photosynthesis in silico - startseite...in advances in photosynthesis and respiration:...
TRANSCRIPT

Photosynthesis in silicoUnderstanding Complexity from Molecules to Ecosystems

Advances in Photosynthesis and Respiration
VOLUME 29
Series Editor:
GOVINDJEEUniversity of Illinois, Urbana, Illinois, U.S.A.
Consulting Editors:Julian EATON-RYE, Dunedin, New Zealand
Christine H. FOYER, Harpenden, U.K.David B. KNAFF, Lubbock, Texas, U.S.A.
Anthony L. MOORE, Brighton, U.K.Sabeeha MERCHANT, Los Angeles, California, U.S.A.
Krishna NIYOGI, Berkeley, California, U.S.A.William PARSON, Seatle, Washington, U.S.A.Agepati RAGHAVENDRA, Hyderabad, India
Gernot RENGER, Berlin, Germany
The scope of our series, beginning with volume 11, reflects the concept that photosynthesis andrespiration are intertwined with respect to both the protein complexes involved and to the entirebioenergetic machinery of all life. Advances in Photosynthesis and Respiration is a book seriesthat provides a comprehensive and state-of-the-art account of research in photosynthesis andrespiration. Photosynthesis is the process by which higher plants, algae, and certain species ofbacteria transform and store solar energy in the form of energy-rich organic molecules. Thesecompounds are in turn used as the energy source for all growth and reproduction in theseand almost all other organisms. As such, virtually all life on the planet ultimately depends onphotosynthetic energy conversion. Respiration, which occurs in mitochondrial and bacterialmembranes, utilizes energy present in organic molecules to fuel a wide range of metabolicreactions critical for cell growth and development. In addition, many photosynthetic organismsengage in energetically wasteful photorespiration that begins in the chloroplast with an oxy-genation reaction catalyzed by the same enzyme responsible for capturing carbon dioxide inphotosynthesis. This series of books spans topics from physics to agronomy and medicine,from femtosecond processes to season long production, from the photophysics of reactioncenters, through the electrochemistry of intermediate electron transfer, to the physiology ofwhole organisms, and from X-ray crystallography of proteins to the morphology or organellesand intact organisms. The goal of the series is to offer beginning researchers, advanced under-graduate students, graduate students, and even research specialists, a comprehensive, up-to-date picture of the remarkable advances across the full scope of research on photosynthesis,respiration and related processes.
For other titles published in this series, go to
www.springer.com/series/5599

Photosynthesis in silicoUnderstanding Complexity from Molecules
to Ecosystems
Edited by
Agu LaiskUniversity of Tartu
Estonia
Ladislav NedbalInstitute of Systems Biology and Ecology
Academy of Sciences of the Czech RepublicNové Hrady
Czech Republic
and
GovindjeeUniversity of Illinois at Urbana-Champaign
UrbanaIllinoisUSA

Library of Congress Control Number: 2009921443
ISBN 978-1-4020-9236-7 (HB)ISBN 978-1-4020-9237-4 (e-book)
Published by Springer,P.O. Box 17, 3300 AA Dordrecht, The Netherlands.
www.springer.com
Cover: Photosynthesis in silico. Photo by Eero Talts, University of Tartu
Printed on acid-free paper
All Rights Reservedc© 2009 Springer Science+Business Media B.V.
No part of this work may be reproduced, stored in a retrieval system, or transmitted in any form or by any means,electronic, mechanical, photocopying, microfi lming, recording or otherwise, without written permission from the Publisher,
with the exception of any material supplied specifi cally for the purpose of being entered and executed on a computersystem, for exclusive use by the purchaser of the work.

From the Series Editor
I am highly obliged to each and everyone of theauthors from fifteen countries(Australia, Austria,Brazil, Canada, China, Czech Republic, Esto-nia, France, Germany, Lithuania, Russia, Switzer-land, The Netherlands, U.K., and U.S.A.) fortheir valuable contributions to the successful andtimely production of this unique book, Volume 29in Advances in Photosynthesis and Respiration:‘Photosynthesis in silico—Understanding Com-plexity from Molecules to Ecosystems’, editedby two of the pioneers in the field (Agu Laisk,of Estonia; and Ladislav Nedbal, of the CzechRepublic).
The authors are: Niels P.R. Anten (TheNetherlands; Chapter 16); Michael J. Behrenfeld(U.S.A.; Chapter 20); Carl J. Bernacchi (U.S.A.;Chapter 10); Joseph Berry (U.S.A.; Chapter 9);Jan Cervený (Czech Republic; Chapter 2); JanP. Dekker (The Netherlands; Chapter 3); GeraldEdwards (U.S.A.; Chapter 14); Hillar Eichelmann(Estonia: Chapter 13); Hadi Farazdaghi (Canada;Chapter 12); Graham Farquhar (Australia; Chap-ters 9 and 10); Arvi Freiberg (Estonia; Chapter4); Andrew Friend (U.K.; Chapter 20); RichardJ. Geider (U.K.; Chapter 20); Jeremy Harbinson(The Netherlands; Chapter 11); HubertHasenauer (Austria; Chapter 19) Michael Hucka(U.S.A.; Chapter 1); Manfred Küppers (Ger-many; Chapter 18); Agu Laisk (Estonia; Chapters13 and 14); Jérôme Lavergne (France; Chapter8); Dušan Lazár (Czech Republic; Chapter 5);Stephen P. Long (U.S.A.; Chapters 10 and 17);Ladislav Nedbal (Czech Republic; Chapter 2);Ülo Niinemets (Estonia; Chapter 16); VladimirI. Novoderezhkin (Russia; Chapter 3); Vello Oja(Estonia; Chapter 13); Michael Pfiz (Germany;Chapter 18); Stephen A. Pietsch (Austria;Chapter 19); Carlos Pimentel (Brazil; Chapter10); Ondrej Prášil (Czech Republic; Chapter 6);Galina Riznichenko (Russia; Chapter 7); DavidM. Rosenthal (U.S.A.; Chapter 10); AndrewRubin (Russia; Chapter 7); James Schaff (U.S.A.;Chapter 1); Gert Schansker (Switzerland; Chap-ter 5); Henning Schmidt (Germany; Chapter 2);
Christopher J. Still (U.S.A.; Chapter 20); Paul C.Struik (The Netherlands; Chapter 11); GediminasTrinkunas (Lithuania; Chapter 4); Rienk vanGrondelle (The Netherlands; Chapter 3); Susannevon Caemmerer (Australia; Chapter 9); WimVredenberg (The Netherlands; Chapter 6); Ian E.Woodrow (Australia; Chapter 15); Xinyou Yin(The Netherlands; Chapter 11), and Xin-GuangZhu (China; Chapter 17).
I also thank Jacco Flipsen, Noeline Gibsonand André Tournois of Springer (Dordrecht, TheNetherlands) for their patience with me duringthe production of this book. I am particularlythankful to Albert André Joseph (of SPi Tech-nologies India Private Limited) for his wonder-ful cooperation in efficiently taking care of thecorrections in the proofs of this book. Finally, Iam grateful to my wife, Rajni Govindjee, and theoffices of the Department of Plant Biology (FengSheng Hu, Head) and of Information Technology,Life Sciences (Jeff Haas, Director), of the Uni-versity of Illinois at Urbana-Champaign, for theirconstant support.
A list of books, published under the Series‘Advances in Photosynthesis and Respiration’ isavailable at the Springer web site: <http://www.springer.com/series/5599>. Members of theISPR (International Society of PhotosynthesisResearch) receive 25 % discount. For the Tableof Content of most of the earlier volumes, see myweb site at: <http://www.life.uiuc.edu/govindjee/Reference-Index.htm>.
GovindjeeFounding Editor of Advances in Photosynthesis
and RespirationDepartments of Biochemistry and Plant Biology
and Center of Biophysics & ComputationalBiology
University of Illinois at Urbana-Champaign,Illinois, USA
E-mail: [email protected]
v

