portal vein branch occlusion induces cell proliferation of cholestatic rat liver
TRANSCRIPT

JOURNAL OF SURGICAL RESEARCH 60, 249–257 (1996)Article No. 0039
Portal Vein Branch Occlusion Induces Cell Proliferationof Cholestatic Rat Liver
SHIN-ICHI MIZUNO,* YUJI NIMURA,* HIDEAKI SUZUKI,* AND SHONEN YOSHIDA†,1
*First Department of Surgery and †Laboratory of Cancer Cell Biology, Research Institute for Disease Mechanism and Control,Nagoya University School of Medicine, Nagoya 466, Japan
Submitted for publication August 16, 1994
have facilitated the development of aggressive liver re-Portal vein branch occlusion induces atrophy of oc- section for biliary tract carcinoma [1–3]. Postoperative
cluded hepatic lobes, concomitantly associated with liver failure is one of the most serious complicationsthe complementary hypertrophy of unoccluded lobes. following extended hepatectomy [4, 5]. To reduce theIn this context, selective embolization of the portal risk of posthepatectomy liver failure, preoperative em-vein branch supplying the area to be resected has been bolization of the portal vein branches feeding the areaperformed prior to extended hepatectomy to reduce to be resected has been introduced [6–9]. Portal veinthe risk of postoperative liver failure. However, carci- branch embolization is a modification of portal veinnoma of the hepatic hilus is often associated with ob- branch ligation which had been initiated as a palliativestructive jaundice. At present it is still obscure therapy of hepatic malignant tumors [10, 11]. It is wellwhether cholestatic hepatocytes can also respond well
established that interruption of the portal blood supplyto the proliferation stimuli caused by the portal veinresults in atrophy of the hepatic part involved (occludedbranch embolization. To clarify this point, we made alobes), with compensatory hypertrophy of the hepaticrat model of portal vein branch ligation in combina-tissue not deprived of its blood supply (unoccludedtion with cholestasis. As an indicator of proliferation,lobes) [12–20]. The portal vein branch embolizationwe determined the activity of DNA polymerase a andapproach is based on this observation. The size of thethe mitotic index. The results demonstrate that, evenresected portion relative to the remaining liver canin the cholestatic liver, the expression of DNA poly-therefore be expected to decrease both by the atrophymerase a in unoccluded lobes was induced by contra-of the occluded lobes and by the hypertrophy of thelateral portal vein branch ligation. The maximal de-
gree of DNA polymerase a induction in the cholestatic unoccluded lobes. In clinical cases, a high incidence ofliver was similar to that in the noncholestatic liver, obstructive jaundice is associated with carcinoma ofi.e., fivefold above that in the resting liver. The level the hepatic hilus, which might increase the risk of liverof DNA polymerase a activity correlated well with the failure after resection. But there have been very fewmitotic index in the same tissue. Furthermore, the cell basic studies on the efficacy of either preoperative por-proliferation after the portal vein branch occlusion is tal vein branch ligation or embolization of the chole-not suppressed by the preceding external biliary static liver [21].drainage, which had been shown to suppress the liver In order to know whether cholestatic hepatocytes canregeneration after partial hepatectomy. From these respond well to the proliferation stimuli caused by theresults, it is concluded that portal vein branch ligation occlusion of portal vein branch, we made a rat modelinduces liver cell proliferation in unoccluded lobes, of the portal vein branch ligation in combination withirrespective of the presence of cholestasis. q 1996 cholestasis. In this model we ligated the left branch ofAcademic Press, Inc.
the portal vein feeding the left lateral and medianlobes. As an indicator of proliferation capacity of livercells, we routinely assayed DNA polymerase a, whichINTRODUCTIONplays a central role in eukaryotic DNA replication. Pre-viously we have shown that the induction pattern ofTechnical progress in hepatic surgery and enhancedDNA polymerase a corresponded well with DNA syn-anatomical data regarding the liver and hepatic hilusthesis measured by the incorporation of [3H]thymidineinto chromosomal DNA in normal human fibloblast [22]
1 To whom correspondence and reprint requests should be ad- as well as regenerating rat liver [23, 24]. In the presentdressed at Laboratory of Cancer Cell Biology, Research Institute study, we evaluated the proliferative capacity of thefor Disease Mechanism and Control, Nagoya University School of
cholestatic rat liver after portal vein branch ligationMedicine, 65 Tsurumai-cho, Showa-ku, Nagoya 466, Japan. Fax: 81-52-741-2596. by monitoring the induction of DNA polymerase a as
249 0022-4804/96 $12.00Copyright q 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

250 JOURNAL OF SURGICAL RESEARCH: VOL. 60, NO. 1, JANUARY 1996
well as the increase in the mitotic index. It was foundthat cholestasis did not suppress the proliferation in-duced by portal vein branch ligation. Furthermore, thisproliferation is not suppressed by preceding externalbiliary drainage, which has been shown to suppressstrongly the liver regeneration after partial hepatec-tomy [24].
MATERIALS AND METHODS
AnimalsSix-week-old adult male rats (200–250 g) of the Donryu strain
were purchased from Chubu Shiryo (Nagoya, Japan). After mainte-nance in an air-conditioned room for 1 week, the operation was per-formed. The animals were fed and watered ad libitum. All surgicalprocedures were performed under light ether anesthesia between 9and 12 A.M. to minimize the influence of diurnal rhythm on thesubsequent liver cell proliferation. Experimental animals were sacri-ficed under light ether anesthesia. The livers were then removedimmediately, weighed, and stored at 0807C until used. The opera-tions performed for each experiment were as follows.
