physiological and immunological effects of coal …
TRANSCRIPT

PHYSIOLOGICAL AND IMMUNOLOGICAL EFFECTS OF COAL COMBUSTION
RESIDUES IN THE YELLOW-BELLIED SLIDER
by
DAVID L. HASKINS
(Under the Direction of Tracey D. Tuberville and Robert B. Bringolf)
ABSTRACT
Freshwater turtles are at increased risk for extinction, and although they have been shown
to accumulate large amounts of pollutants, little is known how contaminants affect their immune
status and overall health. Coal combustion residues (CCRs) contain high amounts of potentially
toxic trace elements (i.e. selenium) and are known to cause metabolic aberrations and
histopathological abnormalities in some reptilian species. My research sought to ascertain if trace
elements associated with CCRs negatively impact the health of the yellow-bellied slider
(Trachemys scripta). We performed a field study to examine bioaccumulation of CCRs in wild T.
scripta and quantified immune responses across site types. We also acutely exposed T. scripta to
Se in a controlled lab study, and we measured Se accumulation, mortality, bactericidal capacity,
hematological profiles, and metabolic rates. In the field study we found that wild T. scripta
captured in CCR-affected wetlands did accumulate large amounts of CCRs, but did not exhibit
diminished immune responses. In the lab study we found that T. scripta exposed to Se exhibit
symptoms common in other vertebrates (mortality and altered hematological profiles). Overall,
my results further our understanding of contaminant accumulation in a widespread chelonian
species, and my results suggest that symptoms associated with Se toxicosis can occur in reptiles.
INDEX WORDS: turtle, bioaccumulation, coal combustion residues, immune system, selenium

PHYSIOLOGICAL AND IMMUNOLOGICAL EFFECTS OF COAL COMBUSTION
RESIDUES IN THE YELLOW-BELLIED SLIDER
by
DAVID L. HASKINS
BS, Maryville College, 2014
A thesis submitted to the Graduate Faculty of The University of Georgia in Partial Fulfillment of
Requirements for the Degree
MASTER OF SCIENCE
ATHENS, GEORGIA
2016

© 2016
David L. Haskins
All Rights Reserved

PHYSIOLOGICAL AND IMMUNOLOGICAL EFFECTS OF COAL COMBUSTION
RESIDUES IN THE YELLOW-BELLIED SLIDER
by
DAVID L. HASKINS
Major Professors: Tracey D. Tuberville
Robert B. Bringolf
Committee: Travis C. Glenn
Terry M. Norton
Electronic Version Approved:
Suzanne Barbour
Dean of the Graduate School
The University of Georgia
August 2016

iv
ACKNOWLEDGEMENTS
I would like to first thank Dr. Tracey Tuberville, who brought me onto her project and
gave me positive encouragement over the course of my study. Thank you Dr. Robert Bringolf for
agreeing to serve as my co-advisor, assisting me with contaminant data preparation and analysis,
and for letting me play basketball with your group. I would like to thank Dr. Travis Glenn for the
invaluable advice he gave for our controlled study and his great taste in music. I want to also
thank Dr. Terry Norton for sharing his expertise in chelonian health throughout my studies.
Thank you to all of the individuals who helped me collect data and conduct my research,
including: Bess Harris, Caitlin Kupar, Katrina Woods, Naya Eady, Jarad Cochran, Dan Quinn,
Megan Winzeler, Sam Dean, Nick Bossenbroek, Chris Murphy, and Kevin Fouts. Also, thanks to
Dr. John Seaman and Dr. Brian Jackson for their assistance with our trace element analyses. In
Athens, I would like to especially thank Drs. Richard Chandler and Michael Yabsley for their
assistance in data analysis and parasite quantification. Thank you to Dr. Nicole Stacy for
working with our hematological data for the selenium exposure chapter. This project was funded
by the Department of Energy under award number DE-FC09-07SR22506 to the University of
Georgia Research Foundation and by the Savannah River Nuclear Solutions – Area Completions
Project.
I would like to thank my best friend and partner - LeeAnn Beam. Thank you for listening
to my numerous rants about my studies. Thanks to my friends, especially Austin Coleman,
Kelsey Solomon, Jacob Daly, Jared Green, Nate Tomczyk, Matt Hamilton, Dan Quinn, and
Tyler Carter. Lastly, thank you to my mother and father for always supporting my passions. I
promise I will eventually become a bug doctor.

v
TABLE OF CONTENTS
Page
ACKNOWLEDGMENTS………………………………………………………………………. iv
LIST OF TABLES……………………………………………………………………………… vii
LIST OF FIGURES……………………………………………………………………………... ix
CHAPTER
1 INTRODUCTION AND LITERATURE REVIEW………………………………… 1
Literature Cited……………………………………………………………….… 10
2 ACCUMULATION AND EFFECTS OF COAL COMBUSTION RESIDUES IN
THE YELLOW-BELLIED SLIDER …………………………………………..…... 22
Introduction……………………………………………………………………... 22
Methods…………………………………………………………………………. 25
Results…………………………………………………………………………... 33
Discussion………………………………………………………………………. 37
Literature Cited…………………………………………………………………. 45
3 EFFECTS OF SELENIUM ON THE HEMATOLOGY, INNATE IMMUNITY,
AND METABOLIC RATE OF YELLOW-BELLIED SLIDERS…………………. 67
Introduction……………………………………………………………………... 67
Methods…………………………………………………………………………. 70
Results…………………………………………………………………………... 78
Discussion………………………………………………………………………. 81

vi
Literature Cited………………………………………………………………… 88
4 CONCLUSIONS………………………………………………………………...... 107
Literature Cited………………………………………………………………... 111

vii
LIST OF TABLES
Page
Table 2.1: Trace element concentrations in claw (mg/kg dw) and blood (mg/kg ww) from
yellow-bellied slider turtles (Trachemys scripta scripta) form D-area and reference
wetlands on the Savannah River Site, South Carolina………….………………………. 54
Table 2.2: Results of analysis of covariance of the effects of site type (D-area vs. reference) and
plastron length (PL) on individual trace element concentration in yellow-bellied slider
(Trachemys scripta scripta) claws…………………………………………………....… 55
Table 2.3: Two types of candidate models with varying correlation structure to explain
differences in toe web swelling response between PHA- and PBS-injections…………. 56
Table 2.4: Prevalence (proportion of individuals infected) and parasitemia levels (± 1 SE) of
hemogregarines in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area
and reference areas on the Savannah River Site, South Carolina……............................. 57
Table 3.1: Differences in selenium accumulation in target tissues (liver, kidney, muscle, and
blood) of Trachemys scripta exposed to one of three selenium treatments (Control, 15
mg/kg, and 30 mg/kg). Selenium concentrations are reported as means (dry weight for
tissues and wet weight for blood) ± 1 SE (range)………………………........................... 97
Table 3.2: Proportional survival of yellow-bellied sliders (Trachemys scripta scripta) exposed to
three different treatments (control, 15 mg/kg, and 30 mg/kg) of selenium (Se). Chi-square
values reported are from a Kruskal-Wallis analysis to compare proportional survival of
treatment groups at the end of the study (37 days)……………………………………..... 98

viii
Table 3.3: Linear mixed model for metabolic rate analysis with varying correlation structure to
explain differences in metabolic rate among selenium exposure groups……………...... 99
Table 3.4: Hematological comparisons of Trachemys scripta exposed to one of three selenium
treatments (Control, 15 mg/kg, and 30 mg/kg). Indices include white blood cell estimate
(WBC), heterophil (HET) to lymphocyte (LYM) ratios (H/L), heterophils (HET),
lymphocytes (LYM), monocytes (MON), eosinophils (EOS), basophils (BAS), packed
cell volume (PCV), and total solids (TS)……………………………….......................... 100

ix
LIST OF FIGURES
Page
Figure 2.1: D-area ash basins and their surrounding wetlands on the Savannah River Site in west-
central South Carolina. Historically, effluent containing coal ash would flow into
retention basins (R1 and R2), then as particulates settled, surface waters would move
into primary and secondary basins. Letters ‘A’ and ‘B’ denote wetlands adjacent to the
primary basin that were also sampled…………………………………………………… 58
Figure 2.2: Relationship between plastron length (PL) and rank-transformed Claw Cd values
(analysis of covariance; PL: F1,52 = 18.21, r2 = - 0.45, p < 0.001; Site: F1,52 = 28.48, p <
0.001) in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and
reference sites on the Savannah River Site, South Carolina…………………………….. 59
Figure 2.3: Relationship between plastron length (PL) and rank-transformed Claw Cu values
(analysis of covariance; PL: F1,52 = 21.69, r2 = - 0.37, p < 0.001; Site: F1,52 = 11.60, p =
0.001) in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and
reference sites on the Savannah River Site, South Carolina……………………………. 60
Figure 2.4: Relationship between plastron length (PL) and rank-transformed Claw As values
(analysis of covariance; PL: F1,52 = 4.90, r2 = 0.37, p = 0.030; Site: F1,52 = 29.37, p <
0.001) in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and
reference sites on the Savannah River Site, South Carolina……………………………. 61
Figure 2.5: Spearman rank correlations between log blood and log claw trace element (As, Cr,
Se, and Sr) concentrations in yellow-bellied sliders (Trachemys scripta scripta). Sample

x
sizes for As and Se (n = 24) were lower than Cr and Sr comparisons, and this was due to
detectability issues………………………………………………………....……………. 62
Figure 2.6: Mean percentage of bacteria (Escherichia coli) killed (± 1 SE) by yellow-bellied
slider turtles (Trachemys scripta. scripta) from D-area (n = 39) and reference sites (n =
42) on the Savannah River Site, South Carolina……………………………....………… 63
Figure 2.7: The effects of PHA injection (dashed black line, solid circles) and PBS injection
(solid line, hollow circles) on mean (± SE) web swelling response (mm) in yellow-bellied
sliders (Trachemys scripta) over 24 hours (n = 44)……………….………………....….. 64
Figure 2.8: Relationship between plastron length (PL) and rank-transformed parasitemia values
(analysis of covariance; PL: F1,40 = 21.87, r2 = 0.32, p < 0.001; Site: F1,40 = 0.00, p =
0.987) in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and
reference sites on the Savannah River Site, South Carolina…………………………….. 65
Figure 2.9: Correlation between log PHA response (stimulation index) and parasitemia values in
yellow-bellied slider turtles (Trachemys scripta scripta) collected during 2014-2015 from
the Savannah River Site, South Carolina (PHA responses; p < 0.001, n = 15)…………. 66
Figure 3.1: Spearman rank correlation between blood selenium (Se) and tissue Se concentrations
(A - kidney, B - liver, and C - muscle) in yellow-bellied sliders (Trachemys scripta
scripta) exposed to three different concentrations of selenium (Se; n = 64)………….. 101
Figure 3.2: Oxygen consumption (ml O2/hr) of Trachemys scripta exposed to one of three
selenium treatments (Control = circles, 15 mg/kg = triangles, 30 mg/kg = squares; N = 5,
6, and 8, respectively). Measurements were taken at two separate temperatures (25°C =
black and 30°C = grey)………………………………………………………………... 102
Figure 3.3: Spearman rank correlation between percent packed cell volume and kidney selenium

xi
(Se, dry weight) concentrations in yellow-bellied sliders (Trachemys scripta scripta)
exposed to three different concentrations of selenium (Se; n = 64)….……………….. 103
Figure 3.4: Spearman rank correlation between total solids and blood selenium (Se, wet weight)
concentrations in yellow-bellied sliders (Trachemys scripta scripta) exposed to three
different concentrations of selenium (Se; n = 64)…………………….…………….... 104
Figure 3.5: Mean percent (± 1 SE) bacterial killing ability of yellow-bellied sliders (Trachemys
scripta scripta) exposed to three different treatments of selenium (Se): control (n = 23),
15 mg/kg (n = 23), and 30 mg/kg (n = 18). Escherichia coli was the bacteria species used
in these assays……………………………………………………………….………... 105
Figure 3.6: Mean heterophil (# of cells x 109) count (± 1 SE) of yellow-bellied sliders
(Trachemys scripta scripta) exposed to three different treatments of selenium (Se):
control (n = 23), 15 mg/kg (n = 24), and 30 mg/kg (n = 18)......................................... 106

1
CHAPTER 1
INTRODUCTION AND LITERATURE REVIEW
In present ecosystems, anthropogenic activities have caused contaminants to be
pervasive, especially in aquatic systems. Pollutants can enter aquatic systems through multiple
pathways, including agricultural run-off, assorted mining methods, and man-made disasters
(Dowd et al. 2008; Mishra et al. 2008; Young et al. 2010). Because of large-scale, industrial
disasters recently occurring in the United States, pollution is again becoming an increasing
public concern. Organisms that make contaminated environments their home are chronically
subjected to pollutants, leading to unfavorable biological outcomes. Although anthropogenic
contaminants are ubiquitous, pollution continues to be one of the lesser-studied man-made
stressors affecting herpetological health and biodiversity in ecosystems (Grillitsch and Schiesari
2010). In particular, reptiles are a class of vertebrates widely underrepresented in
ecotoxicological literature (Hopkins 2000; Grillitsch and Schiesari 2010). To help reduce this
sizable knowledge gap in vertebrate ecotoxicology, my thesis work intended to explore the
sublethal effects of coal combustion residues on chelonians.
Trace Elements and Coal Combustion Residues. In the past 40 years, global energy
usage has increased by more than 100%, and it is dominated by the use of oil, gas, and coal
resources (80% of energy consumption; Nejat et al. 2015). Furthermore, of these energy sources,
coal combustion accounts for approximately 30% of all global energy production (British
Petroleum 2015). Coal combustion comprised 40% of all of the United States’ net electricity

2
generation in 2014 (USDOE 2014). The coal combustion process produces copious amounts of
coal combustion residues (CCRs; i.e., coal fly ash and bottom ash) that are either disposed in
landfills or surface impoundments or are reused for construction (ACAA 2008; Ruhl et al. 2012).
CCRs contain high concentrations of trace elements that are known to be toxic to wildlife as well
as humans (Rowe et al. 2002; George et al. 2015). In the United States, over 130 million tons of
CCRs are produced annually, and roughly 40% of CCRs are placed into surface impoundments
for disposal (ACAA 2010).
A major concern of CCR disposal is the contamination of surrounding environments
through the release of trace elements via leaching, improper discharge, or failure of storage
systems (Izquierdo and Querol 2012; Ruhl et al. 2010). The trace elements that are commonly
found in coal fly ash in high concentrations include: arsenic (As), cadmium (Cd), chromium
(Cr), copper (Cu), mercury (Hg), nickel (Ni), lead (Pb), selenium (Se), strontium (Sr), vanadium
(V), and zinc (Zn) (Rowe et al. 2002). Although each has a complex geochemistry, groups of
these elements tend to follow similar patterns in regards to leachability into the environment. The
leachability of trace elements within CCRs largely depend upon the pH of the ash-water system.
For example, the leachability of Cd, Cu, and Sr is diminished under alkaline conditions;
however, alkaline conditions also amplify the release of oxyanionic species of elements
including As, Cr, and Se (Izquierdo and Querol 2012). It is important to note that the pH of ash
systems is not static and tends to move toward neutrality over time regardless of the initial state.
Understanding how these elements disperse from surface impoundments, into the environment,
and finally into biota is an important step in assessing risks of CCR exposure.
Effects of Coal Combustion Residues on Wildlife. Trace elements contaminants
originating from CCR sources are known to cause deleterious effects on a variety of taxa.

3
Previous field and lab studies demonstrate that acute CCR exposure can be lethal in amphipods,
shrimp, Odonates, fish, and amphibians (Magnuson et al. 1980; Guthrie and Cherry 1976; Rowe
et al. 2002). Guthrie and Cherry’s (1976) outdoor mesocosm study placed shrimp, Odonate spp.,
crayfish, many fish species, and salamanders inside of field-cages at CCR-contaminated areas for
5 days. Shrimp, darters, and salamanders all expressed extreme sensitivities to CCRs, while
catfish, crayfish, and largemouth bass had much lower mortality rates (Guthrie and Cherry
1976). Furthermore, subsequent CCR-exposure studies have shown that fish species that survive
acute exposure may still have high mortality rates when chronically exposed (Hopkins 2001).
In many cases, acute or chronic CCR exposure may not cause direct mortality to the
species of interest, but may result in sublethal effects. One of the most researched CCR disposal
sites in the U.S., D-Area on the Savannah River Site (SRS; Aiken, SC), has provided researchers
with evidence of these sublethal effects. One common response across taxa exposed to CCRs is
the disruption of standard metabolic rates (SMRs), which can alter organismal behavior leading
to modifications in energy expenditure, foraging needs, and predation risk (Hopkins et al. 1999).
Grass shrimp (Palaemonetes paludosus), crayfish (Procambarus acutus), bullfrog larvae (Rana
catesbeiana), and banded water snakes (Nerodia fasciata) all exhibited higher SMRs than their
conspecifics who were not exposed to CCRs (Rowe 1998; Rowe et al. 2001; Rowe et al. 1998;
Hopkins et al. 1999).
Increased SMRs in CCR-exposed wildlife has the potential to impact trade-offs related to
the redistribution and reallocation of an animal’s nutrients. Alterations of energy allocation and
distribution due to oxidative stress may lead to disruptions of other basic life processes that
require large amounts of energy. Because mounting an efficient immune response is costly,
diverting resources to deal with sublethal costs of CCR exposure could weaken the immune

4
system (Martin et al. 2010). Trace elements found in high concentrations within CCRs,
specifically As and Se, are well-known for their potential to cause immunotoxic effects in fish
and birds (Fairbrother and Fowles 1990; Fairbrother et al. 1994; Hermann and Kim 2005; Ghosh
et al. 2006; Ghosh et al. 2007). However, studies that deal specifically with CCR exposure and
its potential effects on the immune system are sparse. In Tennessee, researchers observed no
differences in the immune responses of nestling tree swallows (Tachycineta bicolor) affected by
a remediated CCR spill (Beck et al. 2014). Exploring how CCRs may affect vertebrate immunity
could have global implications for wildlife populations residing within or close to CCR-affected
areas.
Selenium: A major CCR contaminant of concern. Coal combustion residues include
many trace elements that potentially can move into the surrounding environment and accumulate
in resident wildlife. Of these elements, selenium (Se) has received much attention due to its
reproductive toxicity in egg-laying vertebrates and potential to cause mass mortality of fish
populations (Young et al. 2010). Selenium is released into the environment through natural and
anthropogenic sources. Seleniferous deposits from ancient marine basins are a global source of
selenium release (Presser et al. 2004). However, there are many sources of anthropogenic release
including mining, the coal combustion process, atmospheric deposits, electronic wastes, and
even agricultural runoff (Rowe et al. 2002; Wen and Carignan 2007; Young et al. 2010; Seiler et
al. 2003). Agricultural runoff becomes a problem in regions where seleniferous soils are
common. Irrigation of these Se-laden sediments can mobilize Se (in the form of selenate) and
lead to widespread deleterious effects to an ecosystem (i.e., the Kesterson Resevoir disaster;
Ohlendorf 1989).

5
When inorganic Se is released into the environment it is readily transformed by primary
producers (i.e., bacterioplankton and phytoplankton) into an organic form (i.e,
selenomethionine). Selenomethionine (SeMet) is more bioavailable and can accumulate within
wildlife based upon their diet, and oviparous vertebrates tend to be the most sensitive to its
toxicity (Chapman et al. 2009; Young et al. 2010). However, selenium’s effects are not limited to
reproductive toxicity, but can also include histopathological abnormalities, physical deformities,
reduction in growth, and oxidative stress (Janz et al. 2010; Young et al. 2010).
While previous studies have quantified Se burdens in reptiles (Hopkins et al. 1999; Nagle
et al. 2001; Roe et al. 2004), little is known about the sublethal effects of Se or SeMet on
reptilian species. Hopkins et al. (2004) exposed brown house snakes (Lamprophis fuliginosus) to
SeMet to observe the effects of trophic and maternal transfer. Similar studies were conducted on
fence lizards (Sceloporous occidentalis) to ascertain if trophic transfer of SeMet could cause
negative effects (Hopkins et al. 2005). Both studies reported no adverse effects of trophic or
maternal transfer of SeMet on survival, growth, or body condition. It is important to note these
reptiles may not have exhibited effects of Se outwardly, negative effects may be revealed in the
form of tissue abnormalities or oxidative stress (Sorensen et al. 1984; Hopkins et al. 1999;
Ganser et al. 2003). Understanding the potential of Se to cause lethal or sublethal effects in
reptilian species is of utmost importance to the preservation of this threatened group.
Pollutants and Parasitism. Immune suppression in wildlife due to pollutants could also
lead to increased susceptibility to certain pathogens and parasites (King et al. 2007; Kim et al.
2008; Martin et al. 2010). Weakened immune responses may facilitate increased parasitism,
negatively affecting the host population (Kim et al. 2008).