Contents
From the Series Editor v
Preface xv
The Editors xvii
Author Index xxi
Color Plates CP1
Part I: General Problems of Biological Modeling
1 Trends and Tools for Modeling in Modern Biology 3–15Michael Hucka and James Schaff
Summary 3I. Introduction 4II. Representing Model Structure and Mathematics 4III. Augmenting Models with Semantic Annotations 6IV. Connecting Models to Results 9V. Future Directions for Systems Biology Markup Language (SBML) 13VI. Conclusions 13References 14
2 Scaling and Integration of Kinetic Models of Photosynthesis:Towards Comprehensive E-Photosynthesis 17–29
Ladislav Nedbal, Jan Cervený and Henning Schmidt
Summary 17I. Introduction 18II. Mapping Partial Photosynthesis Models into the Comprehensive
Model Space (CMS): The Principles and Strategies 19III. Mapping of Photosystem II Models into the Comprehensive
Model Space (CMS) 21IV. Concluding Remarks 27Acknowledgments 28References 28
vii

Part II: Modeling of Light Harvesting and PrimaryCharge Separation
3 Modeling Light Harvesting and Primary Charge Separationin Photosystem I and Photosystem II 33–53
Rienk van Grondelle, Vladimir I. Novoderezhkin and Jan P. Dekker
Summary 34I. Introduction 34II. Physical Models of Energy Transfer 35III. Exciton Spectra and Energy Transfer in Photosystem I (PS I) Core 40IV. Excitation Dynamics in Major Light Harvesting Complex II (LHCII) 41V. Energy Transfers and Primary Charge Separation in Photosystem II
Reaction Center 44VI. Concluding Remarks 50Acknowledgments 50References 50
4 Unraveling the Hidden Nature of Antenna Excitations 55–82Arvi Freiberg and Gediminas Trinkunas
Summary 55I. Introduction 56II. Disordered Frenkel Exciton Model for Absorbing States
of Circular Antenna Aggregates 59III. Shortcomings of the Disordered Frenkel Exciton Model 63IV. Excitonic Polaron Model of the Antenna Fluorescing States 64V. Evaluation of the Model Parameters from the Experimental Spectra 70VI. Conclusions and Outlook 76Acknowledgments 77References 77
Part III: Modeling Electron Transport and Chlorophyll Fluorescence
5 Models of Chlorophyll a Fluorescence Transients 85–123Dušan Lazár and Gert Schansker
Summary 86I. Fluorescence Induction 86II. Approaches and Assumptions in the Modeling of the Fluorescence Rise 91III. Particular Models for the Fluorescence Rise 100IV. Modeling the Whole Fluorescence Induction 111V. Conclusions and Future Perspectives 115Acknowledgments 115References 115
viii

6 Modeling of Chlorophyll a Fluorescence Kinetics in Plant Cells:Derivation of a Descriptive Algorithm 125–149
Wim Vredenberg and Ondrej Prášil
Summary 126I. Introduction 126II. Variable (Chlorophyll) Fluorescence – Some Basics 128III. Application of Single, Twin and Multiple Turnover Flashes 132IV. Distinguishable Phases of Fluorescence Response
upon Multiturnover Excitation 135V. Fluorescence Induction Algorithm for Experimental Curves 137VI. Concluding Remarks 144Acknowledgments 146References 146
7 Modeling of the Primary Processes in a Photosynthetic Membrane 151–176
Andrew Rubin and Galina Riznichenko
Summary 151I. Introduction 152II. Fluorescence as an Indicator of the Photosystem State 153III. General Kinetic Model of the Processes in Photosynthetic
Thylakoid Membrane 154IV. Multiparticle Modeling of the Processes in the Photosynthetic Membrane 166V. Concluding Remarks and Future Perspectives 171Acknowledgments 171References 171
8 Clustering of Electron Transfer Components:Kinetic and Thermodynamic Consequences 177–205
Jérôme Lavergne
Summary 177I. Introduction 178II. Thermodynamic Performance of Integrated and Diffusive
Photosynthetic Models 179III. Integrated Versus Diffusive Electron Transfer Chain 183IV. The Small Apparent Equilibrium Constant in the Donor Chain
of Rhodobacter sphaeroides 189V. Quinone Domains 195VI. Statistical and Non Statistical Heterogeneities 198VII. Pool Function Test at Steady State 199VIII. Kinetic Analysis: Playing with Inhibitors, Redox Potential and Flash Intensity 201IX. Concluding Remarks 203Acknowledgments 203References 203
ix

Part IV: Integrated Modeling of Light and Dark Reactionsof Photosynthesis
9 Biochemical Model of C3 Photosynthesis 209–230Susanne von Caemmerer, Graham Farquhar and Joseph Berry
Summary 210I. Introduction 210II. The Rate Equations of CO2 Assimilation 211III. Parameters and their Temperature Dependencies 215IV. The Role of Rubisco Activation State 218V. Estimating Chloroplast pCO2 219VI. Predicting Photosynthesis from Chloroplast Biochemistry 220VII. Predicting Chloroplast Biochemistry from Leaf Gas Exchange 223VIII. Concluding Remarks 224References 225
10 Modeling the Temperature Dependence of C3 Photosynthesis 231–246Carl J. Bernacchi, David M. Rosenthal, Carlos Pimentel,Stephen P. Long and Graham D. Farquhar
Summary 232I. Introduction 232II. Processes Limiting to C3 Photosynthesis 233III. Modeling Photosynthesis and the Supply of CO2 240IV. Concluding Remarks 242Acknowledgments 243References 243
11 A Model of the Generalized Stoichiometry of Electron TransportLimited C3 Photosynthesis: Development and Applications 247–273
Xinyou Yin, Jeremy Harbinson and Paul C. Struik
Summary 247I. Introduction 248II. Model Development 250III. Model Applications 255IV. Concluding Remarks 269Acknowledgments 269References 270
12 Modeling the Kinetics of Activation and Reaction of Rubiscofrom Gas Exchange 275–294
Hadi Farazdaghi
Summary 275I. Introduction 276II. Fundamental Photosynthesis Models 276III. Rubisco and Its Sequentially Ordered Reaction 279
x

