phenotypic and functional features of myofibroblasts in sheep fetal wounds
TRANSCRIPT

Differentiation (1994) 56: 173-181 Differentiation On togeny, Neoplasl a and Dlfferentlatlon Therapy
0 Springer-Verlag 1994
Phenotypic and functional features of myofibroblasts in sheep fetal wounds James M. Estes ', Jerry S. Vande Berg ', N. Scott Adzick ', Thomas E. MacGillivray ', Alexis Desmoulih-e 3, Giulio Gabbiani 'Department of Surgery and The Fetal Treatment Center, University of California, San Francisco, California, USA *Core Clinical and Research Electron Microscopy Laboratory, Veterans Administration Medical Center, La Jolla, California, USA 'Department of Pathology, University of Geneva, Switzerland
Accepted in revised form November 22, 1993
Abstract. The myofibroblast is a mesenchymal cell with functional and structural characteristics in common with fibroblasts and smooth muscle cells. These cells play a critical role in wound closure and in the pathologic seque- lae of healing. It has been shown in adult humans and experimental animals that the myofibroblast expresses a -smooth muscle actin (ASMA) temporarily during wound contraction and more persistcotly during fibrocontractive diseases; however, it is unclear whether this cell makes any contribution to tissue repair in utero. Experimental work in fetal animal models has demonstrated that wound repair in fetal skin occurs by reconstitution of epidermal appendages and organized restoration of the dermal colla- gen network. Fetal lamb wound healing studies have shown that a transition from scarless tissue repair to heal- ing with scar formation occurs late i n gestation.
In this study we examined the ontogeny of myofibrob- lasts in fetal lamb wounds at early through late gestation, using transmission electron microscopy (TEM) and AS- MA immunohistochemistry.
Dramatic differences were observed in ASMA content of early as compared to late gestation fetal wound granu- lation tissue: ASMA was absent in wounds made at 75 days gestation but was present in progressively greater amounts in wounds made at 100 and 120 days gestation (term=145 days). TEM studies also demonstrated pro- gressive development and organization of microfilament bundles. Early in development microfilament bundles were sparse and disorganized, but as gestation progressed the bundles became more prevalent and formed tightly parallel arrangements. The organization of microfilament bundles was also accompanied by fibronexus formation. Thus, the ASMA expression in granulation tissue rnyofi- broblasts appears in relation to healing with scar forma- tion.
Additional in vitro experiments demonstrated that the majority of fibroblasts cultured from granulation tissue of
Correspondence to: G. Gabbiani, Department of Pathology, Univer- sity of Geneva, Centre MCdical Universitaire 1 , rue Michel-Servet, CH-1211 Geneva 4, Switzerland
75, 100 and 120 days gestation lambs expressed ASMA, suggesting that factors present in fetal calf serum can in- duce ASMA expression even in fibroblasts not expressing this protein in vivo.
Our observations demonstrate an ontogenic change in the fetal myofibroblast in response to injury in vivo and suggest that local factors may regulate the variable pheno- type of rnyofibroblasts during gestation. These results have implications for the therapy of pathologic scarring.
Introduction
The myofibroblast is a mesenchymal cell sharing ultra- structural features with fibroblasts and smooth muscle (SM) cells [37]. It was first identified in granulation tissue [ 161 and was believed to play an important role in wound closure and contracture [17, 29, 32, 35, 40, 461. The my- ofibroblast was also observed in fibrocontractive diseases such as fibromatoses [15], hepatic cirrhosis [6,33], renal and pulmonary fibrosis [ 1, 3 I], Dupuytren's contracture [47], and neoplasia-induced desmoplastic reactions [38] (for review of all these situations see [37]). The hallmark of the myofibroblastic phenotype is expression of a- smooth muscle actin (ASMA), the actin isoform prevalent in vascular SM cells. ASMA is temporarily expressed dur- ing normal wound healing and more permanently during fibrocontractive diseases (for review see [36]).
The presence of myofibroblasts in normal healing and in the pathologic sequelae of healing, such as scarring and contracture, is well established. However, it is unclear what contributions this cell makes to tissue repair in utero, which appears to occur by a different process than postna- tal tissue repair. The fetus possesses the unique ability to heal skin wounds without scar formation [2, 3, 9, 18, 25, 341. This observation was first extended to clinical situa- tions during the human fetal surgery experience at the Fetal Treatment Center of the University of California, San Francisco, USA [19]. Experimental work in several fetal animal models has demonstrated that this process

174
occurs by reconstitution of epidermal appendages and or- ganized restoration of the dermal collagen network [26, 481.