6
Haemogregarina spp. are intraerythrocytic, protozoan parasites (Order Eucoccidiorida,
suborder Adeleorina) and are commonly found in reptilian species around the globe (Telford
2009). Although these blood parasites do not typically cause clinical signs of infections, they
may cause abnormalities within the turtles’ blood profiles such as anemia (Knotkova et al. 2005).
Although these parasites are common, little is known of how they may affect their host’s
immune function (Davis et al. 2011). Haemogregarina spp. are transferred to intermediate hosts
such as chelonians via leeches (Placobdella spp; Davies & Johnston 2001).
Previous research suggests that pollutants present in CCRs can affect the abundance and
distribution of invertebrate species like leeches. Suchanek et al. (1995) observed that Placobdella
spp. experienced a significant decline in abundance with increasing sediment concentrations of
Hg. In Brazil, Placobdella bistriata was absent in atrazine-contaminated portions of the
Uberabinha River (Brites and Rantin 2004). The North Carolina Biotic Index (NCBI)
characterizes Placobdella parasitica, a common parasite of freshwater turtles, as moderately
sensitive to contaminants (Lenat 1993). These values are calculated by sampling the species
present in the environment and relating presence to distribution of pollutants. If leech species are
sensitive to CCRs, this could mean that reptiles that inhabit CCR-affected may have a decreased
chance of becoming an intermediate host for parasites (i.e., hemogregarines, trypanosomes, etc.).
Furthermore, if leeches are not affected but the immune system of the intermediate host is
weakened, this could lead higher parasite loads and subsequent health issues.
Study species. The yellow-bellied slider (Trachemys scripta scripta) is a subspecies of
pond slider belonging to the family Emydidae. The T. scripta complex includes three subspecies
(Trachemys scripta scripta, Trachemys scripta elegans, Trachemys scripta troostii) that are all
native to North America. In addition, the Red-eared slider (Trachemys scripta elegans), is

7
commonly utilized as a study organism (Burger et al. 1998, Zimmerman et al. 2010, Yu et al.
2011, Zimmerman et al. 2012). Sliders are semiaquatic turtles that can regularly be found in
wetlands with plentiful aquatic vegetation, basking structures, and nearby cleared areas sufficient
for nesting (Gibbons 1990a, Spotila et al. 1990). Sliders are opportunistic omnivores and are
known to consume a plethora of prey such as vertebrates, invertebrates, and plants (Parmenter &
Avery 1990). Juvenile sliders consume a diet higher in protein (carnivorous diet) compared to
adults who transition to a more omnivorous diet. Sliders are common throughout the United
States and can constitute the highest amount of biomass in some chelonian communities
(Congdon et al. 1986). Sliders exhibit behavior (i.e., fidelity to a small home range, opportunistic
carnivorous feeding) that puts them at risk for contaminant exposure, and sliders also commonly
inhabit bodies of water that could subject them to chronic contaminant exposure (Gibbons
1990b). On the Savannah River Site, previous research has measured CCR concentrations in T.
scripta, but the sample sizes were small and sublethal effects were not the focus of the study
(Nagle et al. 2001).
Turtles as Environmental Sentinels. Freshwater turtles in the Southeast represent a
large amount of biomass in their respective ecosystems, and they can be exposed to these
contaminants through several routes (e.g., dietary, dermal, maternal transfer) (Congdon et al.
1986, Nagle et al. 2001, Ernst & Lovich 2009, Van Dyke et al. 2014). Furthermore, these turtles
occupy the middle to upper-level positions in the food chain, have long lifespans. and many have
small home ranges (Gibbons 1990a, Ernst & Lovich 2009). These life-history characteristics
make freshwater turtle species excellent environmental sentinels (Rowe et al. 2008; Steen et al.
2015). Contaminants may be playing a significant role in the decreasing turtle populations and

8
more research is needed to understand lethal and sublethal effects of contaminant exposure
(Gibbons et al. 2000; Yu et al. 2013).
Turtles, Pollutants, and Humans. The accumulation of pollutants by freshwater turtle
species puts them at risk of developing health issues; however, another large concern of exposure
is humans that consume them (Green et al. 2010). Many contaminants found within CCRs (i.e.,
As, Hg, and Se) readily accumulate within turtle species and can be detrimental to human health
(Nagle et al. 2001; George et al. 2015). Although trace elements within CCRs may pose a human
health risk (George et al. 2015), CCR wastes in the United States are not classified as hazardous
waste (US EPA 2015).
As mentioned previously, a large amount of CCRs are produced in the coal combustion
process and are typically disposed of in surface impoundments (aquatic basins) or landfills.
According to Ruhl et al. (2012), more than 1000 aquatic coal ash depositories exist in the United
States. Aquatic settling basins used to store CCRs typically produce habitats that are inviting to
various species of wildlife (Rowe et al. 2002; Bryan et al. 2011). For example, on the SRS,
freshwater turtle species are common inhabitants of aquatic settling basins contaminated with
CCRs (Nagle et al. 2001; Haskins personal observation). Indeed, if these animals spend
considerable amounts of time within these basins, they are likely to accumulate large amounts of
harmful trace elements.
Turtles are a common source of food in many parts of the world, and historically were a
popular food item in the United States. Globally, more than 10 million turtles are purchased
annually for consumption and these trade systems are not regulated by any government agency
(Van Dijk et al. 2000; Green et al. 2010). If trends of global energy consumption through coal
combustion continue, it is increasingly possible that individuals that consume turtle will be at

9
risk for exposure to CCRs. According to the US EPA (2003), only four states have issued
consumption advisories for freshwater turtles: Arizona, Massachusetts, Minnesota, and New
York. Hopkins (2012) found consumption advisories for snapping turtle consumption in
Pennsylvania and Ohio. Risks of human exposure to contaminants through turtle consumption
will undoubtedly be exacerbated in areas where consumers lack access to proper consumption
guidelines and public education.
Evaluation of the Physiological and Immunological Effects of Coal Combustion
Residues and Selenium in Chelonians. Overall, my MS thesis research had two overarching
goals. My first chapter aimed to investigate the accumulation and sublethal effects of chronic
exposure to CCRs in wild-caught yellow-bellied sliders (Trachemys scripta scripta). This was
accomplished through non-destructive tissue sampling (blood and claws), ecoimmunological
assays, and parasite quantification. I hypothesized that turtles from the CCR-affected site would
exhibit higher CCR burdens, impaired immunological responses, and lower parasite loads. My
second chapter aimed to evaluate the lethal and sublethal impacts of acute selenium (Se)
exposure on T. s. scripta via a laboratory experiment. I predicted that turtles experimentally
exposed to higher doses of Se would exhibit altered hematological profiles (indicative of
immunosuppression), weaker immune responses, and depressed metabolic rates.

10
Literature Cited
ACAA [American Coal Ash Association]. 2008 Coal combustion product (CCP) production and
use report. 2008. <https://www.acaa-
usa.org/Portals/9/Files/PDFs/2008_ACAA_CCP_Survey_Report_FINAL_100509.pdf>.
Accessed on 16 Feb 2016.
ACAA [American Coal Ash Association]. 2010 Coal combustion product (CCP) production and
use report. 2010. <https://www.acaa-
usa.org/Portals/9/Files/PDFs/2010_CCP_Survey_FINAL_102011.pdf>. Accessed on 16
Feb 2016.
Beck, M.L., W.A. Hopkins, J.J. Hallagan, B.P. Jackson, D.M. Hawley. 2014. Exposure to
residual concentrations of elements from a remediated coal fly ash spill does not
adversely influence stress and immune responses of nestling tree swallows. Conservation
Physiology 2:1-13.
Brites, V.L., and F.T. Rantin. 2004. The influence of agricultural and urban contamination on
leech infestation of freshwater turtles, Phyrnops geoffroanus, taken from two areas of the
Uberabinha river. Environmental Monitoring and Assessment 96:273-281.
British Petroleum, 2015. BP Statistical Review of World Energy June 2015. British Petroleum,
pp. 48. < http://www.bp.com/en/global/corporate/energy-economics/statistical-review-of-
world-energy.html>. Accessed on 1 Feb 2016.

11
Bryan Jr., A.L., W.A. Hopkins, J.H. Parikh, B.P. Jackson, and J.M. Unrine. Coal fly ash basins
as an attractive nuisance to birds: parental provisioning exposes nestlings to harmful trace
elements. Environmental Pollution 161:170-177.
Buhlmann, K.A., T.S.B. Akre, J.B. Iverson, D. Karapatakis, R.A. Mittermeier, A. Georges,
A.G.J. Rhodin, P.P. van Dijk, and J.W. Gibbons. 2009. A global analysis of tortoise and
freshwater turtle distributions with identification of priority conservation areas.
Chelonian Conservation and Biology 8:116-149.
Burger, J., C. Carruth-Hinchey, J. Ondroff, M. McMahon, J.W. Gibbons, and M. Gochfeld.
1998. Effects of lead on behavior, growth, and survival of hatchling slider turtles. Journal
of Toxicology and Environmental Health. 55:495-502.
Chapman, P.M., W.J. Adams, M.L. Brooks, C.G. Delos, S.N. Luoma, W.A. Maher, H.M.
Ohlendorf, T.S. Presser, D.P. Shaw. 2009. Ecological assessment of selenium in the
aquatic environment: Summary of a SETAC Pellston Workshop. Pensacola, Florida
(USA): Society of Environmental Toxicology and Chemistry (SETAC).
Congdon, J.D., J.L. Greene, and J.W. Gibbons. 1986. Biomass of freshwater turtles: a geographic
comparison. American Midland Naturalist. 115:165-173.
Davies, A.J., and M.R.L. Johnston. 2001. The biology of some intraerythrocytic parasites of
fishes, amphibian, and reptiles. Advances in Parasitology. 45:1-107.
Davis, A.K., and S.C. Sterrett. 2011. Prevalence of haemogregarine parasites in three freshwater
turtle species in a population in Northeast Georgia, USA. International Journal of
Zoological Research. 7:156-163.

12
Dowd B.M., Press D., Huertos M.L. 2008. Agricultural nonpoint source pollution policy:
The case of California’s central coast. Agriculture, Ecosystems, and Environment
128:151-161.
Ernst, C.H., and J.E. Lovich. 2009. Turtles of the United States and Canada. John Hopkins
University Press, Baltimore, Maryland, USA.
Fairbrother, A., and J. Fowles. 1990. Subchronic effects of sodium selenite and selenomethionine
on several immune functions in mallards. Archives of Environmental Contamination and
Toxicology. 19:836-844.
Fairbrother, A., M. Fix, T. O’Hara, and C. Ribic. 1994. Impairment of growth and immune
function of avocet chicks from sites with elevated selenium, arsenic, and boron. Journal
of Wildlife Diseases 30:222-233.
Ganser, L.R., W.A. Hopkins, L. O’Neil, S. Hasse, J.H. Roe, and D.M. Sever. Liver
histopathology of the Southern Watersnake, Nerodia fasciata fasciata, following chronic
exposure to trace element-contaminated prey from a coal ash disposal site. Journal of
Herpetology 37:219-226.
George, J., R.E. Masto, L.C. Ram, T.B. Das, T.K. Rout, and M. Mohan. 2015. Human exposure
risks for metals in soil near a coal-fired power-generating plant. Archives of
Environmental Contamination and Toxicology 68:451-461.
Ghosh, D., S. Bhattacharya, S. Mazumder. 2006. Perturbations in the catfish immune responses
by arsenic: organ and cell specific effects. Comparative Biochemistry and Physiology
143:455-463.

13
Ghosh, D., S. Datta, S. Bhattacharya, and S. Mazumder. 2007. Long-term exposure to arsenic
affects head kidney and impairs humoral immune responses of Clarias batrachus. Aquatic
Toxicology 82:79-89.
Gibbons, J.W. 1990a. The Slider Turtle. Pages 1-18 in Sheffield, R., editor. Life History and
Ecology of the Slider Turtle. Smithsonian Institution Press, Washington, D.C., USA.
Print.
Gibbons J.W. 1990b. Turtle Studies at SREL: A research perspective. Pages 19-45 in Sheffield,
R., editor. Life History and Ecology of the Slider Turtle. Smithsonian Institution Press,
Washington, D.C., USA. Print.
Gibbons, J.W., D.E. Scott, T.J. Ryan, K.A. Buhlmann, T.D. Tuberville, B.S. Metts, J.L. Greene,
T. Mills, M. Leiden, S. Poppy, and C.T. Winne. 2000. The global decline of reptiles, Déjà
vu amphibians. Bioscience 50:653-666.
Green, A.D., K.A. Buhlmann, C. Hagen, C. Romanek, and J.W. Gibbons. 2010. Mercury
contamination in turtles and implications for human health. Journal of Environmental
Health. 72:14-22.
Grillitsch, B., and L. Schiesari. 2010. The ecotoxicology of metals in reptiles. Pages 337-448 in:
Sparling, D.W.,G. Linder, A.C. Bishop, S. Krest, editors. Ecotoxicology of Amphibians
and Reptiles. Pensacola, FL: Society of Environmental Toxicology and Chemistry
(SETAC).
Guthrie, R.K., and D.S. Cherry. Pollutant removal from coal-ash basin effluent. Water Resources
Bulletin 12:889-902.

14
Harden, L., S. Price, M. Dorcas. Terrestrial activity and habitat selection of eastern mud turtles
(Kinosternon subrubrum) in a fragmented landscape: implications for habitat
management of golf courses and other suburban environments. Copeia 2009: 78-84.
Hermann, A.C., and C.H. Kim. 2005. Effects of arsenic on Zebrafish innate immune system.
Marine Biotechnology 7:494-505.
Hopkins, W.A. 2000. Reptile toxicology: Challenges and opportunities on the last frontier
in vertebrate ecotoxicology. Environmental Toxicology and Chemistry. 19:2391-2393.
Hopkins, B.C. 2012. Mercury bioaccumulation and adverse reproductive effects in snapping
turtles inhabiting a historically contaminated river. Master’s thesis, Virginia Polytechnic
Institute and State University, Blacksburg, Virginia, USA.
Hopkins, W.A., C.L. Rowe, and J.D. Congdon. 1999. Elevated trace element concentrations and
standard metabolic rates in banded water snakes (Nerodia fasciata) exposed to coal
combustion wastes. Environmental Toxicology and Chemistry 18:1258-1263.
Hopkins, W.A. 2001. Effects of coal combustion wastes on survival, physiology, and
performance of the benthic-feeding fish, Erimyzon sucetta. PhD Dissertation, University
of South Carolina, Columbia, South Carolina, USA.
Hopkins, W.A., B.P. Staub, J.A. Baionno, B.P. Jackson, J.H. Roe, and N.B. Ford. 2004. Trophic
and maternal transfer of selenium in brown house snakes (Lamprophis fuliginosus).
Ecotoxicology and Environmental Safety 58:285-293.
Hopkins, W.A., B.P. Staub, and J.A. Baionno. 2005. Transfer of selenium from prey to predators
in a simulated terrestrial food chain. Environmental Pollution 134:447-456.
Izquierdo, M., and X. Querol. 2012. Leaching behaviour of elements from coal combustion fly
ash: an overview. International Journal of Coal Geology 94:54-66.

15
Janz, D.M., D.K. DeForest, M.L. Brooks, P.M. Chapman, G. Gilron, D. Hoff, W.A. Hopkins,
D.O. McIntyre, C.A. Mebane, V.P. Palace, J.P. Skorupa, and M. Wayland. 2010.
Selenium toxicity to aquatic organisms. Pages 139-230 in Chapman, P.M., W.J. Adams,
M.L. Brooks, C.G. Delos, S.N. Luoma, W. Maher, H.M. Ohlendorf, T.S. Presser, and P.
Bradshaw, editors. Ecological Assessment of Selenium in the Aquatic Environment.
Pensacola, Florida: Society of Environmental Toxicology and Chemistry.
Kim, Y., E.N. Powell, T.L. Wade, and B.J. Presley. 2008. Relationships of parasites and
pathologies to contaminant body burdens in sentinel bivalves: NOAA Status and trends
‘mussel watch’ program. Marine Environmental Research. 65:101-127.
King, K.C., J.D. McLaughlin, A.D. Gendron, B.D. Pauli, P.I. Giroux, B. Rondeau, M. Boily, P.
Juneau, and D.J. Marcogliese. 2007. Impacts of agriculture on the parasite communities
of northern leopard frogs (Rana pipiens) in southern Quebec, Canada. Parasitology.
134:2063-2080.
Knotkova, Z., S. Mazanek, M. Hovorka, M. Sloboda, and Z. Knotek. 2005. Haematology and
plasma chemistry of Bornean river turtles suffering from shell necrosis and
haemogregarine parasites. Veterinary Medicine Czech. 50:421-426.
Lenat, D.R. 1993. A biotic index for the southeastern United States: derivation and list of
tolerant values criteria for assigning water-quality ratings. Journal of North American
Benthological Society 12:279-290.
Magnuson, J.J., A.M. Forbes, D.M. Harrell, and J.D. Schwarzmeier. 1980. Responses of stream
invertebrates to an ashpit effluent, Wisconsin power plant impact study. Report EPA-
600/3-80-081. U.S. Environmental Protection Agency, Washington, D.C.

16
Martin, L.B., W.A. Hopkins, L.D. Mydlarz, and J.R. Rohr. 2010. The effects of anthropogenic
global changes on immune functions and disease resistance. Annals of the New York
Academy of Sciences 1195:129-148.
Mishra, V.K., A.R. Upadphyaya, S.K. Pandey, and B.D. Tripathi. 2008. Heavy metal pollution
induced due to coal mining effluent on surrounding aquatic ecosystem and its
management through naturally occurring aquatic macrophytes. Biosource Technology
99:930-930.
Nagle, R.D., C.L. Rowe, and J.D. Congdon. 2001. Accumulation and selective maternal
transfer of contaminants in the turtle Trachemys scripta associated with coal ash
deposition. Archives of Environmental Contamination and Toxicology 40:531-536.
Nejat, P., F. Jomehzadeh, M.M. Taheri, M. Gohari, and M.Z.A. Majid. 2015. A global review of
energy consumption, CO2 emissions and policy in the residential sector (with an
overview of the top ten CO2 emitting countries). Renewable and Sustainable Energy
Reviews 43:843-862.
Ohlendorf, H.M. 1989. Bioaccumulation and effects of selenium in wildlife. Pages 15-24 in
Jacobs, L.W., editor. Special Publication: Selenium in Agriculture and the Environment.
Soil Science Society of America, Madison, Wisconsin, USA.
Parmenter, R.R., and H.W. Avery. 1990. The feeding ecology of the slider turtle. Pages 257-266
in Sheffield, R., editor. Life History and Ecology of the Slider Turtle. Smithsonian
Institution Press, Washington, D.C., USA.

17
Presser, T.S., D.Z. Piper, K.J. Bird, J.P. Skorupa, S.J. Hamilton, S.J. Detwiler, and M.A.
Huebner. 2004. The Phosphoria formation: a model for forecasting global selenium
sources in the environment. Pages 299-319 in Hein, J.R., editor. Handbook of
Exploration and Environmental Chemistry, Vol 8.
Roe, J.H., W.A. Hopkins, J.A. Baionno, B.P. Staub, C.L. Rowe, and B.P. Jackson. 2004.
Maternal transfer of selenium in Alligator mississippiensis nesting downstream of a coal-
burning power plant. Environmental Toxicology and Chemistry 23:1969-1972.
Rowe, C.L., O.M. Kinney, R.D. Nagle and J.D. Congdon. 1998. Elevated maintenance costs in
an anuran (Rana catesbeiana) exposed to a mixture of trace elements during the
embryonic and early larval periods. Physiological Zoology 71:27-35.
Rowe, C.L. 2008. The calamity of so long life: life histories, contaminants, and
potential emerging threats to long-lived vertebrates. Bioscience. 58:623-631.
Rowe, C.L. 1998. Elevated standard metabolic rate in a freshwater shrimp (Palaemonetes
paludosus) exposed to trace element-rich coal combustion waste. Comparative
Biochemistry and Physiology 121:299-304.
Rowe, C.L., W.A. Hopkins, C. Zehnder, and J.D. Congdon. 2001. Metabolic costs incurred by
crayfish (Procambarus acutus) in a trace element-polluted habitat: further evidence of
similar responses among diverse taxonomic groups. Comparative Biochemistry and
Physiology 129:275-283.
Rowe, C.L., W.A. Hopkins, and J.D. Congdon. 2002. Ecotoxicological implications of aquatic
disposal of coal combustion residues in the United States: A review. Environmental
Monitoring and Assessment 80:207-276.

18
Ruhl, L., A. Vengosh, G.S. Dwyer, H. Hsu-Kim, and A. Deonarine. 2010. Environmental
impacts of the coal ash spill in Kingston, Tennessee: an 18-month survey. Environmental
Science and Technology 44:9272-9278.
Ruhl, L., A. Vengosh, G.S. Dwyer, H. Hsu-Kim, G. Schwartz, A. Romanski, and S.D. Smith.
2012. The impact of coal combustion residue effluent on water resources: a North
Carolina example. Environmental Science and Technology 46:12226-12233.
Seiler, R.L., J.P. Skorupa, D.L. Naftz, B.T. Nolan. 2003. Irrigation-induced contamination of
water, sediment, and biota in the western United States – synthesis of data from the
National Irrigation Water Quality Program. Professional Paper #1655, Denver (Colorado,
USA): National Irrigation Water Quality Program, U.S. Geological Survey.
Sorensen, E.M.B., P.M. Cumbie, T.L. Bauer, J.S. Bell, and C.W. Harlan. 1984.
Histopathological, hematological, condition-factor, and organ weight changes associated
with selenium accumulation in fish from Belews Lake, North Carolina. Archives of
Environmental Contamination and Toxicology 13:153-162.
Spotila, J.R., R.E. Foley, and E.A. Standora. 1990. Thermoregulation and climate space of the
slider turtle. Pages 288-298 in Sheffield, R., editor. Life History and Ecology of the
Slider Turtle. Smithsonian Institution Press, Washington, D.C., USA. Print.
Steen, D.A., J.U. Van Dyke, B.P. Jackson, and W.A. Hopkins. 2015. Reproduction and hatchling
performance in freshwater turtles associated with a remediated coal fly-ash spill.
Environmental Research. 138:38-48.

19
Suchanek, T.H., P.J. Richerson, L.J. Holts, B.A. Lamphere, C.E. Woodmansee, D.G. Slotton,
E.J. Harner, and L.A. Woodward. Impacts of mercury on benthic invertebrate populations
and communities within the aquatic ecosystem of Clear Lake, California. Water, Air, and
Soil Pollution 80:951-960.
Suchanek, T.H., C.A. Eagles-Smith, D.G. Slotton, E.J. Harner, D.P. Adam, A.E. Colwell,
N.L. Anderson, and D.L. Woodward. 2008. Mine-derived mercury: Effects on lower
trophic species in Clear Lake, California. Ecological Applications. 18:A158-176.
Telfod SR. 2009. Hemoparasites of the Reptilia: Color Atlas and Color Text. CRC Press,
New York, USA.
U.S. Department of Energy. 2015. U.S. Energy Information Administration: Net generation by
energy source – Total.
<https://www.eia.gov/electricity/monthly/epm_table_grapher.cfm?t=epmt_1_1>
Accessed 1 Feb 2016.
U.S. Environmental Protection Agency. 2003. Fact Sheet – Update: National Listing of Fish and
Wildlife Advisories. Report EPA-823-F-03-003.
<http://limnology.wisc.edu/courses/zoo510/jvz%20readings/factsheet.pdf> Accessed 1
Feb 2016.
U.S. Environmental Protection Agency. 2015. Wastes – Non-Hazardous Waste – Industrial
Waste: Coal ash surface impoundment assessments.
<http://www3.epa.gov/epawaste/nonhaz/industrial/special/fossil/coalashletter.htm>
Accessed 1 Feb 2016.

20
Van Dijk, P.P., D.T. Iskandar, T. Palasuwan, A.G.J. Rhodin, Samedi, D.S.K. Sharma, C.R.
Shepherd, O.B. Tisen, and V.R. Genorupa. 2000. Turtle trade in southeast Asia: Regional
summary (Indonesia, Malaysia, Papua New Guinea, and Thailand). Pages 2:145-147 in
Van Dijk, P.P., B.L. Stuart, and A.G.J. Rhodin, editors. Asian Turtle Trade: Proceedings
of a Workshop on Conservation and Trade of Freshwater Turtles and Tortoises in Asia.
Chelonian Research Monographs.
Van Dyke, J.U., D.A. Steen, B.P. Jackson, and W.A. Hopkins. 2014. Maternal transfer and
embryonic assimilation of trace elements in freshwater turtles after remediation of coal
fly ash spill. Environmental Pollution 194:38-49.
Wen, H., and J. Carignan. 2007. Reviews on atmospheric selenium: emissions, speciation and
fate. Atmospheric Environment 41:7151-7165.
Young, T., K. Finley, W. Adams, J. Besser, W.D. Hopkins, D.F. Jolley, E. McNaughton, T.S.
Presser, D. Shaw, and J. Unrine. 2010. What You Need to Know about Selenium. Pages
7-45 in Chapman, P.M., W.J. Adams, M.L. Brooks, C.G. Delos, S.N. Luoma, W. Maher,
H.M. Ohlendorf, T.S. Presser, and P. Bradshaw, editors. Ecological Assessment of
Selenium in the Aquatic Environment. Pensacola, Florida (USA): Society of
Environmental Toxicology and Chemistry.
Yu, S., R.S. Halbrook, and D.W. Sparling. 2011. Metal accumulation and evaluation of effects in
a freshwater turtle. Ecotoxicology 20:1801-1812.
Yu, S., R.S. Halbrook, and D.W. Sparling. 2013. Correlation between heavy metals and turtle
abundance in ponds near the Paducah Gaseous Diffusion Plant, Kentucky, USA.
Archives of Environmental Contamination and Toxicology 65:555-566.

