preventive role of α-lipoic acid, curcumin and ginger …
TRANSCRIPT

1
PREVENTIVE ROLE OF α-LIPOIC ACID, CURCUMIN AND
GINGER IN INHIBITING MYOGLOBIN GLYCATION AND
THE FORMATION OF ADVANCED GLYCATION END
PRODUCTS (AGEs)
Hardik Ghelani (17601883) (BPharm, MPharm (Pharmacology)
A thesis submitted in fulfilment of the requirements for the degree of
Master of Science (Honours)
School of Science and Health,
University of Western Sydney
October, 2014

2
STATEMENT OF AUTHENTICITY
This thesis is submitted in fulfilment of the requirements for the postgraduate research
degree, Master of Science (Hon.), at the School of Science and Health, University of
Western Sydney. The work presented in this thesis is, to the best of my knowledge and
belief, original except as acknowledged in the text. I hereby declare that I have not
previously submitted this material, either in whole or part at any other institution.
_____________________
HARDIK GHELANI
Date: 3rd
October-2014
Place: Campbelltown, NSW, Australia

3
LIST OF PUBLICATIONS
Ghelani, H.S., Razmovski-Naumovski, V., Kyada, A.V., Parekh, H.S., Li, G.Q., &
Nammi, S. (2014) Myoglobin and Low Density Lipoprotein Glycation in Cardiac
Complications of Diabetes: Current Perspectives and Therapeutic Options. Indian
Journal of Medical Research (In Press).
Ghelani, H.S., Razmovski-Naumovski, & Nammi, S. (2014). Ginger inhibits
glucose and fructose-induced myoglobin glycation and advanced glycation end
products formation in vitro. Manuscript under preparation for submission to BMC
Complementary & Alternative Medicine.
Ghelani, H.S., Razmovski-Naumovski, & Nammi, S. (2014). α-lipoic acid, a new
atici-glycation agent inhibits glucose and fructose-induced myoglobin glycation in
vitro. Manuscript under preparation for submission to Basic & Clinical
Pharmacology & Toxicology.
Ghelani, H.S., Razmovski-Naumovski, & Nammi, S. (2014). Curcumin prevents
AGEs formation and protein oxidation against glucose and fructose-induced
myoglobin glycation in vitro. Manuscript under preparation for submission to
Journal of Pharmacy & Pharmacology.

4
AKNOWLEDGEMENTS
Each morning comes with new opportunities in the form of numerous rays of hope.
Choosing a single ray, I decided to carry out something novel which would prove to be
fruitful and beneficial to me and society. It would be a great reward for me if my work
proves to help an endless number of patients. But, it’s not at all possible to walk alone on
this path of research, and it requires a pool of genuine, authentic, brilliant, elegant hands to
be by my side and I take this opportunity to oblige honour and pay gratitude to one and all. I
pay my utmost gratitude to my supervisor, Dr Srinivas Nammi for being a constant
deliverer of hands in the hour of need. I owe him the full share for the successful completion
of my project. I am thankful to him for providing the liberations in correct manner, for
valuable suggestions, for constructive criticism, for all inspiration that he gave me. Without
his guidance, my project would not have seen the ending shores. I take this opportunity to
thank my co-supervisor, Dr Valentina Naumovski for her timely suggestions, generous and
friendly nature, persistent encouragement, critical remarks and counsel during the whole
course of this work. I would like to acknowledge the instructional and handy tips of Mr
Ashish Kyada at the right time. I owe a lot to him for his valuable suggestions, constant
moral support and timely help throughout this project. I sincerely wish to acknowledge the
invaluable technical support extended by the laboratory technicians of the School, Mrs
Kathleen Kyle for her constant help and providing the requirements at the right time for
carrying out the experiments. Finally, I would like to mention my family, especially my
parents and my cousin brother Mr Vipul Ghelani, who have given moral, emotional and
financial support, and have great trust on my academic career.
- HARDIK GHELANI

5
To
Science

6
Table of Contents Page No
Cover page……………………………………………………………………………………………..1
Statement of authentication……………………………………………………………………….......2
List of publications…………………………………………………………………………………….3
Acknowledgements...............................................................................................................................4
Table of contents...................................................................................................................................6
List of tables…………………………………………………………………………………………...9
List of figures………………………………………………………………………………………...10
General abbreviations.........................................................................................................................11
Summary.............................................................................................................................................12
CHAPTER – 1 INTRODUCTION……………………………………………........14
1.1 Pathophysiology of hyperglycaemia-induced biochemical alteration: Focus on advanced
glycation end products………………………………………………………………………...15
1.1.1 Non-enzymatic glycation and advanced glycation end-products ………………….17
1.1.2 Fructose-induced protein glycation…………………………………………….......21
1.1.3 Receptors for advanced glycation end products…………………………………….23
1.1.4 Advanced glycation end products and heart failure………………………………...23
1.1.5 Myoglobin and its physiological role in the heart ……………………………........25
1.1.6 Myoglobin glycation and its possible role in diabetic cardiovascular
complications……………………………………….................................................26
1.2 Therapeutics strategies to inhibit protein glycation and advanced glycation end products
formation………………………………………………………………………………………30
1.2.1 Glycaemic control…………………………………………………………………..30
1.2.2 Synthetic AGEs inhibitors………………………………………………………….31
1.2.3 Natural AGEs inhibitors………………………………………………………........33
1.2.4 Antioxidants………………………………………………………………………...35
1.3 Prospects of alternative medicine therapy…………………………………………………….37
1.3.1 Overview………………………………………………………………………........37
1.3.2 α-Lipoic acid…………………………………………………………………..........37
1.3.3 Curcumin……………………………………………………………………….......38
1.3.4 Ginger……………………………………………………………………………….40
1.4 Rationale and objectives...…………………………………………………………………….42

7
CHAPTER – 2 MATERIALS AND METHODS…………………………………..44
2.1 Chemicals………………………………………………………………………………........45
2.2 Equipment…………………………………………………………………………………...45
2.3 Preparation of reagents……………………………………………………………...............45
2.4 The test substances and dosage selection……………………………………………………49
2.5 Evaluation of myoglobin glycation inhibitory effect of α-lipoic acid, curcumin and ginger
under glucose/fructose overload in vitro…………………………………………………….51
2.5.1 Estimation of total fluorescent advanced glycation end products…………….51
2.5.2 Estimation of liberated free iron (ferrozine test)………………………………..52
2.5.3 Estimation of fructosamine (glycated myoglobin)……………………………...54
2.5.4 Estimation of protein carbonyls…………………………………………………55
2.5.5 Estimation of protein thiols………………………………………………........56
2.5.6 Data and statistical analysis…………………………………………………….57
CHAPTER – 3 RESULTS…………………………………………………………..58
3.1 Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-induced formation of
fluorescent advanced glycation end products……………………………………………........59
3.2 Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-mediated liberated free
iron…………………………………………………………………………………………….65
3.3 Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-induced fructosamine
formation………………………………………………………………....................................71
3.4 Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-induced protein
carbonyls formation ………………………………………......................................................77
3.5 Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-induced protein thiols
oxidation………………………………………........................................................................83
CHAPTER – 4 DISCUSSION………………………………………………….......89
4.1. Anti-glycation activity of α-lipoic acid……………………………………………………...93
4.2. Anti-glycation activity of curcumin…………………………………………………………96
4.3. Anti-glycation activity of ginger extract………………………………………………….....98

8
CHAPTER – 5 CONCLUSION AND FUTURE DIRECTION………..........................102
5.1. General conclusion……………………………………………………………………........103
5.2. Limitations…………………………………………………………………………….........104
5.3. Future direction…………………………………………………………………………….105
BIBLIOGRAPHY…………………………………………………………..........106

9
LIST OF TABLES
Table
No. Chapter Title of the table Page No.
1.1 1 Effects of oral hypoglycaemic agents in glycaemic control 31
1.2 1 Some important synthetic compounds/
phytoconstituents with anti-glycation activity 34
1.3 1 Mechanism of action and clinical significance of known natural
antioxidants 36
2.1 2 Pharmacological activities of the selected test compounds 50
3.1 3 Effect of α-lipoic acid, curcumin and ginger on the formation of
fluorescent AGEs in myoglobin-glucose glycation. 62
3.2 3 Effect of α-lipoic acid, curcumin and ginger on the formation of
fluorescent AGEs in myoglobin-fructose glycation. 63
3.3 3 Effect of α-lipoic acid, curcumin and ginger on free iron release
in myoglobin-glucose glycation 68
3.4 3 Effect of α-lipoic acid, curcumin and ginger on free iron release
in myoglobin-fructose glycation 69
3.5 3 Effects of α-lipoic acid, curcumin and ginger on fructosamine in
myoglobin-glucose glycation 74
3.6 3 Effects of α-lipoic acid, curcumin and ginger on fructosamine in
myoglobin-fructose glycation 75
3.7 3 Effects of α-lipoic acid, curcumin and ginger on carbonyl
content in myoglobin-glucose glycation 80
3.8 3 Effects of α-lipoic acid, curcumin and ginger on carbonyl
content in myoglobin-fructose glycation 81
3.9 3 Effects of α-lipoic acid, curcumin and ginger on the level of thiol
group in myoglobin-glucose glycation 86
3.10 3 Effects of α-lipoic acid, curcumin and ginger on the level of thiol
group in myoglobin-fructose glycation 87
10
LIST OF FIGURES
Fig
No. Chapter Title of the figures Page No.
1.1 1 Formation of AGEs through various biochemical steps 20
1.2 1 Mechanism of fructation for the production of reactive carbonyl
and oxygen species 22
1.3 1 Fructose autoxidation 22
1.4 1 Summary of the AGEs pathways which may cause heart failure. 26
1.5 1 Possible role of myoglobin glycation in diabetes-induced
myocardial damage 29
1.6 1 α-Lipoic acid 38
1.7 1 Curcumin 40
1.8 1 Major bio-active phytoconstituents of ginger 41
3.1 3
Percentage inhibition of fluorescent AGEs formation by α-lipoic
acid (A), curcumin (B) and ginger (C) in myoglobin-glucose
(Mb-Glu) and myoglobin-fructose (Mb-Fru) glycation after 30
days of incubation.
64
3.2 3
Percentage inhibition of free iron release by α-lipoic acid (A),
curcumin (B) and ginger (C) in myoglobin-glucose (Mb-Glu)
and myoglobin-fructose (Mb-Fru) glycation after 30 days of
incubation.
70
3.3 3
Percentage inhibition of fructosamine formation by α-lipoic acid
(A), curcumin (B) and ginger (C) in myoglobin-glucose (Mb-
Glu) and myoglobin-fructose (Mb-Fru) glycation after 30 days of
incubation.
76
3.4 3
Percentage inhibition of protein carbonyl content by α-lipoic acid
(A), curcumin (B) and ginger (C) in myoglobin-glucose (Mb-
Glu) and myoglobin-fructose (Mb-Fru) glycation after 30 days of
incubation.
82
3.5 3
Percentage protection of protein thiol by α-lipoic acid (A),
curcumin (B) and ginger (C) in myoglobin-glucose (Mb-Glu)
and myoglobin-fructose (Mb-Fru) glycation after 30 days of
incubation.
88
4.1 4
Schematic representations of pathways induced by myoglobin
glycation and the inhibitory effects of α-lipoic acid, curcumin
and ginger extract on this pathway
101

11
GENERAL ABBREVIATIONS
Abbreviation Full term
AG Aminoguanidine
AGEs Advanced glycation end products
ALA Alpha-lipoic acid
ANF Anti-nuclear antibodies
CML Carboxymethyllysine
DM Diabetes mellitus
DNHP Dinitrophenylhydrazine
ELISA Enzyme-linked immunosorbent assay
HRP Horseradish peroxidise
LDL Low-density lipoproteins
Mb Myoglobin
MI Myocardial infarction
NBT Nitro-blue tetrazolium
RAGE Receptor for advanced glycation end products
ROS Reactive oxygen species
TCA Trichloro acetic acid
TGF Transforming growth factor

12
SUMMARY
Diabetes mellitus (DM) is a metabolic disorder characterised by constant high blood glucose
(hyperglycaemia) due to an absolute or relative deficiency of insulin and/or insulin
resistance. The precise role of hyperglycaemia in the pathogenesis of long-term
complications of diabetes is still unclear. However, one pathway that has received
considerable interest in the development of diabetic complications is protein glycation.
CHAPTER ONE reviewed the chemistry and pathophysiological implications of
protein glycation and the formation of AGEs. In most studies, glucose-induced protein
glycation and subsequent AGEs formation has been emphasised (Ahmed, N. 2005).
However, fructose-induced glycation (also known as fructation), is also possible and is
considered a highly probable event in hyperglycaemic conditions (Schalkwijk, Stehouwer &
van Hinsbergh 2004). The possible role of myoglobin glycation in pathological implication
of diabetes-induced myocardial injury and the development of myocardial infarction (MI) is
also reviewed in Chapter one. The glycation of myoglobin under chronic hyperglycaemia
compromises its functions which lead to ischaemia and MI (Kyada 2012). Many
pharmacological agents have been developed to reduce the implications of hyperglycaemia
on diabetic complications in both humans and in experimental models however, there are no
anti-glycation therapeutic agents. Many natural products are being sold on the market and
have been shown to be relatively safe for human consumption. A number of naturally-
derived compounds have shown hypoglycaemic, hypolipidaemic, as well as antioxidant
properties (Vasu et al. 2005). However, based on previous research in our laboratory, three
different natural products including α-lipoic acid (ALA), curcumin and an ethanolic extract
of ginger have been selected to evaluate their anti-glycation activity on myoglobin protein.

13
CHAPTER TWO of the thesis describes the methodology for evaluating the anti-glycation
effect of α-lipoic acid, curcumin and an ethanolic extract of ginger. In this study, myoglobin
was incubated for 30 days with high glucose or fructose in the absence or presence of
various concentrations of α-lipoic acid, curcumin or ethanolic extract of ginger. Aliquots of
incubation mixture were taken after each specified period of time (on day-10, day-20 and
day-30) and assayed for various parameters such as the fluorescent AGEs, the liberated free
iron, the Amadori products (fructosamine), the protein carbonyls and the protein thiols.
CHAPTER THREE describes the results of the anti-glycation activity of α-lipoic
acid, curcumin and ginger extract. The results indicate that all the three tested natural
products showed marked reduction in myoglobin glycation in terms of inhibiting the
formation of fluorescent AGEs, reduced levels of fructosamine adducts formation and
reduced free iron release. Furthermore, the test substances significantly prevented protein
oxidative damages, including effects on thiol and protein carbonyl oxidation.
In CHAPTER FOUR, it is proposed that the therapeutic intervention with natural
products possessing multiple pharmacological activities associated with the multi-faceted
complications of diabetes are capable of blocking or reversing the pathological progresses of
myoglobin glycation and subsequently, AGEs formation.
In conclusion, CHAPTER FIVE presents the outcomes of the proposed study and
provides evidence for the protective role of α-lipoic acid, curcumin and ginger extract on
myoglobin glycation, particularly in association with the hyperglycaemic condition. This
study provides further insight for improving the clinical management of diabetes-induced
complications such as MI.

14
CHAPTER-1
INTRODUCTION

15
1.1. Pathophysiology of hyperglycaemia-induced biochemical alterations: focus on
advanced glycation end products
Diabetes mellitus is a significant medical problem and emerging as a major epidemic
worldwide, affecting 1–2% of the population. Diabetic patients are prone to long-term micro
and macrovascular complications such as cardiomyopathy, atherosclerosis, retinopathy,
cataract, neuropathy and nephropathy. As a consequence, their life expectancy is only two-
thirds of that of the general population (Ahmed, N. 2005).
Hyperglycaemia has an important role in the pathogenesis of long-term
complications and diabetic patients with poor blood glucose control are particularly at risk
(Anonymous 1998). Various mechanisms have been studied which show the role of
hypoglycaemia in the pathogenesis of diabetic complications. The first such mechanism that
was discovered was the polyol pathway and increased polyol pathway flux. Physiologically,
aldose reductase has the function of reducing toxic aldehydes in the cell to inactive alcohols.
However, when the glucose concentration in the cell becomes too high (such as in
hyperglycaemic condition), aldose reductase reduces that excess glucose to sorbitol, which
is later oxidised to fructose. In the process of reducing high intracellular glucose to sorbitol,
the aldose reductase consumes the co-factor NADPH (Lee, AY & Chung 1999). NADPH is
an essential co-factor for regenerating a critical intracellular antioxidant such as reduced
glutathione. The reduction in the amount of NADPH and thereby reduced glutathione, the
polyol pathway increases susceptibility to intracellular oxidative stress which is major
pathobiological factor in development of diabetes complications (Brownlee, Michael 2005).
The second mechanism identified is the activation of protein kinase C (PKC)
pathway. Hyperglycaemia increases the synthesis of diacylglycerol intracellularly, which is
a critical activating co-factor for the protein kinase-C (Koya & King 1998). When PKC is

16
activated by intracellular hyperglycaemia, it has a variety of effects on gene expression. In
each case, the things that are “good” for normal function are decreased and the things that
are “bad” are increased. For example, the vasodilator producing endothelial nitric oxide
synthase is decreased, while the vasoconstrictor endothelin-1 is increased (Koya et al. 1997;
Xia et al. 1994).
The third mechanism is the activation of the hexosamine pathway. When glucose is
high inside a cell, most of that glucose is metabolised through glycolysis and converted into
glucose-6 phosphate and fructose-6 phosphate. However, some of that fructose-6-phosphate
is converted to glucosamine-6 phosphate and finally to N-acetyl glucosamine. The
production of N-acetyl glucosamine alters with transcription factors and often results in
pathological changes in gene expression (Kolm-Litty et al. 1998; Sayeski & Kudlow 1996).
The last mechanism is the production of advanced glycation end products through
protein glycation reaction. The high blood glucose reacts with intracellular protein through
the Millard reaction and generates various heterogeneous AGEs. AGEs bind to AGE
receptors and activate them, thereby causing the production of inflammatory cytokines and
growth factors, which in turn cause vascular pathology (Ahmed, N. 2005). Moreover, AGEs
can diffuse out of the cell and modify extracellular matrix molecules nearby, which changes
signalling between the matrix and the cell and causes cellular dysfunction (Smit & Lutgers
2004).
However, each one is an important mechanism in development of diabetic
complications but an attractive mechanism that has received considerable interest is
production of AGEs through protein glycation reaction and its pathophysiological
implications in diabetic complication.

17
1.1.1. Non-enzymatic glycation and advanced glycation end-products
Glucose in the blood is known to react with vital proteins, such as serum albumin
(Schleicher et al. 1993), α-crystallin (Biemel, Friedl & Lederer 2002), collagen (Turk et al.
1999), low-density lipoprotein (Stewart et al. 1994) and haemoglobin (De Rosa et al. 1998),
resulting in chemical modifications to proteins. Such chemical modifications are generally
known as a protein glycation and are more significant in diabetes with persistent
hyperglycaemia (Roy, Sen & Chakraborti 2004).
Protein glycation is initiated by a nucleophilic addition reaction between a free
amino group of a protein and a carbonyl group of a reducing sugar (i.e. glucose, fructose) to
form a freely reversible Schiff’s base via the Maillard reaction (Maillard 1912). This
reaction occurs over a period of hours, and once formed, the labile Schiff’s base rearranges
to a more stable Amadori products (fructosamine). The formation of Amadori products
occurs over a period of days to weeks and, once formed, is practically irreversible. Protein
glycation is a spontaneous reaction and is dependent on the degree and duration of
hyperglycaemia, the half-life of the protein and tissue permeability of free glucose
(Brownlee, M., Cerami & Vlassara 1988). Amadori products (intermediate glycated
products) undergo further rearrangement giving rise to poorly characterised structures called
AGEs (Figure 1.1).
The other mechanism such as autoxidation of glucose and glyoxidation of Amadori
products also involve in AGEs formation. Physiologically, monosaccharides, like glucose,
exist in equilibrium with their enediol, which can oxidise in the presence of transition metals
to form an enediol radical. Furthermore, this enediol oxidised itself to dicarbonyl
compounds which can react with protein amino groups and participate in AGEs formation

18
(Wolff & Dean 1987). This is referred to as autoxidative glycation and is outlined in Figure
1.1. The term glycoxidation is used to describe autoxidation of Amadori products to AGEs
as shown in Figure 1.1. The Amadori products are converted to protein dicarbonyl
compounds which can further participate in AGEs formation and are referred to as
glycoxidation products (Hunt, Bottoms & Mitchinson 1993). The increase in the
concentration of reactive protein carbonyl compounds from autoxidation of glucose and
glycoxidation will lead to carbonyl stress, a situation that aggravates the modification of
proteins and stimulates the formation of AGEs (Miyata 2002; Miyata et al. 1999). AGEs
have been implicated in the development of diabetic complications (Candido et al. 2003) as
they have a tendency to generate damaging reactive oxygen species (ROS) and
subsequently, damage the cellular and extracellular organelles (Ahmed, N. 2005; Bonnefont-
Rousselot 2002; Skovsted et al. 1998).
AGEs are complex and heterogeneous molecules which affect many intracellular
functions through signalling pathways and through extracellular protein cross-linkage
(Ahmed, N. 2005). However, the detailed mechanism underlying AGEs formation, their
characterisation and their role in diabetic complications largely remain unclear (Giardino,
Edelstein & Brownlee 1994). Over the last decade, a number of AGEs have been detected in
tissues and can be divided into three categories as outlined below:
1. Fluorescent cross-linking AGEs such as pentosidine, crossline and glyoxal lysine
dimer (GOLD) or methylglyoxal lysine dimer (MOLD).
2. Non-fluorescent cross-linking AGEs such as imidazolium dilysine cross-links,
alkyl formyl glycosyl pyrrole (AFGP) cross-links and arginine– lysine imidazole
(ALI) cross-links.
3. Non-cross-linking AGEs such as pyrraline and N-carboxymethyllysine (CML).

19
Pentosidine, a well-defined fluorescent AGEs, has been shown to be strongly associated
with diabetic complications, indicating a link between AGEs formation and the development
of diabetic cardiomyopathy, retinopathy and nephropathy (Kerkeni et al. 2013). Previous
studies have reported that serum pentosidine was significantly higher in patients with
diabetes than in subjects without diabetes, and this was associated with an increased
incidence of cardiovascular disease (Sugiyama et al. 1998; Weiss et al. 1998). Moreover, in
a prospective clinical study, Yoshida et al demonstrated that serum pentosidine levels were
significantly higher in diabetic patients with cardiovascular disease than in those without,
and this correlated with increased arterial wall stiffness in diabetic patients (Yoshida,
Okumura & Aso 2005).
GOLD and MOLD are also fluorescent cross-linking AGEs that cross-link with
lysine residues of extracellular proteins (i.e. collagen) leading to stiffening of the extra
cellular matrix. The occurrence GOLD or MOLD results in stiffening of the extracellular
matrix which often compromises organ function and is associated with several chronic
diseases such as diabetes, vascular diseases, retinopathy, arthritis and Alzheimer´s syndrome
(Ahmed, N. 2005). The formation of GOLD on the vascular wall causes the cross-linking of
myocardial collagen molecules to each other. This leads to the loss of collagen elasticity and
subsequently causes arterial stiffening and myocardial diastolic dysfunction which results in
diastolic heart failure in diabetic conditions (Frye et al. 1998).
Non-fluorescent AGEs such as alkyl formyl glycosyl pyrroles and arginine–lysine
imidazole are formed by the reaction between two sugar molecules with a single lysine
residue but are of limited significance in vivo (Farmar, Ulrich & Cerami 1988) (Al-Abed &
Bucala 2000)

20
Pyrraline is a non-cross-linking AGE detectable in human skin, plasma and in brain plaques
(Smith et al. 1994). CML is another non-cross-linking AGE that is formed by the oxidative
breakdown of Amadori products and during metal-catalysed oxidation of polyunsaturated
fatty acids in the presence of protein. CML is a major AGEs formed in vivo (Reddy et al.
1995) and its levels are found to be increased two-fold in the diabetic subject (Dyer et al.
1993).
Figure 1.1 Formation of AGEs through various biochemical steps
In the first step of the Maillard reaction, a sugar adduct such as glucose reacts with a protein amino
(NH2) group to form a Schiff’s base. The Schiff’s base then converts into more stable Amadori
products. The subsequent re-arrangement of Amadori products leads to the formation of stable and
irreversible AGEs compounds. Glucose autoxidation and glycoxidation increase concentration of
carbonyl/dicarbonyl compounds which further participate in AGEs formation.