Experiment 1. The animals were divided into two groups. In theportal vein branch ligation group, the left branch of the portal veinfeeding the left lateral and median lobes (70.0 { 1.6% of the totalliver weight) was ligated following the procedure of Steiner and Mar-tinez [15]. In the sham group, rats underwent laparotomy and theliver was manipulated just as in the portal vein branch ligation groupexcept that the portal vein branch was not ligated. In both groups,five or more animals were sacrificed on Days 0, 1, 2, 3, 5, 7, and 14after the operation. In addition, in the portal vein branch ligationgroup, six animals were sacrificed at 12 and 36 hr after the portalvein branch ligation (Fig. 1).
Experiment 2. To examine the effect of portal vein branch ligationon the cholestatic liver, we induced cholestasis by ligation of thecommon bile duct according to the method of Weinbren [25] withsome modifications [23]. Rats were divided into three groups. In thecommon bile duct ligation group, six animals were sacrificed on Days2, 4, and 5 after bile duct ligation. In the portal vein branch ligationgroup and sham group, rats were relaparotomized on the 5th day FIG. 1. Protocol of experiments. Nonitalic numerals, days afterafter common bile duct ligation and the portal vein branch ligation the bile duct obstruction; italic numerals, days after the portal veinor the sham operation was performed as described above for Experi- branch ligation (PBL) or sham operation; outlined numerals, daysment 1. Five or more animals were sacrificed on Days 1, 2, 3, 5, and after biliary decompression. Solid arrow, portal vein branch ligation7 after the portal vein branch ligation or the sham operation in both or sham operation; open arrow, common bile duct ligation (CBDL);groups (Fig. 1). hatched arrow, production of animal models of external biliary drain-
Experiment 3. We produced an animal model of external biliary age; , sacrifice of five or more animals; À, biliary obstruction period;drainage by the procedure of Iyomasa et al. [24]. A polyethylene tube “, external biliary drainage period.with an outer diameter of 0.9 mm (Portex, Hythe, Kent, UK) wasinserted into the common bile duct by the cut-down technique [24]and the other end of the tube was exposed and fixed at the nape
7.5) containing 10% glycerol, 1 mM dithiothreitol (DTT) (Nakaraithrough the subcutaneous route for protection against bites [24].Tesque, Kyoto, Japan), 1 mM phenylmethylsulfonyl fluoride (SigmaCholestasis and biliary decompression were induced by ligation orChemical Co., St. Louis, MO), 0.5% polyethylene(10)octylphenyletherrelease, respectively, of the end of the tube. Cholestasis was induced(Triton X-100), 0.1 mM EDTA, and 0.5 M KCl. A half volume ofby ligation of the end of the drain tube for 5 days. The animals wereeach sample was used for measuring the DNA content, while thedivided into three groups. In group A, the portal vein branch ligationremainder was clarified by centrifugation at 18,000g for 30 min ator the sham operation was performed simultaneous with biliary de-47C. The supernatant was then collected carefully and used to assaycompression. In group B, the portal vein branch ligation or shamDNA polymerase activity. For determining DNA polymerase a activ-operation was performed 3 days after decompression of the biliaryity, the supernatant was diluted 10-fold with 25 mM Tris–HCl (pHtract. In both groups, six or seven animals were sacrificed 48 hr after7.5) containing 1 mg/ml of bovine serum albumin (BSA), 1 mM DTT,the portal vein branch ligation or the sham operation. In group C,and 10% glycerol. For determining DNA polymerase b activity, theonly external biliary drainage was carried out on the 5th day aftersupernatant was diluted 100-fold with the same solution used forbile duct ligation and five or six animals were sacrificed on Days 0,the DNA polymerase a assay.1, and 3 after external biliary drainage (Fig. 1).
All animals received humane care in compliance with the Guide-Assays of DNA Polymeraseslines for Animal Experiments established at Nagoya University.
The reaction mixture for DNA polymerase a (50 ml) contained 80Enzyme ExtractionmM potassium phosphate (pH 7.2), 8 mM 2-mercaptoethanol, 200mg/ml activated calf thymus DNA, 80 mM each of dATP, dGTP, andOne gram of liver was homogenized with a Dounce homogenizer
and then sonicated for 30 sec in 9 volumes of 50 mM Tris–HCl (pH dCTP, 40 mM [3H]dTTP (1000 cpm/pmol), 8 mM MgCl2, 200 mg/ml
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

251MIZUNO ET AL.: CELL PROLIFERATION AND PORTAL VEIN OCCLUSION
cluded lobes to the whole liver are shown. The ratiowas approximately 30% at the time of the portal veinbranch ligation procedure, either in normal (Experi-ment 1) (30 { 2%) or cholestatic liver (Experiment 2)(29 { 2%). After portal vein branch ligation, the ratioincreased promptly. On Day 7, the ratio in the normalliver increased to 81 { 1%, which was slightly higherthan that in cholestatic liver (66 { 4%). After the shamoperation, the ratio showed no significant change ineither group.
