presence and characteristics of nadp-malic enzyme in soybean nodule bacteroids
TRANSCRIPT

This article was downloaded by: [Laurentian University]On: 05 October 2014, At: 18:49Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office:Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Soil Science and Plant NutritionPublication details, including instructions for authors and subscriptioninformation:http://www.tandfonline.com/loi/tssp20
Presence and characteristics of nadp-malicenzyme in soybean nodule bacteroidsIsao Kimura a & Shigeyuki Tajima ba Kagawa Prefectural Fermentation and Foods Experimental StationTakamatsu Branch , Takamatsu , 761 , Japanb Department of Bioresource Science, Faculty of Agriculture , KagawaUniversity , Miki-cho, Kita-gun, Kagawa, 761-07 , JapanPublished online: 14 May 2012.
To cite this article: Isao Kimura & Shigeyuki Tajima (1989) Presence and characteristics of nadp-malic enzyme in soybean nodule bacteroids, Soil Science and Plant Nutrition, 35:2, 271-279, DOI:10.1080/00380768.1989.10434759
To link to this article: http://dx.doi.org/10.1080/00380768.1989.10434759
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and ourlicensors make no representations or warranties whatsoever as to the accuracy, completeness, orsuitability for any purpose of the Content. Any opinions and views expressed in this publicationare the opinions and views of the authors, and are not the views of or endorsed by Taylor &Francis. The accuracy of the Content should not be relied upon and should be independentlyverified with primary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilitieswhatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantialor systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, ordistribution in any form to anyone is expressly forbidden. Terms & Conditions of access and usecan be found at http://www.tandfonline.com/page/terms-and-conditions

Soil ScL Plant Nutr .. 3S (2). 271-279. 1989
PRESENCE AND CHARACTERISTICS OF NADP-MALIC ENZYME IN SOYBEAN NODULE BACTEROIDS
Isao KIMURA and Shigeyuki T AJIMA *
Kagawa Prefectural Fermentation and Foods Experimental Station Takamatsu Branch. Takamatsu. 76/ Japan
• Department of Bioresource Science. Faculty of Agriculture. Kagawa University. Miki-cho. Kita-gun. Kagawa. 76/-07 Japan
Received July IS. 1988
The localization and characteristics of NADP-malic enzyme [EC 1.1.1.40]. a key enzyme for succinate oxidation in nodule bacteroids. were surveyed in soybean nodules and bacteroids.
Protoplasts prepared from nodules were separated into cortex cell. infected cell. and uninfected cell groups following the determination of NADP-malic enzyme activity in those cells. The enzyme activity was localized in the cortex cells and the infected cells. In the infected cells the malic enzyme activity was not localized in the cytosol but in the bacteroids. The uninfected cells showed no detectable enzyme activity.
The NADP-malic enzyme activity in the bacteroids prepared by Percoll density gradient centrifugation or by differential centrifugation was almost identical with that of the infected cells (mg protein basis). These data indicate that N ADP-malic enzyme in soybean nodules was localized both in the cortex cells and in the bacteroids.
Staining for the detection of the NADP-malic enzyme activity after polyacrylamide gel electrophoresis and the enzyme activity assay after isoelectrofocusing revealed that the malic enzyme in the bacteroids exhibited a unique Rf value from that in the whole nodules. indicating that the NADP-malic enzyme in Bradyrhizobium japonicum J501 bacteroids was from that in the cortex cells.
The NADP-malic enzyme in the bacteroids was partially purified. and the characteristics were surveyed. At least two NADP-malic enzymes were present in the nodule bacteroids. and a major malic enzyme activity in the bacteroids was purified 127 fold. The enzyme showed an optimum pH at 7.5. a Km value at 109 J.lM for malic acid and absolute requirement of NADP for the enzyme reaction. Strong inhibition of the enzyme activity by citrate. malonate. and glyoxylate was observed. suggesting a possible regulatory role of the malic enzyme in the organic acid metabolism in bacteroids.
Key Words: carbon metabolism. malic enzyme. nitrogen fixation. soybean nodules.