21
1.1.2. Fructose-induced protein glycation
Fructose is a reducing monosaccharide, a common component of honey, fruit juice
concentrates, table sugar and high-fructose corn syrup. It has been excessively consumed in
human diets over the last decades, despite the evidence implicating fructose in the
development of metabolic disorders (Bray 2013; Dekker et al. 2010). Compared to glucose,
glycation by fructose (fructation) has not been as thoroughly investigated.
As for glucose-mediated protein glycation (glucation), the initial step of fructation is
the covalent interaction between free carbonyl group of open-chained fructose and amino
groups of protein producing the Schiff’s base (Figure 1.2). The latter is an unstable
compound that is subjected to further isomerisation and forms a more stable Heyns adducts.
The Heyns compounds as well as Amadori products derived from glucation are known as
“early glycation products” or “fructosamine.” The fructose moiety of the Heyns products
undergoes enolisation, followed by dehydration, oxidation and/or fragmentation reactions,
which continually produces a variety of carbonyl compounds (Reihl et al. 2004; Tessier
2010). Like other early glycation products, the Heyns compounds may undergo autoxidation
leading to the formation of reactive carbonyl compounds and ROS (Hunt, Dean & Wolff
1988; Mullarkey, Edelstein & Brownlee 1990). Figure 1.3 demonstrates the mechanism of
fructose autoxidation.
Collectively, fructose is more potent than glucose in protein glycation and AGEs
formation. Thus, fructose-induced glycation may play a significant role in the long-term
complications of diabetes

22
Figure 1.2 Mechanism of fructation for the production of reactive carbonyl and oxygen
species (Edelstein & Brownlee 1990)
Figure 1.3 Fructose autoxidation (Edelstein & Brownlee 1990)

23
1.1.3. Receptors for advanced glycation end products
The expression of AGEs receptors has been demonstrated in various cell-types, including
endothelial cells, monocytes, macrophages and cardiac myocytes (Brett et al. 1993). The
most important AGEs receptor that has been identified is RAGE, a multi-ligand receptor of
the immunoglobulin super family of receptors. The activation of RAGE stimulates second
messenger pathways (Ras, Rac–Cdc42, and Jac–Stat pathways) and the production of ROS
via the NADPH oxidase pathway (Goldin et al. 2006; Hartog et al. 2004). In turn, these
secondary messengers activate or prolong the activation of nuclear factor kappa-B (NF-κB)
transcription factor that subsequently up-regulates the production of endothelin-1, vascular
cell adhesion molecule-1 (VCAM-1), intercellular adhesion molecule-1 (ICAM-1), E-
selectin, plasminogen activator inhibitor-1 (PAI-1), tissue factor and transforming growth
factor-β (TGF-β) (Tanaka et al. 2000).
Besides RAGE, there are other AGE receptors such as macrophage scavenger
receptor Type I and II (SR-A), oligosaccharyl transferase-48 (OST-48), galectin-3, lectin-
like oxidised low density lipoprotein receptor-1 (LOX-1) and cluster of differentiation 36
(CD36) (Chavakis, Bierhaus & Nawroth 2004; Goh & Cooper 2008) which are believed to
bind with AGEs. However, these receptors, unlike RAGE, play a role in the removal of
AGEs rather than in signal transduction.
1.1.4. Advanced glycation end products and heart failure
Heart failure is characterised by a structural or functional cardiac disorder that results in the
incapacity of the heart to fill with or pump out blood. Hartog et al has proposed two major
pathways emphasising the role of AGEs in the development of heart failure (Hartog et al.

24
2007). The first pathway describes the detrimental effect of AGEs on the physiological
properties of proteins in the extracellular matrix by creating cross-links, while the second
pathway shows interaction of AGEs with various AGEs receptors. AGEs may induce
diastolic and systolic dysfunction through these pathways. Subsequently, these abnormalities
may result in the development and progression of heart failure. A summary of these
pathways is presented in Figure 1.2.
The cross-linking of extracellular matrix proteins such as collagen is essential for
physiological functions. It strengthens tissue integrity, without compromising flexibility.
AGEs can covalently bind other AGEs, and form additional cross-links between matrix
proteins like collagen, laminin and elastin (Smit & Lutgers 2004). Excessive cross-linking
caused by AGEs accumulation has been shown to compromise the flexibility of matrix
proteins and increased myocardial stiffness and rigidity which may induce diastolic
dysfunction in the heart (Smit & Lutgers 2004).
Another pathway by which AGEs may contribute to the development of diastolic
dysfunction is via the activation of AGEs receptors. AGE-receptor activation seems to
influence calcium metabolism in cardiac myocytes by delaying calcium reuptake. As a
consequence, the duration of the re-polarisation phase of the cardiac contraction and
relaxation may increase, subsequently causing diastolic/systolic dysfunction (Petrova et al.
2002). AGE-receptor interaction may also induce atherosclerosis and subsequent MI by
modifying low-density lipoprotein (LDL) (Bucala et al. 1994). AGE modified LDL is more
susceptible to macrophage uptake by AGE receptors, creating foam cells which are the hall
mark of the development of atherosclerosis and MI (Witztum & Steinberg 1991).

25
The harmful effects of AGEs are not restricted to specific diabetic patient. Patients with
renal failure are also known to have increased AGEs accumulation. Patients with these
conditions also suffer from an increased prevalence of heart failure (Foley et al. 2005).
AGEs accumulation increases morbidity and mortality rate in patients with diabetes mellitus
and patients with renal failure. This may possibly contribute to the increased prevalence of
heart failure in these conditions (Meerwaldt et al. 2005). Therefore, patients with diabetes
mellitus or renal failure may particularly benefit from intervention with compound(s)
possessing anti-AGEs property.

26
Figure 1.4. Summary of the AGEs pathways which may cause heart failure (Hartog et
al. 2007)
AGE: advanced glycation end-products

27
1.1.5. Myoglobin and its physiological role in the heart
Myoglobin is a cytoplasmic haemoprotein expressed solely in striated muscle (cardiac
myocardium and skeletal muscle fibres) (Ordway & Garry 2004). The diverse functional
roles of myoglobin include buffering intracellular oxygen concentration, facilitating
intracellular oxygen transport, inactivating nitric oxide (NO) and scavenging ROS
(Kanatous et al. 2009). In cardiomyocytes, myoglobin acts as short-time oxygen storage
protein. Iron bound in the heme pocket of myoglobin reversibly binds to oxygen and serves
as an oxygen reservoir. Under certain conditions like hypoxia or in diminished cardiac flow
due to systolic compression, myoglobin releases its bound oxygen for cardiac mitochondrial
oxidative phosphorylation to synthesise ATP. However, under what circumstances the heart
requires myoglobin-bound oxygen for oxidative phosphorylation is currently a subject of
debate (Hendgen-Cotta, Kelm & Rassaf 2014; Roy, Sen & Chakraborti 2004).
Although the primary function of myoglobin has been considered to be cellular
oxygen storage and supply, studies have shown that myoglobin also acts as an intracellular
scavenger of bioactive nitric oxide, regulating its level in the cardiac and skeletal muscle and
thereby, protecting mitochondrial respiration, which is impaired by NO (Brunori 2001;
Flogel et al. 2001; Wunderlich et al. 2003). Flogel and colleagues demonstrated that
myoglobin significantly contributes to the attenuation of oxidative stress in cardiac muscle.
On the contrary, it has been demonstrated that either pharmacological inhibition or genetic
deletion of myoglobin leads to increased vulnerability of cardiac function to oxidative
challenge. Thus, myoglobin is a key element influencing redox pathways in cardiac muscle,
aiding the functional and metabolic protection of the heart from oxidative damage (Flogel et
al. 2004). MI occurs when oxygen supply is reduced to the heart, which eventually causes

28
necrosis of myocardial tissue and release cytoplasmic content into systemic circulation. As
myoglobin is found in cardiomyocytes, it is released into the serum as early as one hour after
MI (heart attack) and so also serves as a cardiac biomarker. (Mercer 1997; Vaidya 1994).
1.1.6. Myoglobin glycation and its possible implications in diabetic cardiovascular
complications
Roy and co-workers have extensively studied glycation-induced structural and functional
modification of myoglobin in vitro in hyperglycaemic condition The authors demonstrated
that glycated myoglobin exhibits structural changes and that it may be a source of free
radicals and oxidative stress in uncontrolled diabetes (Roy, Sen & Chakraborti 2004).
During glycation, iron is liberated from the heme moiety and ligated to another moiety, most
likely distal histidine, in heme pocket. This iron has been termed “mobile reactive iron”,
which can catalyse the Haber–Weiss reaction producing free radicals, particularly hydroxyl
(OH) radicals, and in turn, may damage different cell constituents (Roy, Sen & Chakraborti
2004; Sen et al. 2007). Apart from the iron-mediated oxidative stress, glycation also
modulates peroxidase activity of myoglobin in different ways (Roy, Sil & Chakraborti
2010). During protein glycation, the measurement of protein carbonyl content is the most
general and commonly used biomarker of long-term carbonyl overload or ‘‘carbonyl stress’’
(Dalle-Donne et al. 2003). Glycation-induced and metal-catalysed oxidation may cause
covalent modification of heme proteins by introducing carbonyl groups into amino acid
residues of proteins. Since myoglobin is also a heme protein, it is also susceptible to
carbonyl formation in glycated forms and subsequently, induces carbonyl stress (Roy, Sen &
Chakraborti 2004). Such oxidative modification is an index of oxidative stress and may play

29
a significant role in the pathological processes leading to the complications of diabetes
(Webster et al. 2005).
From the above in vitro studies, it has been confirmed that the glycation of
myoglobin under high glucose environment modifies its structure and may compromise its
functions. Thus, free myoglobin in circulation, if glycated, particularly in uncontrolled
diabetes, may cause a serious threat with respect to oxidative damage. This may, in turn,
aggravate cardiac complications such as ischaemia and MI (Figure 1.5).
Figure 1.5 Possible role of myoglobin glycation in diabetes-induced myocardial damage
Excessive blood glucose may react with myoglobin (Mb) via the Maillard reaction and generate AGEs, which
further potentiates free radical generation via carbonyl compound formation and release of ferrous iron. Mb
glycation in cardiomyocytes may also reduce mitochondrial oxygen delivery and alter ATP phosphorylation.

30
1.2. Therapeutics strategies to inhibit protein glycation and advanced glycation end
products formation
Several anti-AGE intervention strategies have been proposed with the aim of controlling
hyperglycaemia, scavenging ROS, inhibiting and scavenging AGEs, as well as controlling
the consumption of exogenous AGEs.
1.2.1. Glycaemic control
Epidemiological data demonstrates that there is a clear relationship between diabetes and
cardiovascular disease. For newly diagnosed patients of diabetes, glycaemic control with
suitable oral hypoglycaemic agents is the first line approach. However, long term steady
glycaemic control is necessary to circumvent a decline in cardiovascular complications. The
most logical and promising approach for better glycaemic control and thus, improving the
cardiovascular prognosis of diabetes patients would be to use therapies that:
Improve glycaemic control without leading to hypoglycaemia
Effectively lower postprandial glucose excursions (or lower both fasting and
postprandial glucose)
Lower insulin resistance
In prospective trials, therapeutic agents possessing these three features have shown great
promise in reducing the cardiac complications of diabetes (O'Keefe et al. 2011). Examples
of such therapies include metformin, acarbose, bile acid sequestrants, incretinmimetics and
dipeptidylpeptidase-4 inhibitors. The effects of various oral hypoglycaemic drugs on
postprandial glucose, insulin resistance and hypoglycaemia are depicted in Table 1.1.
Pioglitazone, belonging to the thiazolidinedione class of agents used to manage diabetes, has
shown particular benefit in preventing cardiovascular complications. However, this benefit

31
has been overshadowed by its tendency to cause fluid retention, weight gain and reduction in
bone density to the extent of causing fractures (Habib et al. 2010).
Table 1.1.Effect of oral hypoglycaemic agents in glycaemic control
Drug Class
Reducing
postprandial
glucose
Reducing
insulin
resistance
Avoiding
hypoglycaemia
Sulfonylureas + - -
Metformin - + +
Thiazolidinediones + + +
Incretin-based therapies + - +
Bile acid sequestrates + - +
α-Glucosidase inhibitors + - +
1.2.2. Synthetic AGEs inhibitors
Over the last decade, a plethora of compounds have been screened for their AGE-inhibitory
activity, with only a handful progressing to the clinical trials. The first compound which was
extensively studied and found to be a powerful inhibitor of AGE formation was
aminoguanidine (AG) (Brownlee, M., Cerami & Vlassara 1988). Initial studies revealed that
AG displayed inhibitory activity towards the cross-linking of aortic collagen in diabetic rats.
Later, detailed investigation demonstrated that AG inhibited the development of other
diabetic complications such as nephropathy, neuropathy and vasculopathy (Goh & Cooper
2008). Additionally, AG imparted hypolipidaemic and hypoglycaemic actions in diabetic
rats; this was consistent with the effects of AG on dyslipidaemia in humans (Brownlee, M.
1994).

32
During the clinical development of AG, serious adverse events such as pernicious anaemia
and the development of anti-nuclear antibodies (ANF), particularly when AG’s were
administered in high doses, have been reported (Nilsson 1999). Furthermore, pancreatic and
renal-neoplastic tumours were reported in diabetic rats treated with AG (Boel et al. 1995).
Due to the severity of adverse effects, such clinical trials with AG were halted prematurely
due to increased safety concerns. Apart from AG, molecules which have been investigated
include pyridoxamine, OPB-9195 and LR-90 (Goh & Cooper 2008). Pyridoxamine, a
derivative of vitamin B6, has been reported to reduce hyperlipidaemia and prevent AGE
formation in experimental induced animal models. The compound controls the degradation
of protein-Amadori intermediates to protein-AGEs (Degenhardt et al. 2002; Stitt et al. 2002)
and antagonises angiotensin II-induced elevation in serum and renal-derived AGEs (Fioretto
et al. 1998). OPB-9195 is a thiazolidine derivative which has been shown to prevent the
progression of glomerular sclerosis and reduced urinary albumin excretion in association
with decreases in renal transforming growth factor (TGF)-β and vascular endothelial growth
factor (VEGF) expression (Nakamura et al. 1997). However, it has not been tested in
humans, with no mention for planned clinical trials.
Methylene bis-4, 4’-(-2 chloro phenyl ureido-phenoxy iso butyric acid) (LR-90) has
been investigated in a number of animal studies (Figarola et al. 2003). LR-90 inhibited
albuminuria and reduced serum creatinine concentrations, as well as circulating AGEs
levels, in diabetic rats without any effect on glycaemic control. LR-90 prevented
glomerulosclerosis and collagen deposition in association with reduced glomerular AGEs
accumulation. The effect of LR-90 is currently being tested on macrovascular complications

33
in vivo. Interestingly, LR-90 has been shown to inhibit S100b-induced expression of RAGE
and other pro-inflammatory genes in human monocytes (Figarola et al. 2007).
1.2.3. Natural AGEs inhibitors
There has been an accelerated interest in drugs and dietary supplements derived from plants
in recent years. There are numerous crude plant extracts or isolated phyto-constituents
reported to possess anti-glycation or AGEs-crosslink breaking effects. Literature review
demonstrates that phytochemicals isolated from common herbs (e.g. garcinol, quercetin),
spices (e.g. curcumin and ginger), dietary plants (i.e. green tea) and nutraceuticals (e.g. α-
lipoic acid) possess significant anti-glycation properties.
The most popular example of a plant-based product with global health promoting properties
is green tea, derived from the leaves of Camellia sinensis. Green tea is a rich source of
tannins (flavonoids) from which its antioxidant properties are derived. There is now
increasing evidence to suggest that tannins in green tea possess anti-glycation properties
(Nakagawa et al. 2002). In another study, the water-soluble fraction of tomato paste,
containing the flavonoid rutin, inhibited the formation of AGEs and proved to be more
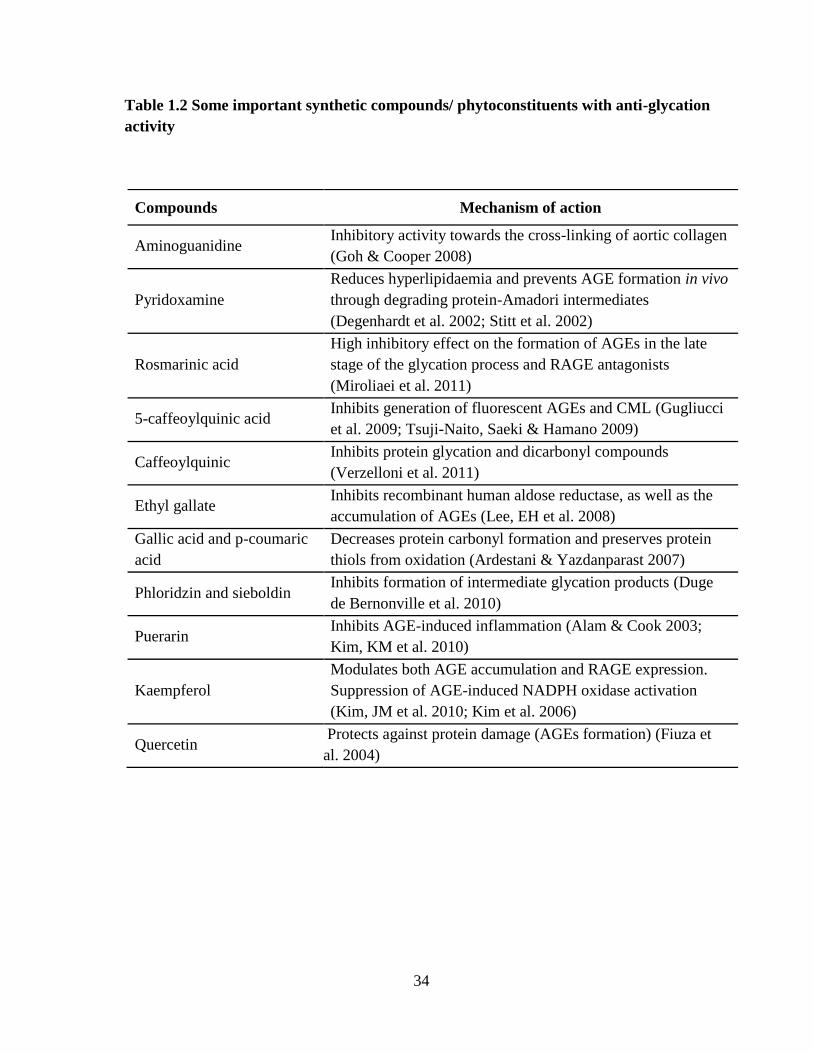
potent than aminoguanidine (Kiho et al. 2004). Table-1.2 lists some important synthetic
compounds/active phyto-constituents with potential anti-glycation activity.
34
Table 1.2 Some important synthetic compounds/ phytoconstituents with anti-glycation
activity
Compounds Mechanism of action
Aminoguanidine Inhibitory activity towards the cross-linking of aortic collagen
(Goh & Cooper 2008)
Pyridoxamine
Reduces hyperlipidaemia and prevents AGE formation in vivo
through degrading protein-Amadori intermediates
(Degenhardt et al. 2002; Stitt et al. 2002)
Rosmarinic acid
High inhibitory effect on the formation of AGEs in the late
stage of the glycation process and RAGE antagonists
(Miroliaei et al. 2011)
5-caffeoylquinic acid Inhibits generation of fluorescent AGEs and CML (Gugliucci
et al. 2009; Tsuji-Naito, Saeki & Hamano 2009)
Caffeoylquinic Inhibits protein glycation and dicarbonyl compounds
(Verzelloni et al. 2011)
Ethyl gallate Inhibits recombinant human aldose reductase, as well as the
accumulation of AGEs (Lee, EH et al. 2008)
Gallic acid and p-coumaric
acid
Decreases protein carbonyl formation and preserves protein
thiols from oxidation (Ardestani & Yazdanparast 2007)
Phloridzin and sieboldin Inhibits formation of intermediate glycation products (Duge
de Bernonville et al. 2010)
Puerarin Inhibits AGE-induced inflammation (Alam & Cook 2003;
Kim, KM et al. 2010)
Kaempferol
Modulates both AGE accumulation and RAGE expression.
Suppression of AGE-induced NADPH oxidase activation
(Kim, JM et al. 2010; Kim et al. 2006)
Quercetin Protects against protein damage (AGEs formation) (Fiuza et
al. 2004)

35
1.2.4. Antioxidants
Oxidative stress is known to potentiate AGEs formation, which in turn, induces further
oxidative stress with a continuum of the cycle (Yamagishi & Matsui 2010). The circulating
levels of oxidisable substrates such as Amadori products, reactive carbony/dicarbonyl-
substrates and polyunsaturated fatty acids are elevated in hyperglycaemia. Herbal
supplements with known antioxidant activity have been proposed as complementary
treatment for the management of protein glycation and may be a prospective first line
therapy to prevent AGEs formation. Epigallocatechin gallate and alpha-lipoic acid, well-
known natural antioxidants, significantly accelerated diabetic cutaneous wound healing
through angiogenesis regulation, anti-inflammatory effects and the inhibition of RAGE
(Chen, SA et al. 2012). Crocetin from the Indian and Chinese herbs Crocus sativus (Saffron)
and Gardenia jasminoides inhibited leukocyte adherence to bovine endothelial cells induced
by AGEs through the up-regulation of the activity of antioxidant enzymes in vitro (Xiang et
al. 2006). Garcinol, isolated from Garcinia indica fruit rind, has been shown to possess
antioxidant properties in an in vitro setting. Reports have suggested that garcinol is more
effective than aminoguanidine in terms of inhibiting AGEs (Yamaguchi et al. 2000). Table
1.3 describes some important natural antioxidants and their mechanism(s) of action.

36
Table 1.3: Mechanism of action and clinical significance for known natural
antioxidants
Antioxidant Mechanism of action Clinical significance
Chatechins Enhance activity of SOD and catalase (Jono et
al. 2002)
Inhibits ROS generation
Carotenoids Act as physical quenchers of ROS (Forbes et
al. 2004)
Prevent tissue
damage
α-Tocopherol Scavenger of lipid peroxy radicals (Bousova
et al. 2005)
Prevent AGEs
formation
Phenolics Inhibitor of oxidation of lipids and proteins
(Ames 1998)
Prevent AGEs formation
Tannins
Enhance synthesis of nitric oxide and relax
vascular segments (Hardy, Parmentier &
Fanni 1999)
Enhance blood
flow towards
vital organ
Glutathione Removes H2O2 and lipid peroxidase (Sekhar et
al. 2011)
Prevents lipid glycation-
induced AGEs formation
Taurine
Acts as an anti-glycative compound, providing
free amino groups that may compete for the
reducing sugars (Devamanoharan, Ali &
Varma 1997; Wright et al. 1986)
Prevent AGEs
formation
Spermine Scavenges free radicals (Ha et al. 1998) Inhibits ROS generation

37
1.3. Prospects of complementary medicine therapy
1.3.1. Overview
In the search for new medicines, the world is moving towards natural product-based
therapeutic options. Plants have been used for medicinal purposes in the traditional
medicinal systems of India, China, Egypt and Africa long before the modern-day recorded
scientific literature came into existence. Relative to pharmaceutical drugs, herbal medicines
are well tolerated, cost effective, have fewer side effects, and may be safer to use over time.
One particular advantage of natural products is their multi-targeted mode of action in
tackling complex diseases. A number of natural products such as α-lipoic acid, ginger and
turmeric have entered the international pharmacopoeia via the ethnopharmacological studies
of traditional medicines.
1.3.2. α-Lipoic acid
α-Lipoic acid (thiotic acid), or 1, 2-dithiolane-3-pentanoic acid, is a naturally occurring
dithiol compound synthesised enzymatically in the mitochondria from octanoic acid. α-
Lipoic acid is a necessary cofactor for mitochondrial α-ketoacid dehydrogenases and thus,
has a critical role in mitochondrial energy metabolism. α-Lipoic acid has been described as
a potent biological antioxidant, a detoxification agent and a diabetes medicine; it has been
used to improve age-associated cardiovascular, cognitive and neuromuscular deficits, and
has been implicated as a modulator of various inflammatory signalling pathways (Shay et al.
2009). It is an inducer of cellular signalling pathways, insulin mimetic, a
hypotriglyceridaemic agent, a vasorelaxant/anti-hypertensive compound, a metal chelator
and an adjuvant for neuro-cognitive function. In this regard, α-lipoic acid stimulates 5'
AMP-activated protein kinase (AMPK) dependent anorectic effect in rodents (Federici et al.

