pneumo virulence factors
TRANSCRIPT

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 1/14
Streptococcus pneumoniae (oftn rfrrd to as thpnuococcus) is th ost coon bactria rspira-tory pathogn in th Unitd Kingdo, and frqunty causs counity-acquird pnuonia, which canhav ortaity rats of or than 20% in patints withconcurrnt pnuococca spticaia1,2. Wordwid,th situation is wors; pnuococca spticaiais a ajor caus of infant ortaity in dvopingcountris, whr it causs approxiaty 25% of aprvntab daths in chidrn undr th ag of 5 andor than 1.2 iion infant daths pr yar3,4. Incountris that hav a high prvanc of HIV-1 infc-tion, thr has bn a significant incras in th ratof pnuococca pnuonia and associatd bactr-aia, and this incras has bn ost arkd inyoung aduts.
Th pnuococcus rsids on th ucosa surfacof th uppr rspiratory tract. It can b radiy cuturdfro th no–orophrynx of huans and, occasion-
ay, othr arg aas that iv in association withhuans. Athough coonization at this sit ss tob asyptoatic, if th organis gains accss to thnoray stri parts of th airway a rapid infaa-tory rspons nsus that rsuts in disas. Th ostcoon anifstations of pnuococca disasincud acut otitis dia, which invovs infctionof th idd-ar spac, and pnuonia, which affctsth trina airways. Bactraic infction, which isassociatd with gratr orbidity and ortaity, typi-cay occurs as a copication of pnuonia (bactr-aic pnuococca pnuonia) or, ss oftn, if thorganis sprads dircty fro its nich in th pharynx
(occut bactraia). Thus, a pnuococca disasbgins with th stabishnt of coonization, that is,th cration of th crrier tte5,6. Non of th coonfors of pnuococca disas, howvr, prootpnuococca transission, which ipis that th
virunc charactristics of th pnuococcus ar prob-aby adaptations that incras its prsistnc within ahost during coonization.
Prson-to-prson sprad is thought to occur throughdirct contact with th scrtions of coonizd individ-uas. Onc acquird, an individua strain can b car-rid for wks to onths bfor its vntua caranc.Howvr, it rains uncar how ffctiv coonizationis as an iunizing vnt. Coonization is ost co-on in ary chidhood; ost infants acquir on orany strains squntiay or siutanousy. Th ratsof carriag vary widy aong th 91 known pnuo-cocca capsuar srotyps, which xprss structuray and antignicay diffrnt capsuar poysaccharids.
Th factors that ar rsponsib for ths diffrncs,and th variations in srotyp distribution btwnrgions and ovr ti, ar not undrstood. Aftr thfirst yars of if, th rats of coonization dcin to ssthan 10% in adut popuations. Th iportanc of car-riag by young chidrn as th ain rsrvoir for thispathogn has bn donstratd by th rcnt wid-sprad us of th pnuococca capsuar poysaccha-rid (CPS) conjugat vaccin (BOX 1). Rducd carriagof th organis in vaccinatd chidrn rsuts in herd
immunity, which can hav an ipact on th frquncy of pnuococca disas in thos ag groups that arnot rciving th vaccin7.
*Department of Infection,Immunity & Inflammation,
University of Leicester,
Leicester LE1 9HN, United
Kingdom.
‡Department of Microbiology,
School of Medicine, University
of Pennsylvania, Pennsylvania
19104-6076, USA.
§School of Molecular and
Biomedical Science,
University of Adelaide, South
Australia 5005, Australia.
Correspondence to A.K.
e-mail: [email protected]
doi:10.1038/nrmicro1871
Naso–oropharynx
The prt of the phrynx tht
re bove nd beow the
pte, repectivey.
Carrier state
The tte of crrying norgnim without
mnifettion of diee.
Herd immunity
Immunity induced by
vccintion tht protect
proportion of unvccinted
individu.
The role of Streptococcus pneumoniaevirulence factors in host respiratorycolonization and disease
Aras Kadioglu*, Jeffrey N. Weiser ‡, James C. Paton§ and Peter W. Andrew*
Abstract | Streptococcus pneumoniae is a Gram-positive bacterial pathogen that colonizes
the mucosal surfaces of the host nasopharynx and upper airway. Through a combination of
virulence-factor activity and an ability to evade the early components of the host immuneresponse, this organism can spread from the upper respiratory tract to the sterile regions of
the lower respiratory tract, which leads to pneumonia. In this Review, we describe how
S. pneumoniae uses its armamentarium of virulence factors to colonize the upper and lower
respiratory tracts of the host and cause disease.
RE V I E W S
288 | APRI 2008 | VOUme 6 www.nau./ws/

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 2/14
Glycocalyx
Extrceur poycchride
mteri tht i excreted by
epithei ce nd form n
outer yer on epithei
urfce.
Mechanisms of colonization
exprinta coonization of aduts has bn usd toinvstigat th host factors that affct suscptibiity to acquisition of S. pneumoniae and its subsquntcaranc8. Ths studis rva that carriag inducsth production of both ucosa and systic iu-nogobuin, which is ainy strain- and typ-spcific.In contrast to th high vs of srotyp-spcificanti-capsuar antibody that ar gnratd foowingadinistration of th conjugat vaccin, it is uncarwhthr th rativy sa aounts that ar inducdby coonization ar sufficint to nhanc caranc. Inthis rgard, th dcin in carriag rats aftr chid-hood is obsrvd widy aong th diffrnt pnu-ococca srotyps, which suggsts that if xposurduring prior coonization vnts ads to iunity it ight not dvop in a srotyp-spcific annr9.
Iunognic ajor c-surfac protins, particuary pnuococca surfac protin A (PspA), aso inducantibody, but this rspons is prdoinaty dirctdat th or variab, xposd rgions of PspA. Thv of pr-xisting cross-ractiv iunogobuin toPspA corrats with suscptibiity to th stabishntof pnuococca carriag8.
Ania ods hav aso bn usd to dfin thhost and bactria factors that contribut to pnuo-cocca coonization (TaBlE 1). For xap, isoats thatwr prviousy xaind in huan xprinta coo-nization studis aso coonizd inbrd adut ic whna siiar inocuu was usd for a siiar duration.
Ths isoats aso icitd a coparab iunrspons to CPS and PspA, thrby donstrating thrvanc of this ania od10. It shoud b notd,howvr, that ic can aso b coonizd by a wid spc-tru of pnuococca srotyps, athough thir sus-cptibiity to disas by diffrnt strains varis graty.Thrfor, for crtain pnuococca srotyps, icar aso usfu ods to study th copications thataris fro bactria coonization, incuding pnuoniaand bactraic infction. Th xprinta findingsfro ous-coonization and airway-infction odswi b suarizd in th foowing sctions.
Interactions with host structures. Within inuts of ntring th nasa cavity, S. pneumoniae cs ncountrucus scrtions (FIG. 1). Th xprssion of a capsurducs ntrapnt in th ucus, thrby aowing
th pnuococcus to accss th pithia surfacs11.Aost a pnuococca CPSs ar ngativy chargd,which coud incras thir rpusion of th siaic acid-rich ucopoysaccharids that ar found in ucus12.Initiay, bactria ar dtctd within th gycocyx ayrthat ovris th rspiratory and ofactory pithiu11 (FIG. 1b). This ight aow bactria accss to rcptorson th apica surfac of th pithia cs that in thnasa spacs. Onc at th pithia surfac, th xprs-sion of a thick capsu ss to b disadvantagous forth pnuococcus, bcaus of its inhibitory ffct onadhrnc. most pnuococca isoats that hav bninvstigatd dispay phe vrition btwn two fors
Box 1 | New approaches to pneumococcal vaccination
Current pneumococcal vaccines are exclusively targeted at the capsular polysaccharide (CPS) ofStreptococcus
pneumoniae, and these vaccines provide strictly serotype-specific protection. Polyvalent purified CPS vaccines were first
licensed in the late 1970s, and the current 23-valent formulation is effective against approximately 90% of disease-
causing serotypes in the United States and Europe. Unfortunately, CPSs are T-cell-independent antigens and are
therefore poorly immunogenic in young children, particularly for the five pneumococcal serotypes that most commonly
cause invasive disease in children145. The poor immunogenicity of CPS antigens has been overcome by conjugation to
protein carriers — this converts them into T-cell-dependent antigens, which are considerably more immunogenic — and apneumococcal CPS–protein conjugate vaccine (PCV) is now licensed in several countries. However, protection is still
serotype specific and, because of the high cost, the number of serotypes that are targeted has been reduced to seven.
Disturbingly, studies of nasopharyngeal colonization with S. pneumoniae during trials of PCV showed that, although the
carriage of vaccine serotypes was reduced, the vacated niche was promptly occupied by non-vaccine pneumococcal
serotypes that are potentially capable of causing disease146. Similar trends were also observed after the general
introduction of the conjugate vaccine in the United States147. Although PCV has dramatically reduced the incidence of
invasive disease that is caused by vaccine serotypes in the United States, increases in the incidence of bacteraemia caused
by non-included serotypes have also been reported148–149. Thus, in the long term, the widespread introduction of PCV
might merely alter the serotype distribution of invasive pneumococcal disease, rather than reducing its overall burden. In
addition, the cost of PCV is high, and therefore it is unlikely that it will be used extensively in developing countries, where
the death rate of children from invasive pneumococcal disease is highest.
Clearly, there is an urgent need to develop alternative pneumococcal vaccines that do not suffer from these
shortcomings. The most promising approach to date is to develop vaccines that are based on pneumococcal proteins that
contribute to virulence and are common to all serotypes. Such vaccines need to be highly immunogenic and elicit
immunological memory even in infants, who respond well to T-cell-dependent protein antigens. Ensuring high expressionlevels of antigenic proteins in recombinant bacteria will enable large-scale vaccine production at low cost, thereby
producing vaccines that are more affordable, particularly for developing countries. Several candidate protein vaccine
antigens have been identified, including non-toxic derivatives of pneumolysin, choline-binding proteins, such as PspA and
CbpA, metal-binding lipoproteins, such as PsaA and PiaA, the poly-histidine triad proteins PhtB and PhtE and the
neuraminidase NanA. Immunization with each of these proteins provided varying degrees of protection against challenge
by one or more S. pneumoniae strains in mouse models of sepsis, pneumonia or carriage146, and there is now clear evidence
that immunization with certain combinations of virulence proteins provides additive, or even synergistic, protection149–151,
the most effective to date being a combination of Ply, PspA and CbpA152,153.
RE V I E W S
NATURe ReVIeWS | microbiology VOUme 6 | APRI 2008 | 289

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 3/14
Phase variation
a moecur mechnim tht
ed to witching of the gene-
expreion tte; for exmpe,
on–off expreion.
Lipoteichoic acid
a teichoic cid pecie tht i
connected to membrne
gycoipid. The
tereochemitry of lTa nd
the bioynthetic origin of the
gycerophophte re
different from thoe of wteichoic cid, which hve
gycero-phophte bckbone.
Human secretory
component
an epithei gycoprotein tht
i required for the ctive
trnport of poymeric
immunogobuin cro
muco urfce.
Glycoconjugate
a crbohydrte tht i
coventy inked to other
chemic pecie.
that can b distinguishd by thir opaqu or transparntcoony orphoogis (discussd bow)13. During thinitia stags of coonization, transparnt variants thatxprss a thinnr capsu and possss othr charactr-istics that proot binding to host tissus prvai ovropaqu variants13.
By anaysing huan pithia cs in cutur, sv-ra rcptor–igand intractions hav bn proposd.Th bactria adhsins that ar invovd incud phos-phorychoin (ChoP), which is a constitunt of bothc-wa-associatd acids and ipoteichoic cid14. ChoP
is an unusua bactria structura coponnt that is ac-surfac fatur of svra othr icroorganiss thatrsid priariy in th uppr rspiratory tract, such asHaemophilus influenzae, Actinobacillus actinomycetem-comitans and consa and pathognic spcis of th gnus Neisseria. As any ChoP-xprssing spcisoccupy a siiar bioogica nich, and hav a coon stof gns (licA–D) for choin uptak and incorporation,bactria ChoP ight b particuary iportant for hostairway coonization15. In H. influenzae, ChoP xprssionnhancs prsistnc in th uppr rspiratory tract16. Inth pnuococcus, ChoP diats bactria adhrncto th rcptor for patt-activating factor (rPAF) and
activats host c signaing through this rcptor. Asth natura igand for rPAF — patt-activating fac-tor (PAF) — aso contains ChoP, th pnuococcusight iic PAF to us its rcptor, which is widy distributd on host tissus such as th pithia surfacof th huan nasopharynx. Anothr adhsin is choin-binding protin A (CbpA; aso rfrrd to as PspC orSpsA), which is non-covanty anchord to ChoP. CbpAbinds to humn ecretory component, which is found onth poyric iunogobuin rcptor, and scrtory fors of iunogobuin17,18. In addition, so isoats
xprss a pius-ik structur that adhrs to an unknownpithia-c rcptor19,20.