Though it is well recognized that dicarboxylic acids. such as succinate or malate. can act as bacteroid in vivo respiratory substrate to promote nitrogen fixation in soybean nodules, the profile of the energy metabolism is poorly documented (KOUCHI and YONEY AMA 1986; STREETER and SALMINEN 1985; T AJIMA et al. 1986; WATERS et al. 1985). Our previous reports showed that the isolated bacteroids activity oxidized exogenous He-succinate as a respiratory substrate even under low oxygen supply if the
271
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4

272 I. KIMURA and S. T AJIMA
system contained 100 pM leghemoglobin, and it was suggested that the citric acid cycle was the main pathway for the oxidation of succinate in the bacteroids (T AJIMA et al. 1986). In the bacteroids the NAD- or NADP-malic enzyme was assumed to be a key enzyme for the citric acid cycle function since acetyl-Co A supply from malic acid under the combined reaction of the malic enzyme and pyruvate dehydrogenase is necessary for citrate formation, and other enzymes concerned with acetyl-CoA formation from malic acid showed very low activities in the bacteroids.
Since the activity of the malic enzyme plays a key role in the organic acid metabolism in mitochondria or anaerobic metabolism, the enzyme activity was studied extensively and found to be widely distributed in animals (T ARONI et aL 1988) and bacteria (BARTOLUCCI et al. 1987). In nodules of Vida Jaba, the NADP-malic enyme activity was already reported to be localized in the plant cytosol (LAWRIE and WHEELER 1975). In this paper the localization and characteristics on NADP-malic enzymes in soybean nodule bacteroids was analyzed, and the possibility that the NADP-malic enzyme could playa role as an alternative pathway to convert dicarboxylic acids to acetyl-Co A was examined.
MA TERIALS AND METHODS
Bradyrhizobium japonicum strain 1501 was supplied from the stock culture of the Institute of Agrobiological Sciences. Yeast extract-mannitol medium (TAJIMA et aL 1985) was modified to obtain a better growth, and it contained polypeptone (5 g/ liter) and sucrose (10 gil iter) instead of mannitol.
Soybean plants were grown for 6 to 7 weeks by hydroponics as previously reported (T AliMA and YAMAMOTO 1975). Bacteroids were prepared from freshly harvested nodules by differential centrifugation as previously reported (EMERICH et al. 1979).
Protoplast preparation. The freshly harvested nodules were sliced and the protoplasts were prepared as reported by KOUCHI et aL (1988). The enzyme extraction from the isolated infected, uninfected, and cortex cells was performed with 0.1 M HEPES buffer, pH 7.5 containing 5% (w Iv) polyclar AT and 2 mM dithiothreitol as previously described (KoucHI et al. 1988).
Biochemical assays. NADP-malic enzyme activity was determined according to the method of Ochoa (OCHOA 1955). The spectrophotometric assay system contained 135 nmol NADP, 1.5 pmol L-malic acid, and 3.0 pmol MnClz in 2.9 ml 25 mM glycylglycine buffer, pH 7.4, and the manometric assay system contained 34 nmol NADP, 20 nmol oxaloacetate, and I ,umol MnCl2 in 0.9 ml I M acetate buffer. pH 4.5. The enzyme reaction was performed at 30°C.
Protein content was determined according to the method of LOWRY et al. (1951), with bovine serum albumin as the standard.
After native polyacrylamide gel electrophoresis at 4°C the gel was stained for the detection of the NADP-malic enzyme activity using the modified method of WATERS
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4

NADP-Malic Enzyme in Soybean Nodule Bacteroids 273
et al. (1985). The isoelectrofocusing was performed according the method of 001 and OHTSURU
(1974). The crude extracts of the nodules and roots were dialyzed by passing through a Sephadex 0-25 column equilibrated with to mM glycine. The eluate was condensed, and subjected to preparative isoelectrofocusing following the NADP-malic enzyme activity assay in each I ml fraction.
RESULTS
Localization of NADP-malic enzyme activities in nodules tissues Table I shows the distribution of the NADP-malic enzyme activity in various
cells or tissues of the nodules. Similar enzyme activities per mg protein were detected in the nodules, roots, and bacteroids. Since a high NADP-malic enzyme activity was present in various tissues and the enzyme activity in the bacteroids may have been due to the contamination of plant enzyme, differential centrifugation, and Percoll density gradient centrifugation were employed for the bacteroid preparation. The malic enzyme activity was detected in the bacteroids in both preparations. Spectrophotometric assay and manometric assay yielded basically identical results.
