phylogenetic relationships of sparassis infer red fro m nuclear
TRANSCRIPT

1015
Mycologia, 96(5), 2004, pp. 1015–1029.! 2004 by The Mycological Society of America, Lawrence, KS 66044-8897
Phylogenetic relationships of Sparassis inferred from nuclear andmitochondrial ribosomal DNA and RNA polymerase sequences
Zheng Wang1
Manfred BinderDepartment of Biology, Clark University, 950 MainStreet, Worcester, Massachusetts 01610
Yu-Cheng DaiInstitute of Applied Ecology, Chinese Academy ofSciences, Shenyang 110016, China
David S. HibbettDepartment of Biology, Clark University, 950 MainStreet, Worcester, Massachusetts 01610
Abstract: Sparassis species show extensive morpho-logical variation, especially when materials from east-ern Asia and Australia are compared with collectionsfrom North America and Europe. We have beenstudying the taxonomy of Sparassis from eastern Asia,North America, Australia and Europe, using bothmorphological and molecular data. DNA was extract-ed from 32 recent collections of Sparassis from Aus-tralia, Canada, China, Finland, France, Germany, Ja-pan, Switzerland, Thailand, the United Kingdom andthe United States. The report of a Sparassis taxonfrom Australia is the first report of this genus fromthe Southern Hemisphere. Sequences of nuclear andmitochondrial rDNA and the gene encoding RNApolymerase subunit II (RPB2) were used to examinerelationships both within the genus Sparassis and be-tween Sparassis species and other members of thepolyporoid clade. Equally weighted parsimony anal-yses and Bayesian analyses were performed using in-dependent datasets and combined datasets of se-quences from different regions. Our results suggestthat: (i) Polyporoid fungi producing a brown rot mayform a clade; (ii) as suggested in a previous study,Sparassis and Phaeolus form a monophyletic group,which is united by the production of a brown rot, thepresence of a bipolar mating system and the frequenthabit of growing as a root and butt rot on living trees;(iii) at least seven lineages are within Sparassis, rep-resented by S. spathulata, S. brevipes, S. crispa, S. rad-icata and three taxa that have not been described,which can be distinguished on the basis of fruitingbody structure, presence or absence of clamp con-
Accepted for publication March 1, 2004.1 Corresponding author. E-mail: [email protected]
nections, presence or absence of cystidia and sporesize.
Key words: polyporoid clade, multigene phylo-geny, MrBayes, biogeography
INTRODUCTION
Sparassis Fr. species frequently are reported fromnorthern temperate forests. Members of this genushave conspicuous cream white or yellow, large andcauliflower-like basidiocarps, which are found arisingfrom tree roots. Sparassis species are variable in bas-idiocarp shape and color and are associated with dif-ferent plant host taxa in different ecosystems. Tax-onomy of this genus has been based mainly on basid-iocarp macromorphology, basidiospore size and hostplant type (Burdsall and Miller 1988a, b). Twelve spe-cies with three varieties have been recorded in thisgenus, according to the CABI Index of Fungi (http://www.indexfungorum.org/). Three species, Spar-assis brevipes Krombh., S. crispa Wulf. : Fr., and S. spa-thulata. Schw. : Fr., have been accepted in recentstudies (Martin and Gilbertson 1976, Kreisel 1983,Burdsall and Miller 1988a, b, van Zanen 1988). Spar-assis radicata Weir was described on Douglas-fir fromwestern North America, but Martin and Gilbertson(1976) suggested that it should be considered con-specific with S. crispa based on di-mon mating testsand culture studies.
Sparassiella longistipitata Schwarzman is groupedin the family Sparassidaceae Herter (Kirk et al 2001),but no collections of this species are available (Burd-sall and Miller 1988a).
The stalked form of the basidiocarp, highlybranched flabellae with hymenium on both sides ininitial stages, and the monomitic hyphal system makeit hard to position the genus Sparassis. Donk (1964)reviewed earlier studies and, apparently based on ba-sidiocarp morphology, concluded that the Sparassi-daceae could be a tribe in the Clavariaceae or a dis-tinct family close to the Clavariaceae. Later, the Spa-rassidaceae was put in the Cantharellales without ex-planation by Julich (1981), and this was accepted inthe Dictionary of Fungi 8th edition by Hawksworthet al (1995). Hibbett et al (1997) included Sparassisspathulata in their molecular study on major groupsof gilled mushrooms, polypores and puffballs, which

1016 MYCOLOGIA
was based on phylogenetic analyses of nuc-ssu andmt-ssu rDNA. Sparassis was resolved as the sistergroup of a clade including two polypores Laetiporussulphureus (Bull. : Fr.) Murr. and Phaeolus schweinitzii(Fr.) Pat. (Hibbett et al 1997). The position of Spa-rassis in the polyporoid clade was supported in otherstudies (Hibbett and Donoghue 2001), and thatplacement is accepted in the Dictionary of Fungi 9thedition (Kirk et al 2001).
Sparassis species are known as brown-rot producerswith a bipolar mating system (Weir 1917, Martin andGilbertson 1976). By analyzing character correlationsamong wood-decay types, mating systems and hostranges in homobasidiomycetes, Hibbett and Dono-ghue (2001) did not support Gilbertson’s hypothesisof a correlation between production of a brown rotand possession of a bipolar mating system. They in-ferred that brown-rot fungi are not monophyletic inthe polyporoid clade and that the pattern of trans-formations between different wood-decay types iscomplicated. Two strongly supported groups of gen-era—one of them is Laetiporus-Phaeolus-Sparassis—were united by the production of brown rot on thebutt or root of living or dead trees and a bipolarmating system (Hibbett and Donoghue 2001). Moregenes, including partial nuclear gene sequences cod-ing for the second largest subunit of RNA polymeraseII (RPB2), were used in this study to test the rela-tionships among Sparassis and other brown-rot poly-pores. Genes that encode the subunits of nuclearRNA polymerase, especially RPB1 and RPB2, arepromising phylogenetic markers in fungal systematics(Liu et al 1999, Matheny et al 2002).
Sparassis crispa has been reported in Asia from Ja-pan to the Tibet highland in western China (Imazekiet al 1988, Teng 1995, Mao et al 1993). In many as-pects, Asian ‘‘S. crispa’’ resembles S. radicata Weirfrom western North America more than ‘‘S. crispa’’from eastern North America and Europe. The rela-tionship between Asian and North American florashas been discussed for many years among botanistsand mycologists (Mueller et al 2001). Early fungalbiogeographic studies were based on specimens anddescriptions in the literature. These studies were lim-ited by the high degree of morphological variationof macrofungi, the existence of huge unexplored ar-eas in eastern Asia and inconsistent applications ofmorphological species concepts. Several distributionpatterns, such as eastern Asia-eastern North Americadisjuncts, eastern North America-western NorthAmerica disjuncts or eastern Asia-western NorthAmerica disjuncts, had been suggested by studies ofvascular plants, bryophytes and lichens (Redhead1989, Zang 1992, Wu and Mueller 1997, Wen 1999,Wu et al 2000, Mueller et al 2001). Molecular phy-
logenetic hypotheses coupled with information frommorphological studies are becoming key to assessingspecies concepts and biogeographic relationships infungi (Hibbett et al 1998, Mueller et al 2001). Forexample, Wu et al (2000) and Mueller et al (2001)tested the distribution pattern of species of Armillar-ia, Suillus and Xerula from eastern Asia and NorthAmerica based on ITS sequences. Given that the ITSsequences can be too variable to be aligned acrossdistantly related taxa, combined data from lsu-rDNA,ITS and partial RPB2 genes were used in this studyto resolve the distribution pattern of Sparassis spe-cies.
