC transporters in resistance
73
5.1 Introduction
Although the molecular mechanisms underlying the development of macrocyclic lactone (ML)-
resistance in helminth parasites remain elusive, the members of the ABC transporter family
(e.g. P-glycoproteins (PGPs), Half-transporters (HAFs) and Multidrug resistant proteins (MRPs))
are thought to play an important role, since they are suspected to affect the absorption,
distribution and elimination of xenobiotics inside nematodes. A large number of ABC
transporter genes has been identified in nematodes. In the free-living model nematode,
Caenorhabditis elegans, 15 pgp genes, 9 haf genes and 8 mrp genes have been identified
[144, 150]. Expression of pgps has been observed in all developmental life stages of C.
elegans [219], in particular in the intestinal cells, but also in the pharynx, the excretory cells
and the chemosensory AWA neurons in the head [219-222]. Functional analyses of mrp-1 and
pgp-1 in C. elegans describe a protective role against the heavy metal ions, cadmium and
arsenite [223]. Cel-pgp-2 has a function in the biogenesis of a lysosome-related fat storage
organelle [224] and Cel-pgp-3 is important in defence against the natural toxins, colchicine
and chloroquine [220]. In parasitic nematode species, the number of identified ABC
transporter genes is still expanding. So far, 9 pgp genes, 1 haf gene and 2 mrp genes have
been described in Haemonchus contortus [140, 144, 151] and 11 partial pgp sequences were
recently identified in Teladorsagia circumcincta [149]. In the human filarial worms, 8 pgp
genes, 8 haf genes and 5 mrp genes are reported for Brugia malayi [225] and, for Onchocerca
volvulus, 4 pgp genes and 3 haf genes have been described [127, 226-229]. The extent to
which the biological role of ABC transporters is conserved between nematode species is still
unclear.
In the first reports that associated ML-resistance in parasitic nematodes with ABC
transporters, higher pgp expression levels or changes in allelic diversity were documented in
resistant H. contortus worms [152, 153]. More recently, a constitutive up-regulation of Hco-
pgp-2 and Hco-pgp-9 was observed in a triple-resistant H. contortus isolate compared to a
susceptible isolate [140]. In T. circumcincta, constitutive differences in gene expression
between a susceptible and a triple-resistant isolate were most notable in Tci-pgp-9, which was
up-regulated in all life cycle stages of the resistant isolate. Also, high levels of polymorphisms
in the partial Tci-pgp-9 nucleotide sequence were identified between the isolates [149]. The
involvement of ABC transporters in the mechanism of ML-resistance has also been described in
C. elegans. Resistant isolates, either generated by ivermectin (IVM)-receptor knock-down (glc-
1/avr-14/avr-15 triple mutant) or through step-wise exposure to non-lethal doses of IVM,
were in vitro cultured with IVM or moxidectin (MOX). Ardelli and Prichard (2008) observed that
IVM and MOX induced similar expression profiles with a marked overexpression of mrp-3, mrp-
5, mrp-7 and mrp-8 [155]. More recently, Yan et al. (2012) described an IVM-induced up-
regulation of pgp-1, pgp-2, pgp-4, pgp-12, pgp-14, mrp-1, mrp-2, mrp-4, mrp-5, mrp-6, mrp-

CHAPTER 5: Cooperia oncophora ABC transporters in resistance
74
7, haf-1, haf-2 and haf-3 [157]. Additionally, the role of PGPs, HAFs and MRPs in protecting C.
elegans from anthelmintic toxicity was investigated in mutant strains (through deletion
mutations or RNAi) exposed to IVM. Cel-mrp-3, Cel-mrp-4 and Cel-mrp-8 may play a role in
protecting the worm from paralysis induced by IVM [155]. Knock-down of Cel-mrp-1 and Cel-
pgp-2 appeared to have the greatest effects in terms of reduced pharyngeal pumping and/or
egg production and motility in response to IVM [157].
Despite the fact that resistance is widespread in C. oncophora, no reports have yet been
published on the potential role of ABC transporters in the development of anthelmintic
resistance in this species. For this reason, the aim of the present study was to investigate the
possible involvement of C. oncophora ABC transporter genes in the resistance mechanism by
analysing constitutive and inducible changes in gene transcription levels between a susceptible
and an IVM-resistant field isolate [19] (CHAPTER 2).
5.2 Materials and methods
5.2.1 Parasite material
Three C. oncophora isolates were used in this trial: � The anthelmintic-susceptible C.
oncophora isolate CoIVSus [132] (CHAPTER 2); � the IVM-resistant C. oncophora field isolate
CoIVR08 [19] and 3 the IVM-resistant C. oncophora field isolate CoIVR09 [132]. All isolates
are maintained in the laboratory by regular passages, without treatment, through helminth-free
calves. Eggs, L3 and adult C. oncophora worms were collected as described in CHAPTER 3. In
vivo exposed resistant adult worms were recovered live at necropsy, 14 days after
subcutaneous treatment with IVM or MOX (0.2 mg/kg bodyweight) during the infection trial
described in CHAPTER 2. Both, IVM and MOX concentrations remain >1 ng/g in gastrointestinal
mucosal tissue for 18 days post-treatment [94, 203]. To obtain in vitro exposed C. oncophora
L3, fresh larvae were harvested from coprocultures and ex-sheathed in 0.5% sodium
hypochlorite (in distilled water) [64]. Third stage larvae were then incubated at 28°C in
deionized water or in 10-8M IVM (8.7 ng/ml IVM) or 10-7M IVM (87 ng/ml IVM). A stock
solution of 10-2M IVM was first prepared in 100% dimethyl sulphoxide (DMSO), while the final
dilutions were made in water. After 24h, the larvae were transferred onto 28 μm sieves
suspended in rows of a 24 well plate. Two hours later, the sieves were carefully lifted out of
the rows, the migrated larvae were collected from the wells and washed 3 times with deionized
water. Pools of larvae (n = 1000) were stored at -80°C until required.
5.2.2 RNA extraction and cDNA synthesis
Total RNA samples were extracted by grinding the parasites in 0.2 ml glass homogenizers
(Wheaton) on ice, followed by a Trizol® extraction (Invitrogen). Residual genomic DNA was
removed by DNase I treatment (Roche). The RNA quality was verified with the Experion™ RNA

CHAPTER 5: Cooperia oncophora ABC transporters in resistance
75
StdSens Starter kit (Bio-Rad) and the RNA concentration was determined using a Nanodrop®
ND-1000 spectrophotometer (NanoDrop Technologies). cDNA was synthesized from 1 μg total
RNA by random priming using the iScript cDNA synthesis kit (Bio-Rad) according to the
manufacturer’s recommendations.
5.2.3 Quantitative real-time PCR
ABC transporter genes were identified as previously described in CHAPTER 3: either by mining
the C. oncophora transcriptome database or by degenerated PCR approach. Quantitative real-
time PCRs were performed to compare the constitutive (without drug exposure) and inducible
(after exposure to IVM or MOX) transcriptional changes of ABC transporter genes between
CoIVSus and CoIVR08 parasite stages. For each biological sample, at least 2 independent RNA
extractions were performed. Total RNA was converted to cDNA and diluted 1/5 or used
undiluted for Con-pgp-11, Con-pgp-12, Con-pgp-16 and Con-mrp-7. Real-time PCR reactions
were prepared with the SYBR Green Master Mix (Applied Biosystems) using 6.4 μl H2O, 0.8 μl of
each amplification primer (10 μM) and 2 μl of cDNA to give a 20 μl reaction volume. All
amplification runs were performed on a StepOnePlus Real-Time PCR System (Applied
Biosystems), under the following conditions: 95°C for 20 s, followed by 40-50 cycles of 95°C
for 5 s, optimal annealing temperature (APPENDIX B) for 20 s and an extension of 72°C for 12
s. A melting curve analysis was performed at the end of the reaction to ensure specificity of
the primers. Each run also included a five-point dilution series of pooled cDNA and a non-
template control. Technical replicates of each sample were performed at least in duplicate
within the same run. For each transcript, the mean Ct value of the replicates was calculated
and then corrected for the run efficiency. Subsequently, Ct values were transformed in relative
quantities using the delta Ct method. The relative quantities were then normalized with the
normalisation factor, obtained by the geNorm software for reference genes Con-gapdh and
Con-tubb [204, 205]. Transcript levels were statistically analysed using an independent-
samples t-test (SPSS Statistics 19). Changes of minimum 2-fold with P<0.05 were regarded as
being significant.
5.3 Results
5.3.1 Analysis of constitutive transcriptional changes of ABC transporter genes
Constitutive differences in transcript levels of the ABC transporter genes between the
susceptible and CoIVR08 resistant isolate were examined in eggs, L3 and adults worm. The
results of the qRT-PCRs are shown in FIGURE 5.1 as the average fold change in mRNA levels
compared to non-exposed susceptible eggs, L3 or adult worms. In the C. oncophora egg stage,
there was considerable variation in the constitutive transcript levels of ABC transporter genes
between biological replicates, but the overall trend showed a higher expression in the eggs of
the CoIVR08 isolate compared to the susceptible eggs. The up-regulation was only significant

CHAPTER 5: Cooperia oncophora ABC transporters in resistance
76
in Con-haf-9 (5.7-fold) and Con-mrp-1 (3.4-fold). None of the genes analysed showed
significant, constitutive differences in transcript levels between susceptible and resistant
CoIVR08 L3 and adult worms.
FIGURE 5.1: Fold changes in constitutive mRNA transcript levels of ABC transporter genes in Cooperia
oncophora eggs (A) L3 (B) and adult worms (C). The transcript levels in susceptible stages have been set
at 1 and the transcript levels ± SD in CoIVR08 eggs, L3 and adult worms expressed relative to this.
Changes of a minimum 2-fold up/down (dotted lines) with P<0.05 were regarded as being statistically
significant (* P<0.05).
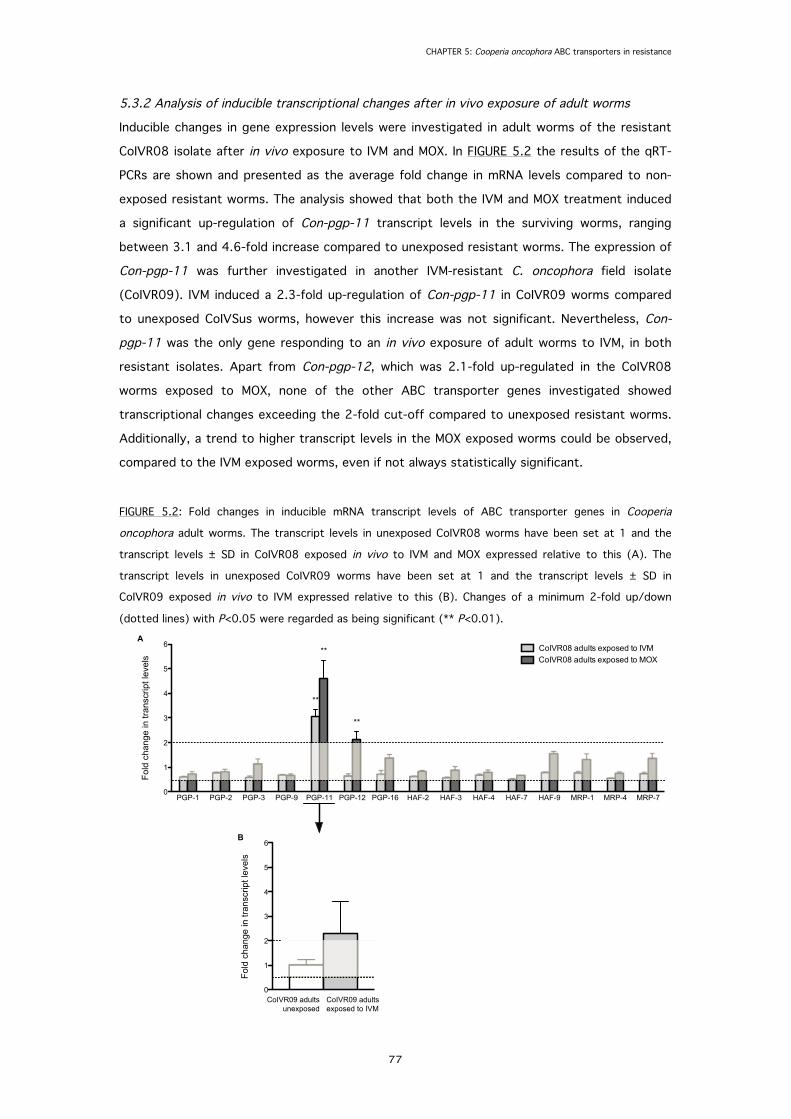
CHAPTER 5: Cooperia oncophora ABC transporters in resistance
77
5.3.2 Analysis of inducible transcriptional changes after in vivo exposure of adult worms
Inducible changes in gene expression levels were investigated in adult worms of the resistant
CoIVR08 isolate after in vivo exposure to IVM and MOX. In FIGURE 5.2 the results of the qRT-
PCRs are shown and presented as the average fold change in mRNA levels compared to non-
exposed resistant worms. The analysis showed that both the IVM and MOX treatment induced
a significant up-regulation of Con-pgp-11 transcript levels in the surviving worms, ranging
between 3.1 and 4.6-fold increase compared to unexposed resistant worms. The expression of
Con-pgp-11 was further investigated in another IVM-resistant C. oncophora field isolate
(CoIVR09). IVM induced a 2.3-fold up-regulation of Con-pgp-11 in CoIVR09 worms compared
to unexposed CoIVSus worms, however this increase was not significant. Nevertheless, Con-
pgp-11 was the only gene responding to an in vivo exposure of adult worms to IVM, in both
resistant isolates. Apart from Con-pgp-12, which was 2.1-fold up-regulated in the CoIVR08
worms exposed to MOX, none of the other ABC transporter genes investigated showed
transcriptional changes exceeding the 2-fold cut-off compared to unexposed resistant worms.
Additionally, a trend to higher transcript levels in the MOX exposed worms could be observed,
compared to the IVM exposed worms, even if not always statistically significant.
FIGURE 5.2: Fold changes in inducible mRNA transcript levels of ABC transporter genes in Cooperia
oncophora adult worms. The transcript levels in unexposed CoIVR08 worms have been set at 1 and the
transcript levels ± SD in CoIVR08 exposed in vivo to IVM and MOX expressed relative to this (A). The
transcript levels in unexposed CoIVR09 worms have been set at 1 and the transcript levels ± SD in
CoIVR09 exposed in vivo to IVM expressed relative to this (B). Changes of a minimum 2-fold up/down
(dotted lines) with P<0.05 were regarded as being significant (** P<0.01).
PGP-1 PGP-2 PGP-3 PGP-9 PGP-11 PGP-12 PGP-16 HAF-2 HAF-3 HAF-4 HAF-7 HAF-9 MRP-1 MRP-4 MRP-70
1
2
3
4
5
6 CoIVR08 adults exposed to IVMCoIVR08 adults exposed to MOX
**
**
**
0
1
2
3
4
5
6
CoIVR09 adults unexposed
CoIVR09 adults exposed to IVM
Fold
cha
nge
in tr
ansc
ript l
evel
s
Fold
cha
nge
in tr
ansc
ript l
evel
s
B
A
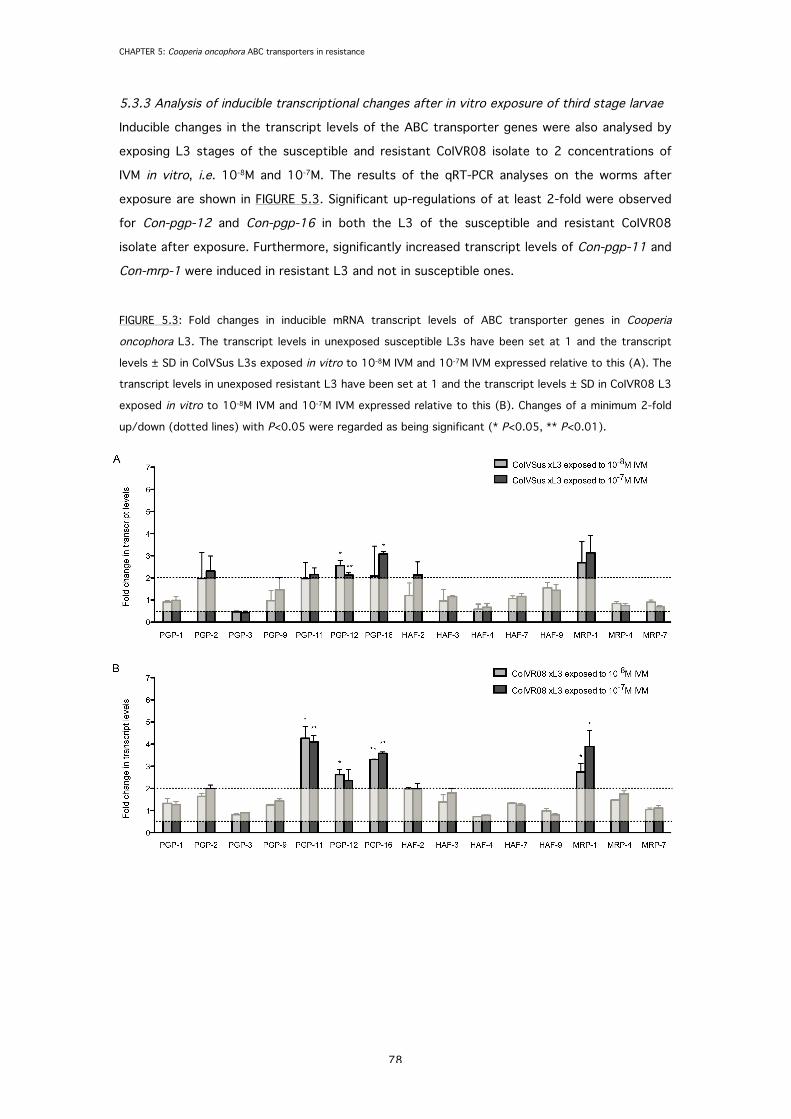
CHAPTER 5: Cooperia oncophora ABC transporters in resistance
78
5.3.3 Analysis of inducible transcriptional changes after in vitro exposure of third stage larvae
Inducible changes in the transcript levels of the ABC transporter genes were also analysed by
exposing L3 stages of the susceptible and resistant CoIVR08 isolate to 2 concentrations of
IVM in vitro, i.e. 10-8M and 10-7M. The results of the qRT-PCR analyses on the worms after
exposure are shown in FIGURE 5.3. Significant up-regulations of at least 2-fold were observed
for Con-pgp-12 and Con-pgp-16 in both the L3 of the susceptible and resistant CoIVR08
isolate after exposure. Furthermore, significantly increased transcript levels of Con-pgp-11 and
Con-mrp-1 were induced in resistant L3 and not in susceptible ones.
FIGURE 5.3: Fold changes in inducible mRNA transcript levels of ABC transporter genes in Cooperia
oncophora L3. The transcript levels in unexposed susceptible L3s have been set at 1 and the transcript
levels ± SD in CoIVSus L3s exposed in vitro to 10-8M IVM and 10-7M IVM expressed relative to this (A). The
transcript levels in unexposed resistant L3 have been set at 1 and the transcript levels ± SD in CoIVR08 L3
exposed in vitro to 10-8M IVM and 10-7M IVM expressed relative to this (B). Changes of a minimum 2-fold
up/down (dotted lines) with P<0.05 were regarded as being significant (* P<0.05, ** P<0.01).