21
Zimmerman, L.M., R.T. Paitz, S.G. Clairardin, L.A. Vogel, and R.M. Bowden. 2010. Variation
in seasonal patterns of innate and adaptive immunity in the red-eared slider (Trachemys
scripta). The Journal of Experimental Biology 213:1477-1483.
Zimmerman, L.M., R.T. Paitz, S.G. Clairardin, L.A. Vogel, and R.M. Bowden. 2012. No
evidence that estrogens affect the development of the immune system in the red-eared
slider turtle (Trachemys scripta elegans). Hormones and Behavior. 62:331-336

22
CHAPTER 2
ACCUMULATION AND EFFECTS OF COAL COMBUSTION RESIDUES IN THE
YELLOW-BELLIED SLIDER
Introduction
Turtles are a group of reptiles whose life history traits make them particularly useful as
environmental sentinels (Meyers-Schöne and Walton 1994; Aguirre and Lutz 2004; Van Dyke et
al. 2013). Many turtle species are of high trophic status, are long-lived, typically have small
home ranges, and can represent a considerable portion of biomass within their respective
ecosystems (Rowe et al. 2008; Steen et al. 2015; Congdon et al. 1986). Furthermore, because
they inhabit a variety of environments, turtle species can serve as indicators of contamination in
terrestrial, freshwater, and marine environments. For instance, previous studies have documented
box turtles (Terrapene carolina), slider turtles (Trachemys scripta), and sea turtles (Chelonia
mydas) with high amounts of pollutants (Beresford et al. 1981; Yu et al. 2011; Faust et al. 2014)
compared to what would be expected in more pristine reference environments.
Although many previous studies have sought to determine the usefulness of reptiles as
bioindicators (Beresford et al. 1981; Yu et al. 2011; Faust et al. 2014), relatively few have
attempted to explore the sublethal effects of contaminants in this threatened group (Grillitsch and
Schiesari 2010). Indeed, chronic sublethal exposure can be just as harmful as lethal exposure to
an individual’s overall fitness and survival (Newman 2014). This is especially concerning due to
the current status of many reptilian species around the globe. For example, the 2013 IUCN Red
List states that of 40% of all turtle species (134/335 species) have some form of conservation

23
status (vunerable, endangered, critically endangered; Turtle Taxonomy Working Group 2014).
Turtles are known to accumulate contaminants through diet and maternal transfer (Rowe et al.
2008; Van Dyke et al. 2014). Even a modest increase in adult mortality can have impacts on
freshwater turtle populations; thus, the possible effects of pollutants and other anthropogenic
stressors (i.e., habitat alteration, poaching) are troubling for freshwater turtle populations
(Harden et al. 2009). Therefore, it is of utmost importance that future research seeks to
understand the toxicological effects of pollutants on turtle species.
Previous work has documented sublethal effects of pollutants on turtles. In marine
species, it has been suggested that organic pollutants (i.e., organochlorine pesticides,
polychlorinated biphenyls) can negatively impact sea turtle kidney and salt glands, metabolic
status, and hematological parameters (Camacho et al. 2013). Research on declining populations
of the western pond turtle (Emys mamorata) in California revealed that mercury exposure can
alter plasma thyroid hormones (Meyer et al. 2014). Nagle et al. (2001) measured coal
combustion residues (CCRs) in yellow-bellied sliders (Trachemys scripta) and their eggs
originating from a coal disposal facility on the Savannah River Site (D-area on the SRS; Aiken,
SC). Turtles from D-area contained elevated CCR burdens and they also maternally transferred
selenium (Se; a CCR trace element of concern) to eggs and hatchlings. The hatchling T. scripta
of contaminated mothers in this study also exhibited lower standard metabolic rates compared to
conspecifics from reference mothers.
Coal combustion residues (CCRs) are common waste products around the globe, are
produced in large quantities, and contain high levels of potentially toxic trace elements (Rowe et
al. 2002). The United States now produces over 130 million tons of CCRs per year, and prior to
the 1980, approximately 66% of CCRs were placed into aquatic settling basins (Rowe et al.

24
2002). As of the 2010, 40% of these wastes were still placed into aquatic settling basins (ACAA
2010). These aquatic basins typically are attractive to many species of wildlife. Wildlife may use
these basins to forage and reproduce, placing them at risk of exposure to potentially toxic levels
of trace elements (Bryan et al. 2012; Lemly and Skorupa 2012).
In spite of the recent advances of reptilian ecotoxicology, it is relatively unknown what
sublethal effects can be caused by chronic exposure to trace element contaminants. Thus, this
study explored the sublethal effects of trace elements on freshwater turtles. The D-area on the
SRS, is one of the most studied coal ash disposal facilities in the United States (Rowe et al.
2002). Previous research at D-area demonstrated that exposure to trace element contaminants of
CCR origin can cause lethal and sublethal effects in biota (Guthrie and Cherry 1976), including
metabolic aberrations in exposed individuals (Rowe 1998; Hopkins et al. 1999; Rowe et al.
2001). Trace elements in CCRs such as arsenic (As) and Se are known to cause immunotoxic
effects in other vertebrates (Ghosh et al. 2006; Fairbrother et al. 1994). Because the immune
system uses copious amounts of energy, disruption of metabolic processes and energy allocation
due to CCR exposure could lead to compromised immune responses or the ability to regulate
parasite loads (Martin et al. 2010). Immune function can also be negatively impacted by parasite
loads in vertebrates (Graham 2002; Martin et al. 2010). Furthermore, little is known overall
regarding how trace element contaminants could negatively impact the reptilian immune system,
and studies assessing the toxic risk of these contaminants in reptiles have only started in the last
two decades (Keller et al. 2006). The reptilian immune system exhibits many similarities to the
immune system of endotherms. Reptiles have all three branches of immunity that are present in
mammals including: innate, cell-mediated, and humoral immune responses (Keller et al. 2006).
Because recent research has suggested that red-eared sliders (Trachemys scripta elegans) rely

25
more heavily on innate immune responses rather than adaptive responses (Zimmerman 2013),
this current study sought to investigate the potential effects of CCR trace elements on the innate
immune responses of T. scripta. The overall goal of this study was to examine the
immunological costs of exposure to trace elements associated with coal combustion in wild
yellow-bellied sliders (Trachemys scripta). Our specific objectives were to: (1) quantify the
accumulation of trace elements (As, Cd, Cr, Cu, Se, Sr) associated with CCRs in tissues
corresponding to different time scales (blood and claw); (2) compare innate immunological
responses of sliders inhabiting CCR-affected wetlands and reference wetlands; and (3) determine
if there are differences in parasite burdens based on capture location.
Methods
Study site
Our study was conducted on the Savannah River Site (SRS) in west-central, South Carolina,
USA. The SRS is an 800-km2 Department of Energy facility located in the southeastern Coastal
Plain and supports a variety of wetland types, including isolated wetlands (Carolina bays), farm
ponds, streams, and bottomland hardwood swamps. A small proportion of these wetlands have
been impacted by site operations (e.g., the production of nuclear materials, power production),
resulting in wetlands with varying contaminant histories. One of the major industrial sources of
contaminants on the SRS and globally is CCRs (Nagle et al. 2001; Lemly and Skorupa 2012). D-
area is one of several coal combustion facilities on the SRS and it is one of the most studied coal
disposal sites in the United States (Rowe et al. 2002). When D-area was actively burning coal,
CCRs were placed into surface impoundments (aquatic settling basins). Coal fly ash would be
discharged from the power plant into receiving basins, then into large primary and secondary

26
basins (Figure 2.1), with particulates separating and settling out as water moved through the
system. Because they offer a permanent water source and become vegetated over time, the basins
can become attractive nuisances (Bryan et al. 2012). Impoundments often attract wildlife because
the habitat offered (i.e., vegetated edges, ample water sources) provides resources necessary for
reproduction and overall survival (Rowe et al. 2002). Turtles and other wildlife have been known
to inhabit coal ash disposal areas and surrounding wetlands (Nagle et al. 2001; Bryan et al.
2012). D-area has several natural wetlands that are within close proximity (< 0.1 km) of the
settling basins. These natural wetlands may receive run-off from the settling basins and are
frequented by animals that may use the settling basins. Thus, for our study D-area animals were
collected from the primary settling basin and from wetlands ‘A’ and ‘B’ (see Figure 2.1).
Reference animals were collected from 10 natural wetlands (encompassing 6 aquatic systems) on
the SRS that have not historically received CCR-effluents and that were 2.4 - 24 km away from
D-area.
Study species
The yellow-bellied slider (Trachemys scripta scripta), is considered the most abundant
freshwater turtle species at SRS (an estimated 61 individuals/ha of aquatic habitat) and accounts
for almost 50% of all freshwater turtle biomass on the site (Congdon et al. 1986). Trachemys
scripta is common throughout southeastern United States and is considered to be a habitat
generalist (Gibbons 1990). Sliders tend to have small home ranges (although they may move
long distances over their lifetime; Morreale et al. 1984), high site fidelity, are long-lived (20-30
years in the wild), and feed in middle to high trophic levels (Gibbons 1990; Tucker 2001;
Parmenter and Avery 1990). These life history characteristics and their wide geographic

27
distribution (North America to South America; Gibbons 1990) make them an excellent receptor
species for contaminant studies. Sliders also experience an ontogenetic diet shift – juveniles tend
to be carnivorous but become more omnivorous as adults (Parmenter & Avery 1990). These diet
shifts may impact dietary exposure and accumulation of contaminants in juvenile sliders relative
to adults. Previous research on the SRS documented accumulation of CCR trace elements and
maternal transfer of Se in T. scripta (Nagle et al. 2001).
Field sampling and sample collection
Trachemys scripta were captured at D-Area wetlands and reference wetlands during
May-September 2014 and May-July 2015 with hoop nets baited with sardines or creamed corn.
Turtles were transported to the Savannah River Ecology Laboratory (SREL) for processing.
Morphological data including plastron length (PL; to nearest 1 mm), weight (to nearest 2 g), and
sex was collected for each individual. Each turtle was individually marked with a permanent ID
by notching or drilling unique combinations of scutes to ensure that individuals were only
sampled once per trapping season for analysis (Cagle 1939; Gibbons 1990b). Whole blood and
claws were collected to provide a short and long-term measurement of contaminant exposure,
respectively (Aresco 2005; Van Dyke et al. 2013). Tips of claws (2-3 mm) from the left forelimb
were collected from each turtle and stored at -60 °C until analysis. Blood samples totaling no
more than 1% of the animal’s body weight, generally <1-1.5 mL, were collected via the
subcarapacial venipuncture site with a 27 or 25-gauge needle (Hernandez-Divers et al. 2002)
Needles used for blood collection were heparinized if needed. After setting aside half of each
whole blood sample for storage at -60 °C for subsequent trace element analysis, a drop of whole
blood was also used to make blood smears, which were fixed with 100% methanol (VWR

28
International, Radnor, PA) for Haemogregarina parasite quantification. Blood smears were
collected opportunistically in 2014 and for all turtles in 2015.
The remaining blood sample was then aliquoted into lithium heparin tubes (Becton
Dickson, San Antonio, TX, USA) or SealRite© 1.5 mL microcentrifuge tubes (USA Scientific
Inc., Ocala, FL, USA) and centrifuged (Heathrow Scientific LLC, Vernon Hills, IL, USA) at
6,000 RPM to collect plasma for the bactericidal assay. Plasma samples were stored at -60°C
until further analysis. Finally, phytohaemagglutinin (PHA) skin assays were performed on
individuals with least 100 mm PL.
Trace Element Analysis: Blood
Blood samples were digested in 2.5 mL of trace metal grade nitric acid (70% HNO3)
using microwave digestion (MarsExpress, CEM Corp., Matthews, NC). Once digestion was
complete, each sample was brought to a final volume of 10 mL with 18-MΩ deionized water.
Blood samples were analyzed in three separate runs using an inductively-coupled plasma mass
spectrometer (ICP-MS, Perkin-Elmer, Norwalk, CT) to quantify trace element concentrations.
Certified reference material (TORT-3, National Research Council of Canada, Ottawa, ON) was
used as a standard for verifying trace metal recoveries for the blood digests. Mean percent
recoveries for trace elements in TORT-3 ranged from 85.7% (Strontium) – 92.3% (Se).
Minimum detection limits in blood samples averaged: As, 0.005; Cadmium, 0.002; Chromium,
0.002; Copper, 0.005; Se, 0.103; and Sr, 0.003 mg/kg. Blood trace element concentrations are
reported as mg/kg on a wet-mass basis.

29
Trace Element Analysis: Claw
Claw trace element concentrations were measured via ICP-MS in one batch at the Trace Element
Analysis Core of Dartmouth College. The claws were first washed to remove any foreign matter.
For washing, claws were placed into individual 7-mL polyethylene vials, 2 mL of acetone were
added, and the mixture was placed into an ultrasonic bath for 20 minutes. Claws were then
washed with 2 mL of 1% Triton X-100 in an ultrasonic bath for 20 minutes, washed 5 more
times with deionized water, and the vial was then placed into a clean box to dry. Once claw
samples were dry, claws were placed into a pre-weighed trace metal clean 15-mL polypropylene
centrifuge tube (VWR, Radnor, PA) and 0.5 mL of 9:1 HNO3:HCl (Optima Grade, Fisher
Scientific) was added. Weights of claws were recorded on a 4 place balance to the nearest 0.1
mg. Samples were prepped for acid digestion along with certified reference materials, digestion
blanks and fortified blanks for quality control. Tubes were then placed into the digestion rack,
lightly capped, and set aside for approximately 3 hours. The digestion rack was then placed into
a CEM MARS6 Express (Matthews, NC) microwave digestion system for digestions. To monitor
the temperature of the MARS unit, a fiber optic probe was placed into a sample tube and the
samples were heated to 95°C (ramp to temperature was 15 minutes) and held at this temperature
for 45 minutes. Once samples were cooled, 0.1 mL of H2O2 (Optima Grade, Fisher Scientific)
was added to samples and the samples were put through a second heating cycle. After cooling,
the samples were brought to 10 mL with deionized water (Element QPod, Millipore, Billarica,
MA) and all measurements were recorded gravimetrically. Digested samples were analyzed for
As, Cd, Cr, Cu, Se, and Sr via collison cell ICPMS (7700x, Agilent, Santa Clara, CA). Selenium
was measured in hydrogen mode with a flow rate of 6 mL/min. All other trace elements were
analyzed in helium mode with a flow rate of ~ 4.5 mL/min. Procedures used in this analysis

30
followed the protocols from EPA 6020A. Calibration checks and blanks were run every 10
samples, and analysis replicates and spikes were run at a ratio of 1 duplicate and spike to each 20
samples. Mean percent recoveries for trace elements in standard reference materials ranged from
77% (Cu) – 99% (Cd). Minimum detection limits (MDLs) in samples averaged: As, 0.016; Cd,
0.031; Cr, 0.155; Cu, 0.466; Se, 0.031; and Sr, 0.031 mg/kg dry mass. All claw concentrations
are reported on a dry mass (mg/kg) basis.
Bacteria-killing assays (BKAs)
A bactericidal assay was used to assess the innate immunity of wild T. scripta. Bacterial
killing assays are used commonly (birds, amphibians, and reptiles) in ecoimmunological research
to quantify an animal’s innate immune response to a species of bacteria (Millet et al. 2007;
French et al. 2010; Zimmerman et al. 2010; Brown et al. 2014). For each turtle, 7μL of plasma
was mixed with a 10μL aliquot of approximately 400 colony-forming units (CFUs) of
Escherichia coli (ATCC #8739; VWR, Radnor, PA) in phosphate buffered saline (PBS), and
137μL of CO2 independent media containing 5% fetal bovine serum and 4mM L-glutamine.
Once mixed, 50μL of the sample mixture was immediately spread with a fish-tail spreader onto
agar plates (Time 0 plates; Teknova Tryptic Soy plates, Fisher Scientific Company LLC,
Suwanee, GA). The remaining sample mixture was allowed to incubate at room temperature for
60 minutes, after which we placed 50μL onto another agar plate (Time 60 plates). Plates were
incubated overnight at 37ºC (Zimmerman et al. 2012). The following morning, the number of
bacterial colonies were counted for each agar plate. We calculated the percentage of bacterial
killed by an individual’s plasma using the following formula: (1 – [Time 60 colony count / Time
0 colony count]) * 100.

31
Phytohaemagglutinin (PHA) skin assays
To further assess innate immunity, animals were subjected to an immunological
challenge known as a PHA skin assay. This method requires an injection of a non-pathogenic,
antigenic lectin isolated from the red kidney bean (Phaseolus vulgaris) and pre- and post-
injection measurements of the injection site to measure swelling response (Finger et al. 2013).
Unlike most studies, we sought only to measure the initial, innate (non-specific) immune
response to the PHA injection. The first injection causes a proliferation of cells associated with
the innate immune response (i.e., granulocytes, lymphocytes, and macrophages; Martin et al.
2006). Due to the seasonal fluctuations in reptilian immunity (Zapata et al. 1992; Zimmerman et
al. 2009), we standardized this assay by only performing the tests during summer months (May-
July). PHA (PHA-P #L8754; Sigma-Aldrich, St. Louis, MO, USA) was dissolved into sterile
phosphate buffered saline (PBS #10010-023; Gibco by Life Technologies, Carlsbad, CA, USA)
to make a 2 mg/mL solution. For each turtle, 20μL was injected subcutaneously between the
webbing of the third and fourth digit of the right rear foot. As a control, the left foot was injected
in the same area with 20μL of PBS. Peak swelling occurs in yellow-bellied sliders (T. scripta) at
24 hours (Jones and Finger, unpublished data). Thickness measurements (to the nearest 0.01
mm) were taken prior to injection (time 0) and at 24 hours using a dial thickness pressure gauge
(Peacock G-1A, Ozaki Manufacturing Ltd, Japan). Three successive measurements were
obtained for each foot and averaged for analysis. Measurements were taken quickly and by the
same individual (AJ) across both years of data collection to minimize sampling errors.

32
Parasites: Hemogregarine quantification
Fixed blood smears were stained with Giesma and viewed in a zig-zag fashion at 1000X
magnification using oil immersion and a standard light microscope. Prevalence of
hemogregarines (proportion of sampled turtles infected with hemoparasites) was calculated. To
quantify total hemogregarine parasite loads, we read an average of approximately 121 fields of
view which contained an averaged ~ 73 ± 7 (mean ± 1 SE) erythrocytes for each smear (Haskins
and Hamilton, unpublished data). Therefore, we counted a total of 8000 erythrocytes per
individual. Total hemogregarine parasite loads were later transformed into a percentage of
infection (i.e., parasitemia; Davis & Sterrett 2011). Parasitemia (the percentage of blood cells
that are infected with hemogregarines) was calculated using the following equation: (# of
parasites / 8000 * 100). Developmental stages of hemogregarines were not characterized.
Statistics
Due to the limited number of turtles collected at individual reference locations, all
individuals captured at reference sites were grouped into a single site type (Reference). Likewise,
turtles captured in D-area’s primary basin and ‘A’ and ‘B’ wetlands were all grouped as D-area
turtles (Figure 2.1). If, for a given trace element, at least 50% of blood samples had detectable
amounts of a trace element, concentrations below the instrument’s minimum detection limits
(<MDL) were replaced with half of the original MDL (based on each sample weight) for
analysis.. All claw trace element concentrations were MDL.
All data were analyzed using program R (R Core Team 2016). Data were tested for
normality and homogeneity of variances using the Shapiro-Wilk and Levene statistic,
respectively. In cases where data did not fit the normality assumption, data were log-transformed

33
or rank-transformed (ANCOVA) prior to analysis. If data still did not fit the proper assumptions,
nonparametric statistics were used. If correlations were tested (parametric methods) and outliers
were suspected, Grubb’s test were used to locate outliers. Blood trace element concentrations
and bacterial killing assays were analyzed with nonparametric statistics (Mann-Whitney U tests).
Trace element concentrations in claws were compared using an ANCOVA with site type (D-
Area vs. Reference) as a main effect and PL as a covariate. Because blood and claw trace
element concentrations reflect different exposure time scales, we examined the correlational
relationship between these values via Spearman’s correlations. We also used ANCOVA to
evaluate possible differences in hemogregarine parasitemia of turtles, and PL was used as a
covariate. In regards to the description of hemogregarine prevalence, individuals were
considered juveniles if they were < 100mm in PL.
PHA responses were analyzed by using a linear mixed model (LMM) repeated measures
analysis with a serial correlation structure. Toe web swelling was the response variable within
the LMM analysis and individual turtle was used as a random effect. A likelihood ratio test was
performed to ensure that the LMM benefitted from the addition of turtle ID as a random effect.
Other variables in the PHA LMM included: plastron length (covariate), site type, and the
interaction of time (0 vs. 24 hours) and treatment (PBS vs. PHA).
Results
D-area vs. Reference populations
Over a 2 year period (2014-2015), a total of 81 T. scripta were collected from D-area and
reference locations (D-area, n = 39; Reference, n = 42). Twenty-six individuals were identified
as female, 27 were identified as male, and the remaining 28 were too immature to be sexed.