MATERIALS AND METHODS
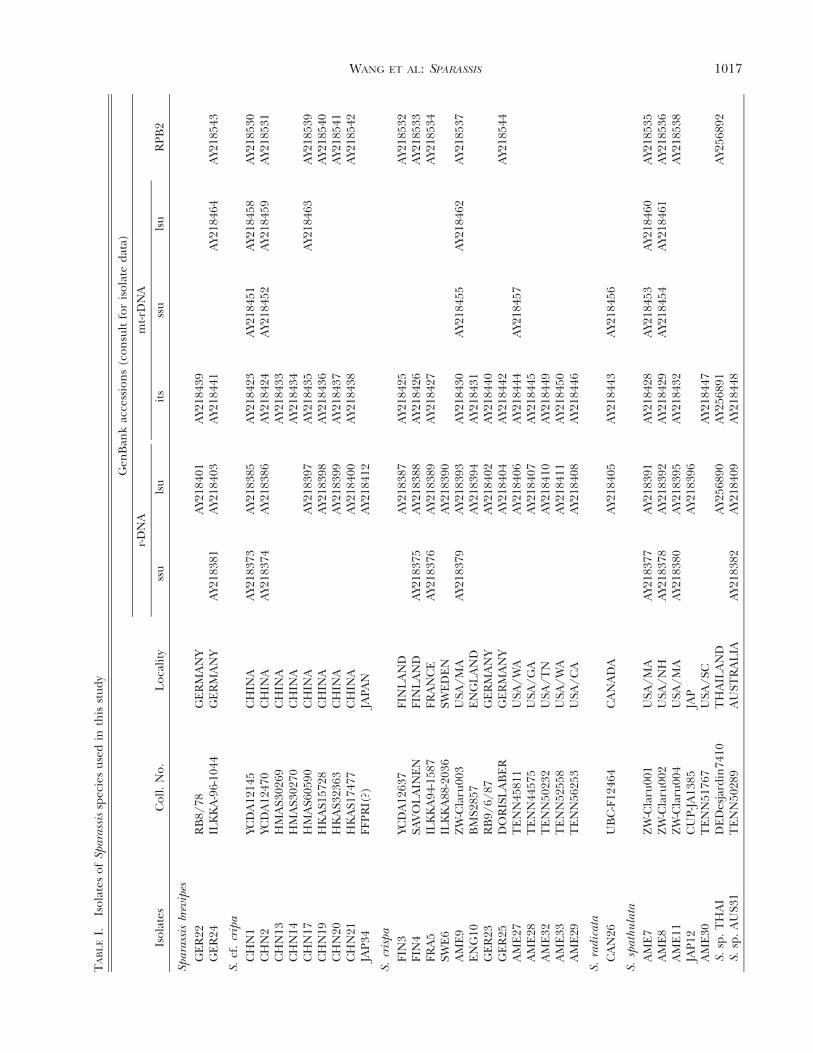
Specimens and morphological studies.—Specimens used inthis study and their GenBank accession numbers are givenin TABLE I and TABLE II. Morphological descriptions arebased on observations of fresh, dried or rehydrated speci-mens. Microscopic studies used squashed tissues and sec-tions cut with a freezing microtome to a thickness of 15–20!m. Measurements were made under 1% Congo red in am-monium hydroxide using bright field (Olympus CH-2).
Molecular techniques.—DNA was isolated from dried fruitingbodies. Approximately 20–30 mg of tissue was ground inliquid nitrogen and extracted in 600 !l of extraction buffer(1% SDS, 0.15 M NaCl, 50 mM EDTA) at 75 C for 1 h andpurified with phenol-chloroform-isoamyl alcohol (25:24:1)and precipitated with 95% ethanol and 3M NaCl overnight.Crude DNA extracts were purified with GeneClean (Bio101, La Jolla, California). Cleaned DNA samples were di-luted with distilled water up to 500-fold for use as PCR tem-plates.
Sequence data of Sparassis species were generated fromsix regions (TABLE I): (1) partial mitochondrial small sub-unit (mt-ssu) rDNA bounded by primers MS1and MS2(White et al 1990), representing four species of Sparassis inseven isolates; (2) partial mitochondrial large subunit (mt-lsu) bounded by primers ML5 and ML6 (White et al 1990),representing four species of Sparassis in five isolates; (3)nearly complete nuclear small subunit (nuc-ssu) rDNAbounded by primers PNS1 and NS8 (White et al 1990, Hib-bett 1996), representing four species of Sparassis in 10 iso-lates; (4) partial nuclear large subunit (nuc-lsu) rDNAbounded by primers LR0R and LR5 (Vilgalys and Hester1990), representing seven species of Sparassis in 29 isolates;(5) complete internal transcribed spacers 1 and 2 and the5.8S rDNA (nuc-its rDNA) bounded by primers ITS4 andITS5 (White et al 1990), representing seven species of Spar-assis in 29 isolates; (6) partial gene encoding RNA poly-merase subunit II (RPB2) bounded by primers RPB2-6F, 5"-TGG GGK WTG GTY TGY CCT GC-3", and RPB2-7R, 5"-CCCAT WGC YTG CTT MCC CAT-3" (http://faculty.washington.edu/benhall/), representing five species ofSparassis in 16 isolates. Additional data of other fungal ma-terials were generated from these six regions or download-
1017WANG ET AL: SPARASSIS
TA
BL
EI.
Isol
ates
ofSp
aras
sis
spec
ies
used
inth
isst
udy
Gen
Ban
kac
cess
ions
(con
sult
for
isol
ate
data
)
Isol
ates
Col
l.N
o.Lo
calit
yr-D
NA
ssu
lsu
mt-r
DN
Aits
ssu
lsu
RPB
2
Spar
assi
sbr
evip
esG
ER22
GER
24R
B8/
78IL
KK
A-9
6-10
44G
ERM
AN
YG
ERM
AN
YAY
2183
81AY
2184
01AY
2184
03AY
2184
39AY
2184
41AY
2184
64AY
2185
43
S.cf
.cri
paC
HN
1C
HN
2YC
DA
1214
5YC
DA
1247
0C
HIN
AC
HIN
AAY
2183
73AY
2183
74AY
2183
85AY
2183
86AY
2184
23AY
2184
24AY
2184
51AY
2184
52AY
2184
58AY
2184
59AY
2185
30AY
2185
31C
HN
13C
HN
14C
HN
17C
HN
19C
HN
20C
HN
21JA
P34
HM
AS3
0269
HM
AS3
0270
HM
AS6
0590
HK
AS1
5728
HK
AS3
2363
HK
AS1
7477
FFPR
I(?)
CH
INA
CH
INA
CH
INA
CH
INA
CH
INA
CH
INA
JAPA
N
AY21
8397
AY21
8398
AY21
8399
AY21
8400
AY21
8412
AY21
8433
AY21
8434
AY21
8435
AY21
8436
AY21
8437
AY21
8438
AY21
8463
AY21
8539
AY21
8540
AY21
8541
AY21
8542
S.cr
ispa
FIN
3FI
N4
FRA
5
YCD
A12
637
SAV
OLA
INEN
ILK
KA
94-1
587
FIN
LAN
DFI
NLA
ND
FRA
NC
EAY
2183
75AY
2183
76
AY21
8387
AY21
8388
AY21
8389
AY21
8425
AY21
8426
AY21
8427
AY21
8532
AY21
8533
AY21
8534
SWE6
AM
E9IL
KK
A88
-203
6ZW
-Cla
ru00
3SW
EDEN
USA
/MA
AY21
8379
AY21
8390
AY21
8393
AY21
8430
AY21
8455
AY21
8462
AY21
8537
ENG
10G
ER23
GER
25A
ME2
7A
ME2
8A
ME3
2A
ME3
3A
ME2
9
BM
S285
7R
B9/
6/87
DO
RIS
LAB
ERT
ENN
4581
1T
ENN
4457
5T
ENN
5023
2T
ENN
5255
8T
ENN
5625
3
ENG
LAN
DG
ERM
AN
YG
ERM
AN
YU
SA/W
AU
SA/G
AU
SA/T
NU
SA/W
AU
SA/C
A
AY21
8394
AY21
8402
AY21
8404
AY21
8406
AY21
8407
AY21
8410
AY21
8411
AY21
8408
AY21
8431
AY21
8440
AY21
8442
AY21
8444
AY21
8445
AY21
8449
AY21
8450
AY21
8446
AY21
8457
AY21
8544
S.ra
dica
taC
AN
26U
BC
-F12
464
CA
NA
DA
AY21
8405
AY21
8443
AY21
8456
S.sp
athu
lata
AM
E7A
ME8
AM
E11
ZW-C
laru
001
ZW-C
laru
002
ZW-C
laru
004
USA
/MA
USA
/NH
USA
/MA
AY21
8377
AY21
8378
AY21
8380
AY21
8391
AY21
8392
AY21
8395
AY21
8428
AY21
8429
AY21
8432
AY21
8453
AY21
8454
AY21
8460
AY21
8461
AY21
8535
AY21
8536
AY21
8538
JAP1
2A
ME3
0S.