IV. Rubisco in Steady State: Biochemical Models 281V. Experimental Evaluation of the Models 287VI. Concluding Remarks 290Acknowledgments 291References 291
13 Leaf C3 Photosynthesis in silico:Integrated Carbon/Nitrogen Metabolism 295–322
Agu Laisk, Hillar Eichelmann and Vello Oja
Summary 295I. Introduction 296II. The Structure of the Model 297III. Mathematics 302IV. Simulations 312V. Concluding Remarks 317Acknowledgments 319References 319
14 Leaf C4 Photosynthesis in silico:The CO2 Concentrating Mechanism 323–348
Agu Laisk and Gerald Edwards
Summary 324I. Introduction 324II. Principles of NADP-ME Type C4 Photosynthesis 325III. The C4 Model 329IV. Simulations 334V. Knowns and Unknowns in Photosynthesis 338Acknowledgments 345References 345
15 Flux Control Analysis of the Rate of PhotosyntheticCO2 Assimilation 349–360
Ian E. Woodrow
Summary 349I. Introduction 350II. Flux Control Coefficients: Theory and Challenges 352III. Reversible Reactions Can Be Flux Limiting 352IV. Small Control Coefficients Are Hard to Detect 354V. Enzymes with Higher Control Coefficients 356VI. Photosynthetic Electron Transport 358VII. Concluding Remarks 358Acknowledgments 359References 359
xi

Part V: From Leaves to Canopies to the Globe
16 Packing the Photosynthetic Machinery: From Leaf to Canopy 363–399Ülo Niinemets and Niels P.R. Anten
Summary 364I. Introduction 364II. Inherent Differences in Microenvironment and Photosynthetic Potentials
Within the Canopy 367III. Scaling Photosynthesis from Leaves to Canopy 379IV. Concluding Remarks 389Acknowledgments 389References 390
17 Can Increase in Rubisco Specificity Increase Carbon Gainby Whole Canopy? A Modeling Analysis 401–416
Xin-Guang Zhu and Stephen P. Long
Summary 401I. Introduction 402II. Theory and Model Description 404III. The Impact of the Inverse Relationship on Leaf and Canopy
Level Photosynthesis 407IV. Current Efforts of Engineering Rubisco for Higher Photosynthesis 410V. Why Has Evolution Failed to Select the Optimal Rubisco? 412VI. Concluding Remarks 413Acknowledgments 413References 413
18 Role of Photosynthetic Induction for Daily and Annual CarbonGains of Leaves and Plant Canopies 417–440
Manfred Küppers and Michael Pfiz
Summary 417I. Introduction 418II. Representation of Plant Architecture by Digital Reconstruction 418III. The Dynamic Light Environment 419IV. Models of Dynamic Photosynthesis 427V. Calculation of Crown Carbon Acquisition 429VI. Annual Carbon Gains from Steady-state and Dynamic
Photosynthesis Simulations 431VII. Concluding Remarks 436Acknowledgments 436References 436
xii

19 Photosynthesis Within Large-Scale Ecosystem Models 441–464Stephan A. Pietsch and Hubert Hasenauer
Summary 441I. Introduction 442II. Biogeochemical Cycles 443III. Models of Biogeochemical Cycles 446IV. Model Application 452V. Examples of Model Application 456VI. Concluding Remarks 461Acknowledgments 462References 462
20 Photosynthesis in Global-Scale Models 465–497Andrew D. Friend, Richard J. Geider, Michael J. Behrenfeldand Christopher J. Still
Summary 466I. Introduction 467II. Description of Model Approaches 469III. Global Simulation 480IV. Concluding Remarks 486Acknowledgments 488References 488
xiii
Index 499

Preface
Scientific perception of nature relies on a pro-cess of transforming data to information, and theninformation into understanding. Data consist ofobservations and measurements and informationis data organized according to some ontology,i.e. some set of assumptions about what entitiesexist and how they should be classified. Under-standing is a model in the investigator’s mind thatdescribes how the entities relate to each other,a model created in the investigator’s mind as aresult of thinking. Thinking is thus a kind of self-programing of the brain, as a result of whichunderstanding is achieved. When it “runs” in ourbrains, it allows us to predict the behavior ofnatural objects, e.g. in their temporal and spatialaspects. For communication within the scientificcommunity, we first share new data, but thenshare the rigorous forms of the models, whichmay be verbal, graphic, or at their best, math-ematical constructions, reflecting essential fea-tures of a natural system. The latter way of pre-sentation of our understanding of photosynthesisis the subject of this book. In many chapters, themodels are represented by differential equationsthat can reproduce the dynamics of the naturalsystem, or in form of linear equations that definesteady state fluxes or stoichiometries of such asystem. A good model can not only reproducealready measured data about the behavior of theinvestigated system, but it can also predict resultsfor future experiments.
By definition, models are approximations ofnature that are by no means capable of capturingall aspects of the investigated system, no matterhow powerful computers we may have used for it.In the early days of photosynthesis research, mod-els were ingenious by their capacity to explaina prominent feature of the investigated process,such as, for example, the photochemical quench-ing of chlorophyll fluorescence. The early modelswere frequently relatively simple, not requiringa complex code or ontology. The closing of thereaction centers of Photosystem II during chloro-phyll fluorescence induction was well describedby Louis N. M. Duysens assuming a single com-ponent – the quencher Q. With increasing experi-mental accuracy and increasing complexity of the
experimental protocols, this simple model was, interms of Karl Popper’s logic of scientific discov-ery, falsified or, in other words, its validity limitswere found. The simple ‘Quencher ‘Q’ model’of Duysens fell short, for example, in explain-ing the periodicity of four that occurs in chloro-phyll fluorescence emission with multiple singleturnover flashes, or in explaining the sigmoidalshape of the chlorophyll fluorescence inductioncurve. This and other models are perpetuallyexpanding to explain new data obtained with newexperimental protocols.
Such an expectation of the linear expansionof the models is by itself a simplified model.Sometimes an established, “generally accepted”,feature of the model is replaced by another modi-fication, a novel mechanism that explains alreadyknown data as well as the previously assumedmechanism, but widens and deepens the predic-tive power of the model. Thus, different mod-els can explain similar or related phenomena,but only those are accepted for wider use thatare able to accommodate new experimental dataand more sophisticated protocols. The ‘falsified’older and simpler models are not necessarilyrejected and forgotten. Much more often theycontinue to be used with reservation about theirrange of applicability. For example, one does notneed to consider the participation of pheophytinfor the understanding of simple chlorophyll fluo-rescence induction curve on the time scale of sec-onds. In the area of whole photosynthetic processthat specifically includes carbon fixation, GrahamFaquhar, Susanne von Caemmerer and JosephBerry have elegantly approximated it with twoenzymatic reactions only. These ontogeneticallyolder (in the sense of model development) modelsare typically easier to solve and can be obtainedfrom the newer models by mathematically rigor-ous or empirical dimensionality reduction.
Photosynthesis is a complex process spanningfrom femtoseconds to days to seasons to centuriesin time domain and from atoms to the globalbiosphere in spatial domain. No single model candescribe photosynthesis in its full complexity andeven approaching such an elusive goal would notbe practical because such a mathematical model
xv

