potential of autochthonous fungal strains isolated from contaminated soils for degradation of...
TRANSCRIPT

f u n g a l b i o l o g y 1 1 7 ( 2 0 1 3 ) 2 6 8e2 7 4
journa l homepage : www.e lsev ier . com/ loca te / funb io
Potential of autochthonous fungal strains isolated fromcontaminated soils for degradation of polychlorinatedbiphenyls
Bello MOUHAMADOUa,*, Mathieu FAUREa, Lucile SAGEa, Johanna MARCAISa,Florence SOUARDb, Roberto A. GEREMIAa
aLaboratoire d’Ecologie Alpine, UMR 5553 CNRS, Universit�e Joseph Fourier, Grenoble 1, BP 53, 38041 Grenoble Cedex 9, FrancebD�epartement de Pharmacochimie Mol�eculaire, UMR 5063 CNRS, Universit�e Joseph Fourier, Grenoble 1, BP 53,
38041 Grenoble Cedex 9, France
a r t i c l e i n f o
Article history:
Received 20 November 2012
Received in revised form
15 February 2013
Accepted 27 February 2013
Available online 14 March 2013
Corresponding Editor:
John Dighton
Keywords:
Ascomycota
Autochthonous fungal strains
Biodegradation
PCBs
Zygomycota
* Corresponding author. Tel.: þ33 476 63 54 4E-mail address: bello.mouhamadou@ujf-g
1878-6146/$ e see front matter ª 2013 The Bhttp://dx.doi.org/10.1016/j.funbio.2013.02.004
a b s t r a c t
Up to now, most studies on polychlorinated biphenyl (PCB) bioremediation have examined
the ability of model fungal strains to biodegrade PCBs. Yet, there is limited information
concerning the potential of autochthonous filamentous fungal strains in the biodegrada-
tion of PCBs and their possible use in the environmental technologies. In this study, we in-
vestigated the capacity of autochthonous fungal strains in the biodegradation of PCBs by
isolating 24 taxa from former industrial sites highly contaminated by PCBs. Microscopic
and molecular analyses using the internal transcribed spacer (ITS) region revealed that
the fungal strains belonged to the phyla Ascomycota (19 strains) and Zygomycota (five
strains). The chromatography gas analysis revealed evidence of degradation of seven
PCB congeners. With the exception of Circinella muscae which presented no degradation po-
tential, the other fungal strains exhibited a rate of biodegradation ranging from 29 to 85 %
after 7 d of incubation in liquid medium. Among these strains, Doratomyces nanus, Dorato-
myces purpureofuscus, Doratomyces verrucisporus, Myceliophthora thermophila, Phoma eupyrena,
and Thermoascus crustaceus showed remarkable degradation ability (>70 %) regardless of
the number of chlorine substituents on the biphenyl nucleus and a high tolerance towards
PCBs. To our knowledge, this is the first study that demonstrates the ability of PCB degra-
dation by these species and indicates the potential effectiveness of some autochthonous
fungal strains in bioremediation systems.
ª 2013 The British Mycological Society. Published by Elsevier Ltd. All rights reserved.
Introduction hydraulic fluids, plasticizers or fire retardant because of their
Polychlorinated biphenyls (PCBs) are a class of compounds
containing a biphenyl molecule that is chlorinated to form
209 different congeners (Beaudette et al. 1998). These mole-
cules have been largely used in industries with numerous ap-
plications mainly in electrical equipment and were used as
1; fax: þ33 476 51 44 63.renoble.fr (B. Mouhamaritish Mycological Societ
physical properties (Pieper & Seeger 2008; Xing et al. 2010). Be-
cause of this widespread use, a large amount of PCBs has been
spread into environment, accumulated in soils and sedi-
ments, and represents a threat to ecosystem health due to
their low degradability and high rate of toxicity (Pointing
2001; Dercova et al. 2009). Through their lipophilicity, PCBs
dou).y. Published by Elsevier Ltd. All rights reserved.

Table 1 e Concentration of the seven PCB congeners inthe studied soil.
PCB congeners mg g�1 Dry matter
28 (2,4,40-Trichlorobiphenyl) 0.45
52 (2,20,5,50-Tetrachlorobiphenyl) 18.35
101 (2,20,4,5,50-Pentachlorobiphenyl) 51.20
118 (20,3,4,40,50-Pentachlorobiphenyl) 33.00
138 (2,20,3,4,40,50-Hexachlorobiphenyl) 44.98
153 (2,20,4,40,5,50-Hexachlorobiphenyl) 101.77
180 (2,20,3,4,40,5,50-Heptachlorobiphenyl) 70.94
Potential of autochthonous fungal strains for degradation of PCBs 269
are able to bioaccumulate and biomagnify along the food
chain (Bedard et al. 2007). They are readily stored in fatty tis-
sues and confer acute or chronic toxicity to animals and
humans and they are also considered as potentially carcino-
genic (McLachlan 1996; Pointing 2001).