Longaker et al. have previously shown that fetal exci- sional wounds in lambs at 100 days gestation contract and contain myofibroblasts, based on morphologic and im- munohistochemical criteria [27]. In this study we exam- ined the development and ultrastructural features of my- ofibroblasts in fetal lamb wounds made at 75 through 120 days gestation (term=145 days) using transmission elec- tron microscopy and ASMA immunohistochemistry. Our purpose was to investigate possible differences between a wound which heals without scar formation (e.g. at 75 days gestation) and another one which heals with scarring (e.g. 100 and 120 days gestation). Our results indicate that AS- MA expression appears only in myofibroblasts of wounds healing with scarring and contracture.
Methods
Surgical procedure. The techniques of fetal lamb surgery have been described previously [20]. Briefly, six time-dated pregnant ewes of mixed Western breed (Torrel Farms, Ukiah, Calif., USA) underwent midline laparotomy at 75, 100 and 120 days gestation (two animals at each gestational age). Fetuses were exposed by hysterotomy of the gravid horn of the bicornuate uterus. Multiple full thickness, cutaneous excisional wounds were created with a 9 mm circular punch and India ink was applied to mark the wound base. The excisional wounds and normal skin were harvested 3 or 7 days later and processed as described below.
Cells and culture conditions. Sheep fibroblasts were obtained from explants of granulation tissue from 75, 100 and 120 days gestation. Cells were cultured in Eagle's Minimum Essential Medium (Gibco AG, Basel, Switzerland) supplemented with 100 units/ml penicillin, 100 Fg/ml streptomycin and 2 mM L-glutamine and containing 10% fetal calf serum (FCS, Gibco AG). Cultures were incubated at 37" C in a humidified atmosphere of 5% C02/95% air with medium changes twice a week. Cells were harvested at confluence by trypsinization and passed using a 1: 2 split ratio. Immunohistochem- ical and biochemical staining of cultured cells were performed from the 3rd to the 8th passage and yielded always similar results.
Antibodies, avidin-biotin-complex peroxidase (ABC-P) and im- munofluorescence staining. We used the following antibodies: anti- aSM-1 [42], an affinity purified rabbit polyclonal anti-desmin anti- body [23], an affinity purified rabbit polyclonal anti-SM myosin antibody [5], a rabbit polyclonal anti-actin antibody recognizing all actin isoforms [42], and a monoclonal antibody (HHF 35) which is also known to react with muscle actin isoforms [32, 461.
Immunoperoxidase staining for ASMA was done on ethanol- fixed and paraffin-embedded tissues. Sections were pretreated with H,O, and subsequently with 0.1 M periodic acid, 0.005 M NaBH, and normal horse serum. They were incubated for 2 h with anti-a SM-1 diluted 1: 200. This first incubation was followed by ABC-P staining using Vectastain kit anti-mouse IgG (Vector Laboratories, Burlingame, Calif., USA). The revelation of peroxidase activity was done with 3-amino-9-ethylcarbazole (Sigma Chemical, St. Louis, Mo., USA). Slides were weakly counterstained with Mayer's hema- toxylin.
Immunofluorescence was performed on fixed cells in culture dishes. For double indirect immunofluorescence, the following combinations were used: anti-aSM- Vanti-actin antibody, and anti-aSM-.l/anti-desmin antibody. Tetramethylrhodamine isothio- cyanate (TR1TC)-labeled goat anti-mouse IgG (Nordic Immunolog- ical Laboratories, Tilburg, The Netherlands) and fluorescein isoth-
iocyanate (F1TC)-conjugated goat anti-rabbit IgG (Cappel Labora- tories, Cochranville, Pa., USA) were used for the second step. The percentage of a-SM actin, desmin and SM-myosin positive cells was evaluated independently by two researchers on 300 to 500 cells.
Controls were performed by using mouse or rabbit non-immune IgGs or by omitting the primary antibody. Preparations were mount- ed and observed on a Zeiss Axiophot microscope equipped with epiillumination and specific filters for rhodamine and fluorescein (Carl Zeiss, Oberkochen, Germany). Photographs were taken, using 10x/0.30 Plan Neofluar to 63~11.40 oil Plan Apochromat objec- tives, on Kodak Ektachrome EPY-64T film or T-Max Kodak black and white film (Kodak, England).
