proteomics of thyroid carcinoma: detection of potential ... · surveillance epidemiology and end...
TRANSCRIPT

Proteomics of Thyroid Carcinoma: Detection of Potential Biomarkers of Aggressive and Non-Aggressive Subtypes
by
Lawrence Kashat
A thesis submitted in conformity with the requirements for the degree of Master of Science
Institute of Medical Science University of Toronto
© Copyright by Lawrence Kashat (2011)

ii
Proteomics of Thyroid Carcinoma: Detection of Potential
Biomarkers of Aggressive and Non-Aggressive Subtypes
Lawrence Kashat
Master of Science
Institute of Medical Science University of Toronto
2011
Abstract
In search of thyroid carcinoma biomarkers, proteins secreted by thyroid cancer cell lines,
papillary-derived TPC-1 and anaplastic-derived CAL62, were analyzed using liquid
chromatography-tandem mass spectrometry. Of forty six high-confidence identifications, six
proteins were considered for verification in thyroid cancer patients’ tissues and blood. The
localization of two proteins, nucleolin and prothymosin-alpha (PTMA), was confirmed in TPC-1
and CAL62 by confocal microscopy and immunohistochemically in xenografts of TPC-1 cells
and human thyroid carcinomas. Increased nuclear and cytoplasmic expression of PTMA was
observed in anaplastic carcinomas compared to normal thyroid tissues, papillary and poorly
differentiated carcinomas. Importantly, six proteins were detected in thyroid cancer patients’
sera, warranting future analysis to confirm their potential as blood-based thyroid cancer markers.
Herein we demonstrate the ability of secretome analysis of thyroid cancer cell lines to identify
proteins that may be studied for application in management of thyroid carcinomas upon future
validation.

iii
Acknowledgments I would like to thank my supervisor, Dr. Paul Walfish, for providing me with such an excellent
opportunity. Words cannot express my gratitude for mentoring me throughout my project and
providing me with such a rewarding research experience. Thank you for allowing me to be
involved in projects with such translational potential and for your strong support during my time
in the lab.
I would also like to thank my committee for sharing their expertise and helping my project
thrive. Dr. Ranju Ralhan, I am truly grateful for the countless hours you have spent mentoring
me and guiding me throughout this project. I would not have been able to succeed without your
support and expertise. Dr. Christina MacMillan, I cannot begin to express my appreciation for
the time you have spent helping me understand all of the intricacies of thyroid pathology and
providing me with ideas to improve the quality of my research. Your insight helped spark many
ideas throughout my research and I will always be appreciative of your wonderful support. Dr.
Jonathan Irish and Dr. Ian Witterick, I am extremely thankful for the time you spent advising me
and helping to ensure my project continued progressed towards its ultimate goals.
I also wish to acknowledge the specific contributions of many brilliant researchers. Tony So,
thank you for all of the time you spent completing the western blots for our paper in the Journal
of Proteome Research. Jun Cao, thank you for providing the xenografted tumours of TPC-1 cells
for IHC analysis. I would like to thank Dr. K.W. Michael Siu, Director, Centre for Mass
Spectrometry, York University, for his support and the members of his laboratory - Leroi
DeSouza, Simon Wang, Ajay Matta, Olena Masui, and Sebastien Voisin for their help with the
MS/MS analysis and Dr. Meng for teaching me about confocal microscopy and his help with the
immunofluorescence of the cell lines. A special thank you to my colleagues, Tony So, Helen He,
Seham Chaker, Jun Cao, Xianwang Meng, Jatinder Kaur, Ipshita Kak and Ajay Matta – I will
always be thankful for your friendship and advice. I also wish to express my profound gratitude
to Gordana, Marianne, Mona, and Cecilia in Special Histology, Mount Sinai Services, for taking
time out of their busy schedule to support me for immunohistochemistry and Visiopharm
microscopy. Finally, I would like to thank my parents, Ghassan and Ghada, and my siblings,
Mary-Anne and George, for their love and support. Thank you for instilling me with the
confidence to pursue my dreams.

iv
Table of Contents
Contents
Acknowledgments.......................................................................................................................... iii
Table of Contents ........................................................................................................................... iv
List of Tables ................................................................................................................................. vi
List of Figures ............................................................................................................................... vii
List of Appendices ....................................................................................................................... viii
List of Abbreviations ..................................................................................................................... ix
Copyright Acknowledgments ..........................................................................................................x
Chapter 1 Literature Review ............................................................................................................1
1 Introduction .................................................................................................................................1
1.1 Thyroid carcinoma – General Overview ..............................................................................1
1.2 Incidence and Mortality .......................................................................................................1
1.3 Prognostic Factors ................................................................................................................4
1.4 Variants of PTC ...................................................................................................................7
1.5 Anaplastic Thyroid Carcinomas ........................................................................................10
1.6 Molecular Diagnostics and Serum Thyroglobulin .............................................................13
1.7 Mutations in Thyroid Carcinoma .......................................................................................17
1.7.1 Papillary Thyroid Carcinoma .................................................................................17
1.7.2 Poorly Differentiated and Anaplastic Carcinomas ................................................17
1.8 Summary ............................................................................................................................17
Chapter 2 Rationale and Objectives ...............................................................................................19
2 Rationale and Objectives...........................................................................................................19
2.1 Goal ....................................................................................................................................19

v
2.2 Specific Aims .....................................................................................................................20
Chapter 3 Methods .........................................................................................................................21
3 Methods .....................................................................................................................................21
3.1 General ...............................................................................................................................21
3.2 Detailed Methods ...............................................................................................................21
Chapter 4 Results ...........................................................................................................................30
4 Results .......................................................................................................................................30
4.1 STR Profile of Cell Lines ..................................................................................................30
4.2 Proteins Identified in Thyroid Carcinoma Cell Lines ........................................................30
4.3 Clinical Verification of Identified Proteins ........................................................................36
4.4 Expanded Proteomic Analysis of Thyroid Carcinoma Cell Lines .....................................48
Chapter 5 Discussion, Conclusion, and Future Directions ............................................................55
5.1 Discussion ..........................................................................................................................55
5.2 Conclusion .........................................................................................................................62
5.3 Future Directions ...............................................................................................................62
References ......................................................................................................................................64
Appendices .....................................................................................................................................81

vi
List of Tables Table 1. Thyroid Carcinoma US Incidence and Mortality by Histologic Subtype and Age – 1985-1995 – in 53 856 thyroid carcinoma Cases. …………………………………………….....2
Table 2. Thyroid Carcinoma, 5-year survival (%) and American Joint Committee on Cancer (AJCC) tumor stage. ………………………………………………………………………….….2
Table 3. The Histologic Variants of PTC. ………………………………………………………7
Table 4. Clinical Trial Agents for ATC Patients. ……………………………………………..13
Table 5. STR profile analysis of cell lines used in this study. ………………………………...30
Table 6. High-Confidence Proteins Identified in the Conditioned Media of TPC-1 and CAL62 Thyroid Carcinoma Cell Lines by Liquid Chromatography-Tandem Mass Spectrometry Analysis. ………………………………………………………………………………………..33
Table 7. High-Confidence Identifications in SW1736, C643, and BCPAP thyroid carcinoma. ………………………………………………………………………………49

vii
List of Figures Figure 1. Schematic for Workflow of Methods. ………………………………………………22
Figure 2. Schematic illustrating strategy for selection of a panel of secreted proteins for verification in thyroid carcinoma cell lines, tumor xenografts, patient tissues and sera..............23
Figure 3. Mass spectrum of prothymosin-alpha (PTMA). ….………………….……………....31
Figure 4. Subcellular locations of high-confidence proteins detected in conditioned serum-free media of CAL62 and TPC-1 thyroid carcinoma cell lines as suggested by Ingenuity Pathway Analysis (www.ingenuity.com). …………..…………………………………………………….32
Figure 5. Biological functions of proteins identified in the conditioned serum-free media of CAL62 and TPC-1 thyroid carcinoma cell lines…………………………….…..…………………………………………………………….32
Figure 6. Determination of subcellular localization of PTMA and nucleolin in cultured TPC-1
and CAL62 cells. ……….………………………………………………………………………38
Figure 7. Xenografts of TPC-1 cells in NOD/SCID/γ mice. ...………………………………..39
Figure 8. Immunodetection of identified proteins in sera of thyroid carcinoma
patients...……………………..……………………………….………………………………….41
Figure 9. Immunodetection of identified proteins in sera of thyroid carcinoma patients, and in
the whole-cell lysates and conditioned-free media of TPC-1 and CAL62 cells. ……..……..… 42
Figure 10. Immunohistochemical analysis of PTMA in human thyroid
carcinoma tissues. …………..……………………………….……………………………..…...44
Figure 11. Immunohistochemical analysis of nucleolin in human thyroid
carcinoma tissues. ……………………………………………………………………………….45
Figure 12. Positive and negative controls – representative photomicrographs for
immunohistochemical analysis of PTMA and nucleolin. ………………..................................46
Figure 13. Scatter plot Analysis of immunohistochemical scoring of PTMA
and nucleolin. …………………………………………………………………………………..47

viii
List of Appendices Appendix I ……………………………………………………………………………………...81
Appendix II ……………………………………………………………………………….........90
Appendix III ……………………………………………………………………………...........93
Appendix IV...……………………………………………………………………………........113
Appendix V...……………………………………………………………………………..........120