38
2002). The AGE production suppression and intermediate production suppression by α-
lipoic acid was also examined using an in vitro experimental model of glycation reactions by
using bovine serum albumin protein. The results showed that α-lipoic acid possesses
fluorescent AGE production inhibitory activity (Akihiko S 2013).
Figure 1.6 α-Lipoic acid
1.3.3. Curcumin
Curcumin [1,7-bis (4- hydroxy-3- methoxy phenyl)-1,6-heptadiene- 3, 5 dione] (Figure 1.4)
is the major bioactive component of turmeric powder, produced from the rhizome of the
plant Curcuma longa (Aggarwal, Kumar & Bharti 2003). Turmeric is one of the most
widely used spices and functional foods and has been used as a traditional medicinal agent
in Ayurvedic medicine for ∼6000 years (Aggarwal, Kumar & Bharti 2003). The
pharmacokinetic, pharmacodynamic and clinical pharmacological properties of curcumin
have been extensively studied over the past several decades. These studies have
demonstrated that curcumin has antioxidant, anti-inflammatory and anti-atherosclerotic
activity and inhibits scarring, cataract and gallstone formation; promotes wound healing and
muscle regeneration; prevents liver injury and kidney toxicity; and exerts medicinal benefits
against psoriasis, diabetes, multiple sclerosis, Alzheimer’s, HIV disease, septic shock,

39
cardiovascular disease, lung fibrosis, arthritis and inflammatory bowel disease (Aggarwal,
Kumar & Bharti 2003; Sharma, Gescher & Steward 2005; Shishodia, Sethi & Aggarwal
2005).
Curcumin also protects against maladaptive tissue repair and improves cardiac
function after ischaemia in non-diabetic condition (Wang et al. 2012). Curcumin treatment
significantly reversed streptozotocin (STZ)-induced hyperglycaemia, glucose intolerance,
hypoinsulinaemia and pancreatic islet damage; attenuated pancreatic lipid peroxidation; up-
regulated antioxidant enzyme activity; and suppressed the serum levels of TNF-α and IL-1β
(El-Azab, Attia & El-Mowafy 2011). Curcumin dose-dependently prevented copper sulfate-
mediated oxidation of low-density lipoproteins (LDL) in vitro, an event that is associated
with thrombosis and atherosclerosis. Curcumin also efficiently inhibited TNF-α-mediated
migration of human aortic smooth muscle cells in vitro (an event associated with
atherosclerosis), and these results were correlated with a down-regulation of MMP-9
expression/activity, ROS production and NF-κB nuclear translocation (Yu & Lin 2010).
The preventive effect of curcumin on the advanced glycation and cross-linking of
collagen was also examined. Results from this report demonstrated that curcumin
administration effectively inhibits the production of AGEs and cross linking of collagen
protein in complications of diabetes mellitus (Sajithlal, Chithra & Chandrakasan 1998).
Furthermore, an anti-glycation study of curcumin demonstrated that curcumin can scavenge
the oxygen radical production generated by high glucose and inhibited the glycation of
proteins in a high glucose-treated erythrocyte cell line model (Jain, Rains & Jones 2006).

40
Figure 1.7 Curcumin
1.3.4. Ginger
For centuries, ginger (Zingiber officinale) has been an important ingredient in Ayurvedic,
Chinese and Tibb-Unani herbal medicine. The major chemical constituents of ginger
rhizome are essential volatile oils and non-volatile pungent compounds (Govindarajan
1982a, 1982b). The volatile oil components mainly consist of various terpenoids. The non-
volatile compounds include the gingerols, shogaols, paradols and zingerone. Among them,
the gingerols and shogaols were identified as the major ginger bioactive constituents that are
found in fresh and dried ginger, respectively (Figure 1.5) (Connell & Sutherland 1969). The
medicinal properties attributed to ginger include anti-arthritic (Srivastava & Mustafa 1989),
anti-migraine (Mustafa & Srivastava 1990), anti-thrombotic (Thomson et al. 2002), anti-
inflammatory (Penna et al. 2003), hypolipidaemic (Bhandari, Kanojia & Pillai 2005) and
hypocholesterolaemic effects (Fuhrman et al. 2000). It was observed that ginger juice
exhibited hypoglycaemic activity in both normal and STZ-induced diabetic rats (Akhani,
Vishwakarma & Goyal 2004). Ginger has shown protective effect in MI and atherosclerosis
in non-diabetic condition (Ansari, Bhandari & Pillai 2006). An aqueous extract of ginger at

41
0.1 and 1.0 mg/mL reduced chemical-derived AGE products in vitro (Saraswat et al. 2009).
The administration of an aqueous extract of ginger (0.5 or 3% in the diet for 2 months)
showed anti-glycation activity in diabetic rats (Saraswat et al. 2010). Ginger also inhibited
the formation of fructose-mediated AGEs of eye lens soluble proteins in vitro and delayed
the progression and onset of cataract (Saraswat et al. 2009; Saraswat et al. 2010).
Figure 1.8 Major bioactive phytoconstituents of ginger

42
1.4. Rationale and objectives
Advanced glycation end products have been implicated in the progression of diabetic
cardiovascular complications such as heart failure in diabetic patients. However, there are no
pharmaceuticals that are currently available for clinical practice. Furthermore, the glucose to
fructose shunt via the polyol pathway becomes more active in diabetic conditions resulting
in the increased concentration of fructose and thereby fructose-induced glycation also
contributes significantly towards diabetic cardiovascular complications. Therefore, fructose-
induced protein glycation, in addition to glucose-induce protein glycation, has been
investigated in this study.
Myoglobin has an important physiological function in the heart, including providing
oxygen for ATP generation and scavenging NO. If these functions are compromised under
certain conditions, ischaemia and MI occurs. Previous studies suggest that glycation of
myoglobin also compromised its function in cardiomyocytes. The immediate effect of
myoglobin glycation is to induce the release of iron from the heme moiety of myoglobin.
Due to the release of iron, myoglobin reduces its oxygen carrying capacity, resulting into
hypoxia in cardiomyocytes which may cause ischaemia or necrosis of the myocardium. The
liberated iron from myoglobin are highly reactive and participate in various free radical
generation reactions. Moreover, the glycation of myoglobin, particularly in hyperglycaemia,
also cause a serious threat with respect to formation of Amadori products, increasing protein
carbonyls and ultimately AGEs formation which may further aggravate ischaemia and MI.
Preventing glucose/fructose-induced glycation of myoglobin in the early stages of diabetes
through the use of agents which possess multiple pharmacological activities may have the
potential of reducing the risk of cardiac ischaemia and MI

43
The specific objectives of this thesis were to:
1. Explore the effectiveness of the selected test substances (α-lipoic acid, curcumin and
ginger) on the prevention of myoglobin glycation and AGEs formation under chronic
high glucose in vitro.
2. Explore the effectiveness of the selected test substances (α-lipoic acid, curcumin and
ginger) on the prevention of myoglobin glycation and AGEs formation under chronic
high fructose in vitro.
The proposed research project is based on the hypothesis that the selected test substances (α-
lipoic acid, curcumin and ginger extract) inhibit glucose/fructose-induced myoglobin
glycation and AGEs formation and thereby, could prevent/delay myocardial damage in
hyperglycaemia

44
CHAPTER-2
MATERIALS AND METHODS

45
2.1. Chemicals
α-Lipoic acid (98%), curcumin (94%), myoglobin, glucose, fructose, nitro-blue tetrazolium
tablets, hydroxylamine hydrochloride, ferrozine, dinitrophenylhydrazine, guanidine
hydrochloride, ethyl acetate, ethanol, tri-chloro acetic acid and aminoguanidine were
purchased from Sigma (St. Louis, MO, USA). The standardised ethanolic ginger extract was
obtained as a generous gift sample from Lipa Pharmaceuticals (Sydney, NSW, Australia).
Fructosamine and iron standards were obtained from PM Separations (Capalaba DC, USA).
Dipotassium phosphate, potassium dihydrogen phosphate, sodium chloride, sodium
carbonate, sodium bicarbonate, sodium acetate, glacial acetic acid, hydrochloride acid
obtained from Astral Scientific Pvt. Ltd. (Sydney, NSW, Australia)
2.2. Equipment
Analytical weighing balance (Mettler Toledo), bench top centrifuge (5424 Eppendorf),
cooling centrifuge (5810 Eppendorf), incubator (Thermo Scientific), pH meter (3505 J
Enway), UV-spectrophotometer (Bio-Rad 680), vortex mixer (Stuart), water bath (Stuart),
spectrofluorometer (Wallac 1420 Victor3 V, PerkinElmer) and magnetic stirrers (IKA) were
used in this study.
2.3. Preparation of reagents
2.3.1. 50 mM Phosphate buffered saline (PBS)
To 26.6 mL of 1 M Dipotassium phosphate (K2HPO4), 23.4 mL of 1 M potassium
dihydrogen phosphate (KH2PO4) and 30 mL of 5 M sodium chloride (NaCl) were added into
a volumetric flask and made up to 1 L with distilled water. The solution (final concentration

46
of 50 mM) was transferred into a beaker and the pH was adjusted to 6.6 and filtered before
use through a 0.2 μM membrane filter.
2.3.2. 1 mg/mL Myoglobin solution
Fifty grams of powdered myoglobin was weighed accurately and transferred into a
volumetric flask and made up to 50 mL with 50 mM PBS (pH 6.6). The solution (1 mg/mL)
was transferred into a beaker and filtered before use through a 0.2 μM membrane filter.
2.3.3. 1 M Glucose
D-(+) glucose (18.01 g) was weighed accurately and transferred into a volumetric flask and
made up to 100 mL with distilled water.
2.3.4. 1 M Fructose
D-(−) fructose (18.01 g) was weighed accurately and transferred into a volumetric flask and
made up to 100 mL with distilled water.
2.3.5. Stock solutions of ginger extract
The ethanolic extract of ginger (lyophilised dried powder – 10 mg, 20 mg and 30 mg) was
weighed accurately and transferred to different volumetric flasks of 10 mL capacity and
made up to the mark with dimethyl sulfoxide to give concentrations of 1 mg/mL, 2 mg/mL
and 3 mg/mL, respectively.

47
2.3.6. Stock solutions of curcumin
Curcumin (368.3 µg, 736.76 µg and 1105.14 µg of dried powder) was weighed accurately
and transferred to different volumetric flasks of 10 mL capacity and made up to the mark
with dimethyl sulfoxide to give concentrations of 100 µM, 200 µM and 300 µM,
respectively
2.3.7. Stock solutions of α-lipoic acid
α-Lipoic acid (20.6 mg, 41.2 mg and 82.4 mg of dried powder) was weighed accurately and
transferred to 10 mL volumetric flasks and made up to the mark with dimethyl sulfoxide to
give concentrations of 10 mM, 20 mM and 40 mM, respectively.
2.3.8. 0.2 M Carbonate buffer (pH 10.8)
To 27.5 mL of 0.2 M sodium carbonate (Na2Co3), 22.5 mL of 0.2 M sodium bicarbonate
(NaHCO3) was added to a volumetric flask and made up to 100 mL with distilled water
(final concentration of 0.2 M). The solution was transferred into a beaker and the pH was
adjusted to 10.8. The solution was filtered before use through a 0.2 μM membrane filter.
2.3.9. 0.5 mM Nitro-blue tetrazolium chloride reagent
Nitro-blue tetrazolium (40.8 g) was weighed accurately and transferred to a volumetric flask
and made up to 100 mL (final concentration of 0.5 mM) with 0.2 M carbonate buffer (pH
10.8). The solution was transferred into a beaker filtered before use through a 0.2 μM
membrane filter. This reagent was freshly prepared prior to use.

48
2.3.10. 0.2 M Sodium acetate buffer (pH 4.5)
Sodium acetate (5.44 g) was weighed accurately and transferred into a volumetric flask. To
this, glacial acetic acid (1.2 mL) was added and made up to 100 mL with distilled water
(final concentration 0.2 M). The solution was transferred into a beaker and the pH was
adjusted to 4.5 and the solution was filtered before use through a 0.2 μM membrane filter.
2.3.11. 1.5% Iron buffer
Hydroxylamine hydrochloride (1.5 g) was weighed accurately and transferred into a
volumetric flask and made up to 100 mL with 0.2 M sodium acetate buffer.
2.3.12. 0.85% Iron colour reagent
Ferrozine (0.85 g) was weighed accurately and transferred into a volumetric flask and made
up to 100 mL with iron buffer.
2.3.13. 6 M Guanidine solution
Guanidine hydrochloride (17.25 g) was weighed accurately and transferred into a volumetric
flask and made up to 30 mL with 20 mM phosphate buffer (to a final concentration of 6 M).
The solution was transferred into a beaker and filtered before use through a 0.2 μM
membrane filter.
2.3.14. 10 mM Dinitrophenylhydrazine (DNPH)
Dinitrophenylhydrazine (0.198 g) was weighed accurately and transferred into a volumetric
flask and made up to 100 mL with 2.5 M hydrochloride acid (final concentration of 10 mM).

49
The solution was transferred into a beaker and filtered before use through a 0.2 μM
membrane filter.
2.3.15. 5 mM 5,5′-dithiobis(2-nitro-benzoicacid) (DTNB)
DTNB (0.198 g) was weighed accurately and transferred into a volumetric flask and made
up to 100 mL with 0.1 M phosphate buffered saline (final concentration 5 mM). The
solution was transferred into a beaker and filtered before use through a 0.2 μM membrane
filter.
2.4. The test substances and dosage selection
α-Lipoic acid, curcumin and ginger extract have been selected as a test substance in the
present investigation. The purity of α-lipoic acid used in the studies is 98% while that of
curcumin is 94% based on the manufacturers product specifications. The standardised
ethanolic ginger extract used in the studies was generously obtained from Lipa
Pharmaceuticals (Sydney, Australia) and further characterised and standardised earlier in our
laboratory as described by Nammi et al. (2010). Briefly, the ginger extract is reported to
contain three pungent compounds 6-shogaol, 6-gingerol, and 8-gingerol as major
compounds, the contents of which are respectively found to be 11.70 mg, 1.56 mg, and 0.24
mg per gram of dried extract. The selected doses of the test substances were rationalised
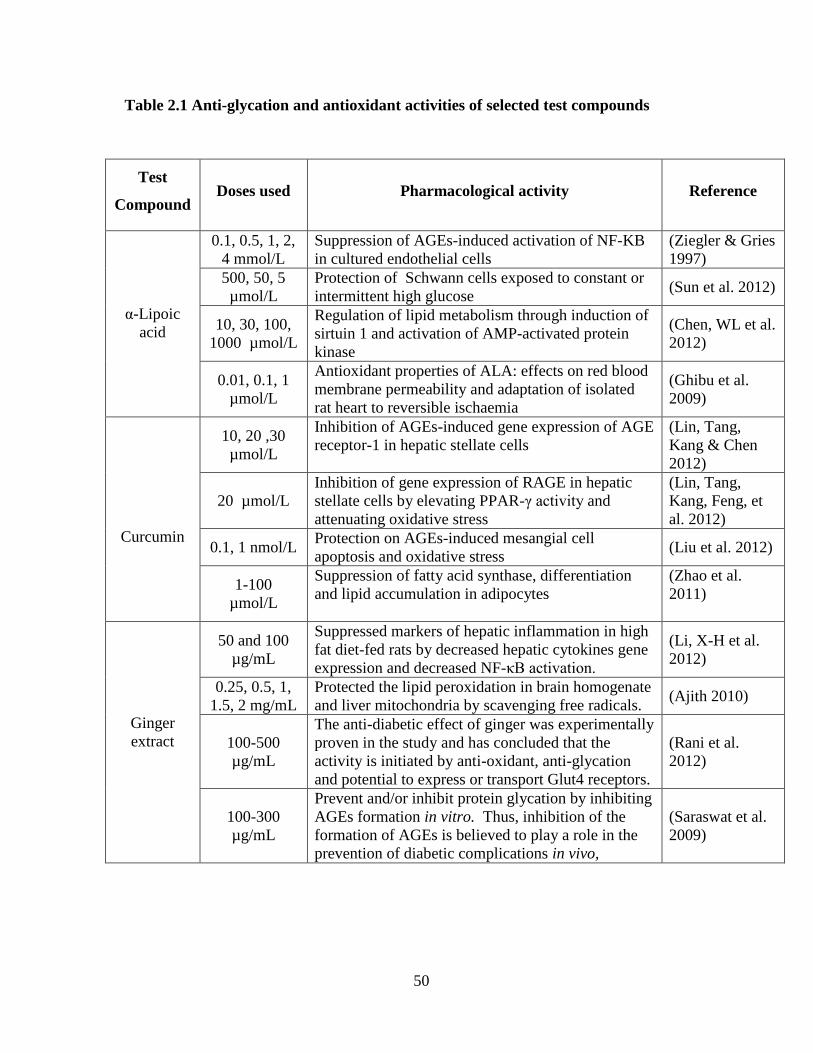
based on published literature. Table 2.1 lists some reported anti-glycation and antioxidant
activities of α-lipoic acid, curcumin and ginger extract. Thus, three different concentrations
of α-lipoic acid (1, 2 and 4 mmol/L), curcumin (10, 20 and 30 µmol/L) and ginger extract
(100, 200 and 300 µg/mL) were selected for the proposed study.
50
Table 2.1 Anti-glycation and antioxidant activities of selected test compounds
Test
Compound Doses used Pharmacological activity Reference
α-Lipoic
acid
0.1, 0.5, 1, 2,
4 mmol/L
Suppression of AGEs-induced activation of NF-KB
in cultured endothelial cells
(Ziegler & Gries
1997)
500, 50, 5
µmol/L
Protection of Schwann cells exposed to constant or
intermittent high glucose (Sun et al. 2012)
10, 30, 100,
1000 µmol/L
Regulation of lipid metabolism through induction of
sirtuin 1 and activation of AMP-activated protein
kinase
(Chen, WL et al.
2012)
0.01, 0.1, 1
µmol/L
Antioxidant properties of ALA: effects on red blood
membrane permeability and adaptation of isolated
rat heart to reversible ischaemia
(Ghibu et al.
2009)
Curcumin
10, 20 ,30
µmol/L
Inhibition of AGEs-induced gene expression of AGE
receptor-1 in hepatic stellate cells
(Lin, Tang,
Kang & Chen
2012)
20 µmol/L
Inhibition of gene expression of RAGE in hepatic
stellate cells by elevating PPAR-γ activity and
attenuating oxidative stress
(Lin, Tang,
Kang, Feng, et
al. 2012)
0.1, 1 nmol/L Protection on AGEs-induced mesangial cell
apoptosis and oxidative stress (Liu et al. 2012)
1-100
µmol/L
Suppression of fatty acid synthase, differentiation
and lipid accumulation in adipocytes
(Zhao et al.
2011)
Ginger
extract
50 and 100
µg/mL
Suppressed markers of hepatic inflammation in high
fat diet-fed rats by decreased hepatic cytokines gene
expression and decreased NF-κB activation.
(Li, X-H et al.
2012)
0.25, 0.5, 1,
1.5, 2 mg/mL
Protected the lipid peroxidation in brain homogenate
and liver mitochondria by scavenging free radicals. (Ajith 2010)
100-500
µg/mL
The anti-diabetic effect of ginger was experimentally
proven in the study and has concluded that the
activity is initiated by anti-oxidant, anti-glycation
and potential to express or transport Glut4 receptors.
(Rani et al.
2012)
100-300
µg/mL
Prevent and/or inhibit protein glycation by inhibiting
AGEs formation in vitro. Thus, inhibition of the
formation of AGEs is believed to play a role in the
prevention of diabetic complications in vivo,
(Saraswat et al.
2009)

51
2.5. Evaluation of myoglobin glycation inhibitory effect of α-lipoic acid, curcumin
and ginger extract under glucose or fructose overload in vitro
The glycation of myoglobin in vitro was performed according to previously described
methods with minor modification (Roy, Sen & Chakraborti 2004; Roy, Sil & Chakraborti
2010). Briefly, 500 µL of myoglobin was incubated with 400 µL of 1 M glucose (1M) or
fructose (1M) solution at 37°C in the dark for 30 days in the presence or absence of different
concentrations of α-lipoic acid, curcumin and ginger extract. Aminoguanidine (5 mM), a
known inhibitor of glycation process and fluorescent AGEs formation, was used as the
positive control for this study. After the specified incubation period (for 10, 20, or 30 days),
aliquots of the glycated reaction mixtures were taken and assayed for fluorescent AGEs,
liberated iron, fructosamine (glycated myoglobin), protein carbonyls and protein thiols as
described below.
2.5.1. Estimation of total fluorescent AGEs formation
The formation total fluorescent AGEs was determined by spectrofluorometrically after 30
days of incubation.
2.5.1.1. Assay principle
The chemical reaction between amino group of protein and carbonyl group of sugar
produced heterogeneous compounds termed as advance glycation end products through
various chemical reactions. AGEs are a very strong fluorescing compound, especially in
dilute acid solution, and thus can be detected in very trace amounts by a spectrofluorometer.
Most fluorescent AGEs have shown fluorescence at excitation wavelengths of 355 nm and
emission wavelengths of 460 nm. The basis for quantitation is that the intensity of

52
fluorescence emission in very dilute solutions is directly proportional to the concentration of
florescent AGEs
2.5.1.2. Assay procedure
To 1 mL of glycated reaction mixture, 250 µL of TCA (100) and 2 mL of chloroform were
added. The resulting mixture was vortexed for 60 seconds and centrifuged at 14,000 rpm for
4 min at 4 0C temperature. The collected supernatant was placed in a 1 mL disposable
polystyrene cuvette and fluorescence intensity was read at an excitation wavelength 355 nm
and emission wavelength 460 nm. The percentage inhibition of fluorescent AGEs formation
was calculated as follows:
Inhibition of fluorescent AGEs (%) = [(FC−FS)/ (FC] x100
Where FC = fluorescent intensity of control; FS = fluorescent intensity of sample
Furthermore, the concentration required for 50% inhibition of fluorescent AGEs formation
relative to the negative control (known as an IC50) was also calculated by using best fit
equation.
2.5.2. Estimation of liberated iron (ferrozine test)
2.5.2.1. Assay principle
The immediate effect of myoglobin glycation is to induce the release of iron from heme
moiety of myoglobin. Due to release of iron, myoglobin reduces its oxygen carrying
capacity, resulting in hypoxia in cardiomyocytes which may cause ischaemia or necrosis of
myocardium. Furthermore, liberated iron increases the free radical generation and
subsequently, increases the oxidative stress. To understand the effect of glycation on iron
release, free iron in myoglobin was estimated by the ferrozine test. In acidic conditions,

53
ferric iron dissociates from transferrin-iron complex and is reduced to ferrous iron. Ferrous
iron then reacts with ferrozine to give a pink-coloured complex:
Transferrin (Fe3+
) + e- 2 Fe2+
+ Transferrin
Fe2+
+ Ferrozine Coloured complex (pink)
The intensity of the color formed is proportional to the concentration of iron in the sample.
2.4.2.2 Assay procedure
The procedure described by Panter (Panter 1994) was followed for detection of free iron in
solution. Briefly, 250 μL of the glycated reaction mixture/iron standard was mixed with 250
μL of cold 20% tri-chloroacetic acid (TCA), centrifuged at 15,000 rpm for 4 minutes and the
supernatant was collected. To 250 μL of supernatant, 2.5 mL of iron buffer and 50 μL iron
colour reagent were added. The resultant mixture was incubated at 37 °C for 30 min and the
absorbance was measured at 560 nm.
Calculation
Concentration of liberated free iron was calculated by the following formula
Concentration of liberated iron (μg/dL) =

54
2.5.3. Estimation of fructosamine (glycated myoglobin)
2.5.3.1. Assay principle
The fructosamine or Amadori products are the first stable products of the Millard reaction.
The amount of formed fructosamine is equivalent to the amount of protein glycated during
the Millard reaction. Thus, the measurement of fructosamine gives an index of glycated
myoglobin. The measurement of fructosamine was carried out according to the method of
Ohkawara (Ohkawara et al. 2002). The assay is based on the ability of fructosamine formed
by the non-enzymatic glycation of myoglobin and glucose/fructose to reduce NBT to the
tetrazinolyl radical NBT+
which dissociates to yield a deep blue coloured complex formazon
(MF+) which was measured colorimetrically at 540 nm.
2.5.3.2. Assay procedure
To 200 µL of glycated reaction mixture or standard fructosamine, 1 mL NBT reagent was
added to a disposable polystyrene cuvette inside a UV-Visible spectrophotometer (presetting
the temperature at 37 0C) and the time was noted. The absorbance values at 540 nm after 10
minutes and 15 minutes were noted and the absorbance difference was calculated.
Calculation
The concentration of fructosamine was calculated by the following fructosamine (µM)
formula:

55
2.5.4. Estimation of protein carbonyls
2.5.4.1. Assay principle
The elevated levels of protein carbonyl content are an index of glycation-induced protein
oxidation. The autoxidation of glucose and glycoxidation of Amadori products generate
highly reactive dicarbonyl compounds which can further generate AGEs. Carbonyl content
is the most general and commonly used biomarker of protein glycation-induced oxidative
modification of protein. Protein carbonyl groups react with dinitrophenylhydrazine (DNHP)
to form 2, 4- dinitrophenylhydrazone. The excess DNHP is removed by extraction with an
ethanol/ethyl acetate solution. After extraction, the 2, 4- dinitrophenylhydrazone can be
determined spectrophotometrically by its absorbance at 370 nm.
2.5.4.2. Assay procedure
Protein carbonyls were measured according to the method of Levine (Levine et al. 1994). To
200 µL of the glycated reaction mixture, 200 μL of 10 mM DNPH was added and mixed
thoroughly. To this mixture, 250 μL of TCA (30%) was added and centrifuged at 15,000
rpm for 4 min at 4 0C temperature. The pellet was collected and washed three times with 1
mL ethanol: ethyl acetate (1:1) mixture to remove any unreacted DNPH. The pellet was then
dissolved in 1 mL of 6 M guanidine hydrochloride solution and incubated at 37 °C for 15
min. After centrifugation at 15, 000 rpm for 4 min at 4 0C temperature, the absorbance of the
supernatant was measured at 375 nm using the molar absorption coefficient of 22,000 M−1
cm−1
for DNPH.