CHAPTER 5: Cooperia oncophora ABC transporters in resistance
79
5.4 Discussion
IVM-resistance has been associated with changes in ABC transporter genes in several parasitic
nematodes [140, 149, 152-154, 226-228, 230, 231], but until now, no reports are available
on the potential role of ABC transporters in the development of ML-resistance in C. oncophora.
In this study, 7 pgp genes, 5 haf genes and 3 mrp genes (CHAPTER 3) were investigated for
their involvement in ML-resistance. It is important to note that since most of the identified
ABC transporter genes were partial, the currently assigned gene names may still change once
more sequence information becomes available.
Differences in constitutive transcript levels were compared between eggs, L3 and adult worms
from a susceptible and resistant isolate. No significant differences in the transcription of ABC
transporter genes were observed between the L3 and adult parasites of the susceptible and
resistant isolates. In the eggs, however, Con-haf-9 and Con-mrp-1 transcript levels were
significantly higher in the resistant isolate compared to the susceptible one. However, further
information on the constitutive expression of these genes in other nematode species is limited.
In laboratory-selected resistant C. elegans worms mrp-1 transcription was highly up-regulated
(20 to 36-fold), this was evident even after 3 months without drug exposure [156]. In H.
contortus and O. volvulus, a constitutive up-regulation of pgp-2 transcription has been
associated with IVM-resistance, whereas in H. contortus and T. circumcincta pgp-9 was
implicated [140, 149, 152, 154, 228]. Interestingly, based on the results of this study,
neither of these PGPs are likely to be involved in the resistance mechanism of C. oncophora,
further suggesting that the resistance mechanism might differ between species.
Although it is often hypothesized that anthelmintic exposure can induce the expression of ABC
transporter genes in nematodes, the experimental evidence is still scarce. In C. elegans,
resistance was associated with the inducible up-regulation of pgp-1, pgp-2, pgp-4, pgp-12,
pgp-14, mrp-1, mrp-2, mrp-3, mrp-4, mrp-5, mrp-6, mrp-7, mrp-8, haf-1, haf-2 and haf-3
after culturing the worms on agar plates with IVM or MOX [155-157]. In H. contortus on the
other hand, no consistent pattern could be discerned in L3 after exposure to IVM or MOX
[140]. In this study, the effect of anthelmintic exposure was investigated in vivo 14 days post
treatment, at which time the worms would still be exposed to active drug [232, 233]. The
most notable change in the surviving worms was observed for pgp-11, in which there was a
significant up-regulation after both IVM and MOX treatment. A smaller but significant effect
was also observed for pgp-12 induced by MOX treatment, but not by IVM. Only one other
study [154] investigated the expression levels of ABC transporters in worms following in vivo
exposure. The authors reported the over-expression of five pgp genes (termed Hco-pgp-A, -B,
-C, -D and –E) in adult H. contortus worms collected 24 hours after the treatment of sheep
with IVM, whereas MOX treatment only resulted in the up-regulation of 2 pgp genes (termed
Hco-pgp-C and -E). Interestingly, according to a phylogenetic analysis, Hco-pgp-B showed most

CHAPTER 5: Cooperia oncophora ABC transporters in resistance
80
homology to the C. elegans pgp-11. Unfortunately, the study did not provide information
regarding the levels of up-regulation of the genes analysed.
Although it is debatable to what extent the in vitro assays can mimic in vivo conditions, the
inducible up-regulation of pgp-11 was also observed in the L3 of the resistant isolate following
in vitro exposure to 2 different concentrations of IVM. Importantly, this up-regulation was not
observed in the larvae of the susceptible isolate, suggesting that the resistant worms have
acquired the ability to up-regulate pgp-11 upon exposure to MLs. On the other hand, pgp-12,
pgp-16 and, to a lesser extent, also mrp-1 were transcriptionally up-regulated in the larvae of
both the susceptible and resistant isolate after exposure, suggesting they are part of a more
general xenobiotic response. The molecular mechanisms involved in the transcriptional
regulation of ABC transporter genes in nematodes are still largely unknown. In mammals, the
transcriptional regulation of the multidrug-resistance ABC transporter gene mdr-1 seems to be
controlled by the nuclear pregnane X receptor (PXR) and the constitutive androstane receptor
(CAR) [234-236]. Several members of the nuclear receptor (NR) superfamily have been
described in C. elegans, of which the nuclear hormone receptor-8 (NHR-8) seems to be
involved in resistance to colchicine and chloroquine [127, 150, 177]. Since both these natural
toxins are substrates for PGP-3 [220], NHR-8 may play some role in the transcriptional
regulation of Cel-pgp-3. However, because of the broad substrate specificity of ABC
transporters, it is plausible that a large number of NRs is involved in the transcriptional
activation of each ABC transporter gene. Besides transcriptional activation, the regulation of
ABC transporters can also occur by post-transcriptional mechanisms such as mRNA
stabilization. Recently, in mouse hepatocytes IVM was shown to prolong the half-life of MDR-1a
and MDR-1b mRNA, leading to the overexpression of P-glycoprotein through post-
transcriptional mRNA stabilization [237].
In summary, the data presented here indicate that resistant C. oncophora worms surviving
exposure to IVM and MOX are able to induce pgp-11 transcription, whereas this is not observed
in susceptible worms. Whether the up-regulation of this particular P-glycoprotein actually helps
to protect the parasites against the toxicity of both MLs is still unclear. Further work is needed
to reveal the genetic basis underpinning this inducible up-regulation and to unravel the
functional role of PGP-11.

CHAPTER 6 General discussion

82

CHAPTER 6: General discussion
83
6.1 Introduction
Macrocyclic lactones (MLs) are the most frequently used family of anthelmintic drugs to
control parasite infections in cattle. Excessive and irrational use of MLs increases the selection
pressure and will eventually lead to ML-resistance. Today, field diagnosis of ML-resistance is
solely based on the faecal egg count reduction test (FECRT). In CHAPTER 2, a comparison
between the FECRT and the gold standard (based on reduction in worm burden) was made to
accurately assess ML-resistance in a field resistant Cooperia oncophora isolate. The results
show that the sensitivity of the FECRT is inferior to the gold standard and that resistance may
be overlooked when treatment suppresses the fecundity of the worms. This makes the
development of a more sensitive molecular test for diagnosing ML-resistance more urgent.
However, despite all the efforts the mechanism of ML-resistance in parasitic nematodes is not
yet understood. It becomes more and more likely that more than 1 gene is responsible for the
resistant phenotype in C. oncophora. This makes it more complicated to elucidate the main
contributor(s) in the mechanism of ML-resistance and slows down the development of a
molecular detection technique for analysing ML-resistance.
In the following paragraphs it is discussed whether the FECRT is still useful in the field, how this
thesis improved our knowledge on the molecular mechanisms of ML-resistance with some
suggestions for future research and whether it will ever be possible to detect ML-resistance in
the field by molecular methods.
6.2 Is the FECRT sti l l useful in the field?
Most Belgian farmers are unaware of the anthelmintic resistance status on their farm, mainly
because they have not encountered any problems yet. Do they need to know? The results of a
field survey, conducted between 2008-2009 on 75 Flemish (Belgium) cattle farms,
demonstrated true resistance on only 1 single farm [18]. This might suggest that farmers
apply sustainable control programs against parasites, which include low treatment frequencies
and maintaining high refugia. Factors such as low parasite pathogenicity and a rapid protective
immune response in the host may also sustain a low prevalence of anthelmintic resistance
[238]. On the other hand, we may not have the adequate means to detect anthelmintic
resistance accurately. It is important to be forethoughtful, since in sheep nematodes it was
demonstrated that once the frequencies of resistance alleles exceed a certain threshold, these
frequencies will increase exponentially. From this stage, the used anthelmintic drugs will be no
longer efficacious against that particular parasite species.
At present, monitoring the efficacy of anthelmintic drugs, such as the MLs, at the farm-level is
solely depending on the FECRT. In CHAPTER 1, several shortcomings of this in vivo method
were elucidated, including a lack of sensitivity, the inability of being species-specific, the

CHAPTER 6: General discussion
84
labour-intensive and time-consuming procedure, confounding factors that bias the outcome of
the test and the difficulty to correctly interpret the results [18, 44, 50, 54-56, 59, 239].
Generally, anthelmintic resistance is declared by a FECR lower than 95% with a lower 95%
confidence limit below 90%. If only 1 of these conditions is met, resistance is suspected [43].
In CHAPTER 2, the accuracy of the FECRT to detect ivermectin (IVM) or moxidectin (MOX)
resistance in a C. oncophora isolate was compared with the controlled efficacy test. The
results clearly showed a big discrepancy between faecal egg counts (FECs) and worms counts,
especially after MOX treatment. This was explained by a significant reduction in the number of
eggs in utero in worms that survived MOX treatment compared to the number of eggs in
worms from untreated controls. MLs are able to paralyse the uterine musculature, however it is
not clear whether the low egg output after MOX treatment is caused by this paralysis or
because a lower number of eggs was produced. Similar observations in C. oncophora after MOX
treatment have been described by [184, 240]. In the early nineties, Scholl et al. [241] also
demonstrated that especially Cooperia species were not fully susceptible to a subcutaneous
injection with MOX, since small numbers of eggs began to appear after 2 weeks post-
treatment, when there had been no opportunity for re-infection. These observations may
suggest that the effect of MOX on the parasite’s fecundity is only temporarily, at least in C.
oncophora. Consequently, the discrepancy between FECs and worm counts could be smaller if
the analyses in CHAPTER 2 were performed at >14 days post MOX treatment.
Although IVM and MOX are believed to exert their anti-parasitic effect by the same mode of
action, some clear pharmacokinetic differences between IVM and MOX have been described, as
well as differences in interactions at the target site in the parasite (i.e. glutamate-gated
chloride channel), which may all play a relevant role on the activity against certain resistant
nematodes [94, 119, 203, 242]. An optimal efficacy against gastrointestinal nematodes might
be achieved following oral administration. This application route achieves the highest drug
concentrations at the target site in the gastrointestinal tract, advances the transcuticular drug
uptake in the worm and hence, enhances the drug efficacy [98-101]. Recently, IVM and MOX
concentrations in the parasitic sheep nematode Haemonchus contortus were determined, after
lambs were intraruminally treated and showed lower MOX concentrations within adult H.
contortus, compared to IVM at 2 days post-treatment [243]. It would be interesting to have
similar data after treatment of infected cattle, to better understand the relationship to the
obtained efficacies. So far, the minimal in vivo drug concentrations required to eliminate
parasites from the host are undetermined.
From the presented data we can conclude that the FECRT is inappropriate for evaluating MOX
efficacy, especially in the cattle nematode C. oncophora, or the efficacy of any other
anthelmintic drug without taking into account the (potentially temporary) sterilizing effect.
Therefore, it would be useful to investigate if MLs other than MOX cause similar effects on the

CHAPTER 6: General discussion
85
parasite’s fecundity, in order to define appropriate timings at which the FECRT can be used for
the detection of ML-resistance.
Additional research is desired in order to optimize and validate the FECRT as a tool to assess
the (lack of) efficacy of anthelmintic drugs. The number of animals sampled and the detection
limit of the test need to be better tailored to the level of infection and the aggregation of egg
excretion. The possibility to use pooled faecal samples should also be examined. Then, the
optimized method should be made uniform in new WAAVP guidelines together with a
standardized FECR formula and statistical analysis [54]. The sooner anthelmintic resistance is
diagnosed, the better. Therefore a promising alternative for the FECRT could be a more
sensitive molecular test, which could also overcome the problem of egg suppression after
treatment, for example by analysing eggs before treatment.
6.3 Molecular background of macrocyclic lactone resistance
To date, no molecular test exists for the detection of ML-resistance. First, a molecular marker
for ML-resistance is required, which distinguishes a genotypically resistant parasite population
from a susceptible one. A molecular marker is a polymorphism that causes structural changes
or changes the transcription patterns of genes that either encode for molecular targets of the
anthelmintic drug or for proteins that affect the concentration or activity of the drug. To be
resistance-associated, the marker needs to be enriched in independently isolated resistant
populations, at least within the same species, compared to susceptible populations. Moreover,
the frequency of the marker should further increase following exposure to the drug [124]. In
this thesis a candidate gene approach was used to investigate the potential involvement of
genes underlying specific and unspecific ML-resistance mechanisms.
6.3.1 The role of glutamate-gated chloride channels in macrocyclic lactone resistance
The pentameric glutamate-gated chloride (GluCl) channels are considered to be the main
targets for MLs to act on. So far, 5 genes have been identified in C. oncophora, encoding for at
least 6 GluCl subunits with putatively different sensitivities to MLs. While investigating the
potential role of these genes in ML-resistance, a remarkable observation was the high amino
acid (AA) heterogeneity of GLC-6 between and within species in comparison with the AVR-14B
protein sequence. Over all C. oncophora isolates investigated, 23 and 96 amino acid (AA)
substitutions were identified, respectively in the Con-AVR-14B and Con-GLC-6 protein
sequences. Genetic diversity provides the raw material for natural selection to act on and leads
to variation in responses to anthelmintic drugs, for example. A possible reason why Con-glc-6
is more polymorphic than Con-avr-14 could be that Con-glc-6 is under more evolutionary
pressure. Recently, Gosh et al. (2012) demonstrated that the elevated level of polymorphism
in glc-1 of the free-living nematode Caenorhabditis elegans played a major role in shaping

CHAPTER 6: General discussion
86
abamectin resistance. It was suggested that this resistance was maintained by a long-term
balancing selection between resistance and worm fitness, rather than an elevated mutation
rate or population subdivision. Exposure to the avermectin-producing bacteria Streptomyces
avermitilis selects for the resistant glc-1 allele, while the absence of toxins confers greater
fitness [129].
According to the criteria for a mutation to be resistance-associated [124], none of the
identified mutations were eligible, since they were either isolate-specific, present in resistant as
well as in susceptible populations, or the frequency of the polymorphism could not be
associated with the level of resistance. Interestingly, compared with the polymorphisms
identified by [132], in the same CoIVSus and CoIVR08 isolates after only one single passage
with or without treatment, the polymorphisms in Con-AVR-14B were all new. This might
suggest a continuous genetic evolution in these worm populations and perhaps in resistance
mechanisms. Generally, the level of polymorphism is related to the mutation rate, the effective
size of the population and migration rates, which are spectacularly high in nematodes [107].
Consequently, it was hypothesized that resistance alleles already existed in the standing
genetic variation of parasite populations and could be selected and increase in frequency after
each drug treatment [244]. In this scenario, all resistance alleles would have a common origin.
However, resistance-conferring polymorphisms could also arise by recurrent spontaneous
mutation, with each event occurring on a different genetic background. Each allele may then
have a widely different set of associated markers [83]. Our findings are more in line with this
last hypothesis.
Further, Con-glc-6 appears to be more transcribed than Con-avr-14. On http://nematode.net
[195], the C. oncophora transcriptome database is partially available (currently only reads
from L3, males and females) and describes 62 reads for Con-glc-6 and only 3 for Con-avr-14B.
Also, from the quantitative real-time experiments it was demonstrated that undiluted cDNA
had to be used for Con-glc-14B to reach similar Ct values as achieved with a 1/5 dilution for
Con-glc-6 cDNA. This finding may suggest that Con-glc-6 is functionally more important for
the worm than Con-avr-14.
Recently, for the first time an X-ray structure was presented of a homomeric GluCl channel
from C. elegans co-cristallized with IVM [111]. IVM binds in the membrane-spanning domains
and stabilizes an open-pore conformation, whereas glutamate binds in the classical
neurotransmitter site in the N-terminal extracellular domain. Considering the structural
differences between IVM and MOX it can be postulated that the interaction of avermectins and
milbemycins is not identical and therefore results in different safety profiles, efficacy profiles
and presumably different resistance-associated markers [242]. Although the identified
polymorphisms in Con-AVR-14B and Con-GLC-6 cannot explain the high level of IVM and/or
MOX resistance in the CoIVR08 and CoIVR09 isolates, it would still be interesting to investigate

CHAPTER 6: General discussion
87
which mutations could affect the GluCl channel functionality or ML-sensitivity. By site-directed
mutagenesis experiments in Con-avr-14B and Con-glc-6, followed by expression of these genes
in Xenopus oocytes, electrophysiological records could be measured in response to IVM, MOX
and glutamate. Alternatively, IVM-resistant (glc-1, avr-14B, avr-15 triple mutant) C. elegans
strains could be transformed with different isoforms of Con-AVR-14B and Con-GLC-6 under a
transcriptional reporter construct to investigate which isoforms are able to restore drug-
sensitivity [123]. Besides, fluorescent protein reporters or antibodies against recombinant
Con-GLC-6 subunits would be helpful to localize the GluCl channel in C. oncophora, since this
parasite-specific subunit has only been described in Haemonchus contortus and C. oncophora
and no functional data is available. Furthermore, the subunit composition of native GluCl
channels in parasite species is still unclear. Now the question arises if GLC-6 is required to
assemble with other subunits to form heteromeric receptors to be fully functional [123], for
example with the IVM-insensitive GLC-2 subunit. In CHAPTER 4, Con-glc-2 mRNA levels were
significantly increased in adult worms of both resistant isolates compared to the susceptible
isolate. Whether this up-regulation of Con-glc-2 transcripts occurs in order to compensate for
a significant constitutive down-regulation, as observed for Con-glc-6 mRNA levels in CoIVR09
worms, is still unclear.
6.3.2 The role of ABC transporters in macrocyclic lactone resistance
IVM and MOX are also known to be substrates for ABC transporters, which are thought to
affect the absorption, distribution and elimination of toxins like ML drugs, and hence, their up-
regulation in parasites could favour the development of anthelmintic resistance [127, 242]. In
CHAPTER 5, the most notable change was observed for Con-pgp-11, which transcript levels
were significantly increased (3- to 5-fold) in resistant C. oncophora worms that survived in
vivo treatment with IVM and MOX. Subsequently, it was shown that this up-regulation was not
caused by a general xenobiotic response, because susceptible L3 were not able to induce this
response following in vitro exposure to IVM. To date, pgp-11 has not been described in any
other parasitic nematode species. Work using fluorescent protein reporters in transgenic C.
elegans has shown that pgp-11 is expressed in the excretory cell and in the intestine, but its
precise role is unknown [150]. It would be interesting to investigate the localization of the
Con-pgp-11 gene product in susceptible and resistant C. oncophora isolates to further
elucidate the role it may play in ML-resistance. This role could also be explored by comparing
the effects of MLs on C. elegans wild-type strains and resistant strains (glc-1, avr-14B, avr-15
triple mutant) both with and without the pgp-11 deletion.
It must be emphasized that the Con-pgp-11 fragment identified in CHAPTER 5 is only 735 bp
long and that its putative name could still change once the full-length sequence is attained.
Sequencing of the full-length coding and genomic sequence of Con-pgp-11 is a priority, now
that this gene has been shown to exhibit statistically significant changes in expression between