Othr studis hav providd vidnc for adhsivintractions with host c gycoconjugte. Pnuococcaadhrnc to pithia cs fro th huan pharynx isinhibitd by N -actygucosain-β-(1,3)-gaactos21.Th pnuococcus is aso on of any pathogns thathav bn rportd to bind to N -actygucosain-β-(1,4)-gaactos, which is a constitunt of sohuan gycosphingoipids22. In addition, S. pneumoniae producs thr surfac-associatd xogycosidass:a nurainidas, NanA, a β-gaactosidas, BgaA, and aβ-N -actygucosainidas,StrH23. Ths nzys act
Table 1 | Pneumococcal virulence factors and their main role in colonization and disease
Pnua unas and dsas
man n nzan
Upper-airway colonization
Capsule Prevents entrapment in the nasal mucus, thereby allowing access to epithelial surfaces.Also inhibits effective opsonophagocytosis.
ChoP Binds to rPAF on the epithelial surface of the human nasopharynx.CbpA (also known as PspC) Binds to human secretory component on a polymeric Ig receptor during the first stage of
translocation across the epithelium.
NanA, BgaA and StrH Act sequentially to cleave terminal sugars from human glycoconjugates, which mightreveal receptors for adherence.
Hyl Breaks down hyaluronan-containing extracellular matrix components.
PavA Binds to fibronectin.
Eno Binds to plasminogen.
Competition in upper airway
Bacteriocin (pneumocin) Small antimicrobial peptide that targets members of the same species.
Respiratory-tract infection and pneumonia
Ply Cytolytic toxin that also activates complement. An important determinant of virulence in
in vivo models of disease. Wide range of effects on host immune components at sub-lyticconcentrations.
PspA Prevents binding of C3 onto pneumococcal surface. Also binds lactoferrin.
LytA Digests the cell wall, which results in the release of Ply.
PsaA Component of the ABC transport system, which is involved in resistance to oxidative stress.
PiaA and PiuA Component of the ABC transport system.
NanA and NanB Aid colonization by revealing receptors for adherence, modifying the surfaces ofcompeting bacteria that are within the same niche and/or modifying the function of hostclearance glycoproteins.
IgA Cleaves human IgA1.
This list of virulence factors is not exhaustive and only selected examples are shown. For detailed descriptions of virulence factorssee the main text. BgaA, β-galactosidase; CbpA, choline-binding protein A; ChoP, phosphorylcholine; Eno, enolase; Hyl,hyaluronate lyase; IgA, IgA1 protease; IgA1, immunoglobulin A1; LytA, autolysin A; Nan, neuraminidase; PavA, pneumococcal
adhesion and virulence A; PiaA, pneumococcal iron acquisition A; PiuA, pneumococcal i ron uptake A; Ply, pneumolysin; PsaA,pneumococcal surface antigen A; PspA, pneumococcal surface protein A; StrH, β-N-acetylglucosaminidase.
RE V I E W S
290 | APRI 2008 | VOUme 6 www.nau./ws/
8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 4/14
a 30 minutes b 1 day
c 3 days d 14 days
Basement membrane
a tiue tructure tht conit
of network of cogen fibre
tht ttch the overying
epithei yer to the
connective tiue underneth.
squntiay to rov th trina sugars that ar foundon any huan gycoconjugats and, thrfor, ightunask rcptors for adhrnc, thrby affcting thfunction of gycosyatd host caranc ocus and/orproviding a nutrint sourc.
Binding to xtracuar atrix coponnts coudaso proot bactria adhrnc onc th organ-is has gaind accss to bement membrne. Thxprssion of a surfac-attachd hyauronidas coudfaciitat bactria sprad through a atrix of th
hyauronan-containing poysaccharid coponntof connctiv tissus24. Pnuococca adhsion and
virunc A (PavA) and noas ar two pnuococ-ca surfac adhsins that bind th xtracuar-atrixcoponnts fibronctin and pasinogn, rspc-tivy 25,26. PavA is ocaizd to th pnuococcaoutr c surfac and has bn shown to b ipor-tant in virunc, as th absnc of PavA significanty rducs systic bactria oads and incrass hostsurviva25. Pnuococca noas utants aso xhibitattnuation in a od of rspiratory infction, whichindicats that pasinogn binding has a ro inrspiratorydisas26.
Th pnuococca factors that nab host pithiaand tissu barrirs to b brachd during th progrs-sion fro coonization to invasiv infction ar poory undrstood. In both ous ods and in huans,opaqu variants that xprss incrasd aounts of CPSand ar or rsistant to opsonophagocytic kiing arsctd for during th transition fro th ucosasurfac to th boodstra27,28.
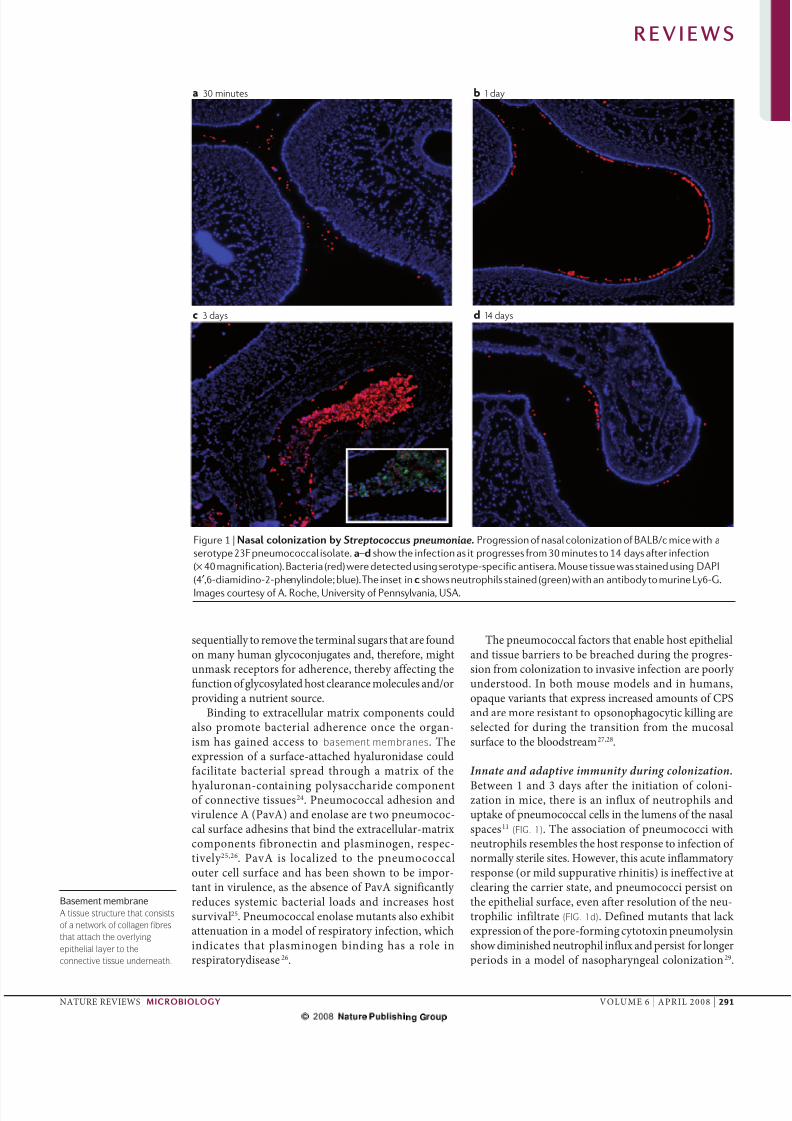
Innate and adaptive immunity during colonization.
Btwn 1 and 3 days aftr th initiation of cooni-zation in ic, thr is an infux of nutrophis anduptak of pnuococca cs in th uns of th nasaspacs11 (FIG. 1). Th association of pnuococci withnutrophis rsbs th host rspons to infction of noray stri sits. Howvr, this acut infaatory rspons (or id suppurativ rhinitis) is inffctiv atcaring th carrir stat, and pnuococci prsist onth pithia surfac, vn aftr rsoution of th nu-trophiic infitrat (FIG. 1d). Dfind utants that ack xprssion of th por-foring cytotoxin pnuoysinshow diinishd nutrophi infux and prsist for ongrpriods in a od of nasopharynga coonization29.
Figure 1 | Nasa nzan Streptococcus pneumoniae. Progression of nasal colonization of BALB/c mice with a
serotype 23F pneumococcal isolate. a–d show the infection as it progresses from 30 minutes to 14 days after infection
(× 40 magnification). Bacteria (red) were detected using serotype-specific antisera. Mouse tissue was stained using DAPI
(4′,6-diamidino-2-phenylindole; blue). The inset in shows neutrophils stained (green) with an antibody to murine Ly6-G.
Images courtesy of A. Roche, University of Pennsylvania, USA.
RE V I E W S
NATURe ReVIeWS | microbiology VOUme 6 | APRI 2008 | 291

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 5/14
Complement
a prt of the innte immune
ytem tht comprie erum
protein which cn protectgint infection.
Fab fragment
The region of n ntibody tht
bind to n ntigen.
Bacteriocin
a bcteriy produced, m,
het-tbe peptide tht i
ctive gint other bcteri
nd to which the producer h
pecific immunity
mechnim. Bcteriocin cn
hve nrrow or brod trget
pectrum.
epithia cs in cutur rspond to th osoticstrss of pnuoysin-diatd por foration by activating th p38 itogn-activatd protin kinas,which rsuts in an incras in chokins that attractnutrophis30. Ths ffcts of pnuoysin on th pi-thia surfac suggst that th pnuococcus ighthav vovd to proot th infaatory rspons,prhaps bcaus th rsuting scrtions incras thikihood of transission, vn though this incrasdhost rspons coud accrat th vntua carancof th pathogn.
Host-diatd kiing of S. pneumoniae is gnr-ay thought to rquir opsonization by a srotyp-spcific antibody togthr with compement, foowdby phagocytosis. Intrstingy, th cours of a coo-nization vnt is unaffctd in ic that fai to gn-rat a spcific antibody, and nithr th dptionof nutrophis nor copnt ipacts on initiacoonization by th organis10. By contrast, icthat ack To-ik rcptor 2 (TR2), an initiator of th infaatory rsponss that foow th rcogni-
tion of ipotichoic acid and/or ipoprotins, xhibitdayd pnuococca caranc29. In addition, icthat do not xprss ajor histocopatibiity copxcass II aso show proongd carriag, which indicats aniportant ro for CD4+ T cs rathr than huoraiunity 31 (BOX 2). Th CD4+ T-c-dpndnt ffc-tor functions that ad to oss of stabishd cooniza-tion hav yt to b idntifid. Thr is aso vidncthat pnuoysin stiuats infaatory rsponssthrough TR4 (REF. 32).
Anothr factor that iits th ffctivnss of thhost huora rspons on ucosa surfacs is bac-tria xprssion of a scrtd zinc taoprotasthat spcificay targts huan iunogobuin A1(IgA1), which constituts or than 90% of th IgAin th huan airway 33. Th cavag of bound IgA1producs bactria surfac antigns that ar boundto Fb frgment, which prvnts infaation frobing initiatd through host rcognition of th Fc rgion of th antibody. Bcaus of th activity of thIgA1 protas, antibody-diatd caranc ightoccur ony aftr sufficint aounts of othr casssand subcasss of spcific antibody hav bn gnr-atd. Indd, th dcras in pnuococca cooniza-tion foowing th us of th CPS conjugat vaccincoud b attributab to its induction of high vs of IgG, which is not targtd by th IgA1 protas. Th
abiity of S. pneumoniae to inactivat IgA1 in iu-nocoptnt hosts coud account for th gnray unatrd incidnc of infction that is obsrvd inost IgA-dficint individuas.
C-ractiv protin (CRP) is anothr iportantcoponnt of th innat iun rspons againstS. pneumoniae. CRP binds spcificay to ChoP and, aftrbinding, CRP intracts with copnt coponntC1q to activat th cassica pathway of copnt.Transgnic ic that xprss th huan crp gn aror rsistant to pnuococca infction34. morovr,at th concntrations found in th airway, CRP can bock pnuococca adhrnc through rPAF35.