Biological technologies using microorganisms or their en-
zymes to promote toxic compounds degradation are widely
investigated because of their low environmental impact and
their economic advantage compared to the physicochemical
methods. Bacteria are the most studied microorganisms for
the biological destruction of PCBs and numerous studies
have shown their capacity to degrade less-chlorinated conge-
ners (Abramowicz 1990; Quensen et al. 1990; Rojas-Avelizapa
et al. 1999). The degradation mechanisms are well known
and involve dehalogenase enzymes which catalyse the selec-
tive removal of meta- and para-chlorine and the dioxygenase
degradative pathway resulting in the degradation or mineral-
ization of low chlorinated congeners (Abramowicz 1990;
Pieper 2005; Pieper & Seeger 2008). However, the efficiency of
bacterial remediation is low in the highly polluted habitats
and negatively affected by the bioavailability of pollutants
making them inaccessible to unicellular organisms (Harms
et al. 2011).
The potential use of fungi able to degrade PCB congeners
can be an attractive alternative compared to the bacterial sys-
tem. Firstly, fungi are ubiquitous microorganisms found in
aquatic sediments, terrestrial habitats, and water surfaces
and their hyphal development promotes access to pollutants
(April et al. 2000). Secondly, many reports have demonstrated
the capacity of fungal species such as Phanerochaete chrysospo-
rium, Lentinus tigrinus, Pleurotus ostreatus, Trametes versicolor or
some filamentous ascomycetous fungi to degrade various PCB
congeners (Kubatova et al. 2001; Ruiz-Aguilar et al. 2002; Kamei
et al. 2006; Tigini et al. 2009; Federici et al. 2012). However, al-
though the nonspecific oxidative exoenzymes such as lignin
peroxidase, manganese-dependent peroxidase, and laccases
have been described as involved in the degradation of numer-
ous organic pollutants, their direct involvement in PCB degra-
dation has not been demonstrated.
Despite the interesting ecological and biochemical proper-
ties of fungal species, their introduction for bioremediation
purposes in environments which differ from their natural
habitats proved to be inefficient (Baldrian 2008; Harms et al.
2011). This phenomenon might be either due to their inability
to compete with indigenousmicrobiota or to their nonadapta-
tion to these new environments. For example, studies on the
degradation of organic pollutants such as fluorene have
shown a greater degradation potential of autochthonous fun-
gal strains isolated from contaminated soils than those from
similar uncontaminated environments (Garon et al. 2004).
Similarly, the role of autochthonous fungi in degradation of
aromatic hydrocarbons has been shown (D’Annibale et al.
2006). Up to now, only few studies have investigated the effec-
tiveness of the autochthonous fungal species in the biodegra-
dation of PCBs (Tigini et al. 2009).
In this study, the potential of fungal strains isolated from
PCB contaminated soils in PCBs biodegradation was evalu-
ated. Fungal strains were isolated from former industrial sites
and identified bymorphological andmolecular methods, with
the latter relying on the nuclear ribosomal internal
transcribed spacer (ITS) sequences. Their capacity to degrade
seven PCB congeners was assessed in liquid cultures of the
identified isolates by gas chromatographic analysis.
Materials and methods
Materials
PCB 28, 52, 101, 118, 138, 153, and 180 that are the most abun-
dant in the environmental and biological matrices, were ob-
tained from SigmaeAldrich Corp. (St. Louis, MI, USA). A
stock solution containing 15 mg of each PCB in 105 ml of iso-
octane was prepared and stored in a freezer (�20 �C). Thecontaminated soil was sampled from a former industrial
site located in Metz (France). The concentrations of the
seven PCB congeners investigated in this study are shown
(Table 1).
Fungal isolation and morphological identification
For the isolation of autochthonous filamentous fungal strains,
1 g of contaminated soil was added to 9ml of sterilewater con-
taining 0.05 % of sodium dodecyl sulphate (w/v). After stirring,
aliquots of the suspension were serially diluted and spread
onto Petri dishes containing malt extract agar (1.5 % w/v) sup-
plemented with 0.05 % chloramphenicol. Cultures were incu-
bated at 25 �C or 37 �C to optimize the isolation of strains
possessing different optimal growth temperatures. The iso-
lated strains were characterized by morphological criteria
and identified according to the general principle of fungal
classification (Domsch et al. 1980; Arx 1981).
DNA extraction and PCR
For themolecular analysis of the isolated strains, total DNA of
each strain was extracted using a Fast DNA Spin Kit (QBIO-
gene, Germany) according to the manufacturer’s recommen-
dations. The PCRs were carried out according to
conventional protocols using Ampli Taq Gold DNA polymer-
ase (Applied Biosystems, USA) and the primers ITS4 and
ITS5 (White et al. 1990) synthesized by Eurogentec (Seraing,
Belgium) were used to amplify the fungal ITS regions. The
PCRs were performed in a programmable thermal cycler
GeneAmps 2720 (Applied Biosystems). Amplifications were
carried out in 50 ml reaction mixtures as described by Molitor
et al. (2010).

