primary factors affecting sturgeon … · mark r. collins, s. gordon rogers, theodore l j. smith...
TRANSCRIPT

BULLETIN OF MARINE SCIENCE, 66(3): 917-928, 2000
PRIMARY FACTORS AFFECTING STURGEON POPULATIONS IN
THE SOUTHEASTERN UNITED STATES: FISHING MORTALITY
AND DEGRADATION OF ESSENTIAL HABITATS
Mark R. Collins, S. Gordon Rogers, Theodore l J. Smith
and Mary L. Moser
ABSTRACT
Atlantic and shortnose sturgeons are highly valued for their eggs ( caviar) and flesh, but stocks of both species are depleted. The shortnose sturgeon is listed as an endangered species, and a long-term (multidecade) moratorium on fishing for Atlantic sturgeon is specified in the recently updated fishery management plan of the Atlantic States Marine Fisheries Commission. Thus, identification, protection, and enhancement of essential habitats are priority issues for management agencies. As anadromous (Atlantic) and nearly anadromous (shortnose) species, these sturgeons use a wide variety of habitats; variations occur latitudinally, seasonally, and among different life stages. Spawning habitats have been identified in two southeastern rivers for shortnose sturgeon, but to date no spawning sites have been verified for Atlantic sturgeon. Some life stages of both species (including young juveniles) concentrate in or are restricted to small portions of estuaries during summer. Deterioration of water quality (especially dissolved oxygen) appears to be degrading the nursery function of these summer refugia, creating a recruitment bottleneck. Protection of essential habitats, especially nursery/summer habitats, from humancaused dissolved-oxygen reductions and other impacts is critical. By-catch, primarily in riverine/estuarine gillnet and estuarine/marine trawl fisheries, appears to be the second major source of mortality for southern sturgeons. Reserves, or no-fishing zones, could protect populations by reducing or eliminating by-catch mortality. This protection would be especially valuable in high salinity foraging areas (used primarily during cool months) and during spawning migrations. Proposed management strategies aimed at conservation and restoration of sturgeon populations therefore focus on two areas: protection of essential habitats and establishment of reserves to provide protection from fishing gears that generate substantial sturgeon by-catch. Specific management recommendations were developed on a system-by-system basis.
Two anadromous or semianadromous sturgeon (Acipenseridae) species occur on the
Atlantic coast of North America, shortnose (Acipenser brevirostrum) and Atlantic (A.
oxyrinchus oxyrinchus) sturgeons (Vladykov and Greeley, 1963). Previously ranging south
ward to northern Florida, all known populations of these species in the South have been
depleted or extirpated. The shortnose sturgeon was listed as an endangered species in the
United States in 1967. North Carolina's fishery for Atlantic sturgeon was closed in 1991,
South Carolina's in 1985, Georgia's in 1996, and Florida's (east coast) in 1983. A
multidecade moratorium on the Atlantic sturgeon fisheries in all states was imposed in
1998 by the Atlantic States Marine Fisheries Commission (ASMFC, 1998).
Biologists have studied shortnose and Atlantic sturgeons in the Southeast for over two
decades. In general these research efforts were initially focused on documenting the com
mercial fishery (e.g., Leland, 1968) but then expanded into the areas of artificial propa
gation ( e.g., Smith et al., 1980), habitat associations (Hall et al., 1991 ), physiology (Jenkins
et al., 1993), and sources of mortality (Collins et al., 1996). However, information in
certain areas, including habitat utilization patterns, has been difficult to obtain because of
917

918 BULLETIN OF MARINE SCIENCE, VOL. 66, NO. 3, 2000
the relative scarcity of these fishes in most systems. It now appears that the ecology of
southern sturgeons differs from that documented for northern populations. Further, al
though the directed fisheries have been closed for up to 25 yrs, at least some populations
in the South continue to decline. Fishing mortality is believed to be an important factor in
at least some localities (Collins et al., 1996), and recent evidence suggests that degrada
tion of essential habitat is another major factor in the declines (Rogers and Weber, 1995a).
Here, we synthesize the habitat-use information collected in river systems and nearshore
waters in Georgia, South Carolina, and North Carolina; integrate existing by-catch infor
mation; and provide management recommendations on a system-by-system basis.
METHODS
The methods used in the studies we synthesize have been described in previous publications and reports and will only be briefly mentioned here. The objectives of some studies included identifying essential habitats of various life stages, whereas other studies focused on other issues but pro
duced information on habitat utilization. Habitat use was determined primarily by collection of
specimens in gill nets, trammel nets, and trawls and through radio and acoustic telemetry studies.