DNA Content
In Experiment 2, 5 days of biliary obstruction in-duced a 20–25% increase in the DNA content of livers(Day 0 in Figs. 3A and 3B), in agreement with ourFIG. 2. Increase in wet weight of the unoccluded lobes of theprevious observations [23, 24]. After the portal veinliver after portal vein branch ligation. Each point represents the
mean{ SD for 5–10 animals. **Põ 0.01 vs sham. —s—, the portal branch ligation, the DNA content of the unoccludedvein branch ligation (PBL) group from Experiment 1; ---s- - -, the lobes increased linearly until Day 5 and thereafter,sham group from Experiment 1; —l—, the portal vein branch liga- when it reached a plateau which was 3.5 times greatertion (PBL) group from Experiment 2; ---l- - -, the sham group from
than that in the corresponding lobes of the resting liverExperiment 2.in both Experiments 1 and 2 (Fig. 3A). On the otherhand, the DNA content of the occluded lobes decreasedBSA, and enzyme. The reaction mixture for DNA polymerase b (50by more than 50% in both Experiments 1 and 2 (Fig.ml) contained 0.1 M Tris–HCl (pH 8.8), 5 mg of (dA)n/(dT)12–18 (base3B). However, on Days 3 to 7, the DNA content of theratio of A/T Å 5), 80 mM [3H]dTTP (1000 cpm/pmol), 0.1 M NaCl, 4
mM N-ethylmaleimide, 0.5 mM MnCl2, and 0.2 mg/ml BSA. After occluded lobes in cholestatic livers was significantlyincubation for 30 min at 377C, the reaction was terminated by theaddition of 0.5 ml of cold 10% trichloroacetic acid containing 0.2%sodium pyrophosphate and acid-insoluble radioactivity was mea-sured as described previously [26, 27].
Measurement of DNA ContentDNA content was measured by fluorescence assay, using a Hoechst
33258 according to the method of Labarca and Paigen [28].
Mitotic IndexA portion of the median lobe and the right lateral lobe of each
sample was fixed in formalin and stained with hematoxylin–eosin.On each slide, 5000–20,000 hepatocytes were examined by light mi-croscopy at 4001 magnification and the number of hepatocytes withmitotic figures was counted [23].
Measurement of Bilirubin and Enzymes in BloodBlood was withdrawn from the vena cava of rats when sacrificed.
Serum was obtained by centrifugation (3000 rpm, 20 min). Totalbilirubin concentration and the activities of glutamic oxaloacetictransaminase (GOT) and glutamic pyruvic transaminase (GPT) weremeasured as described [29–31].
StatisticsThe data were expressed as the mean{ standard deviation. Statis-
tical analyses were performed using Student’s t test. Differences witha P value of less than 0.05 were considered statistically significant.
RESULTS
Wet Weight of the Liver
After portal vein branch ligation, the wet weights ofthe occluded lobes of the liver decreased, while theweight of the unoccluded lobes increased both in the
FIG. 3. Changes in DNA contents of the unoccluded lobes and thenormal and in the jaundiced rats. The total liveroccluded lobes of livers after portal vein branch ligation. Each pointweights in the portal vein branch ligation group were represents the mean { SD for 5–10 animals. (A) Unoccluded lobes.
similar to those of the sham-operated group (data not (B) Occluded lobes. s, Experiment 1; l, Experiment 2; h, Experiment1; j, Experiment 2. *P õ 0.05; **P õ 0.01 vs Experiment 1.shown). In Fig. 2, the ratios of the wet weight of unoc-
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

252 JOURNAL OF SURGICAL RESEARCH: VOL. 60, NO. 1, JANUARY 1996
higher than that in the normal livers (Fig. 3B). Thisfinding might reflect the fact that obstructive jaundicealone induces proliferation of hepatocytes [23].
GOT and GPT Activities and Total Bilirubin Contentin Blood
As shown in Fig. 4A, the level of serum GOT activityrapidly increased after portal vein branch ligation andthen gradually decreased. After Day 5, there was nosignificant difference in serum GOT activity betweenthe portal vein branch ligation and sham groups. Withbiliary obstruction, the serum GOT activity increasedmoderately, increasing further after the portal veinbranch ligation (Fig. 4B). Thereafter, it decreased andreached nearly the same level of that of the sham opera-tion group at Day 7 after portal vein branch ligation(Fig. 4B). Serum GPT showed a pattern similar to serumGOT in both Experiments 1 and 2 (data not shown).
After ligation of the portal vein branch of the nonchole-static liver, the concentration of serum total bilirubinremained low, less than 1.0 mg/dl throughout the experi-mental period (data not shown). On the other hand, theserum total bilirubin concentration exceeded 7 mg/dlafter 5 days’ biliary obstruction and gradually increasedthereafter. This high concentration of bilirubin was notsignificantly affected by the additional portal vein branchligation compared with the sham operation (Fig. 4C).
Activities of DNA Polymerase a
DNA polymerase a activity was measured as an indi-cator of cell proliferation. As shown in Fig. 5A, DNApolymerase a activity in the unoccluded lobes startedto increase on Day 1 after the portal vein branch liga-tion and reached the maximal level on Day 2, thereafterdecreasing. The maximal level was almost fivefoldabove that of the sham-operated control. The DNApolymerase a activity in the sham-operated controlliver and in the occluded lobes of the portal vein branchligation group was nearly identical to that in the rest-ing liver throughout the experimental period.