34
Average plastron size of females (n = 8) collected in D-area was 174.5 ± 17.7 mm (Mean ± SE;
range of 102-227 mm), and average plastron size of males (n = 11) in D-area was 154.4 ± 10.9
mm (106-205 mm). Plastron size of juveniles (n = 20) captured in D-area averaged 66.6 ± 2.9
mm (42-88 mm). Average plastron size of reference females (n = 18) was 202.4 ± 9.7 mm (129-
261 mm), males (n = 16) averaged 158.2 ± 8.5 mm (65-195 mm), and juveniles (n = 8) averaged
72.1 ± 3.8 mm (59-91 mm).
Trace element burdens (Blood)
Chromium, Cu, and Sr were detected in 100% of blood samples. Arsenic and Se were
<MDL in 43% (27/63) and 37% (23/63) of blood samples, respectively. Cadmium was <MDL in
86% (54/63) of all blood samples. Arsenic (U = 657, p = 0.015), Cu (U = 779, p < 0.001), Se (U
= 836, p < 0.001), and Sr (U = 858, p < 0.001) concentrations were all significantly higher in D-
area turtles than in reference turtles. In contract, Cr (U = 291.5, p = 0.006) concentrations were
significantly higher in animals captured in reference areas than in D-area turtles (Table 2.1). Two
individuals from reference areas had high amounts of Cr in their blood; however, even after these
two observations were removed the comparison remained significant (U = 291.5, p = 0.017).
Because the two outliers were captured at wetlands (Ellenton Bay and Risher Pond) ranging 2.4 -
4.3 km from D-area, it is possible that these individuals had spent time at D-area (or another
contaminated area) prior to moving to their capture location.
Trace element burdens (Claw)
Arsenic, Cd, Cr, Cu, Se, and Sr were detected in 100% (n = 56) of claw samples.
ANCOVAs were used to test differences in claw trace element values between site types using

35
PL as a covariate (Table 2.2). Arsenic (F1,52 = 29.37, p < 0.001), Cd (F1,52 = 15.20, p < 0.001),
Cu (F1,52 = 11.60, p = 0.001), and Se (F1,52 = 70.99, p < 0.001), were significantly higher in D-
area turtles compared to reference turtles, while neither Cr (F1,52 = 2.30, p = 0.135) nor Sr claw
concentrations (F1,52 = 1.91, p = 0.169) were significantly different between sites. Plastron length
was a significant covariate in the As (F1,52 = 4.90, R2 = 0.37, p = 0.030), Cd (F1,52 = 18.21, R2 = -
0.45, p < 0.001), and Cu (F1,52 = 21.69, R2 = - 0.37, p < 0.001) ANCOVA models. Cadmium and
Cu were negatively related to PL, whereas As concentrations were positively related to PL
(Figures 2.2-2.4).
Relationship between blood and claw trace element concentrations
To explore the relationship between turtle blood and claw values we tested these data
with Spearman’s correlation statistic. We only included in analyses individuals for which trace
element concentrations for a given element >MDL for both blood and claw. Thus, due to
detectability issues, Cd blood and claw could not be examined and As and Se sample sizes were
reduced. Comparisons of blood and claw As, Cr, Se, and Sr yielded significant relationships
(Figure 2.5). Arsenic trace element values had a moderately positive relationship (n = 24, rho =
0.52, p = 0.009), with 27% of variation in blood [As] explained by claw [As]. Similarly, Sr blood
and claw concentrations (n = 51, rho = 0.30, p = 0.034) had a moderately positive relationship.
However, only 9% of variation in blood [Sr] could be explained by claw [Sr]. Chromium blood
and claw concentrations (n = 51, rho = - 0.29, p = 0.042), exhibited a negative relationship and
only 8% of blood [Cr] variation could be explained by claw [Cr]. Selenium blood and claw
values (n = 24, rho = 0.87, p < 0.001) exhibited a strong positive relationship, with 75% of
variation in blood [Se] explained by claw [Se].

36
Bacterial killing assays (BKAs)
Mean percent bacteria killed by D-area turtles (0.91 ± 0.03) was significantly higher than
reference turtles (0.88 ± 0.03; Figure 2.6), based on Mann-Whitney U test (U = 1077, p = 0.015).
Differences within site types were also explored to examine possible trends based on sex and age
class of turtles. In D-area turtles, BKA response did not differ by age class (adults versus
juveniles; U = 205, p = 0.683); differences in BKA response by sex could not be investigated due
to low sample sizes (10 males, 8 females). In reference turtles, BKA response did not differ by
sex (U = 157, p = 0.666). Relationships between BKA response and trace element burdens could
not be statistically evaluated due to the extreme nonnormality of the BKA data.
Phytohaemagglutinin (PHA) skin assays
Originally we ran three separate models to interpret PHA response between site types,
with each model containing a different ARMA (autoregressive moving average) serial
correlation structure. Another LMM was created to explore if PHA responses were impacted by
sex and the same parameters as the previous model. The same model selection methods were
used for the second LMM. Linear mixed models for PHA response are reported in Table 2.3.
PHA-treated toe webs had significantly higher swelling responses compared to PBS-treated toe
webs (t 1,125 = 5.06, p = 0.019), and this difference varied over time (interaction term: treatment
group * time; t 1,125 = 3.06, p = 0.003; Figure 2.7), indicating that the assay was successful in
producing a measurable swelling response. PHA responses (i.e., differences in swelling response
between PHA- and PBS-injected sites) did not differ between D-area and reference turtles when
compared using a linear mixed model with turtle ID as a random effect (t 1,41 = 0.578, p = 0.566).
The same model suggested that PHA response did not vary with turtle size (i.e., PL; t 1,125 = 1.58,

37
p = 0.116). A second model to investigate sex-related effects on adult T. scripta revealed no
significant difference in swelling response between males and females (t 1,34= - 0.583, p = 0.564).
PHA response did not vary as a function of claw trace element burdens for any trace element
examined (all p > 0.10).
Hemogregarine parasitemia
Mean prevalence of hemogregarine infection was 51.85% and 77.78% for D-area and
reference turtles, respectively (Table 2.4). When combined across all sites, female T. scripta had
the highest rate of prevalence (15/16) at 94%, followed by males (9/14) at 64%, and immatures
at 20% (3/15). Overall prevalence of hemoparasites among all individuals at all sites sampled
was 62.22%. Parasitemia levels were not significantly different between sites based on an
ANCOVA (D-area = 0.030 ± 0.01; Reference = 0.043 ± 0.02; F1,40 = 0.01, p = 0.98). However,
PL was a significant covariate in the model (F1,40 = 21.87, p < 0.001; Figure 2.8), with
parasitemia increasing with increasing body size. Two observations were not included in the
ANCOVA analysis because we did not have plastron length for these individuals. In addition,
parasitemia was positively correlated with log-transformed PHA responses (r = 0.51, p = 0.003).
Grubb’s tests detected four outliers (G = 4.73, p = 0.04; G = 3.55, p < 0.001). After removing
outliers, parasitemia levels were still positively correlated with PHA responses (r = 0.8, p <
0.001; Figure 2.9).
Discussion
Blood trace element concentrations of As, Cu, Se, and Sr in T. scripta collected from D-
area were significantly elevated compared to those captured in reference wetlands. An

38
interesting observation from our data is that contrary to our hypothesis, blood Cr concentrations
were significantly higher in reference turtles. Two reference turtles (an adult female and an
immature individual) had the highest amounts of blood Cr (1.31 and 2.50 mg/kg, respectively) in
our dataset. The wetlands of origin for these turtles are within the dispersal range for D-area T.
scripta (2.4 – 4.3 km), and trace element analysis of blood reflects a short-term measure of
contaminant exposure. Therefore, it is possible that these individuals had migrated from
contaminated sites such D-area prior to their capture. Other blood trace element values (As, Cd,
Cu, Se, and Sr) supported our hypothesis that D-area turtles would exhibit higher blood burdens
of CCR-associated trace elements compared to reference turtles. Our blood trace element data is
similar to recent historical data at the same site (Tuberville et al., unpublished data). Mean blood
As, Cu, and Se were slightly lower in our study (As, 0.07 ± 0.02 mg/kg; Cu, 0.46 ± 0.02 mg/kg;
Se, 1.56 ± 0.43 mg/kg) relative to previous data (As, 0.14 ± 0.01 mg/kg; Cu, 0.52 ± 0.03 mg/kg;
Se, 2.19 ± 0.05 mg/kg) from D-area (Tuberville et al., unpublished data). Blood Sr
concentrations from this study were also similar (Sr, 0.52 ± 0.06 mg/kg) compared to previous
data (Sr, 0.47 ± 0.01 mg/kg; Tuberville et al., unpublished data). The majority of D-area turtles
captured in our study came from wetlands adjacent to the settling basins, and mean blood trace
element concentrations of As, Cu, Se, and Sr were higher in animals caught in the primary
settling basin. Therefore, the slightly lower trace element values in this study are likely due a
large amount of turtles being captured in adjacent wetlands and not the settling basins. Blood As,
Cu, Se, and Sr values in D-area are higher than those reported from T. scripta living in
Tennessee waterways that were heavily impacted by a coal fly ash spill (Van Dyke et al. 2014).
Interestingly, blood As levels in D-area turtles from adjacent wetlands were equal or slightly
elevated compared to T. scripta exposed to the coal fly ash spill in Tennessee. The alkalinity of

39
ash basins influences mobility of trace elements, and As is one of the more mobile trace elements
in CCRs (Izquierdo and Querol 2012).
We hypothesized that claw trace elements, like blood values, would also be elevated in
D-area turtles relative to reference turtles. Although few studies have quantified claw trace
element concentrations in freshwater turtles, claw burdens in this study are among the highest
reported in freshwater turtles. Mean claw As, Cd, Cu, and Se concentrations were significantly
higher in turtles captured in D-area compared to reference turtles. Although not significant, claw
Sr and Cr values were also elevated in D-area turtles compared to turtles from reference sites. As
one might expect, likely due to length and severity of exposure, claw trace element values from
D-area turtles in this study were much higher than those reported from turtles captured in an area
affected by the Tennessee Valley Authority’s (TVA) coal ash spill of 2008 (Van Dyke et al.
2014). In fact, mean concentrations of D-area T. scripta claw Se were five times higher than
mean claw Se values reported for T. scripta collected near the TVA spill site (D-area, 6.65
mg/kg; TVA, ~ 1.25 mg/kg; Van Dyke et al. 2013). Trachemys scripta captured in D-area had
mean claw Sr (47.56 ± 10.49 mg/kg) concentrations 200 times higher than T. scripta captured
near the TVA coal ash spill (0.25 ± 0.04 mg/kg; Steen et al. 2015). Overall, these data confirmed
our hypothesis that claw trace element values would be elevated in turtles that are residents of
the D-area system.
Claw As, Cd, and Cu concentrations were influenced by individual size (PL). Although in
some cases larger turtles were captured in reference areas, younger turtles in this study tended to
accumulate larger amounts of trace elements in claws relative to adults. Previous studies suggest
that younger, immature T. scripta consume a higher protein diet compared to adults, who tend to
consume more plant matter (Clark and Gibbons 1969; Hart 1983; Parmenter and Avery 1990).

40
Unrine et al. (2007) reported that odonate larvae (Tramea and Erythemis spp.) and bullfrog
(Lithobates catesbeiana) larvae that were sampled near D-area accumulated high amounts of As,
Cd, Se, and Cu. Studies of T. scripta have previously recorded odonate and anuran species
within their diets (Parmenter and Avery 1990). Perhaps size-related differences in claw trace
element burdens can be attributed to the ontogenetic diet shift that these turtles experience. Little
is known about the behavioral habits of juvenile freshwater species, and hatchling T. scripta
probably exhibit little movement once they have entered the aquatic environment (Gibbons
1990). In contaminated habitats such as D-area, if juvenile T. scripta exhibit higher site fidelity
than adults it could expose them to larger amounts of CCR trace elements.
To further explore the validity of blood and claw tissues as non-destructive sampling
methods, we investigated correlational relationships in trace element concentrations between the
two sample types. Claw trace element concentrations have a long turnover period (~ 12 months;
Aresco 2005), while blood trace element values are great indicators of recent exposure. Of the
six trace element relationships we examined, As, Cr, Se, and Sr claw concentrations were
significantly and positively correlated with corresponding blood concentrations. Arsenic and Se
(blood and claw) comparisons yielded the strongest relationships. Recent studies of trace element
correlations in freshwater turtles found that some trace elements (Mercury, Se) were significantly
and positively correlated in blood and claw tissues (B.C. Hopkins et al. 2013; Van Dyke et al.
2013). As highlighted in previous studies, there is little information regarding thresholds of trace
element concentrations in reptilian species (Komoroske et al. 2012; Van Dyke et al. 2013).
Therefore, while it is difficult to relate our non-destructive data to toxicity benchmarks, our data
does reveal exposure histories for wild T. scripta,

41
Although reptiles are experiencing global declines, and contaminants have been proposed
as a factor of this decline (Gibbons et al. 2000), few studies have investigated the potential of
contaminants to cause immunosuppressive effects in this taxa (Keller et al. 2006). We predicted
that T. scripta captured in D-area would display weakened BKA responses; however, our results
suggest that BKA responses in T. scripta were elevated in D-area. In studies near the TVA coal
spill, tree swallows sampled at reference and coal ash spill sites were found to have positively
correlated Se burdens and BKA responses, and birds sampled downstream of the spill had
slightly elevated BKA responses compared to other sites (Beck et al. 2014). Because swallow
blood Se values were correlated with BKA response, authors hypothesized that increased Se may
have increased gene expression of genes associated with the innate immune response. In regards
to our turtles, one must also consider the microbial community of the ash basins when
interpreting these results. Previous data from coal ash basins on the SRS found that ash samples
contained higher estimates of bacteria relative to control samples, and that microorganism
populations increase as the ash basins age (Klubek et al. 1992). D-area’s coal ash basins are over
60 years old, which suggests that microbial populations at D-area could be more robust than
those at reference sites. Overall, BKA responses in our turtles were similar between D-area and
reference site types (0.91 ± 0.03 and 0.88 ± 0.03, respectively). Our analysis suggested a
significant difference between site types but any biological consequences of such a small
difference is difficult to interpret.
Assays that measure cell-mediated immune responses, like PHAs, can quantify
components of both the innate and adaptive immune responses in vertebrates (Martin et al. 2006;
Demas et al. 2011). In our current study, as expected, the administration of PHA in T. scripta
caused a significantly greater increase in toe web swelling than PBS, and this difference varied

42
significantly over time. We predicted that T. scripta with high trace element burdens would
exhibit weaker PHA responses relative to individuals with lower trace element burdens.
Although T. scripta from D-area had elevated trace elements in their blood and claws, site type
was not a significant factor in PHA swelling response. Furthermore, PHA swelling response was
not correlated with any claw trace element concentrations in T. scripta. Our results also suggest
that in T. scripta PHA response is not affected by plastron size. Studies of PHA responses in the
western pond turtle (Emys marmorata) suggested that E. marmorata inhabiting a water pollution
control plant had weaker PHA responses than individuals residing in reference habitats (Polo-
Cavia et al. 2009). Adult common eiders (Somateria mollissima) exposed to Se, a trace element
found in high concentrations in coal ash, had weaker PHA responses relative to control birds
(Franson et al. 2007). PHA experiments in saltwater crocodiles (Crocodylus porosus) and
painted turtles (Chrysemys picta) found that PHA swelling response significantly increases
corresponding with an individual’s size (i.e., snout-vent length and plastron length; Schwanz et
al. 2011; Finger et al. 2013). However, Zimmerman et al. (2010) found that swelling response
was not affected by plastron size in T.s. elegans. Perhaps Trachemys spp. do not exhibit size-
related differences to antigenic stimulation, although, more studies need to be performed before
this is determined.
Contrary to our hypothesis, hemogregarine parasite loads did not yield significant
differences based on site type. However, we did note a larger prevalence of infected individuals
in reference sites. In 2015, we observed leeches (Placobdella spp.) on 9 T. scripta captured in D-
area and we also found leeches on 5 T. scripta collected from reference sites. Our analyses did
also find that hemogregarine parasitemia covaried significantly and positively with plastron
length. An interesting observation in the current study is that 3/15 juvenile T. scripta (average PL

43
= 67.3 ± 4.1) were infected with hemogregarines. To our knowledge, this study is the first
reported instance of hemogregarine infection in immature T. scripta. Hemogregarines are
thought to be most commonly transmitted to freshwater turtles by leeches (Davis and Sterrett
2011), which have been shown to be sensitive to contaminants such as mercury and atrazine
(Suchanek et al. 1995; Brites and Rantin 2004), and are moderately sensitive to contamination on
the North Carolina Biotic Index (NCBI; Lenat 1993). Thus, it is intriguing that hemogregarines
loads did not vary between reference site types and D-area. It may be that leeches inhabiting D-
area wetlands can tolerate environments with high amounts of CCR contaminants. In past
studies, larger freshwater turtles, typically females, had larger hemogregarine parasitemia values
and this is likely due to their larger surface area (McCoy et al. 2007; Davis and Sterrett 2011;
Schwanz et al. 2011). Previous observations of hemogregarine prevalence in southeastern
(Georgia, Kentucky, Louisiana, and Tennessee) freshwater turtles range from 45-100% and
average to approximately 75% prevalence (Davis and Sterrett 2011). Thus, our prevalence values
(62.22%) are within the range of what has been reported in freshwater turtles.
Reptiles are threatened globally and little is known regarding how these species respond
if chronically exposed to contaminants of anthropogenic origin. Trachemys scripta captured in
D-area exhibited some of the highest claw trace element concentrations reported in freshwater
turtle species. Arsenic and Se burdens in our study were positively correlated between blood and
claw tissues, suggesting that in spite of the discrepancy between blood and claw tissue turnover,
either may serve as an acceptable non-destructive sample for monitoring specific trace element
burdens. Smaller T. scripta also seemed to accumulate higher amounts of claw Cd and Cu, while
larger T. scripta tended to have elevated claw As concentrations. The observed relationship
between plastron size and trace elements may have interesting implications. Immature T. scripta

44
in sites such as D-area may be exposed to preferred prey that contain high concentrations of toxic
trace elements. Freshwater turtle species exhibit high juvenile mortality and are sensitive to
anthropogenic-based adult mortality (i.e., vehicle collisions, habitat alterations; Gibbons 1990).
Thus, future controlled studies should be used to investigate the toxicological implications of
acute and chronic trace element exposure in multiple age classes. Furthermore, future work
should also aim to discern the nature of relationships among trace element burdens and multiple
tissue types.
The reptilian immune system is complex, is affected by a wide variety of factors (i.e.,
age, population dynamics, stress, temperature, season, gender; Keller et al. 2006), and more
comprehensive studies are necessary to fully explore the potential effects of contaminants on
turtle immune function and host-parasite dynamics. Future studies of contaminant exposure in
wild freshwater turtles should seek to incorporate long-term monitoring of hematological
parameters, nutrient profiles, parasite loads, and further measurements of immunity (i.e.,
adaptive and humoral immunity) to complement innate immunity measures. Field and laboratory
studies of CCR-exposure in T. scripta may allow future researchers to discern if they are affected
by trace element concentrations observed in this study.

45
Literature Cited
American Coal Ash Association [ACAA]. 2010 Coal combustion product (CCP) production and
use report. 2010. <https://www.acaa-
usa.org/Portals/9/Files/PDFs/2010_CCP_Survey_FINAL_102011.pdf
>. Accessed on 16 Feb 2016.
Aguirre, A.A., and P.L. Lutz. 2004. Marine turtles as sentinels of ecosystem health: is
fibropapillomatosis an indicator? EcoHealth 1:275-283.
Aresco, M.J. 2005. Ecological relationships of turtles in northern Florida lakes: a study of
omnivory and the structure of the lake food web. Dissertation, Florida State University,
Tallhassee, FL.
Beck, M.L., W.A. Hopkins, J.J. Hallagan, B.P. Jackson, D.M. Hawley. 2014. Exposure to
residual concentrations of elements from a remediated coal fly ash spill does not
adversely influence stress and immune responses of nestling tree swallows. Conservation
Physiology 2:1-13.
Beresford, W.A., M.P. Donovan, J.M. Henninger, M.P. Waalkes. 1981. Lead in the bone and soft
tissues of box turtles caught near smelters. Bulletins of Environmental Contamination and
Toxicology 27:349-352.
Brites, V.L., and F.T. Rantin. 2004. The influence of agricultural and urban contamination on
leech infestation of freshwater turtles, Phyrnops geoffroanus, taken from two areas of the
Uberabinha river. Environmental Monitoring and Assessment 96:273-281.

46
Brown, G.P., and R. Shine. 2014. Immune response varies with rate of dispersal in invasive cane
toads (Rhinella marina). PLoS ONE 9:e99734. doi:10.1371/journal.pone.0099734.
Bryan Jr., A.L., W.A. Hopkins, J.H. Parikh, B.P. Jackson, and J.M. Unrine. Coal fly ash basins
as an attractive nuisance to birds: parental provisioning exposes nestlings to harmful trace
elements. Environmental Pollution 161:170-177.
Cagle, F.W. 1939. A system of marking turtles for future identification. Copeia 1939:170-173.
Clark, D.B., and J.W. Gibbons. 1969. Dietary shift in the turtle Pseudemys scripta (Schoepff)
from youth to maturity. Copeia 1969:704-706.
Camacho, M., J. Orós, L.D. Boada, A. Zaccaroni, M. Silvi, C. Formigaro, P. López, M.
Zumbado, and O.P. Luzardo. 2013. Potential adverse effects of inorganic pollutants on
clinical paramters of loggerhead sea turtles (Caretta caretta): results from a nesting
colony from Cape Verde, West Africa. Marine Environmental Research 91:15-22.
Davis, A.K., and S.C. Sterrett. 2011. Prevalence of haemogregarine parasites in three freshwater
turtle species in a population in Northeast Georgia, USA. International Journal of
Zoological Research. 7:156-163.
Demas, G.E., D.A. Zysling, B.R. Beechler, M.P. Muehlenbein, and S.S. French. 2011. Beyond
phytohaemagglutinin: assessing vertebrate immune function across ecological contexts.
Journal of Animal Ecology 80:710-730.
Congdon, J.D., J.L. Greene, and J.W. Gibbons. 1986. Biomass of freshwater turtles: a geographic
comparison. American Midland Naturalist. 115:165-173.
Fairbrother, A., M. Fix, T. O’Hara, and C. Ribic. 1994. Impairment of growth and immune
function of avocet chicks from sites with elevated selenium, arsenic, and boron. Journal
of Wildlife Diseases 30:222-233.

47
Faust, D.R., M.J. Hooper, G.P. Cobb, M. Barnes, D. Shaver, S. Ertolacci, and P.N. Smith. 2014.
Inorganic elements in green sea turtles (Chelonia mydas): relationships among external
and internal tissues. Environmental Toxicology and Chemistry 33:2020-2027.
Finger, J., A.L. Adams, P.C. Thomson, C.M. Shilton, G.P. Brown, C. Moran, L.G. Miles, T.C.
Glenn, and S.R. Isberg. 2013. Using phytohaemagglutinin to determine immune
responsiveness in saltwater crocodiles (Crocodylus porosus). Australian Journal of
Zoology 61:301-311.
Franson, J.C., A. Wells-Berlin, M.C. Perry, V. Shearn-Bochsler, D.L. Finley, P.L. Flint, and T.E.
Hollmén. 2007. Effects of dietary selenium on tissue concentrations, pathology, oxidative
stress, and immune function in common eiders (Somateria mollissima). Journal of
Toxicology and Environmental Health A 70:861-874.
French, S.S., D.F. DeNardo, T.J. Greives, C.R. Strand, and G.E. Demas. 2010. Human
disturbance alters endocrine and immune responses in Galapagos marine iguanas
(Amblyrhynchus cristatus). Hormones and Behavior 58:792-799.
Ghosh, D., S. Bhattacharya, S. Mazumder. 2006. Perturbations in the catfish immune responses
by arsenic: organ and cell specific effects. Comparative Biochemistry and Physiology
143:455-463.
Gibbons, J.W. 1990a. The Slider Turtle. Pages 1-18 in Sheffield, R., editor. Life History and
Ecology of the Slider Turtle. Smithsonian Institution Press, Washington, D.C., USA.
Print.
Gibbons J.W. 1990b. Turtle Studies at SREL: A research perspective. Pages 19-45 in Sheffield,
R., editor. Life History and Ecology of the Slider Turtle. Smithsonian Institution Press,
Washington, D.C., USA. Print.