Impact of competition. It is stiatd that or than 700diffrnt icrobia spcis can rsid within th huanpharynx36. Succssfu coonization probaby rquirschaniss that countract th prsnc of icrobiacoptitors. Thr is vidnc that in a ous od of pnuococca infction, co-coonization with anothrbactria spcis that rsids in th sa nich can rsutin th caranc of S. pneumoniae through th induc-tion of copnt-diatd nutrophi kiing37. Thisdonstrats that on icrobia spcis can harnss thinnat iun rspons of its host to prvai ovr a co-ptitor. A furthr ipication is that th pnuococcusis rsistant to th innat iun rsponss it stiuats,but is not ncssariy rsistant to th rsponss that arinducd by its coptitors.
Pnuococca strains aso copt with ach othr.Th incras in th prvanc of prviousy uncoonsrotyps in popuations in which th pnuococca CPSconjugat vaccin is xtnsivy usd (a phnonon thatis rfrrd to as srotyp rpacnt) suggsts that non-
vaccin pnuococca typs ar bing out-coptd by
th srotyps that ar prsnt in th vaccin. On cha-nis that coud undri this intra-spcis coptition isth strain-spcific activity of pnuococca bcteriocin,which ar known as pnuocins38. Ths sa antii-crobia pptids, which ar xprssd by th blp ocus andar undr th contro of a quoru-snsing phroon(BpC), targt brs of th sa spcis that do notproduc th cognat iunity protin. In a ous odof coonization, diffrncs of ony a fw aino acidsin th bipartit pnuocin BpmN ar sufficint to dictatth outco of coptition btwn two isoats.
Anothr consqunc of th highy popuatd icro-bia nvironnt in th huan pharynx is th avaiabiity of xognous DNA fro cosy ratd ora strpto-cocca spcis and co-coonizing pnuococci. Thsnucic acids can b takn up by S. pneumoniae bcausof its natura coptnc and, subsqunty, b usd toincras its ovra fitnss. Th acquisition of gns thatncod atrd pniciin-binding protins, for xap,has faciitatd rsistanc to β-acta antibiotics, whichis now a coon prob in th tratnt of pnuo-cocca infctions39. Th abiity of th pnuococcus totak up DNA fragnts and incorporat hooogoussquncs into its gno is ony obsrvd during aro-bic growth. Indd, this activity coud aso b a ansof copnsating for th high utation rats that rsutfro th oxidativ ifsty of this organis.
Protein virulence factors
Ovr th past 20 yars, th iportanc of protins forS. pneumoniae virunc has bco car. Rsarchin this ara has bn stiuatd by th raization thatpnuococca protins rprsnt a proising avnufor th dvopnt of vaccins that ar coon to apnuococca srotyps. Consqunty, th nubr of potntia virunc factors that hav bn charactrizdhas incrasd graty in rcnt yars, and ar too nur-ous to rviw in dtai hr. W wi thrfor discuss ony a sction of th pnuococca virunc factors that arproising candidats for us as vaccin antigns (FIG. 2).
RE V I E W S
292 | APRI 2008 | VOUme 6 www.nau./ws/

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 6/14
Pneumolysin. many studis hav shown that pnuo-ysin is a potnt, wid-ranging virunc factor. It is foundin virtuay a pnuococca isoats, and its aino acidsqunc is w consrvd, athough a sa nubr of
variants hav bn dscribd40,41. Pnuoysin is a -br of th faiy of chostro-dpndnt cytoysins thatar synthsizd by Gra-positiv bactria.
Pnuoysin is producd as a 52 kDa soub protinthat oigorizs in th bran of targt cs tofor a arg ring-shapd transbran por. Th poris 260 Å in diatr and is coposd of approxiaty 40 onor subunits. During its convrsion fro asoub onor to a bran-insrtd oigor,pnuoysin undrgos a sris of spctacuar structura
changs42. Th oigors ar thought to b rsponsib forth cytoytic activity of th toxin and th pthora of c-oduatory activitis that ar vidnt at sub-ytic con-cntrations. Ths activitis incud: inhibition of ciiary bating on rspiratory pithiu and brain pndya;inhibition of th phagocyt rspiratory burst; and induc-tion of cytokin synthsis and CD4+ T-c activation andchotaxis43,44. Sit-dirctd utagnsis has shown thatpnuoysin activats th cassica copnt pathway indpndnty of its c-oduatory activity 45.
Both th c-oduatory and copnt-activationactivitis of pnuoysin hav bn shown to contrib-ut to pnuococca virunc in urin ods of
pnuonia46–48; howvr, it is notworthy that strainsof a srotyp 1 con that produc a non-cytoyticpnuoysin hav bn isoatd fro cass of invasivpnuococca disas41. Intrstingy, S. pneumoniae that xprssd a utatd pnuoysin protin whichackd haoytic and copnt-activating activity was shown to b or virunt than a pnuococcusin which th pnuoysin gn was dtd47. Thisindicatd that pnuoysin has an additiona, unidn-tifid function. It was suggstd that this activity was anintraction with TR4, bcaus a pnuococca utantthat producd pnuoysin without haoytic activity was rportd to activat TR4-dpndnt rsponss32.In addition, Baba and coagus49 rportd that a
non-cytoytic pnuoysin stiuatd th productionof intrfron-γ . This coud aso b an unidntifidfunction, athough S. pneumoniae strains that xprssthis for of pnuoysin wr no or virunt thanth pnuococcus in which th pnuoysin gnhad bn knockd out (A.K. and P.W.A., unpubishdobsrvations).
Thr is substantia vidnc that pnuoysin is cru-cia for pnuococca virunc in pnuonia46–48,50,51,athough its rativ iportanc can vary fro strainto strain. For xap, Axandr et al.52 found thatic that had bn iunizd with pnuoysinwr protctd significanty fro nin pnuococca
Box 2 | CD4+ T cells and pneumococcal disease
Research into the host immune response to pneumococcal disease has focused primarily on the role of humoral
components of innate and acquired immunity and the cellular aspects of innate immunity. However, an appreciation of
the involvement of CD4+ T cells in the clinical setting, in which HIV-1-infected patients who have lower CD4+ T-cell
counts are significantly more likely to be persistent pneumococcal carriers than non-HIV-1-infected patients and
patients with AIDS frequently develop severe pneumococcal infections, has led to more detailed investigations of the
role of these cells154,155.
Recently, the role of CD4+
T cells in pneumococcal disease has been revisited using mouse models of pneumococcaldisease. To summarize, this work showed that in lungs of mice that are infected intranasally with Streptococcus
pneumoniae, CD4+ T cells contribute to early host resistance to infection, as shown by an early rapid increase in T-cell
infiltration to areas that are subject to increased pneumococcal invasion. In addition, an increased host susceptibility to
infection that was due to significantly increased bacterial loads both in lungs and blood was observed in CD4+-deficient
mice44,57. Interestingly, infections that used pneumolysin-deficient isogenic pneumococcal mutants resulted in a total
lack of T-cell infiltration into infected lungs, which suggests that pneumolysin was responsible for the pattern of T-cell
infiltration57. Studies that used in vitro chemotaxis assays with purified human CD4+ T cells have shown that the CD4+
T-cell migratory process is indeed dependent on the presence of pneumolysin, as pneumolysin-deficient pneumococci
failed to stimulate CD4+ T-cell chemotaxis. Sub-lytic concentrations of purified pneumolysin or pneumolysin-positive
S. pneumoniae were able to successfully stimulate CD4+ T-cell chemotaxis44.
Protection studies in mice have recently confirmed the role of CD4+ T cells as part of an antibody-independent immune
response to pneumococcal infection31. It was demonstrated that intranasal immunization with live pneumococci (or a
killed, non-encapsulated whole-cell vaccine) against subsequent pneumococcal intranasal challenge, protected
antibody-deficient mice, but not CD4+ T-cell-deficient mice. Other researchers have also shown that CD4+ T cells are
required for efficient clearance of nasopharyngeal pneumococcal colonization in naive mice, as CD4+
T-cell-deficientmice failed to clear pathogen colonization29. The inability of CD4+ T-cell-deficient mice to clear nasopharyngeal
colonization was explained by the lack of induction of a T helper 1 (TH1) response, which was previously shown to have a
protective role in the response to pneumococcal disease in humans156. In addition, it has been demonstrated that
pneumolysin promotes CD4+ T-cell-dependent clearance of colonization29.
Recent studies have also found that CD4+ T-cell proliferative responses to pneumolysin are significantly higher in
children who do not have detectable pneumococci in their nasopharynx, which suggests that natural CD4+ T-cell
immunity to pneumococcal protein antigens could modulate nasopharyngeal carriage157. Indeed, previous work had
shown that mucosal antibody production against pneumococcal proteins, such as pneumolysin and CbpA, was T-cell
dependent158. Although it is unclear whether this T-cell immunity cleared existing bacterial carriage or prevented new
colonization, it is worth considering that future vaccines should include conserved pneumococcal protein antigens that
are capable of inducing CD4+ T-cell immunity.
RE V I E W S
NATURe ReVIeWS | microbiology VOUme 6 | APRI 2008 | 293

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 7/14
|
Pneumolysin
LytA
PspA
PspC
PiuAPiaA
Metal-bindingproteins
ATP-bindingcassettetransporter
Choline-bindingproteins
LPXTG-anchoredneuraminindase proteins
Hyl
PsaA
PavA
Eno Sortases
Cell wall
Capsule
Cell membrane
Opsonic antibody
a bcteri-binding (oponizing)
ntibody, uch IgG nd Iga,
tht interct with Fc receptoron phgocytic ce, which
ed to n increed uptke of
bcteri.
Teichoic acid
a ce-enveope gycopoymer
tht i compoed of mny
identic ugr-phophte
repeting unit, which re
uuy modified with d-nine
nd ddition ugr.
Apolactoferrin
The iron-depeted form of the
gycoprotein ctoferrin.
strains, but no protction was affordd against atnth strain. It is iky that th strain-spcific naturof this pnuococca virunc factor wi aso hodtru for any othr, and prhaps a, pnuococca
virunc factors.In a ous od of acut pnuonia, pnu-
oysin was shown to b ssntia for th surviva of S. pneumoniae in both th uppr and owr rspiratory tracts53,54. Thr is aso good vidnc that pnuoysinis rquird for bactria sprad fro th ungs to thboodstra53–56. In bactraic infction, if pnu-oysin is xprssd, high nubrs of pnuococciar found in th boodstra, and th host succubsto infction54,56; howvr, in th absnc of pnuo-ysin, high pnuococca nubrs ar toratd in th
bood without ovrt disas syptos57, and chronicbactraia can dvop58.
For ods of ningitis, howvr, thr is ordbat on th iportanc of pnuoysin. In contrastto a rport that suggstd ony a iitd ro for pnu-oysin in S. pneumoniae ningitis59, subsquntstudis found that pnuoysin was a ky dtri-nant of pnuococca virunc in this disas 60,61.Furthror, pnuoysin has aso bn shown to brquird for pnuococca-inducd dafnss in n-ingitis62 and for pnuococca-inducd daag to thbrain pndya63,64. Additiona vidnc that supportsth ro of pnuoysin in ningitis was discussd in
a rcnt rviw of th ro and action of pnuoysin inpnuococca disass65.
S. pneumoniae is aso a natura pathogn of horss66.Intrstingy, a quin isoats ar srotyp 3 (REF. 66),and hav a ajor dtion within th gn that ncodspnuoysin, as w as th gn that ncods thautoysin ytA (discussd bow)67.
Pneumococcal cell-surface proteins
C-surfac protins hav attractd considrab attn-tion bcaus of thir potntia as vaccin antigns thatcan stiuat th production of oponic ntibodie. Thrajor groups of pnuococca c-surfac protins havbn idntifid: choin-binding protins, ipoprotinsand protins that ar covanty inkd to th bactriac wa by a carboxy (C)-trina sortas (PXTG; inwhich X dnots any aino acid) otif.
Choline-binding proteins. As ntiond prviousy,pnuococci xprss ChoP as a coponnt of thirc-wa teichoic cid and bran-bound ipotichoic
acids. ChoP anchors a group of protins, th choin-binding protins, to th c wa. most of ths protinshav rpat squncs of approxiaty 20 aino acidsthat diat attachnt of th protins to th c surfacthrough phosphorychoin. Th aino (N)-trinasquncs vary widy, and ar th sits of th spcificactivitis of th diffrnt protins68. S. pneumoniae ncods 10–15 choin-binding protins69, incudingPspA, PspC and ytA.
PspA. PspA has thr structura doains, and itsN-trina rgion is coposd of rpatd α-hicsthat protrud fro th bactria c surfac70. Its highy ctrongativ proprtis ar thought to inhibit co-pnt binding70. Btwn th N trinus and thC-trina choin-binding rgion is a proin-richrgion of 60–80 aino acids that probaby confr fx-ibiity 70. PspA is a highy variab ocu that, basdon th squncs of th N trini, can b groupd intothr faiis that, in turn, can b subdividd into sixdiffrnt cads71.