CHAPTER 6: General discussion
88
exposed and unexposed resistant worms. The changes in gene expression of Con-pgp-11 also
need to be investigated in other isolates with differing levels of ML-resistance and from
different geographical regions. The observed up-regulation of Con-pgp-11 could be the result
of polymorphisms in transcription factors, changes in the promoter region or in genes further
upstream of the coding sequence. Consequently, gene clusters could be affected
simultaneously. Because of the broad substrate specificity of ABC transporters, it is plausible
that a large number of transcription factors is involved in the transcription activation of ABC
transporter genes. It would be interesting to gain better insights in the transcriptional
modulation pathways, in order to interfere with regulatory elements and improve the ML
activity in the parasite. By comparing the sequences of the regulatory elements between
unexposed susceptible and resistant worms and resistant worms exposed to MLs, resistance-
conferring polymorphisms could be detected.
Further, it is not sure what the impact of the increase in transcription levels could mean at the
PGP-11 protein level and how this further affects the parasite. It has also been shown that co-
administration of MLs and PGP-interfering agents improved treatment efficacy in animals
infected with ML-susceptible or ML-resistant parasitic nematodes, probably due to enhanced
plasma availability [159, 160, 164]. Also, in vitro experiments demonstrated that the
combination of PGP-inhibitors with IVM could restore IVM-sensitivity in resistant nematodes
[127, 165]. Ideally, the inhibitor should have a higher affinity for the parasite’s transporters
rather than the host’s transporters.
By exploring the molecular mechanisms of ML-resistance, it seems like more than 1
mechanism/gene is involved in the development of resistance. This has the advantage that
resistance will develop more slowly, than if resistance would be determined by a single gene
[2]. On the other hand, the multigenic character of resistance makes it more complicated to
elucidate which genes contribute most to the resistance phenotype.
6.4 Prospects for molecular methods to detect macrocyclic lactone
resistance in the field
Even if (a) molecular marker(s) for resistance are identified, developing a PCR-based assay
incorporating these genes will be a tremendous challenge. Theoretically, resistance-conferring
polymorphisms could be accurately detected and quantified (allele frequencies) by the
pyrosequencing assay, whereas transcriptional differences could be measured by quantitative
real-time PCR. The advantages of the pyrosequencing assay are that it is quick and easy to
perform, suitable for testing multiple single nucleotide polymorphisms, has a higher sensitivity
and requires the parasite’s DNA as starting material. However, the equipment required for a
pyrosequencing assay is more expensive and less widely available than quantitative real-time
PCR equipment. Quantitative real-time PCR can be optimized for multiplex diagnosis of mixed

CHAPTER 6: General discussion
89
infections, but the starting material to detect transcriptional changes is RNA, which is more
difficult to prepare and less stable compared to DNA [73]. Furthermore, we need to include a
reference isolate to measure transcriptional changes or how do we otherwise define a
significant up/down-regulation? Whatever method is preferred, the interpretation of the
results of molecular diagnosis is not straightforward. Which frequencies of polymorphisms
correlate with highly/moderate/low resistant parasite populations? How do transcription data
correlate with sequence polymorphisms? Can a polymorphism be detected before any
significant transcriptional change is observed? Moreover, if inducible changes, such as
observed for Con-pgp-11, need to be measured, the timing of exposure and collection of
surviving parasites may be crucial. Sampling should occur preferentially before treatment to
deliver a preventive strategy and to guarantee a representative parasite population for
investigation, since treatment could have a significant influence on the parasite’s fecundity. It
would become too devious if an additional in vitro exposure experiment needs to be optimized
in order to detect inducible changes in transcription levels. The easiest option would be to
design a molecular test based on a polymorphism instead of a (inducible) transcriptional
change.
From a practical point of view, eggs or L3s are the only life stages that are easily accessible
for diagnosis in the field. It is easier to obtain ‘sufficient’ L3s from coprocultures than
recovering fresh eggs and L3s can also be preserved for a longer time. However, it takes 14
days before you harvest the larvae from a coproculture. To obtain a representative parasite
population, a pooled faecal sample could be taken from first grazing season calves after being
at least 1 month on pasture. However, since nematode populations on pastures grazed by
cattle typically consist of several worm species, the molecular detection technique should be
robust across multiple species and therefore a multiplex PCR design with several species-
specific primers/probes needs to be optimized. Next, an extensive evaluation of field
populations and experimentally mixed populations is needed, using the molecular detection
technique and the FECRT to correlate genotype with the expected phenotype. Subsequently, it
should be investigated how such a PCR-based technique could be implemented at the farm
level.
Once a molecular test will be available, it remains unclear at which resistance allele frequencies
we will recommend a farmer to stop using a drug. Does it make sense for a farmer to stop
using a drug which causes almost 95% reduction in egg counts, if a molecular test indicates a
low level of resistance [5]? The FECRT is quite insensitive according to researchers (only
detecting benzimidazole resistance when the frequency of the resistance alleles is above 25%
in the population [42]), because they aim to detect resistance as early as possible, while for
most farmers the threshold for anthelmintic resistance is only reached when clinical treatment
failure occurs. Although it is not known how fast resistance to a certain anthelmintic drug

CHAPTER 6: General discussion
90
develops, the harsh reality is that resistance is irreversible and alternatives are scarce. For
example, the FECRT has been used to monitor the evolution of IVM-resistance on a Belgian
cattle farm and showed a rapid increase of the resistance level in C. oncophora, with
reductions in FECs of 73%, 40% and 0%, respectively in 2006, 2007 and 2008 [19]. This
observation demonstrated that once a certain level of resistant nematodes is established, the
following treatments result in an exponential increase in drug-resistant nematodes [2].
Nowadays, when a reduced anthelmintic efficacy is confirmed to be ML-resistance by the
FECRT, farmers are advised to change to an anthelmintic drug class with a different mode of
action. Unfortunately, only few anthelmintic classes with a different mode of action are
currently available as alternatives for MLs, i.e. the benzimidazoles and the imidazothiazoles.
Recently, anti-parasitic compounds with a novel mode of action, i.e. monepantel and
derquantel (TABLE 1.1), were introduced on the market, but both products are only registered
for use in sheep until now. Of course, when alternative drugs are advised in cases of ML-
resistance, these anthelmintic drugs should also be used with caution, to prevent the
development of resistance against this drug class too (see paragraph 6.5). Further, it should
be stressed that any adjustments in a worm control program are case-specific, since they
depend on the treatment history and the pasture management of the farm. Decision support
systems (based on computer simulations) could make it easier to improve future decisions on
nematode control at the farm level [245, 246].
6.5 Delaying macrocyclic lactone resistance
Anthelmintic resistance develops mainly because of under-dosing, frequent treatments and low
refugia. Dosing animals according to the manufacturer’s recommendations is the first
requirement to reduce the development of anthelmintic resistance. Secondly, farmers should
integrate preventive anthelmintic treatments in the grazing management in order to reduce
the number of treatments required. The main focus should be on the first-grazing season
calves, since they are most susceptible to gastrointestinal nematode infections. Complete
eradication of gastrointestinal parasites on the pasture is not feasible. Instead, a low level of
parasitism must be tolerated to trigger a protective immune response in the host, which will
protect the animals in the following grazing seasons. Measures that can be taken to reduce the
larval pasture contamination, and hence the number of treatments, include mowing, late
turnout on pasture and reduced stocking density [75]. Recently, the importance of the worm
population in refugia for slowing down the development rate of anthelmintic resistance has
been the focus of attention. This population is believed to be under no anthelmintic pressure
and therefore likely to contain a greater proportion of susceptible alleles to provide a reservoir
in which resistant alleles could be diluted. Higher proportions of refugia may be achieved
through a targeted selective treatment (TST) approach, where anthelmintic drugs are only

CHAPTER 6: General discussion
91
administered to heavily infected individuals in the herd [245, 246]. This strategy is based on
the fact that the majority of the worms are in the minority of the animals [247]. For the
successful implementation of the TST approach, it is essential to identify those animals with
the highest worm burdens. Preferably, a preventive TST approach should be pursued.
Unfortunately, for cattle no convenient diagnostics exist to identify individual animals in the
herd that should be treated. FECs could be determined 2 months after the turnout or the
weight gain per animal could be monitored, but both approaches are too labour intensive to be
widely used. It would be interesting if a sensitive molecular test could be integrated in a TST
approach, to identify the most heavily infected animals and simultaneously defining the
resistance status of the parasites.
Another advice farmers could take into account to reduce the development rate of
anthelmintic resistance, is avoiding the use of the same class of anthelmintic drugs every year,
in this way the efficacy will be maximized and the longevity of the compounds will be
prolonged [248]. Recently, the WAAVP guidelines provided a scientific basis upon which to
recommend globally applicable principles, concerning anthelmintic combination products for
use in ruminant livestock and horses [45]. The use of combination products could maximize
the breadth of spectrum, overcome species-specific resistance profiles (dose-limiting species)
and delay the development and spread of resistance when resistance allele frequencies are still
low. Moreover, research on several alternative measures that reduce the dependence on
anthelmintic drugs is also on-going. For example, nematophagous microfungi, such as
Duddingtonia flagrans, could be given in an oral formulation and after passage through the
bovine gastro-intestinal tract they reduce pasture contamination by preying on the pasture
larvae [249, 250]. Because the impact of infection on the protein metabolism typically
exceeds its impact on energy balance, the resilience and productivity of the infected animals
can be improved by protein supplementation [251]. Another alternative measure is the
selective-breeding of animals that have an enhanced resistance to nematode infections [252].
Also, an immunologic control of worm infections through vaccination could be the answer to
anthelmintic resistance. However, despite the identification of several candidate protective
antigens, no commercially available vaccines against gastrointestinal nematode parasites are
currently available [253, 254].
6.6 Conclusion
Since anthelmintic resistance is inheritable, irreversible once fully fixed within a population and
alternatives are scarce, it is of major importance to detect resistance at an early stage with
the aim to maintain the efficacy of the currently available drug classes as long as possible. Due
to its lack of sensitivity and inconsistent egg outputs, the FECRT is less suitable for the
correct diagnosis of anthelmintic resistance. Molecular testing is considered to be the most

CHAPTER 6: General discussion
92
sensitive for diagnosing anthelmintic resistance at an early stage. However, to date molecular
diagnosis is only available for the analysis of benzimidazole resistance in a few parasite species
[44]. Regarding the molecular detection of ML-resistance, we are not there yet. The genetics
of ML-resistance are complex; it becomes more and more likely that ML-resistance is multigenic
and varies between and even within species. Therefore, the understanding of the mechanisms
of drug action and resistance remains an important prerequisite for the development of
improved and more sensitive diagnostics. Proteomics and functional genomics provide
promising resources for prospective research into diagnostic markers, vaccine and drug
discovery, molecular epidemiology and basic parasite biology [255]. Furthermore, farmers and
veterinarians should be informed about the consequences of anthelmintic resistance and the
importance of routinely monitoring the efficacy of the drugs with the FECRT until better
diagnostics are available. To stimulate the implementation of monitoring for anthelmintic
resistance at the farm level, the results from diagnostic assays (FECRT or future molecular
tests) and the corresponding expert advise on worm control should be integrated into decision
support tools (e.g. computer programs) an farm management systems. Until novel methods of
worm control are developed, the reduction of selection pressure, through a TST approach and
adjusted pasture management, remains a key issue to adopt a more sustainable and less
intensive parasite control strategy.

SUMMARY/SAMENVATTING

94

SUMMARY
95
SUMMARY
Worldwide, infections with gastrointestinal nematodes restrict welfare and productivity of
livestock. In temperate climatic regions, the most common gastrointestinal nematodes
infecting grazing cattle are Ostertagia ostertagi and Cooperia oncophora, respectively
parasitizing the abomasum and the small intestines. The control of these bovine parasites
relies heavily on the use of macrocyclic lactones (MLs), such as ivermectin, abamectin,
doramectin, eprinomectin and moxidectin. However, the intensive and frequent administration
of such anthelmintic drugs has led to the selection for ML-resistance in these economically
important gastrointestinal nematodes and therefore restricts the continued use of these drugs
in the future. If resistance occurs against drugs belonging to the same anthelmintic class, one
speaks of side-resistance, whereas cross- & multidrug-resistance refers to resistance against 2
or multiple drugs belonging to different anthelmintic classes.
CHAPTER 1 provides a general overview of the available methods to evaluate the efficacy of
MLs against ruminant nematodes, the current ML-resistance status on Belgian cattle farms,
factors that affect the development of anthelmintic resistance and our current knowledge of
molecular mechanisms of ML-resistance. Under-dosing, frequent treatments and low refugia
facilitate the development of ML-resistance. Using the faecal egg count reduction test
(FECRT), emerging ML-resistance has been reported on Belgian cattle farms in the dose-limiting
species C. oncophora, but not yet in the more pathogenic O. ostertagi. Although the FECRT is
the most practical and commonly used method for diagnosing ML-resistance in the field, its
lack of sensitivity is a serious drawback. It is of major importance that anthelmintic resistance
can be detected at an early stage, to maintain the efficacy of the currently available drug
classes as long as possible. Therefore, a more sensitive molecular detection technique is
urgently needed. One key requirement for the development of such a test is to understand the
resistance mechanisms at the molecular level, in order to identify (a) resistance
marker(s)/gene(s).
The first objective of this thesis was to evaluate the accuracy of the FECRT to assess ML-
resistance in a C. oncophora field isolate and an O. ostertagi laboratory isolate, compared with
the reduction in worm burden after treatment (controlled efficacy test). The results are
presented in CHAPTER 2. Both the FECRT and the controlled efficacy test demonstrated that
these 2 parasitic isolates were resistant against ivermectin (IVM). The IVM-resistant O.
ostertagi isolate was still susceptible to moxidectin (MOX) treatment, as shown by over 99%
reduction in egg counts and worm burdens. The FECRT suggested borderline side-resistance
against MOX in the IVM-resistant C. oncophora isolate, with egg count reductions between
97% (95% CI: 76; 100) on day 7 and 86% (95% CI: 49; 96) on day 14. However, the
controlled efficacy test irrefutably showed side-resistance against MOX, with a decrease of

SUMMARY
96
only 31% (95% CI: -12; 57) in worm numbers of the IVM-resistant C. oncophora isolate. This
discrepancy between egg counts and worm counts could be explained by the fact that MOX
treatment resulted in a significantly lower number of eggs per female C. oncophora worm
(43% reduction) compared to the untreated control group. From these observations it was
concluded that the sensitivity of the FECRT is inferior to the controlled efficacy test and that
resistance may be overlooked when treatment suppresses the fecundity in resistant female
worms.
In order to develop a more sensitive PCR-based test for the diagnosis of ML-resistance, the C.
oncophora transcriptome database was explored thoroughly for the presence of genes that
could be involved in the development of ML-resistance in C. oncophora (CHAPTER 3). MLs
bind on the invertebrate-specific glutamate-gated chloride (GluCl) channels to exert a
paralysing effect on the nematode’s pharyngeal, somatic and uterine musculature. Therefore,
alterations in the GluCl subunit genes are thought to contribute to the development of ML-
resistance. In addition to these target-specific mechanisms of resistance, unspecific
mechanisms involve broad-spectrum detoxification systems (i.e. ABC transporters), which
enhance the removal of anthelmintic compounds from the target site. Members of the ABC
transporter family include P-glycoproteins (PGPs), the Half-transporters (HAFs) and the
Multidrug resistant proteins (MRPs). Partial sequences of 4 GluCl subunit genes (Con-avr-14B,
Con-glc-3, Con-glc-4 and the parasite-specific Con-glc-6) and 15 ABC transporter genes (Con-
pgp-1, Con-pgp-2, Con-pgp-3, Con-pgp-9, Con-pgp-11, Con-pgp-12, Con-pgp-16, Con-haf-2,
Con-haf-3, Con-haf-4, Con-haf-7, Con-haf-9, Con-mrp-1, Con-mrp-4 and Con-mrp-7) were
identified in this research project, either by mining the C. oncophora transcriptome dataset or
with a degenerated PCR approach. The genes identified from the transcriptome database are
believed to be the most highly transcribed members of their gene families in C. oncophora
under normal conditions, excluding Con-pgp-9, Con-pgp-12 and Con-pgp-16, which were only
identified by PCR. Reverse-transcriptase PCR showed that most ABC transporter and GluCl
subunit genes were constitutively transcribed throughout the complete life cycle of C.
oncophora. Only Con-pgp-1, Con-pgp-9, Con-pgp-16, Con-glc-4, Con-glc-6 and Con-avr-14
showed a lower transcription in the egg stage. Furthermore, full-length cDNA sequences were
generated for the newly identified Con-glc-3, Con-glc-4 and Con-glc-6 genes which encode for
predicted protein sequences of respectively 492 amino acids (AA), 503 AA and 444 AA.
In CHAPTER 4, the possible involvement of the identified C. oncophora GluCl subunit genes in
the mechanism of ML-resistance was investigated. First, constitutive and inducible gene
transcription levels were compared between one susceptible and 2 IVM-resistant field isolates
(CoIVR08 and CoIVR09). Although some transcriptional changes were statistically significant,
they were considered to be too small to account for the high levels of IVM-resistance in these
C. oncophora isolates. Besides, some transcriptional changes were inconsistent between the

SUMMARY
97
IVM-resistant C. oncophora isolates. Only the transcription results of Con-glc-2 and Con-glc-6
showed similar trends in both IVM-resistant isolates. Constitutive expression levels of Con-glc-
2, coding for an IVM-insensitive β-subunit, were significantly up-regulated in the IVM-resistant
isolates, 2.9 and 4.7-fold in CoIVR08 and CoIVR09 worms respectively, compared to
susceptible worms. Further, a significant constitutive decrease (1.8-fold) was observed for the
parasite-specific Con-glc-6 in CoIVR09 worms compared to susceptible worms and was
maintained after in vivo IVM exposure of CoIVR09 worms. Also, in vivo exposure to IVM
resulted in a significant down-regulation of Con-glc-6 transcript levels (2.7-fold) in the
CoIVR08 isolate compared to unexposed CoIVR08 and susceptible worms. The down-regulation
of this subunit suggests that GLC-6 may be IVM-sensitive. Whether the transcriptional changes
of Con-glc-2 and Con-glc-6 are potential resistance mechanisms warrants further investigation.
Secondly, the presence of AA substitutions associated with ML-resistance was investigated by
comparing the full-length protein sequences of AVR-14B (in which previously the L256F
polymorphism has been linked to ML-resistance) and GLC-6 between 3 susceptible C.
oncophora isolates from different regions, the 2 IVM-resistant Belgian field isolates (CoIVR08
and CoIVR09) and both resistant isolates after in vivo exposure to IVM or MOX. At first sight, it
was noticed that Con-GLC-6 was more polymorphic between and within species than Con-AVR-
14B, with 96 and 23 AA substitutions and 101 and 23 isoforms identified respectively over all
isolate sequences. This might suggest that Con-glc-6 is under more evolutionary pressure than
Con-avr-14B. The L256F mutation in AVR-14B, which had previously been accounted for
losses in sensitivity to IVM in a resistant C. oncophora isolate from the UK, could not be
detected in any of the (un)exposed C. oncophora isolates investigated. Most of the identified
mutations were either isolate-specific or present in resistant as well as in susceptible
populations and, therefore, could not explain the high level of resistance in the CoIVR08 and
CoIVR09 isolates. Even passaging the isolates through helminth-free calves, with or without
treatment with IVM or MOX, yielded mutations in the isolates that were different from the ones
identified in the same isolates from the previous passage. These observations suggest a
continuous genetic evolution in these worm populations and perhaps in resistance mechanisms.
In CHAPTER 5, the possible involvement of the 15 identified C. oncophora ABC transporter
genes in unspecific mechanisms of ML-resistance was investigated by analysing constitutive
and inducible changes in gene transcription levels between a susceptible isolate and the IVM-
resistant CoIVR08 isolate. Significant constitutive up-regulations compared to the susceptible
isolate were only observed for Con-haf-9 (5.7-fold) and Con-mrp-1 (3.4-fold) in eggs of the
CoIVR08 isolate. None of the genes analysed showed significant, constitutive differences in
transcript levels between susceptible and resistant third stage larvae (L3) or adult worms. The
in vivo effect of anthelmintic exposure was investigated in resistant adult worms, collected
from calves 14 days post treatment with either IVM or MOX. The most notable change in the
surviving worms was observed for Con-pgp-11, in which IVM and MOX induced a significant