270 B. Mouhamadou et al.
Sequencing and sequence analysis
The PCR product of each strain was sequenced by Beckman
Coulter Genomics (United Kingdom). Comparisons with se-
quences from the GenBank databases were made using the
BLAST search algorithm (Altschul et al. 1990). Alignments of
nucleotide sequences were carried out with the Clustal W
software (Thompson et al. 1994). Phylogenetic analyses were
carried out using the neighbour-joining method based on
Clustal W alignments and the robustness of tree topologies
was evaluated by performing bootstrap analysis of 1000 data
sets using MEGA 3.1 (Tamura et al. 2007). For the alignment
and phylogeny, only the 5.8S ribosomal DNA sequences and
the partial sequences of ITS1 and ITS2 were used because of
the high polymorphism of the ITS regions across the phyloge-
netically distant strains.
Biodegradation test in liquid medium
The mycelium of each isolated strain, cultivated in malt ex-
tract (1.5 % w/v), was scraped with a scalpel and ground
with an Ultra Turrax homogenizer (IKA T8, Germany) in 3 ml
of sterile water. 400 ml aliquots were introduced into 100 ml
Erlenmeyer flasks containing 20 ml of modified GS liquid me-
dium (Galzy & Slonimski 1957) supplemented with glucose
(5 g l�1) and incubated at 25 �C or 37 �C for the thermophilic
strains on a rotary shaker (180 rpm). After 48 h incubation,
the culture of each strain was spiked with 400 ml of a PCB
mix (seven PCB congeners in isooctane) giving a final concen-
tration of 3 mg of each congener per ml. All Erlenmeyer flasks
were sealed with Teflon stoppers (VWR, Fontenay-sous-Bois,
France) to prevent evaporation of PCBs. Each experiment con-
taining each strain was performed in triplicate and included
cell-free flasks for stability assessment. After 7 d incubation
on a rotary shaker at 120 rpm, the cultures were stopped
and the mycelium was separated from the culture medium
by filtering through Whatman 40 filter.
PCB extraction and Gaz chromatography (GC) analyses
Culture medium and fungal mycelium were extracted by us-
ing hexane. Culture medium was extracted three times with
20 ml of hexane with agitation (250 rpm, room temperature)
for 30min. Myceliumwas first homogenized in the serum bot-
tles with a POTTER homogenizer in 20 ml of hexane and incu-
bated 24 h at room temperature with agitation (250 rpm) then
extracted three times with 20 ml of hexane with agitation
(250 rpm, room temperature, 30 min). The organic phases
from culture medium and mycelium were evaporated under
vacuum using a rotary evaporator and adjusted to a volume
of 5 ml.
For the extraction of PCBs from the contaminated soil, 5 g
of dry soil were extracted using hexane acetone (1:1, v/v)
with the extractor ASE (Accelerated Solvent Extraction) (Agi-
lent Technologies, USA) according to the manufacturer’s
recommendations.
For the PCB analyses, aliquots of 200 ml were analysed using
a gas chromatograph (7890A), equipped with a DB-XLB capil-
lary column (ID: 0.25 mm, Length: 60 m, Film: 0.25 mm) and
an electron capture detector (mECD) (Agilent Technologies,
USA). The carrier gas was hydrogen. The injector temperature
was 250 �C. The detector temperature was 310 �C. The column
temperature was 50 �C. PCB 30 (2,4,6-trichlorobiphenyl) was
used as an internal standard.
The PCB recovery yield was determined from the cell-free
controls at initial time. The abiotic losses were determined
from the cell-free controls incubated in parallel with fungal
cultures.
Since the ultimate aim of this study was to provide novel
strains for bioaugmentation applications, the selection of
the fungal strains that we considered as the most effective is
based on two criteria: high rate of degradation and nonpatho-
genicity of the strain.
Statistical analysis
A statistical method to partition the sums of squares of Y
among experimental treatments (Anderson 2001) was
used. Two experimental factors were analysed, namely
strains (A) and PCB congeners (B). The generic form of the
model was thus: Y ¼ A þ B þ ε, where ε is an error term.
This multivariate analysis of variance was carried out using
the function adonis of the package vegan (Anderson 2001;
Oksanen et al. 2009), a method similar to a redundancy anal-
ysis (McArdle & Anderson 2001). Sums of squares and
resulting F-tests from permutations of the raw data were
calculated to test for the significance of experimental fac-
tors on Y. We used the KruskaleWallis tests (Kruskal &
Wallis 1952) to test for each selected strain, significant dif-
ferences between presence and absence of PCB. All statisti-
cal analyses were performed using R package vegan
(Oksanen et al. 2009).
Results
Characterization of species isolated from the contaminatedsoil
Strains isolated from PCB contaminated soil were identified by
their morphological criteria. A total of 24 strains were found
and all these strains belonged to Ascomycota and Zygomycota
phyla. A phylogenetic analysis was performed by adding the
reference sequences available in the GenBank database. The
phylogram on Fig 1 confirmed the morphological identifica-
tion of the strains and showed that the 24 strainswere divided
into 15 and four genera belonging to the Ascomycota (19
strains) and Zygomycota (five strains) phyla respectively.