56
Calculation
With the absorbance of the sample taken at 375 nm, the protein carbonyls content was
calculated as:
Protein Carbonyls (M) = A375 nm / 22,000 M-1
Protein Carbonyls (nmol/mL) = A375 nm x 45.45 (nmol/L)
2.5.5. Estimation of protein thiols
2.5.5.1. Assay principle
The total protein thiols pool constitutes the majority of the total body antioxidants and plays
a major role against ROS (Rossi et al. 2009). Protein glycation increase the oxidation of
protein thiols groups and decrease the antioxidant defense. 5, 5′-Dithiobis (2-nitro-benzoic
acid) (DTNB) reacts with a free sulfhydryl group to yield a mixed disulfide and pink-
coloured 2-nitro-5-thiobenzoic acid (TNB) which is measured at 412 nm.
2.5.5.2. Standard curve of L-cysteine
Various concentrations of L-cysteine ranging from 20 nM to 100 nM were prepared in
distilled water. Theses concentrations (20, 40, 60, 80, 100 nM) were heated with three
volumes of 0.5 mM DTNB at 25 0C for 15 min and the absorbance was taken at 412 nm.
The standard curve was used to calculate the concentration of protein thiols in the glycated
reaction mixtures.

57
2.5.5.3. Assay procedure
The free thiols in the glycated reaction mixture were measured by Ellman’s assay with
minor modification (Ellman 1959). Briefly, 70 μL of glycated reaction mixture was
incubated with 130 μL of 5 mM DTNB at 25 °C for 15 min and the absorbance of the
resulting mixture was measured at 412 nm. The concentration of free thiols was calculated
from the L-cysteine standard and expressed as nmol/mg protein.
2.5.6. Data and statistical analysis
Data are expressed as mean standard error of means. Statistical analysis was done using
one-way analysis of variance (ANOVA) followed by Dunnet’s test and paired t-test using
Graph Pad Prism software (Version 5.0). In all tests, p<0.05 was used as the criterion for
statistical significance when compared to the negative controls.

58
CHAPTER-3
RESULTS

59
3.1. Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-induced
fluorescent AGEs formation
Protein glycation depends on the degree and duration of hyperglycaemia. The Amadori
products (fructosamine) undergo further rearrangement to form both fluorescent and non-
fluorescent AGEs. As mentioned in Chapter one, various fluorescent AGEs such as
pentosidine and crossline are considered the most likely culprits in the development of
diabetic cardiovascular complications.
3.1.1. α-Lipoic acid
As shown in Table 3.1 and 3.2, the formation of fluorescent AGEs was monitored on day-30
after incubation by measuring the fluorescence intensity in the myoglobin-glucose and
myoglobin-fructose glycation models. The addition of α-lipoic acid to the myoglobin-
glucose glycation model at the final concentrations of 1, 2 and 4 mmol/L significantly
(p<0.01; n=6) inhibited the fluorescent AGEs formation by 43.6%, 52.4% and 56.3%,
respectively compared to the negative control, with an IC50 value of 2.1 mmol/L as observed
from a concentration-dependent decrease in the fluorescence intensity (Figure 3.1A). α-
Lipoic acid also showed a significant (p<0.01; n=6) concentration-dependent inhibition of
fluorescent AGEs formation in the myoglobin-fructose glycation model by 42.9%, 46.5%
and 49.7% at the final concentrations of 1, 2 and 4 mmol/L respectively compared to the
negative control, with an IC50 value of 4.0 mmol/L (Figure 3.1A). In comparison,
aminoguanidine (positive control) at the concentration of 5 mmol/L significantly (p<0.01;
n=6) inhibited the fluorescent AGEs formation on day-30 by 71.6% in the myoglobin-
glucose model, and by 78.6% in the myoglobin-fructose model compared to the negative
control.

60
3.1.2. Curcumin
As shown in Tables 3.1 and 3.2, the formation of fluorescent AGEs was monitored on day-
30 after incubation by measuring the fluorescence intensity in myoglobin-glucose and
myoglobin-fructose glycation models. The addition of curcumin to the myoglobin-glucose
glycation model at the final concentrations of 10, 20 and 30 µmol/L significantly (p<0.05 to
p<0.01; n=6) inhibited the fluorescent AGEs formation by 31.4%, 42.4% and 48.5%,
respectively compared to the negative control. An IC50 value of 30.6 µmol/L was observed
from a concentration-dependent decrease in the fluorescence intensity (Figure 3.1B).
Curcumin also showed a significant (p<0.05 to p<0.01; n=6) concentration-dependent
inhibition of fluorescent AGEs formation in the myoglobin-fructose glycation model by
16.1%, 29.5% and 33.1% at the final concentrations of 10, 20 and 30 µmol/L, respectively
compared to the negative control, with an IC50 value of 48.6 µmol/L (Figure 3.1B). On the
other hand, the positive control (aminoguanidine) at the concentration of 5 mmol/L
significantly (p<0.01; n=6) inhibited the fluorescent AGEs formation by 71.6% in the
myoglobin-glucose model and by 78.6% in the myoglobin-fructose model when compared
to the negative control.
3.1.3. Ginger
As shown in Table 3.1 and 3.2, the formation of fluorescent AGEs was monitored on day-30
after incubation by measuring the fluorescence intensity in the myoglobin-glucose and
myoglobin-fructose glycation models. The addition of ginger to the myoglobin-glucose
glycation model at the final concentrations of 100, 200 and 300 µg/mL significantly (p<0.05
to p<0.01; n=6) inhibited the fluorescent AGEs formation by 29.7%, 39.7% and 45.0%,

61
respectively compared to the negative control, with an IC50 value of 355.6 µg/mL as
observed from a concentration-dependent decrease in the fluorescence intensity (Figure
3.1C). Ginger also showed a significant (p<0.05 to p<0.01; n=6) concentration-dependent
inhibition of fluorescent AGEs formation in myoglobin-fructose glycation model by 11.8%,
22.6% and 27.9% at the final concentrations of 100, 200 and 300 µg/mL respectively
compared to the negative control, with an IC50 value of 533.0 µg/mL (Figure 3.1C). The
positive control, aminoguanidine at the concentration of 5 mM significantly (p<0.01; n=6)
inhibited the fluorescent AGEs formation by 71.6% in the myoglobin-glucose model and by
78.6% in the myoglobin-fructose model compared to the negative control.

62
Table 3.1 Effect of α-lipoic acid, curcumin and ginger on the formation of fluorescent
AGEs in myoglobin-glucose glycation
Experimental group Fluorescence
intensity
% inhibition of
AGEs formation IC50
Negative control 7633.33 ± 332.53 - -
α-Lipoic acid (1 mmol/L) 4300.00 ± 309.44** 43.7
2.1 (mmol/L) α-Lipoic acid (2 mmol/L) 3633.33 ± 300.09** 52.4
α-Lipoic acid (4 mmol/L) 3333.33 ± 300.09** 56.3
Curcumin (10 µmol/L) 5233.33 ± 442.80* 31.4
30.6 (µmol/L)
Curcumin (20 µmol/L) 4400.00 ± 409.36** 42.4
Curcumin (30 µmol/L) 3933.33 ± 147.17** 48.5
Ginger (100 µg/mL) 5366.67 ± 300.09* 29.7
355.3
(µg/mL)
Ginger (200 µg/mL) 4600.00 ± 409.36** 39.7
Ginger (300 µg/mL) 4200.00 ± 350.88** 45.0
Aminoguanidine (5 mmol/L) 2166.67 ± 147.17** 71.6
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01.

63
Table 3.2 Effect of α-lipoic acid, curcumin and ginger on the formation of fluorescent
AGEs in myoglobin-fructose glycation
Experimental group Fluorescence
intensity
% inhibition of
AGEs formation IC50
Negative control 9226.33 ± 337.74 -
α-Lipoic acid (1 mmol/L) 5266.67 ± 469.05** 42.92
4.0 (mmol/L) α-Lipoic acid (2 mmol/L) 4933.33 ± 89.33** 46.53
α-Lipoic acid (4 mmol/L) 4633.33 ± 510.93** 49.78
Curcumin (10 µmol/L) 7733.33 ± 276.36* 16.18
48.6 (µmol/L)
Curcumin (20 µmol/L) 6500.00 ± 233.92** 29.55
Curcumin (30 µmol/L) 6166.67 ± 205.37** 33.16
Ginger (100 µg/mL) 8134.33 ± 263.64* 11.84
533.0
(µg/mL)
Ginger (200 µg/mL) 7137.33 ± 510.93** 22.64
Ginger (300 µg/mL) 6648.67 ± 328.62** 27.94
Aminoguanidine (5 mmol/L) 1969.67 ± 81.42** 78.65 -
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01.

64
Figure 3.1 Percentage inhibition of fluorescent AGEs formation by α-lipoic acid (A),
curcumin (B) and ginger (C) in myoglobin-glucose (Mb-Glu) and myoglobin-fructose
(Mb-Fru) glycation after 30 days of incubation.

65
3.2. Effect of α-lipoic acid, curcumin and ginger on free iron release in myoglobin-
glycation
Non-enzymatic glycation of myoglobin has a tendency to release free iron from the heme
moiety of myoglobin. This liberated iron increases the free radical generation and
subsequently, increases oxidative stress which is a high risk factor for myocardium damage.
3.2.1. α-Lipoic acid
The effects of α-lipoic acid on glycation-induced iron release in myoglobin-glucose and
myoglobin-fructose glycation models are shown in Table 3.3 and 3.4, respectively. In the
negative control group, glycation-induced liberated free iron was time-dependently
increased over the studied time periods and was significantly different (p<0.05; n=6) on day-
30 compared to day-10 in both the glycation models. The supplementation of α-lipoic acid
in the myoglobin-glucose model at the final concentrations of 1, 2 and 4 mmol/L displayed a
significant (p<0.05 to p<0.01; n=6) concentration-dependent reduction in liberated free iron
at the corresponding time points when compared to the negative control. On day-30 of
incubation, α-lipoic acid significantly reduced (p<0.01; n=6) the liberation of free iron
compared to the negative control by 21.9%, 42.8% and 45.1% at the three concentrations (1,
2 and 4 mmol/L) studied, with an IC50 value of 4.3 mmol/L. (Figure 3.2A). Similarly, α-
lipoic acid also showed a significant concentration-dependent reduction (p<0.01; n=6) in the
liberated free iron on day-30 compared to the negative control by 23.9%, 43.2% and 44.5%
at the respective concentrations in the myoglobin-fructose glycation model, with an IC50
value of 4.4 mmol/L (Figure 3.2A). These results are statistically comparable to the positive
control aminoguanidine (5 mmol/L) which exhibited a significant reduction (p<0.01; n=6) in

66
the released free iron on day-30 by 48.2% in the myoglobin-glucose model, and by 48.8% in
the myoglobin-fructose model compared to the negative control.
3.2.2. Curcumin
The effects of curcumin on glycation-induced iron release in the myoglobin-glucose and
myoglobin-fructose models are shown in Table 3.3 and 3.4, respectively. In the negative
control group, glycation-induced liberated free iron was time-dependently increased over the
studied time periods and achieved significance (p<0.05; n=6) on day-30 compared to day-10
in both the glycation models. Added curcumin in the myoglobin-glucose model at the final
concentrations of 10, 20 and 30 µmol/L displayed a significant concentration-dependent
reduction (p<0.05 to p<0.01; n=6) in liberated free iron at the corresponding time points
compared to the negative control. On day-30 of incubation, curcumin significantly (p<0.05
to p<0.01; n=6) reduced the liberation of free iron compared to the negative control by
23.3%, 39.2% and 41.3% respectively at the three concentrations studied, with an IC50 value
of 37.1 µmol/L (Figure 3.2B). Similarly, curcumin (10, 20 and 30 µmol/L) also showed a
significant concentration-dependent reduction (p<0.05 to p<0.01; n=6) in the liberated free
iron on day-30 compared to the negative control by 20.4%, 31.3% and 41.5%, respectively
in the myoglobin-fructose glycation model, with an IC50 value of 38.0 µmol/L (Figure
3.2B). These results are statistically comparable with the positive control aminoguanidine (5
mmol/L) which exhibited a significant reduction (p<0.01; n=6) in the released free iron on
day-30 by 48.2% in the myoglobin-glucose model, and by 48.8% in the myoglobin-fructose
model compared to the negative control.

67
3.2.3. Ginger
The effects of ginger on glycation-induced iron release in the myoglobin-glucose and
myoglobin-fructose models are shown in Table 3.3 and 3.4, respectively. In the negative
control group, glycation-induced liberated free iron was time-dependently increased over the
studied time periods and achieved significance (p<0.05; n=6) on day-30 compared to day-10
in both the glycation models. Added ginger in the myoglobin-glucose model at the final
concentrations of 100, 200 and 300 µg/mL displayed a significant concentration-dependent
reduction (p<0.01; n=6) in liberated free iron at corresponding time points when compared
to the negative control. On day-30 of incubation, ginger significantly reduced (p<0.01; n=6)
the liberation of free iron compared to the negative control by 20.0%, 35.1% and 42.1% at
the three concentrations studied, with an IC50 value of 358.2 µg/mL (Figure 3.1C).
Similarly, ginger extract (100, 200 and 300 µg/mL) also showed a significant concentration-
dependent reduction (p<0.01; n=6) in the liberated free iron on day-30 compared to the
negative control by 18.4%, 33.1% and 40.8%, respectively in the myoglobin-fructose
glycation model, with an IC50 value of 374.1 µg/mL (Figure 3.2C). These results are
statistically comparable with the positive control aminoguanidine (5 mmol/L) that also
exhibited a significant reduction (p<0.01; n=6) in the released free iron on day-30 by 48.2%
in the myoglobin-glucose model and by 48.8% in the myoglobin-fructose model compared
to the negative control.
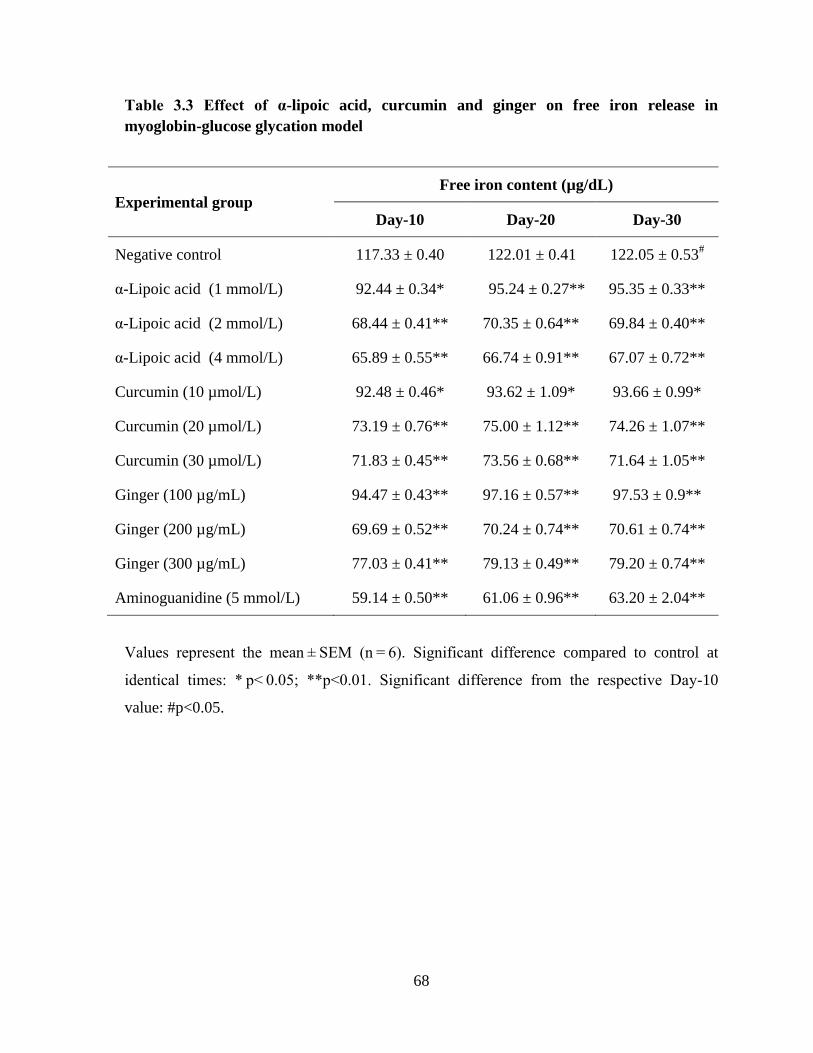
68
Table 3.3 Effect of α-lipoic acid, curcumin and ginger on free iron release in
myoglobin-glucose glycation model
Experimental group Free iron content (µg/dL)
Day-10 Day-20 Day-30
Negative control 117.33 ± 0.40 122.01 ± 0.41 122.05 ± 0.53#
α-Lipoic acid (1 mmol/L) 92.44 ± 0.34* 95.24 ± 0.27** 95.35 ± 0.33**
α-Lipoic acid (2 mmol/L) 68.44 ± 0.41** 70.35 ± 0.64** 69.84 ± 0.40**
α-Lipoic acid (4 mmol/L) 65.89 ± 0.55** 66.74 ± 0.91** 67.07 ± 0.72**
Curcumin (10 µmol/L) 92.48 ± 0.46* 93.62 ± 1.09* 93.66 ± 0.99*
Curcumin (20 µmol/L) 73.19 ± 0.76** 75.00 ± 1.12** 74.26 ± 1.07**
Curcumin (30 µmol/L) 71.83 ± 0.45** 73.56 ± 0.68** 71.64 ± 1.05**
Ginger (100 µg/mL) 94.47 ± 0.43** 97.16 ± 0.57** 97.53 ± 0.9**
Ginger (200 µg/mL) 69.69 ± 0.52** 70.24 ± 0.74** 70.61 ± 0.74**
Ginger (300 µg/mL) 77.03 ± 0.41** 79.13 ± 0.49** 79.20 ± 0.74**
Aminoguanidine (5 mmol/L) 59.14 ± 0.50** 61.06 ± 0.96** 63.20 ± 2.04**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05.

69
Table 3.4 Effects of α-lipoic acid, curcumin and ginger on free iron release in
myoglobin-fructose glycation
Experimental group Free iron content (µg/dL)
Day-10 Day-20 Day-30
Negative control 121.83 ± 2.17 123.21 ± 0.89 125.26 ± 0.62#
α-Lipoic acid (1 mmol/L) 94.91 ± 0.50** 95.03 ± 0.47** 95.32 ± 1.05**
α-Lipoic acid (2 mmol/L) 70.24 ± 0.69** 70.81 ± 0.98** 71.05 ± 1.09**
α-Lipoic acid (4 mmol/L) 68.22 ± 0.44** 69.22 ± 1.12** 69.47 ± 1.19**
Curcumin (10 µmol/L) 92.48 ± 0.46* 93.62 ± 1.09* 93.66 ± 0.99*
Curcumin (20 µmol/L) 73.19 ± 0.76** 75.00 ± 1.12** 74.26 ± 1.07**
Curcumin (30 µmol/L) 71.83 ± 0.45** 73.56 ± 0.68** 71.64 ± 1.05**
Ginger (100 µg/mL) 100.04 ± 0.73** 101.58 ± 1.23** 102.10 ± 0.52**
Ginger (200 µg/mL) 81.08 ± 1.62** 81.93 ± 0.54** 82.45 ± 1.36**
Ginger (300 µg/mL) 72.12 ± 0.92** 73.89 ± 1.56** 74.15 ± 1.17**
Aminoguanidine (5 mmol/L) 62.98 ± 1.08** 63.01 ± 0.10** 64.16 ± 1.50**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05;
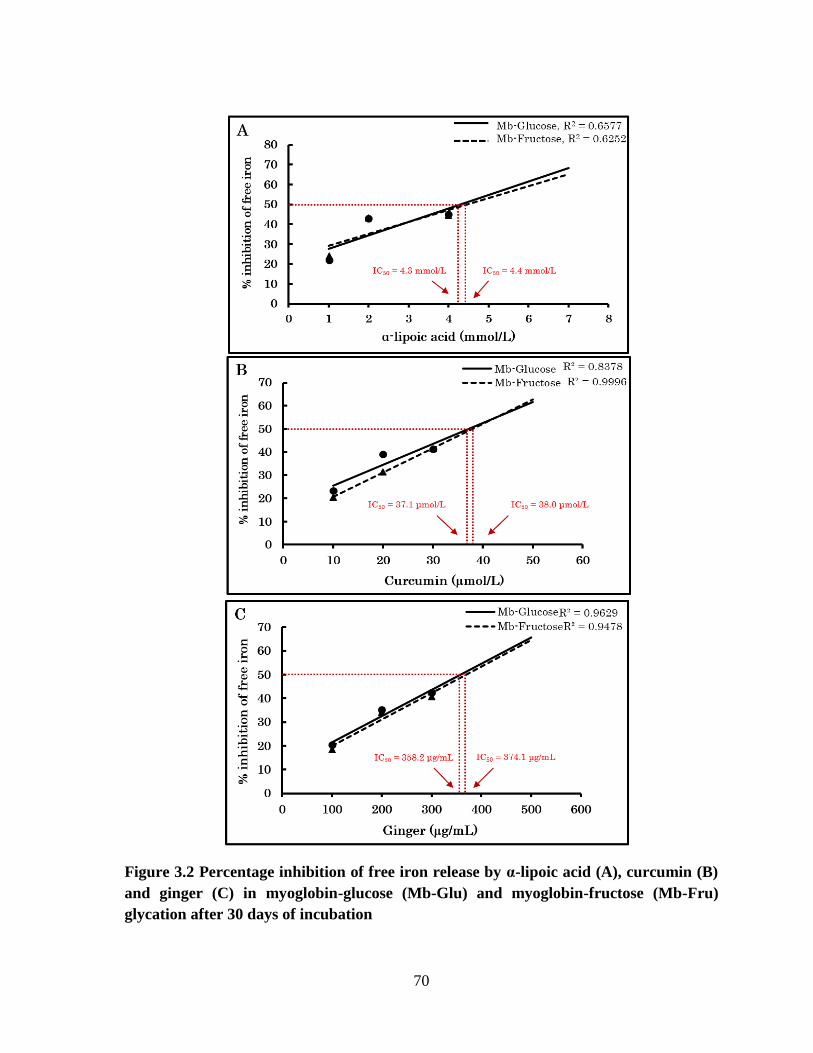
70
Figure 3.2 Percentage inhibition of free iron release by α-lipoic acid (A), curcumin (B)
and ginger (C) in myoglobin-glucose (Mb-Glu) and myoglobin-fructose (Mb-Fru)
glycation after 30 days of incubation