SUMMARY
98
3.1- and 4.6-fold up-regulation respectively, compared to unexposed resistant worms.
Furthermore, MOX exposure induced a significant 2.1-fold increase of Con-pgp-12 transcript
levels in CoIVR08 adults compared to unexposed CoIVR08 adult worms. Whether these induced
up-regulations resulted from a general stress response or were resistance-associated was
investigated by comparing the inducible changes in transcript levels of ABC transporter genes
between in vitro exposed susceptible and resistant L3. Based on the EC50 values of the
susceptible and IVM-resistant isolate, L3 of both isolates were in vitro exposed to 10-8M and
10-7M IVM. Significant up-regulations were observed for Con-pgp-12 and Con-pgp-16 in the L3
of both the susceptible and resistant CoIVR08 isolate. Interestingly, a significant 4-fold
increase in transcript levels of Con-pgp-11 was induced following in vitro exposure to IVM in
resistant L3 and not in exposed susceptible larvae. The results suggest that the worms of this
particular CoIVR08 isolate have acquired the ability to specifically up-regulate Con-pgp-11
upon exposure to MLs.
CHAPTER 6 presents the general discussion. First it is discussed whether the FECRT is useful
to detect ML-resistance in the field. From the presented data we can conclude that the FECRT
is inappropriate for evaluating MOX efficacy, or the efficacy of any other anthelmintic drug
without taking into account the temporary sterilizing effect on the female parasites. It must be
emphasized that the FECRT is able to detect anthelmintic resistance only when resistance allele
frequencies are already high in the worm population. As long as no sensitive molecular
alternatives are available, it is advised to optimize and validate the FECRT for a continued
routinely monitoring of anthelmintic efficacy at the farm level. In the meantime, we should
proceed to unravel the molecular mechanisms underlying ML-resistance in parasitic nematodes
in order to identify a molecular marker, which can be included in a molecular detection
technique for ML-resistance. Therefore, suggestions are made for further investigation of the
potential involvement of Con-glc-6 and Con-pgp-11 in the mechanisms of ML-resistance.
Furthermore, the prospects for a molecular based method to detect ML-resistance in the field
are discussed. Detecting allelic variation with the pyrosequencing assay appears to be the
most promising and feasible detection technique. Moreover, an extensive evaluation of the
molecular test and the FECRT in the field is needed to correlate resistance genotypes with the
expected phenotypes. Most challenging will be the implementation of the molecular test at the
farm level and to define a threshold for resistance allele frequencies at which we should
recommend a farmer to stop using a drug. Once it is decided to change to an anthelmintic
drug class with a different mode of action, it is essential to preserve the efficacy of these
drugs by integrating preventive targeted selective treatment strategies in the grazing
management.

SAMENVATTING
99
SAMENVATTING
Wereldwijd beperken infecties met gastro-intestinale nematoden het welzijn en de
productiviteit van de veestapel. De meest voorkomende gastro-intestinale nematoden bij
rundvee in gematigde klimaatzones zijn de lebmaagnematode Ostertagia ostertagi en de dunne
darm nematode Cooperia oncophora. Controle van deze runderparasieten steunt voornamelijk
op het gebruik van breedspectrum anthelminthica met een persisterende werking, met name
de macrocyclische lactones (ML’s) zoals ivermectine, abamectine, doramectine, eprinomectine
en moxidectine. Het intensieve en frequente gebruik van deze anthelminthica heeft echter
geleid tot de selectie van ML-resistentie in deze economisch belangrijke maag-darm nematoden
en beperkt daarom het verdere gebruik van deze anthelminthica. Wanneer resistentie optreedt
tegen anthelminthica van dezelfde klasse spreekt men van ‘side’-resistentie, terwijl
kruisresistentie en multidrug-resistentie verwijzen naar resistentie tegen twee of meer
anthelminthica van verschillende klassen.
HOOFDSTUK 1 geeft een algemeen overzicht van de beschikbare methoden waarmee de
werkzaamheid van ML’s tegen rundernematoden kan geëvalueerd worden, van de huidige
resistentiestatus tegen ML’s op Belgische rundveebedrijven, de verschillende factoren die de
ontwikkeling van anthelminthicumresistentie beïnvloeden en van onze huidige kennis omtrent
de moleculaire mechanismen van ML-resistentie. De ontwikkeling van ML-resistentie wordt
voornamelijk in de hand gewerkt door het toedienen van sub-therapeutische dosissen, het
veelvuldige behandelen en een te laag aandeel van parasieten in refugia (d.w.z. parasieten die
niet blootgesteld worden aan ontwormingsproducten). Tot nu toe werd ML-resistentie op
Belgische rundveebedrijven enkel nog maar gerapporteerd in het ‘dose-limiting’ species C.
oncophora en nog niet in het meer pathogene species O. ostertagi. Hierbij werd de diagnose
van ML-resistentie gesteld met de ‘faecal egg count reduction test’ (FECRT), die gebaseerd is
op de reductie in uitscheiding van wormeieren na behandeling. Op dit moment is de FECRT de
enige beschikbare test om de doeltreffendheid van anthelminthica in het veld te controleren.
Een ernstig nadeel van de FECRT is het gebrek aan gevoeligheid, waardoor resistentie niet
vroegtijdig kan opgespoord worden. Het is echter van groot belang om anthelminthicum-
resistentie zo vroeg mogelijk te detecteren, om zo de doeltreffendheid van de (beperkte)
beschikbare anthelminthica zo lang mogelijk te behouden. Om die reden is een gevoeligere,
moleculaire detectie techniek meer dan welkom. De belangrijkste vereiste voor het ontwikkelen
van zo een test is een moleculaire merker (gen), die een resistente wormpopulatie
onderscheidt van een gevoelige populatie. Om deze merker te vinden moeten we eerst de
onderliggende moleculaire mechanismen van ML-resistentie verder ontrafelen.
De eerste doelstelling van deze thesis was om de accuraatheid van de FECRT om ML-resistentie
te detecteren te vergelijken met de gouden standaard methode (‘controlled efficacy test’), die

SAMENVATTING
100
gebaseerd is op de reductie in het aantal wormen na behandeling. De doeltreffendheid van
ivermectine (IVM) en moxidectine (MOX) werd met beide testen bepaald in een IVM-resistent C.
oncophora veld-isolaat en een IVM-resistent O. ostertagi labo-isolaat. De resultaten zijn
weergegeven in HOOFDSTUK 2. Zowel de FECRT als de ‘controlled efficacy test’ bevestigden
dat beide parasitaire isolaten resistent waren tegen IVM. Het IVM-resistente O. ostertagi isolaat
was nog gevoelig voor MOX, zoals aangetoond werd met reducties in ei-uitscheiding en aantal
wormen van >99%. In het IVM-resistente C. oncophora isolaat werd met de FECRT een
randgeval van ‘side’-resistentie tegen MOX gedetecteerd, met reducties in ei-uitscheiding van
97% en 86%, respectievelijk 7 en 14 dagen na behandeling met MOX. De ‘controlled efficacy
test’ daarentegen toonde veel duidelijker deze ‘side’-resistentie tegen MOX aan, met slechts
een reductie in aantal wormen van 31% in het IVM-resistente C. oncophora isolaat na MOX
behandeling. Deze wanverhouding tussen de reducties in ei-uitscheiding en aantal wormen werd
verklaard door het feit dat er na MOX behandeling significant minder (43%) eieren aanwezig
waren per overlevende vrouwelijke C. oncophora worm, vergeleken met de wormen uit de
onbehandelde controle dieren. Uit deze waarnemingen werd besloten dat de gevoeligheid van
de FECRT betrekkelijk lager ligt dan die van de ‘controlled efficacy test’ en dat met de FECRT
resistentie over het hoofd gezien kan worden wanneer de behandeling de fecunditeit in
resistente vrouwelijke wormen onderdrukt.
Met het oog op de ontwikkeling van een gevoeligere moleculaire test om ML-resistentie te
detecteren, werd de C. oncophora transcriptoom databank onderzocht op de aanwezigheid van
genen die betrokken kunnen zijn bij de ontwikkeling van ML-resistentie (HOOFDSTUK 3). ML’s
binden op de invertebraat-specifieke ‘glutamate-gated chloride’ (GluCl) kanalen om hun
paralyserend effect uit te oefenen op de faryngeale, somatische en uteriene spieren van de
nematode. Daarom worden wijzigingen in deze GluCl subunit genen verondersteld bij te dragen
aan de ontwikkeling van ML-resistentie. Naast deze doelwit-specifieke mechanismen van ML-
resistentie bestaan er ook niet-specifieke mechanismen. Zo bestaan er breedspectrum
detoxificatie systemen (bijv. ABC transporters), die de verwijdering van anthelminthica uit de
‘target site’ bevorderen. P-glycoproteïnen (PGP’s), Half-transporters (HAF’s) en ‘Multidrug
resistant proteins’ (MRP’s) behoren tot de ABC transporter familie. In dit onderzoeksproject
werden, hetzij door het onderzoeken van de C. oncophora transcriptoom databank ofwel door
een gedegenereerde PCR aanpak, de partiële sequenties van 4 GluCl subunit genen (Con-avr-
14B, Con-glc-3, Con-glc-4 en het parasiet-specifieke Con-glc-6) en 15 ABC transporter genen
(Con-pgp-1, Con-pgp-2, Con-pgp-3, Con-pgp-9, Con-pgp-11, Con-pgp-12, Con-pgp-16, Con-
haf-2, Con-haf-3, Con-haf-4, Con-haf-7, Con-haf-9, Con-mrp-1, Con-mrp-4 en Con-mrp-7)
geïdentificeerd. Met uitzondering van Con-pgp-9, Con-pgp-12 en Con-pgp-16 werden
bovenvermelde genen allemaal opgepikt uit de C. oncophora transcriptoom databank en
worden ze daarom verondersteld van de meest overgeschreven genen uit hun genenfamilies te
zijn onder normale omstandigheden. ‘Reverse-transcriptase’ PCR toonde aan dat de meeste

SAMENVATTING
101
ABC transporter genen en GluCl subunit genen constitutief tot expressie worden gebracht
doorheen de complete levenscyclus van C. oncophora. Alleen Con-pgp-1, Con-pgp-9, Con-pgp-
16, Con-glc-4, Con-glc-6 en Con-avr-14 vertoonden een lagere transcriptie in het ei stadium.
Verder werden ook nog de volledige cDNA sequenties gegenereerd van de nieuw
geïdentificeerde GluCl subunit genen, Con-glc-3, Con-glc-4 en Con-glc-6, die codeerden voor
voorspelde eiwitsequenties van respectievelijk 492 aminozuren (AA), 503 AA en 444 AA.
In HOOFDSTUK 4 werd de mogelijke betrokkenheid van de geïdentificeerde C. oncophora
GluCl subunit genen in ML-resistentie onderzocht. Eerst werden constitutieve en induceerbare
mRNA transcriptieniveaus in adulte wormen vergeleken tussen 1 gevoelig en 2 IVM-resistente
C. oncophora veldisolaten (CoIVR08 en CoIVR09). Over het algemeen waren de veranderingen
in transcriptieniveaus te klein om de hoge graad van IVM-resistentie in deze isolaten te kunnen
verklaren, ook al bleken sommige verschillen statistisch significant. Bovendien waren de
veranderingen in transcriptie niet altijd consistent tussen de 2 IVM-resistente C. oncophora
isolaten. Enkel voor Con-glc-2 en Con-glc-6 werden gelijkaardige trends in de
transcriptieprofielen van CoIVR08 en CoIVR09 waargenomen. Van Con-glc-2, dat codeert voor
de IVM-ongevoelige β-subunit, werden significante, constitutieve 2.9- en 4.7-voudig op-
regulaties waargenomen in respectievelijk CoIVR08 en CoIVR09, vergeleken met de gevoelige
wormen. Daarnaast werd er een significante, constitutieve neer-regulatie (1.8-voudig)
geobserveerd van Con-glc-6 in CoIVR09 adulte wormen, zowel in aan- als afwezigheid van IVM.
Verder werd er ook nog een significante neer-regulatie van Con-glc-6 waargenomen (2.7-
voudig) na IVM blootstelling van het CoIVR08 isolaat, in vergelijking met niet-blootgestelde
CoIVR08 en gevoelige wormen. Deze geïnduceerde neer-regulatie suggereert dat GLC-6
mogelijk een IVM-gevoelige subunit is. Toch blijft verder onderzoek nodig om uit te maken of
de veranderingen in Con-glc-2 en Con-glc-6 transcriptie betrokken zijn in het mechanisme van
ML-resistentie. Ten tweede werden de volledige eiwitsequenties van AVR-14B en GLC-6
vergeleken tussen 3 gevoelige C. oncophora isolaten (uit België en het Verenigd Koninkrijk), 2
Belgische IVM-resistente C. oncophora veldisolaten (CoIVR08 en CoIVR09) en beide resistente
isolaten na in vivo blootstelling aan IVM of MOX. Daarbij werd gezocht naar AA veranderingen
die mogelijk geassocieerd zijn met ML-resistentie. Op het eerste zicht viel het op dat Con-GLC-
6 aanzienlijk meer polymorf was tussen en binnen species dan Con-AVR-14B. Zo werden er
respectievelijk 96 en 23 AA veranderingen en 101 en 23 isovormen geïdentificeerd over alle
onderzochte isolaat sequenties in Con-GLC-6 en Con-AVR-14B. De L256F mutatie in AVR-14B
die eerder al geassocieerd werd met IVM-resistentie in een Brits IVM-resistent C. oncophora
isolaat, werd niet gevonden in de hier onderzochte (niet)-blootgestelde C. oncophora isolaten.
De meeste van de gevonden mutaties waren ofwel isolaat-specifiek ofwel aanwezig in zowel de
gevoelige als resistente isolaten. Geen enkele mutatie kon rechtstreeks in verband gebracht
worden met de hoge graad van IVM-resistentie in de CoIVR08 en CoIVR09 isolaten. Bovendien
verschilden de gevonden mutaties binnen hetzelfde isolaat al na 1 enkele passage (met of

SAMENVATTING
102
zonder IVM/MOX behandeling) in helminth-naïeve kalveren. Deze waarnemingen voorspellen dat
deze worm populaties onder een continue genetische evolutie staan en dat daardoor ook hun
ontwikkelde resistentiemechanismen voortdurend veranderen.
De mogelijke betrokkenheid van de 15 geïdentificeerde C. oncophora ABC transporter genen in
niet-specifieke mechanismen van ML-resistentie werd onderzocht in HOOFDSTUK 5, door het
analyseren van verschillen in constitutieve en induceerbare mRNA transcriptieniveaus tussen
een gevoelig C. oncophora isolaat en het IVM-resistente CoIVR08 veldisolaat. In vergelijking
met het gevoelige C. oncophora isolaat, werd er enkel een significante, constitutieve op-
regulatie van Con-haf-9 (5.7-voudig) en Con-mrp-1 (3.4-voudig) waargenomen in eieren van
het CoIVR08 isolaat. Er werden geen significante transcriptie verschillen gevonden in L3 en
adulte wormen tussen de isolaten. Om het effect van in vivo blootstelling aan anthelminthica
na te gaan, werden resistente adulte wormen verzameld van kalveren 14 dagen na behandeling
met IVM of MOX. De meest opvallende verandering in de overlevende CoIVR08 wormen was
een significante, geïnduceerde 3.1- en 4.6-voudig op-regulatie van Con-pgp-11transcriptie na
behandeling met respectievelijk IVM en MOX, in vergelijking met niet-blootgestelde CoIVR08
wormen. Daarnaast leidde de MOX behandeling ook nog tot een significante 2.1-voudige
toename van Con-pgp-12 transcriptieniveaus in CoIVR08 wormen in vergelijking met niet-
blootgestelde CoIVR08 wormen. Vervolgens werd onderzocht of deze op-regulaties
voortvloeiden uit een algemene stressrespons of geassocieerd konden worden met IVM-
resistentie. Hiervoor werden de induceerbare veranderingen in ABC transporter transcriptie-
niveaus vergeleken tussen in vitro blootgestelde gevoelige en resistente L3. Op basis van de
EC50 waarden van het gevoelige en IVM-resistente C. oncophora isolaat, werden de L3 van
beide isolaten in vitro blootgesteld aan concentraties van 10-8M en 10-7M IVM. Zowel in L3 van
het gevoelige als het CoIVR08 isolaat werden significante op-regulaties geïnduceerd van Con-
pgp-12 en Con-pgp-16 in vergelijking met niet-blootgestelde L3. Interessanter was echter een
significante 4-voudige toename in Con-pgp-11 transcriptie die enkel geïnduceerd werd na IVM
blootstelling in IVM-resistente L3 en niet na IVM blootstelling in gevoelige L3. Deze resultaten
wijzen erop dat de wormen van dit CoIVR08 isolaat de mogelijkheid hebben verworven om Con-
pgp-11 transcriptie op te reguleren na blootstelling aan ML’s.
HOOFDSTUK 6 geeft de algemene discussie weer, waarin eerst besproken werd of de FECRT
wel bruikbaar is om ML-resistentie op te sporen in het veld. Er werd besloten dat de FECRT
ongeschikt is om de doeltreffendheid van MOX of eender welk ander anthelminthicum te
evalueren, zonder het tijdelijke steriliserende effect op fecunditeit van de vrouwelijke
parasieten in acht te nemen. Toch moet benadrukt worden dat de FECRT wel degelijk
anthelminthicumresistentie kan opsporen, maar alleen wanneer de resistentie allelfrequenties al
een hoge prevalentie hebben in de wormpopulatie. Daarom blijft het ook wenselijk om de
FECRT verder te optimaliseren en te valideren zolang er geen gevoelige moleculaire

SAMENVATTING
103
alternatieven beschikbaar zijn, om zo toch voor een routinematige opvolging te zorgen van de
anthelminthicum werkzaamheid op rundveebedrijven. Ondertussen wordt er ook verder
gewerkt aan de ontcijfering van de moleculaire mechanismen die aan de basis kunnen liggen
van ML-resistentie in parasitaire nematoden. Daarom werden er enkele suggesties gemaakt
voor verder onderzoek naar de mogelijke betrokkenheid van Con-glc-6 en Con-pgp-11 in de
mechanismen van ML-resistentie. Op die manier hopen we een moleculaire merker te vinden die
gebruikt kan worden in een moleculaire test om ML-resistentie te detecteren. Verder werden
ook de vooruitzichten van een moleculaire detectietechniek in het veld besproken. Het
opsporen van allelische variatie door middel van ‘pyrosequencing’ lijkt de meest veelbelovende
en haalbare techniek. Vervolgens zal er een uitgebreide evaluatie nodig zijn van de moleculaire
test tegenover de FECRT in het veld, om het resistentie genotype en het verwachte fenotype
met elkaar te kunnen correleren. De grootste uitdaging wordt wellicht een zinvolle
implementatie van de moleculaire test op het landbouwbedrijf. Bij welke limiet van resistente
allelfrequenties gaan we immers een landbouwer aanraden om het gebruik van een
anthelminthicum te stoppen? Als eenmaal beslist is om over te schakelen naar een
anthelminthicumklasse met een ander werkingsmechanisme, is het essentieel om de
doeltreffendheid van deze nieuwe anthelminthica te behouden, door een doelgerichte,
selectieve behandelingsstrategie te integreren in een optimaal begrazingsbeheer.