Salinity, temperature, dissolved oxygen (DO), and often substrate type were determined for most
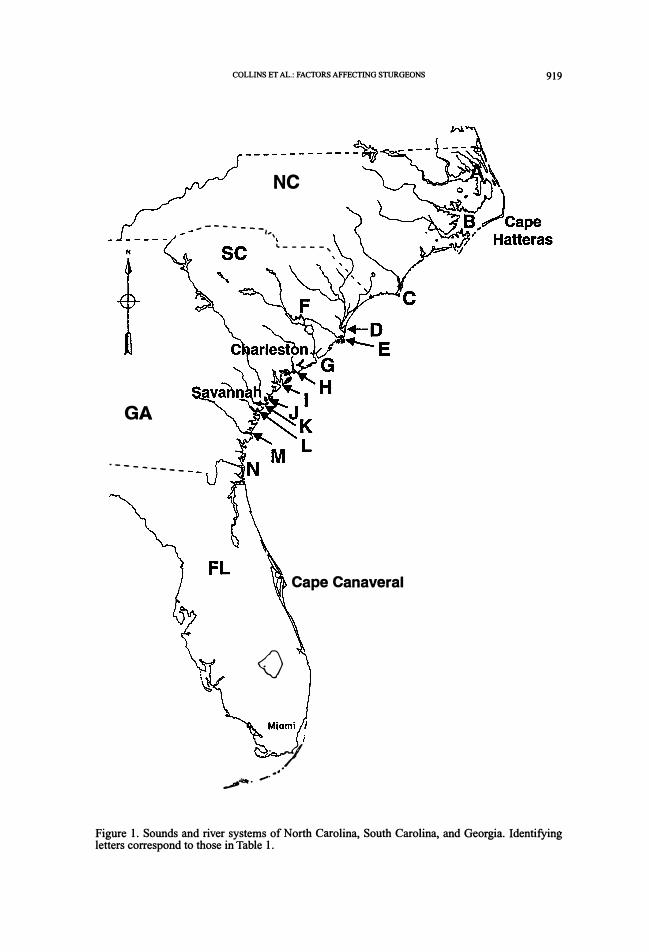
capture and telemetry locations. Study areas included most major rivers and sounds (Fig. 1 ). In the St. Marys River (Georgia) during 1994-1995, and in the Satilla River (Georgia) during
1995 and 1998, stratified random trammel net surveys for sturgeons were conducted during summer (Rogers and Weber, 1995a). In the Altamaha River (Georgia), trammel nets were deployed during 1986-1994 in all seasons at river kilometers (rlan) 0-80. Side-scan sonar was used to survey
areas of summer concentrations of both sturgeon species (Rogers et al., 1994). Shortnose sturgeon
spawning locations were tentatively identified by telemetry studies and gill-net collections (Rogers
and Weber, 1995b ). Winter concentrations of subadultAtlantic sturgeon were located serendipitously during trawling intended to determine sea turtle abundance in the nearshore shipping channel (Rogers et al., 1994).
In the Ogeechee River (Georgia), both stratified random (1993-1994) and nonrandom trammel-net
sampling were used to collect sturgeons. Shortnose sturgeon telemetry studies were conducted
during 1994-1995 (Weber et al., in press). Nonrandom trammel net sampling and standardized
hydrographic surveys were conducted during 1997-1998 (Rogers, unpubl. data). Sturgeons have probably been studied more extensively in the Savannah River (Georgia/South
Carolina) than in any other system in the region. Most research was directed toward shortnose
sturgeon, but information on Atlantic sturgeon ( especially juveniles) was also acquired. From 1985
through 1992, and again 1997 to present, sturgeons were captured by directed sampling and acquired from the by-catch of a network of commercial American shad (Alosa sapidissima) gill-net
fishermen (Hall et al., 1991; Smith et al., 1993, unpubl. data). Radio and acoustic telemetry studies were conducted during 1985-1991 (Hall et al., 1991; Collins and Smith, 1993). Shortnose sturgeon
brood fish from this river were spawned, and about 92,000 progeny of various ages were released into the river at several locations in a pilot stock-enhancement study conducted during 1984-1992 (Smith and Collins, 1996). In addition to the stock-enhancement work, cultured juveniles were used in other studies, such as delineation of environmental tolerances (salinity, temperature, and DO; Jenkins et al., 1993) and identification of optimal tagging techniques (Smith et al., 1990; Collins et al., 1994). Spawning and culture research on Savannah River strain shortnose sturgeon has been conducted since the mid-1980s at several locations, including U.S. Fish and Wildlife Service facilities (e.g., Orangeburg National Fish Hatchery, Bears Bluff National Fish Hatchery) and South Carolina Department of Natural Resources facilities (e.g., Marine Resources Research Institute, Waddell Mariculture Center). Nonrandom trammel-net sampling for sturgeons was conducted in the lower Savannah River during 1997-1998 (Collins, unpubl. data).
GA
. - - - -----
COLLINS ET AL.: FACTORS AFFECTING STURGEONS 919
NC
--------
,,,
"------,
Cape Canaveral
()
·.1,..,,,r"'· -··
Figure 1. Sounds and river systems of North Carolina, South Carolina, and Georgia. Identifying letters correspond to those in Table 1.

920 BULLETIN OF MARINE SCIENCE, VOL. 66, NO. 3, 2000
In the St. Helena Sound system, a mark-recapture study of shortnose and juvenile Atlantic stur
geons has been under way since 1994 (McCord, 1998). Also, former commercial sturgeon fisher
men have assisted in the capture of adult Atlantic sturgeon for an ongoing telemetry study that began in winter-spring 1997 in this river and in the nearby Combahee River (Collins et al., 1999).