By biliary obstruction alone, DNA polymerase a ac-tivity increased approximately twofold above that inthe resting liver (Fig. 5C) in agreement with our previ-ous observations [23, 24]. Due to portal vein branchligation, the activity in the unoccluded lobes started to FIG. 4. Changes in serum GOT and total bilirubin concentra-
tion. (A) Serum GOT in Experiment 1. (B) Serum GOT in Experi-increase on Day 1, reached the maximal level on Dayment 2. (C) Total bilirubin in Experiment 2. Each point represents2, and then decreased thereafter (Fig. 5C). On Day 7the mean { SD for 5–10 animals. *P õ 0.05; **P õ 0.01 vs sham.after the second operation, no significant difference wass, the portal vein branch ligation (PBL) group; l, the sham-oper-
observed between the portal vein branch ligation and ated control group; n, the common bile duct ligation group fromthe sham-operated groups (Fig. 5C). The peak level of Experiment 2; m, nonoperated animals; solid arrow, portal vein
branch ligation or sham operation; open arrow, common bile ductDNA polymerase a activity in the unoccluded lobes inligation.Experiment 2 (47.9 { 7.3 U/mg DNA) was almost com-
parable to that in Experiment 1 (54.1{ 5.6 U/mg DNA).In the occluded lobes, the levels of DNA polymerase a Activity of DNA Polymerase bactivity were the same or lower than that in sham-operated controls, but significantly higher than that in As shown in Fig. 5B, DNA polymerase b activity in
the unoccluded lobes also increased after PBL but in aresting livers (Fig. 5C).
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res
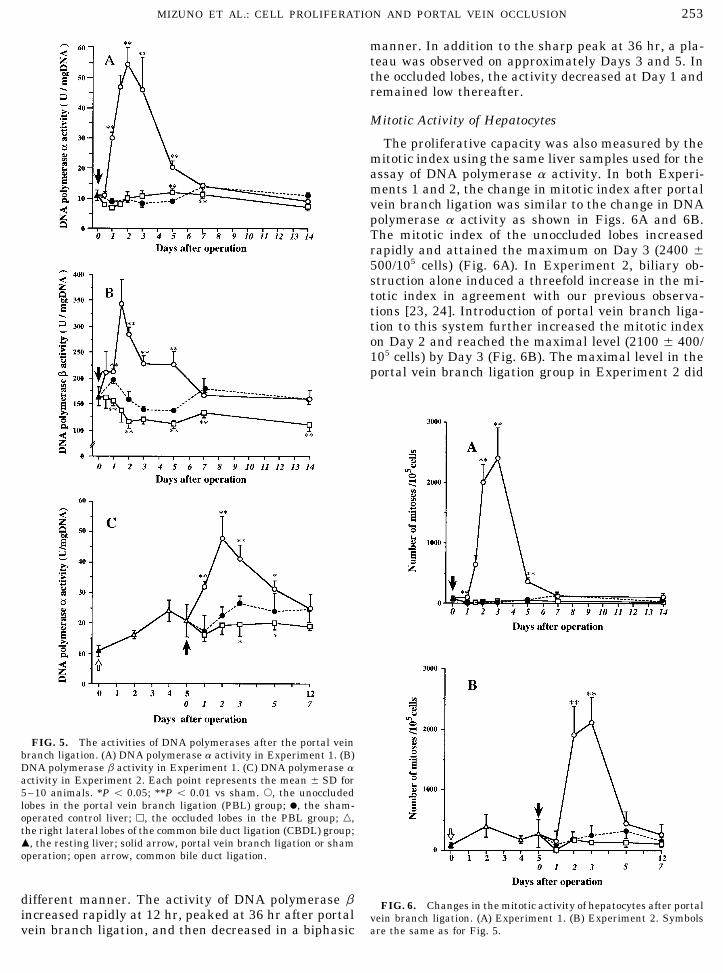
253MIZUNO ET AL.: CELL PROLIFERATION AND PORTAL VEIN OCCLUSION
manner. In addition to the sharp peak at 36 hr, a pla-teau was observed on approximately Days 3 and 5. Inthe occluded lobes, the activity decreased at Day 1 andremained low thereafter.
Mitotic Activity of Hepatocytes
The proliferative capacity was also measured by themitotic index using the same liver samples used for theassay of DNA polymerase a activity. In both Experi-ments 1 and 2, the change in mitotic index after portalvein branch ligation was similar to the change in DNApolymerase a activity as shown in Figs. 6A and 6B.The mitotic index of the unoccluded lobes increasedrapidly and attained the maximum on Day 3 (2400 {500/105 cells) (Fig. 6A). In Experiment 2, biliary ob-struction alone induced a threefold increase in the mi-totic index in agreement with our previous observa-tions [23, 24]. Introduction of portal vein branch liga-tion to this system further increased the mitotic indexon Day 2 and reached the maximal level (2100 { 400/105 cells) by Day 3 (Fig. 6B). The maximal level in theportal vein branch ligation group in Experiment 2 did
FIG. 5. The activities of DNA polymerases after the portal veinbranch ligation. (A) DNA polymerase a activity in Experiment 1. (B)DNA polymerase b activity in Experiment 1. (C) DNA polymerase aactivity in Experiment 2. Each point represents the mean { SD for5–10 animals. *P õ 0.05; **P õ 0.01 vs sham. s, the unoccludedlobes in the portal vein branch ligation (PBL) group; l, the sham-operated control liver; h, the occluded lobes in the PBL group; n,the right lateral lobes of the common bile duct ligation (CBDL) group;m, the resting liver; solid arrow, portal vein branch ligation or shamoperation; open arrow, common bile duct ligation.
different manner. The activity of DNA polymerase b FIG. 6. Changes in the mitotic activity of hepatocytes after portalincreased rapidly at 12 hr, peaked at 36 hr after portal vein branch ligation. (A) Experiment 1. (B) Experiment 2. Symbols
are the same as for Fig. 5.vein branch ligation, and then decreased in a biphasic
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

254 JOURNAL OF SURGICAL RESEARCH: VOL. 60, NO. 1, JANUARY 1996
polymerase a activity in the cholestatic liver decreasedrapidly due to external biliary drainage even on Day 1after decompression, in agreement with our previousfindings [24]. On Day 3 after biliary decompression, theactivity decreased to 80% of that in the resting liver(Fig. 7A). When the portal vein branch ligation wasperformed either on Day 3 after decompression or si-multaneous with decompression (groups A and B ofExperiment 3 of Fig. 1), the activity in the unoccludedlobes increased markedly and there was no significantdifference between groups A and B in the level of in-duced DNA polymerase a (Fig. 7A), which was compa-rable to the peak level in Experiment 1.