sp.T
HA
IS.
sp.A
US3
1
CU
P-JA
1385
TEN
N51
767
DED
esja
rdin
7410
TEN
N50
289
JAP
USA
/SC
TH
AIL
AN
DA
UST
RA
LIA
AY21
8382
AY21
8396
AY25
6890
AY21
8409
AY21
8447
AY25
6891
AY21
8448
AY25
6892

1018 MYCOLOGIA
TABLE II. Isolates of other fungi used in this study
Species
GenBank accessions (consult for isolate data)
18s-rDNA LR0R-LR5 ITS1-ITS4 MS1-MS2 ML5-ML6 RPB2/6.3-7
Agaricus bisporusAlbatrellus flettiAlbatrellus skamaniusAlbatrellus syringaeAmanita muscariaAmylostereum laevigatumAntrodia carbonicaAuriporia aureaAuriscalpium vulgare
AJ012396AF518569AF287829AF026632AF026631AF334901AF026570AF334903AF026581
U11910AF518596AF393044AF393045AF042643AF287843AF287844AF287846AF287847
AF287817AF026674AF026673AF334871U27023AF334873U27024
AD001540AD001542AD001543AD001549AF393088AF393089AF393091AF393092
AF107785AY218465AY218466AY218467AY218468AY218469AY218470AY218471AY218472
Boletus satanasBotryobasidium isabellinumCeriporia purpureaChondrostereum purpureumColtricia perennisConiophora aridaDacrymyces chrysospermusDentocorticium sulphurellumEchinodontium tinctorium
M94337AF026610AF026594AF082851AF026583AY293123L22257AF026604AF026578
AF071528AF393047AF287852AF518607AF287854AF098375AF287855AF393055AF393056
M91009AF393083U27029AF518672U27028AF518674AF026642AF026647U27035
AD001566AF393097AF393100AF518704AF393103
AF393106AF393111AF393112
AY218473AY218475AY218476AY218477AY218526AY218478AY218480AY218481AY218482
Entoloma strictiusFomitopsis pinicolaGanoderma australeGautieria otthiiGrifola frondosa
AF287832AF026599AF026629AF393043AY218383
AF393057AF287858X78780AF336249AY218413 AY218415
AF287820U27038AF026672AF393085
AF393113AF393115AF393116AF393117
AY218483AY218484AY218485AY218486AY218521
Heliocybe sulcataHeterobasidion annosumHydnum repandumHyphodontia alutariaLaeticorticium roseocarneumLaetiporus sulphureusLentinula lateritia
AF334915AF026576AF026641AF026615AF334910AY218384AF026596
AF518619AF139949AJ279576AF393061AF393053AY218414AF287872
AY218417
AF334881U27042AF026683AF026660AF334875
U27047
AD001593AF393124AF393125AF393109
AY218487AY218488AY218489AY218490AY218491AY218522AY218492
Lentinus tigrinusLycoperdon sp.
U59080AF026619
AF518627AF287873
AY218419 U27050AF026663 AF393131
AY218493AY218495
Meripilus giganteusNeolentiporus maculatissimusNia vibrissaOligoporus lacteusOligoporus rennyi
U59082AF334921AF334754AY293152AF334922
AF287874AF518632AF334750AY293205AF287876 AY218416
U27053AF334884AF334753AY293241AF334885
AF334748AY293267AF393132
AY218496AY218497AY218498AY218510AY218499
Peniophora nudaPhaeolus schweinitziiPhlebia radiataPleurotus ostreatusPlicaturopsis crispaPolyporus squamosus
AF026586AF026598AF026606U23544AY293148AF026573
AF287880AF287882AF287885U04160AY293203AF393069
AY218422
AY218421AY218418
U27063U27066AF026649U27064AY293239U27068
AF393134
AF393136AF393137AY293266AF393142
AY218500AY218501AY218502AY218504AY218505AY218508
Pycnoporellus fulgensRussula compactaSarcodon imbricatusSchizophyllum communeScytinostroma alutumSerpula himantioides
AF518587AF026582AY293157X54865AF026607AF518589
AF518643AF287888AF518646AF334751AF393075AF518648
AF518690U27074AF518693AF069639AF026650AF518694
AF393148
AF334747AF393150
AY218527AY218514AY218528AY218515AY218516AY218517
Sistotrema eximumSistotrema muscicolaStereum hirsutumSuillus cavipesThelephora sp.Typhula phacorrhiza
AF334935AF334936AF026588M90828AF026627AF026630
AF393076AF518649AF393078AF071535AF287890AF393079
AF334891AF334892U27076M91016AF026670AF026687
AF393151
AF393153AD001641AD001646AF393154
AY218518AY218519AY218520AY218523AY218529AY218525

1019WANG ET AL: SPARASSIS
ed from GenBank (TABLE II). Strain numbers of some iso-lates were provided in Binder and Hibbett (2002).
PCR reaction mixes (Promega Corp., Madison, Wiscon-sin) contained 2.5 !L 10# PCR buffer, 5 !M dNTP, 12.5pM of each PCR primer and 5 !L DNA in 25 !L. Theamplification program included 40 cycles of 94 C for 30s,45 C for 30s, and 72 C for 1 min. PCR products were pu-rified using GeneClean (Bio 101, Carlsbad, California) andsequenced using the ABI Prism BigDye-terminator cycle se-quencing kit (Applied Biosystems, Foster City, California)according to the manufacturer’s protocols. Primers used forsequencing were PNS1, NS19bc, NS19b, NS41, NS51, NS6,NS8, MS1, MS2, ML5, ML6, RPB2-6F, RPB2-6.3F, RPB2-7R,RPB2-7.1R, LR0R, LR3, LR3R, LR5, ITS4, and ITS5. Se-quencing reactions were purified using Pellet Paint (Nova-gen, Madison, Wisconsin) and were run on an Applied Bio-systems 377XL automated DNA sequencer. Sequences wereedited with Sequencher version 3.1 (GeneCodes Corp., AnnArbor, Michigan). Sequences generated in this study weresubmitted to GenBank (accession numbers AY218373–AY218547).
Phylogenetic analyses.—Two datasets were prepared, one forhigher-level analyses (HLA) and one for lower-level analyses(LLA). The datasets for the HLA included sequences fromfive genes: mt-ssu rDNA (68 isolates), mt-lsu rDNA (59 iso-lates), nuc-ssu rDNA (72 isolates), nuc-lsu rDNA (72 iso-lates) and RPB2 (71 isolates). HLA were intended to resolvethe placement of Sparassis among 61 homobasidiomycetegenera (TABLE I and TABLE II). The datasets for the LLAincluded sequences from three genes nuc-lsu (36 isolates),ITS (37 isolates) and RPB2 (24 isolates). LLA were intend-ed to resolve the relationships among Sparassis species (TA-BLE I and TABLE II). Sequences of nuc-rDNA and mt-rDNAwere aligned by eye in the data editor of PAUP* 4.0b (Swof-ford 1999). Sequences of RPB2 were translated into aminoacid sequences and aligned by eye in Sequencher and thenconverted back to nucleotides for the analyses. Both data-sets were analyzed in PAUP* 4.0b (Swofford 1999) andMrBayes 2.01 (Huelsenbeck and Ronquist 2001), with gapstreated as missing data and ambiguous or unalignable po-sitions excluded. Ambiguous positions were excluded fromthe datasets before performing the analyses.