48
Gibbons, J.W., D.E. Scott, T.J. Ryan, K.A. Buhlmann, T.D. Tuberville, B.S. Metts, J.L. Greene,
T. Mills, M. Leiden, S. Poppy, and C.T. Winne. 2000. The global decline of reptiles, Déjà
vu amphibians. Bioscience 50:653-666.
Graham, A.L. 2002. When T-helper cells don’t help: immunopathology during contomitant
infection. Quarterly Reviews of Biology 77:409-434.
Grillitsch, B., and L. Schiesari. 2010. The ecotoxicology of metals in reptiles. Pages 337-448 in
Sparling, D.W., G. Linder, A.C. Bishop, S. Krest, editors. Ecotoxicology of Amphibians
and Reptiles. Pensacola, FL: Society of Environmental Toxicology and Chemistry
(SETAC).
Guthrie, R.K., and D.S. Cherry. 1976. Pollutant removal from coal-ash basin effluent. Water
Resources Bulletin 12:889-902.
Harden, L.A., S.J. Price, and M.E. Dorcas. 2009. Terrestrial activity and habitat selection of
eastern mud turtles (Kinosternon subrubrum) in a fragmented landscape: implications for
habitat management of golf courses and other suburban environments. Copeia 2009:78-
84.
Hart, D. 1983. Dietary and habitat shift with size of red-eared turtles (Pseudemys scripta) in a
southern Louisiana population. Herpetologica 39:285-290.
Hernandez-Divers, S.M., S.J. Hernandez-Divers, and J. Wyneken. 2002. Angiographic,
anatomic, and clinical technique descriptions of a subcarapacial venipuncture site for
chelonians. Journal of Herpetological Medicine and Surgery 12:32-37.
Hopkins, B.C., Willson, J.D., and W.A. Hopkins. 2013. Mercury exposure is associated with
negative effects on turtle reproduction. Environmental Science and Technology 47:2416-
2422.

49
Hopkins, W.A., C.L. Rowe, and J.D. Congdon. 1999. Elevated trace element concentrations and
standard metabolic rates in banded water snakes (Nerodia fasciata) exposed to coal
combustion wastes. Environmental Toxicology and Chemistry 18:1258-1263.
Izquierdo, M., and X. Querol. 2012. Leaching behaviour of elements from coal combustion fly
ash: an overview. International Journal of Coal Geology 94:54-66.
Keller, J.M., M.M. Peden-Adams, A.A. Aguirre. 2006. Immunotoxicology and implications for
reptilian health. Pages 199-240 in Gardner, S.C., and E. Oberdorster, (eds). Toxicology of
reptiles. CRC Press, Boca Raton, FL.
Klubek, B., C.L. Carison, J. Oliver, and D.C. Adriano. 1992. Characterization of microbial
abundance and activity from three coal ash basins. Soil Biology and Biochemistry
24:1119-1125.
Komoroske, L.M., R.L. Lewison, J.A. Seminoff, D.D. Deustchman, and D.D. Deheyn. 2012.
Trace metals in an urbanized estuarine sea turtle food web in San Diego Bay, CA.
Science of the Total Environment 417:108-116.
Lemly, A.D., and J.P. Skorupa. 2012. Wildlife and the coal waste policy debate: proposed rules
for coal waste disposal ignore lessons from 45 years of wildlife poisoning. Environmental
Science and Toxicology 46:8595-8600.
Lenat, D.R. 1993. A biotic index for the southeastern United States: derivation and list of
tolerant values criteria for assigning water-quality ratings. Journal of North American
Benthological Society 12:279-290.
Martin, L.B. II, P. Han, J. Lewittes, J.R. Kuhlmann, K.C. Klasing, and M. Wikelski. 2006.
Phytohemagglutinin-induced skin swelling in birds: histological support for a classic
immunoecological technique. Functional Ecology 20:290-299.

50
Martin, L.B. II, W.A. Hopkins, L.D. Mydlarz, and J.R. Rohr. 2010. The effects of anthropogenic
global changes on immune functions and disease resistance. Annals of the New York
Academy of Sciences 1195:129-148.
Meyer, E., C.A. Eagles-Smith, D. Sparling, and S. Blumenshine. 2014. Mercury exposure
associated with altered plasma thyroid hormones in the declining Western Pond Turtle
(Emys marmorata) from California mountain streams. Environmental Science and
Technology 48:2989-2996.
Meyers-Schöne, L., and B.T. Walton. 1994. Turtles as monitors of chemical contaminants in the
environment. Pages 93-153 in Ware, G.W., editor. Reviews of Environmental
Contamination and Toxicology, Vol. 135. Springer Publishing, New York City, New
York, USA.
Millet, S., J. Bennett, K.A. Lee, M. Hau, and K.C. Klasing. 2007. Quantifying and comparing
constitutive immunity across avian species. Developmental and Comparative
Immunology 31:188-201.
Morreale, S.J., J.W. Gibbons, and J.D. Congdon. 1984. Significance of activity and movement in
the yellow-bellied slider turtle (Pseudemys scripta). Canadian Journal of Zoology
62:1038-1042.
Nagle, R.D., C.L. Rowe, and J.D. Congdon. 2001. Accumulation and selective maternal
transfer of contaminants in the turtle Trachemys scripta associated with coal ash
deposition. Archives of Environmental Contamination and Toxicology 40:531-536.
Parmenter, R.R., and H.W. Avery. 1990. The feeding ecology of the slider turtle. Pages 257-266
in Sheffield, R., editor. Life History and Ecology of the Slider Turtle. Smithsonian
Institution Press, Washington, D.C., USA.

51
Polo-Cavia, N., T. Engstrom, P. López, and J. Martin. 2009. Body condition does not predict
immunocompetence of western pond turtles in altered veruss natural habitats. Animal
Conservation 13:256-264.
R Core Team. 2016. R: a language and environment for statistical computing. R Foundation for
Statistical Computing, Vienna, Austria. <http://R-project.org>.
Rowe, C.L. 1998. Elevated standard metabolic rate in a freshwater shrimp (Palaemonetes
paludosus) exposed to trace element-rich coal combustion waste. Comparative
Biochemistry and Physiology 121:299-304.
Rowe, C.L. 2008. The calamity of so long life: life histories, contaminants, and
potential emerging threats to long-lived vertebrates. Bioscience. 58:623-631.
Rowe, C.L., W.A. Hopkins, C. Zehnder, and J.D. Congdon. 2001. Metabolic costs incurred by
crayfish (Procambarus acutus) in a trace element-polluted habitat: further evidence of
similar responses among diverse taxonomic groups. Comparative Biochemistry and
Physiology 129:275-283.
Rowe, C.L., W.A. Hopkins, and J.D. Congdon. 2002. Ecotoxicological implications of aquatic
disposal of coal combustion residues in the United States: A review. Environmental
Monitoring and Assessment 80:207-276.
Schwanz, L., D.A. Warner, S. McGaugh, R.D. Terlizzi, and A. Bronikowski. 2011. State-
dependent physiological maintenance in a long-lived ectotherm, the painted turtle
(Chrysemys picta). The Journal of Experimental Biology 214:88-97.
Steen, D.A., J.U. Van Dyke, B.P. Jackson, and W.A. Hopkins. 2015. Reproduction and hatchling
performance in freshwater turtles associated with a remediated coal fly-ash spill.
Environmental Research. 138:38-48.

52
Suchanek, T.H., P.J. Richerson, L.J. Holts, B.A. Lamphere, C.E. Woodmansee, D.G. Slotton,
E.J. Harner, and L.A. Woodward. 1995. Impacts of mercury on benthic invertebrate
populations and communities within the aquatic ecosystem of Clear Lake, California.
Water, Air, and Soil Pollution 80:951-960.
Tucker, J.K. 2001. Nesting red-eared sliders exhibit site fidelity to their nesting areas. Journal of
Herpetology 35:661-664.
Turtle Taxonomy Working Group. 2014. Turtles of the world, 7th edition: annotated checklist of
taxonomy, synonomy, distribution with maps, and conservation status. Pages 329-479 in
Rhodin, A.G.J., P.C.H. Pritchard, P.P. van Dijk, R.A. Saumure, K.A. Buhlmann, J.B.
Iverson, and R.A. Mittermeier.
Unrine, J.M., W.A. Hopkins, C.S. Romanek, and B.P. Jackson. 2007. Bioaccumulation of trace
elements in omnivorous amphibian larvae: Implications for amphibian health and
contaminant transport. Environmental Pollution 149:182-192.
Van Dyke, J.U., W.A. Hopkins, and B.P. Jackson. 2013. Influence of relative trophic position
and carbon source on selenium accumulation in turtles from a coal fly-ash spill site.
Environmental Pollution 182:45-52.
Van Dyke, J.U., D.A. Steen, B.P. Jackson, and W.A. Hopkins. 2014. Maternal transfer and
embryonic assimilation of trace elements in freshwater turtles after remediation of coal
fly ash spill. Environmental Pollution 194:38-49.
Yu, S., R.S. Halbrook, and D.W. Sparling. 2011. Metal accumulation and evaluation of effects in
a freshwater turtle. Ecotoxicology 20:1801-1812.
Zapata, A.G., A. Varas, and M. Torroba. 1992. Seasonal variations in the immune system of
lower vertberates. Immunology Today 13:142-147.

53
Zimmerman, L.M. 2013. Immune strategy of a long-lived ectotherm, the red-eared slider,
Trachemys scripta. Dissertation, Illinois State University, Normal, IL.
Zimmerman, L.M., L.A. Vogel, and R.M. Bowden. 2009. Understanding the vertebrate immune
system: insights from the reptilian perspective. The Journal of Experimental Biology
213:661-671.
Zimmerman, L.M., R.T. Paitz, S.G. Clairardin, L.A. Vogel, and R.M. Bowden. 2010. Variation
in seasonal patterns of innate and adaptive immunity in the red-eared slider (Trachemys
scripta). Journal of Experimental Biology 213:1477-1483.
Zimmerman, L.M., R.T. Paitz, S.G. Clairardin, L.A. Vogel, and R.M. Bowden. 2012. No
evidence that estrogens affect the development of the immune system in the red-eared
slider turtle (Trachemys scripta elegans). Hormones and Behavior. 62:331-336

54
Table 2.1: Trace element concentrations in claw (mg/kg dw) and blood (mg/kg ww) from
yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and reference areas on the
Savannah River Site, South Carolina. Values are reported as means ± 1 SE (ranges of raw values
reported below respective means in parentheses). Significant differences between site types are
marked with bold text. Samples were collected during the summers (May-August) of 2014 and
2015. Note that blood As and Se means are not reported in the table but analyses were performed
using half of the minimum detection limit (MDL) for each sample.
Element D-Area Reference
Claws Blood Claws Blood
As 3.94 ± 1.06 0.07 ± 0.02 0.06 ± 0.01 <MDL
(0.02 – 21.74) (0.00 – 0.42) (0.00 – 0.30) (0.00 – 0.02)
Cd 0.13 ± 0.02 <MDL 0.04 ± 0.01 <MDL
(0.01 – 0.40) <MDL (0.00 – 0.12) <MDL
Cr 0.14 ± 0.02 0.07 ± 0.01 0.10 ± 0.02 0.23 ± 0.09
(0.01 – 0.32) (0.02 – 0.44) (0.02 – 0.50) (0.02 – 2.50)
Cu 3.13 ± 0.56 0.46 ± 0.02 1.45 ± 0.33 0.33 ± 0.02
(0.48 – 14.54) (0.06 – 0.66) (0.41 – 8.37) (0.01 – 0.59)
Se 6.65 ± 1.59 1.56 ± 0.43 0.30 ± 0.03 <MDL
(0.49 – 39.24) (0.08 – 9.57) (0.13 – 0.70) (0.08 – 0.45)
Sr 71.04 ± 0.89 0.52 ± 0.06 47.56 ± 10.49 0.22 ± 0.02
(0.17 – 271.44) (0.05 – 1.45) (0.11 – 245.40) (0.01 – 0.51)
n (sample size) 30 35 25 28

55
Table 2.2: Results of analysis of covariance of the effects of site type (D-area vs. reference) and
plastron length (PL) on individual trace element concentrations in yellow-bellied slider
(Trachemys scripta scripta) claws. Reference sites were grouped into one site type for analysis.
Element Source df MS F p
As Site 1 18.29 29.37 <0.001
PL 1 3.052 4.90 0.030
Error 52 0.623 - -
Cd Site 1 15.203 28.48 <0.001
PL 1 9.717 18.21 <0.001
Error 52 0.534 - -
Cr Site 1 2.149 2.30 0.135
PL 1 2.478 2.65 0.109
Error 52 0.934 - -
Cu Site 1 7.307 11.60 0.001
PL 1 13.67 21.69 <0.001
Error 52 0.630 - -
Se Site 1 31.00 70.99 <0.001
PL 1 0.032 0.074 0.787
Error 52 0.437 - -
Sr Site 1 1.907 1.948 0.333
PL 1 0.936 0.956 0.169
Error 52 0.979 - -

56
Table 2.3: Two types of candidate models with varying correlation structure to explain
differences in toe web swelling response between PHA- and PBS-injections. The most
parsimonious model for each correlation structure is indicated in bold. Model terms included
treatment (PHA vs PBS; Grp), time of sampling (Time; 0 vs 24 hrs), Site (D-area vs reference),
and plastron length (PL). Correlation structures (Corr.) types included ARMA 01 (AR 1) and
none designated (None). Model outputs presented are log likelihood (LL), model degrees of
freedom (K), Akaike Information Criteria (AIC), the difference in AIC between given model and
most parsimonious model (△AIC), and Akaike weights, which displays the weight of each
model in the candidate set.
Model Corr. LL K AIC △AIC AICWt
Grp*Time + Site + PL AR 1 94.62 9 -171.24 0 0.91
Grp*Time + Site + PL None 91.28 8 -166.56 4.68 0.09
Sex + Grp*Time + Site + PL AR 1 66.17 10 -112.34 0 0.63
Sex + Grp*Time + Site + PL AR 2 66.50 11 -111.00 1.34 0.32
Sex + Grp*Time + Site + PL None 62.73 9 -107.47 4.87 0.05

57
Table 2.4: Prevalence (proportion of individuals infected) and parasitemia levels (± 1 SE) of
hemogregarines in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and
reference areas on the Savannah River Site, South Carolina. For all blood smears, approximately
120 fields were observed to examine 8,000 cells per turtle at 1000x magnification. Plastron
length and parasitemia ranges are reported below means in parenthesis. PL = plastron length, and
parasitemia is equivalent to the percent of cells (of 8000) infected by hemogregarine parasites.
Site type n
No.
Infected Mean (range) PL (mm) Prevalence (%)
Mean (range)
parasitemia
Reference 18 14 155a 77.78 0.043 ± 0.02
- - (62 – 234) - (0.000 – 0.413)
D-area 27 14 125 51.85 0.030 ± 0.01
- - (42 – 226) - (0.000 – 0.163)
Total 45 28 136 62.22 0.035 ± 0.01
a Two of 18 observations did not have PL values due to sampling errors in 2014

58
Figure 2.1: D-area ash basins and their surrounding wetlands on the Savannah River Site in
west-central South Carolina. Historically, effluent containing coal ash would flow into retention
basins (R1 and R2), then as particulates settled, surface waters would move into primary and
secondary basins. Letters ‘A’ and ‘B’ denote wetlands adjacent to the primary basin that were
also sampled.

59
Figure 2.2: Relationship between plastron length (PL) and rank-transformed Claw Cd values
(analysis of covariance; PL: F1,52 = 18.21, R2 = - 0.45, p < 0.001; Site: F1,52 = 28.48, p < 0.001)
in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and reference sites on the
Savannah River Site, South Carolina.

60
Figure 2.3: Relationship between plastron length (PL) and rank-transformed Claw Cu values
(analysis of covariance; PL: F1,52 = 21.69, R2 = - 0.37, p < 0.001; Site: F1,52 = 11.60, p = 0.001)
in yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and reference sites on the
Savannah River Site, South Carolina.

61
Figure 2.4: Relationship between plastron length (PL) and rank-transformed Claw As values
(analysis of covariance; PL: F1,52 = 4.90, R2 = 0.37, p = 0.030; Site: F1,52 = 29.37, p < 0.001) in
yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and reference sites on the
Savannah River Site, South Carolina.

62
Figure 2.5: Spearman rank correlations between log blood and log claw trace element (As, Cr,
Se, and Sr) concentrations in yellow-bellied sliders (Trachemys scripta scripta). Sample sizes for
As and Se (n = 24) were lower than Cr and Sr comparisons, and this was due to detectability
issues.

63
Figure 2.6: Mean percentage of bacteria (Escherichia coli) killed (± 1 SE) by yellow-bellied
slider turtles (Trachemys scripta. scripta) from D-area (n = 39) and reference sites (n = 42) on
the Savannah River Site, South Carolina.
80
82
84
86
88
90
92
94
96
Reference D-area
Per
cent
bac
teri
a kil
led
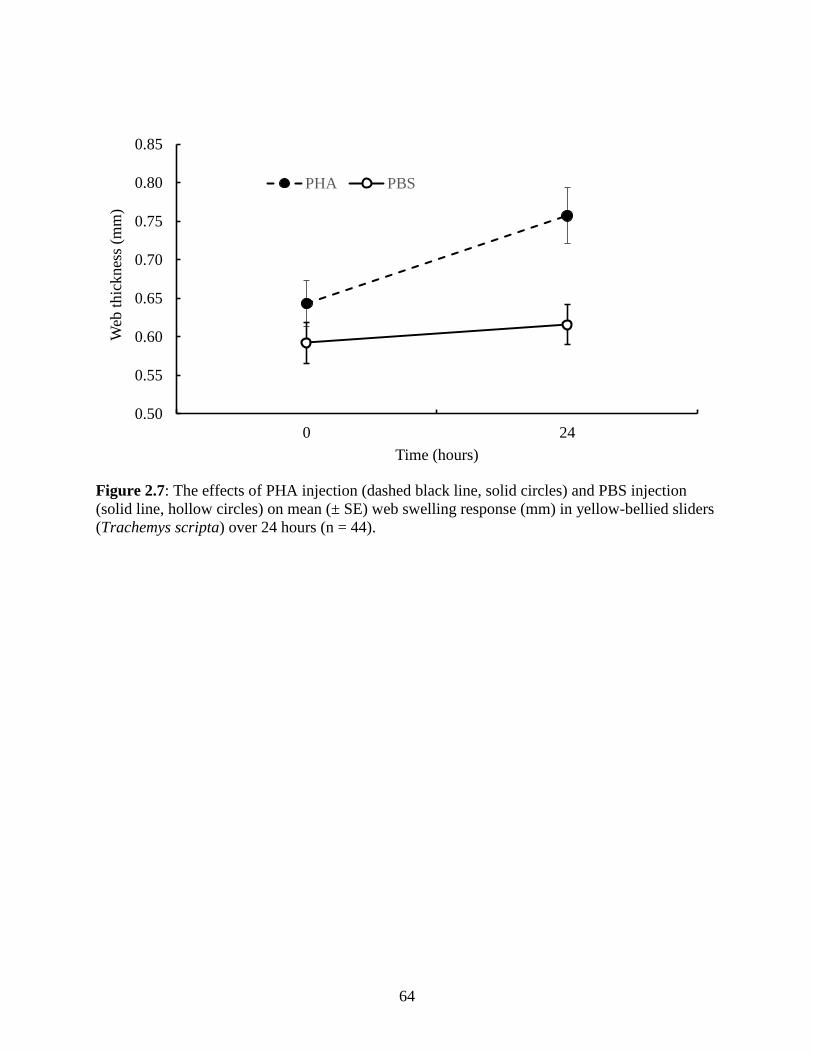
64
Figure 2.7: The effects of PHA injection (dashed black line, solid circles) and PBS injection
(solid line, hollow circles) on mean (± SE) web swelling response (mm) in yellow-bellied sliders
(Trachemys scripta) over 24 hours (n = 44).
0.50
0.55
0.60
0.65
0.70
0.75
0.80
0.85
0 24
Web
th
ick
nes
s (m
m)
Time (hours)
PHA PBS

65
Figure 2.8: Relationship between plastron length (PL) and rank-transformed parasitemia values
(analysis of covariance; PL: F1,40 = 21.87, R2 = 0.32, p < 0.001; Site: F1,40 = 0.00, p = 0.987) in
yellow-bellied slider turtles (Trachemys scripta scripta) from D-area and reference sites on the
Savannah River Site, South Carolina.

66
Figure 2.9: Correlation between log PHA response (stimulation index) and parasitemia values in
yellow-bellied slider turtles (Trachemys scripta scripta) collected during 2014-2015 from the
Savannah River Site, South Carolina (PHA responses; p < 0.001, n = 15).