To determine the localization of the malic enzyme in various cells or tissues in the nodules, protoplasts were prepared from nodule slices, and the NADP-malic enzyme activity was determined after the protoplasts were separated into cortex cell, infected cell, and uninfected cell groups. The contamination by infected cells or by uninfected cells was less than 10% on the basis of marker enzyme distributions (infected cell, {j-hydroxybutyric acid dehydrogenase; uninfected cell, uricase). The enzyme activity was detected in the cortex cells and in the infected cells, although the activity (mg protein basis) in the infected cells was ca. 1/10 of that in the cortex cells (Table I).
Table \. Distribution of NADP-malic enzyme activity in various tissues and cells in soybean nodules. Bacteroids were prepared by Percoll gradient centrifugation (Bacteroids I) and by differential centrifugation (Bacteroids 2). The determination was performed in duplicate of three sample prepara-
tions, and the activity was calculated from the mean of each value.
Nodules
Whole
Infected ceIls
Uninfected cells
Cortex
Bacteroids I
Bacteroids 2
Root
NADP-malic enzyme activity (nmolfmg protein. min)
42.8 12.7 n.d.
49.2
40.3
43.1
49.3
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4

274 I. KIMURA and S. T AJIMA
- 669000 - 440000
- 232000
- 140000
- 67000
~ Front
ABC D
Fig. I. Polyacrylamide gel assay for NADP-malic enzymes from various soy ben plant tissues. Electrophoresis was performed on 7.5% polyacrylamide slab gel, and staining for NADPmalic enzyme activity was carried out at 37'C. Lane A, bacteroid extract; lane B, nodule extract; lane C, extract; lane D, molecular weight standards (from top to bottom: thyroglobulin (Mr =669,000), ferritin (Mr = 440,000), catalase (Mr = 232,000), lactate dehydrogenase (Mr = 140,000), albumin (Mr =67,000».
Since each type of cells requires different incubation times for the separation and the enzyme may lose the activity with time, the enzyme activity was determined at exactly 3 h after nodule slicing. The infected cells were further homogenized with 0.1 M
HEPES buffer, pH 7.S containing 0.4 M sucrose and 2 m M dithiothreitol. The homogenate was loaded on a Percoll density gradient. After centrifugation (100,000 X g, 60 min) the distribution of the activity was determined. The enzyme activity was detected in two fractions (bacteroid fraction, 37.8 nmol NADPH formed per mg protein per min; supernatant fraction, 1.27 nmol NADP formed per mg protein per min), suggesting that the enzyme activity in the infected cells is localized in the bacteroids.
NADP-malic enzyme activities in the extracts from nodules, roots, or bacteroids were assayed by native PAGE, and the gel was stained for the detection of NADPdependent malic enzyme activity. Two bands were seen in the root extract or nodule extract, and one band in the bacteroid extract. Figure I shows that roots and nodules contained NADP-malic enzymes with identical Rf values (roots and whole nodules; 0.2 and 0.31) though the enzyme in the bacteroids exhibited a Rf value of 0.37, suggesting that the NADP-malic enzyme in the bacteroids is different from major malic enzymes in the roots or in the nodules.
