printed in u.s.a. alteration in bacterial morphology by …jb.asm.org/content/97/1/362.full.pdf ·...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Jan. 1969, p. 362-366Copyright @ 1969 American Society for Microbiology
Vol. 97, No. IPrinted in U.S.A.
Alteration in Bacterial Morphology by Optochin andQuinine Hydrochlorides1
ARNOLD S. KREGER,2 D. C. SWARTZENDRUBER,3 AND R. H. OLSENDepartment of Microbiology, University of Michigan, Ann Arbor, Michigan 48104
Received for publication 16 October 1968
Incubation of washed bacterial and ribosomal suspensions with optochin orquinine hydrochloride caused an increase in the turbidity of the suspensions andthe appearance of electron-dense cytoplasmic aggregates in the treated cells. Theseeffects were more pronounced with optochin hydrochloride than with quinine hy-drochloride, and they did not correlate with the relative sensitivities of differentbacteria to growth inhibition by optochin or quinine.
Suspensions of washed pneumococci incubatedfor a short time with high concentrations (1 mg/ml) of optochin hydrochloride or quinine hydro-chloride increase in turbidity (unpublished data).Cellular morphology and chain length, as deter-mined in wet-mount or Gram-strain preparations,were not altered by these treatments. Therefore,to determine whether changes in ultrastructuralmorphology could be correlated with the increasein turbidity, treated and untreated cells were ex-amined in the electron microscope. Experimentswere also done to determine whether the increasein turbidity was specific for the pneumococcus orwhether there was a correlation between thephenomenon and the relative sensitivities of differ-ent bacteria to growth inhibition by optochin orquinine.
This report shows that the increase in turbidityof bacterial suspensions incubated with optochinor quinine hydrochloride is a nonspecific effectof high concentrations of drug, not related tooptochin or quinine sensitivity, and is associatedwith the appearance of electron-dense aggregatesin the cytoplasms of treated cells.
MATERIALS AND METHODSOrganisms. Pseudomonas aeruginosa (1C strain)
was obtained from B. W. Holloway (Department ofBacteriology, University of Melbourne, Victoria,Australia) and was maintained on tryptone glucose-yeast extract (TGE) agar containing 0.5% Tryptone(Difco), 0.25% Yeast Extract (Difco), 0.1% glucose,and 1.5% Agar (Difco). This organism was grown in
I Taken from a thesis submitted by Arnold S. Kreger to theUniversity of Michigan Graduate School in partial fulfillment ofthe requirements for the Ph.D. degree.
2 Present address: Department of Microbiology, New YorkUniversity School of Medicine, New York, N.Y. 10016.
3 Present address: Oak Ridge Associated Universities, OakRidge, Tenn. 37830.
TGE broth or Brain Heart Infusion (BHI, Difco) a37 C.The isolation of the optochin-resistant pneumo-
cocci has been reported (2). The organisms were grownin BHI at 37 C.
All other organisms were obtained from the culturecollection of the Department of Microbiology, Uni-versity of Michigan, Ann Arbor. They were grown at37 C in BHI, with the exception of the Micrococcuslysodeikticus and Lactobacillus acidophilus strainswhich were grown in Trypticase Soy Broth (TSB,BBL) at 37 C.
Determination of minimal growth-inhibitory con-centrations of optochin hydrochloride and quininehydrochloride. Minimal growth-inhibitory concentra-tions of optochin and quinine hydrochlorides weredetermined for M. lysodeikticus and L. acidophilusin TSB and for all other organisms in BHI. Five-milliliter volumes of broth, containing various con-centrations of optochin hydrochloride (K & KLaboratories, Plainview, N.Y;) or quinine hydrochlo-ride (Mallinckrodt Chemical Works, St. Louis, Mo.),were inoculated with approximately 106 late log-phase cells. The presence or absence of growth wasnoted after incubation for 2 days at 37 C.
Extraction of ribosomes. All procedures werecarried out at approximately 5 C. The buffer used was0.01 M tris(hydroxymethyl)aminomethane hydrochlo-ride (pH 7.6) with 0.02 M magnesium acetate. Cellswere harvested from 2 liters of a late log-phase TGEculture of Pseudomonas aeruginosa 1 C. These cellswere washed twice and suspended in 20 ml of buffer.The suspension was subjected to sonic disruption for30 min at full power in a tap water-cooled Raytheon10-kc magnetostriction oscillator, and the sonic-treated material was centrifuged twice at 10,000 X gfor 30 min. The supernatant fluid was centrifugedtwice at 34,000 X g for 30 min, each time discardingthe pellet, and then centrifuged once at 105,000 X gfor 2 hr. The final pellet was washed twice, suspendedin 20 ml of buffer, dialyzed overnight against 3 litersof buffer, and centrifuged at 10,000 X g for 10 min.The supernatant fluid was decanted and stored at 5 C.