The HLA dataset was rooted with Dacrymyces chrysosper-mus Berk. & M.A. Curtis (Hibbett and Donoghue 2001).Ssu-rDNA sequences of Bondarzewia montana (Quel.) Sing-er, Piptoporus betulinus (Bull. : Fr.) P. Karst., Pycnoporus cin-nabarinus ( Jacq. : Fr.) P. Karst., Ramaria formosa (Pers. :Fr.) Quel, Trametes versicolor (L. : Fr.) Pilat, Polyporus varius(Pers.) Fr., Polyporus melanopus (Pers.) Fr., Polyporus arcu-larius (Batsch) Fr., Cryptoporus volvatus (Peck) Shear andLenzites betulina (Fr.) Fr. represented a shorter region thansequences of other taxa. Mt-ssu sequences of Albatrellus flet-ti (Morse) Pouzar, Grifola frondosa (Fr.) S.F. Gray, Sparassisbrevipes GER24, and S. sp. AUS31, and mt-lsu sequences ofS. crispa USA9, S. brevipes GER24, Trametes versicolor, Helio-cybe sulcata (Berk.) Redhead & Ginns, Lentinula lateritia(Berk.) Pegler, Lentinus tigrinus (Fr.) Fr., Neolentiporus ma-culatissimus (Lloyd) Rajchenb., Phaeolus schweinitzii (Fr.)Pat., Piptoporus betulinus, Ramaria formosa, Sparassis sp.
AUS31, Pycnoporellus fulgens (Fr.) Donk and one unidenti-fied resupinate homobasidiomycete were not available, andthose taxa were excluded from the analysis based on mt-DNA. No sequence of RPB2 of S. sp. AUS31 was generated,and it was excluded in analyses using RPB2 data.
Parsimony analyses were performed using equal weight-ing of characters and transformations. Heuristic searcheswere performed with 1000 replicate searches, each with arandom taxon addition sequence, MAXtrees set to autoin-crease, and TBR branch swapping. A bootstrap analysis wasperformed with 1000 replicates, each with 10 random taxonaddition sequences, MAXtrees set to 1000, and TBR branchswapping. Bootstrap consensus trees generated from anal-ysis of each gene were compared to check whether there isan apparent conflict in tree topologies. Bootstrap valueshigher than 80% were used as the criterion for ‘‘strong’’conflict. The combined analyses were performed using nuc-rDNA and RPB2 sequences, which were congruent accord-ing to this criterion.
Bayesian posterior probabilities were computed using Me-tropolis-coupled Markov Chain Monte Carlo (MCMCMC)under the GTR$G model in MrBayes 2.01 (Huelsenbeckand Ronquist 2001) by running four chains with 200 000generations using the program default priors on model pa-rameters. Trees were sampled every 100 generations, and atotal of 2001 trees were saved. Likelihoods converged to astable value after 5000 generations, so the first 50 trees werediscarded as ‘‘burn-in’’ before computing a majority ruleconsensus tree in PAUP*.
The LLA dataset was rooted using Lentinus tigrinus basedon the results of the HLA. The same analytical settings asthat for the HLA were applied to the LLA for separate andcombined data of nuc-lsu rDNA, ITS and RPB2. Isolateswithout RPB2, ITS and nuc-lsu rDNA sequences were ex-cluded from different analyses. Alignments are available atTreeBASE (accession number M1814-M1815).
RESULTS
Phylogenetic relationships.—HLA. The higher-levelplacement of Sparassis was estimated using five dif-ferent datasets: nuc-rDNA; mt-rDNA; RPB2; and, nuc-rDNA $ RPB2 (FIGS. 1–5).
The combined nuc-ssu rDNA and nuc-lsu rDNAgenes had an aligned length of 2694 base pairs (bp)with 374 uninformative variable positions and 628parsimony-informative positions. Parsimony analysisbased on nuc rDNA generated 140 equally parsimo-nious trees of 4416 steps and consistency index (CI)% 0.336 (FIG. 1). A clade including S. spathulata, S.brevipes, S. crispa, S. sp. AUS31, several other brown-rot species and Grifola frondosa was weakly supported(bootstrap % 53%) within the polyporoid clade. Aclade (bootstrap % 90%) containing S. spathulataand S. brevipes and a clade (bootstrap % 100%) con-taining two isolates of S. crispa were grouped togeth-er with the Phaeolus-Laetiporus clade (bootstrap %50%) as the sister group (bootstrap % 72%). Spa-

1020 MYCOLOGIA
FIG
S.1–
3.H
ighe
r-lev
elph
ylog
enet
icre
latio
nshi
psof
Spar
assi
sin
ferr
edw
ithsi
ngle
-gen
eeq
ually
wei
ghte
dpa
rsim
ony
anal
ysis
.1.N
uc-r
DN
Aan
alys
is.1
of14
0eq
ually
pars
imon
ious
tree
s(L
engt
h%
4416
,CI
%0.
336,
RI
%0.
461)
.2.M
t-rD
NA
anal
ysis
.1of
733
equa
llypa
rsim
onio
ustr
ees
(Len
gth
%18
10,C
I%
0.36
4,R
I%
0.49
6).3
.R
PB2
anal
ysis
.1
of3
equa
llypa
rsim
onio
ustr
ees
(Len
gth
%52
53,
CI
%0.
172,
RI
%0.
357)
.B
row
n-ro
tsp
ecie
sar
ein
bold
type
.N
odes
that
colla
pse
inth
est
rict
cons
ensu
str
eear
em
arke
dw
ithan
aste
risk
abov
eth
ebr
anch
.Boo
tstr
apva
lues
grea
ter
than
50%
are
indi
cate
dal
ong
node
s.

1021WANG ET AL: SPARASSIS
rassis sp. AUS31 was grouped with Grifola frondosaand Pycnoporellus fulgens (bootstrap % 100%). Thebrown-rot polypores Fomitopsis, Piptoporus and Neo-lentiporus formed a clade (bootstrap % 67%). Rela-tionships between white-rot and brown-rot species inthe polyporoid clade were not well resolved.
The mt-rDNA had an aligned length of 684 bp with117 uninformative variable positions and 290 parsi-mony-informative positions. Parsimony analysis basedon mt-rDNA (FIG. 2) generated 733 equally parsi-monious trees of 1810 steps with CI % 0.364. A cladecontaining Sparassis spathulata and S. crispa wasweakly supported (bootstrap % 63%) and wasgrouped with white-rot species including Ganoderma,Pycnoporus and Polyporus (bootstrap % 84%).
The RPB2 sequences had an aligned length of 510bp with 79 uninformative variable positions and 333parsimony-informative positions. Parsimony analysisbased on RPB2 (FIG. 3) generated three equally par-simonious trees of 5253 steps with CI % 0.172. Brown-rot species of Oligoporus, Antrodia, Auriporia, Phaeo-lus, Laetiporus, Sparassis, Fomitopsis, Piptoporus, Neo-lentiporus and the white-rot species Albatrellus syrin-gae (Parm.) Pouz. formed a weakly supported clade,within which there was a clade containing S. crispa,S. spathulata and S. brevipes (bootstrap % 93%).
The combined nuc-rDNA and RPB2 genes had analigned length of 3149 bp with 449 uninformativevariable positions and 954 parsimony-informative po-sitions. Parsimony analysis based on this dataset gen-erated 19 equally parsimonious trees of 9866 stepswith CI % 0.240 (FIG. 4). Sparassis species, includingS. spathulata, S. brevipes and S. crispa, were mono-phyletic (bootstrap % 87%) with the Phaeolus-Laeti-porus clade (bootstrap % 64%) as its sister group(bootstrap % 68%). A brown-rot polypore clade, theFomitopsis-Piptoporus-Neolentiporus clade, also was sup-ported (bootstrap % 87%), but the relationshipsamong brown-rot and white-rot species in the poly-poroid clade still were not well resolved.
Bayesian analysis based on combined nuc-rDNAand RPB2 sequences (FIG. 5) strongly supported(posterior probability % 93%) a clade containingSparassis, Phaeolus, Pycnoporellus, and Grifola, inwhich Sparassis species are monophyletic (posteriorprobability % 99%). Two more clades of brown rotpolypores were supported: the Fomitopsis-Piptoporus-Neolentiporus-Laetiporus clade (posterior probability% 85%) and the Oligoporus-Antrodia clade (posteriorprobability % 61%). In addition, the clade includingwhite rot species of Cryptoporus, Ganoderma, Lentin-us, Polyporus, Dentocorticium, Lenzites, Pycnoporusand Trametes is strongly supported (posterior proba-bility % 96%).