The mitotic index in the unoccluded lobes also in-creased similar to DNA polymerase a although the mi-totic index induction exceeded that of DNA polymerasea (Fig. 7B). The mitotic index 2 days after the portalvein branch ligation in group A was comparable to thatin group B (Fig. 7B).
Cholestasis and biliary decompression were moni-tored by blood total bilirubin. In group A, the bloodtotal bilirubin concentration decreased from 7.4 { 0.8mg/dl to less than 1.0 mg/dl 2 days after either theportal vein branch ligation or the sham operation. Ingroup B, the blood total bilirubin was 0.4 { 0.1 mg/dland 0.3 { 0.1 mg/dl 2 days after the portal vein branchligation and the sham operation, respectively. The dif-ference was not statistically significant.
DISCUSSION
In the present study, it was clearly demonstratedthat portal vein branch ligation markedly increased theproliferative capacity of hepatocytes in the unoccluded
FIG. 7. Changes in the activity of DNA polymerase a and the lobes of the cholestatic rat liver as well as the nonchole-mitotic index in Experiment 3. (A) Activity of DNA polymerase a.static one. Previously we have evaluated the prolifera-(B) Mitotic activity of hepatocytes. Each point represents the mean
{ SD for 6 or 7 animals. **P õ 0.01 vs sham. s, the unoccluded tive capacity of the rat liver after partial hepatectomylobes after portal vein branch ligation in group A; l, the sham- and bile duct obstruction by monitoring DNA polymer-operated control liver in group A; h, the unoccluded lobes after portal ase a activity, mitotic index, and [3H]thymidine incor-vein branch ligation in group B; j, the sham-operated control liver
poration [23, 24]. The pattern of DNA polymerase ain group B; n, the right lateral lobes in group C; m, the resting liver;induction corresponded well with DNA synthesis ashatched arrow, insertion point of the external drainage tube and
ligation of it; solid arrow, portal vein branch ligation or sham opera- measured by incorporation of [3H]thymidine in compar-tion; À, the interval of the biliary obstruction. Italic numerals, days ison with elevation of the mitotic index [23, 24]. Inafter portal vein branch ligation or sham operation; outlined numer- Experiment 1 of this study we showed that DNA poly-als, days after biliary decompression. Note that the vertical axis in
merase awas also a reliable marker of cell proliferativeFig. 7B was logarithmically plotted.capacity of the liver after portal vein branch ligation(Figs. 5A and 6A). It should be noted, however, thatthe assay conditions for DNA polymerase a in this
not differ significantly from that in Experiment 1 (Figs. study might also detect polymerase d and e. But our6A and 6B). recent study showed that DNA polymerase activity de-
tected with activated calf thymus DNA (see MaterialsEffect of Biliary Decompression and Methods) in crude extract of rat liver was morethan 80% due to DNA polymerase a (data not shown).After 5 days of biliary obstruction, there was no sig-
nificant difference in DNA polymerase a activity be- Therefore, we tentatively designated the incorporationas DNA polymerase a activity.tween the livers in the common bile duct ligation model
(Experiment 2, Fig. 5C) and the external biliary drain- It is generally accepted that DNA polymerase bmight act in the DNA repair process. So we hypothe-age tube ligation model (Experiment 3, Fig. 7A). The
level of enzyme activity was about threefold above that sized that DNA polymerase b activity in the occludedlobes increased to repair the damage of DNA after por-in the resting liver. As shown in Fig. 7A, the high DNA
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

255MIZUNO ET AL.: CELL PROLIFERATION AND PORTAL VEIN OCCLUSION
tal vein branch ligation. Contrary to our expectations, major factors which supports hepatocyte growth [36–40], the different local concentrations of hepatocytehowever, DNA polymerase b activity did not increase
in the occluded lobes after portal vein branch ligation growth factor might affect the time of proliferation,generated by a paracrine of hepatocyte growth factorbut increased significantly in the unoccluded lobes. The
role of this enzyme has not been fully established and arising from neighboring nonparenchymal cells afterfocal damage to liver produced by portal vein branchsome investigators have proposed that DNA polymer-
ase b also plays a role in DNA replication [32–34]. The ligation or an endocrine of hepatocyte growth factorfrom extrahepatic organs such as the lung and kidneyinduction of enzyme activity in the unoccluded lobes
after portal vein branch ligation might be related to in the case of partial hepatectomy.It was remarkable that the proliferative capacity inDNA replication in portal vein branch ligation-induced
liver cell proliferation. the unoccluded lobes after portal vein branch ligationwas not suppressed by external biliary drainage (Fig.In this study we demonstrated that in the cholestatic
liver the activity of DNA polymerase a was increased 7). This result was entirely different from our previousfindings [24] that the regenerative capacity of the rem-by portal vein branch ligation about fivefold above that
in the resting liver. There was no significant difference nant liver after partial hepatectomy was strongly sup-pressed by external biliary drainage. Recently we alsoin the maximal level of DNA polymerase a between
normal (Experiment 1) and cholestatic liver (Experi- demonstrated that internal biliary drainage, unlike ex-ternal drainage, does not suppress the regeneration ofment 2) in the unoccluded lobes after portal vein
branch ligation. The mitotic index pattern was also rat liver after partial hepatectomy [41]. Therefore, theexistence of bile seems to be essential for liver regener-similar between normal and cholestatic liver. Under
several conditions, DNA polymerase a activity in the ation after partial hepatectomy, while bile is not re-quired for proliferation in the case of portal vein branchunoccluded lobes increased and attained a maximal
level at Day 2 after portal vein branch ligation. The ligation. These results indicate that the partial hepa-tectomy and portal vein branch ligation stimulate livermaximal activity of the enzyme was always about five-
fold higher than that in the resting liver. So it appears cells by different mechanisms, the former requiring anadditional substance as an auxiliary factor, but the con-that this enzyme level represents the upper limit of
hepatocyte proliferative capacity as induced by various clusion must await further studies.Initially portal vein branch ligation was performedstimuli of proliferation.