ix
List of Abbreviations ATA – American Thyroid Association ATC – anaplastic thyroid carcinoma BRAF – B-raf proto-oncogene CT – computerized tomography CYR61 – cysteine rich angiogenic inducer 61 variant DTC – differentiated thyroid carcinoma ELISA – enzyme-linked immunosorbent assay FBS – fetal bovine serum FNA – fine needle aspiration FTC – follicular thyroid carcinoma HAMA – heterophilic anti-mouse antibodies IHC - immunohistochemistry IMA – immunometric assay LC-MS/MS – liquid chromatography tandem-mass spectrometry MNG – multinodular goiter MTC – medullary thyroid carcinoma NCI – national cancer institute NIS – sodium-iodine symporter PBS – phosphate buffered saline PET – positron emission tomography PDC – poorly differentiated carcinoma PTC – papillary thyroid carcinoma PTMA – prothymosin-α RAI – radioactive iodine RIA - radioimmunoassay rhTSH – recombinant human thyroid stimulating hormone SCC – squamous cell carcinoma STR – short tandem repeat SEER – Surveillance Epidemiology and End Results Tg – thyroglobulin TgAb – thyroglobulin autoantibody TSH – thyroid stimulating hormone TTF-1 – thyroid transcription factor-1

x
Copyright Acknowledgments Content in this thesis have been reproduced with permission from:
Kashat, L.; So, A. K. C.; Masui, O.; Wang, X. S.; Cao, J.; Meng, X.; MacMillan, C.; Ailles, L.
E.; Siu, K. W. M.; Ralhan, R.; Walfish, P. G., Secretome-Based Identification and
Characterization of Potential Biomarkers in Thyroid Cancer. Journal of Proteome Research
2010, 9, (11), 5757-5769. Copyright 2010 American Chemical Society.

1
Chapter 1 Literature Review
1 Introduction 1.1 Thyroid carcinoma – General Overview Thyroid carcinoma is the most common endocrine malignancy, with an estimated annual
incidence of 122 800 cases worldwide and consists of a group of tumors with distinct clinical
features.1, 2 Papillary thyroid carcinoma (PTC), follicular thyroid carcinoma (FTC), and Hürthle
cell carcinoma are tumors that originate in thyroid follicular cells and are commonly referred to
as differentiated thyroid carcinoma (DTC).3 While management of these tumors has many
similarities, there are important diagnostic, therapeutic, and prognostic implications to consider
that are dependent on the tumor type.4 Anaplastic thyroid carcinoma (ATC), originating from
thyroid follicular cells and one of the most aggressive human malignancies, and medullary
thyroid carcinoma (MTC), a calcitonin–secreting tumor of the thyroid C cell are the two other
major forms of thyroid carcinoma.3, 5 Well-differentiated thyroid carcinomas often have an
indolent clinical course, with low morbidity and mortality, and with some important exceptions,
are considered among the most curable cancers.3 Distant metastases occur in up to 15% of
patients, with a significant decrease in 10-year disease-specific survival rates (40% compared to
85% in patients without distant metastases).6 In light of this fact, it is important to ensure that
both patients and physicians are vigilant when facing this disease and that long-term follow-up, a
crucial part of disease management, is carried out.
1.2 Incidence and Mortality An important study by Hundahl et al of the United States National Cancer Institute’s
Surveillance Epidemiology and End Results database (SEER) found 53 856 cases of thyroid
carcinoma treated in the USA and summarized incidence and mortality rates from 1985 to 1995
for each thyroid carcinoma subtype (Table 1).4 As thyroid carcinoma comprises only 2% of total
cancer cases in the USA, this study holds considerable importance for thyroid carcinoma
epidemiology. The incidence of PTC is significantly greater than all other tumor types and has
the most favourable survival rates. The 5-year survival rates for PTC, FTC, hurthle cell
carcinoma, and MTC patients sorted by American Joint Committee on Cancer (AJCC) stage (as
reported in the Hundahl et al study) are shown in Table 2.4 For PTC patients, there is a dramatic
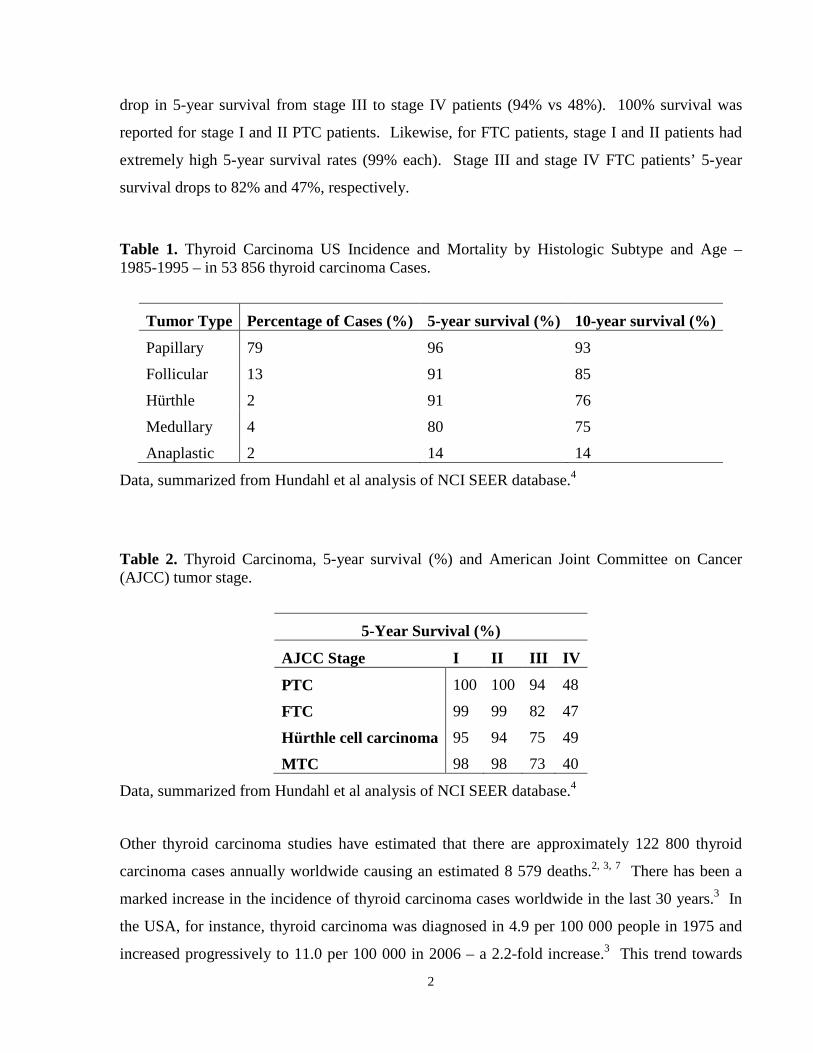
2
drop in 5-year survival from stage III to stage IV patients (94% vs 48%). 100% survival was
reported for stage I and II PTC patients. Likewise, for FTC patients, stage I and II patients had
extremely high 5-year survival rates (99% each). Stage III and stage IV FTC patients’ 5-year
survival drops to 82% and 47%, respectively.
Table 1. Thyroid Carcinoma US Incidence and Mortality by Histologic Subtype and Age – 1985-1995 – in 53 856 thyroid carcinoma Cases.
Tumor Type Percentage of Cases (%) 5-year survival (%) 10-year survival (%)
Papillary 79 96 93
Follicular 13 91 85
Hürthle 2 91 76
Medullary 4 80 75
Anaplastic 2 14 14
Data, summarized from Hundahl et al analysis of NCI SEER database.4
Table 2. Thyroid Carcinoma, 5-year survival (%) and American Joint Committee on Cancer (AJCC) tumor stage.
5-Year Survival (%)
AJCC Stage I II III IV
PTC 100 100 94 48
FTC 99 99 82 47
Hürthle cell carcinoma 95 94 75 49
MTC 98 98 73 40
Data, summarized from Hundahl et al analysis of NCI SEER database.4
Other thyroid carcinoma studies have estimated that there are approximately 122 800 thyroid
carcinoma cases annually worldwide causing an estimated 8 579 deaths.2, 3, 7 There has been a
marked increase in the incidence of thyroid carcinoma cases worldwide in the last 30 years.3 In
the USA, for instance, thyroid carcinoma was diagnosed in 4.9 per 100 000 people in 1975 and
increased progressively to 11.0 per 100 000 in 2006 – a 2.2-fold increase.3 This trend towards