104
APPENDIX A
Individual data for faecal egg counts (FECs) on days 0, 7 and 14 post-treatment (in eggs per gram), % Cooperia oncophora and Ostertagia ostertagi larvae
from coprocultures on days 0, 7 and 14 post-treatment, worm burdens and mean in utero egg counts in 10 surviving female C. oncophora worms per animal.
Animal ID Treatment FECs
D0
% C. onc.
L3
% O. ost.
L3
FECs D7 P-T
% C. onc.
L3
% O. ost.
L3
FECs D14 P-T
% C. onc.
L3
C. onc. worm
burden
Arithm. mean eggs in utero in 10 C. onc.
% O. ost.
L3
O. ost. worm
burden
8446 Control 850 88 12 200 80 20 500 91 6650 85.5 (66-115) 9 15450
8531 Control 850 70 30 300 62 38 250 74 4200 106.1 (77-135) 26 8200
8615 Control 550 42 58 100 10 90 200 13 7000 12.7 (0-47) 87 13950
6871 Control 300 95 5 100 6 94 50 8 9750 4 (0-12) 92 9650
3672 Control 200 56 44 0 8 92 150 73 5450 66.4 (51-89) 27 11950
9914 Control 850 74 26 300 70 30 400 70 7900 55.9 (28-69) 30 6350
2310 Control 1000 65 35 200 88 12 500 85 7450 75.8 (55-92) 15 11300
8872 Control 650 48 52 500 40 60 550 65 7150 92.3 (75-132) 35 11850
8199 Control 950 71 29 350 86 14 350 76 9050 56.3 (34-75) 24 9900
8401 Control 1100 80 20 700 85 15 750 89 6600 114 (69-155) 11 12150
8196 Ivermectin 150 3 97 50 0 100 100 1 550 / 99 4050
0986 Ivermectin 550 86 14 50 60 40 0 2 2600 1.3 (0-9) 98 1850
1920 Ivermectin 550 51 49 50 48 52 200 67 7700 19.6 (0-53) 33 3200
9275 Ivermectin 300 80 20 250 78 22 150 53 6500 42.3 (4-85) 47 3100
0944 Ivermectin 1400 84 16 300 100 0 250 100 3650 114.1 (91-151) 0 50
1591 Ivermectin 450 61 39 0 37 63 0 43 1800 24.9 (0-67) 57 800
4248 Ivermectin 1050 53 47 50 45 55 300 89 4900 103.2 (75-141) 11 1500
3304 Ivermectin 800 83 17 100 97 3 50 96 5650 25.5 (10-38) 4 550
8197 Ivermectin 850 93 7 550 96 4 350 98 5350 89.1 (52-121) 2 1150
2783 Ivermectin 1300 62 38 350 87 13 150 90 5700 93 (49-128) 10 1300
105
Animal ID Treatment FECs
D0
% C. onc.
L3
% O. ost.
L3
FECs D7 P-T
% C. onc.
L3
% O. ost.
L3
FECs D14 P-T
% C. onc.
L3
C. onc. worm
burden
Arithm. mean eggs in utero in 10 C. onc.
% O. ost.
L3
O. ost. worm burden
3555 Moxidectin 800 84 16 0 100 0 0 100 200 18.5 (0-38) 0 0
5522 Moxidectin 1000 77 23 0 99 1 100 100 3300 51.7 (13-98) 0 50
8606 Moxidectin 2000 87 13 50 97 3 0 99 8650 42.5 (0-85) 1 50
5982 Moxidectin 700 75 25 0 0 0 0 0 250 31.7 (0-64) 100 0
5597 Moxidectin 1450 65 35 0 100 0 200 100 6600 47.3 (8-83) 0 0
1888 Moxidectin 900 84 16 0 0 0 0 72 1450 30.25 (1-56) 28 0
8489 Moxidectin 900 70 30 0 100 0 0 99 6450 61 (34-102) 1 0
5289 Moxidectin 750 75 25 0 71 29 0 100 9000 25.4 (0-55) 0 0
2816 Moxidectin 800 67 33 0 98 2 0 98 8100 16.4 (0-64) 2 0
3107 Moxidectin 700 59 41 0 94 6 100 96 5150 57 (20-90) 4 200
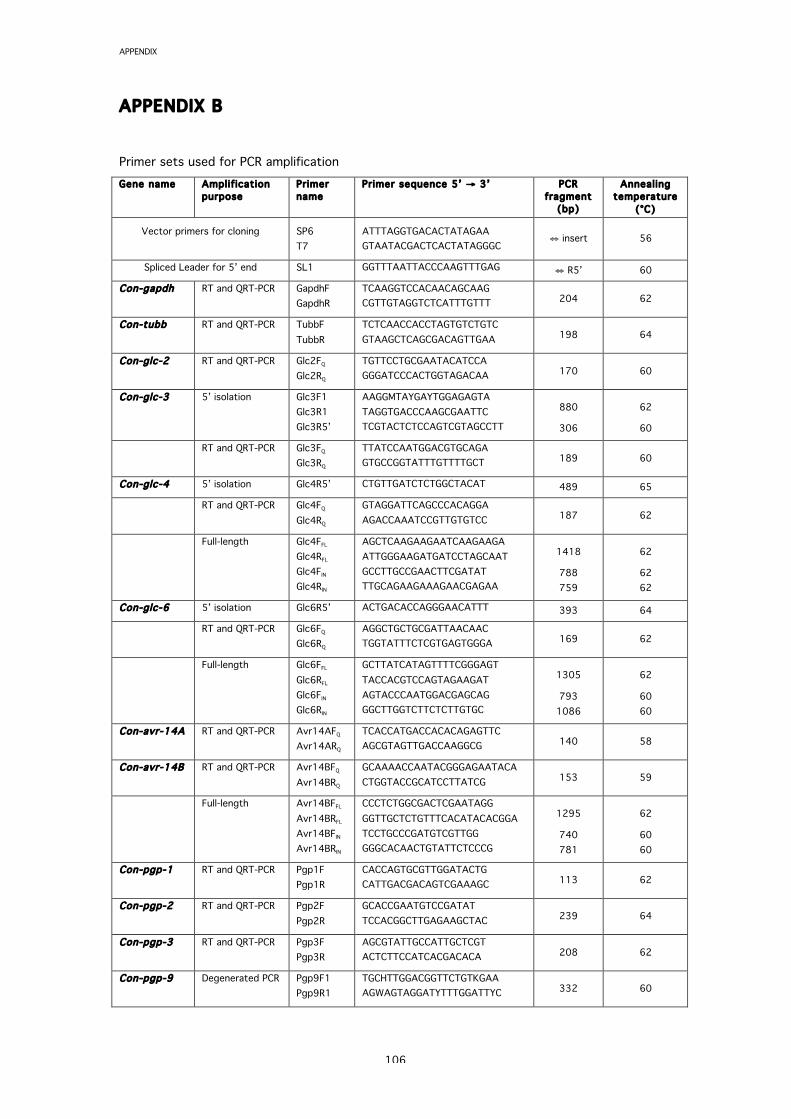
APPENDIX
106
APPENDIX B
Primer sets used for PCR amplification Gene name Amplification
purpose Primer name
Primer sequence 5’ → 3’ PCR fragment
(bp)
Annealing temperature
(°C)
Vector primers for cloning
SP6 T7
ATTTAGGTGACACTATAGAA GTAATACGACTCACTATAGGGC ⇔ insert 56
Spliced Leader for 5’ end SL1 GGTTTAATTACCCAAGTTTGAG ⇔ R5’ 60
Con-gapdh RT and QRT-PCR GapdhF GapdhR
TCAAGGTCCACAACAGCAAG CGTTGTAGGTCTCATTTGTTT 204 62
Con-tubb RT and QRT-PCR TubbF TubbR
TCTCAACCACCTAGTGTCTGTC GTAAGCTCAGCGACAGTTGAA 198 64
Con-glc-2 RT and QRT-PCR Glc2FQ
Glc2RQ TGTTCCTGCGAATACATCCA GGGATCCCACTGGTAGACAA 170 60
Con-glc-3 5’ isolation Glc3F1 Glc3R1 Glc3R5’
AAGGMTAYGAYTGGAGAGTA TAGGTGACCCAAGCGAATTC TCGTACTCTCCAGTCGTAGCCTT
880
306
62
60
RT and QRT-PCR Glc3FQ
Glc3RQ TTATCCAATGGACGTGCAGA GTGCCGGTATTTGTTTTGCT 189 60
Con-glc-4 5’ isolation Glc4R5’ CTGTTGATCTCTGGCTACAT 489 65
RT and QRT-PCR Glc4FQ
Glc4RQ GTAGGATTCAGCCCACAGGA AGACCAAATCCGTTGTGTCC 187 62
Full-length Glc4FFL
Glc4RFL
Glc4FIN
Glc4RIN
AGCTCAAGAAGAATCAAGAAGA ATTGGGAAGATGATCCTAGCAAT GCCTTGCCGAACTTCGATAT TTGCAGAAGAAAGAACGAGAA
1418
788 759
62
62 62
Con-glc-6 5’ isolation Glc6R5’ ACTGACACCAGGGAACATTT 393 64
RT and QRT-PCR Glc6FQ
Glc6RQ AGGCTGCTGCGATTAACAAC TGGTATTTCTCGTGAGTGGGA 169 62
Full-length Glc6FFL
Glc6RFL
Glc6FIN Glc6RIN
GCTTATCATAGTTTTCGGGAGT TACCACGTCCAGTAGAAGAT AGTACCCAATGGACGAGCAG GGCTTGGTCTTCTCTTGTGC
1305
793 1086
62
60 60
Con-avr-14A RT and QRT-PCR Avr14AFQ
Avr14ARQ TCACCATGACCACACAGAGTTC AGCGTAGTTGACCAAGGCG 140 58
Con-avr-14B RT and QRT-PCR Avr14BFQ
Avr14BRQ GCAAAACCAATACGGGAGAATACA CTGGTACCGCATCCTTATCG 153 59
Full-length Avr14BFFL
Avr14BRFL
Avr14BFIN
Avr14BRIN
CCCTCTGGCGACTCGAATAGG GGTTGCTCTGTTTCACATACACGGA TCCTGCCCGATGTCGTTGG GGGCACAACTGTATTCTCCCG
1295
740 781
62
60 60
Con-pgp-1 RT and QRT-PCR Pgp1F Pgp1R
CACCAGTGCGTTGGATACTG CATTGACGACAGTCGAAAGC 113 62
Con-pgp-2 RT and QRT-PCR Pgp2F Pgp2R
GCACCGAATGTCCGATAT TCCACGGCTTGAGAAGCTAC 239 64
Con-pgp-3 RT and QRT-PCR Pgp3F Pgp3R
AGCGTATTGCCATTGCTCGT ACTCTTCCATCACGACACA 208 62
Con-pgp-9 Degenerated PCR Pgp9F1 Pgp9R1
TGCHTTGGACGGTTCTGTKGAA AGWAGTAGGATYTTTGGATTYC 332 60
APPENDIX
107
Gene name Amplification purpose
Primer name
Primer sequence 5’ → 3’ PCR fragment
(bp)
Annealing temperature
(°C) Con-pgp-9 RT and QRT-PCR Pgp9FQ
Pgp9RQ CGATCCTTTTCGACAGATCC GCAATGGCTATCCGTTGTTT 202 60
Con-pgp-11 RT and QRT-PCR Pgp11F Pgp11R
TCGGGAAAGAGTACGATAAT TCGATAGCATCATCCCTTGA 215 60
Con-pgp-12 RT and QRT-PCR Pgp12F Pgp12R
GCGGCACTGATTATCTCGTT CCATTTCCTCTTGACCATTGA 217 60
Con-pgp-16 RT and QRT-PCR Pgp16F Pgp16R
GGAAAGGTATCCGTCGATGA GGTAGCCCTGGCTTAGGTTC 217 62
Con-haf-2 RT and QRT-PCR Haf2F Haf2R
GGTTTGATGGAATGCGTAGG CCAGCCTCAACAACCAAATC 191 62
Con-haf-3 RT and QRT-PCR Haf3F Haf3R
ATAGAAGAGGCTGCGGAACA ATTGAGAGCCTTCCTGACCA 225 62
Con-haf-4 RT and QRT-PCR Haf4F Haf4R
ACAAGAATCTCGATGGCAAA GCAGAATGCGTGAAACTTGA 292 60
Con-haf-7 RT and QRT-PCR Haf7F Haf7R
GTCGGAAGCTCAAGTCCAAG AGGCCTTCAGGTTCTTCCAT 174 64
Con-haf-9 RT and QRT-PCR Haf9F Haf9R
GATGTCAGGCGGTCAAAAAC CGATGGGCGATGAGAACTAC 182 60
Con-mrp-1 RT and QRT-PCR Mrp1F Mrp1R
GCTGAAACCGATTCCCTTCT TTCCATAGAAAACGCCTTCG 190 62
Con-mrp-4 RT and QRT-PCR Mrp4F Mrp4R
ACGCTCTTGAAATGGCAAAC CTCCGAATCGTACGCTGAAT 220 60
Con-mrp-7 RT and QRT-PCR Mrp7F Mrp7R
GATCAACGAAGGTGGTGAAA CAGCAGTGCCTGAGGTGAATC 280 60

REFERENCES
108
REFERENCES
[1] McKellar QA, Jackson F. Veterinary anthelmintics: old and new. Trends Parasitol. 2004 (20:456-61).
[2] Barnes EH, Dobson RJ, Barger IA. Worm control and anthelmintic resistance: adventures with a model. Parasitol Today. 1995 (11:56-63).
[3] Sangster NC. Anthelmintic resistance: past, present and future. Int J Parasitol. 1999 (29:115-24; discussion 37-8).
[4] Chandrawathani P, Waller PJ, Adnan M, Hoglund J. Evolution of high-level, multiple anthelmintic resistance on a sheep farm in Malaysia. Tropical animal health and production. 2003 (35:17-25).
[5] Kaplan RM, Vidyashankar AN. An inconvenient truth: Global worming and anthelmintic resistance. Vet Parasitol. 2012 (186:70-8).
[6] McKenna PB. Update on the prevalence of anthelmintic resistance in gastrointestinal nematodes of sheep in New Zealand. N Z Vet J. 2010 (58:172-3).
[7] Sargison ND, Jackson F, Wilson DJ, Bartley DJ, Penny CD, Gilleard JS. Characterisation of milbemycin-, avermectin-, imidazothiazole- and benzimidazole-resistant Teladorsagia circumcincta from a sheep flock. Vet Rec. 2010 (166:681-6).
[8] Torres-Acosta JF, Mendoza-de-Gives P, Aguilar-Caballero AJ, Cuellar-Ordaz JA. Anthelmintic resistance in sheep farms: Update of the situation in the American continent. Vet Parasitol. 2012 (189:89-96).
[9] Van Wyk JA, Stenson MO, Van der Merwe JS, Vorster RJ, Viljoen PG. Anthelmintic resistance in South Africa: surveys indicate an extremely serious situation in sheep and goat farming. The Onderstepoort journal of veterinary research. 1999 (66:273-84).
[10] Geary TG. Ivermectin 20 years on: maturation of a wonder drug. Trends Parasitol. 2005 (21:530-2).
[11] Kaplan RM. Drug resistance in nematodes of veterinary importance: a status report. Trends Parasitol. 2004 (20:477-81).
[12] Blake N, Coles G. Flock cull due to anthelmintic-resistant nematodes. Vet Rec. 2007 (161:36).
[13] Anziani OS, Suarez V, Guglielmone AA, Warnke O, Grande H, Coles GC. Resistance to benzimidazole and macrocyclic lactone anthelmintics in cattle nematodes in Argentina. Vet Parasitol. 2004 (122:303-6).
[14] Coles GC, Stafford KA, MacKay PH. Ivermectin-resistant Cooperia species from calves on a farm in Somerset. Vet Rec. 1998 (142:255-6).
[15] Coles GC, Watson CL, Anziani OS. Ivermectin-resistant Cooperia in cattle. Vet Rec. 2001 (148:283-4).
[16] Demeler J, Van Zeveren AM, Kleinschmidt N, Vercruysse J, Hoglund J, Koopmann R, Cabaret J, Claerebout E, Areskog M, von Samson-Himmelsjerna G. Monitoring the efficacy of ivermectin and albendazole against gastro intestinal nematodes of cattle in Northern Europe. Vet Parasitol. 2009 (160:109-15).