LLA. The lower-level relationships among Sparas-
sis species were estimated using five different data-sets: nuc-lsu rDNA; ITS; RPB2; nuc-lsu rDNA$ITS;and nuc-lsu rDNA$ITS$RPB2. Trees inferred fromsingle regions showed similar topologies to trees in-ferred from the combined sequences but with weakerbranch support (FIGS. 6–9).
The RPB2 sequences had an aligned length of 642bp with 87 uninformative variable positions and 225parsimony-informative positions. Parsimony analysisbased on the RPB2 gene generated 282 equally par-simonious trees of 789 steps with CI % 0.608 (FIG.6). A clade containing Sparassis crispa, S. cf. crispa,S. spathulata, S. brevipes and S. sp. THAI was sup-ported weakly (bootstrap % 61%) with S. sp. THAIas a basal branch. One North American isolate andfour European isolates of S. crispa and six Chineseisolates of S. cf. crispa formed a clade (bootstrap %100%). Isolates of S. spathulata formed a clade (boot-strap % 96%) with S. brevipes as its sister group (boot-strap % 100%). The relationship among the Sparassisclade and other brown-rot polypores were not re-solved. Unfortunately, no RPB2 sequences were ob-tained for S. radicata, S. sp. AUS31 and isolates of S.crispa collected from Georgia (AME28), Tennessee(AME32) and western North America (TABLE I).
The combined sequences of nuc-lsu rDNA, ITSand RPB2 had an aligned length of 2080 bp with 286uninformative variable positions and 505 parsimony-informative positions. Parsimony analysis based onthis dataset generated 20 equally parsimonious treesof 1818 steps with CI % 0.633 (FIG. 7). Sparassis spe-cies formed a monophyletic group (bootstrap %79%) with S. sp. THAI as a basal branch. Five cladeswere found within the Sparassis clade: Asian S. cf.crispa clade (bootstrap % 96%), European-NorthAmerican S. crispa clade (bootstrap % 64%), S. spa-thulata clade (bootstrap % 100%), S. brevipes and S.sp. THAI. Laetiporus and Phaeolus formed a clade(bootstrap % 98%), which is the sister group of theSparassis clade.
The combined sequences of nuc-lsu rDNA andITS had an aligned length of 1432 bp with 193 un-informative variable positions and 295 parsimony-informative positions. Parsimony analysis based onthis dataset generated 90 equally parsimonious treesof 1061 steps with CI % 0.644 (FIG. 8). Eight cladeswere recognized among Sparassis species: crispaclade (European isolates of S. crispa, bootstrap %69%), radicata clade (western North American iso-lates of S. crispa and S. radicata, bootstrap % 67%),Asian crispa clade (Chinese isolates of S. cf. crispa,bootstrap % 94%), brevipes clade (European isolatesof S. brevipes, bootstrap % 100%), spathulata clade(eastern North American isolates of S. spathulata,bootstrap % 100%), and two unidentified Sparassis

1022 MYCOLOGIA
FIG
S.4–
5.H
ighe
rle
velp
hylo
gene
ticre
latio
nshi
psof
Spar
assi
sin
ferr
edw
ithm
ultig
ene
pars
imon
yan
dB
ayes
ian
anal
ysis
.4.P
arsi
mon
yan
alys
isba
sed
onth
eco
mbi
ned
nuc-
rDN
Aan
dR
PB2
sequ
ence
s.1
of19
equa
llypa
rsim
onio
ustr
ees
(Len
gth
%98
66,C
I%
0.24
0,R
I%
0.38
1).B
row
n-ro
tsp
ecie
sar
ein
bold
type
.Nod
esth
atco
llaps
ein
the
stri
ctco
nsen
sus
tree
are
mar
ked
with
anas
teri
skab
ove
the
bran
ch.B
oots
trap
valu
esgr
eate
rth
an50
%ar
ein
dica
ted
alon
gno
des.
5.B
ayes
ian
anal
ysis
base
don
the
com
bine
dnu
c-rD
NA
and
RPB
2se
quen
ces.
The
maj
ority
rule
cons
ensu
sof
1952
MC
MC
MC
-sam
pled
tree
s.B
row
n-ro
tsp
ecie
sar
ein
bold
type
.Gro
upfr
eque
ncie
sgr
eate
rth
an50
%ar
ein
dica
ted
aspo
ster
ior
prob
abili
ty(%
)al
ong
node
s.

1023WANG ET AL: SPARASSIS
FIG
S.6–
8.Lo
wer
-leve
lph
ylog
enet
icre
latio
nshi
psof
Spar
assi
sin
ferr
edw
ithsi
ngle
-gen
ean
dm
ulti-
gene
anal
ysis
.6.
Rel
atio
nshi
psam
ong
Spar
assi
ssp
ecie
sin
ferr
edfr
omth
eR
PB2
sequ
ence
s.1
of28
2eq
ually
pars
imon
ious
tree
s(L
engt
h%
798,
CI
%0.
608,
RI
%0.
656)
.7.
Rel
atio
nshi
psam
ong
Spar
assi
ssp
ecie
sin
ferr
edfr
omth
eco
mbi
ned
nuc-
lsu-
rDN
A,I
TS,
and
RPB
2se
quen
ces.
One
of20
equa
llypa
rsim
onio
ustr
ees
(Len
gth
%18
18,C
I%
0.63
3,R
I%
0.67
9).8
.Rel
atio
nshi
psam
ong
Spar
assi
ssp
ecie
sin
ferr
edfr
omth
eco
mbi
ned
nuc-
lsu-
rDN
Aan
dIT
Sse
quen
ces.
One
of90
equa
llypa
rsim
onio
ustr
ees
(Len
gth
%10
61,
CI
%0.
644,
RI
%0.
765)
.C
lade
sar
ena
med
and
indi
cate
dby
blac
kba
rs.N
odes
that
colla
pse
inth
est
rict
cons
ensu
str
eear
em
arke
dw
ithan
aste
risk
abov
eth
ebr
anch
.Boo
tstr
apva
lues
grea
ter
than
50%
are
indi
cate
dal
ong
node
s.

1024 MYCOLOGIA
FIG. 9. Lower-level phylogenetic relationships of Sparassis inferred with multigene Bayesian analysis and representativefruiting body morphologies. The majority rule consensus of 1952 MCMCMC-sampled trees. Group frequencies greater than50% are indicated as posterior probability (%) along nodes.
species, S. sp. THAI and S. sp. AUS31. Two easternNorth American isolates of S. crispa (S. crispa AME9and AME28) formed a branch close to the radicataclade and the Asian crispa clade without bootstrapsupport. Relationships among those clades and lin-eages were not completely resolved. The crispaclade, the radicata clade, the Asian crispa clade andtwo eastern North American S. crispa were support-ed as monophyletic (bootstrap % 100%), whereasthe brevipes clade and the spathulata clade weregrouped together (bootstrap % 62%). S. sp. AUS31was grouped with Grifola and Pycnoporellus (boot-strap value less than 50%).
Bayesian analysis based on the combined sequenc-es of nuc-lsu rDNA and ITS provided higher confi-dence for all clades and lineages of Sparassis ob-served in the parsimony analysis (FIG. 9). The crispa
clade (posterior probability % 98%), the radicataclade (posterior probability % 74%), the Asian crispaclade (posterior probability % 100%), the brevipesclade (posterior probability % 100%), and the spa-thulata clade (posterior probability % 100%) wereconfirmed. Two eastern North American isolates ofS. crispa formed a weakly supported clade (posteriorprobability % 64%). Sparassis species were stronglysupported as monophyletic (posterior probability %86%) with Grifola and Pycnoporellus as the sistergroup. All species from the Northern Hemisphereformed a clade (posterior probability % 100%). S.brevipes and S. spathulata were grouped together(posterior probability % 99%), and the crispa clade,the radicata clade, the Asian crispa clade and easternNorth American S. crispa formed a monophyleticgroup (posterior probability % 100%).