Starting in 1998, adult shortnose sturgeon have been captured and are being used in telemetry
studies to identify movements and habitat use (Collins, unpubl. data).
In the Cooper River (South Carolina), adult shortnose sturgeon were captured during the spawn
ing season in 1995-1998 below Pinopolis dam and fitted with radio transmitters to assess spawning
and postspawning habitat use (Cooke, 1998). Information on Atlantic sturgeon was obtained inci
dentally from trawling operations designed to determine sea turtle abundance in the Charleston Harbor shipping channel during 1990-1991 (Van Dolah and Maier, 1993). In the Santee-Cooper
Lakes, a reservoir/canal/river system formed by the damming of the Santee and Cooper rivers,
shortnose sturgeon captured in 1998-1999 by commercial catfish trotline fishermen and by di
rected sampling with trammel nets were implanted with radio transmitters in an attempt to deter
mine whether reproduction took place within the system (Collins, unpubl. data).
The primary sturgeon fishery in the southeast during 1970-1985 was in the Winyah Bay area.
During 1978-1982 fisheries and biological data were collected in an effort to characterize that
segment of the sturgeon fishery (Smith et al., 1984). The fishery was closed in 1985, but since then
by-catch mortality and poaching have become concerns. During 1994-1996, sturgeon by-catch
mortality in Winyah Bay's commercial shad gill net fishery was assessed (Collins et al., 1996).
In the lower Cape Fear River (North Carolina), a 1990-1992 gill-net survey and telemetry study
investigated habitat use and distribution patterns of juvenile Atlantic and adult shortnose sturgeons
(Moser and Ross, 1995). In 1996-1998 this survey was expanded upriver to document effects of
low-elevation dams on sturgeon movements (Moser et al., 1998). Ancillary data on by-catch of sturgeon in the shad fishery were obtained throughout the surveys.
Little is known of sturgeon distribution in other North Carolina drainages. A data base of stur
geon captures by various agencies was compiled in 1997 and is being used to identify the rivers
most likely to support sturgeon populations (Moser et al., 1998). In addition, a telemetry study is in
progress in Albemarle Sound to document habitat use by juvenile Atlantic sturgeon (J. Hightower,
North Carolina State University, pers. comm.).
RESULTS AND DISCUSSION
Adult shortnose sturgeon are especially vulnerable to passive fishing gears during their
extensive up- and downriver spawning movements. In the Savannah River, for example,
adults are common in the by-catch of the American shad gill-net fishery from the lowest
point in the river at which fishing is allowed (about rkm 43) up to at least rkm 278 (the
uppermost of several sturgeon spawning areas; Collins and Smith, 1993). By-catch of
sturgeons in this river was as high as 102 fish fisher1 yr1, and immediate by-catch mor
tality of sturgeons in this gear type was 16%; another 20% offish were injured (Collins et al., 1996). In addition to such accidental mortality, intentional mortality of shortnose
sturgeon captured in the shad fishery is known to occur (McCord, 1998). In the Cooper
River, for example, about 50 shortnose sturgeon (20-50% of the estimated annual spawn
ing aggregation) were taken by a single team of poachers in 1995 (D. Cooke, S.C. Depart
ment ofNatural Resources, pers. comm.). Total fishing mortality is therefore likely to be substantial in rivers containing both a sturgeon population and a shad fishery. Further
more, sublethal effects of by-catch have been documented. Shortnose sturgeon may delay
or even abort upstream spawning migration after gill-net capture (Moser and Ross, 1995).
Valuable shortnose sturgeon spawners can be protected by establishment of no-fishing

COLLINS ET AL.: FACTORS AFFECTING STURGEONS 921
zones for set-net (anchored gillnet) fisheries from the mesohaline estuary (overwintering
area) upriver to spawning areas.
Growth rates of juvenile Atlantic, and probably shortnose, sturgeon are greatest in spring
and fall and much lower in winter and summer (McCord, 1998; unpubl. data), so the fish
are probably inhabiting important foraging areas during cooler seasons. These areas ap
pear to include a large portion of the estuary and, for Atlantic sturgeon, the nearshore
continental shelf and shipping channels (Rogers et al., 1994). Marine and estuarine hook
and-line fisheries have little impact, but sturgeons (especially juvenile and subadult
Atlantics) do occur in the by-catch of trawl fisheries in South Carolina and Georgia,
especially the inshore/nearshore segment of the penaeid shrimp trawl fishery during cool
months. The shrimp trawl fishery produced 39% of 97 reported recaptures of Atlantic
sturgeon tagged in a Georgia study conducted before use of turtle excluder devices be
came mandatory (Collins et al., 1996). Use of turtle excluder devices is thought, but not
proven, to reduce by-catch of sturgeons. In North Carolina, sturgeons are probably caught
in shrimp trawls during much of the year because trawling is permitted in the sounds
shoreward of the Outer Banks. Unfortunately, little is known about the magnitude of
by-catch or by-catch-induced mortality in the inshore segment of the fishery, but the long
tow times normally employed in this fishery suggest that mortality may be high. Marine/
estuarine reserves, in the form of no-trawling zones in bays and sounds shoreward of
established lines, are already in place in South Carolina and Georgia. If future studies
verify that trawling causes significant sturgeon mortality, the mortality could be drasti
cally reduced by extension of existing reserves seaward and by closure of the sounds to
trawling in North Carolina.