Most genera were represented by a single strain with the ex-
ception of Trichoderma (two strains), Doratomyces (three
strains), Penicillium (two strains), and Mucor (two strains)
(Fig 1).
Analysis of PCB reduction
In our experimental procedure, the PCB recovery yield was
78 % and the abiotic losses were 7 %. Considering these data,
we determined the percentage of depletion of PCBs by taking
into account the quantity of residual PCBs in the culture me-
dium and in themycelium (biosorption) of each strain. Results

Doratomyces nanus (S2-12): JX537957
D. purpureofuscus FJ914688.1
D. purpureofuscus (S2-32): JX537967
D. verrucisporus (S2-33): JX537968
D. stemonitis JN104543.1
Pseudallescheria boydii (S2-23): JX537963
Gliomastix roseogrisea (S2-17): JX537960
Fusarium solani (S2-27): JX537966
Trichoderma spirale JF439515.1
Trichoderma harzianum (S2-18): JX537961
T. harzianum (S2-19): JX537962
Myceliophthora thermophila (S2-1): JX537951
Chaetomium murorum JQ946413.1
C. piluliferum AB625587
C. piluliferum (S2-2): JX537970
Thielavia sp (S2-24): JX537964
Geomyces pannorum (S2-11): JX537957
Geomyces pannorum HQ533810.1
Thermoascus crustaceus (S2-7): JX537956
T. crustaceus U18353.1
Aspergillus terreus FJ011538.1
Neosartorya pseudofischeri (S2-14): JX537959
A. fumigatus (S2-9): JX537971
Penicillium chrysogenum (S2-20): JX537974
P. aurantiogriseum (S2-34): JX537969
Phoma eupyrena (S2-26): JX537965
Mortierella elongata (S2-6): JX537954
M. elongata JF439485.1
Galactomyces geotrichum (S2-13): JX537958
G. geotrichum HE799669.1
Mucor circinelloides (S2-3): JX537952
M. plumbeus (S2-5): JX537955
Lichtheimia corymbifera (S2-4): JX537953
Circinella muscae (S2-10): JX537972
100
100
100
100
96
5087
98
60
99
9894
95
90
100
97
95
9390
52
94
99
54
0.05
Fig 1 e Consensus trees obtained using the neighbour-joining method based on aligned ITS sequences. Bootstrap values are
shown on the branches. The accession numbers of reference sequences which have been recovered from GenBank are in-
dicated. The strains isolated in this study and their accession numbers are indicated in bold.
Potential of autochthonous fungal strains for degradation of PCBs 271
showed that the reduction of PCBs was variable according to
the strains (Table 2). For instance, Circinella muscae showed
the lowest reduction of PCBs (9.76 %). Fifteen strains showed
intermediate rates of PCB reduction (20e70 %). Finally the
cultures of eight strains (Thermoascus crustaceus, Neosartorya
pseudofischeri, Doratomyces purpureofuscus, Myceliophthora
thermophila, Aspergillus fumigatus, Doratomyces nanus, Dorato-
myces verrucisporus, and Phoma eupyrena) led to an important
loss of PCB (>70 %). Because A. fumigatus and the related spe-
cies N. pseudofischeri are pathogenic fungi that cannot be used
in the process of bioremediation by bioaugmentation, we used
the six other strains only for further analysis.
Tolerance and efficiency of the selected strains in disruption ofPCB congeners
The tolerance of each of the six selected strains towards PCBs
was assessed by comparing the mycelial dry weight of each
strain incubated in presence or in the absence of PCBs. There
were no statistically significant differences in growth of any
fungal isolate when grown in the presence or absence of
PCBs, as determined by KruskaleWallis analysis (Fig 2).