would not be solvable, being as complex in itsstructure as nature itself. Rather, the process canbe described in a mosaic of models such as theones offered in this book. With increasing com-plexity of the models, we suggest that the read-ers consult the first two chapters of this bookfor a standardization of the model description,so that models become more than abstractionsof individual modelers that are hard to share,merge or even compare with each other. Weexpect this book to be a beginning for creatinga comprehensive modeling space of photosyn-thetic processes that would facilitate an ongoing‘falsification-upgrade’ modeling spiral and wouldallow mergers between related model lines. Theindividual model areas represented here beginwith the absorption of a photon and include elec-tron transport, carbon assimilation, and productsynthesis. With all these molecular models athand one can upscale to cell, organ, plant, canopy,and eventually to global biosphere. Chapters pre-sented in this book show how different levels ofbiological hierarchy overlay and interact in theamazing process of photosynthesis.
Photosynthesis in silico is a unique book inits integrated approach to the understanding ofphotosynthesis processes from light absorptionand excitation energy transfer to global aspects ofphotosynthetic productivity – all interconnectedby the use of mathematical modeling. The bookis written by 44 international authorities from 15countries. Chapters in this book are presentedin a review style with emphasis on the latestbreakthroughs. Instead of providing mathemati-cal details, only the key equations, the basis forthe novel conclusions, are provided, with refer-ences to the original work at the end of eachchapter. Thus, de facto this is not a mathemat-ical book of equations, but dominantly verbaldiscussion showing why the quantitative logic ofmathematics has been so efficient for understand-ing the subject. Yet, in order to exploit the fullpotential of the book, we hope the models willeventually be translated to the universal formatof the Systems Biology Mark-Up Language andmade accessible also in their full mathematicalform on the internet. As argued in Chapter 1,
we stand here at the beginning of a qualitativelynew scientific collaboration with its dynamicslargely dependent on willingness of our researchcommunity to share resources to generate a free-access model database of photosynthesis. Suchan endeavor is fully justified by an increasinglyrecognized role of photosynthesis in nature andlately also as an important alternative for tech-nological solutions of currently surging energyneeds of the humankind.
We thank our families and coworkers in ourlaboratories for their patience with us, and fortheir support during the preparation of this book.We also thank Noeline Gibson, Jacco Flipsen andAndré Tournois, of Springer, for their friendlyand valuable guidance during the typesetting andprinting of this book.
Agu LaiskInstitute of Molecular and Cell Biology
University of Tartu, Riia 23, Tartu 51010Estonia
Telephone: 372 736 6021Fax: 372 742 0286
e-mail: [email protected]
Ladislav NedbalDepartment of Biological Dynamics
Institute of Systems Biology and Ecology CASZámek 136, 37333 Nové Hrady
Czech Republic
Telephone/Fax: 420 386 361 231e-mail: [email protected]
GovindjeeDepartment of Plant Biology
University of Illinois at Urbana-Champaign265 Morrill Hall, 505 South Goodwin Avenue
Urbana, IL 61801-3707, USA
Telephone: 1 217 337 0627Fax: 1 217 244 7246
e-mail: [email protected]
xvi

The Editors
Agu Laisk, born in 1938, obtained BS and MSdegrees in Physics at the University of Tartu(Estonia) in 1961. He then joined the researchgroup of Juhan Ross to study the penetrationof sunlight into plant canopies for the purposeof modeling of plant productivity. His “candidateof science” work (equivalent to Ph.D., in the for-mer Soviet Union), on the ‘statistical distributionof light in the canopy’, was completed in 1966.Since then he became interested in mechanismsthat determine the rate of photosynthesis of aleaf. Together with his former student Vello Oja,he observed that in photosynthesis O2 competeswith CO2 for one and the same acceptor and in1970 published a mathematical model of pho-tosynthesis and photorespiration, based on thecompetition of CO2 and O2 for ribulosebispho-sphate (RuBP). Then he observed that at highCO2 concentrations, O2 enhances photosynthe-sis, showing the importance of the Mehler reac-tion. Soon thereafter, sophisticated experimentson “flashing” a leaf with short pulses of CO2showed that photosynthesis is limited by Rubiscoat low CO2, but by RuBP regeneration at highCO2 levels. For these findings, the degree of Doc-tor of Science in Biology was awarded to him in1976 by the Timiryazev Institute of Plant Phys-iology in Moscow (published as a monograph“Kinetics of Photosynthesis and Photorespirationin C3 Plants” by “Nauka”, Moscow, 1977). The
specific approach of Laisk’s group is in usingonly intact leaves as objects for measurements.This requires original equipment to be built inthe laboratory – now appreciated in several otherlaboratories and, in principle, described in a book(together with Vello Oja) “Dynamics of Leaf Pho-tosynthesis. Rapid-Response Measurements andtheir Interpretations”, edited by Barry Osmond(CSIRO, Australia, 1998). A recent unexpectedresult from Laisk’s laboratory is that cyclic elec-tron transport around Photosystem I is muchfaster than necessary to cover the possible deficitin ATP synthesis – indicating that cyclic elec-tron flow may be largely uncoupled from pro-ton translocation or there must be a controllableproton leak. The interpretation of such kineticexperiments is unthinkable without the applica-tion of mathematical modeling. Agu Laisk is aFellow Member of Estonian Academy of Sci-ence, life-time corresponding member of TheAmerican Society of Plant Biologists (ASPB), amember of the editorial board of PhotosynthesisResearch and of Photosynthetica. He has receivedNational Science Awards from the Estonian Gov-ernment. His international collaborators, whohave deeply influenced his views, include: UlrichHeber (Germany), David Walker (UK), BarryOsmond (Australia), Gerry Edwards (USA) andRichard Peterson (USA). At Tartu University, heteaches Bioenergetics.
xvii

Ladislav Nedbal, born in 1955, studied Bio-physics at the Faculty of Mathematics andPhysics, Charles University in Praha, CzechRepublic. He graduated in 1981 with a thesison the ‘theory of the excitonic energy trans-fer in molecular crystals’. He learned about thefascinating process of photosynthesis from IvanŠetlík, who is one of the founders of algalbiotechnology. He moved over from doing model-ing of energy transfer to research in experimentalphotosynthesis in the early years of his scientificcareer; this led to his present interest in modelingphotosynthesis. Yet, preceding the present déjà vuwith mathematical models were many more yearsof apprenticeship in experimental science thatwere marked with discreet advice from Govind-jee. It was the present Series Editor of Advancesin Photosynthesis and Respiration, who taughthim the principles of technical writing in the late1980s and introduced him, in 1990, to John Whit-marsh of the University of Illinois at Urbana-Champaign, USA. It was in John‘s lab whereNedbal discovered the photoprotective role ofcytochrome b559. His other important tutors wereTjeerd Schaafsma in Wageningen, The Nether-lands, and Anne-Lise Etienne in France. A sig-nificant inspiration came from David Kramer dur-ing the postdoc years in Urbana-Champaign andin Paris, where they collaborated in constructing
a modulated light spectrophotometer and fluo-rometer. In that project he met Martin Trtilek,with whom he founded Photon Systems Instru-ments (PSI), a small company that has createda number of innovative instruments for photo-synthesis research. The most important achieve-ment was the development of the first commercialPulse Amplitude Modulating (PAM) type imag-ing fluorometer – FluorCam. Recently, collabo-ration with Martin resulted in the construction of‘intelligent’ photobioreactor for the cultivation ofalgae and cyanobacteria. The instrument collects,in real time, detailed information on the culture’sphotochemical yields and on its growth dynam-ics. The combination of mathematical modelingwith experimental research in photosynthesis andengineering approaches logically led to anotherdéjà vu in his career, this time with algal biotech-nology. In this area, mathematical models bringto light yet unexplored pathways towards com-mercially viable use of algae and cyanobacte-ria. Further stimulating his interest in modelsare the mysterious dynamic features that he andhis co-workers, including both his co-Editors ofthe present volume Agu Laisk and Govindjee,recently discovered in harmonically modulatedlight. Understanding plant behavior in dynamiclight remains a major challenge that will be tack-led by the current book Photosynthesis in silico.
xviii