No spawning sites of Atlantic sturgeon have been identified in the southeastern United
States, but shortnose sturgeon spawning sites have been identified in two rivers and ten
tatively in another. Sites were characterized by deep, scoured channels with hard sub
strates to which the eggs may adhere. Spawning of shortnose sturgeon occurred at several
widely separated locations with no obvious similarities in at least one river (Hall et al.,
1991; Collins and Smith, 1993), but in other rivers, only a single spawning site is known
(the base of the dam in the Cooper River; Cooke, 1998). Alternatively, spawning appears
to be closely associated with a scarce and easily identifiable landform in some rivers. For
example, in the Altamaha River, shortnose sturgeon appear to spawn at widely separated
locations but only at limestone outcrops (Rogers and Weber, 1995b ). This spawning habi
tat is similar to that used by Gulf sturgeon (A. o. desotoi) in the Choctawhatchee River
(Florida/Alabama; J. Hightower, North Carolina State University, pers. comm.) and in the
Suwanee River (Florida; Sulak and Clugston, 1998). In this case, designating known or
probable spawning areas as essential habitat, and protecting or enhancing them, may help conserve existing sturgeon stocks (Table 1 ).
With rare exceptions, the fresh-brackish water interface area serves as the summer
nursery habitat for Atlantic sturgeon and the summer habitat for all ages of shortnose
sturgeon in the Southeast (Smith et al., 1993; McCord, 1998; Fig. 2). The essential nature
of this habitat is illustrated by patterns of capture during summer for shortnose sturgeon
in the Altamaha (Flournoy et al., 1992), Savannah (Hall et al., 1991; Smith and Collins, 1996), and Edisto (unpubl. data) rivers, where juvenile shortnose sturgeon have been
captured only in the vicinity of the interface. Summer behavior of adults, however, may
differ somewhat among rivers. In the Altamaha River, shortnose sturgeon oversummer in

922 BULLETIN OF MARINE SCIENCE, VOL. 66, NO. 3, 2000
Table 1. Initial management recommendations for Acipenser brevirostrum and A. oxyrinchus population segments in North Carolina, South Carolina, and Georgia. Some systems include more than one river. The option of restocking rivers with hatchery-produced fish is omitted from consideration because its present applicability is in doubt. Letters in parentheses correspond to locations in Figure 1.
System
Albemarle Sound (A)
Pamlico Sound (B)
Cape Fear River (C)
Winyah Bay (D)
Santee River (E)
Santee-Cooper Lakes (F)
Cooper River (G)
St. Helena Sound (H)
Port Royal Sound (I)
Savannah River (J)
Ogeechee River (K)
Altamaha River (L)
Satilla River (M)
St. Marys River (N)
Initial Management Recommendations
Establish estuarine set-net fishery reserve.
None (Presence of sturgeon populations not verified.)
(1) Establish riverine/estuarine reserve closed to set-net fishery.
(2) Provide access to upriver habitats through low-elevation
dams.(3) Reverse degradation of nursery habitat.
(1) Establish riverine/estuarine reserves closed to set-net fishery
in the bay and all four major rivers (except any river shown not
to harbor sturgeon).(2) Maintain continuous, adequate flow from dam on Great Pee
Dee River during spawning season.(1) Establish riverine/estuarine reserve closed to set-net fishery.
(2) Maintain continuous, adequate flow from dam during
spawning season.None (Spawning and nursery habitats not identified. No net
fisheries.)(1) Protect A. brevirostrum spawning habitat at base of dam.
(2) Halt degradation of nursery habitat.
(3) Maintain continuous, adequate flow from dam during
spawning season.(1) Establish riverine reserve closed to set-net fishery for the
Edisto River only.(2) Expand estuarine reserve closed to trawl net fishery seaward
during late fall through winter.None (Presence of sturgeon populations not verified.)
(1) Halt degradation of nursery habitat, especially the increased
salinity and decreased DO from harbor deepening and point
source pollution.(2) Establish riverine reserve closed to set-net fishery to first
dam at river kilometer 300.(3) Maintain continuous, adequate flow from dam during
spawning season.(1) Halt degradation of nursery habitat.
(2) Establish riverine/estuarine reserve closed to set-net fishery.
(1) Halt degradation of nursery habitat.
(2) Establish riverine/estuarine reserve closed to set-net fishery.
(3) Protect apparent A. brevirostrum spawning habitat at
limestone outcrops.(1) Reverse degradation of nursery habitat.
(2) Establish riverine/estuarine reserve closed to set-net fishery.
Reverse degradation of nursery habitat.
a very few deep holes near the interface, which they rarely leave. This behavior has also
been noted for juvenile Atlantic sturgeon in the Cape Fear River (Moser and Ross, 1995).
In the Edisto River, however, telemetry results suggest that shortnose sturgeon follow the
salt front upriver with flood tides and downriver with ebb tides. The narrow range of
COLLINS ET AL.: FACTORS AFFECTING STURGEONS 923
7 0.07
6 Salinity
,• '.