The effectiveness of the selected strains in the biodegrada-
tion of each PCB congener was determined by calculating the
equivalent of the consumption of each PCB per mg of mycelial
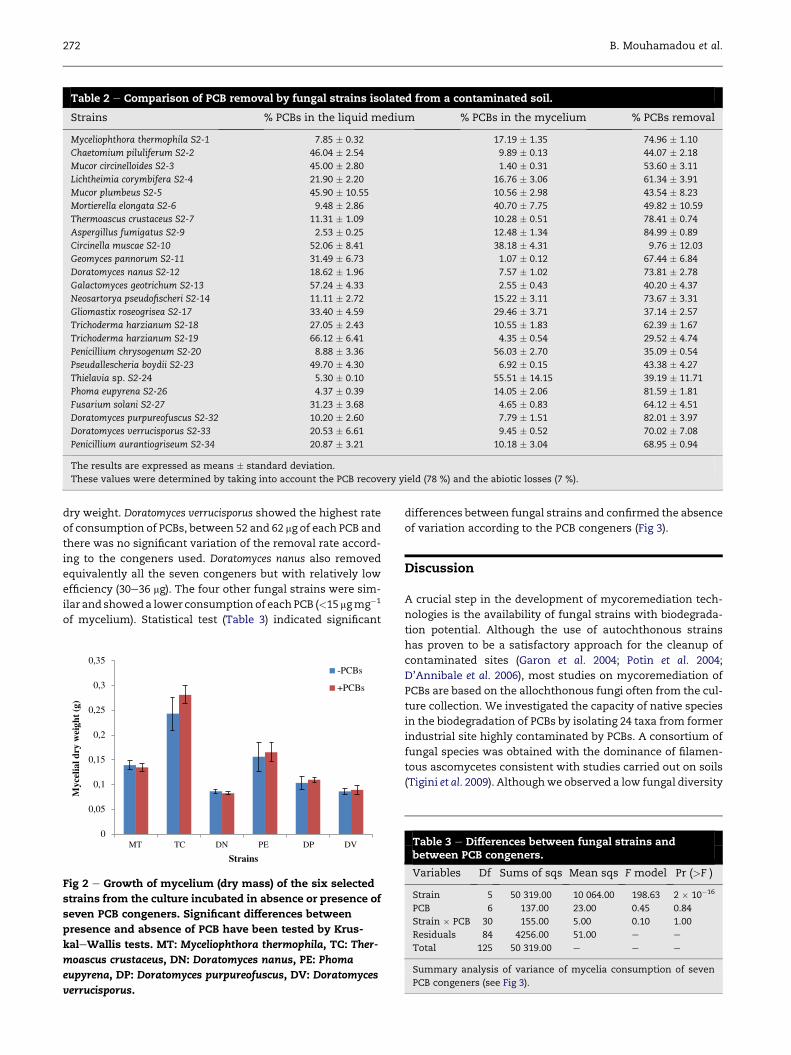
Table 2 e Comparison of PCB removal by fungal strains isolated from a contaminated soil.
Strains % PCBs in the liquid medium % PCBs in the mycelium % PCBs removal
Myceliophthora thermophila S2-1 7.85 � 0.32 17.19 � 1.35 74.96 � 1.10
Chaetomium piluliferum S2-2 46.04 � 2.54 9.89 � 0.13 44.07 � 2.18
Mucor circinelloides S2-3 45.00 � 2.80 1.40 � 0.31 53.60 � 3.11
Lichtheimia corymbifera S2-4 21.90 � 2.20 16.76 � 3.06 61.34 � 3.91
Mucor plumbeus S2-5 45.90 � 10.55 10.56 � 2.98 43.54 � 8.23
Mortierella elongata S2-6 9.48 � 2.86 40.70 � 7.75 49.82 � 10.59
Thermoascus crustaceus S2-7 11.31 � 1.09 10.28 � 0.51 78.41 � 0.74
Aspergillus fumigatus S2-9 2.53 � 0.25 12.48 � 1.34 84.99 � 0.89
Circinella muscae S2-10 52.06 � 8.41 38.18 � 4.31 9.76 � 12.03
Geomyces pannorum S2-11 31.49 � 6.73 1.07 � 0.12 67.44 � 6.84
Doratomyces nanus S2-12 18.62 � 1.96 7.57 � 1.02 73.81 � 2.78
Galactomyces geotrichum S2-13 57.24 � 4.33 2.55 � 0.43 40.20 � 4.37
Neosartorya pseudofischeri S2-14 11.11 � 2.72 15.22 � 3.11 73.67 � 3.31
Gliomastix roseogrisea S2-17 33.40 � 4.59 29.46 � 3.71 37.14 � 2.57
Trichoderma harzianum S2-18 27.05 � 2.43 10.55 � 1.83 62.39 � 1.67
Trichoderma harzianum S2-19 66.12 � 6.41 4.35 � 0.54 29.52 � 4.74
Penicillium chrysogenum S2-20 8.88 � 3.36 56.03 � 2.70 35.09 � 0.54
Pseudallescheria boydii S2-23 49.70 � 4.30 6.92 � 0.15 43.38 � 4.27
Thielavia sp. S2-24 5.30 � 0.10 55.51 � 14.15 39.19 � 11.71
Phoma eupyrena S2-26 4.37 � 0.39 14.05 � 2.06 81.59 � 1.81
Fusarium solani S2-27 31.23 � 3.68 4.65 � 0.83 64.12 � 4.51
Doratomyces purpureofuscus S2-32 10.20 � 2.60 7.79 � 1.51 82.01 � 3.97
Doratomyces verrucisporus S2-33 20.53 � 6.61 9.45 � 0.52 70.02 � 7.08
Penicillium aurantiogriseum S2-34 20.87 � 3.21 10.18 � 3.04 68.95 � 0.94
The results are expressed as means � standard deviation.
These values were determined by taking into account the PCB recovery yield (78 %) and the abiotic losses (7 %).