Electron microscopy. Wound biopsies were immediately removed at surgery and placed into cold (4" C) 0.1 M phosphate buffered (pH 7.4) Karnovsky's fixative and shipped cold by overnight express for further processing. Under a dissecting microscope biopsies were then sliced into small 1-2 mm pieces in Karnovsky's buffered fixa- tive in order to orient the wound margin in the middle of the block face. Fixation was continued for an additional hour before washing three times in cold (4" C) 0.1 M sodium phosphate buffer (pH 7.0). Following three buffer washes, the tissues were post-fixed in 0.1 M phosphate buffered 2% osmium tetroxide (pH 7.4) for 1 h at room temperature. After three buffer washes of 5 min each the tissues were dehydrated through a graded series of ethanol and propylene oxide and embedded in plastic resin (EM-bed 812, EM Sciences, Fort Washington, Pa., USA). Thick sections of all tissue blocks were stained with 1% toluidine blue and oriented by light microscopy. From these sections, areas of the wound margin were thin sectioned (60 nm), mounted on 200 mesh unsupported mesh grids, and stained in uranyl acetate and bismuth subnitrate. Multiple sections were examined and photographed with a Zeiss EM-IOB electron micro- scope.
SDS-PAGE and immunoblotting. For SDS-PAGE, cultured cells were trypsinized at confluence and, after hemacytometer cell counts, were mixed in buffer containing 1% sodium dodecyl sulfate (SDS, Bio-Rad Laboratories AG, Glattbrugg, Switzerland), 1 % dithiothre- itol (Fluka Chemie AG, Buchs, Switzerland), 1 mM phenylmethyl- sulfonyl fluoride (Merck, Darmstadt, Germany), 1 mM N a -P-To- syl-L-arginine methylester (Sigma Chemical) in 0.4 M TRIS HCI pH 6.8 [13]. The samples were immediately sonicated and boiled for 3 min. Protein content was determined according to Bradford [8]. Forty micrograms protein was electrophoresed on a 5-20% gradient gel and stained with Coomassie blue. For quantification of total actin, gels were scanned with a computerized laser beam densito- meter (Genofit SA, Geneva, Switzerland) as previously described [ l l , 241.
Western blotting with anti-total actin and anti-aSM-1 was per- formed according to Skalli et al. [42]. Briefly, 5 to 20 pg protein were electrophoresed on a 5 2 0 % gradient gel. Separated proteins were transferred to nitrocellulose filters [45] which were incubated with a rabbit affinity purified IgG recognizing all actin isoforms or with anti-aSM-1. A second incubation was dpne either with sheep anti-rabbit or a rabbit anti-mouse IgG conjugated with horseradish peroxidase (Nordic Immunological Laboratories). Enhanced chemi- luminescence (Amersham International plc, Buckinghamshire, UK) was used for detection.
Results
The surgical procedures were well tolerated and there was no fetal or maternal morbidity. Analysis of non-wounded skin at all gestational ages showed ASMA staining only in small blood vessels (Fig. l a and b). Immunohistochemical staining for ASMA revealed marked differences between wounds made at 75 days and those at 100 and 120 days gestation. ASMA staining in granulation tissue was absent in 75 day gestation wounds except for pericytes and vascu-

Fig. 1. Alpha-smooth muscle actin (ASMA) staining in fetal wounds. a, b normal skin at 75 and 100 days gestation respectively. Note the preqence of a similar stain around small blood vessels in both cases. Hair follicles are visible at 100 days gestation. A positive staining for ASMA is present in fibroblastic cells surrounding these follicles [22]. c, d 3-day-old wound at 75 and 100 days gestation respectively. In (c), staining is present only around small blood ves-
sels. In (a), there is a substantial staining of fibroblastic cells in the wound bed. e, f ASMA staining in a 7-day-old wound at 75 and 100 days gestation respectively. g, h higher power micrographs of ASMA staining in a 7-day-old wound at 75 and 100 days gestation respec- tively. In (e) and (g), staining remains evident only around small blood vessels. In (0 and (h), most fibroblastic cells in the wound bed are positive to ASMA staining. a-f, x400; g, h, ~800

176
lar smooth muscle (positive control) (Fig. lc, e and g). In contrast, granulation tissue from 100 day gestation wounds revealed substantial ASMA staining in fibroblas- tic cells; this staining increased from 3 to 7 days post wounding. At 3 days post wounding the density of ASMA positive cells was sparse (Fig. Id), while at 7 days ASMA- positive cells comprised approximately 90% of the granu- lation stromal cells (Fig. If and h). ASMA staining was also strongly positive in 7-day-old wounds made at 120 days gestation (data not shown). The presence of ASMA was also confirmed in all wounds by staining with HHF 35 (data not shown), a monoclonal antibody which is known to react with SM actin [46]. Desmin and SM myosin staining were always negative in wound stromal cells.