Govindjee, born in 1932, obtained his B.Sc.(Chemistry, Biology) and M.Sc. (Botany) in 1952and 1954, from the University of Allahabad,India, and his Ph.D. (Biophysics) in 1960, fromthe University of Illinois at Urbana-Champaign(UIUC), IL, USA. His mentors were RobertEmerson and Eugene Rabinowitch. He is bestknown for his research on the excitation energytransfer, light emission, the primary photochem-istry and the electron transfer in PhotosystemII (PS II). His research, with many collabo-rators, has included the discovery of a short-wavelength form of chlorophyll (Chl) a func-tioning in the Chl b-containing system of PS II;of the two-light effects in Chl a fluorescenceand in NADP reduction in chloroplasts (EmersonEnhancement); the basic relationships betweenChl a fluorescence and photosynthetic reactions;the unique role of bicarbonate at the acceptorside of PS II. He provided the theory of thermo-luminescence in plants, made the first picosec-ond measurement on the primary photochem-istry of PS II and used Fluorescence LifetimeImaging Microscopy (FLIM) of Chl a fluores-cence in understanding photoprotection againstexcess light. His current focus is on the historyof photosynthesis research, photosynthesis edu-cation, and possible existence of extraterrestriallife. He has served on the faculty of UIUC forabout 40 years. Since 1999 he has been ProfessorEmeritus of Biochemistry, Biophysics and PlantBiology at the same institution. He is coauthorof ‘Photosynthesis’ (with E. Rabinowitch; JohnWiley, 1969), and editor of several books includ-ing Bioenergetics of Photosynthesis (Academic
Press, 1975); Photosynthesis, Volumes I and II(Academic Press, 1982); Light Emission of Plantsand Bacteria (with J. Amesz and D.C. Fork;Academic Press, 1986); Chlorophyll a Fluores-cence: A Signature of Photosynthesis (with G.C.Papageorgiou, Springer, 2004); and Discoveriesin Photosynthesis (with J.T. Beatty, H. Gest andJ.F. Allen; Springer, 2005). His honors include:Fellow of the American Association of Advance-ment of Science; Distinguished Lecturer of theSchool of Life Sciences, UIUC; Fellow and Life-time member of the National Academy of Sci-ences (India); President of the American Societyfor Photobiology (1980–1981); Fulbright Scholarand Fulbright Senior Lecturer; Honorary Pres-ident of the 2004 International PhotosynthesisCongress (Montréal, Canada); the 2006 Recipi-ent of the Lifetime Achievement Award from theRebeiz Foundation for Basic Biology; the 2007Recipient of the Communication Award of theInternational Society of Photosynthesis Research(ISPR); and the 2008 Liberal Arts and SciencesAlumni Achievement Award of the University ofIllinois. During 2007, Photosynthesis Researchcelebrated Govindjee’s 50 years in Photosynthe-sis, and his 75th birthday through a two-Partspecial volume of the journal (Julian Eaton-Rye,editor). To celebrate his life-long achievement inPhotosynthesis Research, Education, and its His-tory, University of Indore, India, recently helda 3-day International Symposium (Nov. 27–29,2008) on ‘Photosynthesis in Global Perspective’(K.N. Guruprasad, Convener).
Govindjee has trained more than 20 Ph.D. stu-dents and about 10 postdoctoral associates.
xix

Author Index
Anten, Niels P.R. 363–399
Behrenfeld, Michael J. 465–497Bernacchi, Carl. J. 231–246Berry, Joseph 209–230
Cervený, Jan 17–29
Dekker, Jan P. 33–53
Edwards, Gerald 323–348Eichelmann, Hillar 295–322
Farazdaghi, Hadi 275–294Farquhar, Graham 209–230, 231–246Freiberg, Arvi 55–82Friend, Andrew D. 465–497
Geider, Richard J. 465–497
Harbinson, Jeremy 247–273Hasenauer, Hubert 441–464Hucka, Michael 3–15
Küppers, Manfred 417–440
Laisk, Agu 295–322, 323–348Lavergne, Jérôme 177–205Lazár, Dušan 85–123Long, Stephen P. 231–246, 401–416
Nedbal, Ladislav 17–29Niinemets, Ülo 363–399Novoderezhkin, Vladimir I. 33–53
Oja, Vello 295–322
Pfiz, Michael 417–440Pietsch, Stephan A. 441–464Pimentel, Carlos 231–246Prášil, Ondrej 125–149
Riznichenko, Galina 151–176Rosenthal, David M. 231–246Rubin, Andrew 151–176
Schaff, James 3–15Schansker, Gert 85–123Schmidt, Henning 17–29Still, Chrisostopher J. 465–497Struik, Paul C. 247–273
Trinkunas, Gediminas 55–82
Van Grondelle, Rienk 33–53Von Caemmerer, Susanne 209–230Vredenberg, Wim 125–149
Woodrow, Ian E. 349–360
Yin, Xinyou 247–273
Zhu, Xin-Guang 401–416
xxi
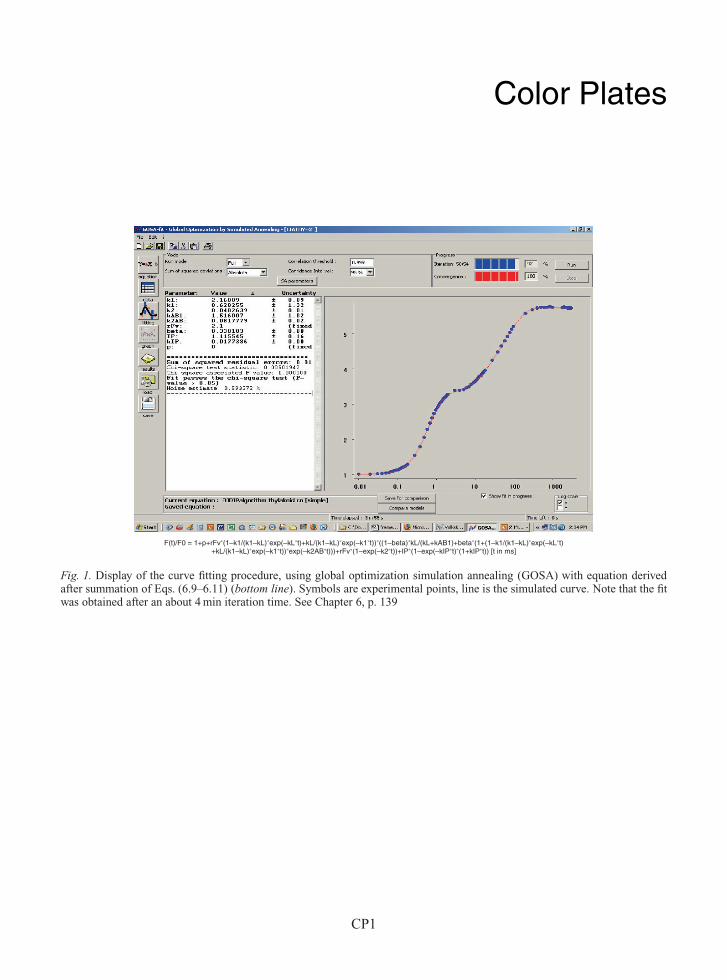
Color Plates
F(t)/F0 = 1+p+rFv∗(1–k1/(k1–kL)∗exp(–kL∗t)+kL/(k1–kL)∗exp(–k1∗t))∗((1–beta)∗kL/(kL+kAB1)+beta∗(1+(1–k1/(k1–kL)∗exp(–kL∗t)+kL/(k1–kL)∗exp(–k1∗t))∗exp(–k2AB∗t)))+rFv∗(1–exp(–k2∗t))+IP∗(1–exp(–kIP∗t)∗(1+kIP∗t)) [t in ms]
Fig. 1. Display of the curve fitting procedure, using global optimization simulation annealing (GOSA) with equation derivedafter summation of Eqs. (6.9–6.11) (bottom line). Symbols are experimental points, line is the simulated curve. Note that the fitwas obtained after an about 4 min iteration time. See Chapter 6, p. 139
CP1