The proliferative capacity in the unoccluded lobes as a palliative operation for hepatic malignancy to in-duce atrophy of the occluded lobes carrying tumor(s)after portal vein branch ligation has been compared
with the regenerative capacity in remnant liver tissue [10, 11]. Progress in interventional radiological tech-niques enables us to perform portal vein branch emboli-after partial hepatectomy [14, 16, 17]. By comparing
the weight of the liver, partial hepatectomy stimulates zation without laparotomy as a modification of portalvein branch ligation [6, 9]. Recently we have performedregeneration more strongly than does portal vein
branch ligation [14]. By comparing the mitotic index, preoperative percutaneous transhepatic portal veinbranch embolization to reduce the risk of posthepatec-however, ligation of the portal vein branch which was
feeding 70% of the liver induced greater proliferation tomy liver failure in high-risk patients [5, 9], especiallypatients with biliary tract carcinoma that frequentlythan did 70% partial hepatectomy [16]. In our previous
work in which DNA polymerase a activity was used as develop obstructive jaundice. Since the safety and effi-cacy of the embolization of the portal vein branch forthe indicator, the proliferative capacity in the remnant
liver reached the maximal level at 24–28 hr after 70% the cholestatic liver are unknown, we generally post-pone the embolization until the serum total bilirubinpartial hepatectomy [24]. This agrees well with results
by Bucher and Swaffield [35] as determined by [14C]- concentration becomes normal after percutaneoustranshepatic biliary drainage. Here we demonstratedthymidine incorporation. The present study clearly in-
dicated that the portal vein branch ligation drastically that portal vein branch ligation could induce a cellularproliferation in the unoccluded lobes of the cholestaticincreased DNA polymerase a activity in the unoccluded
lobes, reaching the maximum at 48 hr after the ligation liver to a similar extent as in the noncholestatic liver.In regard to the proliferation capacity, therefore, embo-(Fig. 5A). This finding corresponded well with Taka-
san’s observation that the unoccluded lobes of rabbit lization of branches of the portal vein might be feasiblefor patients with obstructive jaundice. But many inves-liver showed the highest incorporation of [3H]-
thymidine at 48 hr after portal vein branch ligation tigators have reported that cholestasis inhibits themetabolic functions of liver cells, including mitochon-[19]. At present, the reason for the delay in response to
portal vein branch ligation in comparison with partial drial function [42, 43], which slowly recovers followingrelief of biliary obstruction. Moreover, it is well knownhepatectomy is unclear. Recently, Hamanoue et al. [36]
demonstrated that portal vein branch ligation induced that many organs other than the liver suffer functionaldamage during severe jaundice [44–46]. From a clini-hepatocyte growth factor activity in sera and mRNA of
hepatocyte growth factor in the occluded lobes with cal point of view, Nagino et al. [5, 47] emphasized thatpercutaneous transhepatic biliary drainage was effica-kinetics that differed from that after partial hepatec-
tomy. Since hepatocyte growth factor may be one of the cious for the prevention of cholangitis, which was one
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

256 JOURNAL OF SURGICAL RESEARCH: VOL. 60, NO. 1, JANUARY 1996
12. Rous, P., and Larimore, L. D. Relation of the portal blood toof the major risk factors for liver failure after hepatec-liver maintenance. J. Exp. Med. 31: 609, 1920.tomy. These results indicate that biliary drainage may
13. Weinbren, K. The portal blood supply and regeneration of thebe required to restore liver function. Since the cell pro-rat liver. Br. J. Exp. Pathol. 36: 583, 1955.liferation induced by portal vein branch ligation is re-
14. Lawrence, W., Joly, D., and Brasfield, R. A comparative studyfractory to cholestasis and to the preceding external of various mechanisms of hepatic restoration in the rat. Surgerybiliary drainage, it is suggested that preoperative per- 45: 543, 1959.cutaneous transhepatic portal vein branch emboliza- 15. Steiner, P. E., and Martinez, J. B. Effects on the rat liver of
bile duct, portal vein and hepatic artery ligations. Am. J. Pathol.tion can be performed shortly after percutaneous trans-39: 257, 1961.hepatic biliary drainage, even if serum bilirubin is still
16. Weinbren, K., and Tarsh, E. The mitotic response in the rathigher than the normal level. Although other parame-liver after different regenerative stimuli. Br. J. Exp. Pathol. 45:ters of liver functions must be taken into consideration475, 1964.