362
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VOL. 97, 1969 ALTERATION IN BACTERIAL MORPHOLOGY BY OPTOCHIN
Electron microscopy. Bacterial pellets were fixed for15 min in 2.5% glutaraldehyde in 0.15 M phosphatebuffer (pH 7.2), washed thoroughly with buffer,post-fixed for 2 hr in 1% Palade's fixative (5), de-hydrated in ethyl alcohol, and embedded in Epon812 (3). Thin sections were stained with aqueousuranyl acetate and lead citrate and were examinedwith a Philips EM 200 electron microscope.Ribosome suspensions were pipetted onto Formvar-
coated grids which were stabilized lightly with carbon.Excess fluid was removed by blotting the grids ontofilter paper. Preparations were allowed to air dry priorto examination.
Treatment of washed bacteria. Early stationaryphase cells were washed twice with 0.05 M phosphatebuffer (pH 7.6) and suspended in buffer to an ab-sorbancy at 650 nm of approximately 0.2 (approxi-mately 109 cells per ml). A 0.5-ml portion of glass-distilled water or of the appropriate optochin hydro-chloride or quinine hydrochloride stock solution wasadded to 4.5-ml samples of suspensions, and the mix-tures were incubated at 37 C for 15 to 120 min. Aftermeasurement of the absorbancy of the suspensions at650 nm in a Bausch and Lomb Spectronic-20 colorim-eter, the cells were sedimented by centrifugation inthe cold (5 C) and washed twice with chilled buffer.The pellets were then fixed and examined as described.
Treatment of washed ribosomes. Samples (4.5 ml)of a washed ribosome suspension, with an absorbancyof aproximately 0.2 at 650 nm, were incubated for 30min at 37 C with 0.5 ml of glass-distilled water or ofthe appropriate optochin hydrochloride or quininehydrochloride stock solution. After measurement ofthe absorbancy of the suspensions at 650 nm, smallamounts of suspensions were applied to grids andexamined by electron microscopy.
RESULTS