Fig. 2. Seventy-five day gestation, 7-day-old wound. The myofibrob- last contains patches of small mi- crofilament bundles with electron dense bodies (arrowheads). Mi- crofilament bundles were found near Golgi bodies (G) and the cell membrane. Bar, 0.4 pm Fig. 3. Seventy-five day gestation, 7-day-old wound. The most well differentiated myofibroblast at 75 days displayed bundles that .were composed of loosely arranged mi- crofilaments (MF) among electron dense bodies. Ribosomal clusters (RC) appeared near the rough en- doplasmic reticulum. Polyribo- somes (arrowheads) sometimes associated with small filaments were observed between ribosomal clusters and microfilament bun- dles. Bar, 0.2 pm
Electron microscopic analysis was performed on the excisional wounds and normal skin. Tissue from wounds made at 75, 100 and 120 days gestation revealed consider- able variation in the cytoskeletal features characteristic of myofibroblasts. Seven-day-old wounds made at 75 days gestation presented cells which appeared to be in the early stages of microfilament development. Evidence for this interpretation was derived from the presence of loose clusters of randomly-oriented microfilaments (60 to 80 Angstroms) and electron dense bodies. The hallmark of these organizational sites was represented by clusters of polyribosomes and concentrated areas of microfilaments. In other later stage myofibroblasts, small patches of more organized bundles were observed between Golgi bodies and the cell membrane (Fig. 2). The most developed my-

177
ofibroblasts presented longer microfilament bundles with electron dense bodies along the cell membrane (Fig. 3). Higher magnification showed that these microfilaments remained loosely arranged among electron dense bodies.
In wounds made at 75 days gestation, fibronexi were present but appeared to be very undeveloped (data not shown). Occasionally some cells demonstrated these transmembrane complexes between clusters of organizing intracellular microfilaments along with sparsely dis- tributed collagen fibrils in the extracellular matrix.
Seven-day-old wounds made at 100 days gestation showed increased numbers of myofibroblasts. Bundles of microfilaments with electron dense bodies were well formed along the long axis of the cell (Fig. 4). Unlike 7-day-old wounds made at 75 days gestation, microfila-
Fig. 4. One hundred day gestation, 7-day-old wound. Most observa- tions displayed myofibroblasts with microfilament bundles (MF) composed of parallel, tightly wo- ven microfilaments among elec- tron dense bodies. An undeveloped fibronexus (FN) was seen with at- tachments to collagen fibers (C). Clusters of glycogen (GL) parti- cles were observed in many my- ofibroblasts and small amounts of fibrin (F) were also noted. Bur, 0.4 l m Fig. 5. One hundred twenty day gestation, 7-day-old wound. Net- works of myofibroblasts were joined via fibronexi (FN) between microfilament bundles (MF) of neighboring cells. These syncytial arrangements of myofibroblasts appeared throughout the wound bed. Bur, 2.0 Fm
ments from cells at the same interval of wound repair at 100 days bestation were parallel and tightly packed, form- ing broad microfilament bundles (Fig. 4). Many of these latter cells also displayed noticeable clusters of glycogen throughout their cytoplasm (Fig3. 4). Like their 75 day ges- tation counterparts, myofibroblasts from 100 day gesta- tion wounds contained well developed Golgi bodies and rough endoplasmic reticulum.