272 B. Mouhamadou et al.
dry weight. Doratomyces verrucisporus showed the highest rate
of consumption of PCBs, between 52 and 62 mg of each PCB and
there was no significant variation of the removal rate accord-
ing to the congeners used. Doratomyces nanus also removed
equivalently all the seven congeners but with relatively low
efficiency (30e36 mg). The four other fungal strains were sim-
ilar and showeda lower consumption of eachPCB (<15 mgmg�1
of mycelium). Statistical test (Table 3) indicated significant
Myc
elia
l dry
wei
ght
(g)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35-PCBs
+PCBs
Strains
MT TC DN PE DP DV
Fig 2 e Growth of mycelium (dry mass) of the six selected
strains from the culture incubated in absence or presence of
seven PCB congeners. Significant differences between
presence and absence of PCB have been tested by Krus-
kaleWallis tests. MT: Myceliophthora thermophila, TC: Ther-
moascus crustaceus, DN: Doratomyces nanus, PE: Phoma
eupyrena, DP: Doratomyces purpureofuscus, DV: Doratomyces
verrucisporus.
differences between fungal strains and confirmed the absence
of variation according to the PCB congeners (Fig 3).
Discussion
A crucial step in the development of mycoremediation tech-
nologies is the availability of fungal strains with biodegrada-
tion potential. Although the use of autochthonous strains
has proven to be a satisfactory approach for the cleanup of
contaminated sites (Garon et al. 2004; Potin et al. 2004;
D’Annibale et al. 2006), most studies on mycoremediation of
PCBs are based on the allochthonous fungi often from the cul-
ture collection. We investigated the capacity of native species
in the biodegradation of PCBs by isolating 24 taxa from former
industrial site highly contaminated by PCBs. A consortium of
fungal species was obtained with the dominance of filamen-
tous ascomycetes consistent with studies carried out on soils
(Tigini et al. 2009). Althoughwe observed a low fungal diversity
Table 3 e Differences between fungal strains andbetween PCB congeners.
Variables Df Sums of sqs Mean sqs F model Pr (>F )
Strain 5 50 319.00 10 064.00 198.63 2 � 10�16
PCB 6 137.00 23.00 0.45 0.84
Strain � PCB 30 155.00 5.00 0.10 1.00
Residuals 84 4256.00 51.00 e e
Total 125 50 319.00 e e e
Summary analysis of variance of mycelia consumption of seven
PCB congeners (see Fig 3).

0
10
20
30
40
50
60
70
802852101118153138180
Qua
ntit
y of
eac
h P
CB
con
gene
r (µ
g / m
g of
myc
elia
l dry
wei
ght)
MT TC DN PE DP DV
a
b
c
cddd
Fig 3 e Consumption of each PCB congener by the six se-
lected strains. Lower-case letters indicate significant dif-
ferences between strains but not between congeners within
strains (see Table 3).
Potential of autochthonous fungal strains for degradation of PCBs 273
in comparison to that estimated in unpolluted soils (Thorn
1997), our result is consistent with previous studies
(D’Annibale et al. 2006; Tigini et al. 2009) and suggests that
the high quantity of pollutants (>300mg g�1 of soil dry weight)
may exert a selective effect on soil fungi. This effect could re-
sult from the selection of strains tolerant to pollutants as
shown by Martino et al. (2000) or strains potentially able to de-
grade PCBs by the process of cometabolism. Interestingly,
some isolated strains, including those belonging to the genera
Aspergillus, Penicillium or Pseudallescheria have often been iso-
lated from polluted soils and their tolerance and their remark-
able degradative ability towards different organopollutants
have been reported (Pant & Adholeya 2007; Junghanns et al.
2008; Chang 2008; Tigini et al. 2009; Pinedo-Rilla et al. 2009).
The capacity of the isolated strains to degrade PCBswas de-
termined by taking into account both the remaining PCBs in
the culture medium and those extracted from the mycelium
corresponding to the fungal biosorption. Due to the strong
ability of PCBs to bind to the fungal biomass, the extraction
procedure of PCBs from the mycelium was modified from
that of PCBs present in the liquid medium by incubating the
mycelium of each strain 24 h in hexane with vigorous agita-
tion. We concluded that the substantial losses of PCBs were
essentially due to the fungal biodegradation. With the excep-
tion of Circinella muscae, the other strains exhibited variable
biodegradation capacity, from 29 to 85 %, comparable to the
result obtained by Ruiz-Aguilar et al. (2002) when using the ba-
sidiomycete ligninolytic fungi. This result confirms that most
fungi isolated from the highly polluted substrates are effective
in the pollutant biodegradation and is also consistent with the
reported high capacity for the ascomycetes to metabolize var-
ious organic chemicals (Harms et al. 2011).
We considered that efficient strains which could be used
for bioaugmentation applications should be nonpathogenic
and present at least 70 % of degrading rate. This criterion
was fulfilled by six strains among which three strains belong-
ing to the genus Doratomyces, two thermophilic strains
Myceliophthora thermophila and Thermoascus crustaceus, and
one strain belonging to the genus Phoma. The six selected
strains showed a high tolerance towards PCBs and a remark-
able ability to degrade different PCB congeners. In addition,
there was no significant selective degradation regarding the
number of chlorine on the biphenyl nucleus. This contrasts
with studies on the bacterial systems in which the extent of
degradation seemed to be affected by the degree or the posi-
tion of chlorine (Abramowicz 1990; Ohtsubo et al. 2000), but
is consistent with the study conducted by Federici et al.