67
CHAPTER 3
EFFECTS OF SELENIUM ON THE HEMATOLOGY, INNATE IMMUNITY, AND
METABOLIC RATE OF YELLOW-BELLIED SLIDERS
Introduction
Selenium (Se) is a well-known trace element contaminant that also is an essential
micronutrient in many vertebrate species (Young et al. 2010). For almost a century and a half
(1817-1957) Se was considered to be only a toxic trace element (Wisniak 2000). Selenium is also
intriguing because there is a narrow range between the element’s essentiality and toxicity (Janz
et al. 2010). The essentiality of Se is best understood in mammalian species, where it is now
known that Se is necessary for certain proteins and enzymes (e.g., glutathione peroxidases,
thioredoxin reductases) to function properly (Bellinger et al. 2010). In mammals, specific
selenoproteins provide protection from reactive oxygen species, assist in detoxification and Se
transport, and play a key role in thyroid hormone metabolism (inactivation/activation of
iodothyronine deiodinases) (Chapman et al. 2009; Bellinger et al. 2010; Janz et al. 2010).
Selenium in the environment can originate from both natural and anthropogenic sources.
Global seleniforous deposits are associated with ancient marine basins (Presser et al. 2004).
While natural weathering of these rock formations does slowly release Se, the movement of Se
into the environment has been greatly accelerated by anthropogenic activities. Coal combustion,
atmospheric deposition, various types of mining, agricultural runoff, and electronic wastes all
contribute to Se contamination within the environment (Young et al. 2010). Selenium
contamination of aquatic environments is of particular concern. When Se enters these

68
ecosystems, inorganic Se species are the most common form present (i.e., selenate, SeO32- and
selenite, SeO42; Fan et al. 2002). However, the most common species of Se that accumulates
within wildlife is the organic form, selenomethionine (SeMet), and its toxicity is well-
documented in oviparous vertebrate species (Young et al. 2010). Selenium is readily transformed
to SeMet and other organic forms (i.e., selenocysteine) by algae, bacteria, and plants, which are
then transferred through the food web. Selenomethionine is the main organic form of Se in the
environment and its toxicity to birds and fish is well-documented (Chapman et al. 2009).
Selenium can accumulate within vertebrates through trophic, maternal, and
environmental exposure (Fan et al. 2002; Ohlendorf 2003; Young et al. 2010). One of the most
sensitive endpoints of Se exposure in wildlife is reproductive toxicity in egg-laying vertebrates.
For instance, the Se-contamination of Kesterson Reservoir (CA, USA) and other reservoirs in the
USA provide classic examples of selenium’s deleterious effects on reproduction. At the
Kesterson Reservoir, Se-enriched agricultural runoff and the subsequent accumulation of Se led
to impaired reproduction of avian species (Ohlendorf et al. 1986a; Ohlendorf et al. 1986b).
Selenium’s adverse effects are not limited to reproduction. Exposure to SeMet and sodium
selenite can lead to impaired immune responses in mallards (Fairbrother and Fowles 1990).
Oxidative stress is another sublethal effect that is commonly affiliated with Se exposure in
vertebrates (Chapman et al 2009). The exact mechanism of SeMet toxicity in vertebrates is still
unknown, however, it is hypothesized that toxicity originates from increased cellular oxidative
stress and possibly improper protein substitutions (Janz et al. 2010).
Reptilian species whose life histories tie them to aquatic environments are vulnerable to
Se contamination. Many reptiles, such as turtles and crocodilians, are at risk of Se exposure
because they are long-lived and feed at upper trophic levels within their respective ecosystems

69
(Rowe et al. 2008). The effects of Se on vertebrates have been well documented in birds and fish
(Janz et al. 2010); however, field and laboratory studies of the effects of Se on reptiles are
underrepresented (Young et al. 2010). Research conducted in aquatic settling basins (coal ash)
on the Savannah River Site (SRS; Aiken, SC, USA) previously found that Se is maternally
transferred in yellow-bellied sliders (Trachemys scripta; Nagle et al. 2001). Mothers from Se-
affected areas accumulated significant amounts of Se and hatchlings of contaminated mothers
exhibited lower metabolic rates than reference conspecifics. Another study in the same SRS
aquatic settling basins found that water snakes captured with elevated Se burdens exhibited
increased standard metabolic rates (Hopkins et al. 1999). Rich and Talent (2009) found that
dietary exposure to Se in the leopard gecko (Eublepharis macularius) led to reduced growth and
reduced food consumption. All other available laboratory studies suggest a lack of adverse
effects for the banded water snake (Nerodia fasciata), brown house snake (Lamprophis
fulginosis), and western fence lizards (Sceloporus occidentalis) exposed to concentrations of
dietary SeMet ranging 11-23 mg/kg Se (concentrations based on dry weight of prey provided;
Hopkins et al. 2001; 2002; 2004; 2005). All endpoints assessed within these studies fell within
normal ranges. However, in the Hopkins et al. (2002) study, further analyses of water snake
tissues found histological abnormalities associated with contaminant exposure (Ganser et al.
2003).
The objective of the current study was to examine the effects of SeMet exposure in a
freshwater turtle species, the yellow-bellied slider (Trachemys scripta scripta). We exposed
juvenile T. scripta to SeMet via oral gavage for 6 weeks (5 total doses). We quantified mortality,
Se accumulation in multiple tissue types (blood, kidney, liver, muscle), metabolic rates, and

70
immunologic status. To assess possible immunological effects of Se exposure, we measured
bactericidal capacity and hematological profiles.
Methods
Study species
The yellow-bellied slider (Trachemys scripta scripta) is a subspecies of pond slider
belonging to the family Emydidae. Sliders are common throughout the United States and, in
habitats where they occur, can represent the largest proportion of turtle biomass relative to other
species (Congdon et al. 1986). These turtles can regularly be found in a diversity of wetland
types that have plentiful aquatic vegetation, basking structures, and adjacent open terrestrial
habitats for nesting (Gibbons 1990a, Spotila et al. 1990). Sliders are opportunistic omnivores
and are known to consume a plethora of aquatic prey, including plants, vertebrates, and
invertebrates (Parmenter & Avery 1990). Adults tend to become more herbivorous, while
juvenile sliders are known to consume a diet higher in protein. Sliders commonly inhabit
contaminated bodies of water, and they exhibit behaviors (i.e., fidelity to a small home range,
opportunistic carnivorous feeding) that may put them at an increased risk for contaminant
exposure (Gibbons 1990b). In addition, both yellow-bellied sliders and their close relative, the
red-eared slider (Trachemys scripta elegans), are commonly used in field studies and laboratory
experiments (Burger et al. 1998, Zimmerman et al. 2010, Yu et al. 2011, Zimmerman et al.
2012), thus their ecology and physiology are well-characterized (Gibbons 1990b). This
information about sliders allows researchers to readily place findings in context with known
physiological processes.

71
Husbandry and experimental protocol
Seventy juvenile yellow-bellied slider (Trachemys scripta scripta; < 111 mm plastron
length, PL) were obtained from Concordia Turtle Farms (Jonesville, LA) in September of 2014
and were weighed (to nearest 0.1 g), measured (to nearest 1.0 mm PL), and given a unique
identification number by notching their marginal scutes (Cagle 1939). Each turtle was visually
inspected and appeared healthy (i.e., no visible injuries or abnormalities). Prior to the initation of
the study, turtles were kept in the living streams tanks inside of the SREL’s Aquatic Animal
Holding Facility (AAF). The AAF consists of a concrete block base building with
semitransparent fiberglass upper walls and roof, which allows animals housed inside to be
exposed to natural light (Hamilton et al. 2016). Each turtle was randomly assigned to one of six
270 x 56 x 57-cm living streams tanks (Frigid Units Inc., Toledo, OH, USA). Each living stream
tank was provided two 38 x 38 cm concrete basking platforms, and had at least two 100-W bulbs
that were suspended approximately 30 cm above the cement platforms. Basking lights were set
on a light/dark cycle using automatic timers (0600-1800). Living streams tanks were drained and
cleaned once per week, and turtles were fed Mazuri aquatic turtle chow (Mazuri, PMI Nutrition
International LLC, Arden Hills, MN) 3 times per week and were fed ad libitum. Turtles were
held in the living streams tanks under these conditions for 8 months to ensure acclimation to their
housing facility and the dry food chow.
In May 2015, each turtle was randomly assigned to one of three selenium (Se) treatment
groups: control (n=24), 15 mg/kg Se treatment (n=23), and 30 mg/kg treatment group (n=23).
Turtles were individually housed in ~ 68 L totes (Sterilite Corporation, Clinton, SC) to prevent
confounding interactions among individuals and to allow each individual to represent a true
replicate. Turtles were distributed randomly among three separate stalls in the AAF. Stalls were

72
considered blocks to attempt to control for variation in environmental conditions among stalls.
Turtles were given wood basking platforms that were anchored to the wall of each tote, and were
maintained under ambient temperatures of 25-32ºC. Each turtle tote was cleaned 3-4 times per
week and water was replaced at each cleaning. Turtles were fed aquatic turtle chow 3 times per
week and were fed ad libitum.
Selenium exposure via oral gavage
Originally, we aimed to expose turtles to Se for eight weeks, but due to mortalities in the highest
treatment group, we terminated the study after the fifth week. Juvenile sliders were exposed to
seleno-L-methionine (SeMet, Sigma Aldrich, St. Louis, MO) via oral gavage for a total of five
weeks (~ 37 days). Seleno-L-methionine was used for this study because it is a naturally
occurring form of selenium that has a high bioavailability within food webs (Fan et al. 2002).
Selenium concentrations in our study also are within the range of Se concentrations reported for
contaminated prey species that slider turtles might be expected to consume (Unrine et al. 2007).
The SeMet stock solution was made by mixing deionized water with powdered SeMet to a
concentration of 5000 mg/kg SeMet. Our stock solution was made to be highly concentrated to
minimize the solution volume administered. A minute amount (<15µL) of green food coloring
was also added to the mixture to ensure that regurgitation could be detected during and after the
gavage procedure. The amount of SeMet given to each turtle was calculated based on their
individual body weight (wet weight) and estimated weekly food consumption of young turtles
(Equation 1; 4% of body weight per day, CCPA 2006).

73
Equation (1):
Dose (µL) =Turtle mass (g) * 0.28 * [SeMet]
5000 (mg/kg of stock solution)
* where [SeMet] = 15 or 30 mg/kg
Prior to the initiation of the experiment, turtles were weighed again to ensure that the dose they
received was accurate. Turtles were then weighed every two weeks to ensure that the dose of
SeMet accurately reflected their weight. Control turtles were gavaged with deionized water, and
the total volume of water was equivalent to the dosing volumes that 15 mg/kg turtles received.
Juvenile turtles were given a dose of SeMet weekly via oral gavage (Turner et al. 2011) using an
18-gauge crop feeding tube (Southpointe Surgical Supply Inc., Coral Springs, FL, USA).
Metabolic rate
A total of 8 slider turtles from each treatment group (n = 24) had their standard metabolic
rate (SMR) analyzed using a flow-through respirometry system (Field Metabolic System, FMS;
Sable Instruments, Las Vegas, NV). Metabolic data was collected after the last dose (5th
application) of SeMet was given. Turtles were held without food for a week prior to sampling to
ensure that they were post-absorptive.
To collect data, turtles were placed into Rubbermaid tupperware metabolic chambers (14
x 14 cm, 1.2 L), and were placed into a FMS metabolic cooler to minimize visual disturbances to
animals during measurements. The FMS metabolic cooler provides dark conditions and is
temperature-controlled by a programmable FMS PELT device. Individual turtles were placed
into their own Rubbermaid metabolic chamber, which contained ‘in’ and ‘out’ flow airlines.
Each metabolic trial collected data on four turtles and a baseline chamber simultaneously. All

74
individuals experienced constant airflow and flow rates ranged from 105-175 mL/min
(dependent upon animal mass in g). All metabolic data was collected between 2100 – 0900 hours
(30 June 30 – 2 July; 6 trials for all data).
Metabolic measurements were recorded sequentially for each individual at 25 and 30 C.
Turtles (4 per run) were placed into the FMS metabolic cooler 75 minutes prior to initial
measurements to acclimate turtles to their chambers and temperature. After the first
measurement at 25C, the automated FMS PELT system increased the FMS metabolic cooler
temperature to 30C. Turtles were also allowed 75 minutes to acclimate to 30C after their 25C
measurement. Measurements were taken for each individual in twenty minute intervals (for each
temperature) and five-minute baseline measurements were taken every hour. A trial would
measure the oxygen consumption and CO2 production (mL/hr) of each turtle sequentially, and
control measurements of the baseline chamber would be taken every hour to allow for
corrections due to lag and drift.
Euthanasia and sample collection
After five weekly doses, turtles were euthanized to quantify Se body burdens and to
collect additional samples. Turtles were euthanized via decapitation followed by pithing of brain
(Learly et al. 2013). To minimize lymph contamination, blood was collected from the turtles’
necks immediately following decapitation. Approximately 3 mL of whole blood was transferred
into multiple 1-mL lithium heparin tubes to prevent clotting (Becton Dickson, San Antonio, TX,
USA). Whole blood was then used to prepare 2-3 blood smears per individual, which were fixed
in high-quality methanol (VMW, Radnor, PA) for subsequent leukocyte differentials. Following
blood collection, a small aliquot of whole blood was placed into microhematocrit tubes and

75
centrifuged at 10,000 rpm for 5 minutes to obtain packed cell volume (% PCV) and total solids
(g/dL) by refractometer. Approximately 1 mL of whole blood was also centrifuged to collect
plasma samples from each individual for bactericidal assays. Any abnormalities in plasma color
were noted. Remaining whole blood (~ 1 mL) was saved for subsequent trace element analysis.
Animals were dissected and whole liver, kidney, and muscle samples were weighed to the
nearest 0.01g. After dissection, all samples were stored at -60C and saved for trace element
analysis. All sample collection procedures were approved by the University of Georgia
Institutional Animal Care and Use Committee (IACUC # A2014 08-016-Y1-A2).
Quantification of selenium burdens: Tissues and blood
Turtle tissues (liver, kidneys, muscle) were individually lyophilized and homogenized
prior to digestion and subsequent trace element analysis, which was performed in six runs using
inductively coupled plasma-mass spectrometry (ICP-MS; NexION, Perkin-Elmer, Inc.,
Waltham, MA, USA). Tissue samples were digested in 150µL of a 9:1 nitric acid (HNO3,
70%):hydrochloric acid (HCl, 12.4%) mixture prior to digestion in a Mars Express microwave
system. After digestion, samples were brought to a volume of approximately 15 mL with 18-MΩ
deionized water, and ICP-MS measured samples for Se. The values obtained from analysis were
also compared to certified reference materials (TORT-3, National Research Council of Canada,
Ottawa, ON) to ensure quality of the data. Mean percent recovery for Se in tissue samples was
98.1%. The minimum detection limit for Se in kidney, liver, and muscle samples was 0.47
mg/kg. Tissue Se concentrations are reported on a dry weight basis (mg/kg).
Blood samples were digested in 2.5 mL of trace metal grade HNO3 using microwave
digestion as described above. Once samples were digested, each sample was brought to a volume

76
of 10 mL with 18-MΩ deionized water. Blood samples were analyzed in two runs using ICP-MS.
This protocol used the same certified reference material as described for tissue samples. Mean
percent recovery in blood samples was 92.25%. Instrument detection limit for Se in blood
samples was 0.003 mg/kg. Blood Se concentrations were reported on a wet weight basis (mg/kg).
All Se values reported (including literature) were in dry weight unless noted otherwise. All tissue
samples and >95% of blood samples had Se concentrations above the instrument’s minimum
detection limit (>MDL). Samples with Se concentrations <MDL were replaced with calculated
0.5*MDL values (based on sample weight) for analysis.
Survival
Survival of turtles was monitored daily throughout the duration of the study. Proportional
survival of turtles at the conclusion of the study was compared among treatment groups. If
mortalities occurred, deceased individuals were not included in the analysis of hematological
data. Tissue accumulation data included two 30 mg/kg turtles that died after the last dose of Se.
Hematological and immunological measures
To determine the possible effects of Se exposure on bactericidal capacity of juvenile
sliders we used a bacterial killing assay (BKA). Turtle plasma was mixed with phosphate
buffered saline (PBS), a standardized aliquot of Escherichia coli (ATCC #8739; VWR, Radnor,
PA), and CO2 independent media. Fifty microliters of the mixture was spread onto agar plates
(Time 0 plates; Teknova Tryptic Soy plates, Fisher Scientific Company LLC, Suwanee, GA) and
50 μL of the same mixture was plated again one hour later (Time 60 plates). These plates were

77
incubated overnight and analyzed to determine bactericidal capacity. See Ch. 2 for further
information regarding the bactericidal assay protocol.
Methanol-fixed blood films were stained with Wright-Giemsa and blindly evaluated for
white blood cell (WBC) estimate, WBC differential (200 cells), and evaluation of blood cell
morphology by the same clinical pathologist (N. Stacy). The WBC estimate was performed by
multiplying the average number of WBC in 10 microscope fields × the objective power squared
(Weiss 1984). The erythroid regenerative response in anemic turtles (PCV ≤25%) was
subjectively evaluated by morphologic evaluation of the erythroid cell line for anisocytosis,
polychromasia, mitotic figures, and/or early erythroid stages and classified as absent, mild,
moderate, or marked. The morphologic evaluation of WBC included evaluation for heterophil
toxicity and left-shifting, among other possible morphologic abnormalities.
Statistical analyses
To compare differences in selenium accumulation among treatments groups, one-way
Analysis of Variance (ANOVAs) was used. If differences existed, Tukey’s Honest Significant
Difference (HSD) post-hoc tests were used to see which groups significantly differed. Possible
differences in bacterial killing assays among treatment groups were also tested using ANOVA.
Normality and homogeneity of variances were tested using Shapiro-test statistic and the Levene
statistic. When necessary, data were transformed using log, square root, or rank transformations
to achieve normality and meet assumptions of parametric tests. Due to the nonnormality and
heteroscedasticity of hematological and survival data, these data were compared using
nonparametric Kruskal-Wallis H tests. Significant differences found in Kruskal-Wallis analyses
were tested for multiple pair-wise differences using Dunn’s post-hoc test with Šidák-corrected

78
significance values. All hematological data were tested for correlations with selenium
concentrations in all tissue types. Blood Se data were tested for correlations with all tissue types.
Because these data did not fit the assumptions of Pearson’s correlation test, Spearman rank
correlation (Rs) tests were used.
We used ExpeData-P Data Analysis Software (Sable Systems, Las Vegas, NV, USA) to
calculate oxygen consumption data from the metabolic trials. Oxygen consumption (mL/hr) was
analyzed using a linear mixed effects model for repeated measures (R package lme). Individual
turtle ID was used as a random effect within the model. Stall was included in preliminary models
to account for the possible effect of block group but was not significant. Preliminary models also
examined an interaction term of treatment and temperature (treatment * temperature), but this
was also found to be nonsignificant. Each model investigated was fit with a different kind of
ARMA (autoregressive moving average) serial correlation structure. The final model included Se
treatment group, trial temperature, and plastron length as fixed effects, with turtle ID serving as a
random effect (Table 3.1). All data were analyzed using R (R Core Team 2016).
Results
Selenium body burdens
Accumulation of total Se in all tissue types was dependent on SeMet dose (Table 3.1).
The 30 mg/kg group exhibited the highest Se burdens, followed by 15 mg/kg and control groups.
There was a significant difference in mean Se concentrations among treatment groups (ANOVA,
p < 0.001) for all tissue types. Tukey’s post hoc test found that turtles from the 30 mg/kg group
exhibited the highest Se concentrations in all tissue types (p < 0.001 for all tissue comparisons,
Table 3.1). Blood Se accumulation also significantly differed among treatment groups (Kruskal-

79
Wallis, X 2 = 56.40, p < 0.001). Dunn’s multiple comparisons revealed that all groups differed
significantly (p < 0.005), and accumulation followed trends previously described in tissues (30
mg/kg > 15 mg/kg > control). For all treatment groups Se concentration was highest in the
kidney, followed by liver, muscle, and blood. Blood Se was strongly and positively correlated
with kidney Se (rs = 0.922, p < 0.001), liver, (rs = 0.860, p < 0.001), and muscle (rs = 0.849, p <
0.001; Table 3.2 and Figure 3.1).
Survival
Proportional survival of turtles (Table 3.3) was significantly influenced by treatment (X 2 = 8.55,
p = 0.014). The 30 mg/kg group was the only group that experienced mortality throughout the
duration of our study (82.6% survival). The first mortality occurred after the third weekly dose of
SeMet, another after the fourth weekly dose, and two more individuals died between the fifth
dose and culling of turtles.
Metabolic rates
The most parsimonious model showed that metabolic rates of turtles varied significantly
and positively with temperature (t 2,17 = 5.55, p < 0.001) and plastron length (t 2,17 = 2.28, p =
0.036; Table 3.4). Average metabolic rates of all turtles at 25 and 30°C were 6.47 ± 0.60 mL/hr
and 9.76 ± 1.01 mL/hr, respectively. Se treatment group was not a significant factor in any of our
models, although at both temperatures turtles from 15 mg/kg group tended to have lower
metabolic rates than the other two Se treatment groups (Figure 3.2; t 2,17 = -1.86, p = 0.080).

80
Hematological and immunological effects of selenium exposure
WBC estimates varied significantly among groups (X 2 = 6.96, p = 0.031) and were
higher in the 15 mg/kg group compared to control and 30 mg/kg groups (Table 3.5). Dunn’s
post-hoc test revealed that only the 15 mg/kg and control groups differed significantly (p =
0.013). Treatment groups also significantly differed in H/L ratios (X 2 = 7.83, p = 0.019). The
H/L ratios of turtles in the 30 mg/kg group were five times higher when compared to both
control and 15 mg/kg groups, due to higher heterophil and lower lymphocyte concentrations in
the 30 mg/kg group (Table 3.5). Lymphocytes were significantly (33.72 ± 3.12 x 109 cells/L)
lower in the 30 mg/kg group relative to control and 15 mg/kg groups (Table 3.5). PCV
significantly differed among treatment groups (X 2 = 14.08, p = 0.043), and turtles in the 30
mg/kg group exhibited lower PCV results when compared to the control and 15 mg/kg groups.
Fifty-five percent (10/18) of turtles from the 30 mg/kg group were anemic (PCV ≤ 25), with
PCVs averaging 18.9 ± 1.18. The 30 mg/kg group also had the highest incidence of regenerative
anemia, and one-third of turtles (6/18) had green plasma which could be suggestive of hemolytic
anemia, liver disease, or starvation. Approximately 17% (4/23) of turtles in the 15 mg/kg group
were anemic (mean PCV = 20.8 ± 2.63). Lastly, 13% (3/23) of individuals from the control
group had mild anemia (mean PCV = 24). Anemic individuals from the 15 mg/kg and control
groups mostly exhibited nonregenerative anemia. All other hematological analytes (heterophils,
monocytes, and total solids) did not significantly differ between groups (all p > 0.235), although
some did trend toward significance (absolute basophils and eosinophils). Heterophil toxicity or
left-shifting (a shift towards immaturity in white blood cells) was not observed. Thrombocytes
were adequate in all groups.