71
3.3. Effect of α-lipoic acid, curcumin and ginger on level of fructosamine formation
by myoglobin glycation
Fructosamine (Amadori products) is considered as the first stable product of Millard
reaction. Thus, the amount of fructosamine formed is equivalent to the degree of protein
glycation during the Millard reaction. Therefore, in the present study, time-dependent
measurement of fructosamine which provides an index of glucose/fructose-mediated
myoglobin glycation was monitored at 10, 20 and 30 days of incubation.
3.3.1. α-Lipoic acid
The effect of α-lipoic acid on glycation-induced fructosamine formation in myoglobin-
glucose and myoglobin-fructose glycation models is shown in Table 3.5 and 3.6,
respectively. In the negative control group (n=6), glycation-induced fructosamine formation
was time-dependently increased over the studied time periods and achieved significance on
day-20 (p<0.05) and day-30 (p<0.01) compared to day-10 in both glycation models. When
α-lipoic acid was added to myoglobin-glucose model at the final concentrations of 1, 2 and 4
mmol/L, a significant (p<0.05 to p<0.01; n=6) concentration-dependent reduction in the
fructosamine value was noted at the corresponding time points compared to the negative
control. On day-30 of incubation, α-lipoic acid significantly reduced (p<0.01; n=6) the
fructosamine levels compared to the negative control by 19.5%, 31.2%, and 36.6% at the
three concentrations studied, with an IC50 value of 6.2 mmol/L (Figure 3.3A). Similarly, α-
lipoic acid (1, 2 and 4 mmol/L) also showed a significant concentration-dependent reduction
(p<0.05 to p<0.01; n=6) in fructosamine formation on day-30 compared to the negative
control by 25.1%, 28.3% and 34.5%, respectively in myoglobin-fructose glycation model,
with an IC50 value of 8.9 mmol/L (Figure 3.3A). Furthermore, these results are comparable

72
with the positive control aminoguanidine (5 mmol/L) which significantly (p<0.01; n=6)
inhibited the formation of fructosamine on day-30 by 55.9% in the myoglobin-glucose
glycation model and by a less-marked 37.9% in the myoglobin-fructose glycation model
compared to the negative control.
3.3.2. Curcumin
The effect of curcumin on glycation-induced fructosamine formation in the myoglobin-
glucose and myoglobin-fructose glycation models is shown in Table 3.5 and 3.6,
respectively. In the negative control group (n=6), glycation-induced fructosamine formation
was time-dependently increased over the studied time periods and achieved significance on
day-20 (p<0.05) and day-30 (p<0.01) compared to day-10 in both the glycation models.
When curcumin was added to myoglobin-glucose model at the final concentrations of 10, 20
and 30 µmol/L, a significant (p<0.01; n=6) concentration-dependent reduction in the
fructosamine formation was noted on day-20 and day-30 compared to the negative control.
On day-30 of incubation, curcumin significantly reduced (p<0.05 to p<0.01; n=6) the
fructosamine levels compared to the negative control by 23.3%, 39.2% and 41.3%, at the
three concentrations studied, with an IC50 value of 47.1 µmol/L (Figure 3.3B). Similarly,
curcumin (10, 20 and 30 µmol/L) also showed a significant concentration-dependent
reduction (p<0.05 to p<0.01) in fructosamine formation on day-30 compared to the negative
control by 20.4%, 31.3% and 41.5% in the myoglobin-fructose glycation model, with an
IC50 value of 74.7 µmol/L (Figure 3.3B). Furthermore, these results are comparable to the
positive control aminoguanidine (5 mmol/L) which significantly (p<0.01) inhibited
fructosamine formation on day-30 by 55.9% in the myoglobin-glucose glycation model and
by 37.9% in the myoglobin-fructose glycation model compared to the negative control.

73
3.3.3. Ginger
The effect of ginger on glycation-induced fructosamine formation in myoglobin-glucose and
myoglobin-fructose glycation models is shown in Table 3.5 and 3.6, respectively. In the
negative control group (n=6), glycation-induced fructosamine formation was time-
dependently increased over the studied time periods and achieved significance on day-20
(p<0.05) and day-30 (p<0.01) compared to day-10 in both the glycation models. When
ginger was added to myoglobin-glucose model at the final concentrations of 100, 200 and
300 µg/mL, a significant (p<0.01; n=6) concentration-dependent reduction in fructosamine
formation was noted on day-20 and day30 compared to the negative control. On day-30 of
incubation, ginger significantly reduced (p<0.01; n=6) the fructosamine levels compared to
the negative control by 10.0%, 21.1% and 32.8% at the three concentrations studied, with an
IC50 value of 445.8 µg/mL (Figure 3.3C). Similarly, ginger (100, 200 and 300 µg/mL)
showed a significant concentration-dependent reduction (p<0.01) in fructosamine formation
on day-30 compared to the negative control by 14.2%, 19.6% and 25.2% in the myoglobin-
fructose glycation model, with an IC50 value of 764 µg/mL (Figure 3.3C). Furthermore,
these results are comparable to the positive control aminoguanidine (5 mmol/L) which
significantly (p<0.01) inhibited fructosamine formation on day-30 by 55.9% in the
myoglobin-glucose glycation model, and by 37.9% in the myoglobin-fructose glycation
model compared to the negative control.
.

74
Table 3.5 Effect of α-lipoic acid, curcumin and ginger on fructosamine formation in
myoglobin-glucose glycation
Experimental group Fructosamine (µmol/L)
Day-10 Day-20 Day-30
Negative control 759.52 ± 21.6 1131.80 ± 34.6# 1376.87 ± 27.8
##
α-Lipoic acid (1 mmol/L) 742.69 ± 18.19 959.69 ± 40.96* 1107.48 ± 13.12**
α-Lipoic acid (2 mmol/L) 617.35 ± 16.90* 795.07 ± 36.16** 984.01 ± 11.33**
α-Lipoic acid (4 mmol/L) 501.36 ± 28.34** 632.31 ± 71.15** 871.77 ± 19.77**
Curcumin (10 µmol/L) 723.98 ± 36.05 1088.78 ± 22.72 1201.02 ± 23.66**
Curcumin (20 µmol/L) 695.92 ± 28.98 946.60 ± 21.05** 1043.88 ± 25.44**
Curcumin (30 µmol/L) 662.24 ± 8.69 763.27 ± 32.67** 944.73 ± 17.35**
Ginger (100 µg/mL) 744.55 ± 42.2 1066.32 ± 23.2 1238.43 ± 11.3**
Ginger (200 µg/mL) 692.17 ± 29.7 896.08 ± 13.0** 1085.03 ± 39.3**
Ginger (300 µg/mL) 622.95 ± 43.1 825.00 ± 28.9** 924.14 ± 27.8**
Aminoguanidine (5 mmol/L) 445.24 ± 13.2** 493.88 ± 21.4** 606.12 ± 24.3**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05; ##p<0.01

75
Table 3.6 Effect of α-lipoic acid, curcumin and ginger on fructosamine formation in
myoglobin-fructose glycation
Experimental group Fructosamine (µmol/L)
Day-10 Day-20 Day-30
Negative control 1300.17 ± 29.6 1608.84 ± 34.4# 1754.76 ± 34.0
##
α-Lipoic acid (1 mmol/L) 1023.30 ± 36.70* 1227.21 ± 16.56** 1313.27 ± 16.39**
α-Lipoic acid (2 mmol/L) 991.50 ± 29.50** 1144.90 ± 32.00** 1275.85 ± 27.13**
α-Lipoic acid (4 mmol/L) 952.21 ± 15.98** 1100.00 ± 25.26** 1148.64 ± 33.50**
Curcumin (10 µmol/L) 1128.06 ± 33.79 1350.68 ± 25.86* 1404.93 ± 12.77**
Curcumin (20 µmol/L) 1006.46 ± 28.50** 1259.01 ± 32.19** 1307.65 ± 12.55**
Curcumin (30 µmol/L) 976.53 ± 43.14** 1154.25 ± 32.45** 1242.18 ± 11.83**
Ginger (100 µg/mL) 1204.76 ± 33.9 1457.31 ± 31.4* 1504.08 ± 22.8**
Ginger (200 µg/mL) 1135.54 ± 31.8** 1318.88 ± 33.3** 1410.54 ± 37.4**
Ginger (300 µg/mL) 1032.65 ± 33.8** 1234.69 ± 30.5** 1311/39 ± 13.7**
Aminoguanidine (5 mmol/L) 879.25 ± 14.0** 1013.95 ± 19.5** 1088.78 ± 24.2**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05; ##p<0.01
.

76
Figure 3.3 Percentage inhibition of fructosamine formation by α-lipoic acid (A),
curcumin (B) and ginger (C) in myoglobin-glucose (Mb-Glu) and myoglobin-fructose
(Mb-Fru) glycation after 30 days of incubation.

77
3.4. Effect of α-lipoic acid, curcumin and ginger on myoglobin glycation-induced
protein carbonyls formation.
Protein carbonyl content is the most commonly used biomarker of glycation-induced
oxidation of proteins. Thus, the elevation of protein carbonyls as an index of glycation-
induced protein oxidation was examined.
3.4.1. α-Lipoic acid
The effect of α-lipoic acid on protein carbonyl content of glycated myoglobin in myoglobin-
glucose and myoglobin-fructose glycation models are shown in Table 3.7 and 3.8,
respectively. In the negative control group (n=6), protein carbonyl content was time-
dependently increased over the studied time periods that achieved significance (p<0.05) on
day-20 and on day-30 compared to day-10 in both the glycation models. Adding α-lipoic
acid in the myoglobin-glucose glycation model at the final concentrations of 1, 2 and 4
mmol/L displayed a significant concentration-dependent reduction (p<0.05 to p<0.01; n=6)
in protein carbonyls at the corresponding time points compared to the negative control. On
day-30 of incubation, α-lipoic acid significantly reduced (p<0.01; n=6) the protein carbonyl
content compared to the negative control by 30.6%, 43.2%, and 52.6% at the three
concentrations studied, with an IC50 value of 3.4 mmol/L (Figure 3.4A). Similarly, α-lipoic
acid (1, 2 and 4 mmol/L) also showed a significant concentration-dependent reduction
(p<0.01) in protein carbonyls on day-30 compared to the negative control by 27.2%, 37.6%
and 41.9% in myoglobin-fructose glycation model ,with an IC50 value of 5.5 mmol/L
(Figure 3.4A). In comparison, the positive control aminoguanidine (5 mmol/L) also
significantly (p<0.01; n=6) inhibited the protein carbonyls formation on day-30 by 60.6% in

78
myoglobin-glucose glycation model and by 48.2% in myoglobin-fructose glycation model
compared to the negative control.
3.4.2. Curcumin
The effect of curcumin on protein carbonyl content of glycated myoglobin in the myoglobin-
glucose and myoglobin-fructose glycation models are shown in Table 3.7 and 3.8,
respectively. In the negative control group (n=6), the protein carbonyl content was time-
dependently increased over the studied time periods and achieved significance (p<0.05) on
day-20 and on day-30 compared to day-10 in both the glycation models. Adding curcumin
in the myoglobin-glucose glycation model at the final concentrations of 10, 20 and 30
µmol/L displayed a significant concentration-dependent reduction (p<0.05 to p<0.01; n=6)
in protein carbonyl content at the corresponding time points compared to the negative
control. On day-30 of incubation, curcumin significantly reduced (p<0.01) the protein
carbonyl content compared to the negative control by 25.3%, 37.2%, and 55.2% at the three
concentrations studied, with an IC50 value of 24.4 µmol/L (Figure 3.4B). Similarly,
curcumin (10, 20 and 30 µmol/L) showed a significant concentration-dependent reduction
(p<0.01; n=6) in protein carbonyl content on day-30 compared to the negative control by
23.6%, 28.7% and 37.0% in the myoglobin-fructose glycation model, with an IC50 value of
49.2 µmol/L (Figure 3.4B). In comparison, the positive control aminoguanidine (5 mmol/L)
also significantly (p<0.01; n=6) inhibited protein carbonyls formation on day-30 by 60.6%
in the myoglobin-glucose glycation model and by 48.2% in myoglobin-fructose glycation
model compared to the negative control.

79
3.4.3. Ginger
The effect of ginger on protein carbonyl content of glycated myoglobin in the myoglobin-
glucose and myoglobin-fructose glycation models are shown in Table 3.7 and 3.8,
respectively. In the negative control group (n=6), protein carbonyl content was time-
dependently increased over the studied time periods and achieved significance (p<0.05) on
day-20 and on day-30 compared to day-10 in both the glycation models. Adding ginger in
the myoglobin-glucose glycation model at the final concentrations of 100, 200 and 300
µg/mL displayed a significant concentration-dependent reduction (p<0.05 to p<0.01; n=6) in
the protein carbonyl content at the corresponding time points compared to the negative
control. On day-30 of incubation, ginger significantly reduced (p<0.05 to p<0.01; n=6) the
protein carbonyl content compared to the negative control by 14.0%, 26.3%, and 39.2% at
the three concentrations studied, with an IC50 value of 386.5 µg/mL (Figure 3.4C).
Similarly, ginger (100, 200 and 300 µg/mL) also showed a significant concentration-
dependent reduction (p<0.05 to p<0.01) in the protein carbonyl content on day-30 compared
to the negative control by 23.6%, 28.7% and 37.0% in the myoglobin-fructose glycation
model, with an IC50 value of 475 µg/mL (Figure 3.4C). In comparison, the positive control
aminoguanidine (5 mmol/L) also significantly (p<0.01; n=6) inhibited the protein carbonyl
content formation on day-30 by 60.6% in the myoglobin-glucose glycation model and by
48.2% in the myoglobin-fructose glycation model compared to the negative control.

80
Table 3.7 Effect of α-lipoic acid, curcumin and ginger on protein carbonyl content in
myoglobin-glucose glycation
Experimental group Protein carbonyl content (nmol/mL)
Day-10 Day-20 Day-30
Negative control 5.45 ± 0.17 6.84 ± 0.29# 8.17 ± 0.18
#
α-Lipoic acid (1 mmol/L) 4.48 ± 0.13* 4.99 ± 0.13* 5.67 ± 0.16**
α-Lipoic acid (2 mmol/L) 3.89 ± 0.08** 4.17 ± 0.21** 4.64 ± 0.17**
α-Lipoic acid (4 mmol/L) 3.08 ± 0.16** 3.24 ± 0.34** 4.36 ± 0.10**
Curcumin (10 µmol/L) 4.98 ± 0.19* 5.11 ± 0.20** 6.14 ± 0.12**
Curcumin (20 µmol/L) 4.33 ± 0.12** 4.58 ± 0.19** 5.13 ± 0.17**
Curcumin (30 µmol/L) 3.75 ± 0.06** 3.07 ± 0.23** 4.67 ± 0.22**
Ginger (100 µg/mL) 5.04 ± 0.19 5.74 ± 0.17* 7.02 ± 0.10*
Ginger (200 µg/mL) 4.52 ± 0.18* 4.66 ± 0.18** 6.01 ± 0.11**
Ginger (300 µg/mL) 4.02 ± 0.11* 4.58 ± 0.09** 4.96 ± 0.10**
Aminoguanidine (5 mmol/L) 2.89 ± 0.15** 2.95 ± 0.09** 3.21 ± 0.24**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05.

81
Table 3.8 Effect of α-lipoic acid, curcumin and ginger on protein carbonyl content in
myoglobin-fructose glycation
Experimental group Protein carbonyl content (nmol/mL)
Day-10 Day-20 Day-30
Negative control 7.09 ± 0.24 8.01 ± 0.09# 9.07 ± 0.12
#
α-Lipoic acid (1 mmol/L) 5.16 ± 0.09** 5.89 ± 0.12** 6.90 ± 0.13**
α-Lipoic acid (2 mmol/L) 4.42 ± 0.19** 5.53 ± 0.13** 5.98 ± 0.15**
α-Lipoic acid (4 mmol/L) 4.11 ± 0.25** 4.90 ± 0.13** 5.73 ± 0.23**
Curcumin (10 µmol/L) 5.75 ± 0.15** 6.14 ± 0.20** 7.62 ± 0.11**
Curcumin (20 µmol/L) 5.05 ± 0.19** 5.73 ± 0.13** 6.87 ± 0.15**
Curcumin (30 µmol/L) 4.46 ± 0.21** 5.23 ± 0.20** 6.51 ± 0.16**
Ginger (100 µg/mL) 6.08 ± 0.19* 7.05 ± 0.19* 8.08 ± 0.11*
Ginger (200 µg/mL) 5.15 ± 0.16* 6.22 ± 0.19** 7.03 ± 0.18**
Ginger (300 µg/mL) 4.61 ± 0.16** 5.75 ± 0.17** 6.21 ± 0.15**
Aminoguanidine (5 mmol/L) 3.86 ± 0.23** 4.15 ± 0.16** 4.70 ± 0.24**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05.

82
Figure 3.4 Percentage inhibition of protein carbonyl content by α-lipoic acid (A),
curcumin (B) and ginger (C) in myoglobin-glucose (Mb-Glu) and myoglobin-fructose
(Mb-Fru) glycation after 30 days of incubation

83
3.5. Effect of α-lipoic acid, curcumin and ginger on glycation-induced oxidation of
protein thiol group
Reactive oxygen species are generated during glycation and glycoxidation reactions, and are
able to oxidise side chains of amino acid residues in proteins to form carbonyl derivatives.
Decreased thiol groups diminish the oxidative defence of proteins. Thus, the determination
of protein thiol groups is a clear indication of protein oxidation occurring during protein
glycation.
3.5.1. α-Lipoic acid
The effect of α-lipoic acid on glycation-induce d oxidation of protein thiols in myoglobin-
glucose and myoglobin-fructose glycation models are shown in Table 3.9 and 3.10,
respectively. In the negative control group (n=6), the levels of protein thiols level were time-
dependently decreased over the studied time periods and achieved significance (p<0.05) on
day-30 compared to day-10 in both the glycation models. The addition of α-lipoic acid in the
myoglobin-glucose model at the final concentrations of 1, 2 and 4 mmol/L displayed a
significant concentration-dependent increase (p<0.05 to p<0.01; n=6) in protein thiols on
day-20 and day-30 compared to the negative control. On day-30 of incubation, α-lipoic acid
significantly increased (p<0.01) the protein thiol levels compared to the negative control by
30.4%, 39.5% and 54.7% at the three concentrations studied, with an IC50 value of 3.39
mmol/L (Figure 3.5A). Similarly, α-lipoic acid (1, 2 and 4 mmol/L) also showed a
significant concentration-dependent increase (p<0.01; n=6) in protein thiol levels on day-30
compared to the negative control by 55.3%, 69.35% and 74.8% in the myoglobin-fructose
glycation model, with an IC50 value of 0.7 mmol/L (Figure 3.5A). These results were
comparable to the positive control aminoguanidine (5 mmol/L) which also exhibited

84
significant increase (p<0.01; n=6) in protein thiols on day-30 by 76.1% in the myoglobin-
glucose glycation model and by 78.6% in the myoglobin-fructose glycation model compared
to the negative control.
3.5.2. Curcumin
The effect of curcumin on glycation-induced oxidation of protein thiols in myoglobin-
glucose and myoglobin-fructose models are shown in Table 3.9 and 3.10, respectively. In
the negative control group (n=6), protein thiols were time-dependently decreased over the
studied time periods that achieved significance (p<0.05) on day-30 compared to day-10 in
both the glycation models. The addition of curcumin in the myoglobin-glucose model at the
final concentrations of 10, 20 and 30 µmol/L displayed a significant concentration-
dependent increase (p<0.05 to p<0.01; n=6) in protein thiols on day-20 and day-30
compared to the negative control. On day-30 of incubation, curcumin significantly increased
(p<0.05 to p<0.01) the protein thiols level compared to the negative control by 27.9%,
52.4% and 58.5% at the three concentrations studied, with an IC50 value of 22.3 µmol/L
(Figure 3.5B). Similarly, curcumin (10, 20 and 30 µmol/L) also showed a significant
concentration-dependent increase (p<0.01) in protein thiols level on day-30 compared to the
negative control by 30.3%, 49% and 57% respectively in the myoglobin-fructose glycation
model, with an IC50 value of 23.5 µmol/L (Figure 3.5B). These results are comparable to
the positive control aminoguanidine (5 mmol/L) which exhibited a significant increase
(p<0.01) in protein thiols on day-30 by 76.1% in the myoglobin-glucose glycation model
and 78.6% in the myoglobin-fructose glycation model compared to the negative control.

85
3.5.3. Ginger
The effect of ginger on glycation-induced oxidation of protein thiols in myoglobin-glucose
and myoglobin-fructose models are shown in Table 3.9 and 3.10, respectively. In the
negative control group (n=6), protein thiols were time-dependently decreased over the
studied time periods and achieved significance (p<0.05) on day-30 compared to day-10 in
both the glycation models. The addition of ginger extract in the myoglobin-glucose model
with final concentrations of 100, 200 and 300 µg/mL displayed a significant concentration-
dependent improvement (p<0.05 to p<0.01; n=6) in protein thiols on day-20 and day-30
compared to the negative control. On day-30 of incubation, curcumin significantly increased
(p<0.05 to p<0.01; n=6) the protein thiols compared to the negative control by 23.8%,
34.1% and 55.3% at the three concentrations studied, with an IC50 value of 292.5 µg/mL
(Figure 3.5C). Similarly, ginger (100, 200 and 300 µg/mL) also showed a significant
concentration-dependent increase (p<0.01; n=6) in protein thiols on day-30 compared to the
negative control by 19.6%, 32.9% and 46.33% in myoglobin-fructose glycation model, with
an IC50 value of 328.7 µg/mL (Figure 3.5C). These results are comparable to the positive
control aminoguanidine (5 mmol/L) which exhibited significant increase (p<0.01; n=6) in
protein thiols on day-30 by 76.1% in the myoglobin-glucose glycation model and 78.6% in
the myoglobin-fructose glycation model compared to the negative control.

86
Table 3.9 Effect of α-lipoic acid, curcumin and ginger on the level of thiol group in
myoglobin-glucose glycation
Experimental group Thiol group (nmol/mg protein)
Day-10 Day-20 Day-30
Negative control 2.85 ± 0.21 2.21 ± 0.10 1.92 ± 0.21#
α-Lipoic acid (1 mmol/L) 3.15 ± 0.74 2.68 ± 0.12* 2.50 ± 0.06**
α-Lipoic acid (2 mmol/L) 3.20 ± 0.41 2.85 ± 0.60* 2.68 ± 0.51**
α-Lipoic acid (4 mmol/L) 3.56 ± 0.46 3.15 ± 0.33** 2.97 ± 0.24**
Curcumin (10 µmol/L) 3.09 ± 0.37 2.62 ± 0.06 2.44 ± 0.06*
Curcumin (20 µmol/L) 3.20 ± 0.21 3.03 ± 0.26* 2.91 ± 0.21**
Curcumin (30 µmol/L) 3.38 ± 0.31 3.20 ± 0.28** 3.03 ± 0.12**
Ginger (100 µg/mL) 3.03 ± 0.39 2.74 ± 0.10 2.21 ± 0.10
Ginger (200 µg/mL) 3.15 ± 0.06 2.80 ± 0.21 2.56 ± 0.10*
Ginger (300 µg/mL) 3.20 ± 0.16 3.09 ± 0.10* 2.97 ± 0.21*
Aminoguanidine (5 mmol/L) 4.61 ± 0.41** 3.73 ± 0.16** 3.44 ± 0.47**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05.

87
Table 3.10 Effect of α-lipoic acid, curcumin and ginger on the level of thiol group in
myoglobin-fructose glycation
Experimental group Thiol group (nmol/mg protein)
Day-10 Day-20 Day-30
Negative control 2.56 ± 0.10 2.42 ± 0.31 2.19 ± 0.26#
α-Lipoic acid (1 mmol/L) 3.79 ± 0.18* 3.50 ± 0.16* 3.26 ± 0.10**
α-Lipoic acid (2 mmol/L) 3.85 ± 0.24* 3.73 ± 0.33** 3.56 ± 0.33**
α-Lipoic acid (4 mmol/L) 3.96 ± 0.41** 3.85 ± 0.26** 3.67 ± 0.21**
Curcumin (10 µmol/L) 3.32 ± 0.26 3.20 ± 0.16 2.85 ± 0.16*
Curcumin (20 µmol/L) 3.56 ± 0.21 3.44 ± 0.31** 3.26 ± 0.10**
Curcumin (30 µmol/L) 3.67 ± 0.36 3.61 ± 0.27** 3.44 ± 0.10**
Ginger (100 µg/mL) 3.20 ± 0.36 3.09 ± 0.10 2.62 ± 0.15*
Ginger (200 µg/mL) 3.38 ± 0.16 3.15 ± 0.12 2.91 ± 0.20**
Ginger (300 µg/mL) 3.56 ± 0.16* 3.32 ± 0.21* 3.20 ± 0.21**
Aminoguanidine (5 mmol/L) 4.26 ± 0.93** 3.85 ± 0.85** 3.02 ± 0.82**
Values represent the mean ± SEM (n = 6). Significant difference compared to control at
identical times: * p< 0.05; **p<0.01. Significant difference from the respective Day-10
value: #p<0.05.