(2012) which showed that different chlorinated PCBs were de-
graded to the same extent by the basidiomycetous fungus Len-
tinus tigrinus.
Conclusion
Our results based on GC analyses showed that the strains iso-
lated from a soil heavily contaminated by PCBs are tolerant
and likely to degrade these xenobiotics. The high rates of dis-
appearance of PCBs observed in the cultures of six strains sug-
gested a metabolization of PCBs by these fungi and their
ability to be used in bioremediation through bioaugmentation.
Our results are in line with other previous studies which have
reported a high potential of the autochthonous microflora in
bioremediation processes.
Acknowledgements
This research was financed by the Cluster Axelera (Pole de
comp�etitivit�e Chimie-Environnement Lyon & Rhone-Alpes).
The authors would like to thankMohamed Abdelghafour, Car-
ole Gaignaire, and JacquesMehu for their help on the GC-mass
analysis. We wish to thank Sophie P�erigon for her help con-
cerning the statistical analysis. We also really appreciated
the critical reading of the manuscript by Viviane Barbreau
and address our special thanks to Nael Mouhamadou for his
help.
r e f e r e n c e s
Abramowicz DA, 1990. Aerobic and anaerobic biodegradation ofPCB: a review. Critical Reviews in Biotechnology 10: 241e250.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ, 1990. Basiclocal alignment search tool. Journal of Molecular Biology 215:403e410.
Anderson MJ, 2001. A new method for non-parametric multivar-iate analysis of variance. Austral Ecology 26: 32e46.
April TM, Foght JM, Currah RS, 2000. Hydrocarbon-degrading fil-amentous fungi isolated from flare pit soils in northern andwestern Canada. Canadian Journal of Microbiology 46: 38e49.
Arx JA Von, 1981. The Genera of Fungi Sporulating in Pure Culture, 3rdedn. Cramer, Vaduz.
Baldrian P, 2008. Wood-inhabiting ligninolytic basidiomycetes insoils: ecology and constraints for applicability in bioremedia-tion. Fungal Ecology 1: 4e12.
Beaudette LA, Davies S, Fedorak PM, Ward OP, Pickard PA, 1998.Comparison of gas chromatography and mineralization ex-periments for measuring loss selected polychlorinated

274 B. Mouhamadou et al.
biphenyl congeners in cultures of white rot fungi. Applied andEnvironmental Microbiology 64: 2020e2025.
Bedard DL, Ritalahti KM, L€offler FE, 2007. The Dehalococcoidespopulation in sediment-free mixed cultures metabolicallydechlorinates the commercial polychlorinated biphenyl mix-ture aroclor 1260. Applied and Environmental Microbiology 73:2513e2521.
Chang YS, 2008. Recent developments in microbial biotransfor-mation and biodegradation of dioxins. Journal of Molecular Mi-crobiology and Biotechnology 15: 152e171.
D’Annibale A, Rosetto F, Leonardi V, Federici F, Petruccioli M,2006. Role of autochthonous filamentous fungi in bioremedi-ation of a soil historically contaminated with aromatic hy-drocarbons. Applied and Environmental Microbiology 72: 28e36.
Dercova K, Seligova J, Dudasova H, Mikul�a�sov�a M, �Silh�arov�a K,T�othov�a L, Hucko P, 2009. Characterization of the bottomsediments contaminated with polychlorinated biphenyls:evaluation of ecotoxicity and biodegradability. InternationalBiodeterioration and Biodegradation 63: 440e449.
Domsch KH, Gams W, Anderson TH, 1980. Compendium of SoilFungi. Academic Press, London.
Federici E, Giubilei M, Santi G, Zanaroli G, Negroni A, Fava F,Petruccioli M, D’Annibale A, 2012. Bioaugmentation of a his-torically contaminated soil by polychlorinated biphenyls withLentinus tigrinus. Microbial Cell Factories 23: 11e35.
Galzy P, Slonimski P, 1957. Variations physiologiques de la levureau cours de la croissance sur l’acide lactique comme seulesource de carbone. Comptes Rendus de l’Acad�emie des Sciences245D: 2423e2426.
Garon D, Sage L, Siegle-Murandi F, 2004. Effects of fungal bio-augmentation and cyclodextrin amendment on fluorine deg-radation in soil slurry. Biodegradation 15: 1e8.
Harms H, Schlosser D, Wick LY, 2011. Untapped potential: ex-ploiting fungi in bioremediation of hazardous chemicals. Na-ture Reviews Microbiology 9: 177e192.
Junghanns C, Krauss G, Schlosser D, 2008. Potential of aquaticfungi derived from diverse freshwater environments to de-colourise synthetic azo and anthraquinone dyes. BioresourceTechnology 99: 1225e1235.