REFERENCES
109
[17] Edmonds MD, Johnson EG, Edmonds JD. Anthelmintic resistance of Ostertagia ostertagi and Cooperia oncophora to macrocyclic lactones in cattle from the western United States. Vet Parasitol. 2010 (170:224-9).
[18] El-Abdellati A, Charlier J, Geldhof P, Levecke B, Demeler J, von Samson-Himmelstjerna G, Claerebout E, Vercruysse J. The use of a simplified faecal egg count reduction test for assessing anthelmintic efficacy on Belgian and German cattle farms. Vet Parasitol. 2010 (169:352-7).
[19] El-Abdellati A, Geldhof P, Claerebout E, Vercruysse J, Charlier J. Monitoring macrocyclic lactone resistance in Cooperia oncophora on a Belgian cattle farm during four consecutive years. Vet Parasitol. 2010 (171:167-71).
[20] Gasbarre LC, Smith LL, Lichtenfels JR, Pilitt PA. The identification of cattle nematode parasites resistant to multiple classes of anthelmintics in a commercial cattle population in the US. Vet Parasitol. 2009 (166:281-5).
[21] Loveridge B, McArthur M, McKenna PB, Mariadass B. Probable multigeneric resistance to macrocyclic lactone anthelmintics in cattle in New Zealand. N Z Vet J. 2003 (51:139-41).
[22] Mason PC, McKay CH. Field studies investigating anthelmintic resistance in young cattle on five farms in New Zealand. N Z Vet J. 2006 (54:318-22).
[23] Mejia ME, Fernandez Igartua BM, Schmidt EE, Cabaret J. Multispecies and multiple anthelmintic resistance on cattle nematodes in a farm in Argentina: the beginning of high resistance? Vet Res. 2003 (34:461-7).
[24] Soutello RG, Seno MC, Amarante AF. Anthelmintic resistance in cattle nematodes in northwestern Sao Paulo State, Brazil. Vet Parasitol. 2007 (148:360-4).
[25] Suarez VH, Cristel SL. Anthelmintic resistance in cattle nematode in the western Pampeana Region of Argentina. Vet Parasitol. 2007 (144:111-7).
[26] Vermunt JJ, West DM, Pomroy WE. Inefficacy of moxidectin and doramectin against ivermectin-resistant Cooperia spp. of cattle in New Zealand. N Z Vet J. 1996 (44:188-93).
[27] Waghorn TS, Leathwick DM, Rhodes AP, Jackson R, Pomroy WE, West DM, Moffat JR. Prevalence of anthelmintic resistance on 62 beef cattle farms in the North Island of New Zealand. N Z Vet J. 2006 (54:278-82).
[28] Leland SE, Jr., Drudge JH, Wyant ZN, Elam GW. Strain variation in the response of sheep nematodes to action of phenothiazine. III. Field observations. American journal of veterinary research. 1957 (18:851-60).
[29] Drudge JH, Lyons ET, Tolliver SC, Lowry SR, Fallon EH. Piperazine resistance in population-B equine strongyles: a study of selection in Thoroughbreds in Kentucky from 1966 through 1983. American journal of veterinary research. 1988 (49:986-94).
[30] Drudge JH, Szanto J, Wyant ZN, Elam G. Field Studies on Parasite Control in Sheep: Comparison of Thiabendazole, Ruelene, and Phenothiazine. American journal of veterinary research. 1964 (25:1512-8).
[31] Berger J. The resistance of a field strain of Haemonchus contortus to five benzimidazole anthelmintics in current use. Journal of the South African Veterinary Association. 1975 (46:369-72).

REFERENCES
110
[32] Drudge JH, Lyons ET, Tolliver SC, Swerczek TW. Use of oxibendazole for control of cambendazole-resistant small strongyles in a band of ponies: a six-year study. American journal of veterinary research. 1985 (46:2507-11).
[33] Cawthorne RJ, Whitehead JD. Isolation of benzimidazole resistant strains of Ostertagia circumcincta from British sheep. Vet Rec. 1983 (112:274-7).
[34] Boersema JH, Lewing-van der Wiel PJ. Benzimidazole resistance in a field strain of Haemonchus contortus in the Netherlands. Vet Rec. 1982 (110:203-4).
[35] Le Jambre LF, Prichard PK, Hennessy DR, Laby RH. Efficiency of oxfendazole administered as a single dose or in a controlled release capsule against benzimidazole-resistant Haemonchus contortus, Ostertagia circumcincta and Trichostrongylus colubriformis. Research in veterinary science. 1981 (31:289-94).
[36] Sangster NC, Whitlock HV, Russ IG, Gunawan M, Griffin DL, Kelly JD. Trichostrongylus colubriformis and Ostertagia circumcincta resistant to levamisole, morantel tartrate and thiabendazole: occurrence of field strains. Research in veterinary science. 1979 (27:106-10).
[37] Chapman MR, French DD, Monahan CM, Klei TR. Identification and characterization of a pyrantel pamoate resistant cyathostome population. Vet Parasitol. 1996 (66:205-12).
[38] Wooster MJ, Woodgate RG, Chick BF. Reduced efficacy of ivermectin, abamectin and moxidectin against field isolates of Haemonchus contortus. Aust Vet J. 2001 (79:840-2).
[39] van Wyk JA, Malan FS. Resistance of field strains of Haemonchus contortus to ivermectin, closantel, rafoxanide and the benzimidazoles in South Africa. Vet Rec. 1988 (123:226-8).
[40] Leathwick DM. A case of moxidectin failing to control ivermectin resistant Ostertagia species in goats. Vet Rec. 1995 (136:443-4).
[41] Borgsteede FH, Dercksen DD, Huijbers R. Doramectin and albendazole resistance in sheep in The Netherlands. Vet Parasitol. 2007 (144:180-3).
[42] Martin PJ, Anderson N, Jarrett RG. Detecting benzimidazole resistance with faecal egg count reduction tests and in vitro assays. Aust Vet J. 1989 (66:236-40).
[43] Coles GC, Bauer C, Borgsteede FH, Geerts S, Klei TR, Taylor MA, Waller PJ. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Vet Parasitol. 1992 (44:35-44).
[44] Coles GC, Jackson F, Pomroy WE, Prichard RK, von Samson-Himmelstjerna G, Silvestre A, Taylor MA, Vercruysse J. The detection of anthelmintic resistance in nematodes of veterinary importance. Vet Parasitol. 2006 (136:167-85).
[45] Geary TG, Hosking BC, Skuce P, von Samson-Himmelstjerna G, Maeder S, Holdsworth P, Pomroy W, Vercruysse J. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) Guideline: Anthelmintic combination products targeting nematode infections of ruminants and horses. Vet Parasitol. 2012 (190:306-16).
[46] Wood IB, Amaral NK, Bairden K, Duncan JL, Kassai T, Malone JB Jr, Pankavich JA, Reinecke RK, Slocombe O, Taylor SM, Vercruysse J. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) second edition of guidelines for evaluating the efficacy of anthelmintics in ruminants (bovine, ovine, caprine). Vet Parasitol. 1995 (58:181-213).

REFERENCES
111
[47] Presidente PJA. Methods for the detection of resistance to anthelmintics. In: Anderson and Waller (Eds): Resistance in nematodes to anthelmintic drugs Division of Animal Health, CSIRO, Australia. 1985 (13-27).
[48] Taylor MA, Hunt KR, Goodyear KL. Anthelmintic resistance detection methods. Vet Parasitol. 2002 (103:183-94).
[49] Sutherland IA, Leathwick DM. Anthelmintic resistance in nematode parasites of cattle: a global issue? Trends Parasitol. 2011 (27:176-81).
[50] Levecke B, De Wilde N, Vandenhoute E, Vercruysse J. Field validity and feasibility of four techniques for the detection of Trichuris in simians: a model for monitoring drug efficacy in public health? PLoS Negl Trop Dis. 2009 (3:e366).
[51] Cringoli G. [Coprological diagnosis: what's new?]. Parassitologia. 2004 (46:137-9).
[52] Jackson F. New technique for obtaining nematode ova from sheep faeces. Laboratory practice. 1974 (23:65-6).
[53] Christie M, Jackson F. Specific identification of strongyle eggs in small samples of sheep faeces. Research in veterinary science. 1982 (32:113-7).
[54] Levecke B, Dobson RJ, Speybroeck N, Vercruysse J, Charlier J. Novel insights in the faecal egg count reduction test for monitoring drug efficacy against gastrointestinal nematodes of veterinary importance. Vet Parasitol. 2012 (188:391-6).
[55] Eysker M, Ploeger HW. Value of present diagnostic methods for gastrointestinal nematode infections in ruminants. Parasitology. 2000 (120 Suppl:S109-19).
[56] Kotze AC, Kopp SR. The potential impact of density dependent fecundity on the use of the faecal egg count reduction test for detecting drug resistance in human hookworms. PLoS Negl Trop Dis. 2008 (2:e297).
[57] Cabaret J, Berrag B. Faecal egg count reduction test for assessing anthelmintic efficacy: average versus individually based estimations. Vet Parasitol. 2004 (121:105-13).
[58] Dash KM, Hall E, Barger IA. The role of arithmetic and geometric mean worm egg counts in faecal egg count reduction tests and in monitoring strategic drenching programs in sheep. Aust Vet J. 1988 (65:66-8).
[59] Dobson RJ, Sangster NC, Besier RB, Woodgate RG. Geometric means provide a biased efficacy result when conducting a faecal egg count reduction test (FECRT). Vet Parasitol. 2009 (161:162-7).
[60] Lifschitz A, Sallovitz J, Imperiale F, Pis A, Jauregui Lorda J, Lanusse C. Pharmacokinetic evaluation of four ivermectin generic formulations in calves. Vet Parasitol. 2004 (119:247-57).
[61] Vercruysse J, Deprez P, Everaert D, Bassissi F, Alvinerie M. Breed differences in the pharmacokinetics of ivermectin administered subcutaneously to Holstein and Belgian Blue calves. Vet Parasitol. 2008 (152:136-40).
[62] Gonzalez Canga A, Sahagun Prieto AM, Jose Diez Liebana M, Martinez NF, Vega MS, Vieitez JJ. The pharmacokinetics and metabolism of ivermectin in domestic animal species. Vet J. 2009 (179:25-37).
[63] Sangster NC, Gill J. Pharmacology of anthelmintic resistance. Parasitol Today. 1999 (15:141-6).

REFERENCES
112
[64] Demeler J, Kuttler U, von Samson-Himmelstjerna G. Adaptation and evaluation of three different in vitro tests for the detection of resistance to anthelmintics in gastro intestinal nematodes of cattle. Vet Parasitol. 2010 (170:61-70).
[65] Folz SD, Pax RA, Thomas EM, Bennett JL, Lee BL, Conder GA. Detecting in vitro anthelmintic effects with a micromotility meter. Vet Parasitol. 1987 (24:241-50).
[66] Gill JH, Redwin JM, van Wyk JA, Lacey E. Avermectin inhibition of larval development in Haemonchus contortus--effects of ivermectin resistance. Int J Parasitol. 1995 (25:463-70).
[67] Tandon R, Kaplan RM. Evaluation of a larval development assay (DrenchRite) for the detection of anthelmintic resistance in cyathostomin nematodes of horses. Vet Parasitol. 2004 (121:125-42).
[68] Alvarez-Sanchez MA, Perez Garcia J, Bartley D, Jackson F, Rojo-Vazquez FA. The larval feeding inhibition assay for the diagnosis of nematode anthelmintic resistance. Exp Parasitol. 2005 (110:56-61).
[69] Coles GC. Anthelmintic resistance--looking to the future: a UK perspective. Research in veterinary science. 2005 (78:99-108).
[70] Gruber JD, Colligan PB, Wolford JK. Estimation of single nucleotide polymorphism allele frequency in DNA pools by using Pyrosequencing. Human genetics. 2002 (110:395-401).
[71] Ronaghi M. Pyrosequencing sheds light on DNA sequencing. Genome research. 2001 (11:3-11).
[72] Kwa MS, Veenstra JG, Roos MH. Benzimidazole resistance in Haemonchus contortus is correlated with a conserved mutation at amino acid 200 in beta-tubulin isotype 1. Mol Biochem Parasitol. 1994 (63:299-303).
[73] von Samson-Himmelstjerna G, Walsh TK, Donnan AA, Carriere S, Jackson F, Skuce PJ, Rohn K, Wolstenholme AJ. Molecular detection of benzimidazole resistance in Haemonchus contortus using real-time PCR and pyrosequencing. Parasitology. 2009 (136:349-58).
[74] Agneessens J, Dorny P, Hollanders W, Claerebout E, Vercruysse J. Epidemiological observations on gastrointestinal nematode infections in grazing cow-calf pairs in Belgium. Vet Parasitol. 1997 (69:65-75).
[75] Charlier J, Demeler J, Hoglund J, von Samson-Himmelstjerna G, Dorny P, Vercruysse J. Ostertagia ostertagi in first-season grazing cattle in Belgium, Germany and Sweden: general levels of infection and related management practices. Vet Parasitol. 2010 (171:91-8).
[76] Vermunt JJ, West DM, Pomroy WE. Multiple resistance to ivermectin and oxfendazole in Cooperia species of cattle in New Zealand. Vet Rec. 1995 (137:43-5).
[77] Vercruysse J, Claerebout E. Immunity development against Ostertagia ostertagi and other gastrointestinal nematodes in cattle. Vet Parasitol. 1997 (72:309-16; discussion 16-26).
[78] Sangster NC, Dobson RJ. Anthelmintic resistance. In: The biology of nematodes Eds D Lee, Taylor and Francis. 2002 (531-67).
[79] Leathwick DM, Pomroy WE, Heath AC. Anthelmintic resistance in New Zealand. N Z Vet J. 2001 (49:227-35).
[80] Leathwick DM, Hosking BC, Bisset SA, McKay CH. Managing anthelmintic resistance: is it feasible in New Zealand to delay the emergence of resistance to a new anthelmintic class? N Z Vet J. 2009 (57:181-92).

REFERENCES
113
[81] Vercruysse J, Rew RS. Macrocyclic lactones in antiparasitic therapy. CAB International, Wallingford, UK. 2002.
[82] Wolstenholme AJ, Fairweather I, Prichard R, von Samson-Himmelstjerna G, Sangster NC. Drug resistance in veterinary helminths. Trends Parasitol. 2004 (20:469-76).
[83] Skuce P, Stenhouse L, Jackson F, Hypsa V, Gilleard J. Benzimidazole resistance allele haplotype diversity in United Kingdom isolates of Teladorsagia circumcincta supports a hypothesis of multiple origins of resistance by recurrent mutation. Int J Parasitol. 2010 (40:1247-55).
[84] Coles GC. Resistance in cattle worms. Vet Rec. 2004 (155:312).
[85] Le Jambre LF, Gill JH, Lenane IJ, Baker P. Inheritance of avermectin resistance in Haemonchus contortus. Int J Parasitol. 2000 (30:105-11).
[86] Sangster NC, Redwin JM, Bjorn H. Inheritance of levamisole and benzimidazole resistance in an isolate of Haemonchus contortus. Int J Parasitol. 1998 (28:503-10).
[87] van Wyk JA. Refugia--overlooked as perhaps the most potent factor concerning the development of anthelmintic resistance. The Onderstepoort journal of veterinary research. 2001 (68:55-67).
[88] Kaminsky R. Drug resistance in nematodes: a paper tiger or a real problem? Current opinion in infectious diseases. 2003 (16:559-64).
[89] Pomroy WE. Anthelmintic resistance in New Zealand: a perspective on recent findings and options for the future. N Z Vet J. 2006 (54:265-70).
[90] Van Zeveren AM, Casaert S, Alvinerie M, Geldhof P, Claerebout E, Vercruysse J. Experimental selection for ivermectin resistance in Ostertagia ostertagi in cattle. Vet Parasitol. 2007 (150:104-10).
[91] Laffont CM, Bousquet-Melou A, Bralet D, Alvinerie M, Fink-Gremmels J, Toutain PL. A pharmacokinetic model to document the actual disposition of topical ivermectin in cattle. Vet Res. 2003 (34:445-60).
[92] Bisset SA, Brunsdon RV, Forbes S. Efficacy of a topical formulation of ivermectin against naturally acquired gastro-intestinal nematodes in weaner cattle. N Z Vet J. 1990 (38:4-6).
[93] Shoop WL, Mrozik H, Fisher MH. Structure and activity of avermectins and milbemycins in animal health. Vet Parasitol. 1995 (59:139-56).
[94] Lifschitz A, Virkel G, Sallovitz J, Sutra JF, Galtier P, Alvinerie M, Lanusse C. Comparative distribution of ivermectin and doramectin to parasite location tissues in cattle. Vet Parasitol. 2000 (87:327-38).
[95] Hennessy DR. Modifying the formulation or delivery mechanism to increase the activity of anthelmintic compounds. Vet Parasitol. Vet Parasitol. 1997 (72:367-82; discussion 82-90).
[96] Imperiale F, Sallovitz J, Farias C, Lifschitz A, Lanusse C. Licking induced changes to the pattern of moxidectin milk elimination after topical treatment in dairy cows. J Vet Pharmacol Ther. 2009 (32:534-40).
[97] Sallovitz JM, Lifschitz A, Imperiale F, Virkel G, Larghi J, Lanusse C. Doramectin concentration profiles in the gastrointestinal tract of topically-treated calves: Influence of animal licking restriction. Vet Parasitol. 2005 (133:61-70).

REFERENCES
114
[98] Alka, Gopal RM, Sandhu KS, Sidhu PK. Efficacy of abamectin against ivermectin-resistant strain of Trichostrongylus colubriformis in sheep. Vet Parasitol. 2004 (121:277-83).
[99] Gopal RM, West DM, Pomroy WE. The difference in efficacy of ivermectin oral, moxidectin oral and moxidectin injectable formulations against an ivermectin-resistant strain of Trichostrongylus colubriformis in sheep. N Z Vet J. 2001 (49:133-7).
[100] Lloberas M, Alvarez L, Entrocasso C, Virkel G, Lanusse C, Lifschitz A. Measurement of ivermectin concentrations in target worms and host gastrointestinal tissues: influence of the route of administration on the activity against resistant Haemonchus contortus in lambs. Exp Parasitol. 2012 (131:304-9).
[101] Alvarez LI, Mottier ML, Lanusse CE. Drug transfer into target helminth parasites. Trends Parasitol. 2007 (23:97-104).
[102] Kotze AC, Hines BM, Ruffell AP. A reappraisal of the relative sensitivity of nematode pharyngeal and somatic musculature to macrocyclic lactone drugs. International journal for parasitology: Drugs and Drugs Resistance. 2012 (2:29-35).
[103] Geary TG, Sims SM, Thomas EM, Vanover L, Davis JP, Winterrowd CA, Klein RD, Ho NF, Thompson DP. Haemonchus contortus: ivermectin-induced paralysis of the pharynx. Exp Parasitol. 1993 (77:88-96).
[104] Klager S, Whitworth JA, Post RJ, Chavasse DC, Downham MD. How long do the effects of ivermectin on adult Onchocerca volvulus persist? Trop Med Parasitol. 1993 (44:305-10).
[105] Le Jambre LF, Gill JH, Lenane IJ, Lacey E. Characterisation of an avermectin resistant strain of Australian Haemonchus contortus. Int J Parasitol. 1995 (25:691-8).
[106] Yates DM, Portillo V, Wolstenholme AJ. The avermectin receptors of Haemonchus contortus and Caenorhabditis elegans. Int J Parasitol. 2003 (33:1183-93).
[107] Prichard R. Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol. 2001 (17:445-53).
[108] Arena JP, Liu KK, Paress PS, Cully DF. Avermectin-sensitive chloride currents induced by Caenorhabditis elegans RNA in Xenopus oocytes. Molecular pharmacology. 1991 (40:368-74).
[109] Arena JP, Liu KK, Paress PS, Schaeffer JM, Cully DF. Expression of a glutamate-activated chloride current in Xenopus oocytes injected with Caenorhabditis elegans RNA: evidence for modulation by avermectin. Brain research Molecular brain research. 1992 (15:339-48).
[110] Wolstenholme AJ, Rogers AT. Glutamate-gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitology. 2005 (131 Suppl:S85-95).
[111] Hibbs RE, Gouaux E. Principles of activation and permeation in an anion-selective Cys-loop receptor. Nature. 2011 (474:54-60).
[112] Tsetlin V, Kuzmin D, Kasheverov I. Assembly of nicotinic and other Cys-loop receptors. Journal of neurochemistry. 2011 (116:734-41).
[113] Cully DF, Vassilatis DK, Liu KK, Paress PS, Van der Ploeg LH, Schaeffer JM, Arena JP. Cloning of an avermectin-sensitive glutamate-gated chloride channel from Caenorhabditis elegans. Nature. 1994 (371:707-11).
[114] Dent JA, Davis MW, Avery L. avr-15 encodes a chloride channel subunit that mediates inhibitory glutamatergic neurotransmission and ivermectin sensitivity in Caenorhabditis elegans. Embo J. 1997 (16:5867-79).