1025WANG ET AL: SPARASSIS
MORPHOLOGY AND TAXONOMY
The lack of type specimens hampers the taxonomicand nomenclatural studies of the genus Sparassis.Martin and Gilbertson (1976), Kreisel (1983), andBurdsall and Miller (1988a, b), have produced au-thoritative taxonomies of Sparassis. This study usedrecent collections because they provide high qualityDNA for molecular studies. For nomenclatural pur-poses, however, it would be useful to attempt to ex-tract DNA from the older specimens studied by Gil-bertson and others. All the collections were studiedmorphologically following widely accepted speciesconcepts, which are based on Weir (1917), Kreisel(1983) and Burdsall and Miller (1988a, b). Simplifiedtaxonomic descriptions and habit illustrations (FIG.9) based on our own observations are provided inaddition to the major clades found in phylogeneticanalyses (as in FIG. 8). Commentary is provided re-garding morphological incongruence or conflictamong the members of each clade.
brevipes clade (represented by Sparassis brevipesKrombh.)Base of basidiocarps composed of a round mass.
Flabellae spathulate, all originating directly from acommon base, split into pieces and then fused withothers several times on the way to the top, azonateto not distinctively zonate, margin entire. Hyphal sys-tem monomitic, simple septate, no clamp connectionobserved. Basidiospores hyaline, thin-walled, smooth,broadly elliptic to subglobose, (3.9–)4.4–5.2(–5.5) #(6.2–)6.8–7.8(–8.7) !m. Distribution: Europe.
Specimens examined. GERMANY. BADEN-WURTTEM-BERG: In mixed forest of Abies, Picea, Pinus and Fagus onlimestone, 24-IX-1995, MBUH (ILKKA96-1044); VIII-1978,RB8/78.
Commentary. Sparassis brevipes has been docu-mented in Europe without precise typification (Burd-sall and Miller 1988a). Morphological studies showedthat there are at least two distinct Sparassis species inEuropean samples. We followed Kreisel’s (1983) con-cept and accepted the widely used name S. brevipesto represent the European species mostly growing onAbies, Fagus and Quercus. S. brevipes is similar to theeastern North American S. spathulata in spore sizeand possession of simple septate hyphae, but the fla-bellae of S. brevipes are azonate.
crispa clade (represented by Sparassis crispa Wulf. :Fr.)Base of basidiocarps branched and elongated to
form several flattened branches. Flabellae extendfrom the branches, broad but short, dissected andcontorted, azonate, margin entire. Hyphal system
monomitic, clamp connections present, gelatinoushyphae rarely present. Basidiospores hyaline, thin-walled, smooth, subglobose, 4.0–4.9 # 4.9–6.0(–6.9)!m. Distribution: Europe, eastern North America.
Specimens examined. USA. MASSACHUSETTS: PurgatoryChasm State Park, 15-IX-2001, FH (ZW-Clarku003); GEOR-GIA: Fort Top Mountains, 13-IX-1983, TENN44575. FIN-LAND. North Kotinen Forest: on Pinus, 12-IX-1997, MBUH(YCD2637); PAINO TUOREENAN: 25-IX-1983, MBUH(Savolaninen). FRANCE. LOZERE: in mixed forest of Fa-gus, Picea and Quercus, 21-X-1994, MBUH (PIR-JO&ILKKA94-1587). SWEDEN. NARKE: on Pinus, 26-X-1988, MBUH (ILKKA88-2036). UK. ESHER COMMON,SURREY: collected by P.M. Kirk, on Pinus sylvestris, 29-X-2000, BMS2857. GERMANY. 6-IX-1987, RB9/6/87; collect-ed by Doris Laber, 27-IX-1996, MBUH.
Commentary. Neotypification of Sparassis crispa wasbased on a European collection (Burdsall and Miller1988b). S. crispa has been reported from Europe,eastern Asia and North America (Breitenbach andKranzlin 1986; Burdsall and Miller 1988a, 1988b; Gil-bertson 1980; Imazeki et al 1988; Mao et al 1993;Martin and Gilbertson 1976). Asian collections underthe name of S. crispa are morphologically differentfrom European S. crispa. European S. crispa collec-tions are characterized by branched basidiocarps,strongly dissected and contorted flabellae and arestrictly growing on conifers. Two North American iso-lates, FH-ZW-Clarku003 (S. crispa AME9) andTENN44575 (S. crispa AME28), were not nested inthe crispa clade in two analyses (FIGS. 8–9) and needfurther study. FH-ZW-Clarku003 collected from Mas-sachusetts is similar to the European S. crispa exceptfor possessing gelatinous hyphae. TENN44575 col-lected from Georgia is similar to the western NorthAmerican S. radicata, which has bigger, slightly dis-sected and contorted flabellae.
Asian crispa clade (represented by Sparassis cf. crispaWulf. : Fr.)Base of basidiocarps branched or not. Flabellae
mostly extend from a common central mass, broad,dissected and slightly contorted, azonate, marginssometimes tooth-like. Hyphal system monomitic,clamp connections present. Basidiospores hyaline,thin-walled, smooth, subglobose, 4.0–5.0 # 5.0–6.0(–6.6) !m. Distribution: eastern Asia ( Japan and Chi-na).
Specimens examined. CHINA. JILIN, ANTU: Changbais-han Mountains, on Larix sp., 16-IX-1995, MBUH(YCD2145); on Larix sp., 14-VIII-1997, MBUH(YCD2470);on conifers, 25-VII-1960, HMAS30269; on conifers, 1-VIII-1960, HMAS30270; NEIMENGU: Arxan Mountains, 14-VIII-1991, HMAS60590; SICHUAN, DAOCHENG: on Picea,11-VIII-1984, HKAS15728; SICHUAN, XIANGCHENG: onQuercus sp., 17-VII-1998, HKAS 32363; YUNNAN, ZHONG-

1026 MYCOLOGIA
DIAN: on Larix, 25-VII-1986, HKAS17477. JAPAN. NAGA-NO PREF, USUDA: on Larix, VII-1988, FFPRI(Tsengoku); 5-IX-1989, FFPRI(F-15706).
Commentary. Sparassis crispa reported and collect-ed from Japan and China are macro-morphologicallydistinct from European S. crispa and are referred toas S. cf. crispa in this study. Flabellae of S. cf. crispaare similar to those of S. radicata, but they are biggerand not as contorted as in European S. crispa collec-tions. Chinese collections show a host range from co-nifers such as Larix, Pinus to hardwood plants (dac-tyls) such as Quercus. S. radicata so far has been re-ported only on Douglas-fir and Pines (Martin andGilbertson 1976).
radicata clade (represented by Sparassis radicataWeir)Basidiocarps expand from a round mass. Flabellae
anastomosed, vertically subdivided, broad, dissectedand slightly contorted, azonate. Hyphal system mon-omitic, clamp connections present. Basidiospores hy-aline, thin-walled, smooth, broadly elliptic to subglo-bose, 3.9–5.0 # 6.3–7.0 !m. Distribution: westernNorth America.
Specimens examined. CANADA. VANCOUVER: StanleyPark, on Tsuga heterophylla, collected by P. Koreger, 6-X-1985, UBC(F12464). USA. WASHINGTON: on the base ofTsuga sp., 4-X-1984, TENN45811; on base of conifer, 3-X-1992, TENN52558; CALIFORNIA: 13-II-1998, TENN56253;TENNESSEE: Great Smoky Mountains National Park, onbase of Pinus sp., 28-VII-1991, TENN50232.
Commentary. S. radicata is well studied and wasthought to be conspecific with the European andAsian S. crispa, mainly based on culture studies anddikaryon-monokaryon mating tests (Martin and Gil-bertson 1976). Morphologically, S. radicata is similarto Asian S. cf. crispa in having large but not stronglycontorted flabellae. The flabellae arise radially froma common center rather than from a branched baseas in the European S. crispa.
spathulata clade (represented by Sparassis spathulataSchw. : Fr.)Base of basidiocarps composed of a round mass.
Flabellae spathulate, all originating from a commonbase, anastomosing and vertically subdivided, distinc-tively zonate, margin entire. Hyphal system monom-itic, simply septate, no clamp connections observed.Basidiospores hyaline, thin-walled, smooth, broadlyelliptic to subglobose, 4.7–5.8 # (5.9–)6.9–8.0 !m.Distribution: eastern North America and Japan (?).