Kamei I, Kogura R, Kondo R, 2006. Metabolism of 4,40-dichlorobi-phenyl by white-rot fungi Phanerochaete chrysosporium andPhanerochaete sp. MZ142. Applied Microbiology and Biotechnology72: 566e575.
Kruskal W, Wallis A, 1952. Use of ranks in one-criterion varianceanalysis. Journal of the American Statistical Association 47:583e621.
Kubatova A, Erbanova P, Eichlerova I, Homolka L, Nerud F,Sasek V, 2001. PCB congener selective biodegradation by thewhite rot fungi Pleurotus ostreatus in contaminated soil. Che-mosphere 43: 207e215.
Martino E, Turnau K, Girlanda M, Bonfante P, Perotto S, 2000. Er-icoid mycorrhizal fungi from heavy metal polluted soils: theiridentification and growth in the presence of zinc ions. Myco-logical Research 104: 338e344.
McArdle BH, Anderson MJ, 2001. Fitting multivariate models tocommunity data: a comment on distance-based redundancyanalysis. Ecology 82: 290e297.
McLachlan MS, 1996. Bioaccumulation of hydrophobic chemicalsin agricultural food chains. Environmental Science and Technol-ogy 30: 252e259.
Molitor C, Inthavong B, Sage L, Geremia RA, Mouhamadou B, 2010.Potentiality of the cox1 gene in the taxonomic resolution ofsoil fungi. FEMS Microbiology Letters 302: 76e84.
Ohtsubo Y, Nagata Y, Kimbara K, Takagi M, Ohta A, 2000. Ex-pression of the bph genes involved in biphenyl/PCB degrada-tion in Pseudomonas sp. KKS102 induced by the biphenyldegradation intermediate, 2-hydroxy-6-oxo-6-phenylhexa-2,4-dienoic acid. Gene 256: 223e228.
Oksanen J, Kindt R, Legendre P, O’Hara B, Gavin L, 2009. vegan:Community Ecology Package. R Package Version 1.15-4.
Pant D, Adholeya A, 2007. Identification, ligninolytic enzyme ac-tivity and decolorization potential of two fungi isolated froma distillery effluent contaminated site. Water Air and Soil Pol-lution 183: 165e176.
Pieper DH, 2005. Aerobic degradation of polychlorinated biphe-nyls. Applied Microbiology and Biotechnology 67: 170e191.
Pieper DH, Seeger MJ, 2008. Bacterial metabolism of polychlori-nated biphenyls. Journal of Molecular Microbiology and Biotech-nology 15: 121e138.
Pinedo-Rilla C, Aleu J, Collado IG, 2009. Pollutants biodegradationby fungi. Current Organic Chemistry 13: 1194e1214.
Pointing SB, 2001. Feasibility of bioremediation by white rot fungi.Applied Microbiology and Biotechnology 57: 20e33.
Potin O, Rafin C, Veignie E, 2004. Bioremediation of an agedpolycyclic aromatic hydrocarbons (PAHs)-contaminated soilby filamentous fungi isolated from the soil. International Bio-deterioration and Biodegradation 54: 45e52.
Quensen III JF, Boyd SA, Tiedje JM, 1990. Dechlorination of fourcommercial polychlorinated biphenyl mixtures (Aroclors) byanaerobic microorganisms from sediments. Applied and Envi-ronmental Microbiology 56: 2360e2369.
Rojas-Avelizapa NG, Rodriguez-Vazquez R, Enriquez-Villanueva F, Martinez-Cruz J, Poggi-Varaldo HM, 1999.Transformer oil degradation by an indigenous microflora iso-lated from a contaminated soil. Resources, Conservation andRecycling 27: 15e26.
Ruiz-Aguilar GML, Fernandez-Sanchez JM, Rodriguez-Vazquez R,Poggio-Varaldo H, 2002. Degradation by white-rot fungi of highconcentrations of PCB extracted from a contaminated soil.Advances in Environmental Research 6: 559e568.
Tamura K, Dudley J, Nei M, Kumar S, 2007. MEGA4: molecularevolutionary genetics analysis (MEGA) software version 4.0.Molecular Biology and Evolution 24: 1596e1599.
Thompson JD, Higgins DG, Gibson TJ, 1994. CLUSTAL W: im-proving the sensitivity of progressive multiple sequencealignment through sequence weighting, position specific gappenalties and weight matrix choice. Nucleic Acids Research 22:4673e4680.
Thorn G, 1997. The fungi in soil. In: Modern Soil Microbiology.Marcel Dekker, pp. 63e127.
Tigini V, Prigione V, Di Toro S, Fava F, Varese GC, 2009. Isolationand characterization of polychlorinated biphenyl (PCB) de-grading fungi from a historically contaminated soil. MicrobialCell Factories 8: 5.
White TJ, Bruns T, Lee STJ, 1990. PCR Protocols: A Guide to Methodsand Applications.
Xing GH, Wu SH, Wong MH, 2010. Dietary exposure to PCBs basedon food consumption survey and food basket analysis atTaizhou, China e the World’s major site for recycling trans-formers. Chemosphere 81: 1239e1244.