3
increasing incidence has been widely observed throughout the world, including countries
throughout Europe, Asia, South America, and Oceania.8 The only countries to have reported
decreases are Sweden (18% reduction in both men and women), Norway (5.8% reduction for
women), and Spain (25.9% reduction in women). Other European countries reported increases
from 5.3% (Switzerland) to 155.6% (France). This increasing incidence in thyroid carcinoma is
primarily attributable to rising rates in PTC.3 The rates of FTC, MTC, and ATC have all
decreased in the same time period.3 The incidence in women is about 3-fold higher than in men,
but the disease appears to be more aggressive in men.3, 9 This is demonstrated in the finding that
while overall mortality rates have stabilized or improved from 1973 to 2001 according to records
in the SEER database, relative-survival comparisons show a significant decline in female
mortality (P<0.05) and increase in mortality in men (P<0.05).
The reasons behind the rising incidence of thyroid carcinoma are not yet known. Davies and
Welch examined the NCI SEER database and found that 87% of the increase in incidence from
1988-2002 were papillary cancers measuring 2 cm or less, with no significant increase in the
incidence of either FTC or MTC.10 In their study, nearly the entire rise in thyroid carcinoma
incidence from 1973-2002 was attributable to papillary carcinomas. They suggested, based on
previous autopsy studies which have indicated a large number of asymptomatic incidental
papillary carcinomas, this increase thyroid carcinoma incidence is due to greater diagnostic
scrutiny and not an actual increase in occurrence. If true, that the reported increase in incidence
is due to an increase in diagnosis of subclinical tumors, then the authors argued that the clinical
management of these tumors may need to be reconsidered.
In contrast, Mazafferri stressed the fact that small, asymptomatic cancers in the thyroid have the
potential to metastasize.11 A later study by Enewold et al examined 48 403 patients diagnosed
with thyroid carcinoma from 1980-2005 from the SEER database and found incidence varied
according to histology, gender, and ethnicity.9 If the increase in thyroid carcinoma incidence
was due solely to greater detection, the authors expected a more rapid increase in the incidence
of small early-stage tumors than large late-stage tumors and also expected that the rates for
larger, more advanced tumors would decline because of earlier detection and treatment. In the
39 706 PTC patients identified in their study, they found that half of the increase in incidence
was attributable to papillary thyroid microcarcinomas (<1 cm), 30% due to tumors measuring
1.1-2 cm, and 20% due to tumors >2 cm. They noted that incidence increased most rapidly

4
among women and that among White women, the rate of increase for PTC >5 cm was equal to
that for the smallest cancers. Although the greatest increases were seen in the smallest tumors
(≤1 cm), no corresponding decline in larger, more advanced tumors were observed.
Furthermore, the incidence rates of tumors of all sizes were found to increase. Furthermore, only
PTC was found to increase consistently over time and not tumors of all histologic types. If
improved diagnostics alone was to account for the increased incidence, then it would have been
expected that tumors of all histologic types should increase, except for anaplastic tumors, due to
its typical symptomatic presentation and aggressive course. Based on these findings, the authors
suggested that increased diagnostic scrutiny alone cannot account for the entire increase in
thyroid carcinoma incidence and other factors should also be explored. These findings were also
supported by another later study of the SEER database by Chen et al, which found that the
incidence of well-differentiated thyroid carcinomas of all sizes has been increasing in both males
and females from 1998-2005.3, 12
Another important study by Aschebrook-Kilfoy et al examined the incidence of thyroid
carcinoma among patients of varying ethnic backgrounds.13 The authors hypothesized that if
improved detection was primarily responsible for the increasing incidence of thyroid carcinoma,
then there would be demographic differences in incidence and different age-specific incidence
patterns, as those patients from poorer socioeconomic backgrounds would be expected to be
diagnosed later in life.8 Furthermore, some geographic-based variation in detection would also
be expected.8 None of these findings were observed in their study, leading the authors to
conclude that other factors in addition to improved diagnostics must be considered to account for
the rising incidence of thyroid carcinomas. Importantly, while changes in the classification of
thyroid carcinomas has resulted in many carcinomas being classified as follicular variants of
PTC rather than FTC, accounting for some of this increased incidence, it cannot account for all.3
Other possible sources include increased body mass index, radiation, and iodine intake.3
1.3 Prognostic Factors Histology
Tumor histology is crucial to patient outcome. The well-differentiated carcinomas generally
have excellent survival rates, while MTC and ATC have a much poorer prognosis.3 There are 15
histologic variants of PTC classified in the World Health Organization’s monograph on
endocrine tumors.14 Despite having remarkably varying histologic features, many of these

5
tumors have similar clinical behavior.3 Of the variants, tall cell, insular diffuse sclerosing, and
columnar variants of PTC are often found to be more aggressive.15 Insular and tall cell variants
of PTC are associated with numerous aggressive clinical parameters.3 Insular carcinomas have
been suggested to be classified as ‘poorly differentiated’ thyroid carcinomas as their clinical
course is an intermediate between PTC and ATC.3 In contrast, follicular, microfollicular,
pseudo-Warthin, clear cell, and cancers with lymphocytic stromal reactions are generally thought
to have similar prognosis to classical PTC.15 Oncocytic, solid, and trabecular variants of PTC
exhibit variable prognosis.15 The clinical features of the aggressive papillary thyroid carcinoma
variants and the variants that have been examined in our study will be described in more detail in
Section 1.4.
Tumor Size Among patients with PTC, tumor size correlates with outcome as larger tumors are more likely
to have metastasized at the time of presentation.16 In a study by Mazzaferri et al, the risk of
recurrence and cancer-specific mortality was found to increase linearly with tumor size, while
tumors <1.5 cm were found to have a 30-year cancer-specific mortality rate of 0.4%, in stark
contrast to the 22% mortality rate in tumors >4.5 cm.16
Lymph Node Metastasis About 15-30% of patients were found to have locoregional lymph node involvement at
presentation, but this has increased with the use of ultrasonography and its ability to identify
smaller nodal metastasis.3, 17 With prophylactic neck dissections, up to 50% of patients are found
to have locoregional disease; in children, even greater number of patients present with nodal
disease.3 There has been controversy with regards to the clinical significance of these findings.
Some studies have found nodal metastasis leads to reduced survival18 or increased risk of
recurrence16, while others found no difference in survival in patients with or without lymph node
metastasis.19 A 2008 analysis of the SEER database confirmed the associations of lymph node
metastasis with more aggressive clinical course.20 This study, however, determined the crucial
factor to be the age of the patient. In patients younger than 45 years, no effect on survival was
seen for patients with metastasis. In contrast, patients ≥45 years presenting with lymph node
metastasis had a 46% higher risk of death (p<0.001).
Extrathyroidal Extension

6
Gross extension of thyroid carcinoma into surrounding muscle, esophagus, or trachea is
associated with high-risk of recurrence.3 For these patients, aggressive surgical debridement is
suggested, and some studies have suggested beneficial effects of external beam radiation
therapy.3 Microscopic extrathyroidal extension is also associated with aggressive disease and
higher mortality.3, 16
Distant Metastasis This is the leading cause of death in patients with PTC.3 Mortality is high with distant disease
(50% at 3.5 years).21 Fewer than 10% of patients present with distant metastases and an
additional 2.5-5% will later develop them following initial radioiodine ablation.3, 21 In this
group, survival improves in younger patients and patients with iodine-avid tumors.21 Patients
who are older than 40 years have a greater extent of metastases and frequently have poorly
differentiated carcinomas and low 131I-uptake.22 A study of 444 patients of papillary and
follicular thyroid carcinoma patients with distant metastasis identified five variables that were
significant for survival: female, younger age, well-differentiated tumors, limited extent of
disease, and iodine-avid tumors.22 In patients <40 years of age with metastasis that were not
visible on radiographs or that were micronodular, 10-year survival was 95%. Importantly,
survival in patients with 131I-uptake was 92% at 10 years, compared to 29% in patients with 131I-
uptake and persistent abnormalities, and only 10% in patients with no initial 131I-uptake.22
Radioiodine ablation therapy was most efficient in patients younger than 40 years, and those
with well-differentiated subtypes and limited extent of disease.22
Oncogenes The most thoroughly investigated oncogenes is the B-raf proto-oncogene (BRAF).3 The
BRAFV600E mutation’s importance in diagnostics, prognostics, and therapeutics is still not
entirely clear.23 Over thirty studies on BRAFV600E and its characteristics in PTC have been
reported; the majority suggesting it is associated with advanced disease stage and aggressive
phenotypes.23 The presence of this mutation has been correlated with the worst outcomes for
patients with PTC and is associated with numerous other parameters of aggressive disease
(extrathyroidal invasion, multifocal tumor, nodal metastases, late-stage disease, older age, and
increased likelihood for recurrence).3 While the majority of studies have indicated this mutation
is involved in the aggressive behavior of tumors, some have not shown a significant association
and further work is required to provide clarity the matter.23

7
The BRAFV600E mutation can also be used to diagnose PTC patients.23 Since the mutation is
known to be exclusive to PTC (or ATC originating from PTC), DNA extracted from fine needle
aspiration (FNA) specimens can be used to improve diagnostics.23 For instance, Cohen et al
confirmed the mutation in 72% of carcinomas within the malignant samples they examined and
were able to establish a diagnosis for 16% of FNA specimens that had resulted in an
indeterminate result.24 Similar findings were observed in a study by Salvatore et al, where 5 of
15 indeterminate samples had a refined diagnosis by mutational analysis.25 The occurrence of
the mutation has varied in different studies, with pooled analysis showing up to 39% of tumors
harbouring this mutation.3
1.4 Variants of PTC Table 3. The Histologic Variants of PTC.
Follicular Macrofolliclar Oncocytic cell Clear cell
Diffuse sclerosing Columnar cell Tall cell PTC with insular
Cribriform Solid/Trabecular PTC with squamous cell carcinoma
PTC with medullary carcinoma
PTC with /focal anaplastic spindle and giant cell carcinoma
PTC with mucoepidermoid carcinoma
PTC with fasciitis or fibromatosis-like stroma
Table adapted from Albores-Saavedra et al14
PTC commonly presents as a thyroid nodule/mass, possibly in the background of a multinodular
goiter (MNG) or as a solitary cold nodule.26 The size of the tumor may be <1 cm to several
centimeters in diameter.26 The cyotologic diagnosis is based on characteristic nuclear features of
PTC, which show irregular nuclear membranes with “raisin-like” ovular (or ovoid) contours,
nuclear overlapping, powdery chromatin, micronucleoli, nuclear grooves, nuclear
pseudoinclusions, and optically clear nuclei dubbed “Orphan Annie eye nuclei.”14, 26 Classical
PTC exhibits papillary structures, which are lined up with characteristic nuclei.26 Many
histological variants of PTC have been described, some of which carry prognostic significance,
making them important to distinguish from classical PTC. While the aggressiveness of many
variants remains undetermined due to the relatively small number of published reports examining
their clinical course, the following section will review variants examined in this study and those
known to be associated with more aggressive disease.
Follicular Variant