PspA intrfrs with th fixation of copntcoponnt C3 on th pnuococca c surfac,and thus inhibits copnt-diatd opsoniza-tion. PspA is aso a actofrrin-binding protin and,through this activity, is thought to protct th bact-riu fro th bactricida activity of poctoferrin.
PspA-knockout utants ar or snsitiv to kiingby apoactofrrin, and anti-PspA antibodis nhancth bactricida activity of apoactofrrin72,73. Thrar conficting data, howvr, on th in vivo ffct of dting th pspA gn. Athough studis of srotyp 3(REFs 74,75) and 4 (REF. 20) pnuococci showdthat PspA is rquird for in vivo growth, siiarxprints that usd a oss-of-function utant of a srotyp 2 strain faid to show an ffct54,76,77. Aninvstigation by Abyta and coagus77 suggstdthat th contribution of PspA to virunc ightdpnd on th proprtis of th pnuococcapoysaccharid capsu.
Figure 2 | Pnua un as.Streptococcus pneumoniae virulence is multi-faceted.
Important pneumococcal virulence factors include: the
capsule; the cell wall; choline-binding proteins;
pneumococcal surface proteins A and C (PspA and PspC);
the LPXTG-anchored neuraminidase proteins; hyaluronate
lyase (Hyl); pneumococcal adhesion and virulence A (PavA);
enolase (Eno); pneumolysin; autolysin A (LytA); and the
metal-binding proteins pneumococcal surface antigen A
(PsaA), pneumococcal iron acquisition A (PiaA) and
pneumococcal iron uptake A (PiuA).
RE V I E W S
294 | APRI 2008 | VOUme 6 www.nau./ws/

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 8/14
Factor H
a component of the terntive
compement pthwy tht i
invoved in the regution of
compement ctivtion.
PspC. PspC is a utifunctiona c-surfac protin thatis known by svra othr nas that rfct its diffrntactivitis. For xap, it is aso known as choin-bindingprotin A (CbpA), bcaus it was th prdoinant ntity to b purifid by ChoP-affinity chroatography 78. A pspC-knockout utant binds ss w to pithia csand siaic acid in vitro, and shows rducd nasopharyngacoonization copard with th wid typ78.
PspC aso binds th poyric iunogobuinrcptor that noray transports scrtory IgA; hncit is aso cad SpsA (scrtory pnuococca surfacprotin A, as ntiond abov)79,80. This activity coudb th first stag of transocation across th rspiratory pithiu, which is consistnt with th rducd viru-nc of a pspC utant in a ous od of pnuonia48.Ianni et al.81 rportd that PspC utants of srotyp 2and 3 S. pneumoniae wr ss virunt in a spsisod. Additiona studis hav supportd this finding82.Howvr, Orihua et al.54 faid to find any ffct onspsis or pnuonia using a pspA utant of th sasrotyp 2 strain, athough an ipairnt in pathogn
surviva in th nasopharynx was obsrvd.An additiona proprty of PspC is its abiity to bind to
fctor H83,84, which has bn shown to prvnt foration of C3b though th atrnativ copnt pathway, and thusprvnt pnuococca opsonization82. PspC protins arhighy poyorphic54, and can b dividd into two struc-tura groups. In addition to th PspC ocu that bindsto phosphorychoin, th variant that was first dscribdas Hic83 is anchord to th bactria c wa using thcassica PXTG otif. Howvr, both th Hic variantand th cassic choin-binding for of th protin arab to squstr factor H. PspC aso binds copntcoponnt C3 (REF. 85).
LytA. ytA is an aidas that cavs th N -acty-uraoy-l-aanin bond of pnuococca pptidog-ycan86. Th autoytic action of this nzy ads to thc ysis that is typica of pnuococca cs growing inbatch cutur, but this nzy aso participats in c-wa growth and turnovr. ytA-ngativ S. pneumoniae utants wr shown to hav rducd virunc in urinods of pnuonia and bactraia51,54,76,87. Th con-tribution of ytA to virunc is thought to b diatd,in part, by its function in th ras of pnuoysin andinfaatory pptidogycan and tichoic acids froysd bactria cs.
Divalent metal-ion-binding lipoproteinsBtwn 42 and 45 pnuococca c-surfac ipopro-tins hav bn dscribd to dat69. Ths incud thpptid isorass PpA and SrA, which hav bnshown to b invovd in S. pneumoniae virunc88,89. Thta-binding ipoprotins pnuococca surfac anti-gn A (PsaA), pnuococca iron acquisition A (PiaA)and pnuococca iron uptak A (PiuA) ar additionac-surfac ipoprotins that hav bn shown to bssntia for pnuococca virunc.
PsaA. Dting psaA aboishs virunc in urinods of pnuonia, bactraia and coonization90–92.
Originay, PsaA was proposd to b a pnuococcaadhsin bcaus of its squnc siiaritis to puta-tiv adhsins fro othr strptococci93. Subsquntinvstigations sd to support this assrtion. psaA pnuococca utants wr dficint in binding toaaian cs in vitro90,94 and, or rcnty, it wasrportd that anti-PsaA antibodis inhibitd pnu-ococca adhrnc95. Howvr, PsaA is actuay thdivant ta-ion-binding ipoprotin coponnt of an ATP-binding casstt (ABC) transport syst thathas spcificity for angans96,97. Structura anaysis of PsaA shows a potntia divant ta-ion-binding sitthat can accoodat zinc or angans ions98, and psaA utants rquir angans for nora growth96.Thrfor, th obsrvd ffcts on bactria adhrncar prsuaby a rsut of th piotropic ipact of adficincy in angans transport on th xprssionof othr pnuococca gns, incuding adhsins. Thisproposa is consistnt with structura studis which indi-cat that PsaA is uniky to protrud byond th pnu-ococca c wa68,98. Furthror, psaA is transcribd
as part of th psaBCA opron, and if othr gns of thopron ( psaB or psaC) ar utatd, coparab dfi-cincis in bactria adhrnc ar obsrvd99. A oriportant outco of th psaA utant studis, how-vr, is that angans uptak ss to b ssntia forpnuococca rsistanc to oxidativ strss, which canrsut fro th production of hydrogn proxid duringpnuococca tabois, as w as th gnration of ractiv oxygn spcis during th host innat iunrspons. It is this proprty that probaby accounts forth avirunc of psaA utants in ous ods of coonization and invasiv disas97,100.
PiaA and PiuA. Thr ABC transportr oprons thatdiat iron uptak, pia, piu and pit , hav bn dscribdin S. pneumoniae101,102. each opron copriss gns thatncod a ta-binding protin, a bran prasand an ATPas, but th pia and piu systs s to bparticuary iportant102. PiaA and PiuA ar th ipopro-tin ta-binding coponnts of ths systs, andiunization with ths protins is protctiv103. Thrss to b rdundancy in th iron-uptak systs,howvr, as ony a piu– pia doub-utant strain had sig-nificanty rducd growth in an iron-dficint diuand, athough a sing utation in pia or piu dcrasd
virunc in ods of pnuonia and bactraia,th doub utant was attnuatd to a significanty
gratr xtnt102.
LPXTG-anchored proteins
Th bst-charactrizd chanis of anchoring pro-tins to th pptidogycan of Gra-positiv bactria isthrough th sortas transpptidas that rcognizs thaino-acid squnc PXTG in surfac-ocatd pro-tins. So pnuococca strains ncod a sing sortasgn, whras in othrs utip sortas-ik gns arprdictd69. Whr utip sortas gns ar prsnt,it is proposd that StrA is rsponsib for anchoringost XPTG-containing protins and th othr sortasprotins diat th anchoring of a subst of surfac
RE V I E W S
NATURe ReVIeWS | microbiology VOUme 6 | APRI 2008 | 295

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 9/14
C-type lectin
a crbohydrte-binding
protein tht i found in wide
rnge of nim. C-type ectin
hre highy conerved
ccium-dependent
crbohydrte-recognition
domin tht i ued to
ditinguih them from other
nim ectin.
protins, prhaps in rspons to spcific nvironntacus20. StrA has bn rportd to hav a ro in pnuo-cocca coonization, pnuonia and bactraia, andin adhsion to nasopharynga cs104–106. It is stiatdthat up to 20 S. pneumoniae protins ar anchord by anPXTG otif 69, incuding th nurainidass.
Neuraminidases. Nurainidass, aso known as siai-dass, cav trina siaic acid rsidus fro gyco-protins, gycoipids and c-surfac oigosaccharids.A rcnt study showd that nurainidass can rovsiaic acid fro soub protins, such as actofrrin, IgA2and scrtory coponnt107. S. pneumoniae ncods atast thr nurainidas gns: nanA, nanB and nanC.A strains ncod nanA, and ost aso ncod nanB;howvr, ony approxiaty 50% of strains ncodnanC108. Nurainidass ar scrtd fro th c,but ony NanA contains th PXTG squnc, whichsuggsts that ths nzys hav diffrnt functionsin vivo. This hypothsis is supportd by th obsrva-tion that NanB has a uch owr acidic pH optiu
than NanA109. exprints that usd oss-of-functionutants in a ous od of acut pnuonia havshown that NanA and NanB ar iportant for pnu-ococca surviva in th rspiratory tract and bood-stra110. Ths xprints aso indicatd that thtwo nzys hav diffring ros; th NanA utant wasrapidy card fro th rspiratory tract, but th NanButant prsistd, athough th nubr of organissdid not incras110. Howvr, othr work that usd aous nasopharynga-coonization od showd norduction in th abiity of a NanA utant to coonizth nasopharynx ovr a ongr priod23. Athough thris no dirct xprinta vidnc of a bioogica rofor NanC, an anaysis of th distribution of nanC aongS. pneumoniae isoats suggstd a tissu-spcific ro —nanC was or coon in isoats fro crbrospinafuid than in carriag isoats108.
Tissue specificity of virulence
It is car that th contribution of ach virunc fac-tor varis, individuay and coctivy, according toth in vivo ocation of th bactriu. Using rvrs-transcription PCR, a rcnt study obsrvd two pattrnsof in vivo gn xprssion by S. pneumoniae: on pattrnwas charactristic of pnuococci in th boodstraand th othr of pnuococci in th ungs and brain111.In th boodstra, for xap, th xprssion of ply
(which ncods pnuoysin) and pspA was incrasd,whras in ung and brain tissu th xprssion of nanA and nanB, and th coptnc gns comA, comE andcomX , was highr. Th invovnt of th coptncsyst in virunc was confird by adinistr-ing coptnc-stiuating pptid (CSP) and usinga coptnc-ngativ S. pneumoniae utant. CSPoduatd virunc, but in a tissu-spcific annr.Thus, th adinistration of CSP nhancd virunc inpnuonia ods, and a comD utant had rducd
virunc. Ths obsrvations contrast dircty withthos of bactraia, in which th adinistration of CSP dcrass virunc. Finay, CSP was aso shown
to induc pnuococca biofi foration in vitro, and th pattrn of pnuococca-gn xprssion inbiofis parad that obsrvd in th ungs, whrasth gn-xprssion profi in th boodstra chodthat obsrvd in panktonic cutur in vitro111.
The pneumococcal capsule
Structure, function and role in virulence. Th poysaccha-rid capsu fors th outrost ayr of S. pneumoniae cs, and is approxiaty 200–400 n thick 112. With thxcption of srotyp 3, and possiby othrs, th capsuis covanty attachd to th outr surfac of th c-wapptidogycan113. A tota of 90 structuray and sroogi-cay distinct CPS typs hav bn rcognizd to dat114.Capsu production is indispnsab for pnuococca
virunc and is strongy anti-phagocytic in non-iunhosts115 Athough non-ncapsuatd strains hav bnassociatd with suprficia infctions, such as conjunctivi-tis116,117, cinica isoats fro othr stri sits ar ncap-suatd, and spontanous non-ncapsuatd drivativs of ths strains ar argy avirunt. most CPS srotyps ar
highy chargd at physioogica pH, and this can dircty intrfr with intractions with phagocyts118. Th capsuaso fors an inrt shid that ss to prvnt ithr thFc rgion of IgG or copnt coponnt iC3b, whichis bound to dpr c-surfac structurs (for xap,tichoic acids and c-surfac protins), fro intract-ing with thir rvant rcptors on phagocytic cs119,120.mor rcnt data suggst that th capsu can aso rducth tota aount of copnt that is dpositd on thbactria surfac121. It aso rducs th trapping of pnuococca cs in nutrophi xtracuar traps122.