REFERENCES
115
[115] Dent JA, Smith MM, Vassilatis DK, Avery L. The genetics of ivermectin resistance in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2000 (97:2674-9).
[116] Horoszok L, Raymond V, Sattelle DB, Wolstenholme AJ. GLC-3: a novel fipronil and BIDN-sensitive, but picrotoxinin-insensitive, L-glutamate-gated chloride channel subunit from Caenorhabditis elegans. British journal of pharmacology. 2001 (132:1247-54).
[117] Vassilatis DK, Arena JP, Plasterk RH, Wilkinson HA, Schaeffer JM, Cully DF, Van der Ploeg LH. Genetic and biochemical evidence for a novel avermectin-sensitive chloride channel in Caenorhabditis elegans. Isolation and characterization. J Biol Chem. 1997 (272:33167-74).
[118] Vassilatis DK, Elliston KO, Paress PS, Hamelin M, Arena JP, Schaeffer JM, Van der Ploeg LH, Cully DF. Evolutionary relationship of the ligand-gated ion channels and the avermectin-sensitive, glutamate-gated chloride channels. Journal of molecular evolution. 1997 (44:501-8).
[119] Ardelli BF, Stitt LE, Tompkins JB, Prichard RK. A comparison of the effects of ivermectin and moxidectin on the nematode Caenorhabditis elegans. Vet Parasitol. 2009 (165:96-108).
[120] Laughton DL, Lunt GG, Wolstenholme AJ. Reporter gene constructs suggest that the Caenorhabditis elegans avermectin receptor beta-subunit is expressed solely in the pharynx. The Journal of experimental biology. 1997 (200:1509-14).
[121] Jagannathan S, Laughton DL, Critten CL, Skinner TM, Horoszok L, Wolstenholme AJ. Ligand-gated chloride channel subunits encoded by the Haemonchus contortus and Ascaris suum orthologues of the Caenorhabditis elegans gbr-2 (avr-14) gene. Mol Biochem Parasitol. 1999 (103:129-40).
[122] Portillo V, Jagannathan S, Wolstenholme AJ. Distribution of glutamate-gated chloride channel subunits in the parasitic nematode Haemonchus contortus. The Journal of comparative neurology. 2003 (462:213-22).
[123] Glendinning SK, Buckingham SD, Sattelle DB, Wonnacott S, Wolstenholme AJ. Glutamate-gated chloride channels of Haemonchus contortus restore drug sensitivity to ivermectin resistant Caenorhabditis elegans. PLoS One. 2011 (6:e22390).
[124] McCavera S, Walsh TK, Wolstenholme AJ. Nematode ligand-gated chloride channels: an appraisal of their involvement in macrocyclic lactone resistance and prospects for developing molecular markers. Parasitology. 2007 (134:1111-21).
[125] Raymond V, Sattelle DB. Novel animal-health drug targets from ligand-gated chloride channels. Nature reviews Drug discovery. 2002 (1:427-36).
[126] James CE, Hudson AL, Davey MW. Drug resistance mechanisms in helminths: is it survival of the fittest? Trends Parasitol. 2009 (25:328-35).
[127] Lespine A, Ménez C, Bourguinat C, Prichard R. P-glycoproteins and other multidrug resistance transporters in the pharmacology of anthelmintics: Prospects for reversing-dependent anthelmintic resistance. International journal for parasitology: Drugs and Drugs Resistance. 2012 (2:58-75).
[128] Prichard R, Tait A. The role of molecular biology in veterinary parasitology. Vet Parasitol. 2001 (98:169-94).
[129] Ghosh R, Andersen EC, Shapiro JA, Gerke JP, Kruglyak L. Natural variation in a chloride channel subunit confers avermectin resistance in Caenorhabditis elegans. Science. 2012 (335:574-8).

REFERENCES
116
[130] Njue AI, Prichard RK. Genetic variability of glutamate-gated chloride channel genes in ivermectin-susceptible and -resistant strains of Cooperia oncophora. Parasitology. 2004 (129:741-51).
[131] Njue AI, Hayashi J, Kinne L, Feng XP, Prichard RK. Mutations in the extracellular domains of glutamate-gated chloride channel alpha3 and beta subunits from ivermectin-resistant Cooperia oncophora affect agonist sensitivity. Journal of neurochemistry. 2004 (89:1137-47).
[132] El-Abdellati A, De Graef J, Van Zeveren A, Donnan A, Skuce P, Walsh T, Wolstenholme AJ, Tait A, Vercruysse J, Claerebout E, Geldhof P. Altered avr-14B gene transcription patterns in ivermectin-resistant isolates of the cattle parasites, Cooperia oncophora and Ostertagia ostertagi. Int J Parasitol. 2011 (41:951-7).
[133] Martinez-Valladares M, Geldhof P, Jonsson N, Rojo-Vasquez FA, Skuce P. Teladorsagia circumcincta: Molecular characterisation of the avr-14B subunit and its relatively minor role in ivermectin resistance. International journal for parasitology: Drugs and Drugs Resistance. 2012 (2:154-61).
[134] McCavera S, Rogers AT, Yates DM, Woods DJ, Wolstenholme AJ. An ivermectin-sensitive glutamate-gated chloride channel from the parasitic nematode Haemonchus contortus. Molecular pharmacology. 2009 (75:1347-55).
[135] Blackhall WJ, Pouliot JF, Prichard RK, Beech RN. Haemonchus contortus: selection at a glutamate-gated chloride channel gene in ivermectin- and moxidectin-selected strains. Exp Parasitol. 1998 (90:42-8).
[136] Forrester SG, Beech RN, Prichard RK. Agonist enhacement of macrocyclic lactone activity at a glutamate-gated chloride channel subunit from Haemonchus contortus. Biochem Pharmacol. 2004 (67:1019-24).
[137] Forrester SG, Hamdan FF, Prichard RK, Beech RN. Cloning, sequencing, and developmental expression levels of a novel glutamate-gated chloride channel homologue in the parasitic nematode Haemonchus contortus. Biochem Biophys Res Commun. 1999 (254:529-34).
[138] Forrester SG, Prichard RK, Beech RN. A glutamate-gated chloride channel subunit from Haemonchus contortus: expression in a mammalian cell line, ligand binding, and modulation of anthelmintic binding by glutamate. Biochem Pharmacol. 2002 (63:1061-8).
[139] Forrester SG, Prichard RK, Dent JA, Beech RN. Haemonchus contortus: HcGluCla expressed in Xenopus oocytes forms a glutamate-gated ion channel that is activated by ibotenate and the antiparasitic drug ivermectin. Mol Biochem Parasitol. 2003 (129:115-21).
[140] Williamson SM, Storey B, Howell S, Harper KM, Kaplan RM, Wolstenholme AJ. Candidate anthelmintic resistance-associated gene expression and sequence polymorphisms in a triple-resistant field isolate of Haemonchus contortus. Mol Biochem Parasitol. 2011 (180:99-105).
[141] Cheeseman CL, Delany NS, Woods DJ, Wolstenholme AJ. High-affinity ivermectin binding to recombinant subunits of the Haemonchus contortus glutamate-gated chloride channel. Mol Biochem Parasitol. 2001 (114:161-8).
[142] Hollenstein K, Frei DC, Locher KP. Structure of an ABC transporter in complex with its binding protein. Nature. 2007 (446:213-6).
[143] Jones PM, George AM. Multidrug resistance in parasites: ABC transporters, P-glycoproteins and molecular modelling. Int J Parasitol. 2005 (35:555-66).

REFERENCES
117
[144] Lespine A, Alvinerie M, Vercruysse J, Prichard RK, Geldhof P. ABC transporter modulation: a strategy to enhance the activity of macrocyclic lactone anthelmintics. Trends Parasitol. 2008 (24:293-8).
[145] Lespine A, Martin S, Dupuy J, Roulet A, Pineau T, Orlowski S, Alvinerie M. Interaction of macrocyclic lactones with P-glycoprotein: structure-affinity relationship. Eur J Pharm Sci. 2007 (30:84-94).
[146] Mealey KL, Bentjen SA, Gay JM, Cantor GH. Ivermectin sensitivity in collies is associated with a deletion mutation of the mdr1 gene. Pharmacogenetics. 2001 (11:727-33).
[147] Roulet A, Puel O, Gesta S, Lepage JF, Drag M, Soll M, Alvinerie M, Pineau T. MDR1-deficient genotype in Collie dogs hypersensitive to the P-glycoprotein substrate ivermectin. Eur J Pharmacol. 2003 (460:85-91).
[148] Schinkel AH, Smit JJ, van Tellingen O, Beijnen JH, Wagenaar E, van Deemter L, Mol CA, van der Valk MA, Robanus-Maandag EC, te Riele HP, Berns AJ, Borst P. Disruption of the mouse mdr1a P-glycoprotein gene leads to a deficiency in the blood-brain barrier and to increased sensitivity to drugs. Cell. 1994 (77:491-502).
[149] Dicker AJ, Nisbet AJ, Skuce PJ. Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int J Parasitol. 2011 (41:935-42).
[150] Zhao Z, Sheps JA, Ling V, Fang LL, Baillie DL. Expression analysis of ABC transporters reveals differential functions of tandemly duplicated genes in Caenorhabditis elegans. J Mol Biol. 2004 (344:409-17).
[151] Williamson SM, Wolstenholme AJ. P-glycoproteins of Haemonchus contortus: development of real-time PCR assays for gene expression studies. J Helminthol. 2011(1-7).
[152] Blackhall WJ, Liu HY, Xu M, Prichard RK, Beech RN. Selection at a P-glycoprotein gene in ivermectin- and moxidectin-selected strains of Haemonchus contortus. Mol Biochem Parasitol. 1998 (95:193-201).
[153] Xu M, Molento M, Blackhall W, Ribeiro P, Beech R, Prichard R. Ivermectin resistance in nematodes may be caused by alteration of P-glycoprotein homolog. Mol Biochem Parasitol. 1998 (91:327-35).
[154] Prichard RK, Roulet A. ABC transporters and beta-tubulin in macrocyclic lactone resistance: prospects for marker development. Parasitology. 2007 (134:1123-32).
[155] Ardelli BF, Prichard R. Effects of ivermectin and moxidectin on the transcription of genes coding for multidrug resistance associated proteins and behaviour in Caenorhabditis elegans. The Journal of Nematology. 2008 (40:290-8).
[156] James CE, Davey MW. Increased expression of ABC transport proteins is associated with ivermectin resistance in the model nematode Caenorhabditis elegans. Int J Parasitol. 2009 (39:213-20).
[157] Yan R, Urdaneta-Marquez L, Keller K, James CE, Davey MW, Prichard RK. The role of several ABC transporter genes in ivermectin resistance in Caenorhabditis elegans. Vet Parasitol. 2012 (190:519-29).
[158] Alvinerie M, Dupuy J, Kiki-Mvouaka S, Sutra JF, Lespine A. Ketoconazole increases the plasma levels of ivermectin in sheep. Vet Parasitol. 2008 (157:117-22).

REFERENCES
118
[159] Lifschitz A, Entrocasso C, Alvarez L, Lloberas M, Ballent M, Manazza G, Virkel G, Borda B, Lanusse C. Interference with P-glycoprotein improves ivermectin activity against adult resistant nematodes in sheep. Vet Parasitol. 2010 (172:291-8).
[160] Lifschitz A, Virkel G, Sallovitz J, Imperiale F, Pis A, Lanusse C. Loperamide-induced enhancement of moxidectin availability in cattle. J Vet Pharmacol Ther. 2002 (25:111-20).
[161] Lifschitz AL, Virkel GL, Sallovitz JM, Pis A, Imperiale FA, Lanusse CE. Loperamide modifies the tissue disposition kinetics of ivermectin in rats. J Pharm Pharmacol. 2004 (56:61-7).
[162] Molento MB, Lifschitz A, Sallovitz J, Lanusse C, Prichard R. Influence of verapamil on the pharmacokinetics of the antiparasitic drugs ivermectin and moxidectin in sheep. Parasitol Res. 2004 (92:121-7).
[163] Molento MB, Prichard RK. Effects of the multidrug-resistance-reversing agents verapamil and CL 347,099 on the efficacy of ivermectin or moxidectin against unselected and drug-selected strains of Haemonchus contortus in jirds (Meriones unguiculatus). Parasitol Res. 1999 (85:1007-11).
[164] Bartley DJ, Morrison AA, Dupuy J, Bartley Y, Sutra JF, Menez C, Alvinerie M, Jackson F, Devin L, Lespine A. Influence of Pluronic 85 and ketoconazole on disposition and efficacy of ivermectin in sheep infected with a multiple resistant Haemonchus contortus isolate. Vet Parasitol. 2012 (187:464-72).
[165] Bartley DJ, McAllister H, Bartley Y, Dupuy J, Menez C, Alvinerie M, et al. P-glycoprotein interfering agents potentiate ivermectin susceptibility in ivermectin sensitive and resistant isolates of Teladorsagia circumcincta and Haemonchus contortus. Parasitology. 2009 (136:1081-8).
[166] Beech RN, Skuce P, Bartley DJ, Martin RJ, Prichard RK, Gilleard JS. Anthelmintic resistance: markers for resistance, or susceptibility? Parasitology. 2011 (138:160-74).
[167] Blackhall WJ, Prichard RK, Beech RN. Selection at a gamma-aminobutyric acid receptor gene in Haemonchus contortus resistant to avermectins/milbemycins. Mol Biochem Parasitol. 2003 (131:137-45).
[168] Feng XP, Hayashi J, Beech RN, Prichard RK. Study of the nematode putative GABA type-A receptor subunits: evidence for modulation by ivermectin. Journal of neurochemistry. 2002 (83:870-8).
[169] Rao VT, Siddiqui SZ, Prichard RK, Forrester SG. A dopamine-gated ion channel (HcGGR3*) from Haemonchus contortus is expressed in the cervical papillae and is associated with macrocyclic lactone resistance. Mol Biochem Parasitol. 2009 (166:54-61).
[170] Freeman AS, Nghiem C, Li J, Ashton FT, Guerrero J, Shoop WL, Schad GA. Amphidial structure of ivermectin-resistant and susceptible laboratory and field strains of Haemonchus contortus. Vet Parasitol. 2003 (110:217-26).
[171] Bourguinat C, Pion SD, Kamgno J, Gardon J, Gardon-Wendel N, Duke BO, Prichard RK, Boussinesq M. Genetic polymorphism of the beta-tubulin gene of Onchocerca volvulus in ivermectin naive patients from Cameroon, and its relationship with fertility of the worms. Parasitology. 2006 (132:255-62).
[172] Eng JK, Blackhall WJ, Osei-Atweneboana MY, Bourguinat C, Galazzo D, Beech RN, Unnasch TR, Awadzi K, Lubega GW, Prichard RK. Ivermectin selection on beta-tubulin: evidence in Onchocerca volvulus and Haemonchus contortus. Mol Biochem Parasitol. 2006 (150:229-35).

REFERENCES
119
[173] Mottier ML, Prichard R. Genetic analysis of a relationship between macrocyclic lactone and benzimidazole anthelmintic selection on Haemonchus contortus. Pharmacogenetics and Genomics. 2008 (18:129-40).
[174] Cvilink V, Lamka J, Skalova L. Xenobiotic metabolizing enzymes and metabolism of anthelminthics in helminths. Drug metabolism reviews. 2009 (41:8-26).
[175] Kawalek JC, Rew RS, Heavner J. Glutathione-S-transferase, a possible drug-metabolizing enzyme, in Haemonchus contortus: comparative activity of a cambendazole-resistant and a susceptible strain. Int J Parasitol. 1984 (14:173-5).
[176] Kerboeuf D, Aycardi J. Unexpected increased thiabendazole tolerance in Haemonchus contortus resistant to anthelmintics by modulation of glutathione activity. Parasitol Res. 1999 (85:713-8).
[177] Lindblom TH, Dodd AK. Xenobiotic detoxification in the nematode Caenorhabditis elegans. Journal of experimental zoology Part A, Comparative experimental biology. 2006 (305:720-30.)
[178] Sotirchos IM, Hudson AL, Ellis J, Davey MW. A unique thioredoxin of the parasitic nematode Haemonchus contortus with glutaredoxin activity. Free radical biology & medicine. 2009 (46:579-85).
[179] Smith G, Grenfell BT, Anderson RM. The regulation of Ostertagia ostertagi populations in calves: density-dependent control of fecundity. Parasitology. 1987 (95 ( Pt 2):373-88).
[180] van Wyk JA, Cabaret J, Michael LM. Morphological identification of nematode larvae of small ruminants and cattle simplified. Vet Parasitol. 2004 (119:277-306).
[181] Kochapakdee S, Pandey VS, Pralomkarn W, Choldumrongkul S, Ngampongsai W, Lawpetchara A. Anthelmintic resistance in goats in southern Thailand. Vet Rec. 1995 (137:124-5).
[182] Anderson RC. Nematode parasites of vertebrates. Their development and transmission. CABI Publishing Wallingford UK. 2002 (672 pp).
[183] Paiement JP, Leger C, Ribeiro P, Prichard RK. Haemonchus contortus: effects of glutamate, ivermectin, and moxidectin on inulin uptake activity in unselected and ivermectin-selected adults. Exp Parasitol. 1999 (92:193-8).
[184] Condi GK, Soutello RG, Amarante AF. Moxidectin-resistant nematodes in cattle in Brazil. Vet Parasitol. 2009 (161:213-7).
[185] Ranjan S, Trudeau C, Prichard RK, von Kutzleben R, Carrier D. Efficacy of moxidectin against naturally acquired nematode infections in cattle. Vet Parasitol. 1992 (41:227-31).
[186] Schnyder M, Torgerson PR, Schonmann M, Kohler L, Hertzberg H. Multiple anthelmintic resistance in Haemonchus contortus isolated from South African Boer goats in Switzerland. Vet Parasitol. 2005 (128:285-90).
[187] McKenna PB. The detection of anthelmintic resistance by the faecal egg count reduction test: an examination of some of the factors affecting performance and interpretation. N Z Vet J. 1990 (38:142-7).
[188] de Soutello RV, Coelho WM, de Oliveira FP, Fonzar JF, Luquetti BC, de Souza RF, Seno MC, Amarante AF. Evaluation of reduction in egg shedding of gastrointestinal nematodes in cattle following administration of anthelmintics. Revista brasileira de parasitologia veterinaria: Orgao Oficial do Colegio Brasileiro de Parasitologia Veterinaria. 2010 (19:183-5).