Ochroma lagopus
March 1996 October 1996
October 1996
March 1997
1 m
March 1997March 1996
Time of investigation
Tree height [m]
Total leaf area [m2] (gain/ loss)
Total leaf biomass [g] (gain/ loss)
Supporting tissue [g] (gain/ loss)
Aboveground biomass [g] (gain/ loss)
Total carbon gain
Carbon allocation to supporting tissue
Carbon allocation to leaves
Resulting carbon allocation to roots
2.79
2.21
185.5
374
560
via abovegroundbiomass monitoring
3561 g
1359 g
2202 g
4.54
6.39 (9.46/5.28)
537 (794/443)
1267 (1075/182)
1804 (1869/625)
380 d
via ‘steady-state’ model
6393 g
35%
21%
2832 g (44%) 975 g (21%)
4.89
3.56 (6.70/9.54)
299 (564/802)
2028 (1127/366)
2327 (1691/1168)
4536 g
30%
49%
via ‘dynamic’ model
a
b
c
d
Fig. 2. Development of an individual of the shade-intolerant pioneer Ochroma lagopus from an open site and deduction of itsannual carbon allocation. Light green leaf area: sun exposed, dark green: (self-)shaded (from Timm et al., 2004). (a) Above-ground architectural development as reconstructed via the method described in Fig. 18.1; (b) change in the individual’s lightenvironment as indicated by hemispherical photography immediately above its uppermost leaves; (c) growth and biomassparameters of the respective individual; (d) deduction of annual assimilate flux balances (carbon allocation) as percentageof total annual crown carbon gain, either via a steady-state or a dynamic photosynthesis model. Carbon gain and allocation ofbiomass are given in equivalents of dry matter (CH2O)n. See Chapter 18, p. 423
CP2

Billia colombiana
March 1996 October 1996
October 1996
March 1997
1 m
March 1997March 1996
Time of investigation
Tree height [m]
Total leaf area [m2] (gain/ loss)
Total leaf biomass [g] (gain/ loss)
Supporting tissue [g] (gain/ loss)
Aboveground biomass [g] (gain/ loss)
Total carbon gain
Carbon allocation to supporting tissue
Carbon allocation to leaves
Resulting carbon allocation to roots
2.1
0.42
27.6
58.7
86.3
via abovegroundbiomass monitoring
64.9 g
19.1 g
45.8 g
2.26
0.581 (0.204/0.045)
38 (13.3/2.9)
90.0 (32.1/0.76)
128.0 (45.5/3.7)
380 d
via ‘steady-state’ model
194.1 g
24%
10%
129.2 g (66%) 16.9 g (21%)
2.45
0.632 (0.089/0.038)
41.3 (5.8/2.5)
103.0 (13.7/0.68)
144.3 (19.5/3.2)
81.8 g
23%
56%
via ‘dynamic’ model
a
b
c
d
Fig. 3. The same as Fig. 18.4, but for an individual of the mid- to late-successional shade-tolerant Billia colombiana below aclosed canopy. Red leaf area: newly developed (from Timm et al., 2004). See Chapter 18, p. 424
CP3

1 m
71–85
25–39
>67>63>59>55>51>47>43
>39
Daily carbon gain mmol m–2 d– –1
Fig. 4. Daily carbon balance of each individual leaf in the crown of a Salacia petenensis plant. Crown carbon gain wasdetermined by summing up the individual balances. In the mean over 380 days carbon gain amounted to 426 mg day−1 (fromTimm et al., 2004). See Chapter 18, p. 432
CP4

Fig. 5. Modeled pools and fluxes for the virgin forest reserve Rothwald using model parameters for Common beech forests.Upper graph: Comparison of the temporal development of modeled soil, necromass, stem and total ecosystem carbon content for600 simulation years at landscape level steady state. Lower graph: Corresponding annual C fluxes from heterotrophic respiration(Rh), net primary production (NPP) and net ecosystem exchange (NEE). I – optimum phase; II – breakdown/regeneration phase;III – juvenescence (Pietsch and Hasenauer, 2006). See Chapter 19, p. 457
Fig. 6. Attractor of modeled NEE for the successional cycle evident within the virgin forest reserve Rothwald. a: NEE-Attractorfor the virgin forest successional cycle reconstructed from model results using site and climate conditions of 18 research plots.b: Attractor reconstructed for one single plot and one successional cycle. Arrows indicate the trends of model behavior duringdifferent phases of the successional cycle. (S.A. Pietsch, unpublished) See Chapter 19, p. 460
CP5

Net Primary Productivity (kg Cm−2 yr−1)
0 0.03 0.1 0.3 0.5 0.8 1.0 1.5
Fig. 7. Mean annual net primary productivity (NPP) simulated by Hybrid6.5 (land) and the CbPM (ocean) for the period 2000–2007. Total mean annual NPP is 107.3 Pg C year−1, with 51.1% coming from land and 48.9% from the oceans. Land pixelssimulated with 1/4
◦ resolution and ocean pixels with 1/12◦ resolution. Land leaf area dynamics prescribed from MODIS satellite
retrievals, ocean production calculated using data from the SeaWiFS instrument. Full simulation details are given in the text.See Chapter 20, p. 486
CP6

Part I
General Problems of Biological Modeling

Chapter 1
Trends and Tools for Modeling in Modern Biology
Michael Hucka∗Control and Dynamical Systems, California Institute of Technology, Pasadena, CA 91125,
USA
James SchaffRichard D. Berlin Center for Cell Analysis and Modeling, University of Connecticut Health
Center, Farmington, CT 06030, USA
Summary...............................................................................................................................................................................................3I. Introduction...................................................................................................................................................................................4II. Representing Model Structure and Mathematics................................................................................................................4III. Augmenting Models with Semantic Annotations.................................................................................................................6
A. Systems Biology Ontology (SBO)...........................................................................................................................6B. Minimum Information Requested in the Annotation of Biochemical Models (MIRIAM)..............................7
IV. Connecting Models to Results.................................................................................................................................................9A. Common Experimental and Modeling Activities..................................................................................................9B. Supporting Modeling Activities Through Software Environments.................................................................10
V. Future Directions for Systems Biology Markup Language (SBML)...............................................................................13VI. Conclusions................................................................................................................................................................................13References.........................................................................................................................................................................................14
Summary
Computational modeling in biology requires sophisticated software tools. Precise communication andeffective sharing of the models developed by researchers requires standard formats for storing, annotat-ing, and exchanging models between software systems. Developing such standards is the driving visionbehind the Systems Biology Markup Language (SBML) and several related efforts that we discuss inthis chapter. At the same time, such standards are only enablers and ideally should be hidden “under thehood” of modeling environments that provide users with high-level, flexible facilities for working withcomputational models. As an example of the modern software systems available today, we discuss theVirtual Cell and illustrate its support for typical modeling activities in biology.
∗ Author for correspondence, e-mail: [email protected]
A. Laisk, L. Nedbal and Govindjee (eds.), Photosynthesis in silico: Understanding Complexity from Molecules to Ecosystems, pp. 3–15.c© 2009 Springer Science+Business Media B.V.