Th virunc of S. pneumoniae is ratd to ththicknss of th capsu in a particuar strain andsrotyp123. Howvr, pnuococci fro th diffr-nt CPS srotyps diffr arkdy in thir capacity tocaus disas115. This prsuaby rfcts thir rativcapacity to rsist phagocytosis, as w as diffrncs inthir abiity to icit a huora iun rspons. Forxap, Hostttr124 rportd srotyp-dpndntdiffrncs in both th aount and sit of covanty bound C3b on th surfac of opsonizd pnuococcacs, as w as diffrncs in th dgradativ procssingof bound C3b to iC3b and C3d. Rcnt studis in icaso suggst an additiona potntia contributing factor.SIGN-R1, a C-type ectin that is xprssd on acrophagsin th argina zon of th spn was shown to diatth uptak of both purifid CPS and S. pneumoniae125.
Th spn is known to pay a crucia part in th car-anc of bood-born pnuococci, and ic that ardficint in SIGN-R1 ar hyprsnsitiv to invasivpnuococca disas126. C-typ ctins, such asSIGN-R1 and its huan hooogus, xhibit oigosac-charid spcificity, and diffrncs in thir affinity forindividua CPS pnuococca typs woud undoubtdy infunc phagocytosis and bactria caranc. SIGN-R1coud aso infunc th prsntation of CPS antigns toth iun syst, thrby affcting th host’s capacity to ount an antibody rspons. Finay, Frnbro et al.127 rportd that th capsu provids a dgr of rsistancto spontanous or antibiotic-inducd autoysis, thrby
RE V I E W S
296 | APRI 2008 | VOUme 6 www.nau./ws/

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 10/14
Wzxflippase
Ligase?
+
Wzy capsular polysaccharidepolymerase
Clonal type
a pneumococc con inege
i determined by equence
nyi of et of
houekeeping gene,
oppoed to it cpur
erotype.
contributing to antibiotic toranc in cinica isoats.Intrstingy, this capacity aso varid significanty btwn capsuar srotyps.
Athough isognic S. pneumoniae xprsss diffr-nt CPS srotyps that xhibit arkd diffrncs inurin virunc, non-capsuar factors ar aso cary iportant128,129. mocuar pidioogica anaysishas donstratd that proprtis which ar associatdwith a particuar con type, in addition to capsuarsrotyp, infunc th potntia of S. pneumoniae tocaus invasiv disas in huans. Th contributionof host factors was donstratd in a subsquntstudy in which isoats that had high huan-invasionpotntia xhibitd significanty diffrnt viruncand disas kintics in BAB/c ic copard withC57B/6 ic130.
Regulation of CPS production. Th transition fronasopharynga coonization to invasiv disas is cary a watrshd in th rationship btwn S. pneumoniae and its host, and invovs a ajor switch in th xprs-
sion of iportant virunc dtrinants as th pathognadapts to its atrd icronvironnt. maxia xprs-sion of capsu is ssntia for systic virunc, but thxtnt of xposur of othr iportant pnuococcasurfac structurs, such as adhsins, is aso infuncdby capsuar thicknss. Non-ncapsuatd pnuococcixhibit gratr adhrnc to huan rspiratory pith-ia cs (A549) in vitro copard with isognic driva-tivs that xprss ithr srotyp 3 or 19F capsus 131. Furthror, prviousy ncapsuatd pnuococci
s to xprss itt CPS during intiat contact withrspiratory pithia cs in vitro or in vivo132.
Th capacity to rguat CPS production at th tran-scriptiona, transationa or post-transationa v isiportant for th surviva of S. pneumoniae in diffrnthost nvironnts. To dat, no transcriptiona-contronts hav bn idntifid in association with thpnuococca σ70 cps prootr133. Howvr, thr isso vidnc to suggst that th v of xprssion of th cps ocus diffrs btwn th transparnt and opaquphas variants, as or CpsD was dtctab by wstrniunobotting in th opaqu phas variants28. Th vof S. pneumoniae cps RNA, rativ to 16S ribosoaRNA, that was isoatd fro th bood of infctd icwas shown to b approxiaty fourfod highr copardwith th vs in S. pneumoniae that had bn grownin vitro134. Howvr, significant diffrncs in cps RNAvs coud not b dtctd btwn pnuococci thatwr isoatd fro th nasopharynx, bood or ungs of infctd ic135.
Th first four gns of th cps ocus (cpsA–D) ar co-
on to a pnuococca srotyps, with th xcptionof srotyps 3 and 37. Th protins that ar ncodd by ths gns, CpsA–D, hav bn shown to affct th vof CPS xprssion136. Th cntra rgion of th cps ocuscopriss gns that ncod spcific gycosy transfrassthat assb th srotyp-spcific oigosaccharid-rpat unit on a ipid carrir. This rgion aso incudsa fippas (Wzx) that transports th rpat unit to thxtrna fac of th bran and a poyras (Wzy)that inks th units togthr. Th fina rgion of th ocuscopriss gns that ncod th synthsis of activatdsugar prcursors (FIG. 3).
Intrstingy, a CpsA hooogu in group B strp-tococci ss to function as a transcriptiona activa-tor137; howvr, to dat, th cpsA gn has not bnshown to ipact on cps transcription in S. pneumoniae.Nvrthss,cpsA-dtion utants producd rducdvs of CPS138–140. CPS biosynthsis of a but two pnu-ococca srotyps has bn shown to b dpndntupon a rguatory syst that is dtrind by CpsB,CpsC and CpsD. CpsB is a angans-dpndnt phos-photyrosin-protin phosphatas, CpsC is a branprotin that is ratd to poysaccharid co-poyrassand CpsD is an autophosphoryating protin-tyrosinkinas136,138,139. CpsC is rquird for CpsD tyrosin auto-phosphoryation; a cpsC-dtion utant is rough, andCpsD dos not bco phosphoryatd. mutation of th
cpsD gn to inactivat th ATP-binding sit iinatdCPS production. It has aso bn shown that CpsB isrquird to dphosphoryat CpsD; in cpsB-dtionutants, th proportion of CpsD that is phosphor-yatd incrass draaticay, and thr is a significantdcras in th aount of CPS that is producd. Thsobsrvations suggst that th non-phosphoryatd forof CpsD is activ in CPS biosynthsis136,138,139. Rcnty,a nov ro for CpsC in th attachnt of CPS to thpnuococca c wa was aso idntifid141. Thrfor,CpsB, CpsC and CpsD function togthr to rguatCPS assby, xport and attachnt to th c waby tyrosin phosphoryation of CpsD (FIG. 4).
Figure 3 | Sps n Streptococcus pneumoniae apsu snss. Capsule
biosynthesis is regulated by the cps locus, which encodes specific glycosyltransferases
that assemble the oligosaccharide repeats on the cytoplasmic face of the membrane, and
a flippase (Wzx), which transports the repeat units to the external surface of the
membrane. At this location, the repeat units are polymerized by Wzy to form high-
molecular-weight capsular polysaccharides, which are then, presumably, ligated to the
cell wall. The cps locus also encodes enzymes for the synthesis of activated sugar
precursors, as well as conserved genes that are involved in regulation.
RE V I E W S
NATURe ReVIeWS | microbiology VOUme 6 | APRI 2008 | 297

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 11/14
P P
P P
ATP ATP
CpsC CpsC
CpsD CpsD
Ligase
P P
Ligase
PP
Cell wall Cell wall
a b
WzyWzx WzyWzx
P P P P
1
2
3
4
CpsB
CpsC CpsC
CpsB
CpsD CpsD
Intrstingy, Rgm, a hooogu of th staphyococcacataboit contro protin CcpA, which is invovd in thrguation of sugar-tabois pathways, has bn shownto affct transcription of th cps ocus, which suggsts thata carbon sourc ight aso infunc capsuar xprs-sion142. Indd, two protins that ar invovd in sugartabois hav bn shown to affct CPS production.Pg is th phosphogucoutas that catayss th con-
vrsion of gucos-6-phosphat to gucos-1-phosphat,and GaU is a gucos-1-phosphat uridyytransfrasthat catayss th foration of uridin diphosphat-gucos (UDP-Gc) fro gucos-1-phosphat. mutantsof S. pneumoniae in which ithr th galU or pgm gn wasdisruptd producd aost no CPS and xhibitd growthdfcts137,143. Additionay, pnuococca strains in whichth pgm gn had dfind point utations that signifi-canty rducd but did not iinat nzyatic activity sti producd rducd aounts of CPS, vn thoughth utants no ongr xhibitd growth dfcts144. BothPg and GaU ar rquird for th synthsis of UDP-Gc,which is a prcursor for th biosynthsis of a 90 pnuo-cocca CPS typs, as w as othr cuar structurs, suchas tichoic acid. Thus, iiting th suppy of this prcursorwoud b xpctd to ipact haviy upon CPS produc-
tion in th pnuococcus. Indirct oduation of CPS
production by controing th avaiabiity of prcursors orco-factors coud b on of th rguatory chaniss thatis usd by S. pneumoniae.
Conclusions
Th pnuococcus has provn to b a particuary for-idab fo, and can vad iination by sru thrapy,chothrapy, utip casss of antibiotic and, orrcnty, conjugat poysaccharid vaccins. For ach of ths approachs, thr was considrab initia confi-dnc that Sir Wiia Osr’s ‘captain of th n of dath’had finay bn dat a fata bow. Th adaptabiity of thorganis in rspons to an array of sctiv prssurs,howvr, has aowd it to rtain its capacity to coonizits huan host without dcrasing its fitnss to th pointof diinishd virunc. Th abiity of S. pneumoniae toadapt sts fro th rarkab pasticity and htrog-nity of its gno. Who-gno squncing of argnubrs of pnuococca isoats is now rvaing bothth xtnt of strain-to-strain variation in gn contntand aso th siz of th S. pneumoniae pan-gno, as
w as othr strptococci that provid an additionarsrvoir of gntic inforation.
For th ost part, th initia focus of invstigationsof S. pneumoniae was iitd to th ros of its variouscapsu srotyps and th chaniss that undrpin itsnatura coptnc for th uptak of DNA. As outindin this Rviw, advancs in ocuar bioogy and bact-ria gntics hav not ony faciitatd a fur undrstand-ing of th bioogy of its capsu and coptnc, but havaso aowd charactrization of th any attributs thatcontribut to bactria–host intractions. In addition, rp-rsntativ ods of pnuococca infction ar nowbing appid in gnticay odifid ic to dfin thhost factors and othr aspcts of th iun rsponsthat contribut to coonization and disas. Ths studishav, for xap, phasizd th iportanc of CD4+ T cs, rathr than antibody rsponss, in th diationof iunity to S. pneumoniae (BOX 2). Th copxity of th bactria and host factors that hav bn iuinatdby ths studis has aso providd insight into novstratgis for antiicrobias and a nxt gnration of or broady ffctiv pnuococca vaccins that wius cobinations of protin antigns (BOX 1). Dspit ourprogrss, ky aspcts of pnuococca bioogy, such asth bactria and host factors that ipact on pathogntransission, rain argy unxpord. It is sti pr-atur, thrfor, to assu that th pnuococcus has
yidd a of its ost iportant scrts.
Figure 4 | md swn uan apsua psaad (cPS)
pdun sn pspan cpsD. a | CpsC, CpsD and ATP interact to
promote CPS biosynthesis by the polysaccharide polymerase (step 1). CpsD
autophosphorylates, which causes a change in protein interactions and slows CPS
biosynthesis (step 2). | The CPS polymer is then transferred to the putative CPS cell-wall
ligase, and is ligated to the cell wall (step 3). Finally, CpsB dephosphorylates CpsD,thereby allowing the cycle to be repeated (step 4).
1. Balakrishnan, I., Crook, P., Morris, R. & Gillespie, S. H.
Early predictors of mortality in pneumococcal
bacteraemia. J. Infect. Dis. 40, 256–261 (2000).
2. Lim, W. S. et al. Study of community acquired
pneumonia aetiology (SCAPA) in adults admitted to
hospital: implications for management guidelines.
Thorax 50, 296–301 (2001).
3. Denny, F. W. & Loda, F. A. Acute respiratory infections
are the leading cause of death in children in developing
countries. Am. J. Trop. Med. Hyg . 35, 1–2 (1986).
4. Berkley, J. A. et al. Bacteremia among children
admitted to a rural hospital in Kenya. N. Engl. J. Med .
352, 39–47 (2005).
5. Bogaert, D., de Groot, R. & Hermans, P.
Streptococcus pneumoniae colonisation: the key to
pneumococcal disease. Lancet Infect. Dis. 4, 144–154
(2004).
6. Regev-Yochay, G. et al. Association between carriage
of Streptococcus pneumoniae and Staphylococcus
aureus in children. JAMA 292, 716–720 (2004).
7. Lexau, C. et al. Changing epidemiology of invasive
pneumococcal disease among older adults in the era
of pediatric pneumococcal conjugate vaccine. JAMA
294, 2043–2051 (2005).