for the safety of each patient, the early embolization of17. Lieberman, I., and Short, J. Hepatic blood supply and controlthe portal vein branch might benefit icteric patients of deoxyribonucleic acid synthesis in liver. Am. J. Physiol. 208:
with carcinoma of the hepatic hilus to curtail the preop- 896, 1965.erative preparation period for extended liver resection. 18. Ozawa, K., Takasan, H., Kitamura, O., Mizukami, T., Kamano,
T., Takeda, H., Ohsawa, T., Murata, T., and Honjo, I. Effect ofligation of portal vein on liver mitochondrial metabolism. J.
ACKNOWLEDGMENTS Biochem. 70: 755, 1971.19. Takasan, H., Mizukami, T., Yamada, T., Ozawa, K., and Honjo,
This work was supported in part by a Grant-in-Aid from the Ministry I. Morphological correlates of functional enhancement of liverof Education, Science and Culture of Japan and by the Uehara Memo- mitochondria nourished with excess portal blood. J. Electronrial Life Science Foundation. Microsc. 24: 235, 1975.
20. Katoh, T., Tanaka, M., Nimura, Y., Kanai, M., Nagino, M., andOzawa, T. Enhancement of rat liver mitochondrial function byREFERENCES portal branch ligation secures subsequent extended hepatec-tomy. Biochem. Int. 24: 107, 1991.
1. Nimura, Y., Hayakawa, N., Kamiya, J., Kondo, S., and Shio- 21. Ishikawa, M., Yogita, S., Komi, N., and Makuuchi, M. Experi-noya, S. Hepatic segmentectomy with caudate lobe resection for mental studies on liver regeneration following transcatheterbile duct carcinoma of the hepatic hilus. World J. Surg. 14: 535, portal embolization in obstructive jaundice (in Japanese). Jpn.1990. J. Gastroenterol. Surg. 22: 2615, 1989.
2. Nimura, Y., Hayakawa, N., Kamiya, J., Maeda, S., Kondo, S., 22. Nakamura, H., Morita, T., Masaki, S., and Yoshida, S. Intracel-Yasui, A., and Shionoya, S. Hepatopancreatoduodenectomy for lular localization and metabolism of DNA polymerase a in hu-advanced carcinoma of the biliary tract. Hepatogastroenterology man cells visualized with monoclonal antibody. Exp. Cell Res.38: 170, 1991. 151: 123, 1984.
3. Nimura, Y., Hayakawa, N., Kamiya, J., Maeda, S., Kondo, S., 23. Terasaki, M., Kuriki, H., Nimura, Y., Shionoya, S., Kojima, K.,Yasui, A., and Shionoya, S. Combined portal vein and liver and Yoshida, S. Induction of DNA replication and cell growthresection for carcinoma of the biliary tract. Br. J. Surg. 78: 727, in rat liver by obstructive jaundice. Jpn. J. Cancer Res. 82: 170,1991. 1991.
4. Bengmark, S., Ekberg, H., Evander, A., Klofver-stahl, B., and 24. Iyomasa, S., Terasaki, M., Kuriki, H., Nimura, Y., Shionoya, S.,Tranberg, K. G. Major liver resection for hilar cholangiocarci- Kojima, K., and Yoshida, S. Decrease in regeneration capacity ofnoma. Ann. Surg. 207: 120, 1988. rat liver after external biliary drainage. Eur. Surg. Res. 24:
265, 1992.5. Nagino, M., Nimura, Y., Hayakawa, N., Kamiya, J., Kondo, S.,Sasaki, R., and Hamajima, N. Logistic regression and discrimi- 25. Weinbren, K. The effect of bile duct obstruction on regenerationnant analyses of hepatic failure after liver resection for carci- of the rat’s liver. Br. J. Exp. Pathol. 34: 280, 1953.noma of the biliary tract. World J. Surg. 17: 250, 1993. 26. Yoshida, S., Modak, M. J., and Yagi, K. DNA polymerase associ-
6. Kinoshita, H., Sakai, K., Hirohashi, K., Igawa, S., Yamasaki, ated with nuclear membrane of calf thymus. Biochem. Biophys.O., and Kubo, S. Preoperative portal vein embolization for hepa- Res. Commun. 43: 1408, 1971.tocellular carcinoma. World J. Surg. 10: 803, 1986. 27. Yoshida, S., Kondo, T., and Ando, T. Multiple molecular species
7. Makuuchi, M., Thai, B. L., Takayasu, K., Takayama, T., Ko- of cytoplasmic DNA polymerase from calf thymus. Biochim. Bio-suge, T., Gunven, P., Yamazaki, S., Hasegawa, H., and Ozaki, phys. Acta 353: 463, 1974.H. Preoperative portal embolization to increase safety of major 28. Labarca, C., and Paigen, K. A simple, rapid, and sensitive DNAhepatectomy for hilar bile duct carcinoma: A preliminary report. assay procedure. Anal. Biochem. 102: 344, 1980.Surgery 107: 521, 1990. 29. Michaelsson, M., Nosslin, B., and Sjolin, S. Plasma bilirubin
8. Makuuchi, M., Kosuge, T., and Lygidakis, N. J. New possibilit- determination in the newborn infant: A methodological studyies for major liver surgery in patients with Klatskin tumors or with special reference to the influence of hemolysis. Pediatricsprimary hepatocellular carcinoma—An old problem revisited. 35: 925, 1965.Hepatogastroenterology 38: 329, 1991. 30. Wroblewski, F., and LaDue, J. S. Serum glutamic pyruvic trans-