88
Figure 3.5 Percentage protection of protein thiol by α-lipoic acid (A), curcumin (B) and
ginger (C) in myoglobin-glucose (Mb-Glu) and myoglobin-fructose (Mb-Fru) glycation
after 30 days of incubation.

89
CHAPTER-4
DISCUSSION

90
Diabetes mellitus is a common endocrine disorder characterised by hyperglycaemia and
predisposes to chronic complications. Hyperglycaemia is a major culprit in the pathogenesis
of long-term complications in diabetic patients The four mechanisms that that have received
high appreciation which have been studied extensively in the past few decades include
activation of polyol pathways, activation of protein kinase C, increased hexosamine pathway
flux and increased formation of AGEs through protein glycation reactions (Brownlee,
Michael 2005). In particular, the role of protein glycation and the formation of AGEs in
diabetic complications have received considerable attention among scientific community.
Protein glycation, also known as the Maillard reaction, is a complex biochemical
reaction beginning with the non-enzymatic interaction of a reducing sugar (i.e. glucose or
fructose) or its derivative to an amine group of a protein (Rabbani et al. 2010). Molecular
rearrangements (Schiff base formation and Amadori rearrangements) lead to the formation
of AGEs that trigger pathogenic signalling pathways and cross-link extracellular matrix
proteins (Wells-Knecht et al. 1996). Furthermore, in hyperglycaemic condition, the polyol
pathway becomes active and facilitates the conversion of intracellular glucose to fructose
and thereby increases intracellular fructose levels. In the context of intracellular protein
glycation, the rate of fructose-induced glycation is faster than glucose-induced glycation and
thus, fructose and its metabolites are considered as important precursors in the intracellular
formation of AGEs (Ruderman, Williamson & Brownlee 1992; Sakai, Oimomi & Kasuga
2002; Suarez et al. 1989).
The AGEs have been shown to have a negative impact on the development of diabetic
cardiomyopathy (Kerkeni et al. 2013). It has been demonstrated that serum fluorescent
AGEs (such as pentosidine) was significantly higher in diabetic patients and also associated

91
with an increased incidence of cardiovascular disease (Sugiyama et al. 1998; Weiss et al.
1998). Moreover, AGEs can also cross-link with extracellular matrix proteins such as
collagen and increase arterial wall and myocardium stiffness which further leads to systolic
and diastolic dysfunction of heart and precipitate heart failure in diabetic patients (Yoshida,
Okumura & Aso 2005).
In addition to this, AGEs intracellularly couple with the receptors for advanced
glycation end products (RAGE). The exposure of AGEs to RAGE causes a significant delay
in calcium reuptake which leads to diastolic dysfunction by altering the duration of the re-
polarisation phase of the cardiac contraction (Petrova et al. 2002). Furthermore, AGEs are
also known to modify LDL creating foam cells and subsequently, play a role in the
development of atherosclerosis and possibly even MI (Bucala et al. 1994; Forbes et al. 2004;
Witztum & Steinberg 1991). Therefore, preventing the formation of AGEs or removal of
breaking of crosslink AGEs is an efficient way to interrupt the glycation cascade and
prevent the above mentioned potential pathological consequences of AGEs.
In the past, there has been a general lack of understanding of the biochemistry of
protein glycation and the associated role of AGEs in various diabetic complications.
Nevertheless, extensive studies on protein glycation and AGEs formation in the last decade
have now revealed the pathophysiological implications of AGEs in diabetic complications
(Ahmed, Nessar 2005; Jandeleit-Dahm & Cooper 2008; Li, D & Mehta 2005; Peppa &
Vlassara 2005). These complications include (but are not limited to) retinopathy (Ibrahim et
al. 2011), nephropathy (Thomas, Forbes & Cooper 2005), neuropathy (Sasaki et al. 1998)
and atherosclerosis (Del Turco & Basta 2012). Bookchin and Gallop first described the
glycation of haemoglobin, a heme protein found extensively in red blood cells (largely

92
known as an HbA1c) and its increase in persons with diabetes was reported by Rahbar
(Bookchin & Gallop 1968; Rahbar 1968). In the hyperglycaemic condition, glucose in the
blood reacts with haemoglobin via the Millard reaction which leads to irreversible
modification. Several studies demonstrated the glycation-induced structural and functional
modification of haemoglobin (Kar & Chakraborti 1999; Khoo et al. 1994; Watala,
Gwozdzinski & Malek 1992). Similarly, apolipoprotein B (a component of LDL) becomes
glycated in the hyperglycaemic environment and is involved in atherogenesis.
In cardiomyocytes, it is known that myoglobin acts as a short-time oxygen storage
protein where myoglobin reversibly binds with oxygen with help of iron and serves as an
oxygen reservoir (Hendgen-Cotta, Kelm & Rassaf 2014; Roy, Sen & Chakraborti 2004).
Myoglobin also acts as an intracellular scavenger of bioactive nitric oxide, regulating its
level in the cardiac and skeletal muscle and thereby protecting mitochondrial respiration,
which is impaired by nitric oxide (Brunori 2001; Flogel et al. 2001; Wunderlich et al. 2003).
As myoglobin is a vital protein in cardiomyocytes, any functional dysregulation may
precipitate pathological events. In a high glucose environment, myoglobin reacts with sugar
through the Millard reaction to form AGEs which may compromise its functions (Hendgen-
Cotta, Kelm & Rassaf 2014; Roy, Sen & Chakraborti 2004). Recently, Banerjee and
Chakraborti demonstrated that myoglobin incubated with methylglyoxal, a known AGEs
precursor, induced structural modifications of myoglobin and generated various AGEs
(Banerjee & Chakraborti 2013; Banerjee & Chakraborti 2014). Furthermore, the glycation of
myoglobin also released iron from the heme moiety which may reduce the binding capacity
of cellular oxygen with heart myoglobin in vivo (Roy, Sen & Chakraborti 2004; Roy, Sil &
Chakraborti 2010). Thus, reduced oxygen supply to cardiac myocytes will result in cardiac

93
ischaemia and necrosis. In addition, the liberated free iron can produce free radicals which
have been suggested to contribute to the pathogenesis of diabetic cardiovascular
complications (Sil et al. 2004).
The protective effects of various natural products against diabetes have been
predominantly studied with reference to their hypoglycaemic activity. Information regarding
whether natural products can protect against diabetic complications, particularly the
prevention of glycation and AGEs formation is limited. In the last decade, a plethora of
natural products have been reported to alleviate various diabetic complications by inhibiting
protein glycation and subsequently AGEs formation (Lin, Tang, Kang & Chen 2012;
Saraswat et al. 2010). These latter studies clearly suggest that, as well as having a
hypoglycaemic property, natural products can protect against diabetic complications in other
ways such as the suppression of protein glycation pathway and the generation of harmful
AGEs.
In light of the above mentioned background, the proposed research project examined
the preventive effects of three different natural products, including α-lipoic acid, curcumin
and ginger extract, on glucose and fructose-induced myoglobin glycation and AGEs
formation using in vitro glycation models. To our knowledge, this is the first study which
investigates the protective effects of α-lipoic acid, curcumin and ginger on glucose and
fructose-induced myoglobin glycation.
4.1. Anti-glycation activity of α-lipoic acid
Protein glycation depends on the degree and duration of hyperglycaemia. The Amadori
products (fructosamine) undergo further rearrangement to form both fluorescent and non-
fluorescent AGEs. Various fluorescent AGEs such as pentosidine and crossline are more

94
likely to be implicated in the development of diabetic cardiovascular complications (Ahmed,
N. 2005). Based on the fluorescence property of AGEs, the effect of α-lipoic acid on the
formation of fluorescent AGEs was studied. Our results demonstrated that α-lipoic acid
efficiently inhibited glucose- and fructose- mediated fluorescent AGEs formation which
supports previously reported inhibitory effects of α-lipoic acid on fluorescent AGEs
formation in vivo (Veresiu 2009). The possible mechanism of action of α-lipoic acid in
inhibiting the formation of fluorescent AGEs include the following and could be at one or
more steps in the biochemical pathways of AGEs generation, i.e., (i) blocking the amino
groups of protein, thus preventing the glycation by free sugar, (ii) blocking the carbonyl
groups of reducing sugars, (iii) preventing the formation of Amadori products by blocking
the Schiff’s base to Amadori products conversion, (iv) blocking the Amadori products and
dicarbonyl intermediates which may reduce glycation, as well as AGEs formation, and/or
(v) preventing autoxidation of sugars and glyoxidation of Amadori products.
In the early stage of glycation, unstable Schiff’s bases are formed and turned into Amadori
products such as fructosamine, which are clinically used as an indicator for short term
control of blood sugar in diabetic patients (Ardestani & Yazdanparast 2007). The reduction
of fructosamine, therefore, is a therapeutic way to delay the occurrence of vascular
complications (Shield et al. 1994). In the present investigation, α-lipoic acid displayed a
significant, time- dependant inhibition of fructosamine formation. α-Lipoic acid may inhibit
the formation of Amadori products by competing with sugar molecules or protecting the
protein amino group from nucleophilic addition of the carbonyl group of sugar. Moreover,
the inhibition of fructosamine formation in the early glycation stage will be beneficial in the
suppression of AGEs formation and subsequent harmful effects of AGEs. Aminoguanidine,

95
a hydrazine derivative, acts by blocking the formation of Amadori products or AGEs
(Khalifah, Baynes & Hudson 1999). In the present study, aminoguanidine also effectively
inhibited the formation of Amadori products and thereby, prevented the formation of AGEs.
In comparison to aminoguanidine, α-lipoic acid displayed similar results and could have a
similar mechanism.
Furthermore, Roy and co-workers demonstrated that in vitro non-enzymatic
glycation of myoglobin induces the release of free iron from the heme pocket of myoglobin,
and the iron release was found to be proportional to the extent of myoglobin glycation (Roy,
Sen & Chakraborti 2004). The released iron, can produce free radicals, particularly hydroxyl
(OH-) radicals, which increases oxidative stress and subsequently causes necrosis of tissue
(Sil et al. 2004). In these studies, similar results were found whereby myoglobin-glucose or
myoglobin-fructose glycation effectively released free iron from the heme moiety of
myoglobin in a time-dependant manner in the negative control group. The addition of α-
lipoic acid suppressed free iron released from myoglobin due to glucose or fructose-
mediated glycation. Additionally, preventing the release of iron from myoglobin with α-
lipoic acid may also preserve its oxygen-carrying capacity and thereby, protect
cardiomyocytes from ischaemia.
Aside from the formation of AGEs, reactive carbonyl intermediaries and protein
carbonyl derivatives also cause protein modifications that are particularly prone to oxidative
reaction to amino acid. ROS are generated during glycation and glycoxidation and are able
to oxidise side chains of amino acid residues in protein to form carbonyl derivatives, and
also diminish the oxidative defence of protein by decreasing thiol groups (Shield et al. 1994;
Singh et al. 2001). Thus, these phenomena are reflective of high oxidative stress, protein

96
oxidative damage and the formation of AGEs, which are a direct reflection of the excess of
free radical generation. In the present investigation, the significant elevation of protein
carbonyl content and oxidation of thiols (reduced total thiol pool) in myoglobin-glucose and
myoglobin-fructose glycation models were observed. In contrast, when α-lipoic acid was
added to the same glycation models, the compound significantly suppressed formation of
protein carbonyl content and oxidation of thiols (increased total thiol pool). Furthermore,
during the early stage of glycation, Schiff bases are prone to oxidation, generating free
radicals, and reactive carbonyl compounds (Miyata 2002; Miyata et al. 1999). Scavenging
hydroxyl radicals and superoxide radicals can alleviate oxidative stress and reduce the
generation of reactive carbonyl compounds (Wu et al. 2011). α-Lipoic acid is a potent
biological antioxidant and is able to scavenge various free radicals such as hydroxyl groups
(Shay et al. 2009) (Perricone et al. 1999). Therefore, the possible mechanism of α-lipoic acid
in suppressing the formation of protein dicarbonyl is scavenging the highly reactive free
radicals generated during chronic glycation.
4.2. Anti-glycation activity of curcumin
In a present study, the effect of curcumin on glucose and fructose-induced glycation of
myoglobin protein was investigated. In this study we found that curcumin inhibited the
formation of fructosamine-derived fluorescent AGEs in myoglobin-glucose and myoglobin-
fructose glycation. Previously reported in vivo studies showed that curcumin reduced serum
AGEs level by inducing AGE-R1 (AGE receptor-1) gene expression which is responsible
for the detoxification and clearance of AGEs (Lin, Tang, Kang & Chen 2012). Curcumin
also suppressed the gene expression of RAGE by elevating the activity of PPAR-γ and

97
attenuating oxidative stress, leading to the elimination of the AGEs (Lin, Tang, Kang, Feng,
et al. 2012).
Furthermore, it was found that the ferrozine-detected free iron level was increased
proportionally to glucose and fructose-induced myoglobin glycation in vitro, while curcumin
produced a significant reduction in the free iron release from myoglobin and thereby
diminishing harmful effect of free iron. The possible consequence of free iron, apart from
increasing oxidative stress, is the reduced binding capacity of cellular oxygen with heart
myoglobin in vivo resulting in cardiac ischaemia and necrosis (Kyada 2012).
In this study, curcumin attenuated the glycation-induced protein carbonyls formation and
subsequently, blocked the conversion of dicarbonyl intermediates to AGEs. The finding is in
agreement with previously reported studies where curcumin significantly lowered protein
carbonyls and subsequently, overcame oxidative stress (Dkhar & Sharma 2010;
Kukongviriyapan et al. 2014; Suryanarayana et al. 2005). Dicarbonyl intermediates have
received considerable attention as mediators of AGEs formation and are known to react with
lysine, arginine and cysteine residues in proteins to form protein cross-links (Monnier 2003).
Therefore, the suppression of carbonyl/dicarbonyl compound formation could be beneficial
in reduction of AGEs
Furthermore, curcumin inhibited high glucose or fructose-induced oxidative damages
to myoglobin protein by preserving protein thiols groups. This indicates that curcumin may
be effective in preventing oxidative protein damages by decreasing oxidative stress which is
believed to occur during the glycation processes. The results showed a significant decrease
in the number of free thiol groups when myoglobin was incubated with glucose or fructose
(negative control group). Himmelfarb et al demonstrated that curcumin, as an anti-oxidative

98
therapy, showed significantly reduced plasma protein oxidation through the inhibition of the
carbonyls formation and preserving the protein thiol groups (Himmelfarb et al. 2004).
4.3. Anti-glycation activity of ginger extract
In this study, the concentration and time- dependent influence of ethanolic extracts of ginger
rhizomes on glucose and fructose- mediated myoglobin glycation in vitro was investigated.
Myoglobin-glucose and myoglobin-fructose incubation at different time periods
(negative control groups) showed a significant increase in fluorescent AGEs formation and
liberation of free iron from myoglobin. The present study revealed that the ethanolic extract
of ginger possessed marked anti-glycation property by suppressing the fructosamine
formation, protein carbonyls formation and protecting protein thiols group oxidation.
In this study, fluorescent AGEs were measured in myoglobin-glucose and
myoglobin-fructose glycation. The results showed that ginger extract efficiently inhibited
fluorescent AGEs formations which supports previously reported anti-AGEs activity of
ginger extract (Rani et al. 2012). In addition, ginger extract significantly inhibited glycation-
induced release of free iron from myoglobin in a time and concentration dependent manner.
These liberated free iron participate in generating free radicals, particularly hydroxyl (OH)
radicals, which can damage various cellular constituents. Moreover, the iron released from
the destruction of the heme moiety of myoglobin also enhances AGEs formation and
elevates the level of free radicals, which have been suggested to contribute to the
pathogenesis of diabetic cardiovascular complications (Sil et al. 2004).
Kazeem et al. found that ginger extract inhibited protein glycation by scavenging free
radical scavenging. This may also inhibit fructosamine formation by modifying the amino

99
group of the protein or carbonyl groups of glucose/fructose in the Maillard reaction (Kazeem
et al. 2012). These studies are in agreement with the results obtained in our study. It was
found that ginger extract markedly inhibited the fructosamine formation. Ginger extract may
act as an anti-oxidant which further efficiently removed fructosamine adducts and scavenges
the free radical generated during conversion of fructosamine to AGEs (Stoilova et al. 2007).
The determination of the carbonyl content and thiol groups was used to assess the
protein oxidation that occurs during the process of glycation (Webster et al. 2005). In this
study, the incubation of myoglobin with glucose/fructose significantly elevated the protein
carbonyls formation in the reaction media. The ginger extract significantly inhibited the
glucose and fructose-induced protein carbonyl formation which is consistent with previously
reported inhibitory effects of ginger extract on the formation of protein carbonyls (Khadem
Ansari et al. 2008; Saraswat et al. 2009). The elevated levels of protein carbonyl content as
an index of glycation-induced protein oxidation have been reported in several diseases.
Carbonyl content is the most general and commonly used biomarker of long-term carbonyl
overload or ‘‘carbonyl stress’’ (Dalle-Donne et al. 2003). It has been proposed that carbonyl
stress, i.e. the increase in reactive carbonyl compounds derived from oxidative and non–
oxidative reactions, leads to increased chemical modification of proteins and, at a later stage,
oxidative stress and tissue damage (Baynes & Thorpe 1999). Such oxidative modification of
protein is an index of oxidative stress and may play a significant role in diabetic
complications pathology (Webster et al. 2005).
The determination of free thiol groups in myoglobin was performed according to the
Ellman’s method. The results showed a significant decrease in the free thiol groups when
myoglobin was incubated with glucose or fructose. However, ginger extract in the presence

100
of glucose or fructose significantly inhibited the oxidation of these thiol groups due to its
anti-oxidant activity. These results are supported by a previous study regarding protein
oxidation in brain regions during ethanol withdrawal under the influence of ginger extract
treatment prior to the abstinence from chronic ethanol consumption (Marella 2012).
The chemical characterisation showed that the ethanolic ginger extract contained 6-
shogaol as the major constituent and followed the order 6-shogaol > 6-gingerol > 8-gingerol.
The data obtained from chemical characterisation suggests that 6-shogaol could be the major
bioactive constituent responsible for the observed anti-glycation activity including inhibition
of formation of fluorescent AGEs, reduction in fructosamine formation, reduced the
liberation of free iron and prevent protein oxidation by reducing protein carbonyl content
and thiol oxidation. Therefore, 6-shogaol and 6-gingerol should be investigated for their
potential benefits in diabetic cardiovascular complication, including inhibiting myoglobin
glycation and formation of advanced glycation of end products.
Collectively, all three selected natural products, including α-lipoic acid, curcumin and ginger
extract, showed marked reduction in glucose and fructose-induced myoglobin glycation. The
possible anti-glycation mechanisms of all three natural compounds include breaking the
cross-linking structures in the formed AGEs, the reduced levels of fructosamine adducts
formation, reduced free iron release, blocking the carbonyl or dicarbonyl groups in reducing
sugars and protecting the thiol group oxidation of myoglobin (Figure 4.1.)

101
Figure 4.1 Schematic representations of pathways induced by myoglobin glycation and
the inhibitory effects of α-lipoic acid, curcumin and ginger extract on this pathway.

102
CHAPTER-5
CONCLUSION AND FUTURE
DIRECTION

103
5.1. General conclusion
Protein glycation has received substantial concern in the development of diabetic
complications. Myoglobin is a cellular protein mainly found in cardiac myocardium and
serves as an oxygen reservoir and releases its bound oxygen for cardiac mitochondrial ATP
synthesis during the oxidative phosphorylation. Previous reports suggest that glycated
myoglobin (with glucose or fructose) in circulation may cause a serious threat with respect
to oxidative damage, which may, in turn, aggravate the pathophysiology of diabetic
complications. This could have dire consequences in the pathogenesis of diabetic
complications such as atherosclerosis and MI.
In most studies, glucose-induced protein glycation and subsequent AGEs formation
has been emphasised. However, fructose-induced glycation is a highly probable event in
hyperglycaemic conditions as the conversion of glucose to fructose via the polyol pathway
becomes more active. Thus, in the present study, fructose-induced myoglobin glycation was
also studied
This thesis examined two main pathways in the glycation of myoglobin:
(1) Glucose and
(2) Fructose.
Although there are many compounds that possess anti-glycation activity, none are used in
clinical practice. Much of the pharmacological needs of today’s chronic diseases require the
use of agents with multiple bioactivities. Herbal supplements have long been used and
accepted primarily as complementary treatments for managing diabetes, and unless
individual phytoconstituents that possess both hypoglycaemic and anti-oxidative properties
can be identified, they are likely to retain their adjunct role in medicinal therapy.

104
In the present study, three natural products including α-lipoic acid, curcumin and
ginger extract were selected, and their effectiveness in the prevention of myoglobin
glycation (under chronic high glucose and high fructose) and AGEs formation in vitro was
explored.
In previous literature, α-lipoic acid, curcumin and ginger extract have previously
shown anti-diabetic, hypolipidaemic, as well as antioxidant activity. In this proposed thesis,
α-lipoic acid, curcumin and ginger extract have been evaluated first time for their anti-
glycation property on myocardial myoglobin protein.
These findings demonstrated that α-lipoic acid, curcumin and ginger extract
protected against fructose- and glucose-mediated myoglobin glycation in vitro. Additionally,
α-lipoic acid, curcumin and ginger extract reduced the level of fructosamine formation.
Furthermore, all three natural products decreased the release of free iron from the heme
moiety and protein carbonyl content modification in myoglobin. The beneficial effects of
these natural products may be applied to the prevention or management of AGE-mediated
pathologies; particularly chronic hyperglycaemic-induced cardiovascular complications such
as MI.
5.2. Limitations
The limitations to this study include:
Comparing the ginger extract to other extracts in literature, especially if chemical
profiling is absent.
The main compounds of the extract were not examined to determine their
effectiveness in the assays.