8. McCool, T. L., Cate, T. R., Moy, G. & Weiser, J. N. The
immune response to pneumococcal proteins during
experimental human carriage. J. Exp. Med. 195,
359–365 (2002).
One of the few studies of pneumococcal
pathogenesis in humans. This paper characterizes
pneumococcal colonization and the mucosal
immune response in human volunteers. It also
provides strong circumstantial evidence for the
importance of PspA in the colonization of the
human nasopharynx by pneumococci.
9. Lipsitch, M. et al. Are anticapsular antibodies
the primary mechanism of protection against
invasive pneumococcal disease? PLoS Med. 2, e15
(2005).
RE V I E W S
298 | APRI 2008 | VOUme 6 www.nau./ws/

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 12/14
10. McCool, T. & Weiser, J. Limited role of antibody in
clearance of Streptococcus pneumoniae in a murine
model of colonisation. Infect. Immun. 72, 5807–5813
(2004).
First paper to clearly show that antibodies are not
required for the clearance of pneumococcal
colonization based on studies that used mice with
genetic defects in humoral immunity.
11. Nelson, A. et al. Capsule enhances pneumococcal
colonisation by limiting mucus-mediated clearance.
Infect. Immun. 75, 83–90 (2007).
12. Kamerling, J. in Streptococcus pneumoniae: Molecular Biology & Mechanisms of Disease (ed. Tomasz, A.)
81–114 (Mary Ann Liebert, New York, 2000).
13. Weiser, J. N., Austrian, R., Sreenivasan, P. K. &
Masure, H. R. Phase variation in pneumococcal
opacity: relationship between colonial morphology
and nasopharyngeal colonisation. Infect. Immun. 62,
2582–2589 (1994).
Describes pneumococcal phase variation and its
important role in nasopharyngeal colonization and
invasive disease, thereby providing an interesting
insight into the interaction of the pneumococcus
with its host.
14. Cundell, D. R., Gerard, N. P., Gerard, C., Idanpaan-
Heikkila, I. & Tuomanen, E. I. Streptococcus
pneumoniae anchor to activated human cells by the
receptor for platelet-activating factor. Nature 377,
435–438 (1995).
Shows that the pneumococcus binds to a receptor
that is now recognized as being used by several
other important pathogens that reside in the airway.
15. Weiser, J. in Colonisation of Mucosal Surfaces
(ed. Nataro, J.) 61–72 (ASM, Washington DC, 2005).
16. Weiser, J. N. et al. Phosphorylcholine on the
lipopolysaccharide of Haemophilus influenzae
contributes to persistence in the respiratory tract and
sensitivity to serum killing mediated by C-reactive
protein. J. Exp. Med. 187, 631–640 (1998).
17. Rosenow, C. et al. Contribution of novel choline-
binding proteins to adherence, colonisation and
immunogenicity of Streptococcus pneumoniae. Mol.
Microbiol. 25, 819–829 (1997).
18. Hammerschmidt, S., Talay, S. R., Brandtzaeg, P. &
Chhatwal, G. S. SpsA, a novel pneumococcal surface
protein with specific binding to secretory
immunoglobulin A and secretory component. Mol.
Microbiol. 25, 1113–1124 (1997).
19. Barocchi, M. et al. A pneumococcal pilus influences
virulence and host inflammatory responses. Proc. Natl
Acad. Sci. USA 103, 2857–2862 (2006).20. Hava, D. & Camilli, A. Large-scale identification of
serotype 4 Streptococcus pneumoniae virulencefactors. Mol. Microbiol. 45, 1389–1406 (2002).
Important study that used signature-tagged
mutagenesis to identify 387 pneumococcal mutants
that were attenuated in murine models of pneumonia.
21. Andersson, B. et al. Identification of an active
dissaccharide unit of a glycoconjugate receptor for
pneumococci attaching to human pharyngeal epithelial
cells. J. Exp. Med. 158, 559–570 (1983).22. Krivan, H. C., Roberts, D. D. & Ginsberg, V. Many
pulmonary pathogenic bacteria bind specifically to the
carbohydrate sequence GalNAcβ1–4Gal found in
some glycolipids. Proc. Natl Acad. Sci. USA 85,
6157–6161 (1988).
23. King, S., Hippe, K. & Weiser, J. Deglycosylation of
human glycoconjugates by the sequential activities of
exoglycosidases expressed by Streptococcus
pneumoniae. Mol. Microbiol. 59, 961–974 (2006).
24. Jedrzejas, M., Mello, L., de Groot, B. & Li, S.
Mechanism of hyaluronan degradation by
Streptococcus pneumoniae hyaluronate lyase.Structures of complexes with the substrate. J. Biol.
Chem. 277, 28287–28297 (2002).25. Holmes, A. et al. The pavA gene of Streptococcus
pneumoniae encodes a fibronectin-binding protein
that is essential for virulence. Mol. Microbiol. 41,
1395–1408 (2001).
26. Bergmann, S., Rohde, M., Chhatwal, G. &
Hammerschmidt, S. α-Enolase of Streptococcus
pneumoniae is a plasmin(ogen)-binding protein
displayed on the bacterial cell surface. Mol. Microbiol.
40, 1273–1287 (2001).
27. Kim, J. O. et al. Relationship between cell-surface
carbohydrates and intrastrain variation on
opsonophagocytosis of Streptococcus pneumoniae.
Infect. Immun. 67, 2327–2333 (1999).
28. Weiser, J. et al. Changes in availability of oxygen
accentuate differences in capsular polysaccharide
expression by phenotypic variants and clinical isolates
of Streptococcus pneumoniae. Infect. Immun. 69,
5430–5439 (2001).29. van Rossum, A., Lysenko, E. & Weiser, J. Host and
bacterial factors contributing to the clearance of
colonisation by Streptococcus pneumoniae in a murine
model. Infect. Immun. 73, 7718–7726 (2005).
30. Ratner, A. et al. Epithelial cells are sensitive detectors
of bacterial pore-forming toxins. J. Biol. Chem. 281,
12994–12998 (2006).31. Malley, R. et al. CD4+ T cells mediate antibody-
independent acquired immunity to pneumococcal
colonisation. Proc. Natl Acad. Sci. USA 102,4848–4853 (2005).
32. Malley, R. et al. Recognition of pneumolysin by Toll-
like receptor 4 confers resistance to pneumococcal
infection. Proc. Natl Acad. Sci. USA 100, 1966–1971
(2003).
Demonstrates that the pneumococcal toxin
pneumolysin triggers inflammatory responses in
host macrophages by interacting with TLR4. Such
signalling is crucial for the innate immune response
to the pneumococcus, and is a paradigm of the fine
balance between the protective and deleterious
effects of innate inflammatory responses to
mucosal pathogens.
33. Wani, J., Gilbert, J., Plaut, A. & Weiser, J.
Identification, cloning and sequencing of the
immunoglobulin A1 protease gene of Streptococcus
pneumoniae. Infect. Immun. 64, 3967–3974 (1996).
34. Szalai, A. J., Briles, D. E. & Volanakis, J. E. Human
C-reactive protein is protective against fatal
Streptococcus pneumoniae infection in transgenic
mice. J. Immunol. 155, 2557–2563 (1995).
35. Gould, J. & Weiser, J. The inhibitory effect of C-reactive
protein on bacterial phosphorylcholine-platelet
activating factor receptor mediated adherence is blocked
by surfactant. J. Infect. Dis. 186, 361–371 (2002).36. Aas, J. A. et al. Defining the normal bacterial flora of
the oral cavity. J. Clin. Microbiol. 43, 5721–5732
(2005).37. Lysenko, E. S., Ratner, A. J., Nelson, A. L. & Weiser,
J. N. The role of innate immune responses in the
outcome of interspecies competition for colonisation
of mucosal surfaces. PLoS Pathog. 1, 1–9 (2005).
38. Dawid, S., Roche, A. & Weiser, J. The blp bacteriocins
of Streptococcus pneumoniae mediate intraspecies
competition both in vitro and in vivo. Infect. Immun.
75, 443–451 (2007).
39. Dowson, C., Coffey, T. & Spratt, B. Origin and
molecular epidemiology of penicillin-binding-protein-
mediated resistance to β-lactam antibiotics. Trends
Microbiol. 2, 361–366 (1994).
40. Lock, R. A., Zhang, Q. Y., Berry, A. M. & Paton, J. C.Sequence variation in the Streptococcus pneumoniae
pneumolysin gene affecting haemolytic activity and
electrophoretic mobility of the toxin. Infect. Immun.
21, 71–83 (1996).
41. Kirkham, L. A. S. et al. Identification of invasive
serotype 1 pneumococcal isolates that express
nonhemolytic pneumolysin. J. Clin. Microbiol. 44,
151–159 (2006).42. Tilley, S., Orlova, E., Gilbert, R., Andrew, P. & Saibil, H.
Structural basis of pore formation by the bacterial
toxin pneumolysin. Cell 121, 247–256 (2005).43. Hirst, R., Kadioglu, A., O’Callaghan, C. & Andrew, P.
The role of pneumolysin in pneumococcal pneumonia
and meningitis. Clin. Exp. Immunol. 138, 195–201
(2004).44. Kadioglu, A., Coward, W., Colston, M., Hewitt, C. &
Andrew, P. CD4-T-lymphocyte interactions with
pneumolysin and pneumococci suggest a crucial
protective role in the host response to pneumococcal
infection. Infect. Immun. 72, 2689–2697 (2004).The first study to demonstrate an early protective
role for CD4+ T cells during pneumococcal
pneumonia in vivo.
45. Mitchell, T. J., Andrew, P. W., Saunders, F. K., Smith,
A. N. & Boulnois, G. J. Complement activation and
antibody binding by pneumolysin via a region
homologous to a human acute phase protein. Mol.
Microbiol. 5, 1883–1888 (1991).
46. Rubins, J. et al. Distinct roles for pneumolysin’s
cytotoxic and complement activities in the
pathogenesis of pneumococcal pneumonia. Am.
J. Respir. Crit. Care Med. 153, 1339–1346 (1996).47. Alexander, J. E. et al. Amino acid changes affecting
the behaviour of pneumococci in pneumonia. Microb.
Pathog. 24, 167–174 (1998).
48. Jounblat, R., Kadioglu, A., Mitchell, T. & Andrew, P.
Pneumococcal behavior and host responses during
bronchopneumonia are affected differently by the
cytolytic and complement-activating activities of
pneumolysin. Infect. Immun. 71, 1813–1819 (2003).49. Baba, H. et al. Induction of gamma interferon and
nitric oxide by truncated pneumolysin that lacks pore-
forming activity. Infect. Immun. 70, 107–113 (2002).
50. Berry, A. M. et al. Effect of defined point mutations in
the pneumolysin gene on the virulence of
Streptococcus pneumoniae. Infect. Immun. 63,
1969–1974 (1995).51. Canvin, J. R. et al. The role of pneumolysin and
autolysin in the pathology of pneumonia and
septicaemia in mice infected with a type 2pneumococcus. J. Infect. Dis. 172, 119–123 (1995).
52. Alexander, J. E. et al. Immunization of mice with
pneumolysin toxoid confers a significant degree of
protection against at least nine serotypes of
Streptococcus pneumoniae. Infect. Immun. 62,
5683–5688 (1994).
53. Kadioglu, A. et al. Upper and lower respiratory tract
infection by Streptococcus pneumoniae is affected by
pneumolysin deficiency and differences in capsule
type. Infect. Immun. 70, 2886–2890 (2002).
54. Orihuela, C. J. , Gao, G. L., Francis, K. P., Yu, J. &
Tuomanen, E. I. Tissue-specific contributions of
pneumococcal virulence factors to pathogenesis.
J. Infect. Dis. 190, 1661–1669 (2004).55. Berry, A. M., Yother, J., Briles, D. E., Hansman, D. &
Paton, J. C. Reduced virulence of a defined
pneumolysin-negative mutant of Streptococcus
pneumoniae. Infect. Immun. 57, 2037–2042 (1989).
56. Berry, A. M., Ogunniyi, A. D., Miller, D. C. & Paton,
J. C. Comparative virulence of Streptococcus
pneumoniae strains with insertion-duplication, point,
and deletion mutations in the pneumolysin gene.
Infect. Immun. 67, 981–985 (1999).
57. Kadioglu, A. et al. Host cellular immune response to
pneumococcal lung infection in mice. Infect. Immun.
68, 1557–1562 (2000).
Demonstrated the central role of pneumolysin in
driving the pattern of inflammation and cellular
infiltration into the lungs in vivo. Also the first to
show an early, pneumolysin-dependent involvement
of T cells in respiratory infection.
58. Benton, K. A., Everson, M. P. & Briles, D. E.
A pneumolysin negative mutant of Streptococcus
pneumoniae causes chronic bacteremia rather than
acute sepsis in mice. Infect. Immun. 63, 448–455
(1995).