In areas where new collagen fibrils and fibrin appeared near the cell membrane, fibronexus junctions appeared to be more developed. Some myofibroblasts displayed these transmembrane complexes parallel to the long axis of the cell as well as near the tips of cell processes (Fig. 4).
Seven-day-old fetal wounds made at 120 days gestation exhibited microfilament bundles that appeared to be more
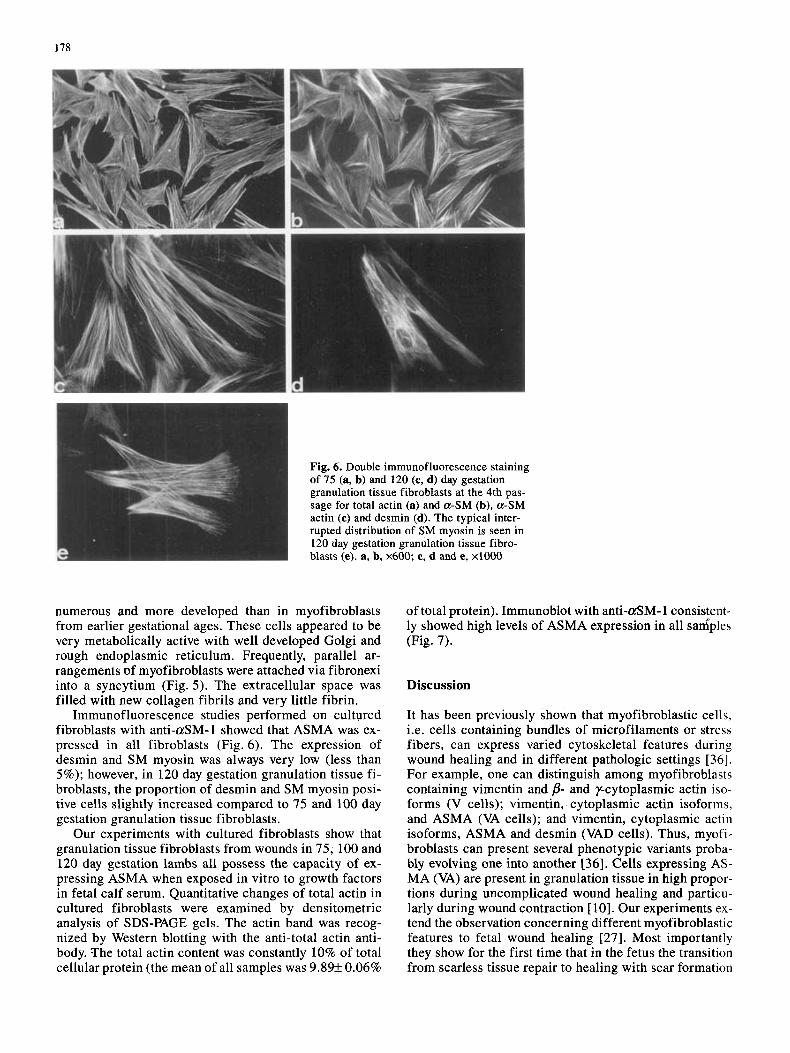
178
numerous and more developed than in myofibroblasts from earlier gestational ages. These cells appeared to be very metabolically active with well developed Golgi and rough endoplasmic reticulum. Frequently, parallel ar- rangements of myofibroblasts were attached via fibronexi into a syncytium (Fig. 5 ) . The extracellular space was filled with new collagen fibrils and very little fibrin.
Immunofluorescence studies performed on cultured fibroblasts with anti-aSM-1 showed that ASMA was ex- pressed in all fibroblasts (Fig. 6). The expression of desmin and SM myosin was always very low (less than 5%); however, in 120 day gestation granulation tissue fi- broblasts, the proportion of desmin and SM myosin posi- tive cells slightly increased compared to 75 and 100 day gestation granulation tissue fibroblasts.
Our experiments with cultured fibroblasts show that granulation tissue fibroblasts from wounds in 75, 100 and 120 day gestation lambs all possess the capacity of ex- pressing ASMA when exposed in vitro to growth factors in fetal calf serum. Quantitative changes of total actin in cultured fibroblasts were examined by densitometric analysis of SDS-PAGE gels. The actin band was recog- nized by Western blotting with the anti-total actin anti- body. The total actin content was constantly 10% of total cellular protein (the mean of all samples was 9.89+ 0.06%
of total protein). Immuaoblot with anti-aSM- 1 consistent- ly showed high levels of ASMA expression in all sadples (Fig. 7).
Discussion
It has been previously shown that myofibroblastic cells, i.e. cells containing bundles of microfilaments or stress fibers, can express varied cytoskeletal features during wound healing and in different pathologic settings [36]. For example, one can distinguish among myofibroblasts containing vimentin and p- and y-cytoplasmic actin iso- forms (V cells); vimentin, cytoplasmic actin isoforms, and ASMA (VA cells); and vimentin, cytoplasmic actin isoforms, ASMA and desmin (VAD cells). Thus, myofi- broblasts can present several phenotypic variants proba- bly evolving one into another [36]. Cells expressing AS- MA (VA) are present in granulation tissue in high propor- tions during uncomplicated wound healing and particu- larly during wound contraction [ 101. Our experiments ex- tend the observation concerning different myofibroblastic features to fetal wound healing [27]. Most importantly they show for the first time that in the fetus the transition from scarless tissue repair to healing with scar formation

179
a b c d e f
Fig. 7. ASMA expression in 75 (a, d), 100 (b, e), and 120 (c, f) day gestation granulation tissue fibroblasts. Densitometric scanning of Coomassie blue-stained gels (a-c) shows that the percentage of actin per total protein is not modified (for value, see Results). Immuno- blotting after SDS-PAGE (d-f) shows the high level of ASMA ex- pression (A=actin)
coincides with the expression of ASMA by myofibrob- lasts and thus support the possibility that ASMA is not only a marker of granulation tissue contraction but partic- ipates actively in this phenomenon.