5 N ; , Shortnose
-
I ,'/\ '. sturgeon C. C. I \ ,,. \ '. - 4 I V; \ •
I \ C: 3
Atlantic sturgeon / \ 'ii \
2 I \ I \
0.06
0.05 'i:' .c:
0.04 C:
0.03 .c:
0.02
0.01
1 ---------'-. - .. J. \ 0
0 -0.01
16 20 24 28 32 36 40 44 48 52 54 56 60 64 68 72
River Kilometer
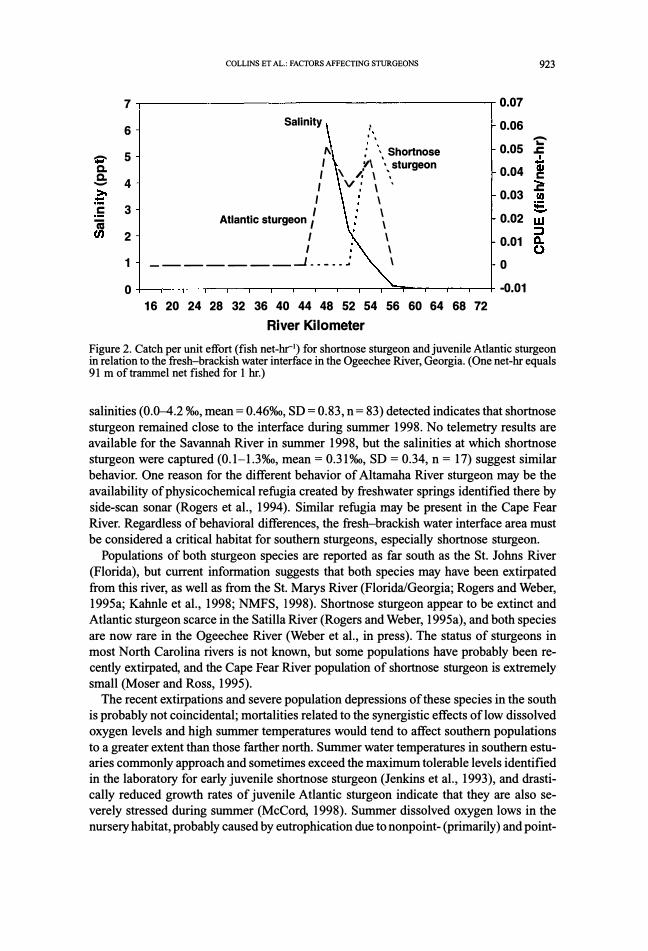
Figure 2. Catch per unit effort (fish net-hr1) for shortnose sturgeon and juvenile Atlantic sturgeon in relation to the fresh-brackish water interface in the Ogeechee River, Georgia. (One net-hr equals 91 m of trammel net fished for 1 hr.)
salinities (0.0--4.2 %o, mean = 0.46%0, SD = 0.83, n = 83) detected indicates that shortnose
sturgeon remained close to the interface during summer 1998. No telemetry results are
available for the Savannah River in summer 1998, but the salinities at which shortnose
sturgeon were captured (0.1-1.3%0, mean = 0.31 %o, SD = 0.34, n = 17) suggest similar
behavior. One reason for the different behavior of Altamaha River sturgeon may be the
availability of physicochemical refugia created by freshwater springs identified there by
side-scan sonar (Rogers et al., 1994). Similar refugia may be present in the Cape Fear
River. Regardless of behavioral differences, the fresh-brackish water interface area must
be considered a critical habitat for southern sturgeons, especially shortnose sturgeon.
Populations of both sturgeon species are reported as far south as the St. Johns River
(Florida), but current information suggests that both species may have been extirpated
from this river, as well as from the St. Marys River (Florida/Georgia; Rogers and Weber,
1995a; Kahnle et al., 1998; NMFS, 1998). Shortnose sturgeon appear to be extinct and
Atlantic sturgeon scarce in the Satilla River (Rogers and Weber, 1995a), and both species
are now rare in the Ogeechee River (Weber et al., in press). The status of sturgeons in
most North Carolina rivers is not known, but some populations have probably been re
cently extirpated, and the Cape Fear River population of shortnose sturgeon is extremely
small (Moser and Ross, 1995).