8
The most common PTC variant is the follicular variant.27 The tumor contains few to no papillary
features and is mainly composed of small-to-medium sized follicles.14, 26 The cells lining the
follicles have enlarged ovular nuclei with intranuclear grooves.14 Diagnosing follicular variant
of PTC is complicated by the findings that many of these tumors arise in the background of
adenoma-resembling nodular goiters that lack capsular and vascular invasion.28 Furthermore,
some tumors show a multifocal, rather than diffuse distribution of typical PTC nuclear features.
Among this subtype, there are two distinctive groups: an encapsulated subvariant and an
infiltrative/diffuse one.26 The majority of the encapsulated follicular variants of PTC are
solitary, lack invasive characteristics, and confined to the thyroid.28 There have been many
reports suggesting the clinical behavior of these tumors is statistically similar to that of classical
PTC.15, 29-31
Oncocytic Variant
The oncocytic (Hürthle cell, or oxyphilic) PTC variant is composed of large polygonal cells with
granular eosinophilic cytoplasm and PTC nuclear characteristics.26 The granular appearance is
due to the predominant presence of mitochondrial-rich cells.14 Although the tumor contains
mitochondrial-rich cells, their nuclei have similar features as in classical PTC.14 Occasionally,
the tumor arises in Hashimoto’s thyroiditis and contains extensive lymphoplasmacytic
infiltrate.14, 32 Oncocytic variants of PTC should not be confused with Hurthle cell follicular
adenoma, which is benign or with Hürthle cell follicular carcinoma which may show focal
papillary architecture and are more clinically aggressive due to propensity for angioinvasion.14
Non-papillary carcinoma Hürthle cell tumors typically contain round and vesicular nuclei with
macronucleoli and dark chromatin, in contrast to the clear nuclei with intranuclear inclusions,
grooves and micronucleoli that typically are found in oncocytic PTC.32 The clinical
characteristics of the tumor are not clear, but there are reports it may be more aggressive than
classical PTC.33
Diffuse Sclerosing Variant
This variant of PTC is characterized by diffuse involvement of both thyroid lobes without
forming a localized mass and is usually seen in young patients.14, 26 Most patients are young
adult females and it has been reported in children and adolescents.26 Also characteristic is
diffuse stromal fibrosis, with dense sclerosis, lymphoid infiltration and squamous metaplasia.26
Extensive lymphatic space invasion and numerous psammoma bodies are also seen. This variant

9
is usually aggressive with many patients presenting with lung metastasis. With regards to
metastases, some studies have found that patients have similar long-term survival rates, while
others have found decreased disease-free survival.14, 15, 26, 34 While the number of cases
examined in the literature has not been large enough to draw strong conclusions about the
aggressiveness of this variant, the consistent findings of increased cervical lymph node and lung
metastases suggests clinicians should aggressively manage these patients to achieve the best
long-term clinical outcome.15
Columnar Cell Variant
The columnar cell variant is considered more aggressive than classical PTC because of its high
recurrence rate, quick growth, and frequent distant metastases.15 These findings are often based
on case reports, case series, and limited reviews, and as with the data on many other variants, is
not sufficient to draw firm conclusions regarding the aggressiveness of the variant. This variant
consists of tall cells with elongated hyperchromatic pseudostratified nuclei which lack the
cytologic features characteristic of the tall cell variant and classical PTC.15 Cells with
supranuclear and subnuclear cytoplasmic vacuoles are also common findings.14 Some studies
have shown that tumors with extracapsular invasion have poor prognosis.15
Tall Cell Variant
This variant of PTC has a worse prognosis and higher recurrence rate, usually presenting in older
male individuals.26 These tumors tend to be highly papillary in architecture with elongated
papillae, containing large eosinophilic cells.35 The tumor has been described as having a tall
columnar shape with cells whose height is at least three times their width15 and usually presents
with aggressive features such as vascular and extrathyroidal invasion.26 It is generally accepted
that these tumors have a higher recurrence, distant metastases, and death rate compared to
classical PTC and require more aggressive treatment.15, 35
Papillary Carcinoma with Insular Pattern
This aggressive variant of PTC is characterized by the presence of defined nests of tumor cells
containing monomorphic, dark, and round nuclei; i.e. they lack the classic nuclear features of
papillary carcinoma.15 The insular islands show numerous mitoses and foci of necrosis. Insular
tumors are often found to be >4 cm with other invasive features, such as extracapsular invasion,
lymph node metastases, and distant metastases.15, 36 It has been proposed that insular carcinomas

10
should be classified as “poorly-differentiated” thyroid carcinomas; however, this term remains
controversial and is not consistently used as a diagnosis by all pathologists.37 This designation
would also include other tumors, thought to be of aggressiveness between that of PTC and ATC,
such as those with a solid or trabecular growth pattern, again with loss of typical papillary
carcinoma nuclei, mitoses and necrosis. Distant metastases have been reported in as much as
70% of patients.38 Patients with a predominantly insular histology in a papillary or follicular
carcinoma have been shown to have a much more aggressive clinical course, including increased
mortality.36 While the tumor has been described as poorly-differentiated, its ability to take up I-
131 has been reported.36 One study found that 60% of patients with a predominantly insular
tumor pattern were able to take up I-131, and although none of these patients were cured, 44% of
patients benefitted from the treatment.36 Uptake of I-131 was higher in tumors immediately
following surgery, than in patients being treated for recurrent disease. These authors also
stressed the importance of the degree of insular pattern within the tumor, as they, along with
others have reported no difference in prognosis between patients with a focally insular
carcinoma.36 In contrast, other authors have found that all insular tumors are aggressive
regardless of the extent.39-41
1.5 Anaplastic Thyroid Carcinomas Anaplastic thyroid carcinoma is one of the most aggressive human malignancies and, with very
few exceptions, is almost always fatal.42 Of the 1200 thyroid carcinoma deaths in the USA in
2006, over 50% were due to ATCs although it accounts for less than 2% of all thyroid
carcinomas.42 The prognosis for patients with ATC is bleak, with studies showing a median
survival time that from 4-12 months.5 A review of numerous studies that followed the outcome
of a total of 1771 ATC patients treated from 1947-2007 found the median survival of all series to
be 5 months.5 In concordance with these findings, a review of 516 patients in the SEER database
found a 19.3% 1-year survival.43 Although there are rare descriptions of long-term survivors,
diagnosis is often questioned in these reports, especially in the few cases of survival >5-years.44
ATC usually affects older patients, with the mean age at diagnosis 55-65 years old, but can also
affect younger patients.44
Clinical Presentation
Patients with ATC usually present with a rapidly growing thyroid mass and symptoms related to
the mechanical compression on or invasion of surrounding structures.45 The mean tumor size

11
has been reported to be 7-8 cm, but can range from 3 cm – 20 cm.44 In addition to the rapidly
growing thyroid mass, mechanical compression on surrounding structures may lead to dyspnea,
stridor, dysphagia, neck pain, and hoarseness.44 Approximately half of patients will have
metastases to lymph nodes or distant sites upon presentation and another 25% will develop them
over the course of the disease.44, 45 Metastatic sites include lungs (80%), bone (6-15%), and
brain (5-13%).42 Cardiac and intra-abdominal metastases have also been reported.44 The
tumor’s aggressive nature is highlighted in the finding that it is not unusual for its volume can
double in the span of one week.46
The histological variants of ATC – spindle cell, giant cell, and squamoid – all have similar
prognosis.44 It is hypothesized that well-differentiated thyroid carcinomas may progress towards
ATC through the dedifferentiation of insular variants of PTC/FTC.44 This notion is further
supported by the finding that up to 90% of ATCs have co-existing regions of differentiated
thyroid carcinoma.46 Immunohistochemically, overexpression of p53 has been detected, may
reflect altered function of the protein, and may also play a role in the de-differentation of well-
differentiated tumors.46, 47 Mutations that have been described in ATC include the following
genes: p53, RAS, BRAF, β-catenin, PIK3CA, Axin, APC, and PTEN.47 Furthermore,
abnormalities in chromosome integrity and number have been identified in nearly every
chromosome illustrating the high level of genomic disarray in ATC and complicating the search
for potential therapeutic targets.47
Prognostics
As mentioned previously, some ATC patients have demonstrated longer survival times although
the disease is nearly always fatal. Younger age (<45), female sex, smaller lesions, small foci of
ATC, no evidence of metastatic disease, and surgery for locoregional disease are considered
favourable prognostic factors.44, 45 When analyzed by stage according to Sixth AJCC edition for
ATC, stage IVA, IVB, and IVC patients had 22.9%, 10.1%, and 0% 5-year overall survival
respectively.45
Treatment
There is currently no consensus on the management of ATC patients due to the lack of
convincing evidence of various therapeutic approaches. In general, a combination of surgery,
radiation therapy, and chemotherapy may possibly improve survival.45