Our in vitro data, showing that fibroblasts cultured from 75,100 and 120 day granulation tissues express sim- ilar amounts of ASMA, suggest that the difference in AS- MA expression by granulation tissue fibroblasts observed in vivo between 75 and 100 or 120 day gestation lambs is due to different microenvironmental conditions in vitro and in vivo rather than to intrinsic fibroblastic properties. Further studies will be necessary to know whether in wounds made at 75 days gestation factors inhibiting AS- MA expression after cloning are produced and/or whether cultured fibroblasts from 75 day gestation animals are stimulated to produce ASMA by serum factors. It has been shown previously that different fibroblastic popula- tions and clones in vitro contain a variable proportion of ASMA-expressing cells [12]. However, it is unclear what microenvironmental factors are implicated in ASMA ex- pression. Recently we have shown that heparin [13] and transforming growth factor-Pl (TGF-P1) [ 141, albeit with different mechanisms, stimulate the appearance of AS- MA-rich fibroblasts both in vivo and in vitro. Other inves- tigators have demonstrated that TGF-P increases the ex- pression of ASMA in cultured smooth muscle cells [7] and a short communication has suggested that this cy- tokine exerts the same action on cultured fibroblasts [30]. TGF-P has been shown to play a role in the production of wound contraction and scars in vivo [39]. Moreover the role of TGF-P in the regulation of collagen synthesis dur- ing wound healing is well established [44]. All these data
point to TGF-P as a regulator of myofibroblast differenti- ation [14]. This cytokine appears to act during wound healing in a coordinated fashion on the metabolism of extracellular matrix components (e.g. collagen) and on the regulation of cellular contractility (e.g. ASMA ex- pression). Work is in progress in our laboratories in order to evaluate TGF-P expression during wound healing at different gestation times. Several other cytokines and ex- tracellular matrix components have also been proposed to modulate ASMA expression in myofibroblasts (for review see [36]).
Other lines of investigation have demonstrated the in- fluence of cell adhesion on the expression of actin in gen- eral as well as ASMA. Yamada et al. [50] and Ali et al. [4] showed that addition of a matrix glycoprotein could re- store cell adhesion and actin synthesis in transformed chick embryo fibroblasts. Willingham et al. [49] also showed that normal fibroblasts produce well organized actin bundles when cultured on an adherent substratum and that this property was lost after dissociation. Later, Singer et al. [40] demonstrated that fibronectin is one of the adhesion molecules that induces actin synthesis in fi- broblasts.
In our study, fully formed microfilament bundles, typi- cally found in normal adult granulating wounds, were not found in wounds made at 75 days gestation. This may be due to an insufficient level of growth factor stimulation in the microenvironment or decreased levels of cell adhesion factors. Further studies addressing these questions are in progress.
The appearance of ASMA at 100 days gestation re- flects a progression of the myofibroblast phenotype from fibroblastic to smooth muscle-like. This transition may have important implications for wound healing as the per- sistence in wounds of ASMA-positive cells is associated with hypertrophic scarring and fibromatoses [41, 431. Progressive cellular differentiation may occpr throughout gestation in wound myofibroblasts, thus influencing tis- sue repair by causing more wound contraction and eventu- ally scar formation. Experimental evidence has demon- strated such a wound healing transition in fetal lambs at approximately 100 days gestation [27]. A similar transi- tion has been described in fetal rat wounds [21], in fetal rhesus monkey wounds [28], and in mesenchymal cells of human embryonic and adult colonic muscularis mucosae W I .
Our study characterizes an ontogenic transition of the myofibroblast phenotype in fetal lamb wounds. The ac- quisition and organization of ASMA rich microfilaments in wound myofibroblasts later in gestation suggests an increasingly important role for wound contraction during fetal development. This phenomenon may be related to the development of scar in the third trimester. A better under- standing of factors modulating the fibroblastic phenotype during wound healing could lead to therapeutic strategies directed against myofibroblast proliferation or differenti- ation, and these may be useful in the prevention of patho- logic scarring and fibrosis.
Acknowledgements. Supported by NIH grants HD 25505 and GM 27348 (N.S.A.), an NIH National Research Service Award

180
HD 07462 (J.M.E.), Veterans Administration Medical Research De- partment (J.S.V.), and the Swiss National Science Foundation Grant No. 31-30796.91 (G.G.). We thank Mrs. G. Chakroun for excellent technical assistance.