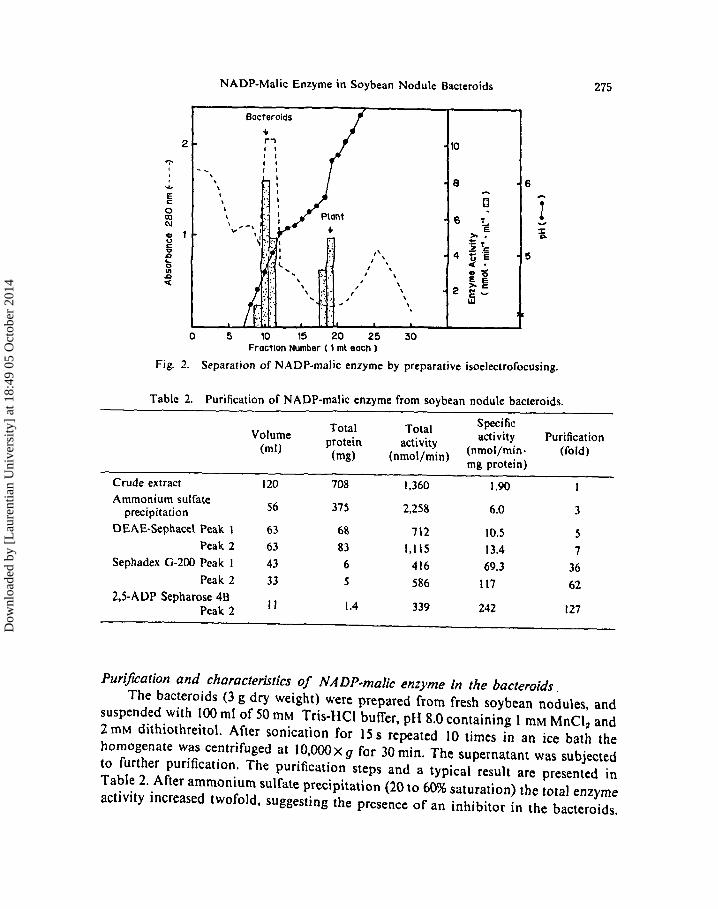
Crude extracts from roots, nodules, and bacteroids were subjected to the electrofocusing separately or as a mixed sample, and the results are shown in Fig. 2. After electrophoresis the malic enzyme activity in each ml was determined. The results revealed that mixture of the two tissue extracts exhibited two different NADP-malic enzyme activities at different pI (roots 6.1; bacteroids 5.2). The pI value of the N ADP-malic enzyme in the nodule extract was similar to that of the roots, suggesting that the major NADP-malic enzyme in roots and nodules gave an identical Rf value.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4
e c o co C\I
2
II> 1 u G n ... o III n <
Fig. 2.
o
Table 2.
--,
NADP-Malic Enzyme in Soybean Nodule Bacteroids
, \ \ \
\ \
Bacteroicls
.. r, I I I I I I
I
"
;. ,.
, ~:: .. , " ',} , :' .
" , '. , , , \
\ \
5 10 15 20 25 30 Fraction Number ( 1 ml each)
10
8
[]
6 !... E
>-,
4 ~ 'ic t; 'e < Ole> E E
2 >-c ~~ W
6
I :r .,.
5
Separation of NADP-malic enzyme by preparative isoelectrofocusing.
Purification of NADP-malic enzyme from soybean nodule bacteroids.
Specific
275
Volume (ml)
Total protein (mg)
activity Purification activity (nmol/min' (fold)
Total
(nmol/min) mg protein)
Crude extract 120 708 1,360 1.90 Ammonium sulfate 56 375 2,258 6,0 3 precipitation
DEAE-SephaceJ Peak I 63 68 712 10.5 5 Peak 2 63 83 1,115 13.4 7
Sephadex G-200 Peak 1 43 6 416 69.3 36 Peak 2 33 5 586 117 62
2,S-ADP Sepharose 4B 11 1.4 339 242 127 Peak 2
Purification and characteristics of NADP-malic enzyme in the bacteroids. The bacteroids (3 g dry weight) were prepared from fresh soybean nodules, and
suspended with 100 ml of 50 mM Tris-HCI butTer, pH 8.0 containing 1 mM MnCl, and 2 roM dithiothreitol. After sonication for 15 s repeated 10 times in an ice bath the homogenate was centrifuged at 10,000 X g for 30 min. The supernatant was subjected to further purification. The purification steps and a typical result are presented in Table 2. After ammonium sulfate precipitation (20 to 60% saturation) the total enzyme activity increased twofold, suggesting the presence of an inhibitor in the bacteroids.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4
276
E c o co (\j 1.0 II u ~
~ 1/1
~ 0.5
o
I. KIMURA and S. TAJIMA
Peck 2
, ,
r I'
, I
I
Peak 1 . I ;:
, I
-- . If
,,' f
I
50 100 150 200 Fraction Number ( 5 ml tach)
0.5 , 25 I ~