Washed cell studies. Although there was somevariation in the extent of the increases in absorb-ance observed among the different bacteria, nodistinct correlation was seen between the sensitiv-ity to growth inhibition of a bacterium and theability of the drugs to affect the absorbance of thebacterial suspension (Table 1). The effect wasgreater with optochin hydrochloride than withquinine hydrochloride. Maximal increases wereobserved by 30 min postincubation, and no de-creases were observed over periods up to 120 minpostincubation.
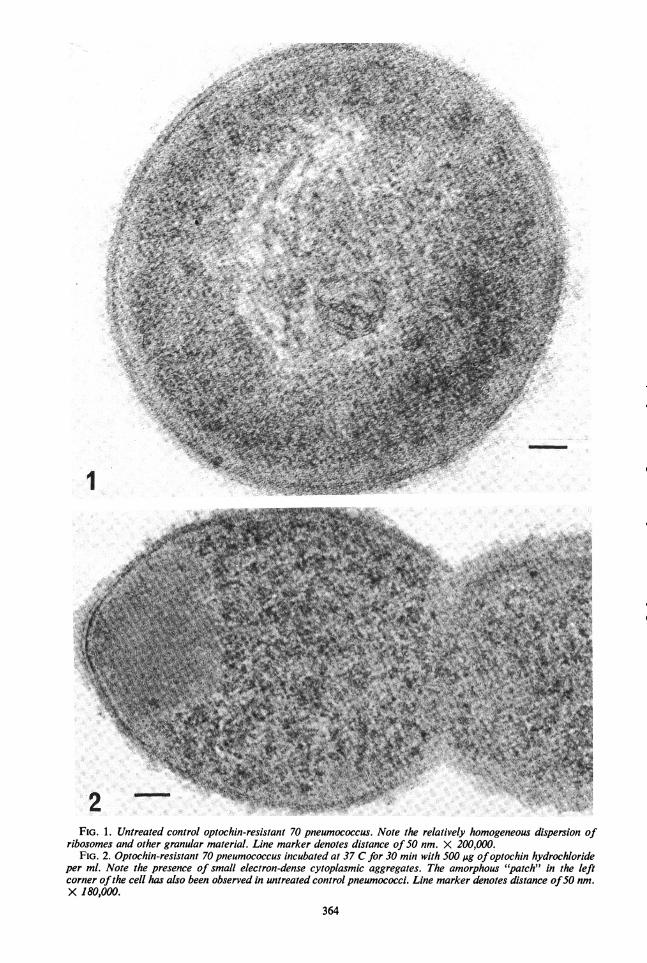
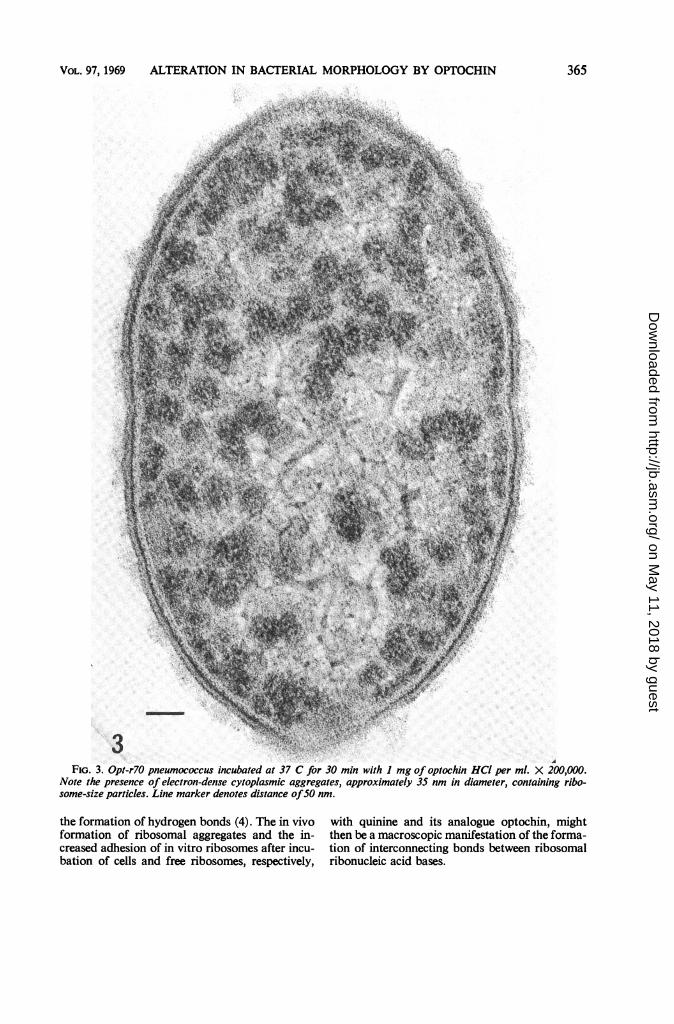
Untreated pneumococci had a relatively homo-geneous dispersion of ribosomes and othergranular material in their cytoplasms (Fig. 1);however, the incubation of pneumococci with 1mg of optochin hydrochloride or quinine hydro-chloride per ml resulted in the appearance ofelectron-dense aggregates in their cytoplasms(Fig. 3). The approximate diameter of these aggre-gates in cells incubated with optochin hydrochlo-ride was 35 nm and, in cells incubated with qui-nine hydrochloride, approximately 20 nm. Smaller
TABLE 1. Increase in absorbance and growth in-hibition of various bacteria by optochin and
quinine hydrochlorides
Per cent increase (MIC)a
OrganismOptochin Quinine
hydrochloride hydrochloride
Optochin-sensitivepneumococcus..... 90 (0.5) 40 (3)
Optochin-resistant70 pneumococcus.. 80 (70) 45 (300)
Optochin-resistant160 pneumococcus. 90 (160) 35 (600)
Staphylococcus aureus(P209)............ 83 (400) 48 (700)
Staphylococcus albus. 71 (400) 55 (700)Streptococcus mitis... 100 (70) 69 (300)Micrococcus lysodeik-
ticus.............. 63 (200) 30 (400)Lactobacillus aci-
dophilus........... 50 (80) 25 (200)Bacillus subtilis ...... 30 (400) 20 (600)Pseudomonas aerugi-nosa (lC strain).... 85 (1,000) 40 (>1,000)
Escherichia coli(Snyder strain).... 72 (400) 25 (600)
Salmonella typhosa(Rawlings strain) . 60 (300) 30 (300)
Shigella flexneri...... 45 (300) 0 (300)Aerobacter aerogenes. 80 (500) 25 (600)Serratia marcescens.. 50 (1,000) 20 (> 1,000)Bacterium anitratum.. 60 (60) 20 (200)
a Per cent increase in absorbance at 650 nmafter incubation at 37 C for 30 min with 1 mg ofdrug/ml [minimal growth-inhibitory concentra-tion (MIC) in ,ug/ml].