59. Friedland, I. R. et al. The limited role of pneumolysin
in the pathogenesis of pneumococcal meningitis.
J. Infect. Dis. 172, 805–809 (1995).
60. Braun, J. et al. Pneumococcal pneumolysin and H2O
2
mediate brain cell apoptosis during meningitis. J. Clin. Invest. 109, 19–27 (2002).
61. Wellmer, A. et al. Decreased virulence of a
pneumolysin deficient strain of Streptococcus
pneumoniae in murine meningitis. Infect. Immun. 70,
6504–6508 (2002).
62. Winter, A. J. et al. in Proc. 7th Intern. Cong. Infect.
Dis. Abstr. 73.004 (International Society for Infectious
Diseases, Brookline,1996).
63. Hirst, R. A. et al. Relative roles of pneumolysin and
hydrogen peroxide from Streptococcus pneumoniae in
inhibition of ependymal ciliary beat frequency. Infect.
Immun. 68, 1557–1562 (2000).
64. Hirst, R., Mohammed, B., Mitchell, T., Andrew, P. &
O’Callaghan, C. Streptococcus pneumoniae-induced
inhibition of rat ependymal cilia is attenuated by
antipneumolysin antibody. Infect. Immun. 72,
6694–6698 (2004).
65. Hirst, R. A., Kadioglu, A., O’Callaghan, C. & Andrew,
P. W. The role of pneumolysin in pneumococcal
pneumonia and meningitis. Clin. Exp. Immunol. 138,195–201 (2004).
66. Chanter, N. Streptococcus pneumoniae and equine
disease. Equine Vet. J. 26, 5–6 (1994).
67. Whatmore, A. M. et al. Molecular characterization of
equine isolates of Streptococcus pneumoniae: natural
disruption of genes encoding the virulence factors
pneumolysin and autolysin. Infect. Immun. 67,
2776–2782 (1999).68. Jedrzejas, M. J. Pneumococcal virulence factors:
structure and function. Microbiol. Mol. Biol. Rev. 65,
187–207 (2001).
69. Bergmann, S. & Hammerschmidt, S. Versatility of
pneumococcal surface proteins. Microbiology 152,
295–303 (2006).
70. Jedrzejas, M. J., Lamani, E. & Becker, R. S.
Characterization of selected strains of pneumococcal
surface protein A. J. Biol. Chem. 276, 33121–33128
(2001).
RE V I E W S
NATURe ReVIeWS | microbiology VOUme 6 | APRI 2008 | 299

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 13/14
71. Hollingshead, S., Becker, R. & Briles, D. Diversity of
PSDPA: mosaic genes and evidence for past
recombination in Streptococcus pneumoniae. Infect.
Immun. 68, 5889–5900 (2000).
72. Shaper, M., Hollingshead, S. K., Benjamin, W. H. &
Briles, D. E. PspA protects Streptococcus pneumoniae
from killing by apolactoferrin, and antibody to PspA
enhances killing of pneumococci by apolactoferrin.
Infect. Immun. 72, 5031–5040 (2004).73. Briles, D. E. & Mirza, S. PspA inhibits the antibacterial
effect of lactoferrin on Streptococcus pneumoniae.
Biochem. Cell Biol. 84, 401 (2006).74. McDaniel, L. S. et al. Use of insertional inactivation to
facilitate studies of biological properties of
pneumococcal protein A (PspA). J. Exp. Med. 165,
381–394 (1987).75. Ren, B., Szalai, A. J., Hollingshead, S. K. & Briles, D. E.
Effects of PspA and antibodies to PspA on activation
and deposition of complement on the pneumococcal
surface. Infect. Immun. 72, 114–122 (2004).
76. Berry, A. & Paton, J. Additive attenuation of virulence
of Streptococcus pneumoniae by mutation of genes
encoding pneumolysin and other putative
pneumococcal virulence proteins. Infect. Immun. 68,
133–140 (2000).
77. Abeyta, M., Hardy, G. G. & Yother, J. Genetic
alteration of capsule type but not PspA type affects
accessibility of surface-bound complement and surface
antigens of Streptococcus pneumoniae. Infect. Immun.
71, 218–225 (2003).
78. Rosenow, C. et al. Contribution of novel choline-
binding proteins to adherence, colonisation and
immunogenicity of Streptococcus pneumoniae. Mol.
Microbiol. 25, 819–829 (1997).
79. Hammerschmidt, S., Tillig, M., Wolff, S. & Chaatwal, J.
Species specific binding of human secretory
component to SpsA protein of Streptococcus
pneumoniae via a hexapeptide motif. Mol. Microbiol.
36, 726–736 (2000).
80. Zhang, J. et al. The polymeric immunoglobulin
receptor translocates pneumococci across human
nasopharyngeal epithelial cells. Cell 102, 827–837
(2000).81. Iannelli, F., Chiavolini, D., Ricci, S., Oggioni, M. R. &
Pozzi, G. Pneumococcal surface protein C contributes
to sepsis caused by Streptococcus pneumoniae in
mice. Infect. Immun. 72, 3077–3080 (2004).82. Quin, L. R. et al. In vivo binding of complement
regulator factor H by Streptococcus pneumoniae.
J. Infect. Dis. 192, 1996–2003 (2005).83. Janulczyk, R., Iannelli, F., Sjoholm, A. G., Pozzi, G. &
Bjorck, L. Hic, a novel surface protein of Streptococcus
pneumoniae that interferes with complement function. J. Biol. Chem. 275, 37257–37263 (2000).
84. Dave, S., Carmicle, S., Hammerschmidt, S.,
Pangburn, M. & McDonald, L. Dual roles of PspC, a
surface protein of Streptococcus pneumoniae, in
binding human secretory IgA and factor H. J. Immunol.
173, 471–477 (2004).
85. Cheng, Q., Finkel, D. & Hostetter, M. K. Novel
purification scheme and functions for a C3-binding
protein from Streptococcus pneumoniae. Biochemistry
39, 5450–5457 (2000).
86. Howard, L. V. & Gooder, H. Specificity of autolysin of
Streptococcus (Diplococcus) pneumoniae. J. Bacteriol.
117, 796–804 (1974).
87. Berry, A. M., Lock, R. A., Hansman, D. & Paton, J. C.
Contribution of autolysin to virulence of Streptococcus
pneumoniae. Infect. Immun. 57, 2324–2330 (1989).
88. Overweg, K. et al. The putative proteinase maturation
protein A of Streptococcus pneumoniae is a conserved
surface protein with potential to elicit protective immune
responses. Infect. Immun. 68, 4180–4188 (2000).89. Hermans, P. W. M. et al. The streptococcal lipoprotein
rotamase A (SlrA) is a functional peptidyl-prolyl
isomerase involved in pneumococcal colonisation.
J. Biol. Chem. 281, 968–976 (2006).
90. Berry, A. M. & Paton, J. C. Sequence heterogeneity of
PsaA, a 37-kilodalton putative adhesin essential for
virulence of Streptococcus pneumoniae. Infect.
Immun. 64, 5255–5262 (1996).91. Marra, A., Lawson, S., Asundi, J. S., Brigham, D. &
Hromockyj, A. E. In vivo characterization of the psa
genes from Streptococcus pneumoniae in multiple
models of infection. Microbiology 148, 1483–1491
(2002).
92. Johnson, S. E. et al. Inhibition of pneumococcal
carriage in mice by subcutaneous immunization with
peptides from the common surface protein
pneumococcal surface adhesin A. Infect. Immun. 185,
489–496 (2002).
93. Sampson, J. S., O’Connor, S. P., Stinson, A. R.,
Tharpe, J. A. & Russell, H. Cloning and nucleotide-
sequence analysis of psaA, the Streptococcus
pneumoniae gene encoding a 37-kilodalton protein
homologous to previously reported Streptococcus sp.
adhesins. Infect. Immun. 62, 319–324 (1994).
94. Briles, D. E. et al. Intranasal immunization of mice
with a mixture of the pneumococcal proteins PsaA and
PspA is highly protective against nasopharyngeal
carriage of Streptococcus pneumoniae. Infect. Immun.
68, 796–800 (2000).
95. Romero-Steiner, S. et al. Inhibition of pneumococcaladherence to human nasopharyngeal epithelial cells
by anti-PsaA antibodies. Clin. Diagn. Lab. Immunol.
10, 246–251 (2003).
96. Dintilhac, A., Alloing, G., Granadel, C. & Claverys, J. P.
Competence and virulence of Streptococcus
pneumoniae: Adc and PsaA mutants exhibit a
requirement for Zn and Mn resulting from inactivation
of putative ABC metal permeases. Mol. Microbiol. 25,
727–739 (1997).
97. McAllister, L. J. et al. Molecular analysis of the psa
permease complex of Streptococcus pneumoniae.
Mol. Microbiol . 53, 889–901 (2004).
98. Lawrence, M. C. et al. The crystal structure of
pneumococcal surface antigen PsaA reveals a metal-
binding site and a novel structure for a putative ABC-
type binding protein. Structure 15, 1553–1561
(1998).
99. Johnston, J. W. et al. Lipoprotein PsaA in virulence of
Streptococcus pneumoniae: surface accessibility and
role in protection from superoxide. Infect. Immun. 72,
5858–5867 (2004).100. Tseng, H. J., McEwan, A. G., Paton, J. C. & Jennings,
M. P. Virulence of Streptococcus pneumoniae: PsaA
mutants are hypersensitive to oxidative stress. Infect.
Immun. 70, 1635–1639 (2002).
101. Brown, J. S., Gilliland, S. M. & Holden, D. W.
A Streptococcus pneumoniae pathogenicity island
encoding an ABC transporter involved in iron uptake
and virulence. Mol. Microbiol. 40, 572–585 (2001).
102. Brown, J. S., Gilliland, S. M., Ruiz-Albert, J. & Holden,
D. W. Characterization of Pit, a Streptococcus
pneumoniae iron uptake ABC transporter. Infect.
Immun. 70, 4389–4398 (2002).
103. Brown, J. S., Ogunniyi, A. D., Woodrow, M. C., Holden,
D. W. & Paton, J. C. Immunization with components of
two iron uptake ABC transporters protects mice
against systemic Streptococcus pneumoniae infection.
Infect. Immun. 69, 6702–6706 (2001).104. Chen, S., Paterson, G. K., Tong, F. H., Mitchell, T. J. &
DeMaria, T. F. Sortase A contributes to pneumococcal
nasopharyngeal colonisation in the chinchilla model.FEMS Microbiol. Lett. 253, 151–154 (2005).
105. Paterson, G. K. & Mitchell, T. J. The role of
Streptococcus pneumoniae sortase A in colonisation
and pathogenesis. Microbes Infect. 8, 145–153
(2006).
106. Kharat, A. S. & Tomasz, A. Inactivation of the srtA
gene affects localization of surface proteins and
decreases adhesion of Streptococcus pneumoniae to
human pharyngeal cells in vitro. Infect. Immun. 71,
2758–2765 (2003).
107. King, S. J. et al. Phase variable desialylation of host
proteins that bind to Streptococcus pneumoniae
in vivo and protect the airway. Mol. Microbiol. 54,
159–171 (2004).108. Pettigrew, M. M., Fennie, K. P., York, M. P., Daniels, J.
& Ghaffar, F. Variation in the presence of
neuraminidase genes among Streptococcus
pneumoniae isolates with identical sequence types.
Infect. Immun. 74, 3360–3365 (2006).
109. Berry, A. M., Lock, R. A. & Paton, J. C. Cloning andcharacterization of nanB, a second Streptococcus
pneumoniae neuraminidase gene, and purification of
the NanB enzyme from recombinant Escherichia coli .
J. Bacteriol. 178, 4854–4860 (1996).
110. Manco, S. et al. Pneumococcal neuraminidases A and
B both have essential roles during infection of the
respiratory tract and sepsis. Infect. Immun. 74,
4014–4020 (2006).111. Oggioni, M. R. et al. Switch from planktonic to sessile
life: a major event in pneumococcal pathogenesis.
Mol. Microbiol . 61, 1196–1210 (2006).
Describes the interesting role of CSP in
pneumococcal virulence and biofilm formation.
Also demonstrates different patterns of
pneumococcal-gene expression during infection in
the host: one that is typical of bacteria in blood
and one that is typical of bacteria in tissue, such as
brain and lung.
112. Sorensen, U. B. S., Blom, J., Birch-Andersen, A. &
Henrichsen, J. Ultrastructural localization of capsules,
cell wall polysaccharide, cell wall proteins, and F
antigen in pneumococci. Infect. Immun. 56,
1890–1896 (1988).