12
The vast majority of patients have disease that is so invasive, it is beyond meaningful resection.44
While some reports have suggested that potentially curative complete surgical resection of tumor
along with post-operative external beam radiation therapy and/or chemotherapy, led to increased
survival there are also reports of neither extent nor the completeness of surgery had bearing on
survival.44 Surgery is also considered for potential palliative benefits, including to help prevent
death by asphyxiation.44
The majority of patients die from uncontrolled local symptoms and local control has been shown
to improve the short-term survival of patients.44 One study found that among the 51 patients
treated with radiation therapy over 25 years, median survival improved to 7.5 months when local
control was achieved (compared to 1.6 months without local control) – even in the presence of
distant metastases.48 Many studies have also demonstrated that combining radiation therapy with
chemotherapy may significantly prolong short-term survival in patients, as evidenced by some
studies which have shown patients surviving >2 years.44, 49 Based on these findings, it appears
that while radiation therapy cannot alter the course of the disease, it can have a beneficial effect
in a select population of ATC patients.
To date, the outcomes of patients with chemotherapy have largely been disappointing. Efforts
have been made using doxyrubicin monotherapy, combination therapy (cisplastin, bleomycin,
melphalan), and newer agents such as paclitaxel.44 Most studies have reported only a few
patients with partial responses and almost no patients with a complete response.
ATC treatment relies on multimodality therapy since no single treatment option has succeeded.
For instance, in the studies demonstrating improved survival in ATC patients with radiation
therapy, many have used chemotherapy to improve sensitivity to radiation therapy.
Nevertheless, treatment efficacy has been dismal and there is a dire need for new, innovative
therapeutic techniques. Several clinical trials are underway or are recruiting patients with ATC,
some of which are listed in Table 4.

13
Table 4. Clinical Trial Agents for ATC Patients.
Imatinib Bcr-Abl protein tyrosine kinase inhibitor
Sorafenib Tyrosine kinase inhibitor
Combrestatin (in combination with
paclitaxel/carboplatin)
Natural stilbenoid phenol that inhibits β-
tubulin
Bevacizumab (in combination with
doxorubicin)
Monoclonal antibody to vascular endothelial
growth factor
Pazopanib Tyrosine kinase inhibitor with anti-angiogenic
properties
Pemetrexed (in combination with paclitaxel) Folate antimetabolite
As reviewed in Pitt et al45
Interestingly, gene therapy targeting a sodium iodine symporter might allow for the application
of RAI therapy to dedifferentiated carcinomas. The sodium-iodine symporter (NIS) a membrane
protein that mediates follicular cells to actively transport iodine into the thyroid and some
extrathryoidal tissues loses its function in ATC due to decreased expression.42 Accordingly,
increasing expression of the protein in ATC may allow patients to benefit from RAI therapy.
There are a few sources of evidence suggesting this approach may be feasible. Transfection of
human NIS into a NIS-deficient FTC cell line led to an increase in vivo iodide accumulation in
xenografted tumors.50 Likewise, ATC cells transfected with human NIS accumulated
radioiodide in vitro and in vivo.51 I-131 effectively inhibited tumor growth in mice bearing these
tumors compared to controls. Furthermore, gene therapy targeting TTF-1 and Pax8 which
upregulate thyroid-specific genes including NIS, may help cells “redifferentiate” and improve
their ability to uptake radioiodine.52 Experimental approaches have the promise of greatly
improving in the survival of ATC patients.
1.6 Molecular Diagnostics and Serum Thyroglobulin Thyroglobulin (Tg) is synthesized as a prohormone for thyroid hormone in follicular cells of the
thyroid and released into blood as a byproduct of normal secretion of thyroid hormone.53 This is
also the case for most differentiated thyroid carcinomas, although the secreted Tg may have
different molecular conformations.54, 55 Since Tg is a thyroid-specific antigen, persistent

14
elevated readings may be the result of residual malignancy.56 Three factors affect post-operative
thyroglobulin levels in a patient: 1) secretions from remaining normal thyroid tissue and tumor,
2) effects of any thyroid injury secondary to FNA, RAI therapy, thyroidectomy, or inflammation
associated with thyroiditis, and 3) TSH-receptor stimulation from endogenous TSH, recombinant
human TSH (rhTSH), human chorionic gonadotrophin during pregnancy, or stimulating anti-
TSH receptor antibodies.53
There are two types of assays used to determine serum Tg concentrations, competitive
radioimmunoassay (RIA) and non-competitive two-stage immunometric (IMA).53 Briefly, in the
RIA, patient serum competes with I-125-labelled human Tg for binding to a limited amount of
high-affinity, rabbit polyclonal antibody. In this assay, the antibody must be excessively dilute
resulting in incubation times that last for days before the Tg-antibody complex can be
precipitated with an anti-rabbit secondary antibody. The amount of radioactivity precipitated in
the complex is inversely proportional to serum Tg concentrations. In the much more commonly
used, non-competitive “sandwich” two stage IMA, Tg in patient serum binds to an excessive
amount of anti-human-Tg mouse monocolonal antibody that is already bound to solid support. A
second (chemilumincently) labelled mouse monoclonal antibody to a different epitope of Tg is
then added following the washing of unbound constituents. After a brief incubation (<1 hour),
unbound antibodies are washed away. The detected chemilumniescence is proportional to the
serum Tg concentration.
There are important considerations which may negatively affect results and must be considered.
Due to interassay variation, serial samples from the same patient must be measured using the
same assay.57, 58 Variations are due to variations in antithyroglobulin antibodies used and
heterogeneity of Tg and splice variants; this heterogeneity can be even greater in Tg secreted by
cancer cells. In addition to this interference from heterophilic anti-mouse antibodies (HAMA) or
autoantibodies to Tg can cause false positive and false negative readings. Although rare, if a
patient has HATA, they can cross-link both the “capture” and secondary antibody in the absence
of Tg and produce false positive readings or also, false negatives, by preventing Tg binding.59
Further complicating matters is the presence of thyroglobulin autoantibody (TgAb), which is the
most serious problem limiting clinical utility of Tg testing.53 TgAb is found in approximately
20% of patients with DTC (vs 10% in the general population) and it is recommended to also
measure their levels with each measurement of serum Tg.60, 61 By binding to serum Tg, anti-Tg

15
autoantibodies may reduce the amount of unbound Tg circulating in the serum of patients and
prevent detection – an effect that still exists when using monoclonal antibodies directed against
epitopes of Tg that do not react with the autoantibodies.62 The effect of TgAb exists even in low
concentrations of the autoantibodies.53
Importantly, TgAb concentrations that persist more than one year after thyroidectomy and RAI
remnant ablation indicate a possible risk of recurrence.60 For example, in two studies of thyroid
carcinoma patients with undetectable serum Tg concentrations, 18% and 49% of patients with
serum anti-Tg antibody concenrtations >100 U/mL had a recurrence, with only 1% and 3% of
patients with serum anti-Tg antibody concentrations <100 U/mL.63, 64 No patients with TgAb
concentrations falling >50% within a year of RAI remnant ablation had a recurrence, while 37%
of patients experiencing an increase had a recurrence.64
The functional sensitivity of most serum Tg assays has been about 0.9 ng/mL, but several recent
assays that are commercially available have improved this to 0.1 ng/mL or slightly lower.58, 65
To enhance the test’s sensitivity, Tg levels are measured following TSH stimulation (by thyroid
hormone withdrawal or the administration of rhTSH). When using the less sensitive assays, TSH
stimulation can make measurable the previously undetectable levels of serum Tg in as many as
25% of patients.66 The need for TSH stimulation may decrease with the use of the more
sensitive assays.58 Importantly, “hook effect” may contribute to a failure to detect serum Tg
because extremely high concentrations of Tg may bind to each antibody and prevent the
formation of the two-antibody sandwich. When this is the case, the sample should be diluted and
repeated.67
Some of the clinical uses of serum Tg testing are summarized below.
Detection of Persistent Disease
A meta-analysis identified the sensitivity and specificity of Tg to detect persistent thyroid
carcinoma following thyroid hormone withdrawal to be 96% and 93% respectively, and 93% and
88%, respectively, after rhTSH adminstration.68 In another study of 340 patients found that
when combined with cervical ultrasound, sensitivity and negative predictive value for serum Tg
following rhTSh administration were 93% and 99%, respectively.69 Some studies have also
suggested that repeating rhTSH-stimulated Tg testing in patients whose first reading was
undetectable may not be useful. For instance, Castanga et al found that of 68 patients with