81
Spearman correlations revealed a negative relationship between kidney Se concentrations
and PCV (rs = - 0.36, p = 0.003; Figure 3.3), and also found a positive relationship between
blood Se concentrations and total solids (rs = 0.26, p = 0.035; Figure 3.4). All other correlational
comparisons of hematological data did not yield significant differences (p > 0.05). Mean percent
bacteria killed did not significantly differ among treatment groups (X 2 = 0.39, p = 0.819).
Relationships between BKA responses and other variables were not explored due to the extreme
nonnormality of the BKA data.
Discussion
The current study shows that T. scripta do accumulate SeMet in a dose-dependent
manner. Kidney Se concentrations in the current study are among the highest reported in reptiles,
if not the highest. The kidneys (renal system) are the main route of elemental elimination in
vertebrates, and turtles in all experimental groups partitioned the largest amounts of Se to their
kidneys. Correlational relationships in Se accumulation between blood and other tissue types
(kidney, liver, and muscle) were strong and positive (all p < 0.001, all Rs > 0.849), suggesting Se
burdens of sensitive organs may be estimated with non-destructive samples in T. scripta. The
significant correlation between blood and muscle Se concentrations also may assist in predicting
the risks of consuming turtle meat. The accumulation patterns (kidney > liver > muscle > blood)
of Se in this study are similar to what little information exists for other reptilian species. In
reptilian exposure studies, snake species (Nerodia and Lamprophis spp.) accumulated the highest
amounts of Se in their kidney tissues, although these differences were not as dramatic as the
amount found in the turtles’ kidneys reported in this study (Hopkins et al. 2002; 2004). However,
some hawksbill sea turtles (Eretmochelys imbricata) caught in Japanese waters had higher

82
amounts of Se in their liver relative to their kidneys (Anan et al. 2001). In contrast to reptiles,
birds are known to accumulate equal or larger amounts of Se in their liver tissues (Ohlendorf et
al. 1988; Ohlendorf and Heinz 2011). The differences in Se accumulation among turtles, snakes,
and birds could be of significance when considering these species in ecological risk assessments.
While our treatment concentrations of 15 and 30 mg/kg may reflect a chronic dose, the
weekly dose of SeMet was an acute exposure to a week’s worth of SeMet (based on turtle
weight). Although the turtles in our study were acutely exposed to SeMet, their Se concentrations
are similar to data from previous field studies. Turtles in both of our treatment groups had mean
Se liver concentrations similar to those measured in dead birds at the Kesterson Reservoir, and
liver Se concentrations of dead birds ranged from 26-86 mg/kg Se (Ohlendorf and Heinz 2011).
The highest liver Se concentrations in the 30 mg/kg group (162.01 mg/kg Se) exceeded those
reported in banded water snakes (Nerodia fasciata) in CCR settling basins on the SRS (Hopkins
et al. 1999). Turtles in the 15 mg/kg group had blood and liver Se concentrations similar to those
reported in wild T. scripta captured in CCR settling basins on the SRS. Mean liver Se
concentrations of wild T. scripta were 37.18 mg/kg (Nagle et al. 2001), while liver Se in our 15
mg/kg group averaged 34.89 mg/kg. Blood Se concentrations of wild T. scripta in my field study
(see chapter 2) were as high as 9.57 mg/kg (ww) Se, and this value falls within the ranges of both
SeMet treatment groups (Haskins et al. unpublished data; Table 3.1).
Turtles in both SeMet-treatment groups accumulated large amounts of Se in their tissues,
and the Se liver burdens described in the 30 mg/kg turtles are well above known thresholds for
lethal responses in mallards (Anas platyrhynchos; Ohlendorf and Heinz 2011). A review of
exposure studies suggests that mallard liver concentrations of > 66 mg/kg SeMet can cause
mortality (Ohlendorf and Heinz 2011). Turtles in the 30 mg/kg group had mean liver Se

83
concentrations of 86.37 mg/kg and a mortality rate of 17.4% (4/23) after 5 treatments of SeMet.
Furthermore, turtles from the 30 mg/kg group exhibited signs of selenium toxicosis (selenosis).
Ohlendorf and Heinz’s (2011) recommended assessment values for selenosis in avian species is
> 20 mg/kg liver Se, and selenosis in mallards was characterized by emaciation, hepatic lesions,
and the necrosis of maxillary nail tissues. Histopathological analysis of the 30 mg/kg turtles
revealed necrosis of maxillary nail tissues, necrosis of kidney tissues, and hepatic lesions
(Haskins et al., unpublished data). To our knowledge, this study is the first to report symptoms
suggesting selenosis in a reptilian species.
Stressors such as CCRs, which often contain elevated levels of Se, can cause
physiological changes in vertebrates that may result in increased energy expenditure (Hopkins et
al. 1999; Holliday et al. 2009). Field studies of water snakes, bullfrogs, shrimp, and crayfish
showed that all species had increased metabolic demands in response to CCR accumulation
(Rowe 1998; Rowe et al. 1998; Hopkins et al. 1999; Rowe et al. 2001). In contrast, Nagle et al.
(2001) found metabolic rates of hatchling T. scripta from CCR-exposed mothers were depressed
compared to hatchlings from unexposed females. Because these data come from individuals
exposed to a mixture of trace elements present in CCRs, it is difficult to discern if Se was the
sole cause of metabolic aberrations in these species. In the present study oxygen consumption
did not significantly differ among treatment groups. However, turtles from the 15 mg/kg group
did have lower oxygen consumption rates than any group, and this difference trended toward
significance (p = 0.080). These trends appear to agree with Nagle et al.’s (2001) findings in T.
scripta, but it is strange that the 30 mg/kg group did not experience metabolic aberrations.
Although metabolic rates did not differ at the conclusion of the 37-d experiment, it could be that
the turtles were not exposed long enough to elucidate the changes in metabolic status.

84
Diamondback terrapins (Malaclemys terrapin) exposed to polychlorinated biphenyls (PCBs) did
not show a significant decrease in metabolic status until 90 days after exposure (Holliday et al.
2009). Perhaps if individuals in the 15 mg/kg were to be chronically exposed (8-12 weeks) a
larger difference would be observed. It is also possible that our sampling period did not observe
the initial stress response in the 30 mg/kg group, and that these turtles were so stressed by Se that
they could not regulate their metabolic response. Lastly, it is important to note that sampling
errors could have impacted our data such as spontaneous animal activity during sampling, which
is common when measuring metabolic rates in ectotherms.
Selenium accumulation had a significant influence on numerous hematological
parameters including packed cell volume, total white blood cell counts, lymphocyte counts, and
heterophil to lymphocyte ratios. Packed cell volume decreased with increasing Se dose. Previous
studies exposed broiler chickens to inorganic Se (selenite) or SeMet, and chickens that were
supplemented with SeMet had a significantly lower hematocrit count than birds that were fed
selenite (da Silva et al. 2010). Fish that were exposed to Se contamination through coal ash
wastewater also had lower hematocrit values than fish from reference sites (Lemly 2002). The
mean PCV value (24.33 ± 1.67) for 30 mg/kg turtles is below the threshold of mild anemia (<
25), and over half (55%) of the 30 mg/kg turtles were anemic. However, total solids were
actually higher in the 30 mg/kg turtles. It is interesting that the 30 mg/kg turtles were anemic but
did not show a significant change in total solids, but this observation could be due to hemolysis.
One-third (6/18) of turtles in the 30 mg/kg group presented with green plasma; this observation,
combined with the significant decreases in PCV, could suggest Se-induced hemolytic anemia.
Studies in mammals have shown that hemolysis can alter hematological parameters including
albumin and alpha and beta-globulins, which are among the proteins that are quantified when

85
measuring total solids (Di Martino et al. 2015). Therefore, it is possible that the total solids
measured in this study were impacted by hemolysis.
The higher WBC count in the 15mg/kg group this cannot be attributed to heterophilia, as
the 30 mg/kg group is the only group that experienced an increase in heterophils. Although the
mean heterophil difference among groups was not statistically significant, turtles from the 30
mg/kg group did have elevated heterophil counts (Figure 3.6). Turtles in the 30 mg/kg group also
exhibited lymphopenia. Lymphopenia is known to occur when reptiles are stressed,
immunosuppressed, or are malnourished (Campbell 2012). Because most Se-treated turtles
continued to consume food throughout the study and discernable weight loss was not readily
observed, it is unlikely that malnourishment would have caused lymphopenia in the 30 mg/kg
group without affecting the 15 mg/kg group. Studies in birds have found that exposure to
excessive amounts of dietary Se can cause lymphopenia and cause atrophy in the thymus (Peng
et al. 2011). T-lymphocytes are produced in the thymus and are involved in cell-mediated
immunity and cytokine responses (Weiser 2012); thus, it could be that damage to the thymus
reduced T-lymphocyte counts in the peripheral blood of our turtles. Unfortunately, we did not
investigate Se-related effects on the thymus so we cannot speak to the possibility that Se
exposure damaged the turtles’ thymuses, leading to lymphopenia.
Heterophil to lymphocyte ratios were about five times higher in the 30 mg/kg group
relative to the other two groups in this study. These findings are expected, as the 30 mg/kg turtles
experienced heterophilia and lymphopenia. Stress can alter hematological profiles and
subsequent H/L ratios. In the current study all turtles were handled, processed, and sacrificed in
an identical manner, which suggests it is unlikely these actions would have altered only one
group’s hematological profile. Heterophil to lymphocyte ratios have been suggested to be a

86
useful biomarker to measure stress in avian species (Gross and Seigel 1983). Hormones
produced (glucocorticoids) during adrenal stress responses increase heterophil counts and reduce
lymphocyte counts (Gross and Seigel 1983; Davis et al. 2008). Studies in avian species have
shown that exposure to trace element contaminants (arsenic, boron, Se) can result in increased
H/L ratios (Grassman 2002). Furthermore, Keller et al. (2004) found that H/L ratios were
significantly elevated in loggerhead sea turtles, and these ratios were positively correlated with
organochlorine contaminant burdens. The H/L ratios of the 30 mg/kg group indicates that these
turtles also had high glucocorticoid levels, suggesting that prolonged acute exposure to Se causes
considerable stress in T. scripta.
Surprisingly, turtles across all treatment groups displayed strong bactericidal capacity (all
means ≥ 98%). BKA is a useful tool to measure a turtle’s innate immune capacity in response to
microbial invaders. The assay measures the innate immune response that involves phagocytes,
opsonizing proteins, and natural antibodies (French et al. 2010). Studies in reptiles and
amphibians have shown that anthropogenic stress, activity patterns, and time of year can all
impact bactericidal response (French et al. 2010; Brown and Shine 2014; Zimmerman et al.
2010). Although the turtles in this experiment were exposed to potentially lethal amounts of
SeMet, their bactericidal capacity did not seem to be affected by the SeMet treatments. Bacteria
are tolerant of heavy metal exposure and are known to be extremely tolerant to Se-exposure
(Janz et al. 2010). Therefore, it is unlikely that the bacteria used in the BKAs were killed by Se
exposure (through the SeMet turtles’ plasma). Our results suggest that the innate bactericidal
abilities of our turtles were not compromised by Se accumulation.
This experiment is the first to induce selenosis in a reptile. Turtles exposed to a Se-dose
of 30 mg/kg had a mortality rate of 17.4%. Turtles in the 30 mg/kg group also exhibited

87
lymphopenia, heterophilia, increased H/L ratios, and a significant decrease in packed cell
volume. The hematological findings of this study support evidence provided by previous Se
exposure experiments in vertebrates (fish and birds). Surprisingly, we did not observe
statistically significant changes in metabolic status or bactericidal capacity. The current
knowledge of Se toxicity in reptiles is insufficient (Young et al. 2010), and further research is
necessary to understand what sublethal impacts are caused by Se exposure. Turtles, like T.
scripta, are known to inhabit Se-contaminated areas such as coal ash basins on the SRS (Nagle
2001). While the Se burdens in our 30 mg/kg group would likely not reflect wild T. scripta Se
burdens, these reptiles are year-round residents in these contaminated systems, and little is
known how Se and other CCRs impact their overall health. Future controlled studies should aim
to investigate chronic, sublethal exposure to Se in chelonians and should also seek to study the
potential impacts of Se exposure on chelonian reproduction.

88
Literature Cited
Anan, Y., T. Kunito, I. Wantanabe, H. Sakai, S. Tanabe. 2001. Trace element accumulation in
hawksbill turtles (Eretmochelys imbricata) and green turtles (Chelonia mydas) from
Yaeyama Islands, Japan. Environmental Toxicology and Chemistry 20:2802-2814.
Bellinger, F.P., A.V. Raman, M.A. Reeves, and M.J. Berry. 2010. Regulation and function of
selenoproteins in human disease. Biochemical Journal 422:11-22.
Brown, G.P., and R. Shine. 2014. Immune response varies with rate of dispersal in invasive cane
toads (Rhinella marina). PLoS ONE 9:e99734. doi:10.1371/journal.pone.0099734.
Burger, J., C. Carruth-Hinchey, J. Ondroff, M. McMahon, J.W. Gibbons, and M. Gochfield.
1998. Effects of lead on behavior, growth, and survival of hatchling slider turtles. Journal
of Toxicology and Environmental Health, Part A: Current Issues 55:495-502.
Cagle, F.W. 1939. A system of marking turtles for future identification. Copeia 1939:170-173.
Campbell, T.W. Hematology of reptiles. 2012. Pages 277-297 in Thrall, M.A., G. Weiser, R.W.
Allison, and T.W. Campbell, editors. Veterinary Hematology and Clinical Chemistry.
Wiley-Blackwell, Ames, IA, USA.
CCPA. 2006. Recommendations of the CCAC: Species-specific recommendations on
amphibians and reptiles. Canadian Council of Animal Care. Ottawa, Ontario.
Chapman, P.M., W.J. Adams, M.L. Brooks, C.G. Delos, S.N. Luoma, W.A. Maher, H.M.
Ohlendorf, T.S. Presser, and D.P. Shaw. 2009. Ecological assessment of selenium in the
aquatic environment: Summary of a SETAC Pellston Workshop. Pensacola, FL, USA.
Society of Environmental Toxicology and Chemistry.

89
Congdon, J.D., J.L. Greene, and J.W. Gibbons. 1986. Biomass of freshwater turtles: a geographic
comparison. American Midland Naturalist. 115:165-173.
da Silva, I.C.M., A.M.L. Riberio, C.W. Canal, L. Trevizan, M. Macagnan, T.A. Gonçalves,
N.R.C. Hlavac, L.L. de Almeida, and R.A. Pereira. 2010. The impact of organic and
inorganic selenium on the immune system of growing broilers submitted to immune
stimulation and heat stress. Brazilian Journal of Poultry Science 12:247-254.
Davis, A.K., D.L. Maney, and J.C. Maerz. 2008. The use of leukocyte profiles to measure stress
in vertebrates: a review for ecologists. Functional Ecology 22:760-772.
Di Martino, G., A.L. Stefani, G. Lippi, L. Gagliazzo, W. McCormick, G. Gabai, and L. Bonfanti.
2015. The degree of acceptability of swine blood values at increasing levels of hemolysis
evaluated through visual inspection versus automated quantification. Journal of
Veterinary Diagnostic Investigation 27:306-312.
Fairbrother, A., and J. Fowles. 1990. Subchronic effects of sodium selenite and selenomethionine
on several immune functions in mallards. Archives of Environmental Contamination and
Toxicology. 19:836-844.
Fan, T.W.M., S.J. Teh, D.E. Hinton, R.M. Higashi. 2002. Selenium biotransformations into
proteinaceous forms by foodweb organisms of selenium-laden drainage waters in
California. Aquatic Toxicology 57:65-84.
Finley, K.A. 1985. Observations of bluegills fed selenium-contaminated Hexagenia nymphs
collected from Belews Lake, North Carolina. Bulletin of Environmental Contamination
and Toxicology 35:816-825.

90
French, S.S., D.F. DeNardo, T.J. Greives, C.R. Strand, and G.E. Demas. 2010. Human
disturbance alters endocrine and immune responses in Galapagos marine iguanas
(Amblyrhynchus cristatus). Hormones and Behavior 58:792-799.
Ganser, L.R., W.A. Hopkins, L. O’Neil, S. Hasse, J.H. Roe, and D.M. Sever. 2003. Liver
histopathology of the southern watersnake, Nerodia fasciata fasciata, following exposure
to chronic trace element-contaminated prey from a coal ash disposal site. Journal of
Herpetology 37:219-226.
Gibbons, J.W. 1990a. The Slider Turtle. Pages 1-18 in Sheffield, R., editor. Life History and
Ecology of the Slider Turtle. Smithsonian Institution Press, Washington, D.C., USA.
Print.
Gibbons, J.W. 1990b. Turtle Studies at SREL: A research perspective. Pages 19-45 in Sheffield,
R., editor. Life History and Ecology of the Slider Turtle. Smithsonian Institution Press,
Washington, D.C., USA.
Grasman, K.A. 2002. Assessing immunological function in toxicological studies of avian
wildlife. Integrative and Comparative Biology 42:34-42.
Gross, W.B., and H.S. Siegel. 1983. Evaluation of the heterophil/lymphocyte ratio as a measure
of stress in chickens. Avian Diseases 27:972-979.
Hamilton, M.T., J.W. Finger Jr., M.E. Winzeler, and T.D. Tuberville. 2016. Evaluating the effect
of sample type on American alligator (Alligator mississippiensis) analyte values in a
point-of-care blood analyzer. Conservation Physiology 4:1-7.
Hamilton, S.J. 2004. Review of selenium toxicity in the aquatic food chain. Science of the Total
Environment 326:1-31.

91
Heinz, H.G., and C.J. Sanderson. 1990. Avoidance of selenium-treated food by mallards.
Environmental Toxicology and Chemistry 9:1155-1158.
Holliday, D.K., A.A. Elskus, and W.M. Roosenburg. 2009. Impacts of multiple stressors on
growth and metabolic rate of Malaclemys terrapin. Environmental Toxicology and
Chemistry 28:338-345.
Hopkins, W.A., C.L. Rowe, and J.D. Congdon. 1999. Elevated trace element concentrations and
standard metabolic rates in banded water snakes (Nerodia fasciata) exposed to coal
combustion wastes. Environmental Toxicology and Chemistry 18:1258-1263.
Hopkins, W.A., J.H. Roe, J.W. Snodgrass, B.P. Jackson, D.E. Kling, C.L. Rowe, and J.D.
Congdon. 2001. Nondestructive indices of trace element exposure in squamate reptiles.
Environmental Pollution 115:1-7.
Hopkins, W.A., J.H. Roe, J.W. Snodgrass, B.P. Staub, B.P. Jackson, and J.D. Congdon. 2002.
Effects of chronic dietary exposure to trace elements on banded water snakes (Nerodia
fasciata). Environmental Toxicology and Chemistry 21:906-913.
Hopkins, W.A., B.P. Staub, J.A. Baionno, B.P. Jackson, J.H. Roe, and N.B. Ford. 2004. Trophic
and maternal transfer of selenium in brown house snakes (Lamprophis fuliginosus).
Ecotoxicology and Environmental Safety 58:285-293.
Hopkins, W.A., B.P. Staub, and J.A. Baionno. 2005. Transfer of selenium from prey to predators
in a simulated terrestrial food chain. Environmental Pollution 134:447-456.

92
Janz, D.M., D.K. DeForest, M.L. Brooks, P.M. Chapman, G. Gilron, D. Hoff, W.A. Hopkins,
D.O. McIntyre, C.A. Mebane, V.P. Palace, J.P. Skorupa, and M. Wayland. 2010.
Selenium toxicity to aquatic organisms. Pages 139-230 in Chapman, P.M., W.J. Adams,
M.L. Brooks, C.G. Delos, S.N. Luoma, W. Maher, H.M. Ohlendorf, T.S. Presser, and P.
Bradshaw, editors. Ecological Assessment of Selenium in the Aquatic Environment.
Pensacola, FL, USA: Society of Environmental Toxicology and Chemistry.
Julius, A.D., M.H. Davies, and O.F. Birt. 1983. Toxic effects of dietary selenium in the Syrian
hamster. Annals of Nutrition and Metabolism 27:296-305.
Keller, J.M., J.R. Kucklick, M.A. Stamper, C.A. Harms, P.D. McClennan-Green. 2004.
Association between organochlorine contaminant concentrations and clinical health
parameters in loggerhead sea turtles from North Carolina, USA. Environmental Health
Perspectives 112:1074-1079.
Leary, S., W. Underwood, R. Anthony, S. Cartner, D. Corey, T. Grandin, C. Greenacre, S.
Gwaltney-Brant, M.A. McCrackin, R. Meyer, D. Miller, J. Shearer, R. Yanong. 2013.
AVMA Guidelines for the Euthanasia of Animals: 2013 edition. American Veterinary
Medical Association, Schaumburg, IL, USA.
Lemly, A.D. 2002. Symptoms and implications of selenium toxicity in fish: the Belews Lake
case example. Aquatic Toxicology 57:39-49.
Lockard, L., C.L. Rowe, and A. Heyes. 2013. Dietary selenomethionine exposure induces
physical malformations and decreases growth and survival to metamorphosis in an
amphibian (Hyla chrysoscelis). Archives of Environmental Contamination and
Toxicology 64:504-513.

93
Nagle, R.D., C.L. Rowe, and J.D. Congdon. 2001. Accumulation and selective maternal
transfer of contaminants in the turtle Trachemys scripta associated with coal ash
deposition. Archives of Environmental Contamination and Toxicology 40:531-536.
Ohlendorf, HM. 2003. Ecotoxicology of selenium. In: Hoffman DJ, Rattner BA, Burton GA,
Cairns J, editors. Handbook of Ecotoxicology. Lewis Publishers, Boca Raton, FL. Pp.
465-500.
Ohlendorf, H.M., and G.H. Heinz. 2011. Selenium in birds. Pages 668-701 in Beyer, W.N., and
J.P. Meador, editors. Environmental Contaminants in Biota. CRC Press, Boca Raton, FL,
USA.
Ohlendorf, H.M., Hoffman, H.J., Saiki, M.K., and Aldrich, T.W. 1986a. Embryonic mortality
and abnormalities of aquatic birds: apparent impacts by selenium from irrigation drain-
water. Science of the Total Environment 52:49-63.
Ohlendorf, H.M., Hothem, R.L., Bunck, C.M., Moore, J.F., and Alridch T.W. 1986b.
Relationships between selenium concentrations and avian reproduction. Transactions of
the North American Wildlife and Natural Resources Conference 51:330-342.
Ohlendorf, H.M., A.W. Kilness, J.L. Simmons, R.K. Stroud, D.J. Hoffman, and J.F. Moore.
1988. Selenium toxicosis in wild aquatic birds. Journal of Toxicology and Environmental
Health 24:67-92.
Parmenter, R.R., and H.W. Avery. 1990. The feeding ecology of the slider turtle. Pages 257-266
in Sheffield, R., editor. Life History and Ecology of the Slider Turtle. Smithsonian
Institution Press, Washington, D.C., USA.

94
Peng, X., C. Hengmin, Y. Cui, J. Deng, Z. Zuo, and J. Fang. 2011. Lesions of thymus and
decreased percentages of the peripheral blood T-cell subsets in chickens fed on diets
excess in selenium. Human and Experimental Toxicology 0:1-7.
Presser, T.S., D.Z. Piper, K.J. Bird, J.P. Skorupa, S.J. Hamilton, S.J. Detwiler, and M.A.
Huebner. 2004. The phosphoria formation: A model for forecasting global selenium
sources to the environment. Pages 299-319 in Hein, J.R., editor. Life Cycle of the
Phosphoria Formation: From Deposition to Post-Mining Environment. Elsevier B.V.,
Philadelphia, PA, USA.
R Core Team. 2016. R: a language and environment for statistical computing. R Foundation for
Statistical Computing, Vienna, Austria. <http://R-project.org>.
Rich, C.N, and L.G. Talent. 2009. Soil ingestion may be an important route for the uptake of
contaminants by some reptiles. Environmental Toxicology and Chemistry 28:311-315.
Rowe, C.L. 1998. Elevated standard metabolic rate in a freshwater shrimp (Palaemonetes
paludosus) exposed to trace element-rich coal combustion waste. Comparative
Biochemistry and Physiology 121:299-304.
Rowe, C.L. 2008. The calamity of so long life: life histories, contaminants, and
potential emerging threats to long-lived vertebrates. Bioscience. 58:623-631.
Rowe, C.L., O.M. Kinney, R.D. Nagle, and J.D. Congdon. 1998. Elevated maintenance costs in
an anuran (Rana catesbeiana) exposed to a mixture of trace elements during the
embryonic and early larval periods. Physiological Zoology 71:27-35.