105
The dose-response curve did not have six doses and thus, a R2 value would not
reflect the dose increase.
An extract/or compound which is not known for its antioxidant effect should have
been examined for its anti-AGEs activity.
Incubation of myoglobin with AGEs precursor (such as glyoxal and methylglyoxal)
which would give more insight of the anti-glycation mechanism of the proposed
compounds.
5.3. Future directions
Future work should focus on:
Investigating the anti-glycation activity of active phytoconstituents of ginger extract
Whether the observed changes can be reproduced in cardiomyocyte cell line when
incubated with high glucose or fructose
Additional research in animal models is required to establish the role of myoglobin
glycation in the pathogenesis of hyperglycaemic-induced MI and the protective role
of α-lipoic acid, curcumin and ethanolic extract of ginger in myoglobin glycation.
Which physiological dosages would be feasible in clinical practice

106
BIBLIOGRAPHY

107
Aggarwal, BB, Kumar, A & Bharti, AC 2003, 'Anticancer potential of curcumin:
preclinical and clinical studies', Anticancer Res, vol. 23, no. 1A, pp. 363-98.
Ahmed, N 2005, 'Advanced glycation endproducts—role in pathology of diabetic
complications', Diabetes Research and Clinical Practice, vol. 67, no. 1, pp. 3-21.
Ajith, TA 2010, 'Ameliorating reactive oxygen species-induced in vitro lipid peroxidation
in brain, liver, mitochondria and DNA damage by Zingiber officinale Roscoe', Indian
Journal of Clinical Biochemistry, vol. 25, no. 1, pp. 67-73.
Akhani, SP, Vishwakarma, SL & Goyal, RK 2004, 'Anti-diabetic activity of Zingiber
officinale in streptozotocin-induced type I diabetic rats', J Pharm Pharmacol, vol. 56, no.
1, pp. 101-5.
Akihiko S, MY, Satoshi H, Takayuki N, Masafumi I, Seigo K, Yoshikazu Y 2013, 'Anti-
Glycation Activity of Alpha-Lipoic Acid Derivatives and Vitamin E Derivatives', Anti-
Aging Medicine, vol. 10, pp. 42-54.
Al-Abed, Y & Bucala, R 2000, 'Structure of a synthetic glucose derived advanced
glycation end product that is immunologically cross-reactive with its naturally occurring
counterparts', Bioconjug Chem, vol. 11, no. 1, pp. 39-45.
Alam, J & Cook, JL 2003, 'Transcriptional regulation of the heme oxygenase-1 gene via
the stress response element pathway', Curr Pharm Des, vol. 9, no. 30, pp. 2499-511.
Ames, J 1998, 'Applications of the Maillard reaction in the food industry', Food
Chemistry, vol. 62, no. 4, pp. 431-9.
Anonymous 1998, 'Intensive blood-glucose control with sulphonylureas or insulin
compared with conventional treatment and risk of complications in patients with type 2
diabetes (UKPDS 33). UK Prospective Diabetes Study (UKPDS) Group', Lancet, vol.
352, no. 9131, pp. 837-53.
Ansari, MN, Bhandari, U & Pillai, KK 2006, 'Ethanolic Zingiber officinale R. extract
pretreatment alleviates isoproterenol-induced oxidative myocardial necrosis in rats',
Indian J Exp Biol, vol. 44, no. 11, pp. 892-7.
Ardestani, A & Yazdanparast, R 2007, 'Cyperus rotundus suppresses AGE formation and
protein oxidation in a model of fructose-mediated protein glycoxidation', Int J Biol
Macromol, vol. 41, no. 5, pp. 572-8.
Banerjee, S & Chakraborti, A 2013, 'In Vitro Study on Structural Alteration of
Myoglobin by Methylglyoxal', The Protein Journal, vol. 32, no. 3, pp. 216-22.
Banerjee, S & Chakraborti, AS 2014, 'Structural alterations of hemoglobin and
myoglobin by glyoxal: A comparative study', International Journal of Biological
Macromolecules, vol. 66, no. 0, pp. 311-8.

108
Baynes, JW & Thorpe, SR 1999, 'Role of oxidative stress in diabetic complications: a
new perspective on an old paradigm', Diabetes, vol. 48, no. 1, pp. 1-9.
Bhandari, U, Kanojia, R & Pillai, KK 2005, 'Effect of ethanolic extract of Zingiber
officinale on dyslipidaemia in diabetic rats', J Ethnopharmacol, vol. 97, no. 2, pp. 227-
30.
Biemel, KM, Friedl, DA & Lederer, MO 2002, 'Identification and quantification of major
maillard cross-links in human serum albumin and lens protein. Evidence for glucosepane
as the dominant compound', J Biol Chem, vol. 277, no. 28, pp. 24907-15.
Boel, E, Selmer, J, Flodgaard, HJ & Jensen, T 1995, 'Diabetic late complications: will
aldose reductase inhibitors or inhibitors of advanced glycosylation endproduct formation
hold promise?', J Diabetes Complications, vol. 9, no. 2, pp. 104-29.
Bonnefont-Rousselot, D 2002, 'Glucose and reactive oxygen species', Curr Opin Clin
Nutr Metab Care, vol. 5, no. 5, pp. 561-8.
Bookchin, RM & Gallop, PM 1968, 'Structure of hemoglobin AIc: nature of the N-
terminal beta chain blocking group', Biochem Biophys Res Commun, vol. 32, no. 1, pp.
86-93.
Bousova, I, Vukasovic, D, Juretic, D, Palicka, V & Drsata, J 2005, 'Enzyme activity and
AGE formation in a model of AST glycoxidation by D-fructose in vitro', Acta Pharm,
vol. 55, no. 1, pp. 107-14.
Bray, GA 2013, 'Energy and Fructose From Beverages Sweetened With Sugar or High-
Fructose Corn Syrup Pose a Health Risk for Some People', Advances in Nutrition: An
International Review Journal, vol. 4, no. 2, pp. 220-5.
Brett, J, Schmidt, AM, Yan, SD, Zou, YS, Weidman, E, Pinsky, D, Nowygrod, R,
Neeper, M, Przysiecki, C, Shaw, A & et al. 1993, 'Survey of the distribution of a newly
characterized receptor for advanced glycation end products in tissues', Am J Pathol, vol.
143, no. 6, pp. 1699-712.
Brownlee, M 1994, 'Lilly Lecture 1993. Glycation and diabetic complications', Diabetes,
vol. 43, no. 6, pp. 836-41.
Brownlee, M 2005, 'The Pathobiology of Diabetic Complications: A Unifying
Mechanism', Diabetes, vol. 54, no. 6, pp. 1615-25.
Brownlee, M, Cerami, A & Vlassara, H 1988, 'Advanced glycosylation end products in
tissue and the biochemical basis of diabetic complications', N Engl J Med, vol. 318, no.
20, pp. 1315-21.
Brunori, M 2001, 'Nitric oxide moves myoglobin centre stage', Trends Biochem Sci, vol.
26, no. 4, pp. 209-10.

109
Bucala, R, Makita, Z, Vega, G, Grundy, S, Koschinsky, T, Cerami, A & Vlassara, H
1994, 'Modification of low density lipoprotein by advanced glycation end products
contributes to the dyslipidemia of diabetes and renal insufficiency', Proc Natl Acad Sci U
S A, vol. 91, no. 20, pp. 9441-5.
Candido, R, Forbes, JM, Thomas, MC, Thallas, V, Dean, RG, Burns, WC, Tikellis, C,
Ritchie, RH, Twigg, SM, Cooper, ME & Burrell, LM 2003, 'A breaker of advanced
glycation end products attenuates diabetes-induced myocardial structural changes', Circ
Res, vol. 92, no. 7, pp. 785-92.
Chavakis, T, Bierhaus, A & Nawroth, PP 2004, 'RAGE (receptor for advanced glycation
end products): a central player in the inflammatory response', Microbes Infect, vol. 6, no.
13, pp. 1219-25.
Chen, SA, Chen, HM, Yao, YD, Hung, CF, Tu, CS & Liang, YJ 2012, 'Topical treatment
with anti-oxidants and Au nanoparticles promote healing of diabetic wound through
receptor for advance glycation end-products', Eur J Pharm Sci, vol. 47, no. 5, pp. 875-83.
Chen, WL, Kang, CH, Wang, SG & Lee, HM 2012, 'alpha-Lipoic acid regulates lipid
metabolism through induction of sirtuin 1 (SIRT1) and activation of AMP-activated
protein kinase', Diabetologia, vol. 55, no. 6, pp. 1824-35.
Connell, D & Sutherland, M 1969, 'A re-examination of gingerol, shogaol, and zingerone,
the pungent principles of ginger (<I>Zingiber officinale</I> Roscoe)', Australian Journal
of Chemistry, vol. 22, no. 5, pp. 1033-43.
Dalle-Donne, I, Rossi, R, Giustarini, D, Milzani, A & Colombo, R 2003, 'Protein
carbonyl groups as biomarkers of oxidative stress', Clin Chim Acta, vol. 329, no. 1-2, pp.
23-38.
De Rosa, MC, Sanna, MT, Messana, I, Castagnola, M, Galtieri, A, Tellone, E, Scatena,
R, Botta, B, Botta, M & Giardina, B 1998, 'Glycated human hemoglobin (HbA1c):
functional characteristics and molecular modeling studies', Biophysical Chemistry, vol.
72, no. 3, pp. 323-35.
Degenhardt, TP, Alderson, NL, Arrington, DD, Beattie, RJ, Basgen, JM, Steffes, MW,
Thorpe, SR & Baynes, JW 2002, 'Pyridoxamine inhibits early renal disease and
dyslipidemia in the streptozotocin-diabetic rat', Kidney Int, vol. 61, no. 3, pp. 939-50.
Dekker, MJ, Su, Q, Baker, C, Rutledge, AC & Adeli, K 2010, Fructose: a highly
lipogenic nutrient implicated in insulin resistance, hepatic steatosis, and the metabolic
syndrome, vol. 299.
Del Turco, S & Basta, G 2012, 'An update on advanced glycation endproducts and
atherosclerosis', Biofactors, vol. 38, no. 4, pp. 266-74.
Devamanoharan, PS, Ali, AH & Varma, SD 1997, 'Prevention of lens protein glycation
by taurine', Mol Cell Biochem, vol. 177, no. 1-2, pp. 245-50.

110
Dkhar, P & Sharma, R 2010, 'Effect of dimethylsulphoxide and curcumin on protein
carbonyls and reactive oxygen species of cerebral hemispheres of mice as a function of
age', Int J Dev Neurosci, vol. 28, no. 5, pp. 351-7.
Duge de Bernonville, T, Guyot, S, Paulin, JP, Gaucher, M, Loufrani, L, Henrion, D,
Derbre, S, Guilet, D, Richomme, P, Dat, JF & Brisset, MN 2010, 'Dihydrochalcones:
Implication in resistance to oxidative stress and bioactivities against advanced glycation
end-products and vasoconstriction', Phytochemistry, vol. 71, no. 4, pp. 443-52.
Dyer, DG, Dunn, JA, Thorpe, SR, Bailie, KE, Lyons, TJ, McCance, DR & Baynes, JW
1993, 'Accumulation of Maillard reaction products in skin collagen in diabetes and aging',
Journal of Clinical Investigation, vol. 91, no. 6, pp. 2463-9.
El-Azab, MF, Attia, FM & El-Mowafy, AM 2011, 'Novel role of curcumin combined
with bone marrow transplantation in reversing experimental diabetes: Effects on
pancreatic islet regeneration, oxidative stress, and inflammatory cytokines', Eur J
Pharmacol, vol. 658, no. 1, pp. 41-8.
Ellman, GL 1959, 'Tissue sulfhydryl groups', Arch Biochem Biophys, vol. 82, no. 1, pp.
70-7.
Farmar, JG, Ulrich, PC & Cerami, A 1988, 'Novel pyrroles from sulfite-inhibited
Maillard reactions: insight into the mechanism of inhibition', The Journal of Organic
Chemistry, vol. 53, no. 10, pp. 2346-9.
Federici, M, Menghini, R, Mauriello, A, Hribal, ML, Ferrelli, F, Lauro, D, Sbraccia, P,
Spagnoli, LG, Sesti, G & Lauro, R 2002, 'Insulin-dependent activation of endothelial
nitric oxide synthase is impaired by O-linked glycosylation modification of signaling
proteins in human coronary endothelial cells', Circulation, vol. 106, no. 4, pp. 466-72.
Figarola, JL, Scott, S, Loera, S, Tessler, C, Chu, P, Weiss, L, Hardy, J & Rahbar, S 2003,
'LR-90 a new advanced glycation endproduct inhibitor prevents progression of diabetic
nephropathy in streptozotocin-diabetic rats', Diabetologia, vol. 46, no. 8, pp. 1140-52.
Figarola, JL, Shanmugam, N, Natarajan, R & Rahbar, S 2007, 'Anti-inflammatory effects
of the advanced glycation end product inhibitor LR-90 in human monocytes', Diabetes,
vol. 56, no. 3, pp. 647-55.
Fioretto, P, Steffes, MW, Sutherland, DE, Goetz, FC & Mauer, M 1998, 'Reversal of
lesions of diabetic nephropathy after pancreas transplantation', N Engl J Med, vol. 339,
no. 2, pp. 69-75.
Fiuza, SM, Gomes, C, Teixeira, LJ, Girao da Cruz, MT, Cordeiro, MN, Milhazes, N,
Borges, F & Marques, MP 2004, 'Phenolic acid derivatives with potential anticancer
properties--a structure-activity relationship study. Part 1: methyl, propyl and octyl esters
of caffeic and gallic acids', Bioorg Med Chem, vol. 12, no. 13, pp. 3581-9.

111
Flogel, U, Godecke, A, Klotz, LO & Schrader, J 2004, 'Role of myoglobin in the
antioxidant defense of the heart', Faseb j, vol. 18, no. 10, pp. 1156-8.
Flogel, U, Merx, MW, Godecke, A, Decking, UK & Schrader, J 2001, 'Myoglobin: A
scavenger of bioactive NO', Proc Natl Acad Sci U S A, vol. 98, no. 2, pp. 735-40.
Foley, RN, Murray, AM, Li, S, Herzog, CA, McBean, AM, Eggers, PW & Collins, AJ
2005, 'Chronic kidney disease and the risk for cardiovascular disease, renal replacement,
and death in the United States Medicare population, 1998 to 1999', J Am Soc Nephrol,
vol. 16, no. 2, pp. 489-95.
Forbes, JM, Yee, LT, Thallas, V, Lassila, M, Candido, R, Jandeleit-Dahm, KA, Thomas,
MC, Burns, WC, Deemer, EK, Thorpe, SR, Cooper, ME & Allen, TJ 2004, 'Advanced
glycation end product interventions reduce diabetes-accelerated atherosclerosis',
Diabetes, vol. 53, no. 7, pp. 1813-23.
Frye, EB, Degenhardt, TP, Thorpe, SR & Baynes, JW 1998, 'Role of the Maillard
reaction in aging of tissue proteins. Advanced glycation end product-dependent increase
in imidazolium cross-links in human lens proteins', J Biol Chem, vol. 273, no. 30, pp.
18714-9.
Fuhrman, B, Rosenblat, M, Hayek, T, Coleman, R & Aviram, M 2000, 'Ginger extract
consumption reduces plasma cholesterol, inhibits LDL oxidation and attenuates
development of atherosclerosis in atherosclerotic, apolipoprotein E-deficient mice', J
Nutr, vol. 130, no. 5, pp. 1124-31.
Ghibu, S, Lauzier, B, Delemasure, S, Amoureux, S, Sicard, P, Vergely, C, Muresan, A,
Mogosan, C & Rochette, L 2009, 'Antioxidant properties of alpha-lipoic acid: effects on
red blood membrane permeability and adaptation of isolated rat heart to reversible
ischaemia', Mol Cell Biochem, vol. 320, no. 1-2, pp. 141-8.
Giardino, I, Edelstein, D & Brownlee, M 1994, 'Nonenzymatic glycosylation in vitro and
in bovine endothelial cells alters basic fibroblast growth factor activity. A model for
intracellular glycosylation in diabetes', J Clin Invest, vol. 94, no. 1, pp. 110-7.
Goh, SY & Cooper, ME 2008, 'Clinical review: The role of advanced glycation end
products in progression and complications of diabetes', J Clin Endocrinol Metab, vol. 93,
no. 4, pp. 1143-52.
Goldin, A, Beckman, JA, Schmidt, AM & Creager, MA 2006, 'Advanced glycation end
products: sparking the development of diabetic vascular injury', Circulation, vol. 114, no.
6, pp. 597-605.
Govindarajan, VS 1982a, 'Ginger--chemistry, technology, and quality evaluation: part 1',
Crit Rev Food Sci Nutr, vol. 17, no. 1, pp. 1-96.
—— 1982b, 'Ginger-chemistry, technology, and quality evaluation: part 2', Crit Rev
Food Sci Nutr, vol. 17, no. 3, pp. 189-258.

112
Gugliucci, A, Bastos, DH, Schulze, J & Souza, MF 2009, 'Caffeic and chlorogenic acids
in Ilex paraguariensis extracts are the main inhibitors of AGE generation by
methylglyoxal in model proteins', Fitoterapia, vol. 80, no. 6, pp. 339-44.
Ha, HC, Sirisoma, NS, Kuppusamy, P, Zweier, JL, Woster, PM & Casero, RA, Jr. 1998,
'The natural polyamine spermine functions directly as a free radical scavenger', Proc Natl
Acad Sci U S A, vol. 95, no. 19, pp. 11140-5.
Habib, ZA, Havstad, SL, Wells, K, Divine, G, Pladevall, M & Williams, LK 2010,
'Thiazolidinedione use and the longitudinal risk of fractures in patients with type 2
diabetes mellitus', J Clin Endocrinol Metab, vol. 95, no. 2, pp. 592-600.
Hardy, J, Parmentier, M & Fanni, J 1999, 'Functionality of nutrients and thermal
treatments of food', Proc Nutr Soc, vol. 58, no. 3, pp. 579-85.
Hartog, JW, Smit, AJ, van Son, WJ, Navis, G, Gans, RO, Wolffenbuttel, BH & de Jong,
PE 2004, 'Advanced glycation end products in kidney transplant patients: a putative role
in the development of chronic renal transplant dysfunction', Am J Kidney Dis, vol. 43, no.
6, pp. 966-75.
Hartog, JW, Voors, AA, Bakker, SJ, Smit, AJ & van Veldhuisen, DJ 2007, 'Advanced
glycation end-products (AGEs) and heart failure: pathophysiology and clinical
implications', Eur J Heart Fail, vol. 9, no. 12, pp. 1146-55.
Hendgen-Cotta, UB, Kelm, M & Rassaf, T 2014, 'Myoglobin functions in the heart', Free
Radical Biology and Medicine, vol. 73, no. 0, pp. 252-9.
Himmelfarb, J, McMonagle, E, Freedman, S, Klenzak, J, McMenamin, E, Le, P, Pupim,
LB, Ikizler, TA & Group, TP 2004, 'Oxidative Stress Is Increased in Critically Ill Patients
with Acute Renal Failure', Journal of the American Society of Nephrology, vol. 15, no. 9,
pp. 2449-56.
Hunt, JV, Bottoms, MA & Mitchinson, MJ 1993, 'Oxidative alterations in the
experimental glycation model of diabetes mellitus are due to protein-glucose adduct
oxidation: Some fundamental differences in proposed mechanisms of glucose oxidation
and oxidant production', Biochemical Journal, vol. 291, no. 2, pp. 529-35.
Hunt, JV, Dean, RT & Wolff, SP 1988, 'Hydroxyl radical production and autoxidative
glycosylation. Glucose autoxidation as the cause of protein damage in the experimental
glycation model of diabetes mellitus and ageing', Biochem J, vol. 256, no. 1, pp. 205-12.
Ibrahim, AS, El-Remessy, AB, Matragoon, S, Zhang, W, Patel, Y, Khan, S, Al-Gayyar,
MM, El-Shishtawy, MM & Liou, GI 2011, 'Retinal Microglial Activation and
Inflammation Induced by Amadori-Glycated Albumin in a Rat Model of Diabetes',
Diabetes, vol. 60, no. 4, pp. 1122-33.

113
Jain, SK, Rains, J & Jones, K 2006, 'Effect of curcumin on protein glycosylation, lipid
peroxidation, and oxygen radical generation in human red blood cells exposed to high
glucose levels', Free Radical Biology and Medicine, vol. 41, no. 1, pp. 92-6.
Jandeleit-Dahm, K & Cooper, ME 2008, 'The role of AGEs in cardiovascular disease',
Curr Pharm Des, vol. 14, no. 10, pp. 979-86.
Jono, T, Kimura, T, Takamatsu, J, Nagai, R, Miyazaki, K, Yuzuriha, T, Kitamura, T &
Horiuchi, S 2002, 'Accumulation of imidazolone, pentosidine and N(epsilon)-
(carboxymethyl)lysine in hippocampal CA4 pyramidal neurons of aged human brain',
Pathol Int, vol. 52, no. 9, pp. 563-71.
Kanatous, SB, Mammen, PP, Rosenberg, PB, Martin, CM, White, MD, Dimaio, JM,
Huang, G, Muallem, S & Garry, DJ 2009, 'Hypoxia reprograms calcium signaling and
regulates myoglobin expression', Am J Physiol Cell Physiol, vol. 296, no. 3, pp. C393-
402.
Kar, M & Chakraborti, AS 1999, 'Release of iron from haemoglobin--a possible source of
free radicals in diabetes mellitus', Indian J Exp Biol, vol. 37, no. 2, pp. 190-2.
Kashiwagi, A, Obata, T, Suzaki, M, Takagi, Y, Kida, Y, Ogawa, T, Tanaka, Y, Asahina,
T, Ikebuchi, M, Saeki, Y & et al. 1992, 'Increase in cardiac muscle fructose content in
streptozotocin-induced diabetic rats', Metabolism, vol. 41, no. 10, pp. 1041-6.
Kazeem, MI, Akanji, MA, Hafizur, RM & Choudhary, MI 2012, 'Antiglycation,
antioxidant and toxicological potential of polyphenol extracts of alligator pepper, ginger
and nutmeg from Nigeria', Asian Pac J Trop Biomed, vol. 2, no. 9, pp. 727-32.
Kerkeni, M, Saïdi, A, Bouzidi, H, Letaief, A, Ben Yahia, S & Hammami, M 2013,
'Pentosidine as a biomarker for microvascular complications in type 2 diabetic patients',
Diabetes and Vascular Disease Research, vol. 10, no. 3, pp. 239-45.
Khadem Ansari, M, Karimipour, M, Salami, S & Shirpoor, A 2008, 'The Effect of Ginger
(Zingiber officinale) on Oxidative Stress Status in the Small Intestine of Diabetic Rats',
Int J Endocrinol Metab, vol. 6, no. 3, pp. 144-50.
Khalifah, RG, Baynes, JW & Hudson, BG 1999, 'Amadorins: novel post-Amadori
inhibitors of advanced glycation reactions', Biochem Biophys Res Commun, vol. 257, no.
2, pp. 251-8.
Khoo, UY, Newman, DJ, Miller, WK & Price, CP 1994, 'The influence of glycation on
the peroxidase activity of haemoglobin', Eur J Clin Chem Clin Biochem, vol. 32, no. 6,
pp. 435-40.
Kiho, T, Usui, S, Hirano, K, Aizawa, K & Inakuma, T 2004, 'Tomato paste fraction
inhibiting the formation of advanced glycation end-products', Biosci Biotechnol Biochem,
vol. 68, no. 1, pp. 200-5.