References
1. Adler KB, Craighead JE, Vallyathan NV, Evans JN (1981) Actin containing cells in human pulmonary fibrosis. Am J Pathol
2. Adzick NS, Longaker MT (1992) Scarless fetal wound healing: Therapeutic implications. Ann Surg 215: 3-7
3. Adzick NS, Harrison MR, Glick PL, Beckstaed JL, Villa RL, Scheuenstuhl H, Goodson WH (1985) Comparison of fetal, newborn, and adult wound healing by histologic, enzyme-histo- chemical, and hydroyoproline determinations. J Pediatr Surg
4. Ali IU, Mautner R, Lanza R, Hynes RO (1977) Restoration of normal morphology; adhesion and cytoskeleton in transformed cells by addition of a transformation-sensitive surface protein. Cell 11: 115-126
5. Benzonana G, Skalli 0, Gabbiani G (1988) Correlation between the distribution of smooth muscle or non muscle myosins and a -smooth muscle actin in normal and pathological soft tissues. Cell Motil Cytoskeleton 11: 260-274
6. Bhathal PS (1972) The presence of modified fibroblasts in cir- rhotic livers in man. Pathology 4: 139-144
7. Bjorkerud S (1991) Effects of transforming growth factor-p 1 on human arterial smooth muscle cells in vitro. Arterioscler Thromb 1 1 : 892-902
8. Bradford MM (1976) A rapid and sensitive method for the quan- titation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 72: 248-254
9. Burrington JD (1971) Wound healing in the fetal lamb. J Pediatr Surg 6: 523-527
10. Darby I, Skalli 0, Gabbiani G (1990) a-Smooth muscle actin is transiently expressed by myofibroblasts during experimental wound healing. Lab Invest 63: 21-29
11. Desmoulitre A, Rubbia-Brandt L, Gabbiani G (1991) Modula- tion of actin isoform expression in cultured arterial smooth mus- cle cells by heparin and culture conditions. Arterioscler Thromb
12. Desmoulikre A, Rubbia-Brandt L, Abdiu A, Walz T, Macieira- Coelho A, Gabbiani G (1992) a-Smooth muscle actin is ex- pressed in a sub-population of cultured and cloned fibroblasts and is modulated by yinterferon. Exp Cell Res 201: 64-73
13. Desmoulikre A, Rubbia-Brandt L, Grau G, Gabbiani G (1992) Heparin induces a-smooth muscle actin expression in cultured fibroblasts and in granulation tissue myofibroblasts. Lab Invest
14. Desmoulikre A, Geinoz A, Gabbiani F, Gabbiani G (1993) Transforming growth factor$] induces a-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J Cell Biol 122: 103-1 11
15. Gabbiani G, Majno G (1972) Dupuytren’s contractwe: Fibrob- last contraction? An ultrastructural study. Am J Pathol 66: 131- 146
16. Gabbiani G, Ryan GB, Majno G (1971) Presence of modified fibroblasts in granulation tissue and their possible role in wound contracture. Experientia 27: 549-552
17. Gabbiani G, Hirschel BJ, Ryan GB, Statkov PR, Majno G (1972) Granulation tissue as a contractile organ: A study of structure and function. J Exp Med 135: 719-734
18. Goss AN (1977) Intrauterine healing of fetal rat oral mucosa, skin, and cartilage. J Oral Pathol 6: 35-38
19. Harrison MR, Adzick NS (1991) The fetus as a patient: Surgical implications. Ann Surg 2 13: 279-290
20. Harrison MR, Jester JA, Ross NA (1980) Correction of congen- ital diaphragmatic hernia in utero. I. The model: Intrathoracic
102: 427-437
20:315-319
11: 244-253
67: 716-726
balloon produces fatal pulmonary hypoplasia. Surgery 88: 174- 182
21. Ihara S, Motobayashi Y (1992) Wound closure in foetal rat skin. Development 114: 573-582
22. Jahoda CAB, Reynolds AJ, Chaponnier C, Forester JC, Gabbiani G (1991) Smooth muscle a-actin is a marker for hair follicle dermis in vivo and in vitro. J Cell Sci 99: 627-636
23. Kocher 0, Skalli 0, Bloom WS, Gabbiani G (1984) Cytoskele- ton of rat aortic smooth muscle cells: Normal conditions and experimental intimal thickening. Lab Invest 50: 645-652
24. Kocher 0, Skalli 0, Cerutti D, Gabbiani F, Gabbiani G (1985) Cytoskeletal features of rat aortic cells during development: An electron microscopic, immunohistochemical, and biochemical study. Circ Res 56: 829-838
25. Krummel TM, Nelson JM, Diegelmann RF, Lindblad WJ, Salzberg AM, Greenfield LJ, Cohen IK (1987) Fetal response to injury in the rabbit. J Pediatr Surg 22: 640-644
26. Longaker MT, Whitby DJ, Adzick NS, Crombleholme TM, Langer JC, Duncan BW, Bradley SM, Stern R, Ferguson MWJ, Harrison MR (1990) Studies in fetal wound healing. VI. Second and early third trimester fetal wounds demonstrate rapid colla- gen deposition without scar formation. J Pediatr Surg 25: 63- 69