113. Sorensen, U. B. S., Henrichsen, J., Chen, H. C. & Szu,
S. C. Covalent linkage between the capsular
polysaccharide and the cell wall peptidoglycan of
Streptococcus pneumoniae revealed by
immunochemical methods. Microb. Pathog . 8,
325–334 (1990).
114. Henrichsen, J. Six newly recognized types of Streptococcus pneumoniae. J. Clin. Microbiol . 33,
2759–2762 (1995).
115. Austrian, R. Some observations on the pneumococcus
and on the current status of pneumococcal disease
and its prevention. Rev. Infect. Dis. 3, S1–S17
(1981).
116. Martin, M. et al. An outbreak of conjunctivitis due to
atypical Streptococcus pneumoniae. N. Engl. J. Med .
348, 1112–1121 (2003).
117. Crum, N. F., Barrozo, C. P., Chapman, F. A., Ryan,
M. A. & Russell, K. L. An outbreak of conjunctivitis
due to a novel unencapsulated Streptococcus
pneumoniae among military trainees. Clin. Infect. Dis.
39, 1148–1154 (2004).118. Lee, C. J., Banks, S. D. & Li, J. P. Virulence, immunity
and vaccine related to Streptococcus pneumoniae.
Crit. Rev. Microbiol. 18, 89–114 (1991).
119. Winkelstein, J. A. The role of complement in the host’s
defense against Streptococcus pneumoniae. Rev.
Infect. Dis. 3, 289–298 (1981).
120. Musher, D. M. Infections caused by Streptococcus
pneumoniae: clinical spectrum, pathogenesis,
immunity and treatment. Clin. Infect. Dis. 14,
801–807 (1992).
121. Abeyta, M., Hardy, G. G. & Yother, Y. Genetic
alteration of capsule type but not PspA type affects
accessibility of surface-bound complement and surface
antigens of Streptococcus pneumoniae. Infect. Immun.
71, 218–225 (2003).
122. Wartha, F. et al . Capsule and d-alanylated lipoteichoic
acids protect Streptococcus pneumoniae against
neutrophil extracellular traps. Cell. Microbiol . 9,
1162–1171 (2007).
123. MacLeod, C. M. & Krauss, M. R. Relation of virulence
of pneumococcal strains for mice to the quantity of
capsular polysaccharide formed in vitro. J. Exp. Med.
92, 1–9 (1950).
Important early paper that demonstrates the in
vivo role of the capsule in pneumococcal virulence.
124. Hostetter, M. K. Serotypic variations among virulent
pneumococci in deposition and degradation of covalently bound C3b: implications for phagocytosis
and antibody production. J. Infect. Dis. 153,
682–693 (1986).
125. Kang, Y. S. et al. The C-type lectin SIGN-R1 mediates
uptake of the capsular polysaccharide of
Streptococcus pneumoniae in the marginal zone of the
mouse spleen. Proc. Natl Acad. Sci. USA 101,
215–220 (2004).
126. Lanoue, A. et al. SIGN-R1 contributes to protection
against lethal pneumococcal infection in mice. J. Exp.
Med . 200, 1383–1393 (2004).127. Fernebro. J. et al. Capsular expression in
Streptococcus pneumoniae negatively affects
spontaneous and antibiotic-induced lysis and
contributes to antibiotic tolerance. J. Infect. Dis. 189,
328–338 (2004).
128. Kelly, T., Dillard, J. P. & Yother, J. Ef fect of genetic
switching of capsular type on virulence of
Streptococcus pneumoniae. Infect. Immun. 62,
1813–1819 (1994).129. Nesin, M., Ramirez, M. & Tomasz, A. Capsular
transformation of a multidrug-resistant Streptococcus
pneumoniae in vivo. J. Infect. Dis. 177, 707–713
(1998).
130. Sandgren, A. et al. Virulence in mice of pneumococcal
clonal types with known invasive disease potential in
humans. J. Infect. Dis. 192, 791–800 (2005).
131. Talbot, U., Paton, A. W. & Paton, J. C. Uptake of
Streptococcus pneumoniae by respiratory epithelial
cells. Infect. Immun. 64, 3772–3777 (1996).
132. Hammerschmidt, S. et al. Illustration of pneumococcal
polysaccharide capsule during adherence and invasion
of epithelial cells. Infect. Immun. 73, 4653–4667
(2005).
133. Muñoz, R., Mollerach, M., López, R. & García, E.
Molecular organization of the genes required for the
synthesis of type 1 capsular polysaccharide of
Streptococcus pneumoniae: formation of binary
RE V I E W S
300 | APRI 2008 | VOUme 6 www.nau./ws/

8/3/2019 Pneumo Virulence Factors
http://slidepdf.com/reader/full/pneumo-virulence-factors 14/14
encapsulated pneumococci and identification of
cryptic dTDP-rhamnose biosynthesis genes. Mol.
Microbiol . 25, 79–92 (1997).
134. Ogunniyi, A. D. , Giammarinaro, P. & Paton, J. C. The
genes encoding virulence-associated proteins and the
capsule of Streptococcus pneumoniae are upregulated
and differentially expressed in vivo. Microbiology
148, 2045–2053 (2002).
135. LeMessurier, K. S., Ogunniyi, A. D. & Paton, J. C.
Differential expression of key pneumococcal virulence
genes in vivo. Microbiology 152, 305–311 (2006).
136. Morona, J. K., Paton, J. C., Miller, D. C. & Morona, R.Tyrosine phosphorylation of CpsD negatively regulates
capsular polysaccharide biosynthesis in Streptococcus
pneumoniae. Mol. Microbiol. 35, 1431–1442
(2000).
Provides the first evidence for the post-
translational regulation of capsule expression
through tyrosine phosphorylation of a conserved
capsule-biosynthesis protein. The mechanism
probably operates in diverse encapsulated
bacteria, and is a potential drug target.
137. Cieslewicz, M. J., Kasper, D. L., Wang, Y. & Wessels,
M. R. Functional analysis in type Ia group B
Streptococcus of a cluster of genes involved in
extracellular polysaccharide production by diverse
species of streptococci. J. Biol. Chem. 276, 139–146
(2001).
138. Morona, J. K., Morona, R., Miller, D. C. & Paton, J. C.
Mutational analysis of the carboxy-terminal (YGX)4
repeat domain of CpsD, an autophosphorylating
tyrosine kinase required for capsule biosynthesis in
Streptococcus pneumoniae. J. Bacteriol. 185,
3009–3019 (2003).
139. Morona, J. K., Miller, D. C., Morona, R. & Paton,
J. C. The effect that mutations in the conserved
capsular polysaccharide biosynthesis genes cpsA,
cpsB and cpsD have on virulence of Streptococcus
pneumoniae. J. Infect. Dis. 189, 1905–1913
(2004).140. Bender, M. H., Cartee, R. T. & Yother, J. Positive
correlation between tyrosine phosphorylation of CpsD
and capsular polysaccharide production in
Streptococcus pneumoniae. J. Bacteriol.185,
6057–6066 (2003).
141. Morona, J. K., Morona, R. & Paton, J. C. Attachment
of capsular polysaccharide to the cell wall of
Streptococcus pneumoniae type 2 is required for
invasive disease. Proc. Natl Acad. Sci. USA 103,
8505–8510 (2006).
Demonstrates the involvement of conserved
capsule-biosynthesis genes in the attachment of
polysaccharide to the cell wall and the importanceof this participation for progression from
pneumonia to bacteraemia.
142. Giammarinaro, P. & Paton, J. C. Role of RegM, a
homologue of the catabolite repressor protein CcpA,
in the virulence of Streptococcus pneumoniae. Infect.
Immun. 70, 5454–5461 (2002).
143. Mollerach, M., López, R. & García, E. Characterization
of the galU gene of Streptococcus pneumoniae
encoding a uridine diphosphoglucose
pyrophosphorylase: a gene essential for capsular
polysaccharide biosynthesis. J. Exp. Med . 188,
2047–2056 (1998).
144. Hardy, G. G., Magee, A. D., Ventura, C. L., Caimano,
M. J. & Yother, J. Essential role for cellularphosphoglucomutase in virulence of type 3
Streptococcus pneumoniae. Infect. Immun. 69,
2309–2317 (2001).
145. Douglas, R. M., Paton, J. C., Duncan, S. J. &
Hansman, D. Antibody response to pneumococcal
vaccination in children younger than five years of age.
J. Infect. Dis. l48, 131–137 (l983).146. Paton, J. C. in The Pneumococcus (eds Tuomanen,
E. I., Mitchell, T. J., Morrison, D. A. & Spratt, B. G.)
382–402 (ASM, Washington DC, 2004).
147. Huang, S. S. et al. Post-PCV7 changes in colonizing
pneumococcal serotypes in 16 Massachusetts
communities, 2001 and 2004. Pediatrics 116,
408–413 (2005).148. Reingold, A. et al. Direct and indirect effects of routine
vaccination of children with 7-valent pneumococcal
conjugate vaccine on incidence of invasive
pneumococcal disease — United States, 1998–2003.
MMWR 54, 893–897 (2005).
149. Steenhoff, A. P., Shah, S. S., Ratner, A. J., Patil, S. M.
& McGowan, K. L. Emergence of vaccine-related
pneumococcal serotypes as a cause of bacteremia.
Clin. Infect. Dis. 42, 907–914 (2006).
150. Ogunniyi, A. D., Folland, R. L., Hollingshead, S., Briles,
D. E. & Paton, J. C. Immunization of mice with
combinations of pneumococcal virulence proteins
elicits enhanced protection against challenge with
Streptococcus pneumoniae. Infect. Immun. 68,
3028–3033 (2000).
The first paper to demonstrate that immunization
with combinations of pneumococcal virulence
proteins elicits greater levels of protection
against challenge than if the same antigens are
used individually. Thus, combinations of protein
antigens may be capable of eliciting robust
protection against diverse pneumococcal strains.
151. Briles, D. E. et al. Intranasal immunization of mice
with a mixture of the pneumococcal proteins PsaA and
PspA is highly protective against nasopharyngeal
carriage of Streptococcus pneumoniae. Infect. Immun.
68, 796–800 (2000).152. Briles, D. E. et al. Immunizations with pneumococcal
surface protein A and pneumolysin are protective
against pneumonia in a murine model of pulmonary
infection with Streptococcus pneumoniae. J. Infect.
Dis. 188, 339–348 (2003).
153. Ogunniyi, A. D., Grabowicz, M., Briles, D. E., Cook, J.
& Paton, J. C. Development of a vaccine against
invasive pneumococcal disease based on combinations
of virulence proteins of Streptococcus pneumoniae.
Infect. Immun. 75, 350–357 (2007).
154. Redd, S. C. et al. The role of human immunodeficiency
virus infection in pneumococcal bacteremia in San
Francisco residents. J. Infect. Dis . 162, 1012–1017
(1990).155. Rodriguez-Barradas, M. C. et al. Colonisation by
Streptococcus pneumoniae among human
immunodeficiency virus-infected adults prevalence
of antibiotic resistance, impact of immunization, and
characterization by polymerase chain reaction with
BOX primers of isolates from persistent
S. pneumoniae carriers. J. Infect. Dis. 175,
590–597 (1997).
156. Kemp, K., Bruunsgaard, H., Skinhoj, P. & Klarlund
Pedersen, B. Pneumococcal infections in humans are
associated with increased apoptosis and trafficking of
type 1 cytokine-producing T cells. Infect. Immun. 70,
5019–5025 (2002).
157. Zhang, Q. et al. Low CD4 T cell immunity to
pneumolysin is associated with nasopharyngeal
carriage of pneumococci in children. J. Infect. Dis.
195, 1194–1202 (2007).
158. Zhang, Q. et al. Regulation of production of mucosal
antibody to pneumococcal protein antigens by
T-cell-derived gamma interferon and interleukin-10
in children. Infect. Immun. 74, 4735–4743 (2006).
DATABASESEntrez Gene: http://www.ncbi.nlm.nih.gov/entrez/query.
fcgi?db=gene
comA | comD | comE | cpsB | galU | nanA | nanB | pgm | psaA |
psaB | psaC | pspA | pspC
Entrez Genome Project: http://www.ncbi.nlm.nih.gov/
entrez/query.fcgi?db=genomeprj
Actinobacillus actinomycetemcomitans | Haemophilus
influenzae | Streptococcus pneumoniae
Entrez Protein: http://www.ncbi.nlm.nih.gov/entrez/query.
fcgi?db=protein
BgaA | CcpA | CpsC | CpsD | CRP | LytA | PiaA | StrH
FURTHER INFORMATIONAras Kadioglu’s homepage: http://www.le.ac.uk/iii/staff/
ak13.html
Jeffrey N. Weiser’s homepage: http://www.med.upenn.edu/
micro/faculty/weiser.html
All liNkS Are Active iN the oNliNe PDf
RE V I E W S