16
stimulated Tg <1 ng/mL at the time of RAI remnant ablation, only 1 of 67 had a detectable
rhTSH-stimulated Tg reading up to 3 years later.70
Detecting Disease Recurrence
Since serum Tg concentration depends on secretions from both normal thyroid tissue and
differentiated cancerous remnants, the sensitivity and specificity of serum Tg values are highest
following total thyroidectomy and RAI ablation of any residual normal thyroid cells (as
previously discussed in Section 1.3.3).71, 72 Serum thyroglobulin is cleared with a half-life of
about 30 hours following thyroidectomy and are expected to become undetectable if a patient is
cured, although this may take a year or longer.71, 73, 74
Predicting Clinical Outcome
Serum Tg has been shown to have the potential to predict disease-free remission during
treatment.75 Likewise, the serum Tg concentration in low risk patients following initial surgery
while hypothyroid, prior to administration of RAI, has been correlated with clinical course.76
Radioiodine Remnant Ablation Treatment Selection
As discussed in Section 1.3.3, RRA decision making is often complicated by numerous factors,
including uncertainty of the impact of RRA on disease recurrence in low risk patients. While
some advocate for its extensive use, others have suggested a more conservative approach,
particularly in the low-risk patient group. A study by our group at Mount Sinai Hospital
presented a novel suggested use for stimulated-Tg measurements as an objective parameter to
assist in RAI remnant ablation decision making.77 In this study, patients with <1 ug/L
stimulated Tg did not receive RAI remnant ablation and would be followed up (ex. neck
ultrasound, repeat yearly stimulated-Tg). Patients with 1-5 ug/L stimulated-Tg were considered
for RAI remnant ablation based upon factors such as aggressive histology, nodal metastases,
gender, neck ultrasounds, and the consideration of patient’s individual attitude towards RAI
remnant ablation (fertility goals, comorbidity concerns) – follow-up with yearly stimulated-Tg
tests would also occur for these patients. Finally, patients with stimulated thyroglobulin >5ug/L
received RAI remnant ablation. In the study of 104 patients, 59 patients had undetectable
stimulated-Tg following thyroidectomy, 35 had stimulated-Tg values of 1-5 ug/mL and 10 had
stimulated-Tg values >5 ug/mL. RRA was administered to one patient with undetectable Stim-
Tg, 6 patients with 1-5 ug/mL, and 9 patients with stimulated-Tg >5 ug/mL. The use of

17
stimulated-Tg helped to significantly reduce the need for RAI remnant ablation and provided an
objective tool for patient assessment and decision-making.
1.7 Mutations in Thyroid Carcinoma 1.7.1 Papillary Thyroid Carcinoma The most essential pathway involved in the pathogenesis of papillary carcinoma is the mitogen-
activated protein kinase (MAPK) pathway, a regulator of cell differentiation, survival, and
growth.78 This pathway is activated due to point mutations in the BRAF and RAS genes, and also
because of rearrangement of RET and NTRK1 genes.79 One of these mutations is found in over
70% of PTCs and they rarely overlap within the same tumor, with the BRAF being the most
common mutation in papillary tumors.79 Studies have suggested that this mutation is rare in
follicular variants of PTC, and common to classical papillary carcinomas and its tall cell
variant.80, 81 As discussed earlier (Section 1.3.1), the BRAF mutation appears to correlate with
aggressive tumor characteristics.
1.7.2 Poorly Differentiated and Anaplastic Carcinomas It is thought that poorly differentiated carcinoma may arise from the partial de-differentiation of
PTC, FTC, and de novo .82 While most PTC usually possesses a normal karyotype, ATC is
characterized by multiple numerical and structural chromosomal aberrations.83, 84 Interestingly,
suggesting they may not be involved in the dedifferentiation of tumors.79 On the other hand,
BRAS and BRAF appear to be common to both well-differentiated and undifferentiated tumors,
suggesting they are early-mutational events in thyroid tumorigenesis.79 Mutations in the
TP53and β-catenin genes frequently occur only in poorly-differentiated and anaplastic
carcinomas and accordingly, are believed to be late-events in thyroid tumorigenesis involved in
the progression of PTC and FTC tumors.82
1.8 Summary Although a lot is known about thyroid carcinoma, there remains a strong need for biological
markers to help in the management of patients. One of the greatest limitations of current
techniques is the profound lack of biochemical markers (particularly, serum-based markers) to
help in prognostication and diagnostics during treatment. Serum Tg, although a key tool in the
management of patients, has many limitations including limited ability to aid in prognostication,
the interference problem posed by autoantibodies, and the fact secretion is limited to

18
differentiated carcinomas. Because of these limitations, patient follow-up currently relies
heavily on diagnostic imaging, which is both expensive and time-consuming. Furthermore, there
is significant ambiguity and controversy regarding the extent of treatment required to provide
patients with optimal benefits – particularly in lower-risk patients with more favourable tumors.
Are lobectomies enough in some patients? Which patients require RAI remnant ablation?
Biochemical markers may help bring a degree of objective analysis to decision-making may help
in this regard. Finally, as highlighted here, the treatment of poorly and undifferentiated
carcinomas poses a strong therapeutic challenge. New therapeutic targets and strategies are
urgently needed to improve the dismal outlook for many of these patients, particularly those with
anaplastic thyroid carcinomas.

19
Chapter 2 Rationale and Objectives
2 Rationale and Objectives As discussed in the literature review there is a lack of molecular markers to predict the
aggressiveness of thyroid carcinomas. Currently fine needle aspiration (FNA) is the most
accurate preoperative technique for diagnosis of thyroid nodules. However, even using
ultrasound-guided FNA, inconclusive biopsy results are quite common (10-20% of all cases).85
Many of these patients may undergo surgery to remove their thyroid gland – a procedure that is
sometimes unnecessary as many suspected nodules are benign.82 Additionally, though most
papillary thyroid carcinomas are non-aggressive and often non-metastatic, a small percentage are
in-fact aggressive and may develop distant metastasis leading to higher mortality.86 This
establishes an urgent need for identifying biomarkers to distinguish benign thyroid nodules from
malignant and aggressive carcinomas. Serum-based biochemical markers are of particular
interest because they are minimally-invasive, cost-effective, and may be used throughout
treatment to monitor patients for recurrence and to aid in treatment decision-making.
The tumor cells and their interactions with the host’s microenvironment play vital roles in tumor
growth, invasion, and metastasis.87 The cancer cells and the host’s microenvironment secrete
and shed proteins or their fragments extracellularly and into bodily fluids, including blood.
These proteins and their fragments constitute the “cancer secretome”.88 Sampling of bodily
fluids is minimally invasive and multiple samples drawn over a period of time can provide
longitudinal data during the course of disease investigation or treatment. In view of this,
analyses of proteins in serum and saliva using mass spectrometry (MS)-based proteomic
technologies have been carried out.89-91 Proteins secreted by cancer cells into their culture media
(“secretome” proteins) make especially appealing targets for study because they may be
detectable in bodily fluids.92-98
2.1 Goal The goal of this study is to use proteomics for secretome analysis of cultured thyroid cancer cells
to identify candidate secreted proteins that may serve as biological markers for aggressive
thyroid carcinomas, to aid in the management of these patients.

20
2.2 Specific Aims AIM 1: Proteomic analysis of secretomes of thyroid carcinoma cell lines for identification of
candidate biological markers for aggressive thyroid carcinomas.
AIM 2: Verification of a panel of secreted proteins in thyroid carcinoma cell lines and their
tumor xenografts.
AIM 3: Verification of select proteins in thyroid carcinoma patients’ tissues and sera.

21
Chapter 3 Methods
3 Methods
3.1 General In this study, we have analyzed the conditioned-media of five thyroid carcinoma cell lines using
one-dimensional LC-MS/MS to identify putative secreted biological markers. We have validated
many of these proteins in the cell lines and their xenografts in immunocompromised mice,
patient tissue samples and sera and done so using numerous means including
immunohistochemistry (IHC), western blot, and immunofluorescence. Our first analysis was
with TPC-1 (papillary-derived) and CAL62 (anaplastic-derived) cells. Based upon our findings
in our first analysis, we later completed a second proteomic analysis of additional cell lines:
BCPAP (papillary-derived) and SW1736, C643 (anaplastic-derived) cells. The workflow of this
study is shown in Figure 1.
3.2 Detailed Methods
Cell Lines
Five thyroid carcinoma cell lines, TPC-1, BCPAP (derived from a human papillary thyroid
carcinoma) and CAL62, SW1736, C643 (derived from a human anaplastic thyroid carcinoma)
were used in this study.99-101 TPC-1 cell line was kindly provided by Dr. S. Jiang (The Ohio
State University, Columbus, Ohio) and CAL62 by Dr. J. Knauf (Sloan-Kettering Institute, New
York, NY) with permission from Dr. M. Santoro (Medical School, University “Federico II” of
Naples, Naples, Italy). SW1736 was provided by Dr. E. Heldin (Rudbeck Laboratory, Uppsala
University, Finland), C643 by Dr. G. Salvatore (University of Naples, Italy). BCPAP is
available for purchase from the German Collection of Microorganisms and Cell Cultures
(Braunschh, Germany). To ensure the problem of cross-contamination and misidentification of
cell lines was avoided, short tandem repeat (STR) profiles of each cell line were determined to
match those of the original thyroid-derived cell lines as reported in previous studies by Schweppe
et al101 and / or in the American Type Culture Collection (ATCC) and German Collection of
Microorganisms and Cell Cultures (DSMZ). Previously published studies with some of these

22
cell lines have demonstrated the expression of thyroid specific genes in these cell lines
confirming their thyroid origin.101, 102
Figure 1. Schematic for Workflow of Methods. Revised figure reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.