0.4 -I 20 T~ E
0.3 :E 15 is E c
>. :t
0.2 10 ~ .. u -< II
5 E >. N
tij
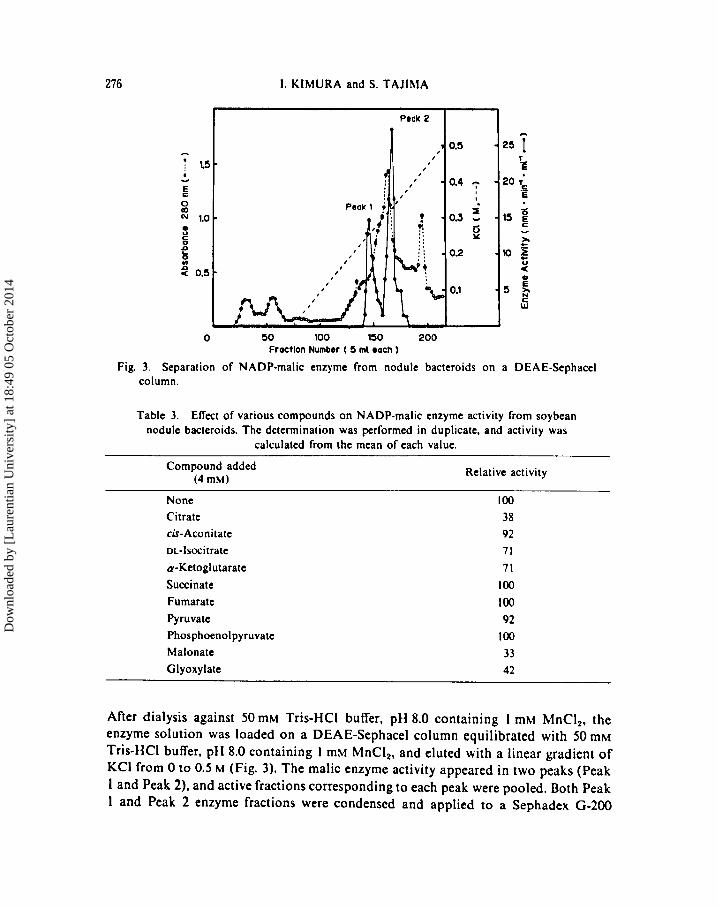
Fig. 3. Separation of NADP-malic enzyme from nodule bacteroids on a DEAE-Sephacel column.
Table 3. Effect of various compounds on NADP-malic enzyme activity from soybean nodule bacteroids. The determination was performed in duplicate. and activity was
calculated from the mean of each value.
Compound added Relative activity (4mM)
None 100 Citrate 38 cif-Aconitate 92
DL-Isocitrate 71 a-Ketoglutarate 71 Succinate 100 Fumarate 100 Pyruvate 92 Phosphoenol pyruvate 100 Malonate 33 Glyoxylate 42
After dialysis against 50 mM Tris-HCI buffer, pH 8.0 containing I mM MnCI 2 , the enzyme solution was loaded on a DEAE-Sephacel column equilibrated with 50 mM Tris-HCI buffer, pH 8.0 containing I mM MnCI2, and eluted with a linear gradient of KCI from 0 to 0.5 M (Fig. 3). The malic enzyme activity appeared in two peaks (Peak 1 and Peak 2), and active fractions corresponding to each peak were pooled. Both Peak 1 and Peak 2 enzyme fractions were condensed and applied to a Sephadex G-200
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4

NADP-MaJic Enzyme in Soybean Nodule Bacteroids 277
column (3.0 X 70 cm) that was equilibrated with 50 mM Tris-HCl buffer, pH 8.0. The activity of the two enzymes showed identical elution profiles by Sephadex G-200 column chromatography, suggesting that both malic enzymes have a similar molecular weight (data are not shown). The major NADP-malic enzyme (Peak 2) was loaded on a 2,5-ADP Sepharose 4B column (0.7 X 5 em), and the enzyme activity was eluted as a single peak with a linear gradient of NAD (0-5 mM) after washing the column with to mt of 50 mM Tris-HCI buffer, pH 8.0. This active fraction (Peak 2) was purified 127-fold (mg protein basis). The enzyme preparation showed no activity with NAD, stoichiometrical conversion of malate to pyruvate, and no detectable OAA formation from malate. These data indicate that the NADP-malate dehydrogenase activity in the enzyme preparation was negligible.
Table 3 shows the effects of various organic acids on the NADP-malic enzyme activity (Peak 2). A few organic acids (citrate, malonate, and glyoxylate) showed 60 to 70% inhibition at 4 mM, and isocitrate, a-ketoglutarate, and pyruvate inhibited the enzyme activity slightly. Succinate and malate had no effect on the malic enzyme activity.