114
Kim, JM, Lee, EK, Kim, DH, Yu, BP & Chung, HY 2010, 'Kaempferol modulates pro-
inflammatory NF-kappaB activation by suppressing advanced glycation endproducts-
induced NADPH oxidase', Age (Dordr), vol. 32, no. 2, pp. 197-208.
Kim, JM, Lee, YM, Lee, GY, Jang, DS, Bae, KH & Kim, JS 2006, 'Constituents of the
roots of Pueraria lobata inhibit formation of advanced glycation end products (AGEs)', A
Kim, KM, Jung, DH, Jang, DS, Kim, YS, Kim, JM, Kim, HN, Surh, YJ & Kim, JS 2010,
'Puerarin suppresses AGEs-induced inflammation in mouse mesangial cells: a possible
pathway through the induction of heme oxygenase-1 expression', Toxicol Appl
Pharmacol, vol. 244, no. 2, pp. 106-13.
Kolm-Litty, V, Sauer, U, Nerlich, A, Lehmann, R & Schleicher, ED 1998, 'High glucose-
induced transforming growth factor beta1 production is mediated by the hexosamine
pathway in porcine glomerular mesangial cells', J Clin Invest, vol. 101, no. 1, pp. 160-9.
Koya, D, Jirousek, MR, Lin, YW, Ishii, H, Kuboki, K & King, GL 1997,
'Characterization of protein kinase C beta isoform activation on the gene expression of
transforming growth factor-beta, extracellular matrix components, and prostanoids in the
glomeruli of diabetic rats', J Clin Invest, vol. 100, no. 1, pp. 115-26.
Koya, D & King, GL 1998, 'Protein kinase C activation and the development of diabetic
complications', Diabetes, vol. 47, no. 6, pp. 859-66.
Kukongviriyapan, U, Pannangpetch, P, Kukongviriyapan, V, Donpunha, W, Sompamit,
K & Surawattanawan, P 2014, 'Curcumin protects against cadmium-induced vascular
dysfunction, hypertension and tissue cadmium accumulation in mice', Nutrients, vol. 6,
no. 3, pp. 1194-208.
Kyada, C 2012, 'A Protective Role of Telmisartan and Metformin Against Myoglobin
Glycation in Isoproterenol-induced Acute Myocardial Infarction in Diabetic Rats', Master
of Pharmacy in Pharmacology thesis, Gujarat Technological University.
Lee, AY & Chung, SS 1999, 'Contributions of polyol pathway to oxidative stress in
diabetic cataract', Faseb j, vol. 13, no. 1, pp. 23-30.
Lee, EH, Song, DG, Lee, JY, Pan, CH, Um, BH & Jung, SH 2008, 'Inhibitory effect of
the compounds isolated from Rhus verniciflua on aldose reductase and advanced
glycation endproducts', Biol Pharm Bull, vol. 31, no. 8, pp. 1626-30.
Levine, RL, Williams, JA, Stadtman, ER & Shacter, E 1994, 'Carbonyl assays for
determination of oxidatively modified proteins', Methods Enzymol, vol. 233, pp. 346-57.
Li, D & Mehta, JL 2005, 'Oxidized LDL, a critical factor in atherogenesis',
Cardiovascular Research, vol. 68, no. 3, pp. 353-4.
Li, X-H, McGrath, KCY, Nammi, S, Heather, AK & Roufogalis, BD 2012, 'Attenuation
of Liver Pro-Inflammatory Responses by Zingiber officinale via Inhibition of NF-kappa

115
B Activation in High-Fat Diet-Fed Rats', Basic & Clinical Pharmacology & Toxicology,
vol. 110, no. 3, pp. 238-44.
Lin, J, Tang, Y, Kang, Q & Chen, A 2012, 'Curcumin eliminates the inhibitory effect of
advanced glycation end-products (AGEs) on gene expression of AGE receptor-1 in
hepatic stellate cells in vitro', Lab Invest, vol. 92, no. 6, pp. 827-41.
Lin, J, Tang, Y, Kang, Q & Chen, A 2012, 'Curcumin eliminates the inhibitory effect of
advanced glycation end-products (AGEs) on gene expression of AGE receptor-1 in
hepatic stellate cells in vitro', Lab Invest, vol. 92, no. 6, pp. 827-41.
Lin, J, Tang, Y, Kang, Q, Feng, Y & Chen, A 2012, 'Curcumin inhibits gene expression
of receptor for advanced glycation end-products (RAGE) in hepatic stellate cells in vitro
by elevating PPARgamma activity and attenuating oxidative stress', Br J Pharmacol, vol.
166, no. 8, pp. 2212-27.
Lin, J, Tang, Y, Kang, Q, Feng, Y & Chen, A 2012, 'Curcumin inhibits gene expression
of receptor for advanced glycation end-products (RAGE) in hepatic stellate cells in vitro
by elevating PPARγ activity and attenuating oxidative stress', British Journal of
Pharmacology, vol. 166, no. 8, pp. 2212-27.
Liu, JP, Feng, L, Zhu, MM, Wang, RS, Zhang, MH, Hu, SY, Jia, XB & Wu, JJ 2012,
'The in vitro protective effects of curcumin and demethoxycurcumin in Curcuma longa
extract on advanced glycation end products-induced mesangial cell apoptosis and
oxidative stress', Planta Med, vol. 78, no. 16, pp. 1757-60.
Maillard, L 1912, 'Action of Amino Acids on Sugars. Formation of Melanoidins in a
Methodical Way.', Compt. Rend, vol. 154, pp. 66-71.
Marella, S, Reddy KS 2012, 'Evaluation of Protein Oxidation in Brain Regions During
Ethanol Withdrawal under the Influence of Ginger Extract Treatment Prior to Abstinence
from Chronic Ethanol Consumption', European Journal of Biological Sciences, vol. 4,
pp. 136-45.
Meerwaldt, R, Hartog, JW, Graaff, R, Huisman, RJ, Links, TP, den Hollander, NC,
Thorpe, SR, Baynes, JW, Navis, G, Gans, RO & Smit, AJ 2005, 'Skin autofluorescence, a
measure of cumulative metabolic stress and advanced glycation end products, predicts
mortality in hemodialysis patients', J Am Soc Nephrol, vol. 16, no. 12, pp. 3687-93.
Mercer, DW 1997, 'Role of cardiac markers in evaluation of suspected myocardial
infarction. Selecting the most clinically useful indicators', Postgrad Med, vol. 102, no. 5,
pp. 113-7, 21-2.
Miroliaei, M, Khazaei, S, Moshkelgosha, S & Shirvani, M 2011, 'Inhibitory effects of
Lemon balm (Melissa officinalis, L.) extract on the formation of advanced glycation end
products', Food Chemistry, vol. 129, no. 2, pp. 267-71.

116
Miyata, T 2002, 'Alterations of non-enzymatic biochemistry in uremia, diabetes, and
atherosclerosis ("carbonyl stress")', Bull Mem Acad R Med Belg, vol. 157, no. 3-4, pp.
189-96; discussion 96-8.
Miyata, T, van Ypersele de Strihou, C, Kurokawa, K & Baynes, JW 1999, 'Alterations in
nonenzymatic biochemistry in uremia: origin and significance of "carbonyl stress" in
long-term uremic complications', Kidney Int, vol. 55, no. 2, pp. 389-99.
Monnier, VM 2003, 'Intervention against the Maillard reaction in vivo', Arch Biochem
Biophys, vol. 419, no. 1, pp. 1-15.
Mullarkey, CJ, Edelstein, D & Brownlee, M 1990, 'Free radical generation by early
glycation products: a mechanism for accelerated atherogenesis in diabetes', Biochem
Biophys Res Commun, vol. 173, no. 3, pp. 932-9.
Mustafa, T & Srivastava, KC 1990, 'Ginger (Zingiber officinale) in migraine headache', J
Ethnopharmacol, vol. 29, no. 3, pp. 267-73.
Nakagawa, T, Yokozawa, T, Terasawa, K, Shu, S & Juneja, LR 2002, 'Protective activity
of green tea against free radical- and glucose-mediated protein damage', J Agric Food
Chem, vol. 50, no. 8, pp. 2418-22.
Nakamura, S, Makita, Z, Ishikawa, S, Yasumura, K, Fujii, W, Yanagisawa, K, Kawata, T
& Koike, T 1997, 'Progression of nephropathy in spontaneous diabetic rats is prevented
by OPB-9195, a novel inhibitor of advanced glycation', Diabetes, vol. 46, no. 5, pp. 895-
9.
Nilsson, BO 1999, 'Biological effects of aminoguanidine: an update', Inflamm Res, vol.
48, no. 10, pp. 509-15.
O'Keefe, JH, Abuannadi, M, Lavie, CJ & Bell, DS 2011, 'Strategies for optimizing
glycemic control and cardiovascular prognosis in patients with type 2 diabetes mellitus',
Mayo Clin Proc, vol. 86, no. 2, pp. 128-38.
Ohkawara, E, Nohara, Y, Kanno, Y, Suzuki, H, Matsumoto, G, Kinoshita, T &
Watanabe, M 2002, 'Fructosamine assay using albumin extracted from serum', Biol
Pharm Bull, vol. 25, no. 9, pp. 1121-4.
Ordway, GA & Garry, DJ 2004, 'Myoglobin: an essential hemoprotein in striated muscle',
J Exp Biol, vol. 207, no. Pt 20, pp. 3441-6.
Panter, SS 1994, 'Release of iron from hemoglobin', Methods Enzymol, vol. 231, pp. 502-
14.
Penna, SC, Medeiros, MV, Aimbire, FS, Faria-Neto, HC, Sertie, JA & Lopes-Martins,
RA 2003, 'Anti-inflammatory effect of the hydralcoholic extract of Zingiber officinale
rhizomes on rat paw and skin edema', Phytomedicine, vol. 10, no. 5, pp. 381-5.

117
Peppa, M & Vlassara, H 2005, 'Advanced glycation end products and diabetic
complications: a general overview', Hormones (Athens), vol. 4, no. 1, pp. 28-37.
Perricone, N, Nagy, K, Horvath, F, Dajko, G, Uray, I & Zs -Nagy, I 1999, 'Alpha lipoic
acid (ALA) protects proteins against the hydroxyl free radical-induced alterations:
rationale for its geriatric topical application', Arch Gerontol Geriatr, vol. 29, no. 1, pp.
45-56.
Petrova, R, Yamamoto, Y, Muraki, K, Yonekura, H, Sakurai, S, Watanabe, T, Li, H,
Takeuchi, M, Makita, Z, Kato, I, Takasawa, S, Okamoto, H, Imaizumi, Y & Yamamoto,
H 2002, 'Advanced glycation endproduct-induced calcium handling impairment in mouse
cardiac myocytes', J Mol Cell Cardiol, vol. 34, no. 10, pp. 1425-31.
Rabbani, N, Chittari, MV, Bodmer, CW, Zehnder, D, Ceriello, A & Thornalley, PJ 2010,
'Increased glycation and oxidative damage to apolipoprotein B100 of LDL cholesterol in
patients with type 2 diabetes and effect of metformin', Diabetes, vol. 59, no. 4, pp. 1038-
45.
Rahbar, S 1968, 'An abnormal hemoglobin in red cells of diabetics', Clin Chim Acta, vol.
22, no. 2, pp. 296-8.
Rani, MP, Krishna, MS, Padmakumari, KP, Raghu, KG & Sundaresan, A 2012, 'Zingiber
officinale extract exhibits antidiabetic potential via modulating glucose uptake, protein
glycation and inhibiting adipocyte differentiation: an in vitro study', Journal of the
Science of Food and Agriculture, vol. 92, no. 9, pp. 1948-55.
Reddy, S, Bichler, J, Wells-Knecht, KJ, Thorpe, SR & Baynes, JW 1995, 'Nε-
(carboxymethyl)lysine is a dominant advanced glycation end product (AGE) antigen in
tissue proteins', Biochemistry, vol. 34, no. 34, pp. 10872-8.
Reihl, O, Rothenbacher, TM, Lederer, MO & Schwack, W 2004, 'Carbohydrate carbonyl
mobility––the key process in the formation of α-dicarbonyl intermediates', Carbohydrate
Research, vol. 339, no. 9, pp. 1609-18.
Rossi, R, Giustarini, D, Milzani, A & Dalle-Donne, I 2009, 'Cysteinylation and
homocysteinylation of plasma protein thiols during ageing of healthy human beings', J
Cell Mol Med, vol. 13, no. 9B, pp. 3131-40.
Roy, A, Sen, S & Chakraborti, AS 2004, 'In vitro nonenzymatic glycation enhances the
role of myoglobin as a source of oxidative stress', Free Radic Res, vol. 38, no. 2, pp. 139-
46.
Roy, A, Sil, R & Chakraborti, AS 2010, 'Non-enzymatic glycation induces structural
modifications of myoglobin', Mol Cell Biochem, vol. 338, no. 1-2, pp. 105-14.
Ruderman, NB, Williamson, JR & Brownlee, M 1992, 'Glucose and diabetic vascular
disease', Faseb j, vol. 6, no. 11, pp. 2905-14.

118
Sajithlal, GB, Chithra, P & Chandrakasan, G 1998, 'Effect of curcumin on the advanced
glycation and cross-linking of collagen in diabetic rats', Biochem Pharmacol, vol. 56, no.
12, pp. 1607-14.
Sakai, M, Oimomi, M & Kasuga, M 2002, 'Experimental studies on the role of fructose in
the development of diabetic complications', Kobe J Med Sci, vol. 48, pp. 125 - 36.
Saraswat, M, Reddy, PY, Muthenna, P & Reddy, GB 2009, 'Prevention of non-enzymic
glycation of proteins by dietary agents: prospects for alleviating diabetic complications',
Br J Nutr, vol. 101, no. 11, pp. 1714-21.
Saraswat, M, Suryanarayana, P, Reddy, PY, Patil, MA, Balakrishna, N & Reddy, GB
2010, 'Antiglycating potential of Zingiber officinalis and delay of diabetic cataract in
rats', Mol Vis, vol. 16, pp. 1525-37.
Sasaki, N, Fukatsu, R, Tsuzuki, K, Hayashi, Y, Yoshida, T, Fujii, N, Koike, T,
Wakayama, I, Yanagihara, R, Garruto, R, Amano, N & Makita, Z 1998, 'Advanced
glycation end products in Alzheimer's disease and other neurodegenerative diseases', Am
J Pathol, vol. 153, no. 4, pp. 1149-55.
Sayeski, PP & Kudlow, JE 1996, 'Glucose metabolism to glucosamine is necessary for
glucose stimulation of transforming growth factor-alpha gene transcription', J Biol Chem,
vol. 271, no. 25, pp. 15237-43.
Schalkwijk, CG, Stehouwer, CD & van Hinsbergh, VW 2004, 'Fructose-mediated non-
enzymatic glycation: sweet coupling or bad modification', Diabetes Metab Res Rev, vol.
20, no. 5, pp. 369-82.
Schleicher, ED, Olgemoller, B, Wiedenmann, E & Gerbitz, KD 1993, 'Specific glycation
of albumin depends on its half-life', Clin Chem, vol. 39, no. 4, pp. 625-8.
Sekhar, RV, McKay, SV, Patel, SG, Guthikonda, AP, Reddy, VT, Balasubramanyam, A
& Jahoor, F 2011, 'Glutathione Synthesis Is Diminished in Patients With Uncontrolled
Diabetes and Restored by Dietary Supplementation With Cysteine and Glycine', Diabetes
Care, vol. 34, no. 1, pp. 162-7.
Sen, S, Bose, T, Roy, A & Chakraborti, AS 2007, 'Effect of non-enzymatic glycation on
esterase activities of hemoglobin and myoglobin', Mol Cell Biochem, vol. 301, no. 1-2,
pp. 251-7.
Sharma, RA, Gescher, AJ & Steward, WP 2005, 'Curcumin: the story so far', Eur J
Cancer, vol. 41, no. 13, pp. 1955-68.
Shay, KP, Moreau, RF, Smith, EJ, Smith, AR & Hagen, TM 2009, 'Alpha-lipoic acid as a
dietary supplement: molecular mechanisms and therapeutic potential', Biochim Biophys
Acta, vol. 1790, no. 10, pp. 1149-60.

119
Shield, JP, Poyser, K, Hunt, L & Pennock, CA 1994, 'Fructosamine and glycated
haemoglobin in the assessment of long term glycaemic control in diabetes', Arch Dis
Child, vol. 71, no. 5, pp. 443-5.
Shishodia, S, Sethi, G & Aggarwal, BB 2005, 'Curcumin: getting back to the roots', Ann
N Y Acad Sci, vol. 1056, pp. 206-17.
Sil, S, Bose, T, Roy, D & Chakraborti, AS 2004, 'Protoporphyrin IX-induced structural
and functional changes in human red blood cells, haemoglobin and myoglobin', J Biosci,
vol. 29, no. 3, pp. 281-91.
Singh, R, Barden, A, Mori, T & Beilin, L 2001, 'Advanced glycation end-products: a
review', Diabetologia, vol. 44, no. 2, pp. 129-46.
Skovsted, IC, Christensen, M, Breinholt, J & Mortensen, SB 1998, 'Characterisation of a
novel AGE-compound derived from lysine and 3-deoxyglucosone', Cell Mol Biol (Noisy-
le-grand), vol. 44, no. 7, pp. 1159-63.
Smit, AJ & Lutgers, HL 2004, 'The clinical relevance of advanced glycation endproducts
(AGE) and recent developments in pharmaceutics to reduce AGE accumulation', Curr
Med Chem, vol. 11, no. 20, pp. 2767-84.
Smith, MA, Taneda, S, Richey, PL, Miyata, S, Yan, SD, Stern, D, Sayre, LM, Monnier,
VM & Perry, G 1994, 'Advanced Maillard reaction end products are associated with
Alzheimer disease pathology', Proc Natl Acad Sci U S A, vol. 91, no. 12, pp. 5710-4.
Srivastava, KC & Mustafa, T 1989, 'Ginger (Zingiber officinale) and rheumatic
disorders', Med Hypotheses, vol. 29, no. 1, pp. 25-8.
Stewart, JM, Kilpatrick, ES, Cathcart, S, Small, M & Dominiczak, MH 1994, 'Low-
density lipoprotein particle size in type 2 diabetic patients and age matched controls', Ann
Clin Biochem, vol. 31 ( Pt 2), pp. 153-9.
Stitt, A, Gardiner, TA, Alderson, NL, Canning, P, Frizzell, N, Duffy, N, Boyle, C,
Januszewski, AS, Chachich, M, Baynes, JW & Thorpe, SR 2002, 'The AGE inhibitor
pyridoxamine inhibits development of retinopathy in experimental diabetes', Diabetes,
vol. 51, no. 9, pp. 2826-32.
Stoilova, I, Krastanov, A, Stoyanova, A, Denev, P & Gargova, S 2007, 'Antioxidant
activity of a ginger extract (Zingiber officinale)', Food Chemistry, vol. 102, no. 3, pp.
764-70.
Suarez, G, Rajaram, R, Oronsky, AL & Gawinowicz, MA 1989, 'Nonenzymatic glycation
of bovine serum albumin by fructose (fructation). Comparison with the Maillard reaction
initiated by glucose', J Biol Chem, vol. 264, no. 7, pp. 3674-9.

120
Sugiyama, S, Miyata, T, Ueda, Y, Tanaka, H, Maeda, K, Kawashima, S, Van Ypersele de
Strihou, C & Kurokawa, K 1998, 'Plasma levels of pentosidine in diabetic patients: an
advanced glycation end product', J Am Soc Nephrol, vol. 9, no. 9, pp. 1681-8.
Sun, L-Q, Chen, Y-Y, Wang, X, Li, X-J, Xue, B, Qu, L, Zhang, T-T, Mu, Y-M & Lu, J-
M 2012, 'The protective effect of Alpha lipoic acid on Schwann cells exposed to constant
or intermittent high glucose', Biochemical Pharmacology, vol. 84, no. 7, pp. 961-73.
Suryanarayana, P, Saraswat, M, Mrudula, T, Krishna, TP, Krishnaswamy, K & Reddy,
GB 2005, 'Curcumin and turmeric delay streptozotocin-induced diabetic cataract in rats',
Invest Ophthalmol Vis Sci, vol. 46, no. 6, pp. 2092-9.
Tanaka, N, Yonekura, H, Yamagishi, S, Fujimori, H, Yamamoto, Y & Yamamoto, H
2000, 'The receptor for advanced glycation end products is induced by the glycation
products themselves and tumor necrosis factor-alpha through nuclear factor-kappa B, and
by 17beta-estradiol through Sp-1 in human vascular endothelial cells', J Biol Chem, vol.
275, no. 33, pp. 25781-90.
Tessier, FJ 2010, 'The Maillard reaction in the human body. The main discoveries and
factors that affect glycation', Pathol Biol (Paris), vol. 58, no. 3, pp. 214-9.
Thomas, MC, Forbes, JM & Cooper, ME 2005, 'Advanced glycation end products and
diabetic nephropathy', Am J Ther, vol. 12, no. 6, pp. 562-72.
Thomson, M, Al-Qattan, KK, Al-Sawan, SM, Alnaqeeb, MA, Khan, I & Ali, M 2002,
'The use of ginger (Zingiber officinale Rosc.) as a potential anti-inflammatory and
antithrombotic agent', Prostaglandins Leukot Essent Fatty Acids, vol. 67, no. 6, pp. 475-
8.
Tsuji-Naito, K, Saeki, H & Hamano, M 2009, 'Inhibitory effects of Chrysanthemum
species extracts on formation of advanced glycation end products', Food Chemistry, vol.
116, no. 4, pp. 854-9.
Turk, Z, Misur, I, Turk, N & Benko, B 1999, 'Rat tissue collagen modified by advanced
glycation: correlation with duration of diabetes and glycemic control', Clin Chem Lab
Med, vol. 37, no. 8, pp. 813-20.
Vaidya, H 1994, 'Myoglobin: an early biochemical marker for the diagnosis of acute
myocardial infarction.', J Clin Immunoassay, vol. 17, pp. 35-9.
Vasu, VT, Modi, H, Thaikoottathil, JV & Gupta, S 2005, 'Hypolipidaemic and
antioxidant effect of Enicostemma littorale Blume aqueous extract in cholesterol fed rats',
J Ethnopharmacol, vol. 101, no. 1-3, pp. 277-82.
Veresiu, I 2009, 'Intravenous alpha-lipoic acid improves fructosamine level in type 2
diabetes patients ', Acta Endocrinol (Buc), vol. 5, pp. 501-6.

121
Verzelloni, E, Tagliazucchi, D, Del Rio, D, Calani, L & Conte, A 2011, 'Antiglycative
and antioxidative properties of coffee fractions', Food Chemistry, vol. 124, no. 4, pp.
1430-5.
Wang, N-P, Wang, Z-F, Tootle, S, Philip, T & Zhao, Z-Q 2012, 'Curcumin promotes
cardiac repair and ameliorates cardiac dysfunction following myocardial infarction',
British Journal of Pharmacology, vol. 167, no. 7, pp. 1550-62.
Watala, C, Gwozdzinski, K & Malek, M 1992, 'Direct evidence for the alterations in
protein structure and conformation upon in vitro nonenzymatic glycosylation', Int J
Biochem, vol. 24, no. 8, pp. 1295-302.
Webster, J, Urban, C, Berbaum, K, Loske, C, Alpar, A, Gartner, U, de Arriba, SG,
Arendt, T & Munch, G 2005, 'The carbonyl scavengers aminoguanidine and tenilsetam
protect against the neurotoxic effects of methylglyoxal', Neurotox Res, vol. 7, no. 1-2, pp.
95-101.
Weiss, MF, Rodby, RA, Justice, AC & Hricik, DE 1998, 'Free pentosidine and neopterin
as markers of progression rate in diabetic nephropathy. Collaborative Study Group',
Kidney Int, vol. 54, no. 1, pp. 193-202.
Wells-Knecht, KJ, Brinkmann, E, Wells-Knecht, MC, Litchfield, JE, Ahmed, MU,
Reddy, S, Zyzak, DV, Thorpe, SR & Baynes, JW 1996, 'New biomarkers of Maillard
reaction damage to proteins', Nephrol Dial Transplant, vol. 11 Suppl 5, pp. 41-7.
Witztum, JL & Steinberg, D 1991, 'Role of oxidized low density lipoprotein in
atherogenesis', J Clin Invest, vol. 88, no. 6, pp. 1785-92.
Wolff, SP & Dean, RT 1987, 'Glucose autoxidation and protein modification. The
potential role of 'autoxidative glycosylation' in diabetes', Biochemical Journal, vol. 245,
no. 1, pp. 243-50.
Wright, CE, Tallan, HH, Lin, YY & Gaull, GE 1986, 'Taurine: biological update', Annu
Rev Biochem, vol. 55, pp. 427-53.
Wu, C-H, Huang, S-M, Lin, J-A & Yen, G-C 2011, 'Inhibition of advanced glycation
endproduct formation by foodstuffs', Food & Function, vol. 2, no. 5, pp. 224-34.
Wunderlich, C, Flogel, U, Godecke, A, Heger, J & Schrader, J 2003, 'Acute inhibition of
myoglobin impairs contractility and energy state of iNOS-overexpressing hearts', Circ
Res, vol. 92, no. 12, pp. 1352-8.
Xia, P, Inoguchi, T, Kern, TS, Engerman, RL, Oates, PJ & King, GL 1994,
'Characterization of the mechanism for the chronic activation of diacylglycerol-protein
kinase C pathway in diabetes and hypergalactosemia', Diabetes, vol. 43, no. 9, pp. 1122-
9.

122
Xiang, M, Qian, ZY, Zhou, CH, Liu, J & Li, WN 2006, 'Crocetin inhibits leukocyte
adherence to vascular endothelial cells induced by AGEs', J Ethnopharmacol, vol. 107,
no. 1, pp. 25-31.
Yamagishi, S & Matsui, T 2010, 'Advanced glycation end products, oxidative stress and
diabetic nephropathy', Oxid Med Cell Longev, vol. 3, no. 2, pp. 101-8.
Yamaguchi, F, Ariga, T, Yoshimura, Y & Nakazawa, H 2000, 'Antioxidative and anti-
glycation activity of garcinol from Garcinia indica fruit rind', J Agric Food Chem, vol.
48, no. 2, pp. 180-5.
Yoshida, N, Okumura, K & Aso, Y 2005, 'High serum pentosidine concentrations are
associated with increased arterial stiffness and thickness in patients with type 2 diabetes',
Metabolism, vol. 54, no. 3, pp. 345-50.
Yu, YM & Lin, HC 2010, 'Curcumin prevents human aortic smooth muscle cells
migration by inhibiting of MMP-9 expression', Nutr Metab Cardiovasc Dis, vol. 20, no.
2, pp. 125-32.
Zhao, J, Sun, XB, Ye, F & Tian, WX 2011, 'Suppression of fatty acid synthase,
differentiation and lipid accumulation in adipocytes by curcumin', Mol Cell Biochem, vol.
351, no. 1-2, pp. 19-28.
Ziegler, D & Gries, FA 1997, 'α-Lipoic Acid in the Treatment of Diabetic Peripheral and
Cardiac Autonomic Neuropathy', Diabetes, vol. 46, no. Supplement 2, pp. S62-S6.