aggregates were observed in cells exposed to 500,ug of optochin hydrochloride per ml (Fig. 2).Particles with the appearance of ribosomes wereseen in the cytoplasmic aggregates. Similar mor-phological changes were observed with P.aeruginosa cells.Washed ribosome studies. The absorbance of
ribosomal suspensions incubated with 1 mg and500 ,ug of optochin hydrochloride per ml increased59 and 27%, respectively. Incubation with 1 mgand 500 Mg of quinine hydrochloride per ml in-creased the absorbance by 38 and 16%, respec-tively.No ribosomal aggregates were seen; however,
electron microscopy showed the optochin-treatedribosomes to be retained on the grid more heavilythan untreated ribosomes.
DISCUSSIONO'Brien et al. have reported that quinine binds
in vitro to deoxyribonucleic acid bases through
363
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
1
FIG. 1. Untreated control optochin-resistant 70 pneumococcus. Note the relatively homogeneous dispersion ofribosomes and other granular material. Line marker denotes distance of50 nm. X 200,000.
FIG. 2. Optochin-resistant 70 pneumococcus incubated at 37 C for 30 min with 500 lAg ofoptochin hydrochlorideper ml. Note the presence of small electron-dense cytoplasmic aggregates. The amorphous "patch" in the leftcorner ofthe cell has also been observed in untreated control pneumococci. Line marker denotes distance of50 nm.X 180,000.
364
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
VOL. 97, 1969 ALTERATION IN BACTERIAL MORPHOLOGY BY OPTOCHIN
FIG. 3. Opt-r70 pneumococcus incubated at 37 C for 30 min with I mg of optochin HCI per ml. X 200,000.Note the presence of electron-dense cytoplasmic aggregates, approximately 35 nm in diameter, containing ribo-some-size particles. Line marker denotes distance ofSO nm.
the formation of hydrogen bonds (4). The in vivoformation of ribosomal aggregates and the in-creased adhesion of in vitro ribosomes after incu-bation of cells and free ribosomes, respectively,
with quinine and its analogue optochin, mightthen be a macroscopic manifestation of the forma-tion of interconnecting bonds between ribosomalribonucleic acid bases.
365
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

KREGER, SWARTZENDRUBER, AND OLSEN
Although the concentrations of optochinhydrochloride which are required to produce theincrease in absorbance and the appearance ofthe electron-dense cytoplasmic aggregates are alsogrowth-inhibitory, it seems unlikely that thephenomena are indicative of the primary anti-pneumococcal action of optochin. If they were,there should have been a correlation between thesensitivity of the different bacteria to growthinhibition by optochin and the amount of drugrequired to elicit the phenomena in the differentbacteria. It is possible, however, that the phenom-ena are indicative of a secondary mechanism ofaction of optochin for the pneumococcus or of aprimary mechanism for growth inhibition of themore optochin-resistant bacteria.
Brzin (1) reported that optochin hydrochloridecaused swelling and elongation of growing Bac-terium anitratum. No alteration in morphologywas observed with other gram-negative bacteria,such as Escherichia coli, Aerobacter aerogenes,Proteus sp., Salmonella typhi, Klebsiella pneu-
moniae, Serratia marcescens, Pseudomonas pyo-cyanea, and Pasteurella pseudotuberculosis. Weobserved that B. anitratum was the most sensitiveof all the gram-negative bacteria tested to growthinhibition by optochin and quinine hydrochlo-rides.
ACKNOWLEDGMENT
We thank John Freer for helpful suggestions during prepara-
tion of the manuscript.
LITERATURE CITED
1. Brzin, B. 1965. Induction of morphological changes in bacteriaby optochin. Experientia 21:700-702.
2. Kreger, A. S., R. H. Olsen, and M. B. Talmadge. 1968. Altereddiaphorase activity in optochin-resistant pneumococci.J. Bacteriol. 96:1021-1028.
3. Luft, J. H. 1961. Improvements in epoxy resin embeddingmethods. J. Biophys. Biochem. Cytol. 9:409-414.
4. O'Brien, R. L., J. G. Olenick, and F. E. Hahn. 1966. Reactionsof quinine, chloroquine, and quinacrine with DNA andtheir effects on the DNA and RNA polymerase reactions.Proc. Natl. Acad. Sci. U.S. 55:1511-1517.
5. Palade, G. E. 1952. A study of fixation for electron microscopy.J. Exptl. Med. 95:285-298.
366 J. BACTERIOL.
on May 11, 2018 by guest
http://jb.asm.org/
Dow
nloaded from