Specimens examined. USA, MASSACHUSETTS: on Quer-cus sp., 13-IX-2001, FH (ZW-Clarku001); WORCESTER: onQuercus sp., 1-X-2000, FH (ZW-Clarku004); NEWHAMPSPHERE: Walpole, 7-IX-2001, FH (ZW-Clarku002);
SOUTH CAROLINA: Oconee, 18-VIII-1992, TENN51767.JAPAN. HONSHU, TODAI-JI: collected by Kumiko andKorf, 22-VIII-1959, CUP(JA-1385).
Commentary. The large, zonate, vertically orientedflabellae of S. spathulata distinguish this species fromother Sparassis species. S. spathulata is similar to theEuropean S. brevipes (synonym: S. laminosa) in someaspects, and therefore validity of S. spathulata as aseparate species was doubted by Burdsall and Miller(1988a). Eastern North American S. spathulata havebeen found on both Pinus and Quercus, whereas S.brevipes has been found mostly on Quercus. One Jap-anese collection (CUP JA1385) was identified as S.spathulata, which might be introduced from NorthAmerica and so far is the only report of this speciesfrom Asia. Basidiocarp development of S. spathulatabegins with central daedaloid ridges on the surface.These ridges expand somewhat parallel to form fla-bellae with an amphigenous hymenium.
Sparassis sp. AUSBase of basidiocarps complex, flabellae narrow and
highly branched, azonate. Branch tips flattened, of-ten daedalioid arranged, then crested. Lower branch-es light ochraceous buff, upper branches pinkish buffto light ochraceous salmon. Margin with 2–4 fork-liketeeth when dried. Hyphal system monomitic, simplyseptate, no clamp connections observed. Basidio-spores hyaline, thin-walled, smooth, broadly ellipticto subglobose, 4.8–5.9 # 5.8–7.5 !m. Distribution:Australia.
Specimens examined. Australia. TASMANIA, VIS. GEES-TON: Tahune Forest Reserve, 5-VI-1991, by R. Peterson andA. Mills, TENN50289, Alan Mills (3996).
Commentary. This fungus shows a unique macro-morphology and was identified as Sparassis sp. by thecollectors. The amphigenous hymenium and the ar-rangement of the flabellae make this fungus a Spar-assis species. There is only one collection of this fun-gus, and the base was rotten away. More and bettermaterials from the southern hemisphere are neces-sary for further studies.
Sparassis sp. THAIBasidiocarps composed of a rosette of flabellae.
Flabellae loosely arranged, up to 120 mm broad, mar-gin not dissected, wavy, azonate. Hyphal system mon-omitic, clamp connections present. Hymenium layercomposed of basidia and cystidia. Basidiospores hya-line, thin-walled, smooth, subglobose to broadly ellip-soid-ovoid, 6–7 # 7–9 !m. Distribution: Thailand.
Specimens examined. THAILAND, CHIANG MAI PROV-INCE: Doi Inthanon National Park, at the base of a livingoak tree (Quercus eumorpha), 27-VI-2002, SFSU (D.E. Des-jardin 7410).

1027WANG ET AL: SPARASSIS
Commentary. Sparassis sp THAI is distinct fromother Sparassis species in possessing cystidia and larg-er basidiospores. Taxonomy of this fungus is dis-cussed elsewhere (Desjardin et al 2004).
DISCUSSION
Sequences from five loci (nuc-ssu rDNA, nuc-lsurDNA, mt-ssu rDNA, mt-lsu rDNA and RPB2) wereused to infer the higher-level phylogenetic relation-ships of Sparassis. Not all the possible combinationsof these loci have been analyzed, and nuc-rDNA(nuc-ssu rDNA $ nuc-lsu rDNA), mt-rDNA (mt-ssurDNA $ mt-lsu rDNA) and RPB2 genes have beentreated as three unlinked units. Sparassis species werenested within the polyporoid clade in all the analyses.
Parsimony analyses, except for the one based onthe RPB2 gene alone, showed a similar tree topologyfor the major clades in homobasidiomycetes. The nu-cleotide sequences of the RPB2 gene, between theprimer pairs bRPB2-6f and bRPB2-7r, are highly var-iable, which suggests that nucleotide sequences inthis part of the gene are saturated among taxa sam-pled here. Analyses based on the amino acid se-quences (data not shown) suggest that amino acidsequences in this part of the gene are too conservedto be informative for HLA, however. CombiningRPB2 sequences with nuc rDNA sequences increasedthe number of resolved clades (FIGS. 1, 3, 4) andreduced the number of equally parsimonious trees.
Sparassis species were grouped with some white-rotpolypores in the parsimony analysis based only onmitochondrial rDNA, whereas other analyses suggest-ed a close relationship between Sparassis species andother brown-rot polypores. Much of the mt-rDNA hasmultiple deletions and insertions and was excludedfrom the dataset. The mt-lsu rDNA sequences of one-fifth of the taxa sampled, including Phaeolus schwein-itzii, were not available, so inferences regarding thehigher-level phylogenetic relationships of Sparassisspecies based mostly on the combined nuc-rDNA andRPB2 sequences.
Sparassis species form a monophyletic group with-in the polyporoid clade supported by multilocus mo-lecular data (FIGS. 1–5). Sparassis is closely related toPhaeolus, Laetiporus, Pycnoporellus, Auriporia, Oligo-porus and other brown-rot polypores. Phaeolus andLaetiporus were confirmed as the sister group of Spa-rassis using equally weighted parsimony (FIGS. 1, 3,4) as in previous studies (Hibbett et al 1997). Phaeo-lus, Laetiporus and Sparassis are united by the pro-duction of a brown rot, bipolar mating system andthe habit of growth as a root and butt rot on livingtrees (rarely on dead trees and logs). Bayesian anal-ysis (FIG. 5) suggests a close relationship among Spar-
assis species, Phaeolus, Grifola and Pycnoporellus. Pyc-noporellus was thought to be closely related to Phaeo-lus (Gilbertson and Ryvarden 1987), and the cystidiasimilar to those found in Phaeolus and Pycnoporelluswere found in Sparassis sp. THAI as well (Desjardinet al 2004). Bayesian analysis supports a clade includ-ing Fomitopsis, Piptoporus, Neolentiporus and Laetipo-rus, which are all brown rot polypores with bipolarmating systems.
Sequences from three loci, nuc-lsu rDNA, ITS andRPB2, have been used to infer the lower-level phy-logenetic relationships of Sparassis. ITS sequences ofsampled taxa are divergent and difficult to align, andparsimony analysis based on ITS sequences alonegenerated more than 53 000 equally parsimonioustrees with little resolution (data not shown). As ex-pected, RPB2 were easier to align and provided moreinformative sites than ITS sequences. CombiningRPB2 gene with nuc-lsu rDNA and ITS providedhigher support to branches and better resolution(FIGS. 6–8). Unfortunately, RPB2 sequences were notobtained for most Sparassis collections from NorthAmerica.
Except for two unidentified Sparassis species, S. sp.AUS and S. sp. THAI, five well-supported clades werefound among Sparassis species. Asian isolates of S. cf.crispa show morphological differences comparedwith European S. crispa. Western North Americancollections of Sparassis, including three isolates iden-tified as S. crispa and one as S. radicata, formed aclade, and they morphologically are similar to theAsian S. cf. crispa. Sparassis spathulata, S. brevipes andthe European S. crispa are distinct species delimitedby both molecular data and morphological charac-ters. The recently collected cystidioid Sparassis sp.THAI was grouped with all Sparassis species andmight provide clues to the plesiomorphic form ofSparassis (Desjardin pers comm). Recognizing theAustralian collection (S. sp. AUS31) as a species ofSparassis denotes that this genus is distributed world-wide, but more material from the Southern Hemi-sphere is necessary to describe this species and toresolve the distribution pattern.
The relationships among members of Sparassis cris-pa sensu lato, which include European and easternNorth American S. crispa, western North AmericanS. radicata and Asian S. cf. crispa, were not well re-solved in all the analyses. Asian S. cf. crispa and west-ern North American S. radicata are morphologicallysimilar, but they are not strongly supported to forma monophyletic group. The eastern North AmericanS. crispa morphologically is identical to the EuropeanS. crispa, but molecular data do not suggest that thecollections from both locations are conspecific. Thiseastern North America-Europe distribution pattern

1028 MYCOLOGIA
was supported by the close relationship between east-ern North American S. spathulata and European S.brevipes. North American S. crispa and S. radicata col-lections show a morphological transition between Eu-ropean S. crispa and Asian S. cf. crispa (i.e., easternNorth American S. crispa is similar to European spe-cies whereas western North American S. radicata issimilar to Asian S. cf. crispa). However, the lack ofthe RPB2 sequences from western North Americancollections make it premature to draw conclusionsabout biogeographic relationships among S. crispasensu lato.