REFERENCES
120
[189] Fiel CA, Saumell CA, Steffan PE, Rodriguez EM. Resistance of Cooperia to ivermectin treatments in grazing cattle of the Humid Pampa, Argentina. Vet Parasitol. 2001 (97:211-7).
[190] Yazwinski TA, Tucker CA, Hornsby JA, Powell JG, Reynolds JL, Johnson ZB, Lindsey W, Silver TK. Effectiveness evaluation of several cattle anthelmintics via the fecal egg count reduction test. Parasitol Res. 2009 (105:71-6).
[191] Heizer E, Zarlenga DS, Gasser RB, De Graef J, Geldhof P, Mitreva M. Transcriptome analyses reveal protein and domain families that delineate stage-related development in the economically important parasitic nematodes, Ostertagia ostertagi and Cooperia oncophora. BMC Genomics. 2012 (Submitted).
[192] Li W, Godzik A. Cd-hit: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics. 2006 (22:1658-9).
[193] Wasmuth JD, Blaxter ML. prot4EST: translating expressed sequence tags from neglected genomes. BMC bioinformatics. 2004 (5:187).
[194] Quevillon E, Silventoinen V, Pillai S, Harte N, Mulder N, Apweiler R, Lopez, R. InterProScan: protein domains identifier. Nucleic acids research. 2005 (33:W116-20).
[195] Martin J, Abubucker S, Heizer E, Taylor CM, Mitreva M. Nematode.net update 2011: addition of data sets and tools featuring next-generation sequencing data. Nucleic acids research. 2012 (40:D720-8).
[196] Elsworth B, Wasmuth J, Blaxter M. NEMBASE4: the nematode transcriptome resource. Int J Parasitol. 2011 (41:881-94).
[197] Demeler J, Krücken J, AlGusbi S, Ramünke S, De Graef J, Kerboeuf D, Geldhof P, Pomroy W, von Samson-Himmelstjerna G. Contribution of P-glycoproteins to macrocyclic lactone resistance in the cattle parasitic nematode Cooperia oncophora. Mol Biochem Parasitol. 2012 (Submitted).
[198] Sheps JA, Ralph S, Zhao Z, Baillie DL, Ling V. The ABC transporter gene family of Caenorhabditis elegans has implications for the evolutionary dynamics of multidrug resistance in eukaryotes. Genome Biol. 2004 (5:R15).
[199] Beech RN, Wolstenholme AJ, Neveu C, Dent JA. Nematode parasite genes: what's in a name? Trends Parasitol. 2010 (26:334-40).
[200] Mes TH. Purifying selection and demographic expansion affect sequence diversity of the ligand-binding domain of a glutamate-gated chloride channel gene of Haemonchus placei. Journal of molecular evolution. 2004 (58:466-78).
[201] Williamson SM, Walsh TK, Wolstenholme AJ. The cys-loop ligand-gated ion channel gene family of Brugia malayi and Trichinella spiralis: a comparison with Caenorhabditis elegans. Invertebrate neuroscience. 2007 (7:219-26.)
[202] Yates DM, Wolstenholme AJ. Dirofilaria immitis: identification of a novel ligand-gated ion channel-related polypeptide. Exp Parasitol. 2004 (108:182-5).
[203] Lifschitz A, Virkel G, Imperiale F, Sutra JF, Galtier P, Lanusse C, et al. Moxidectin in cattle: correlation between plasma and target tissues disposition. J Vet Pharmacol Ther. 1999 (22:266-73).
[204] Van Zeveren AM, Visser A, Hoorens PR, Vercruysse J, Claerebout E, Geldhof P. Evaluation of reference genes for quantitative real-time PCR in Ostertagia ostertagi by the coefficient of variation and geNorm approach. Mol Biochem Parasitol. 2007 (153:224-7).

REFERENCES
121
[205] Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002 (3:RESEARCH0034).
[206] Egea PF, Stroud RM, Walter P. Targeting proteins to membranes: structure of the signal recognition particle. Curr Opin Struct Biol. 2005 (15:213-20).
[207] Janda CY, Li J, Oubridge C, Hernandez H, Robinson CV, Nagai K. Recognition of a signal peptide by the signal recognition particle. Nature. 2010 (465:507-10).
[208] Stroud RM, Walter P. Signal sequence recognition and protein targeting. Curr Opin Struct Biol. 1999 (9:754-9).
[209] Wollmuth LP, Sobolevsky AI. Structure and gating of the glutamate receptor ion channel. Trends in neurosciences. 2004 (27:321-8).
[210] Breitinger HG, Villmann C, Melzer N, Rennert J, Breitinger U, Schwarzinger S, Becker CM. Novel regulatory site within the TM3-4 loop of human recombinant alpha3 glycine receptors determines channel gating and domain structure. J Biol Chem. 2009 (284:28624-33).
[211] Lo WY, Botzolakis EJ, Tang X, Macdonald RL. A conserved Cys-loop receptor aspartate residue in the M3-M4 cytoplasmic loop is required for GABAA receptor assembly. J Biol Chem. 2008 (283:29740-52).
[212] Melzer N, Villmann C, Becker K, Harvey K, Harvey RJ, Vogel N, Kluck CJ, Kneussei M, Becker CM. Multifunctional basic motif in the glycine receptor intracellular domain induces subunit-specific sorting. J Biol Chem. 2010 (285:3730-9).
[213] Lynagh T, Lynch JW. Molecular mechanisms of Cys-loop ion channel receptor modulation by ivermectin. Frontiers in molecular neuroscience. 2012 (5:60).
[214] Dermauw W, Ilias A, Riga M, Tsagkarakou A, Grbic M, Tirry L, Van Leeuwen T, Vontas J. The cys-loop ligand-gated ion channel gene family of Tetranychus urticae: implications for acaricide toxicology and a novel mutation associated with abamectin resistance. Insect biochemistry and molecular biology. 2012 (42:455-65).
[215] Kane NS, Hirschberg B, Qian S, Hunt D, Thomas B, Brochu R, Ludmerer SW, Zheng Y, Smith M, Arena JP, Cohen CJ, Schmatz D, Warmke J, Cully DF. Drug-resistant Drosophila indicate glutamate-gated chloride channels are targets for the antiparasitics nodulisporic acid and ivermectin. Proc Natl Acad Sci U S A. 2000 (97:13949-54).
[216] Kwon DH, Yoon KS, Clark JM, Lee SH. A point mutation in a glutamate-gated chloride channel confers abamectin resistance in the two-spotted spider mite, Tetranychus urticae Koch. Insect molecular biology. 2010 (19:583-91).
[217] Lynagh T, Lynch JW. A glycine residue essential for high ivermectin sensitivity in Cys-loop ion channel receptors. Int J Parasitol. 2010 (40:1477-81).
[218] Yamaguchi M, Sawa Y, Matsuda K, Ozoe F, Ozoe Y. Amino acid residues of both the extracellular and transmembrane domains influence binding of the antiparasitic agent milbemycin to Haemonchus contortus AVR-14B glutamate-gated chloride channels. Biochem Biophys Res Commun. 2012 (419:562-6).
[219] Lincke CR, Broeks A, The I, Plasterk RH, Borst P. The expression of two P-glycoprotein (pgp) genes in transgenic Caenorhabditis elegans is confined to intestinal cells. Embo J. 1993 (12:1615-20).

REFERENCES
122
[220] Broeks A, Janssen HW, Calafat J, Plasterk RH. A P-glycoprotein protects Caenorhabditis elegans against natural toxins. Embo J. 1995 (14:1858-66).
[221] Lincke CR, The I, van Groenigen M, Borst P. The P-glycoprotein gene family of Caenorhabditis elegans. Cloning and characterization of genomic and complementary DNA sequences. J Mol Biol. 1992 (228:701-11).
[222] Nunes F, Wolf M, Hartmann J, Paul RJ. The ABC transporter PGP-2 from Caenorhabditis elegans is expressed in the sensory neuron pair AWA and contributes to lysosome formation and lipid storage within the intestine. Biochem Biophys Res Commun. 2005 (338:862-71).
[223] Broeks A, Gerrard B, Allikmets R, Dean M, Plasterk RH. Homologues of the human multidrug resistance genes MRP and MDR contribute to heavy metal resistance in the soil nematode Caenorhabditis elegans. Embo J. 1996 (15:6132-43).
[224] Schroeder LK, Kremer S, Kramer MJ, Currie E, Kwan E, Watts JL, Lawrenson AL, Hermann GJ. Function of the Caenorhabditis elegans ABC transporter PGP-2 in the biogenesis of a lysosome-related fat storage organelle. Mol Biol Cell. 2007 (18:995-1008).
[225] Ardelli BF, Stitt LE, Tompkins JB. Inventory and analysis of ATP-binding cassette (ABC) systems in Brugia malayi. Parasitology. 2010 (137:1195-212).
[226] Ardelli BF, Guerriero SB, Prichard RK. Genomic organization and effects of ivermectin selection on Onchocerca volvulus P-glycoprotein. Mol Biochem Parasitol. 2005 (143:58-66).
[227] Ardelli BF, Guerriero SB, Prichard RK. Characterization of a half-size ATP-binding cassette transporter gene which may be a useful marker for ivermectin selection in Onchocerca volvulus. Mol Biochem Parasitol. 2006 (145:94-100).
[228] Bourguinat C, Ardelli BF, Pion SD, Kamgno J, Gardon J, Duke BO, Boussinesq M, Prichard RK. P-glycoprotein-like protein, a possible genetic marker for ivermectin resistance selection in Onchocerca volvulus. Mol Biochem Parasitol. 2008 (158:101-11).
[229] Huang YJ, Prichard RK. Identification and stage-specific expression of two putative P-glycoprotein coding genes in Onchocerca volvulus. Mol Biochem Parasitol. 1999 (102:273-81).
[230] Ardelli BF, Prichard RK. Reduced genetic variation of an Onchocerca volvulus ABC transporter gene following treatment with ivermectin. Trans R Soc Trop Med Hyg. 2007 (101:1223-32).
[231] Blackhall WJ, Prichard RK, Beech RN. P-glycoprotein selection in strains of Haemonchus contortus resistant to benzimidazoles. Vet Parasitol. 2008 (152:101-7).
[232] Alvinerie M, Dupuy J, Eeckhoutte C, Sutra JF. Enhanced absorption of pour-on ivermectin formulation in rats by co-administration of the multidrug-resistant-reversing agent verapamil. Parasitol Res. 1999 (85:920-2).
[233] Lifschitz A, Virkel G, Pis A, Imperiale F, Sanchez S, Alvarez L, Kujanek R, Lanusse C. Ivermectin disposition kinetics after subcutaneous and intramuscular administration of an oil-based formulation to cattle. Vet Parasitol. 1999 (86:203-15).
[234] Maglich JM, Parks DJ, Moore LB, Collins JL, Goodwin B, Billin AN, Stoltz CA, Kliewer SA, Lambert MH, Willson TM, Moore JT. Identification of a novel human constitutive androstane receptor (CAR) agonist and its use in the identification of CAR target genes. J Biol Chem. 2003 (278:17277-83).

REFERENCES
123
[235] Synold TW, Dussault I, Forman BM. The orphan nuclear receptor SXR coordinately regulates drug metabolism and efflux. Nature medicine. 2001 (7:584-90).
[236] Wei P, Zhang J, Egan-Hafley M, Liang S, Moore DD. The nuclear receptor CAR mediates specific xenobiotic induction of drug metabolism. Nature. 2000 (407:920-3).
[237] Ménez C, Mselli-Lakhal L, Foucaud-Vignault M, Balaguer P, Alvinerie M, Lespine A. Ivermectin induces P-glycoprotein expression and function through mRNA stabilization in murine hepatocyte cell line. Biochem Pharmacol. 2012 (83:269-78).
[238] Coles GC. Cattle nematodes resistant to anthelmintics: why so few cases? Vet Res. 2002 (33:481-9).
[239] Torgerson PR, Schnyder M, Hertzberg H. Detection of anthelmintic resistance: a comparison of mathematical techniques. Vet Parasitol. 2005 (128:291-8).
[240] Bartley DJ, McArthur CL, Devin LM, Sutra JF, Morrison AA, Lespine A, Matthews JB. Characterisation of macrocyclic lactone resistance in two field-derived isolates of Cooperia oncophora. Vet Parasitol. 2012 (190:454-60).
[241] Scholl PJ, Guillot FS, Wang GT. Moxidectin: systemic activity against common cattle grubs (Hypoderma lineatum) (Diptera: Oestridae) and trichostrongyle nematodes in cattle. Vet Parasitol. 1992 (41:203-9).
[242] Prichard R, Ménez C, Lespine A. Moxidectin and the avermectins: Consanguinity but not identity. International journal for parasitology: Drugs and Drugs Resistance. 2012 (2:134-53).
[243] Lloberas M, Alvarez L, Entrocasso C, Virkel G, Ballent M, Mate L, et al. Comparative tissue pharmacokinetics and efficacy of moxidectin, abamectin and ivermectin in lambs infected with resistant nematodes: Impact of drug treatments on parasite P-glycoprotein expression. International journal for parasitology: Drugs and Drugs Resistance. 2013 (3:20-7).
[244] Gilleard JS, Beech RN. Population genetics of anthelmintic resistance in parasitic nematodes. Parasitology. 2007 (134:1133-47)
[245] Charlier J, Levecke B, Devleesschauwer B, Vercruysse J, Hogeveen H. The economic effects of whole-herd versus selective anthelmintic treatment strategies in dairy cows. Journal of dairy science. 2012 (95:2977-87).
[246] Greer AW, Kenyon F, Bartley DJ, Jackson EB, Gordon Y, Donnan AA, McBean DW, Jackson F. Development and field evaluation of a decision support model for anthelmintic treatments as part of a targeted selective treatment (TST) regime in lambs. Vet Parasitol. 2009 (164:12-20).
[247] Stafford KA, Morgan ER, Coles GC. Weight-based targeted selective treatment of gastrointestinal nematodes in a commercial sheep flock. Vet Parasitol. 2009 (164:59-65).
[248] Dobson RJ, Besier RB, Barnes EH, Love SC, Vizard A, Bell K, Le Jambre LF. Principles for the use of macrocyclic lactones to minimise selection for resistance. Aust Vet J. 2001 (79:756-61).
[249] Assis RC, Luns FD, Araujo JV, Braga FR. Biological control of Trichostrongyles in beef cattle by the nematophagous fungus Duddingtonia flagrans in tropical southeastern Brazil. Exp Parasitol. 2012 (132:373-7).
[250] Waller PJ, Larsen M, Faedo M, Hennessy DR. The potential of nematophagous fungi to control the free-living stages of nematode parasites of sheep: in vitro and in vivo studies. Vet Parasitol. 1994 (51:289-99).

REFERENCES
124
[251] Stear MJ, Doligalska M, Donskow-Schmelter K. Alternatives to anthelmintics for the control of nematodes in livestock. Parasitology. 2007 (134:139-51).
[252] Li RW, Choudhary RK, Capuco AV, Urban JF, Jr. Exploring the host transcriptome for mechanisms underlying protective immunity and resistance to nematode infections in ruminants. Vet Parasitol. 2012 (190:1-11).
[253] Claerebout E, Knox DP, Vercruysse J. Current research and future prospects in the development of vaccines against gastrointestinal nematodes in cattle. Expert review of vaccines. 2003 (2:147-57).
[254] Vercruysse J, Schetters TP, Knox DP, Willadsen P, Claerebout E. Control of parasitic disease using vaccines: an answer to drug resistance? Rev Sci Tech. 2007 (26:105-15).
[255] Gilleard JS. Understanding anthelmintic resistance: the need for genomics and genetics. Int J Parasitol. 2006 (36:1227-39).

ACKNOWLEDGEMENTS/DANKWOORD
125
ACKNOWLEDGEMENTS/DANKWOORD
Na ruim 5 jaar is het dan zover, mijn doctoraat is klaar. Als afsluiter van deze fantastische en
leerrijke periode is een welgemeende dankjewel zeker op zijn plaats. Want zonder de
rechtstreekse of onrechtstreekse medewerking van een heus team schitterende mensen, had ik
dit eindresultaat nooit tot stand kunnen brengen.
In eerste instantie wil ik onze vakgroepvoorzitter Prof. Dr. Jozef Vercruysse bedanken om mij
de kans en het vertrouwen te geven te doctoreren in het labo Parasitologie. Bedankt om mij
steevast de meest pragmatische en realistische manier van aanpakken bij te brengen.
Daarnaast wil ik u ook bedanken voor de leuke labo-uitstapjes waarmee u ons plezierde.
Mijn 2 promotoren, Prof. Dr. Peter Geldhof en Prof. Dr. Edwin Claerebout, wil ik bedanken voor
hun wetenschappelijke input in mijn project en om te blijven geloven in het resistentie
onderzoek. Jullie kritische bijdragen waren van wezenlijk belang voor het tot stand komen van
dit doctoraat. Bedankt voor jullie steun, begeleiding en constructieve opmerkingen. Dankzij
jullie heb ik mijn kennis over parasitologie en anthelminthicumresistentie kunnen uitbreiden en
kreeg ik opportuniteiten om op internationaal niveau mijn werk te presenteren.
I would also like to thank the other members of my reading and exam committee, Prof. Dr. F.
Pasmans, Prof. Dr. A. Wolstenholme, Dr. A. Lespine and Dr. D. Bartley, to find the time to add
some critical and intersting notes to this thesis. All your suggestions really improved my work.
Ook wil ik Prof. Dr. R. Ducatelle bedanken voor het voorzitten van mijn examencommissie en
mijn openbare verdediging.
Annelies en Abdel, op jullie werk mocht ik verder bouwen, bedankt om mij als groentje op te
vangen en wegwijs te maken doorheen het resistentie onderzoek. Now my project has come to
an end, I pass the torch to Kasia and wish her all the best!
Dankzij de geniale uitvindingen van de ‘poopbag’ en de ‘metalen-trechter-bak’ konden we
steeds genoeg eieren en L3’s verzamelen! Petje af voor de ‘Masters in Coprology’, Stijn en
Nathalie!
Dankjewel Prisca en Iris voor jullie technische ondersteuning in het labo. Ook een dikke merci
aan Ellen om zo vaak mijn sequenties en bestellingen door te voeren en de bijkomende
problemen op te lossen. Dirk, Mieke en Louise bedankt voor de hulp bij alle IT, logistieke en
administratieve zaken.
Aangezien ik enorm gesteld ben op orde en netheid J wil ik Annie, Marijke, Erna, Vera en Jana
van ISS bedanken voor hun poetsinspanningen! Alsook Rudy voor het afwassen en autoclaveren
van het labomateriaal.

ACKNOWLEDGEMENTS/DANKWOORD
126
Gedurende mijn doctoraat heb ik ongeveer 155 000 km afgelegd en gelukkig niet alleen!
Bedankt Johannes, Mélissa, Abdel en Femke om te carpoolen wanneer het mogelijk was, dankzij
jullie werden het aangename ritten.
Natuurlijk wil ik ook alle andere (ex-)collega-parasitologen bedanken voor de toffe werksfeer,
de gezellige babbels en jullie helpende handen tijdens dierproeven. Maar ook na de werkuren
bracht jullie aanstekelijk enthousiasme de ambiance er in tijdens de onvergetelijke ‘ladies-
nights’, ‘trip to Italy’, paintball sessies, PARA-weekendjes, ... Bedankt daarvoor!
Tenslotte een bijzondere dankjewel aan mijn vrienden, familie, ouders en Steve! Bedankt voor
de interesse die jullie toonden in mijn werk, jullie aanmoedigingen, de tijd die jullie vrijmaakten
voor mij, maar ik niet altijd had voor jullie, ... Alle plannen die we hadden opgeborgen omwille
van mijn doctoraat, die gaan we nu waarmaken!
Jessie
This research was funded by a Ph.D. grant of the Agency for Innovation by Science and
Technology (IWT)

127
The more you are motivated by love
The more fearless and free your action will be
Dalai Lama