The recent extirpations and severe population depressions of these species in the south
is probably not coincidental; mortalities related to the synergistic effects oflow dissolved
oxygen levels and high summer temperatures would tend to affect southern populations
to a greater extent than those farther north. Summer water temperatures in southern estuaries commonly approach and sometimes exceed the maximum tolerable levels identified
in the laboratory for early juvenile shortnose sturgeon (Jenkins et al., 1993), and drasti
cally reduced growth rates of juvenile Atlantic sturgeon indicate that they are also se
verely stressed during summer (McCord, 1998). Summer dissolved oxygen lows in the
nursery habitat, probably caused by eutrophication due to nonpoint- (primarily) and point-
924 BULLETIN OF MARINE SCIENCE, VOL. 66, NO. 3, 2000
7
6
E s I r
1989
Cl) en 4
'C 3 Cl)
2U) U)
1
o.l.,_ ___________________________ _
� T'" T'"
""' 10
....: 0 N C'>
c,i (')
10
cci (')
N 0
River Kilometer
� T'" It)
CIO It)
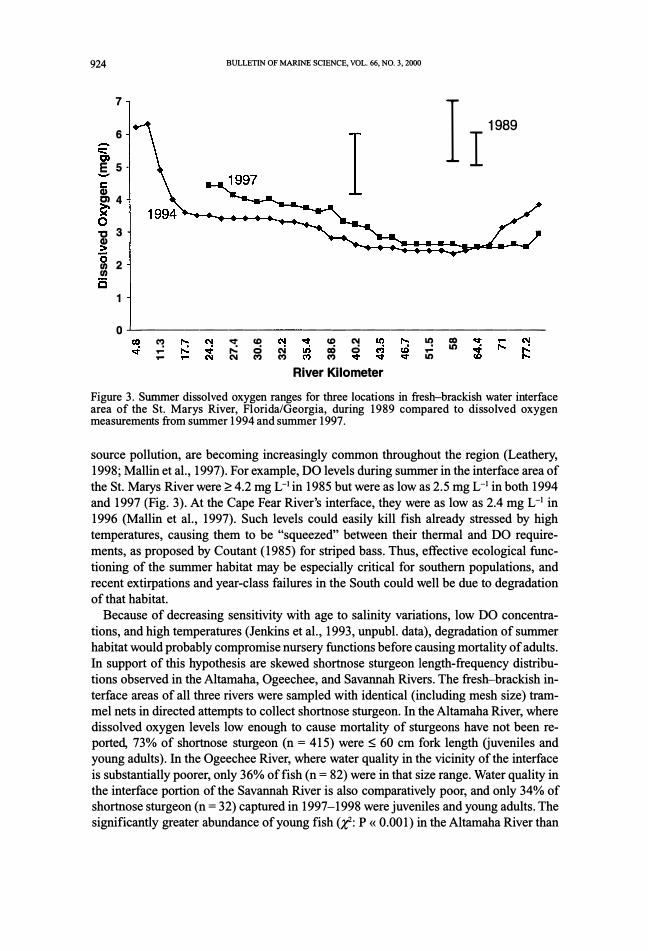
Figure 3. Summer dissolved oxygen ranges for three locations in fresh-brackish water interface area of the St. Marys River, Florida/Georgia, during 1989 compared to dissolved oxygen measurements from summer 1994 and summer 1997.
source pollution, are becoming increasingly common throughout the region (Leathery,
1998; Mallin et al., 1997). For example, DO levels during summer in the interface area of
the St. Marys River were �4.2 mg L-1 in 1985 but were as low as 2.5 mg L-1 in both 1994
and 1997 (Fig. 3). At the Cape Fear River's interface, they were as low as 2.4 mg L-1 in
1996 (Mallin et al., 1997). Such levels could easily kill fish already stressed by high
temperatures, causing them to be "squeezed" between their thermal and DO require
ments, as proposed by Coutant (1985) for striped bass. Thus, effective ecological func
tioning of the summer habitat may be especially critical for southern populations, and
recent extirpations and year-class failures in the South could well be due to degradation
of that habitat.
Because of decreasing sensitivity with age to salinity variations, low DO concentra
tions, and high temperatures (Jenkins et al., 1993, unpubl. data), degradation of summer
habitat would probably compromise nursery functions before causing mortality of adults.
In support of this hypothesis are skewed shortnose sturgeon length-frequency distribu
tions observed in the Altamaha, Ogeechee, and Savannah Rivers. The fresh-brackish in
terface areas of all three rivers were sampled with identical (including mesh size) tram
mel nets in directed attempts to collect shortnose sturgeon. In the Altamaha River, where
dissolved oxygen levels low enough to cause mortality of sturgeons have not been reported, 73% of shortnose sturgeon (n = 415) were � 60 cm fork length (juveniles and
young adults). In the Ogeechee River, where water quality in the vicinity of the interface
is substantially poorer, only 36% offish (n = 82) were in that size range. Water quality in
the interface portion of the Savannah River is also comparatively poor, and only 34% of shortnose sturgeon (n = 32) captured in 1997-1998 were juveniles and young adults. The
significantly greater abundance of young fish (X2: P « 0.001) in the Altamaha River than

/ /
COLLINS ET AL.: FACTORS AFFECTING STURGEONS
Interface
E
925
Figure 4. Generalized southern river system showing locations of essential habitats of Atlantic and shortnose sturgeons in relation to areas where sturgeons are likely to occur in the by-catch of commercial shrimp and shad fisheries . (A= upriver spawning habitat for both species; B = nursery habitat for both species and summer habitat for adult shortnose; C
1 and C, = late fall-winter habitat
for adult shortnose and juvenile Atlantic, respectively; D = late fall-wmter habitat for subadult Atlantic; E = area of potential threat from the shad fishery; F = area of potential threat from the shrimp fishery.)
in the Ogeechee and Savannah rivers may be a result of degradation of the summer habi
tat for these life stages in the latter two rivers.