23
Figure 2. Schematic illustrating strategy for selection of a panel of secreted proteins for verification in thyroid carcinoma cell lines, tumor xenografts, patient tissues and sera.

24
Cell Culture and Serum Free Media Collection
TPC-1, BCPAP, C643, and SW1736 cells were propagated in 25 mL of RPMI-1640 containing
100 µg/mL streptomycin and 100 U / mL penicillin, 10% fetal bovine serum (FBS) and 1% non-
essential amino acids in 150 mm dishes to about 65% confluence. CAL62 cells were propagated
in 25 mL of Dulbecco's Modified Eagle's Medium (DMEM) with high glucose containing
streptomycin, penicillin and 10% FBS. Cells were incubated at 37°C in a humidified atmosphere
of 5% CO2 - 95% air. The culture media were then aspirated and cells were washed three times
with phosphate-buffered saline (PBS). Thereafter, cells were washed once with serum-free
culture media that was collected as a time 0 h control. Cells were then incubated in the serum-
free culture media for 48 h. Thereafter, the conditioned media were collected, centrifuged at
5000 g for 5 minutes at 4°C, filtered using a 0.2 µm nylon filter, snap frozen and stored at -80°C
until further use. Media was collected from at least 15 plates. Trypan blue staining was
performed following collection of the conditioned media at 24 h and 48 h to estimate the number
of dead cells. Since more than 98% cells were viable at 48 h, this time period was chosen for
further study.
Optimization of Cell Culture Conditions for Collection of Conditioned Media
Cells are routinely grown in cell-culture media containing fetal bovine serum, however, the high
abundance proteins present in serum would interfere with the detection of secreted proteins. For
this reason, cell culture conditions needed to be optimized for conditioned media collection. To
avoid this interference, the cells were washed thoroughly four times (three times with PBS and
once with serum-free media) and then grown in conditioned media for 48 h, allowing secreted
proteins to accumulate. To limit cellular stress under these conditions, cells were only placed in
serum-free culture media when they reached about 60% confluence.
Protein Precipitation from Conditioned Medium and LC-MS/MS Analysis
Proteins were precipitated from the pooled conditioned media using 0.2% sodium deoxycholate
(Sigma Aldrich, St.Louis, MO) and 10% trichloroacetic acid (Sigma Aldrich, MO) as described
earlier.103 Following 2 h incubation on ice, the samples were centrifuged at 11 000 g for 30
minutes and washed twice with ice-cold acetone. The precipitated proteins were then dissolved
in 50 mM NH4HCO3 buffer, pH 7.5. The protein concentration was determined using the
Bradford assay (Bio-Rad, Hercules, CA). Protein samples were then heated for 1 h at 65°C in the

25
presence of 5 mM dithiothreitol, cooled to room temperature, and incubated in dark for 1 h with
10 mM iodoacetamide for alkylation. Sequencing grade trypsin (Promega, WI) at 1:20 (w/v) in
50 mM ammonium bicarbonate was subsequently added and the samples were incubated at 37°C
overnight. The trypsin digested samples were then dried under vacuum and dissolved in 10 µL of
0.1% formic acid. Experiments were repeated twice and each set was analyzed separately using
LC-MS/MS.
Liquid Chromatography – Tandem Mass Spectrometry
The trypsin digested samples were analyzed using online LC-MS/MS. The nanobore LC system
(LC Packings, Amsterdam, The Netherlands) and mass spectrometer (QSTAR Pulsar, Applied
Biosystems/MDS SCIEX, Foster City, CA) have been described by some of us earlier.92, 104 One
µL aliquot of the sample was loaded onto a C18 reverse-phase precolumn (LC Packings: 300 µm
x 5 mm) and desalted before separation on an RP analytical column (75 µm x 150 mm packed
in-house with 3-µm Kromasil C18 beads with 100 Å pores, The Nest Group). We used a
nonlinear binary gradient: eluant A consisting of 94.9% deionized water, 5.0% acetonitrile, and
0.1% formic acid (pH 3); and eluant B consisting of 5.0% deionized water, 94.9% acetonitrile,
and 0.1% formic acid for the separation. Eluant A was used to load the sample onto the C18
precolumn at a flow rate of 25 µL min-1. After 8 min, the C18 precolumn was switched inline
with the reverse-phase analytical column; separation was performed at 200 nL min-1 using a 180-
min binary gradient shown below.
Time (min) 0 5 10 120 140 145 155 157 189
B (%) 5 5 15 35 60 80 80 5 Stop
MS data were acquired in information-dependent acquisition (IDA) mode with the Analyst QS
1.1 and Bioanalyst Extension 1.1 software (Applied Biosystems/MDS SCIEX). MS cycles
comprised a TOF MS survey scan with a mass range of 400-1500 Da for 1 s, followed by five
product-ion scans with a mass range of 80-2000 Da for 2 s each. The collision energy (CE) was
automatically controlled by the IDA CE Parameters script. Switching criteria were set to ions
with m/z ≥ 400 and ≤1500, charge states of 2-4, and abundances of ≥10 counts. Former target
ions were excluded for 30 s, and ions within a 6-Da window were ignored. Additionally, the
IDA Extensions II script was set to “no repetition” before dynamic exclusion and to select a
precursor ion nearest to a threshold of 10 counts on every fourth cycle.

26
Bioinformatics – SignalP and SecretomeP – Determination of Secretory Proteins
LC-MS/MS data were searched using the ProteinPilot software (Applied Biosystems, Foster
City, CA), which uses a Paragon Algorithm105 against a Celera human protein database (CDS
KBMS 2004112009) containing 178239 protein sequences. The cut-off for significance used for
this search was set for a score of 1.3, which corresponds to a confidence score of 95%. We used
Signal Peptide Predictor (SignalP, http://www.cbs.dtu.dk/services/SignalP 3.0) to analyze the
secretion features of identified proteins.106 SignalP uses amino acid sequences to predict the
existence and location of signal peptide cleavage sites. SignalP determines the likelihood a
protein is a signaling peptide using numerous artificial neural networks and hidden Markov
model algorithms to detect signal peptides in protein sequences. A protein is considered
classically secreted if it receives a signal peptide probability ≥ 0.9.
In order to identify non-classical, or leaderless, protein secretion SecretomeP
(http://www.cbs.dtu.dk/ services/SecretomeP 2.0) was used.107 SecretomeP uses a neural
network that combines six protein characteristics to determine if a protein is non-classically
secreted. These characteristics include: the number of atoms, number of positively charged
residues, presence of transmembrane helices, presence of low-complexity regions, presence of
pro-peptides, and subcellular localization. A protein is considered non-classically secreted if it
receives an NN-score ≥0.5 (note: only proteins that were not considered classically secreted, i.e.
received SignalP scores <0.900, were analyzed using SecretomeP). It is important to note that
some secretory proteins may not make the SignalP and SecretomeP score cutoffs. Ingenuity
Pathway Analysis (IPA, Ingenuity Systems, www.ingenuity.com) was used to determine the
subcellular localization and biological functions of the identified.
Selection of a Panel of Secreted Proteins
As presented in Figure 2, only proteins identified with a minimum of 2 peptides ≥95 confidence
were considered for this study. This narrowed our proteins of interest from 154 identifications to
46. An exception was made for PTMA, which has been found to play a role in many aggressive
cancers including a head and neck carcinoma study by some in our group.108 Careful literature
reviews were performed using the U.S. National Center for Biotechnology Information PubMed
database (http://www.ncbi.nlm.nih.gov/pubmed/) using common names of the identified
proteins, as suggested by uniprot database (http://www.uniprot.org/). This search was combined

27
with terms including: thyroid carcinoma(s), thyroid cancer, and thyroid nodule(s). Six proteins
were selected for consideration on the basis of reported associations with cancer aggressiveness,
reported detection in the sera of cancer patients, and potentially important biological roles in
cancer progression.
Immunofluorescence
The detectability of prothymosin-alpha (PTMA) and nucleolin in TPC-1 and CAL62 cells was
determined using immunofluorescence to confirm these secretome proteins originated from
thyroid carcinoma cell lines. Furthermore, the subcellular localization of these proteins in TPC-1
and CAL62 cells was compared with their localization in xenografts and human thyroid
carcinoma tissues to demonstrate that their expression patterns are similar in these systems and
human thyroid carcinoma tissues. Cells were grown on glass slides up to 60% confluence. The
cells were then incubated with a primary antibody: nucleolin mouse monoclonal antibody
(Invitrogen, Camarillo, CA, 1:100 dilution) or PTMA rabbit polyclonal antibody (Santa Cruz
Biotechnology, Santa Cruz, CA, 1:100 dilution). The secondary antibody used was a fluorescein
isothiocyanate (FITC)-conjugated anti-mouse antibody or a tetramethyl rhodamine
isothiocyanate (TRITC)-conjugated anti-rabbit antibody (Sigma-Aldrich, 1:200 dilution). Slides
were viewed using an Olympus Upright fluorescence microscope (BX61) and images were
analyzed using Volocity software (PerkinElmer, Waltham, MA).
Detection of Potential Protein Biomarkers in Thyroid Carcinoma Patients’ Sera by
Western Blotting
Western blots were used to verify the expression of selected secretory proteins in thyroid
carcinoma patients’ sera, nucleolin, cysteine rich angiogenic inducer 61 variant (CYR61),
clusterin, enolase 1, biotinidase and PTMA. Further, the conditioned serum-free media and
whole-cell lysates of TPC-1 and CAL62 cells were also examined by immunoblotting to confirm
the detection of these proteins. Cell lysates were prepared by resuspending cells in lysis buffer
(100 mM Tris-HCl, pH 6.8; 100 mM dithiothreitol; 50 mM sodium dodecylsulfate; 0.7 M
glycerol) and boiling for 5 min. Sera (8 μL) from thyroid carcinoma patients were treated and
concentrated using the ProteoPrep 20 Immunodepletion Kit (Sigma). Proteins were separated by
denaturing electrophoresis on 10% (v/v) polyacrylamide and blotted onto polyvinylidene fluoride
membrane (Millipore). Blots were pre-incubated overnight at 4°C in a blocking solution