9. Nagino, M., Nimura, Y., and Hayakawa, N. Percutaneous trans- aminase in cardiac and hepatic disease. Proc. Soc. Exp. Biol.hepatic portal embolization using newly devised catheters: Pre- Med. 91: 569, 1956.liminary report. World J. Surg. 17: 520, 1993. 31. Bergmeyer, H. U., Kreutz, F. H., Pilz, W., Schmidt, F. W., But-
10. Kozaka, S. Extensive hepatectomy in two stages an experimen- tner, H., Lang, H., Rick, W., Stamm, D., Hillmann, G., Laue,tal study. Nippon Geka Hokan 32: 99, 1963. D., Schmidt, E., and Szasz, G. Recommendations of the German
Society for Clinical Chemistry. Z. Klin. Chem. Klin. Biochem.11. Honjo, I., Suzuki, T., Ozawa, K., Takasan, H., Kitamura, O.,10: 182, 1972.and Ishikawa, T. Ligation of a branch of the portal vein for
carcinoma of the liver. Am. J. Surg. 130: 296, 1975. 32. Suzuki, H., Menegazzi, M., Prati, A. C. D., Ogura, T., Esumi,
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res

257MIZUNO ET AL.: CELL PROLIFERATION AND PORTAL VEIN OCCLUSION
H., Matsukage, A., and Libonati, M. Induction of DNA polymer- tion of hepatocyte growth factor from injured liver of carbontetrachloride-treated rats. J. Biochem. 109: 8, 1991.ase b during proliferation of mitogen-stimulated human lym-
phocytes. Biochem. Biophys. Res. Commun. 181: 623, 1991. 40. Kinoshita, T., Hirao, S., Matsumoto, K., and Nakamura, T. Pos-sible endocrine control by hepatocyte growth factor of liver re-33. Jenkins, T. M., Saxena, J. K., Kumar, A., Wilson, S. H., andgeneration after partial hepatectomy. Biochem. Biophys. Res.Ackerman, E. J. DNA polymerase b and DNA synthesis in Xeno-Commun. 177: 330, 1991.pus oocytes and in a nuclear extract. Science 258: 475, 1992.
41. Suzuki, H., Iyomasa, S., Nimura, Y., and Yoshida, S. Internal34. Sweasy, J. B., and Loeb, L. A. Mammalian DNA polymerase b biliary drainage, unlike external drainage, does not suppress
can substitute for DNA polymerase I during DNA replication the regeneration of cholestatic rat liver after partial hepatec-in Escherichia coli. J. Biol. Chem. 267: 1407, 1992. tomy. Hepatology 20: 1318, 1994.
35. Bucher, N. L. R., and Swaffield, M. N. The rate of incorporation 42. Koyama, K., Takagi, Y., Ito, K., and Sato, T. Experimental andof labeled thymidine into the deoxyribonucleic acid of regenerat- clinical studies on the effect of biliary drainage in obstructiveing rat liver in relation to the amount of liver excised. Cancer jaundice. Am. J. Surg. 142: 293, 1981.Res. 24: 1611, 1964. 43. Miyata, K. Delayed recovery of mitochondrial function in rat
liver after releasing biliary obstruction. Nagoya J. Med. Sci. 45:36. Hamanoue, M., Kawaida, K., Takao, S., Shimazu, H., Noji, S.,97, 1983.Matsumoto, K., and Nakamura, T. Rapid and marked induction
44. Wait, R. B., and Kahng, K. U. Renal failure complicating ob-of hepatocyte growth factor during liver regeneration after isch-structive jaundice. Am. J. Surg. 157: 256, 1989.emic or crush injury. Hepatology 16: 1485, 1992.
45. Katz, S., Grosfeld, J. L., Gross, K., Plager, D. A., Ross, D., Rosen-37. Nakamura, T., Nawa, K., and Ichihara, A. Partial purificationthal, R. S., Hull, M., and Weber, T. R. Impaired bacterial clear-and characterization of hepatocyte growth factor from serumance and trapping in obstructive jaundice. Ann. Surg. 199: 14,of hepatectomized rats. Biochem. Biophys. Res. Commun. 122:1984.1450, 1984.
46. Green, J., Beyar, R., Sideman, S., Mordechovitz, D., and Better,38. Kinoshita, T., Tashiro, K., and Nakamura, T. Marked increase O. S. The ‘‘jaundiced heart’’: A possible explanation for postoper-
of HGF mRNA in non-parenchymal liver cells of rats treated ative shock in obstructive jaundice. Surgery 100: 14, 1986.with hepatotoxins. Biochem. Biophys. Res. Commun. 165: 1229, 47. Nagino, M., Hayakawa, N., Nimura, Y., Dohke, M., and Kita-1989. gawa, S. Percutaneous transhepatic biliary drainage in patients
39. Asami, O., Ihara, I., Shimidzu, N., Shimizu, S., Tomita, Y., with malignant biliary obstruction of the hepatic confluence.Hepatogastroenterology 39: 296, 1992.Ichihara, A., and Nakamura, T. Purification and characteriza-
/ m4725$4529 12-13-95 12:12:37 srgas AP: Surg Res