4 Michael Hucka and James Schaff
I. Introduction
Understanding the dynamic processes that arethe essence of a living cell stands as one of themost important and most difficult challenges oftwenty-first century biology. Today, it is widelyappreciated that we can only hope to meet thatchallenge through the development and applica-tion of computational methods (Hartwell et al.,1999; Fraser and Harland, 2000; Arkin, 2001;Tyson et al., 2001; Noble, 2002; Alm and Arkin,2003; Zerhouni, 2003), particularly the creationof mechanistic, explanatory models illuminat-ing the functional implications of the data uponwhich they are built.
Models are not substitutes for experiments anddata; rather, they are faithful teammates in theprocess of scientific discovery. A realistic com-putational model represents a modeler’s dynamicunderstanding of the structure and function ofpart of a biological system. As the number ofresearchers constructing realistic models contin-ues to grow, and as the models become evermore sophisticated, they collectively representa significant accumulation of knowledge aboutthe structural and functional organization of thesystem. Moreover, using them, the assimilationof new hypotheses and data can be done in amore systematic way because the additions mustbe fitted into a common, consistent framework.Once properly constructed, the models becomea dynamic representation of our current state ofunderstanding of a system in a form that can facil-itate communication between researchers andhelp to direct further experimental investigations(Bower and Bolouri, 2001).
Today’s models are large (and growing everlarger) and complex (and getting ever morecomplex). We are now long past the point ofbeing able to communicate and exchange real-world models effectively by simply summariz-
Abbreviations: DOI – digital object identifier; MIASE –minimum information about a simulation experiment;MIRIAM – minimum information requested in the anno-tation of biochemical models; SBGN – systems biologygraphical notation; SBML – systems biology markup lan-guage; SBO – systems biology ontology; SSA – stochasticsimulation algorithm; UML – unified modeling language;URN – uniform resource name; VCell – virtual cell; XML –eXtensible markup language
ing them in written narratives featuring a fewequations. The precise communication of com-putational models between humans and betweensoftware is critical to being able to realize mod-eling’s promise. Achieving this requires standard-izing the electronic format for representing com-putational models in a way independent of anyparticular software – after all, different researchgoals are often best served by different softwaretools, yet modelers still need to share their resultswith their colleagues. At the same time, today’sresearchers need powerful software environmentsthat offer a range of capabilities to support thecreation, analysis, storage and communication ofmodels, all the while hiding the details of themodel representation format and providing bio-logical modelers with high-level user interfacesand capabilities matched to the tasks they needto do.
In this chapter, we discuss both standards andsoftware for computational modeling in biology.We summarize the de facto standard format, theSystems Biology Markup Language (SBML), aswell as ongoing related efforts to standardizethe representation of model annotations throughMIRIAM (the Minimum Information RequestedIn the Annotation of biochemical Models) andSBO (the Systems Biology Ontology). As criticalas they are, however, such standards are in theend only enablers; they are (hopefully) not whatusers interact with directly. We therefore also dis-cuss software systems, focusing on one in partic-ular, the Virtual Cell, as a way to present typi-cal modeling activities in the context of one oftoday’s most full-featured, interactive modelingenvironments. The advanced capabilities of sys-tems such as Virtual Cell also help drive furtherdevelopment of SBML and adjunct efforts, andso we close with a summary of present work toextend SBML as well as standardize other areasof modeling and simulation exchange, such as thedescription of simulations.
II. Representing Model Structureand Mathematics
Until the late 1980s, publication of a compu-tational model almost universally involved pub-lishing only the equations and parameter values,usually with some narrative descriptions of how

1 Modeling in Modern Biology 5
the model was coded in software and how it wassimulated and analyzed. The systems of equa-tions were, with few exceptions, directly imple-mented in software: in a very direct sense, theprogram was the model. Authors sometimes evenwrote their own numerical integration code. Thisgeneral approach was necessary because of theprimitive state of computational platforms andelectronic data exchange, and it was fraught withproblems. The most significant problem is sim-ply the opportunities for errors that arise when amodel must be recapitulated by humans into andback out of natural language form. The degree towhich this is a real problem is startling. Cura-tors for databases of published models such asBioModels Database (Le Novère et al., 2006) andJWS Online (Snoep and Olivier, 2003; Olivierand Snoep, 2004), report by personal commu-nication that when they first began operation inthe 2000–2004 timeframe, over 95% of publishedmodels they encountered had something wrongwith them, ranging from typographical errors tomissing information (even today, the problem rateis greater than 60%). A second problem is that,when a model is inextricably intertwined with itssoftware implementation, it is difficult to examineand understand the precise details of the actualmodel (rather than artifacts of its particular real-ization in software). A third problem is that hav-ing to reconstruct a model from a paper is anextremely tall hurdle to fast, efficient and error-free reuse of research results.
Some areas of biological modeling improvedon this situation in the 1990s. The field of compu-tational neuroscience was particularly advancedin this regard, having two freely-available sim-ulation packages, GENESIS (Bower and Bee-man, 1995; Bower et al., 2002) and NEURON(Hines and Carnevale, 1997), supported on avariety of operating systems. These simulationplatforms made it possible for modelers to dis-tribute abstract definitions of their models andsimulation procedures in the form of scripts thatcould be interpreted automatically by the plat-form software. The approach vastly improved thereusability of models. However, there remainedthe limitation that the formats were specific tothe simulation package in which they were devel-oped. Whoever wanted to reuse the models hadto run the same software in order to reuse themodel (assuming they were able to get the nec-
essary files from the model’s authors – electronicpublishing of models as supplements to journalarticles was still rare).
With the surge of interest in computationalsystems biology at the beginning of this cen-tury, software tools evolved one step further withthe creation of application-independent modeldescription formats such as CellML (Hedleyet al., 2001) and SBML (Hucka et al., 2003,2004). This form of representation is not an algo-rithm or a simulation script; it is a declarativedescription of the model structure that is theninterpreted and translated by each individual soft-ware system into whatever internal format it actu-ally uses. No longer tied to a particular softwaresystem, such software-independent formats per-mit a wider variety of experimentation in algo-rithms, user interfaces, services, and many otheraspects of software tool development, by virtue ofallowing multiple software authors to explore dif-ferent facilities that all use the same input/outputrepresentation. In addition, and even more signif-icantly, it enables practical publication of modelsin public databases.
The Systems Biology Markup Language(SBML; http://sbml.org) has become the de factostandard for this purpose, supported by over120 software systems at the time of this writ-ing. SBML is a machine-readable lingua francadefined neutrally with respect to software toolsand programming languages. It is a model def-inition language intended for use by software –humans are not intended to read and write SBMLdirectly. By supporting SBML as an input andoutput format, different software tools can alloperate on the identical representation of a model,removing opportunities for errors in translationand assuring a common starting point for anal-yses and simulations. SBML is defined using asubset of UML, the Unified Modeling Language(Booch et al., 2000), and in turn, this is usedto define how SBML is expressed in XML, theeXtensible Markup Language (Bray et al., 1998).Software developers can make use of a numberof resources for incorporating SBML support intheir applications (Bornstein et al., 2008).
SBML can encode models consisting of bio-chemical entities (species) linked by reactions toform biochemical networks. An important prin-ciple in SBML is that models are decomposedinto explicitly-labeled constituent elements, the