DISCUSSION
The localization and characteristics of the NADP-malic enzyme, a key enzyme for succinate oxidation, were analyzed in soybean nodules.
For in vivo respiration of the nodule bacteroids, succinate or malate oxidation through the citric acid cycle has been recognized as a major pathway that can supply reductant and ATP for the nitrogenase activity (KoucHi and YONEYAMA 1986; STREETER and SALMINEN 1985; T AJIMA et al 1986; WATERS et aL 1985). For the continuous supply of citrate, however, succinate or malate must be converted to acetyl-CoA through an alternative pathway, in which the NAD, NADP-malic enzymes are considered to be involved in Bradyrhizobium japonicum J501 bacteroids.
NADP-malic enzyme activity was detected in the bacteroids prepared by differential centrifugation or by Percoll density gradient centrifugation (Table 1). The malic enzyme activity in the bacteroids was determined both by the manometric method and by the spectrophotometric method, resulting in identical activity values. These data indicate that the nodule bacteroids exhibited the NADP-malic enzyme activity, and the effect of the NADP-malate dehydrogenase activity was negligible.
Protoplasts were prepared from the nodule slices, and they were separated into cortex cell, infected cell, and un infected cell groups following the determination ofthe enzyme activity in the cells. The malic enzyme activity in the infected cells was mainly localized in the bacteroids, and no malic enzyme activity (Table 1) was detected in uninfected cells. These data suggest that the NADP-malic enzyme activity in the nodules was localized in the cortex cells and bacteroids. KOUCHI et aL (1988) also reported a similar localization of the NADP-malic enzyme activity in protoplasts from soybean nodules.
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4

278 I. KIMURA and S. T AliMA
One of the characteristics of the regulation of the organic acid metabolism is the extremely low oxygen concentration in the nodules (LAWRIE and WHEELER 1975). DAVIES (1980) suggested that flood-tolerant plants can accumulate malic acid in the vacuoles under conditions of anoxia since these plants lack the malic enzyme activity, though other plants accumulate ethanol. In the soybean nodules the cytosol contained pyruvate dehydrogenase and alcohol dehydrogenase (T AliMA and LARUE 1982), and in uninfected cells the specific activities of these enzymes were high (KOUCHI et al. 1988). These data suggest that anaerobic metabolism may be operative in uninfected cells in the nodules. However, the ethanol concentration in intact nodules was very low (T AJIMA and LARUE 1982), suggesting that the uninfected cells in the nodules accumulate malate instead of ethanol. The difference in the NADP-malic enzyme activity between the inner region (infected cells and uninfected cells) and outer region (cortex cells) of the nodules may result from anoxia in the nodules, and could playa key role in the organic acid metabolism and the maintenance of a very low ethanol concentration.
Staining for the detection of the malic enzyme activity after polyacrylamide gel electrophoresis showed that the malic enzyme activity was present in the roots or in the nodules and appeared as two bands on the gel with identical Rf values, which the enzyme activity in the bacteroids exhibited one band with different Rf values from those of the roots or nodules (Fig. I). The enzyme activity assay after isoelectrofocusing also revealed that the NADP-malic enzyme in the bacteroids was eluted in different fractions from those of the roots or whole nodules, indicating that the NADP-malic enzyme in Bradyrhizobium japonicum J501 bacteroids was different from the malic enzyme in the cortex cells and roots (Fig. 2).
The characteristics of the NADP-malic enzyme in the nodule bacteroids are comparable to those in animals (T ARONI et al 1988) and in bacteria (BARTOLUCCI et aL 1987). The malic enzyme activity in the bacteroids was stable during the purification and at least two NADP-malic enzymes were present in the bacteroids (Fig. 3). The overall purification of the major isozyme (Peak 2) was 127-fold, and the characteristics of the final preparation were analyzed. The optimum pH at 7.5, Km value for malic acid (109 pM), and absolute requirement of NADP for the enzyme reaction were similar to those of bacteria (BARTOLUCCI et aL 1987).
Table 3 shows that various organic acids in the citric acid cycle or glyoxylate cycle strongly inhibited the enzyme activity. Though citrate and glyoxylate were present in the bacteroids at low concentrations, the accumulation of these organic acids in the bacteroids could slow down the pyruvate formation from malic acids following spontaneous stalling from the citric acid cycle operation. Details of the regulatory role of the malic enzyme in the organic acid metabolism are under investigation in our laboratory.