Rivers north of the Ogeechee River in which the summer habitat is especially vulner
able at present include the Savannah River, the Cooper River, and a number of drainages
in North Carolina including the Cape Fear River. In all cases, water quality (especially
DO) is already low enough to be of concern, and major dredging projects through the
nursery habitats are scheduled for several systems. In the Savannah River, water-quality
models predict that planned channel deepening will reduce dissolved oxygen concentra
tions in the nursery habitat below its present minimally adequate level (GPA, 1998), and
the predicted concentrations are likely to be lethal (Jenkins et al., 1993). In the Cooper
River, water-quality change predictions have not yet been quantified, but the low adult
population size (about 100-300 spawners annually; D. Cooke, S. C. Department of Natu
ral Resources, pers. comm.) makes even a slight reduction in juvenile survival particu
larly deleterious. The Cape Fear River population of shortnose sturgeon is probably even
smaller than that of the Cooper River, and deepening is scheduled to begin in 1998. Fur
ther, existing water quality problems in North Carolina systems have reportedly been
exacerbated by substantial expansion of the hog farming industry, and low-DO events
and harmful algal blooms have become more frequent there. Protection and restoration of
Estuary

926 BULLETIN OF MARINE SCIENCE, VOL. 66, NO. 3, 2000
the essential summer/nursery habitat, primarily through reduction of nutrient loading in
the region's rivers, is critical if populations of sturgeons are to survive in the South.
Habitat-use patterns of most life stages of Atlantic and shortnose sturgeons have been
identified in the South. Sturgeon management and recovery efforts in this region should
incorporate this information into river-specific policies (Table 1) that focus on two goals:
(1) establishing reserves to reduce fishing mortality and (2) protection of essential habi
tats. Marine, estuarine, and riverine reserves aimed at shad set-net and shrimp trawl fish
eries could reduce or eliminate by-catch mortality during spawning migrations and dur
ing cool months when populations shift seaward. Exempting hook-and-line fisheries, which
do not affect sturgeons, could increase popular support for these reserves. Protection and
restoration of spawning and especially summer habitats are also critically needed man
agement actions (Fig. 4).
ACKNOWLEDGMENTS
We thank D. Cooke, S. C. Department of Natural Resources, for providing information from the Cooper River, SC, J. W. McCord, S. C. Department ofNatural Resources, for discussions pertaining to
McCord (1998), and I. Wirgin, N.Y.U. Medical Center, for collaborating in a recent study that pro
duced information incorporated here. The many studies upon which this synthesis is based were funded
by a number of agencies, including the Cape Fear River Program, Georgia Department of Natural
Resources (Coastal Resources Division, and Nongame and Endangered Species Section), Nature Con
servancy of Georgia, National Marine Fisheries Service, North Carolina Fisheries Resources, South
Carolina Department ofNatural Resources, U.S. Air Force (Warner Robins Air Logistics Center), U.S.
Army (Ft. Stewart, GA), U.S. Army Corps ofEngineers, and U.S. Fish and Wildlife Service. This is
contribution number 438 of the South Carolina Marine Resources Center.
LITERATURE CITED
ASMFC (Atlantic States Marine Fisheries Commission). 1998. Amendment l to the interstate fish
ery management plan for Atlantic sturgeon (Fisheries Management Rpt. 17). ASMFC, Wash
ington, D.C.
Collins, M. R. and T. I. J. Smith. 1993. Characteristics of the adult segment of the Savannah River
population of shortnose sturgeon. Proc. Ann. Conf. Southeastern Assoc. Fish and Wildlife Agen
cies 47: 485491.
_____ and L. D. Heyward. 1994. Effectiveness of six methods for marking juvenile shortnose
sturgeons. Prog. Fish-Cult. 56: 250-254.
____ _, S. G. Rogers and T. I. J. Smith. 1996. Bycatch of sturgeons along the southern Atlan
tic coast of the USA. N. Am. J. Fish. Manage. 16: 24-29.
____ _, T. I. J. Smith and W. Post. 1999. Critical habitats of Atlantic sturgeon. Final report to
the National Marine Fisheries Service (Saltonstall-Kennedy Program), Grant No. NA 77FD0063. Cooke, D. W. 1998. Santee-Cooper blueback herring studies-rediversion project. Annual Progress
Report, SCR 1-21, So. Carolina Dept. Natural Resources, Columbia, South Carolina. Coutant, C. C. 1985. Striped bass, temperature, and dissolved oxygen: a speculative hypothesis for
environmental risk. Trans. Am. Fish. Soc. 114: 31-61.
Flournoy, P. H., S. G. Rogers and P. S. Crawford. 1992. Restoration of shortnose sturgeon in the Altamaha River, Georgia. Final report to the U.S. Fish Wildl. Serv., project AFS-2.
GPA (Georgia Ports Authority). 1998. Savannah Harbor Expansion, Draft Environmental Impact Statement. Savannah, Georgia.

COLLINS ET AL.: FACWRS AFFECTING STURGEONS 927
Hall, J. W., T. I. J. Smith and S. D. Lamprecht. 1991. Movements and habitats ofshortnose sturgeon,
Acipenser brevirostrum in the Savannah River. Copeia 1991: 695-702.