6 Michael Hucka and James Schaff
set of which resembles a verbose rendition ofchemical reaction equations; the representationdeliberately does not cast the model directly into aset of differential equations or other specific inter-pretation of the model. This explicit, modeling-framework-agnostic decomposition makes it eas-ier for a software tool to interpret the model andtranslate the SBML form into whatever internalform the tool actually uses. The main constructsprovided in SBML include the following:
Compartment and compartment type: a com-partment is a container for well-stirred substanceswhere reactions take place, while a compartmenttype is an SBML construct allowing compart-ments with similar characteristics to be classifiedtogether.
Species and species type: a species in SBMLis a pool of a chemical substance located in aspecific compartment, while species types allowpools of identical kinds of species located in sep-arate compartments to be classified together.
Reaction: a statement describing some trans-formation, transport or binding process that canchange one or more species (each reaction ischaracterized by the stoichiometry of its productsand reactants and optionally by a rate equation).
Parameter: a quantity that has a symbolicname.
Unit definition: a name for a unit used in theexpression of quantities in a model.
Rule: a mathematical expression that is addedto the model equations constructed from the setof reactions (rules can be used to set parame-ter values, establish constraints between quanti-ties, etc.).
Function: a named mathematical function thatcan be used in place of repeated expressions inrate equations and other formulae.
Event: a set of mathematical formulae evalu-ated at a specified moment in the time evolutionof the system.
The simple formalisms in SBML allow a widerange of biological phenomena to be modeled,including cell signaling, metabolism, gene regu-lation, and more. Significant flexibility and powercomes from the ability to define arbitrary formu-lae for the rates of change of variables as well asthe ability to express other constraints mathemat-ically.
SBML is being developed in “levels”. Eachhigher level adds richness to the model defini-
tions that can be represented by the language. Bydelimiting sets of features at incremental stages,the SBML development process provides soft-ware authors with stable standards and the com-munity can gain experience with the languagedefinitions before new features are introduced.Two levels have been defined so far, named(appropriately enough) Level 1 and Level 2. Theformer is simpler (but less powerful) than Level2. The separate levels are intended to coexist;SBML Level 2 does not render Level 1 obsolete.Software tools that do not need or cannot supporthigher levels can go on using lower levels; toolsthat can read higher levels are assured of alsobeing able to interpret models defined in the lowerlevels. Open-source libraries such as libSBML(Bornstein et al., 2008) allow developers to sup-port both Levels 1 and 2 in their software with aminimum amount of effort.
III. Augmenting Models with SemanticAnnotations
The ability to have meaningful exchange ofcomplex mathematical models of biological phe-nomena turns out to require a deeper levelof semantic encoding and knowledge manage-ment than is embodied by a format such asSBML, which encompasses only syntax anda limited level of semantics. This realizationcame early in the context of CellML, whosedevelopers added a standard scheme for meta-data annotations soon after CellML was devel-oped (Lloyd et al., 2004). CellML’s metadatascheme was adopted by SBML at the begin-ning of the development of SBML Level 2,but limitations with the scheme later led theSBML community to seek alternatives. Thesewere found in the form of the Systems BiologyOntology (SBO; http://www.ebi.ac.uk/SBO; LeNovère et al., 2006), and the Minimum Informa-tion Requested in the Annotation of BiochemicalModels (MIRIAM; Le Novère et al., 2005).
A. Systems Biology Ontology (SBO)
The rationale for SBO is to provide controlledvocabularies for terms that can be used to anno-tate components of a model in SBML (or indeed,any other formal model representation format).

1 Modeling in Modern Biology 7
It requires no change to the form of the basicmodel in SBML; rather, it provides the option toaugment the basic model with machine-readablelabels that can be used by software systems to rec-ognize more of the semantics of the model. SBOprovides terms for identifying common reactionrate expressions, common participant types androles in reactions, common parameter types andtheir roles in rate expressions, common model-ing frameworks (e.g., “continuous”, “discrete”,etc.), and common types of species and reactions.Recent versions of SBML Level 2 provide anoptional attribute on every element where an SBOterm may be attached. Table 1.1 lists the corre-spondences between major components of SBMLand SBO vocabularies.
The relationship implied by the attribute valueon an SBML model component is “is a”: thething defined by that SBML component “is an”instance of the thing defined in SBO by indi-cated SBO term. By adding SBO term referenceson the components of a model, a software toolcan provide additional details using independent,shared vocabularies that can enable other soft-ware tools to recognize precisely what the compo-nent is meant to be. Those tools can then act onthat information. For example, if the SBO iden-tifier SBO:0000049 is assigned to the conceptof “first-order irreversible mass-action kinetics,continuous framework”, and a given reaction ina model has an SBO attribute with this value,then regardless of the identifier and name given to
Table 1.1. Correspondence between major SBML compo-nents and controlled vocabulary branches in the SystemsBiology Ontology (SBO)SBML component SBO vocabularyModel InteractionFunction definition Mathematical expressionCompartment type Material entitySpecies type Material entityCompartment Material entitySpecies Material entityReaction InteractionReaction’s kinetic law Mathematical expression
→ Rate lawParameter Quantitative parameterInitial assignment Mathematical expressionRule Mathematical expressionEvent Interaction
the reaction itself, a software tool could use thisto inform users that the reaction is a first-orderirreversible mass-action reaction.
As a consequence of the structure of SBO, notonly children are versions of the parents, but themathematical expression associated with a childis a version of the mathematical expressions ofthe parents. This enables a software applicationto walk up and down the hierarchy and inferrelationships that can be used to better interpreta model annotated with SBO terms. Simulationtools can check the consistency of a rate lawin an SBML model, convert reactions from onemodeling framework to another (e.g., continuousto discrete), or distinguish between identicalmathematical expressions based on differentassumptions (e.g., Henri-Michaelis-Menten vs.Briggs-Haldane). Other tools like SBMLmerge(Schulz et al., 2006) can use SBO annotations tointegrate individual models into a larger one.
SBO adds a semantic layer to the formalrepresentation of models, resulting in a morecomplete definition of the structure and mean-ing of a model. The presence of an SBOlabel on a compartment, species, or reaction,can also help map SBML elements to equiva-lents in other standards, such as (but not lim-ited to) BioPAX (http://www.biopax.org) or theSystems Biology Graphical Notation (SBGN,http://www.sbgn.org). Such mappings can beused in conversion procedures, or to build inter-faces, with SBO becoming a kind of “glue”between standards of representation.
B. Minimum Information Requestedin the Annotation of BiochemicalModels (MIRIAM)
While SBO annotations help add semantics, thereremains a different kind of impediment to effec-tive sharing and interpretation of computationalmodels. Figure 1.1 illustrates the issue.
When a researcher develops a model, theyoften use simple identifiers for chemical sub-stances, or at best, only one of a multitude of pos-sible synonyms for the substance. The situation iseven worse when it comes to the chemical reac-tion and other processes: these are often givennames such as “R1”, “R2”, etc., or at best, generic