Acknowledgments. This work was supported by a Grant-in-Aid from the Ministry of Agriculture. Forestry and Fisheries of Japan (GEP-87-I1).
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4

NADP-Malic Enzyme in Soybean Nodule Bacteroids 279
REFERENCES
BARTOLUCCI, S., RELLA, R., GUAGLIARDI, A., RAIA, C.A., GAMBACORTA, A., ROSA, M.D., and ROSSI, M. 1987: Malic enzyme from Archaebacterium. Sulfolobus solfataricus. J. BioL Chem., 262, 7725-7731
DA VIS, D.O. 1980: Anaerobic metabolism and the production of organic acids. In The Biochemistry of Plants, Vol. 2, Ed. D.O. Davies, p. 581-611, Academic Press, New York
001, E. and OHTSURU, C. 1974: Simplified isoelectric focusing columns of small scales. Agric. Bioi. Chem., 38, 1947-1948
EMERICH, D.W., ARUGUESO, T.R., CHING, T.M., and EVANS, H.J. 1979: Hydrogen-dependent nitrogenase activity and ATP formation in Rhizobium japonicum bacteroids. J. Bacteriol .• 137, 153-160
KOUCHI, H., FUKAI, H., KATAGIRI, K., MINAMISAWA, H., and TAJIMA, S. 1988: Isolation and enzymological characterization of infected and uninfected cell protoplasts from soybean (Glycine max L. Merr.) root nodules. Physiol. Plant., 73, 327-334
KOUCHI, H., YONEYAMA, T. 1986: Metablism of [l3C]-labeled photosynthate in plant cytosol and bacteroids of root nodules of Glycine max. Physiol. Plant., 68, 238-244
LAWRIE, A.C. and WHEELER, C.T. 1975: Nitrogen fixation in the root nodules of Vicia faba in relation to the assimilation of carbon. II. Dark fixation of carbon dioxide. New Phytol., 74, 437-445
LOWRY, O.H., ROUSEBROUGB, N.J., FARR, A.L., and RANDALL, RJ. 1951: Protein measurement with the Folin phenol reagent. J. Bioi. Chern., 193, 265-275
OCHOA, S. 1955: Malic enzyme. In Methods in Enzymology, Vol. I, Ed. S.P. Colowick and N.O. Kaplan, p. 739-753, Academic Press, New York
STREETER, J.G. and SALMINEN, S.O. 1985: Carbon metabolism in legume nodules. In Nitrogen Fixation Research Progress, Ed. B.J. Evans, P.J. Bottomley, and W.E. Newton, p. 277-284, Martinus Nijhoff Publishers, Dordrecht
TAJIMA, S., KIMURA, I., and SASAHARA, H. 1986: Succinate metabolism of isolated soybean nodule bacteroids at low oxygen concentration. Agric. BioL Chem., 50, 1009-1014
T AJIMA, S. and LARuE, T.A. 1982: Enzymes for acetaldehyde and ethanol formation in legume nodules. Plant PhysioL, 70, 388-392
TAJIMA, S., SASAHARA, K., KOUCHI, H., YONEYAMA, T., and ISHIZUKA, J. 1985: Effect of oxygen concentration and leghemoglobin on organic acid degradation by isolated soybean nodule bacteroids. Agric. BioL Chem., 49, 3473-3479
T AJlMA, S. and YAMAMOTO, Y. 1975: Enzymes of purine catabolism in soybean plants. Plant Cell Physiol., 16, 27 I -282
TARONI, F., GELLERA, c., and DONATO, S.D. 1988: Evidence for two distinct mitochondrial malic enzymes in human skeletal muscle: Purification and properties of the NAD(P)+ -dependent enzyme. Biochirn. Biophys. Acta, 916, 446-454
WATERS, J.K., KARR, D.B., and EMERICH, D.W. 1985: Malate dehydrogenase from Rhizobium japonicum 31\b-143 bacteroids and Glycine max root-nodule mitochondria. Biochemistry, 24, 6479-6486
Dow
nloa
ded
by [
Lau
rent
ian
Uni
vers
ity]
at 1
8:49
05
Oct
ober
201
4