27. Longaker MT, Burd DAR, Gown AM, Yen TSB, Jennings RW, Duncan BW, Harrison MR, Adzick NS (1991) Midgestational excisional fetal lamb wounds contract in utero. J Pediatr Surg
28. Lorenz HP, Whitby DJ, Longaker MT, Adzick NS (1993) Fetal wound healing: The ontogeny of scar formation in the non-hu- man primate. Ann Surg 217: 391-396
29. Majno G, Gabbiani G, Hirschel BJ, Ryan GB, Statkov PR (197 1) Contraction of granulation tissue in vitro: Similarity to smooth muscle. Science 173: 548-550
30. Mitchell JJ, Woodcock-Mitchell J, Low RB, Absher M (1991) Modulation of a-smooth muscle actin by growth state and TGF- p in lung myofibroblasts. J Cell Biol 115: 161a
31. Nagle RB, Kneiser MR, Bulger RE, Benditt EP (1973) Induction of smooth muscle characteristics in renal interstitial fibroblasts during obstructive nephropathy. Lab Invest 29: 422-427
32. Oda D, Gown AM, Vande Berg JS, Stern R (1988) The fibro- blast-like nature of myofibroblasts. Exp Mol Pathol 49: 316- 329
33. Ramadori G (1991) The stellate cell (Ito-cell, fat-storing cell, lipocyte, perisinusoidal cell) of the liver. New insights into pathophysiology of an intriguing cell. Virchows Arch [B]
34. Rowsell AR (1986) The intra-uterine healing of fetal muscle wounds: Experimental study in the rat. Br J PlastSurg 37: 635- 642
35. Sappino AP, Dietrich PY, Widgren S, Gabbiani G (1989) Colonic pericryptal fibroblasts: Differentiation patterns in em- bryogenesis and phenotypic modulation in epithelial prolifera- tive lesions. Virchows Arch [A] 415: 551-557
36. Sappino AP, Schurch W, Gabbiani G (1990) Differentiation repertoire of fibroblastic cells: Expression of cytoskeletal proteins as markers of phenotypic modulations. Lab Invest
37. Schurch W, Seemayer TA, Gabbiani G (1992) Myofibroblast. In: Sternberg SS (ed) Histology for pathologists. Raven Press, New York, pp 109-144
38. Seemayer TA, Schurch W, Lagact R, Tremblay G (1979) Myofi- broblasts in the stroma of invasive and metastatic carcinoma. Am J Surg Pathol3: 525-533
39. Shah M, Foreman DM, Ferguson MWJ (1992) Control of scar- ring in adult wounds by neutralising antibody to transforming growth factor p. Lancet 339: 213-214
40. Singer 11, Kawka DW, Kazazis DM, Clark RAF (1984) In vivo codistribution of fibronectin and actin fibers in granulation tis- sue: Immunofluorescence and electron microscope studies of the fibronexus at the myofibroblast surface. J Cell Biol
26: 942-948
61: 147-158
63: 144-161
98: 2091-2106

181
41. Skalli 0, Gabbiani G (1988) The biology of the myofibroblast. Relationship to wound contraction and fibroconnective diseases. In: Clark RAF, Henson PM (eds) The molecular and cellular biology of wound repair. Plenum Publishing Corporation, New York, pp 373-402
42.Skalli 0, Ropraz P, Trzeciak A, Benzonana G, Gillessen D, Gabbiani G (1986) A monoclonal antibody against a-smooth muscle actin: A new probe for smooth muscle differentiation. J Cell Biol 103: 2787-2796
43. Skalli 0, Schurch W, Seemayer TA, Lagace R, Montandon D, Pittet B, Gabbiani G (1989) Myofibroblasts from diverse patho- logic settings are heterogeneous in their content of actin iso- forms and intermediate filament proteins. Lab Invest 60: 275- 285
44. Sporn MB, Roberts AB (1992) Transforming growth factor-p: Recent progress and new challenges. J Cell Biol 119: 1017-1021
45. Towbin H, Staehelin T, Gordon J (1979) Electrophoretic trans- fer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc Natl Acad Sci USA 76: 4350-4354
46. Tsukada T, McNutt MA, Ross R, Gown AM (1987) HHF 35, a muscle actin-specific monoclonal antibody. 11. Reactivity in normal, reactive, and neoplastic human tissues. Am J Pathol
47.Vande Berg JS, Rudolph R, Gelberman R, Woodward MR (1982) Ultrastructural relationship of skin to nodule and cord in Dupuytren’s contracture. Plast Reconstr Surg 69: 835-844
48. Whitby DJ, Ferguson MWJ (1991) The extracellular matrix of lip wounds in fetal, neonatal, and adult mice. Development
49. Willingham MC, Yamada KM, Yamada SS, Pousyssegur J, Pas- ten I(1977) Microfilament bundles and cell shape are related to adhesiveness in substratum and are dissociable from growth con- trol in cultured fibroblasts. Cell 10: 375-380
50. Yamada KM, Yamada SS, Plasten 1(1976) Cell surface protein partially restores morphology, adhesiveness, and contact inhibi- tion of movement in transformed fibroblasts. Proc Natl Acad Sci
127: 389-402
112~651-668
USA 73: 1217-1221