Sparassis is characterized morphologically by fla-bellae with an amphigenous hymenium and a centralmass giving rise to the flabellae. The presence ofclamp connections in Sparassis crispa, S. radicata, S.cf. crispa and S. sp. THAI suggests that clamp con-nections might have been lost in S. spathulata and S.brevipes. The spore size range in Sparassis species isoverlapping and therefore not always reliable to usefor recognizing species. Basidia with 2–4 sterigmatahave been observed in all examined collections andbasidiospores generally are larger when fewer sporesare produced on single basidium.
Host shifts must have occurred in the evolution ofSparassis species. S. sp. THAI was thought to growstrickly on Quercus, and this trait has been kept inthe lineages of S. brevipes, S. spathulata and Asian S.cf. crispa, but they all are known to have conifer hostsas well. European S. crispa and western North Amer-ican S. radicata are found strictly on conifers. Addi-tional ecological and phylogenetic studies could re-solve patterns of host shifts in Sparassis.
ACKNOWLEDGMENTS
We thank R.H. Petersen, D.E. Desjardin, D.H. Pfister, R.J.Bandoni, T. Hattori, P.M. Kirk, G. Ringer, Z.L. Yang, HelmutBesl, and the curators of CUP, FH, HKAS, HMAS, MBUH,BMS, TENN and the herbaria in the University of Regens-burg (REG) for providing specimens, and two reviewers fortheir helpful comments. This work was supported by Na-tional Sciences Foundation Grant DEB-9903835 to DSH andMB.
LITERATURE CITED
Binder M, Hibbett DS. 2002. Higher-level phylogenetic re-lationships of Homobasidiomycetes (mushroom-form-ing fungi) inferred from four rDNA regions. Mol Phy-logenet Evol 22:76–90.
Breitenbach J, Kranzlin F. 1986. Fungi of Switzerland. Vol.2: Nongilled fungi. 412 p.
Burdsall HH Jr, Miller OK Jr. 1988a. Type studies and no-menclatural considerations in the genus Sparassis. My-cotaxon 31:199–206.
. 1988b. Neotypification of Sparassis crispa. Mycotax-on 31:591–593.
Desjardin DE, Wang Z, Binder M, Hibbett DS. 2004. Spa-rassis cystidiosa sp. nov. from Thailand is described us-ing morphological and molecular data. Mycologia 96:1008–1012.
Donk MA. 1964. A conspectus of the families of the Aphyl-lophorales. Persoonia 3:199–324.
Gilbertson RL. 1980. Wood-rotting fungi of North America.Mycologia 72:1–49.
. 1981. North American wood-rotting fungi thatcause brown rots. Mycotaxon 12:372–416.
, Ryvarden L. 1987. North American polypores. Vol.2. Fungiflora, Oslo.
Hawksworth DL, Kirk PM, Sutton BC, Pegler DN. 1995. Ain-sworth & Bisby’s dictionary of the fungi. 8th ed. Wal-lingford, UK: CAB International. 616 p.
Hibbett DS. 1996. Phylogenetic evidence for horizontaltransmission of group I introns in the nuclear ribosom-al DNA of mushroom-forming fungi. Mol Biol Evol 13:903–917.
, Donoghue MJ. 1994. Progress toward a phyloge-netic classification of the Polyporaceae through parsi-mony analysis of mitochondrial ribosomal DNA se-quences. Can J Bot 73 (Suppl. 1):S853–S861.
, Pine EM, Langer E, Langer G, Donoghue MJ. 1997.Evolution of gilled mushroom and puffballs inferredfrom ribosome DNA sequences. Proc Nat Acad Sci USA94:12002–12006.
, Hansen K, Donoghue MJ. 1998. Phylogeny and bio-geography of Lentinula inferred from an expandedrDNA data set. Myc Res 102:1041–1049.
, Donoghue MJ. 2001. Analysis of character correla-tions among wood decay mechanisms, mating system,and substrate ranges in Homobasidiomycetes. Syst Biol50:215–242.
, Binder M. 2002. Evolution of complex fruiting-bodymorphologies in homobasidiomycetes. Proc R SocLond B269:1963–1969.
Hirt RP, Logsdn JM, Healy B, Dorey MW, Doolittle WF, Em-bley TM. 1999. Microsporidia are related to fungi: ev-idence from the largest subunit of RNA polymerase IIand other proteins. Proc Natl Acad Sci, USA 96:580–585.
Huelsenbeck JP, Ronquist F. 2001. MrBayes: Bayesian infer-ence of phylogenetic trees. Bioinformatics 17:754–755.
Imazeki R, Otani Y, Hongo T. 1988. Fungi of Japan. Tokyo,Japan: Yama-kei Publishers Co. Ltd. 623 p.
Julich W. 1981. Higher taxa of basidiomycetes. J Cramer,Vaduz. 485 p.
Kirk PM, Cannon PF, David JC, Stalpers JA. 2001. Ainsworth& Bisby’s dictionary of the fungi. 9th ed. Wallingford,UK: CAB International. 655 p.
Kreisel H. 1983. Zur Taxonomie von Sparassis laminosa Fr.s. l. Fed Report 94:675–682.
Liu YJ, Whelen S, Hall BD. 1999. Phylogenetic relationshipsamong ascomycetes: evidence from a RNA polymeraseII subunit. Mol Biol Evol 16:1799–1808.
Mao XL, Jiang CP, Ciwang OZ. 1993. Economic macrofungi

1029WANG ET AL: SPARASSIS
of Tibet. Beijing: Beijing Science & Technology Press.651 p.
Martin KJ, Gilbertson RL. 1976. Cultural and other mor-phological studies of Sparassis radicata and related spe-cies. Mycologia 68:622–639.
Matheny PB, Liu YJ, Ammirati JF, Hall BD. 2002. UsingRPB1 sequences to improve phylogenetic inferenceamong mushroom (Inocybe, Agaricales). Am J Bot 89:688–698.
Mueller GM, Wu QX, Huang YQ, Guo SY, Aldana-GomezR, Vilgalys R. 2001. Accessing biogeographic relation-ships between North American and Chinese macrofun-gi. Journal of biogeography 28:271–281.
Redhead SA. 1989. A biogeographical overview of the Ca-nadian mushroom flora. Can J Bot 67:3003–3062.
Swofford DL. 1999. PAUP*. Phylogenetic analysis using par-simony (*and other methods). Version 4. Sunderland,Massachusetts: Sinauer Associates.
Teng SC. 1995. Fungi of China. Mycotaxon, LTD.van Zanen GCN. 1988. Sparassis laminosa versus Sparassis
crispa. Coolia 31:93–95.Vilgalys R, Hester M. 1990. Rapid genetic identification and
mapping of enzymatically amplified ribosomal DNAfrom several species of Cryptococcus. J Bacteriol 172:4238–4246.
Weir JR. 1917. Sparassis radicata, an undescribed fungus onthe roots of conifers. Phytopathology 7:166–177.
Wen J. 1999. Evolution of eastern Asia and eastern NorthAmerican disjunct distributions in flowering plants.Ann Rev Ecol Syst 30:421–455.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification anddirect sequencing of fungal ribosomal RNA genes forphylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ,White TJ, eds. PCR protocols. San Diego, California:Academic Press. p 315–322.
Wu QX, Mueller GM. 1997. Biogeographic relationships be-tween the macrofungi of temperate eastern Asia andeastern North America. Can J Bot 75:2108–2116.
, , Lutzoni FM, Huang YQ, Guo SY. 2000. Phy-logenetic and biogeographic relationships of easternAsian and eastern North American disjunct Suillus spe-cies (fungi) as inferred from nuclear ribosomal RNAITS sequences. Mol Phylogenet Evol 17:37–47.
Zang M. 1992. Endemic higher fungi with a note on Chinaand adjacent areas. Acta Bot Yunnanica 14:385–400.