95
Rowe, C.L., W.A. Hopkins, C. Zehnder, and J.D. Congdon. 2001. Metabolic costs incurred by
crayfish (Procambarus acutus) in a trace element-polluted habitat: further evidence of
similar responses among diverse taxonomic groups. Comparative Biochemistry and
Physiology 129:275-283.
Spotila, J.R., R.E. Foley, and E.A. Standora. 1990. Thermoregulation and Climate Space of the
Slider Turtle. Pages 288-298 in Sheffield, R., editor. Life History and Ecology of the
Slider Turtle. Smithsonian Institution Press, Washington, D.C., USA.
Turner, P.V., T. Brabb, C. Pekow, and M.A. Vasbinder. 2011. Administration of substances to
laboratory animals: Routes of administration and factors to consider. Journal of the
American Association for Laboratory Animal Science 50:600-613.
Unrine, J.M., W.A. Hopkins, C.S. Romanek, and B.P. Jackson. 2007. Bioaccumulation of trace
elements in omnivorous amphibian larvae: Implications for amphibian health and
contaminant transport. Environmental Pollution 149:182-192.
Wisniak, J. 2000. Jons Jacob Berzelius: A guide to the perplexed chemist. The Chemical
Educator 5:343-350.
Weiser, G. 2012. Introduction to leukocytes and the leukogram. Pages 118-122 in Thrall, M.A.,
G. Weiser, R.W. Allison, and T.W. Campbell, editors. Veterinary Hematology and
Clinical Chemistry. Wiley-Blackwell, Ames, IA, USA.
Weiss, D.J. 1984. Uniform evaluation and semiquantitative reporting of hematologic data in
veterinary laboratories. Veterinary Clinical Pathology 13:27-31.

96
Young, T., K. Finley, W. Adams, J. Besser, W. Hopkins, D. Jolley, E. McNaughton, T. Presser,
P. Shaw, and J. Unrine. 2010. What you need to know about selenium? Pages 7-46 in
Chapman, P.M., W.J. Adams, M.L. Brooks, C.G. Delos, S.N. Luoma, W.A. Maher, H.M.
Ohlendorf, T.S. Presser, and D.P. Shaw, editors. Ecological Assessment of Selenium in
the Aquatic Environment. SETAC Press, Pensacola, FL, USA. ISBN 978-1-4398-2677-5.
Yu, S., R.S. Halbrook, and D.W. Sparling. 2011. Metal accumulation and evaluation of effects in
a freshwater turtle. Ecotoxicology 20:1801-1812.
Zimmerman, L.M., R.T. Paitz, S.G. Clairardin, L.A. Vogel, and R.M. Bowden. 2010. Variation
in seasonal patterns of innate and adaptive immunity in the red-eared slider (Trachemys
scripta). Journal of Experimental Biology 213:1477-1483.
Zimmerman, L.M., R.T. Paitz, S.G. Clairardin, L.A. Vogel, and R.M. Bowden. 2012. No
evidence that estrogens affect the development of the immune system in the red-eared
slider turtle (Trachemys scripta elegans). Hormones and Behavior. 62:331-336

97
Table 3.1: Differences in selenium accumulation in target tissues (liver, kidney, muscle, and
blood) of Trachemys scripta exposed to one of three selenium treatments (Control, 15 mg/kg,
and 30 mg/kg). Selenium concentrations are reported as means (dry weight for tissues and wet
weight for blood) ± 1 SE (range). Letters in the ‘Tukeys’ column designate significant
differences among groups within a tissue type.
Tissue Treatment n Se (mg/kg) F-value, df, p valuea Tukeysb
Liver Control 23* 2.04 ± 0.12 501.8, 2, < 0.001 A
(1.16 – 3.31)
15 mg/kg 23 34.89 ± 3.09 B
(15.08 – 66.18)
30 mg/kg 21 86.37 ± 8.03 C
(25.85 – 162.01)
Kidney Control 24 4.38 ± 0.12 2733, 2, < 0.001 A
(3.00 – 5.48)
15 mg/kg 23 122.74 ± 5.54 B
(73.75 – 180.77)
30 mg/kg 21 229.30 ± 0.12 C
(141.60 – 306.33)
Muscle Control 24 1.90 ± 0.07 1322, 2, < 0.001 A
(1.41 – 2.75)
15 mg/kg 23 27.70 ± 1.40 B
(13.50 – 39.92)
30 mg/kg 21 42.54 ± 2.10 C
(26.56 – 64.75)
Bloodc Control 24 0.29 ± 0.02 56.40, 2, < 0.001c Ac
(0.09 – 0.39)
15 mg/kg 23 6.76 ± 0.48 B
(2.77 – 10.89)
30 mg/kg 21 14.85 ± 0.90 C
(7.68 – 22.36) aOne-way analysis of variance (ANOVA) b Results from post-hoc Tukey’s (adjusted) Honest Significant Difference tests. cStatistical analysis of blood used nonparametric methods (Kruskal-Wallis H test) and Dunn’s
Multiple Comparisons Tests using Šidák-corrected significance values for post-hoc analysis.
* One less sample was available for ‘control’ liver samples due to processing errors

98
Table 3.2: Proportional survival of yellow-bellied sliders (Trachemys scripta scripta) exposed to
three different treatments (control, 15 mg/kg, and 30 mg/kg) of selenium (Se). Chi-square values
reported are from a Kruskal-Wallis analysis to compare proportional survival of treatment groups
at the end of the study (37 days). ‘Dunn’ letters designate significant differences between groups.
Treatment n Proportional Survival X 2 , df, p-value Dunna
12 days 24 days 37 days
Control 24 100 100 100a 8.55, 2 , 0.014 A
15 mg/kg 23 100 100 100a A
30 mg/kg 23 100 95.7 82.6b B
a Results from post-hoc Dunn’s tests using using Šidák-corrected significance values

99
Table 3.3: Linear mixed model for metabolic rate analysis with varying correlation structure to
explain differences in metabolic rate among selenium exposure groups. The most parsimonious
model is indicated in bold. Model terms included treatment group (Control vs. 15 mg/kg vs. 30
mg/kg), trial temperature at time of sampling (25°C vs. 30°C), and plastron length (PL in mm).
Correlation structure (Corr.) types included ARMA 1 (AR 1), ARMA 2 (AR 2), and none
designated (None). Model outputs presented are log likelihood (LL), model degrees of freedom
(K), Akaike Information Criteria (AIC), the difference in AIC between given model and most
parsimonious model (△AIC), and Akaike weights, which displays the weight of each candidate
model in the set.
Model Corr. LL K AIC △AIC AICWt CumWt
Treatment + Temp + PL None -89.26 7 192.52 0 0.67 0.67
Treatment + Temp + PL AR 1 -89.26 8 194.52 2 0.24 0.91
Treatment + Temp + PL AR 2 -89.26 9 196.52 4 0.09 1

100
Table 3.4: Hematological comparisons of Trachemys scripta exposed to one of three selenium
treatments (Control, 15 mg/kg, and 30 mg/kg). Indices include white blood cell estimate (WBC),
heterophil (HET) to lymphocyte (LYM) ratios (H/L), heterophils (HET), lymphocytes (LYM),
monocytes (MON), eosinophils (EOS), basophils (BAS), packed cell volume (PCV), and total
solids (TS). ‘Dunn’ letters designate significant differences between groups.
Indices Treatment n Meana Mean % count X 2, df, p-value Dunnb
WBC Control 24 9.24 ± 0.45 - 6.96, 2, 0.031 A
15 mg/kg 23 12.07 ± 0.80 - B
30 mg/kg 18 10.03 ± 1.23 - A
H/L Control 24 0.28 ± 0.07 - 7.83, 2, 0.019 A
15 mg/kg 23 0.25 ± 0.05 - A
30 mg/kg 18 1.36 ± 0.45 - B
HET Control 24 1.05 ± 0.21 10.88 ± 1.77 2.49, 2, 0.286 -
15 mg/kg 23 1.21 ± 0.20 9.87 ± 1.56 -
30 mg/kg 18 2.93 ± 0.85 27.78 ± 5.36 -
LYM Control 24 4.30 ± 0.28 46.92 ± 2.63 14.08, 2, <0.001 A
15 mg/kg 23 5.26 ± 0.34 45.13 ± 2.45 A
30 mg/kg 18 3.10 ± 0.43 33.72 ± 3.12 B
MON Control 24 0.43 ± 0.06 4.54 ± 0.54 1.17, 2, 0.558 -
15 mg/kg 23 0.51 ± 0.06 4.65 ± 0.56 -
30 mg/kg 18 0.52 ± 0.10 5.00 ± 0.83 -
EOS Control 24 2.59 ± 0.25 27.50 ± 2.09 5.60, 2, 0.061 -
15 mg/kg 23 3.05 ± 0.34 24.26 ± 1.70 -
30 mg/kg 18 2.04 ± 0.41 19.56 ± 2.89 -
BAS Control 24 0.92 ± 0.10 10.17 ± 1.14 5.80, 2, 0.054 -
15 mg/kg 23 2.12 ± 0.36 16.09 ± 2.17 -
30 mg/kg 18 1.49 ± 0.28 13.94 ± 1.77 -
PCV Control 23* 29.78 ± 0.83 - 6.29, 2, 0.043 A
15 mg/kg 23 28.91 ± 1.16 - A
30 mg/kg 18 24.33 ± 1.67 - B
TS Control 23* 4.07 ± 0.14 - 2.90, 2, 0.235 -
15 mg/kg 23 4.46 ± 0.13 - -
30 mg/kg 18 4.34 ± 0.21 - - aNumber of cells x 109/L whole blood bResults from post-hoc Dunn’s tests using Šidák-corrected significance values.
* One PCV and TS sample from a control individual was unusable

101
Figure 3.1: Spearman rank correlation between blood selenium (Se) and tissue Se concentrations
(A - kidney, B - liver, and C - muscle) in yellow-bellied sliders (Trachemys scripta scripta)
exposed to three different concentrations of selenium (Se; n = 64).
A B
C

102
Figure 3.2: Oxygen consumption (ml O2/hr) of Trachemys scripta exposed to one of three
selenium treatments (Control = circles, 15 mg/kg = triangles, 30 mg/kg = squares; N = 5, 6, and
8, respectively). Measurements were taken at two separate temperatures (25°C = black and 30°C
= grey).
0
2
4
6
8
10
12
14
16
18
85 90 95 100 105 110
Ox
ygen
consu
med
(m
L/h
r)
Plastron length (mm)
Control, 25C
15 mg/kg, 25C
30 mg/kg, 25C
Control, 30C
15 mg/kg, 30C
30 mg/kg, 30C

103
Figure 3.3: Spearman rank correlation between percent packed cell volume and kidney selenium
(Se, dry weight) concentrations in yellow-bellied sliders (Trachemys scripta scripta) exposed to
three different concentrations of selenium (Se; n = 64).

104
Figure 3.4: Spearman rank correlation between total solids and blood selenium (Se, wet weight)
concentrations in yellow-bellied sliders (Trachemys scripta scripta) exposed to three different
concentrations of selenium (Se; n = 64).

105
Figure 3.5: Mean percent (± 1 SE) bacterial killing ability of yellow-bellied sliders (Trachemys
scripta scripta) exposed to three different treatments of selenium (Se): control (n = 23), 15
mg/kg (n = 23), and 30 mg/kg (n = 18). No statistical differences were observed (X 2 = 0.39, p =
0.819). Escherichia coli was the bacteria species used in these assays.
97.5
98
98.5
99
99.5
Control 15 mg/kg 30 mg/kg
Per
cen
t b
acte
ria
kil
led

106
Figure 3.6: Mean heterophil (# of cells x 109) count (± 1 SE) of yellow-bellied sliders
(Trachemys scripta scripta) exposed to three different treatments of selenium (Se): control (n =
23), 15 mg/kg (n = 24), and 30 mg/kg (n = 18). Kruskal-Wallis analysis found that there were not
significant differences among groups (X 2 = 2.49, p = 0.286).
0
0.5
1
1.5
2
2.5
3
3.5
4
Control 15 mg/kg 30 mg/kg
Mea
n h
eter
op
hil
s

107
CHAPTER 4
CONCLUSIONS
Freshwater turtles are globally declining due to numerous factors, such as invasive
species, destruction of suitable habitat, exploitation, and anthropogenic pollutants (Gibbons
2000; Hopkins 2000a). Turtles’ evolutionary history extends over 300 million years, and they
have evolved a unique suite of life history characteristics (e.g., including delayed maturity,
ontogenetic diet shifts, long life spans, and low adult mortality; Ernst & Lovich 2009). Because
of these life history traits, freshwater turtles are particularly susceptible to anthropogenic
alterations of their environment (Steen et al. 2015). Turtle population dynamics typically rely on
the high survival rates of adult females rather than being dependent upon egg viability and
hatchling survival (Congdon et al. 1994). Studies have suggested that compounding stressors,
such as increased road mortality and overharvesting, can increase the importance of survival
across age classes (Cunningham and Brooks 1996). Contaminants might have a significant role
in the decrease of freshwater turtle populations; further research is needed to establish
contaminants as a detrimental factor (Gibbons et al. 2000; Grillitsch and Schiesari 2010). Little
toxicological information is available for chelonians, and future studies should aim to better
assess the sublethal and lethal impacts of contaminants on multiple age classes.
Future field studies of contaminants in freshwater turtles should seek to evaluate the true
differences of contaminant accumulation among age classes. Immature slider turtles in my field
study tended to accumulate higher amounts of cadmium (Cd) and copper (Cu), though adults
tended to accumulate more arsenic (As). The differences in blood and claw trace element

108
concentrations may be due to the ontogenetic diet shift that slider turtles experience, and
subsequent research should seek to confirm this. The consequences of trace element exposure in
immature slider turtles may not mirror effects in adults. Results from my field study suggest
exposure to coal combustion residues in sliders caused little to no effects on immunological
status — an effect opposite to my once-held hypothesis. Previous studies on reptilian species
have noted that reptiles seem to be less affected by contaminants that typically cause deleterious
effects in other vertebrates (Hopkins et al. 2005; Chin et al. 2013; Perrault et al. 2014). Perhaps
the contaminant loads presented in my field study were not enough to cause observable
aberrations in slider immune function; however, I sampled only non-destructive tissue types, so
it is difficult to estimate the true body burdens of sensitive organs.
Researchers should continue to investigate the effects of controlled contaminant exposure
in freshwater turtles. Although my Se study used ecologically unrealistic techniques (oral
gavage) to ensure accurate application of Se, we were able to expose slider turtles to two planned
concentrations of Se and discern what impacts Se can have on these reptiles. Hematological
parameters were impacted by Se treatment, but metabolic rates and bactericidal capacity were
not. Turtles in the low dose group (15 mg/kg Se) did not seem to be significantly impacted by the
Se exposure. This is fascinating given that these turtles’ Se burdens were much higher than those
known to cause mortality in other species (Ohlendorf and Heinz 2011). Turtles in the high dose
group (30 mg/kg) exhibited symptoms indicative of excess Se exposure (anemia and increased
heterophil:lymphocyte ratios) and Se toxicosis (mortality and histopathological abnormalities;
Haskins unpublished data). My Se study also suggests that non-destructive sampling of blood
can be a reliable indicator of other tissue burdens. Overall, this study lays a foundation for the
investigation of selenium toxicosis in turtles.

109
The findings of my chapters also have implications that are relevant to chelonian
conservation and human health. Forty percent of the world’s turtle species are considered
threatened, and further investigation of the effects of contaminants on chelonian populations is
warranted (Turtle Taxonomy Working Group 2014). Overall, the immunological status of turtles
we captured in D-area did not seem to be negatively impacted by trace element contaminants.
This is interesting given the concentrations of selenium (Se) in some D-area turtles’ blood were
comparable to individuals in my Se study. I did not collect samples for leukocyte differentials in
the field turtles, and perhaps there would have been differences in hematological parameters. The
15 mg/kg Se group in my Se study accumulated liver Se concentrations that resembled previous
liver Se data from slider turtles in D-area. Nagle et al. (2001) found that adult, female slider
turtles had a mean liver Se concentration of 37.18 (± 10.92; n = 4; dry weight), comparable to the
turtles in my 15 mg/kg group with a mean liver Se concentration of 34.89 (± 3.09; n = 23; dry
weight). To relate my similar liver Se values to human consumption (0.79 g/kg of body weight
per day; Zhang 2014), I controlled for moisture content in the liver samples and used a tolerable
daily intake limit equation (TDIL; JEFCA 2006; Moses et al. 2009). The output of this equation
suggests that an average-sized human adult (70 kg) could only eat approximately 3 grams of 15
mg/kg turtle liver before exceeding their daily Se limit. In regard to human health, this could
have exposure implications in the United States where turtle consumption still occurs near
aquatic coal ash basins. It most certainly has implications for harmful levels of consumption in
regions where turtles are more likely to be consumed (e.g., Asia, Central and South America),
and these regions include some countries whose coal use rivals or exceeds the United States’.
Toxicological studies of freshwater turtles have shown that these reptiles do indeed
accumulate high amounts of contaminants known to cause deleterious effects in other taxa

110
(Nagle et al. 2001; Hopkins et al. 2013). My results corroborated past studies’ findings, as slider
claw trace element concentrations from coal ash sites are among the highest reported in
freshwater turtles. In fact, mean claw trace element concentrations were up to approximately 66
times (Arsenic) higher in turtles captured in coal ash sites when compared to turtles from
reference sites. Slider turtles in my Se exposure study also accumulated some of the highest
amounts of Se reported in a reptilian species. To my knowledge the Se experiment is the first
study to report Se-induced mortality and Se toxicosis in a reptilian species. Historically, most
reptilian toxicological studies sought to quantify body burdens through destructive methods and
did not focus on the sublethal impacts of contaminant exposure (Hopkins 2000b). Further
investigation is necessary to not only discern how and where freshwater turtles accumulate
pollutants, but also to properly interpret the sublethal effects of toxicological exposure in these
species.

111
Literature Cited
Chin, S.Y., J.D. Willson, D.A. Cristol, D.V.V. Drewitt, and W.A. Hopkins. 2013. High levels of
maternally transferred mercury do not affect reproductive output or embryonic survival
of northern watersnakes. Environmental Toxicology and Chemistry 32:619-626.
Congdon, J.D., A.E. Dunham, and R.C. Van Loben Sels. 1994. Demographics of common
snapping turtles (Chelydra serpentina): Implications for conservation and management of
long-lived organisms. American Zoology 34:397-408.
Cunningham, D.C., and R.J. Brooks. Bet-hedging theory and eigenelasticity: A comparison of
life histories of loggerhead sea turtles (Caretta caretta) and snapping turtles (Chelydra
serpentina). Copeia 1996:622-627.
Ernst, C.H., and J.E. Lovich. 2009. Turtles of the United States and Canada. John Hopkins
University Press, Baltimore, Maryland, USA.
Gibbons, J.W., D.E. Scott, T.J. Ryan, K.A. Buhlmann, T.D. Tuberville, B.S. Metts, J.L. Greene,
T. Mills, M. Leiden, S. Poppy, and C.T. Winne. 2000. The global decline of reptiles, Déjà
vu amphibians. Bioscience 50:653-666.
Grillitsch, B., and L. Schiesari. 2010. The ecotoxicology of metals in reptiles. In: Sparling, D.W.,
G. Linder, A.C. Bishop, S. Krest, editors. Ecotoxicology of Amphibians and Reptiles.
Pensacola, FL: Society of Environmental Toxicology and Chemistry (SETAC).
Hopkins, B.C., J.D. Willson, and W.A. Hopkins. 2013. Mercury exposure is associated with
negative effects on turtle reproduction. Environmental Science and Technology 47:2416-
2422.

112
Hopkins, W.A. 2000a. Reptile toxicology: Challenges and opportunities on the last frontier
in vertebrate ecotoxicology. Environmental Toxicology and Chemistry. 19:2391-2393.
Hopkins, W.A. 2000b. Use of tissue residues in reptile ecotoxicology: A call for integration and
experimentalism. Pages 35-62 in Gardner, S.C., and E. Oberdörster, editors. Toxicology
of Reptiles. CRC Press, Boca Raton, FL, USA.
Hopkins, W.A., B.P. Staub, and J.A. Baionno. 2005. Transfer of selenium from prey to predators
in a simulated terrestrial food chain. Environmental Pollution 134:447-456.
Joint FAO/WHO Expert Committee on Food Additives (JECFA). 2006. Evaluation of certain
food additives and contaminants. WHO Technical Report Series, No. 940.
Moses, S.K., A.V. Whiting, G.R. Bratton, R.J. Taylor, and T.M. O’Hara. 2009. Inorganic
nutrients and contaminants in subsistence species of Alaska: Linking wildlife and human
health. International Journal of Circumpolar Health. 68:53-74.
Nagle, R.D., C.L. Rowe, and J.D. Congdon. 2001. Accumulation and selective maternal
transfer of contaminants in the turtle Trachemys scripta associated with coal ash
deposition. Archives of Environmental Contamination and Toxicology 40:531-536.
Ohlendorf, H.M., and G.H. Heinz. Selenium in birds. 2011. Pages 668-701 in Beyer, W.N., and
J.P. Meador, editors. Environmental Contaminants in Biota. CRC Press, Boca Raton, FL,
USA.
Perrault, J. 2014. Mercury and selenium ingestion rates of Atlantic leatherback sea turtles
(Dermochelys coriacea): A cause for concern in this species? Marine Environmental
Research 99:160-169.

113
Steen, D.A., J.U. Van Dyke, B.P. Jackson, and W.A. Hopkins. 2015. Reproduction and hatchling
performance in freshwater turtles associated with a remediated coal fly-ash spill.
Environmental Research. 138:38-48.
Turtle Taxonomy Working Group. 2014. Turtles of the world, 7th edition: annotated checklist of
taxonomy, synonymy, distribution with maps, and conservation status. Pages 000.329-
479 in Rhodin, A.G.J., P.C.H. Pritchard, P.P. van Dijk, R.A. Saumure, K.A. Buhlmann,
J.B. Iverson, and R.A. Mittermeier, editors. Conservation Biology of Freshwater Turtles
and Tortoises: A Compilation Project of the IUCN/SSC Tortoise and Freshwater Turtle
Specialist Group. Chelonian Research Monographs.
Zhang, H., X. Feng, H.M. Chan, and T. Larssen. 2014. New insights into traditional health
assessments of mercury exposure: implications of selenium. Environmental Science and
Technology 48:1206-1202.