Jenkins, W. E., T. I. J. Smith, L. D. Heyward and D. M. Knott. 1993. Tolerance of shortnose stur
geon, Acipenser brevirostrum, juveniles to different salinity and dissolved oxygen concentra
tions. Proc. Ann. Conf. Southeastern Assoc. Fish and Wildlife Agencies 47: 476--484.
Kahnle, A. W., K. A. Hattala, K. A. McKown, C. A. Shirey, M. R. Collins, T. S. Squiers, Jr. and T.
Savoy. 1998. Stock status of Atlantic sturgeon of Atlantic Coast estuaries. Report for the Atlan
tic States Marine Fisheries Commission, Washington, D.C.
Leathery, S. 1998. Eutrophication primary nonpoint pollution problem. Fisheries (Bethesda) 23(9):38.
Leland, J. G., II. 1968. A survey of the sturgeon fishery of South Carolina. Contrib. 47. Bears Bluff
Laboratories, Wadmalaw Island, South Carolina. 27 p.
McCord, J. W. 1998. Investigation of fisheries parameters for anadromous fishes in South Carolina.
Completion report to the National Marine Fisheries Service, project AFC-53.
Mallin, M.A., M. H. Posey, M. L. Moser, G. C. Shank, M. R. Mciver, T. D. Alphin, S. H. Ensign and
J. F. Merritt. 1997. Environmental assessment of the lower Cape Fear River system, 1996--1997.
CMSR Rpt. no. 97-01. Center for Marine Science Research, UNC-Wilmington, Wilmington,
North Carolina.
Moser, M. L. and S. W. Ross. 1995. Habitat use and movements of shortnose and Atlantic sturgeons
in the lower Cape Fear River, North Carolina. Trans. Am. Fish. Soc. 124: 225-234.
____ , J. B. Bichy and S. B. Roberts. 1998. Sturgeon distribution in North Carolina. Final
Report to the U.S. Army Corps Engrs., Wilmington District.
NMFS (National Marine Fisheries Service). 1998. Final recovery plan for the shortnose sturgeon
Acipenser brevirostrum. Prepared by the Shortnose Sturgeon Recovery Team for the National
Marine Fisheries Service, Silver Spring, Maryland.
Rogers, S. G. and W. Weber. 1995a. Status and restoration of Atlantic and shortnose sturgeons in
Georgia. Final report to the National Marine Fisheries Service, project NA46FA102-01.
_____ and ___ . 1995b. Movements of shortnose sturgeon in the Altamaha River sys-
tem, Georgia. Contrib. Ser. #57, Coastal Resources Division, Georgia Department of Natural
Resources, Brunswick, Georgia.
Rogers, S. G., P. H. Flournoy and W. Weber. 1994. Status and restoration of Atlantic sturgeon in Geor
gia. Final report to the National Marine Fisheries Service, project NA16FA0098-01, -02, -03.
Smith, T. I. J. and M. R. Collins. 1996. Shortnose sturgeon stocking success in the Savannah River.
Proc. Ann. Conf. Southeastern Assoc. Fish and Wildlife Agencies 50: 112-121.
____ _, E. K. Dingley and D. E. Marchette. 1980. Induced spawning and culture of Atlantic
sturgeon. Prog. Fish-Cult. 42: 147-151.
____ _, D. E. Marchette, and G. F. Ulrich. 1984. The Atlantic sturgeon fishery in South
Carolina. N. Am. J. Fish. Manage. 4: 164-176. ____ _, S. D. Lamprecht and J. W. Hall. 1990. Evaluation of tagging techniques for shortnose
sturgeon and Atlantic sturgeon. Am. Fish. Soc. Symp. 7:134-141.
____ _, M. R. Collins, and E. Kennedy. 1993. Identification of critical habitat requirements of shortnose sturgeon in South Carolina. Final report to the U.S. Fish Wild!. Serv., Project No.
AFS-17. Sulak, K. J., and J. P. Clugston. 1998. Early life history stages of Gulf sturgeon in the Suwannee
River, Florida. Trans. Am. Fish. Soc. 127: 758-771. Van Dolah, R. F., and P. P. Maier. 1993. The distribution of loggerhead turtles ( Caretta caretta) in the
entrance channel of Charleston Harbor, South Carolina, U.S.A. J. Coastal Res. 9: 1004-1012.
Weber, W., C. A. Jennings and S. G. Rogers. (in press). Population size and movements patterns of
shortnose sturgeon (Acipenser brevirostrum) in the Ogeechee River system, Georgia. Proc.
52nd Ann. Meeting Southeastern Assoc. Fish and Wildlife Agencies.

928 BULLETIN OF MARINE SCIENCE, VOL. 66, NO. 3, 2000
ADDRESSES: (M.R.C., T.I.J.S.) South Carolina Department of Natural Resources, Marine Resources
Research Institute, P.O. Box I 2559, Charleston, South Carolina 29422; (S.G.R.) Satilla Management
Associates, Inc., P.O. Box 177, Waynesville, Georgia 31566; (M.L.M.) University of North Carolina
Wilmington, Center for Marine Science Research, 7205 Wrightsville Ave., Wilmington, North Carolina
28403.