28
consisting of 5% (w/v) skimmed milk powder in tris-buffered saline tween-20 (20 mM Tris-HCl
pH 7.6, 140 mM sodium chloride, 0.1% Tween 20, TBST). Primary antibody dilutions were
prepared with 2% (w/v) gelatin in TBST prior to incubation (1 h) with the following antibodies
from SantaCruz Biotechnology: CYR61 rabbit polyclonal antibody (1:100 dilution); clusterin
mouse monoclonal antibody (1:200 dilution); nucleolin mouse monoclonal antibody (Invitrogen;
1:100 dilution); enolase 1 mouse monoclonal antibody (1:100 dilution); actin mouse monoclonal
antibody (1:500 dilution); and PTMA mouse monoclonal antibody (1:50 dilution) obtained from
Lifespan Biosciences, Seattle, WA. Following incubation, blots were washed (3 x 5 min) with
TBST and incubated (45 min) with horseradish peroxidase-conjugated goat anti-mouse or anti-
rabbit IgG (Santa Cruz Biotechnology) at a dilution of 1:5000 in TBST containing 2% gelatin.
Blots were washed again with TBST and developed using enhanced chemiluminescence
detection (GE Healthcare, Mississauga, Ontario, Canada). Each experiment was repeated at least
twice.
Immunohistochemical Analysis of PTMA and Nucleolin Proteins in Thyroid Carcinomas
Immunohistochemistry was performed on archived human thyroid carcinoma tissues and the
adjacent normal thyroid tissue from patients with benign thyroid disease, in order to determine
the expression profiles of PTMA and nucleolin. For PTMA, a total of 55 thyroid carcinoma
tissues were examined (39 PTC, 6 insular, and 10 ATC) and 20 normal tissues. For nucleolin, 48
thyroid carcinoma tissues were examined (37 PTC, 6 insular, 5 ATC) and 20 normal tissues.
Serial thyroid carcinoma tissue sections (4μm thickness) were deparaffinized and hydrated in
xylene and graded alcohol series as described earlier by some of us.109 The slides were treated
with 0.3% H2O2 at room temperature for 30 minutes to block the endogenous peroxidase activity.
After blocking the non-specific binding with normal horse or goat serum, the sections were
incubated with anti-human antibodies - nucleolin mouse monoclonal antibody (1:200 dilution);
or PTMA mouse monoclonal antibody (1:700 dilution, Lifespan Biosciences, Seattle, WA) for
30 minutes and subsequently with a biotinylated secondary antibody (horse anti-mouse or goat
anti-rabbit) for 30 minutes. The sections were finally incubated with VECTASTAIN Elite ABC
Reagent (Vector labs, Burlingame, CA) and diaminobenzidine was used as the chromogen.
Evaluation of Immunohistochemistry

29
The immunostaining scoring was based on percentage positivity and staining intensity. Sections
were scored as positive if epithelial cells showed immunoreactivity in the plasma membrane,
cytoplasm, and/or nucleus when observed by an evaluator who was blinded to the clinical history
and outcome. Percentage positive scores were assigned according to the following scale: 0, <
10% cells; 1, 10-30% cells; 2, 30-50% cells; 3, 50-70% cells; and 4, >70%. Staining intensity
was then also scored semi-quantitatively as follows: 0, none; 1, mild; 2, moderate; and 3, intense.
A total score was then obtained (ranging from 0 to 7) by adding the percentage positivity scores
and intensity scores for each section. Scatter plots were used to determine the distribution of
total score of membranous, nuclear, and cytoplasmic nucleolin and PTMA in thyroid
carcinomas.
Xenografts of Thyroid Carcinoma TPC-1 Cell Line in NOD/SCID/γ mice
NOD/SCID/ γ (c)(null) mice, (SCID) mutation and interleukin-2Rgamma (IL-2Rgamma) allelic
mutation (gamma(c)(null)), were originally generated by 8 backcross matings of C57BL/6j-
gamma(c)(null) mice and NOD/Shi-scid mice.110 The breeding colony of these mice is
maintained by the Ontario Institute of Cancer Research for use of its researchers in the
University Health Network Max Bell Animal Facility, Toronto, Canada. Tumor xenografts of
thyroid carcinoma cell line TPC-1 were established in these immunocompromised mice to
evaluate the in vivo expression profiles of PTMA and nucleolin. One million TPC-1 cells in
matrigel were implanted subcutaneously on the flanks of the mice and the animals were
monitored for 4-6 months. The tumors appeared within 4-6 weeks; mice were sacrificed after
10-24 weeks, tumors were excised, fixed in formalin and embedded in paraffin. Tissue sections
(5 µm) were cut, stained with hematoxylin and eosin and reviewed by the pathologist. Serial
sections were used for immunohistochemical analysis of PTMA and nucleolin as described
above.

30
Chapter 4 Results
4 Results 4.1 STR Profile of Cell Lines Table 5. STR profile analysis of cell lines used in this study. Amelogenin D3S1358 D8S1179 D21S11 D18S51 D5S818 D13S317 FGA
TPC-1 X 16 17 11 17 30 31.2 13 16 8 10 11 12 20 21
CAL62 X 16 13 32.2 16 9 12 12 19
C643 X Y 15 11 13 28 14 18 11 12 8 10 18 21
BCPAP X 16 17 12 13 30 31 13 17 10 11 12 20 23
SW1736 X 16 17 13 14 29 31 14 12 13 11 12 22
To ensure the problem of cross-contamination and misidentification of cell lines was avoided,
STR profiles of each cell line were determined to match those of the original thyroid-derived cell
lines as reported in previous studies by Schweppe et al101 and/or the known STR profiles
reported in American Type Culture Collection (ATCC) or German Collection of Microorganisms
and Cell Cultures (DSMZ). Detailed STR profile results for all cell lines used in this study are
presented in Table 5 for select loci that were examined. All loci for all cell lines were found to
match the STR profiles reported in the ATCC and DSMZ database with one exception. As was
also reported in the Schweppe study, the SW1736 cell line was a match at all loci examined with
except for a minor difference at the D3S1538 locus. 101 The Schweppe et al study also found the
cell line continues to express thyroid-specific genes (Pax-8, TIF-1) and possesses the V600E
BRAF mutation at one allele.
4.2 Proteins Identified in Thyroid Carcinoma Cell Lines The conditioned media of the thyroid carcinoma cell lines TPC-1 and CAL62 were analyzed by
one-dimensional LC-MS/MS twice. The two blood proteins albumin and globulin were
identified in the 0 h controls and in the test samples and were excluded from the list of identified
proteins. The proteins that were identified based on one peptide were excluded from further
analysis. Proteins identified with at least two peptides with ≥95% confidence were considered as
high-confidence identifications (Appendix I). In addition, PTMA was also considered as its
identification was based on a 99% confidence peptide (Figure 2) and has previously been

31
reported to have an important role in human cancers, including earlier studies by our group.109,
111-114
Figure 3. Mass spectrum of prothymosin-alpha (PTMA). MS/MS spectrum of the detected peptide for identification of PTMA by liquid chromatography-tandem mass-spectrometry. Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.
After applying the high-confidence threshold to the identified protein list, 46 proteins remained
as candidates for further analysis (Table 6). Eighty percent of the high-confidence proteins were
identified in at least two of the four separate analyses. Literature searches revealed that 31 of the
high confidence proteins have not yet been reported in thyroid carcinoma. A comparative
analysis between these 46 high-confidence proteins and their identification in other cancer
secretome datasets is available in the appendix (Appendix II). In both cell lines, investigation
into the reported localization of the identified proteins using Ingenuity Pathway Analysis
revealed that membrane and extracellular proteins were predominantly detected (Figure 3).
Similarly, reported functions of the identified proteins suggest many are involved in metabolic
processes and signal transduction pathways (Figure 4). It is important to note that this
information is based on models generated by IPA on the basis of updated database knowledge
and has not been experimentally verified. It should be considered with caution, but provides
useful information about the potential role of the proteins identified by secretome analysis. In
total, 17 proteins were identified in the conditioned media from both cell lines, 18 were found
only in TPC-1, and 11 proteins only in CAL62 (Table 6). The majority (40/46) of these high-
confidence identifications were deemed secretory according to their SignalP and SecretomeP
scores. Six of these 46 high-confidence protein identifications were considered for further
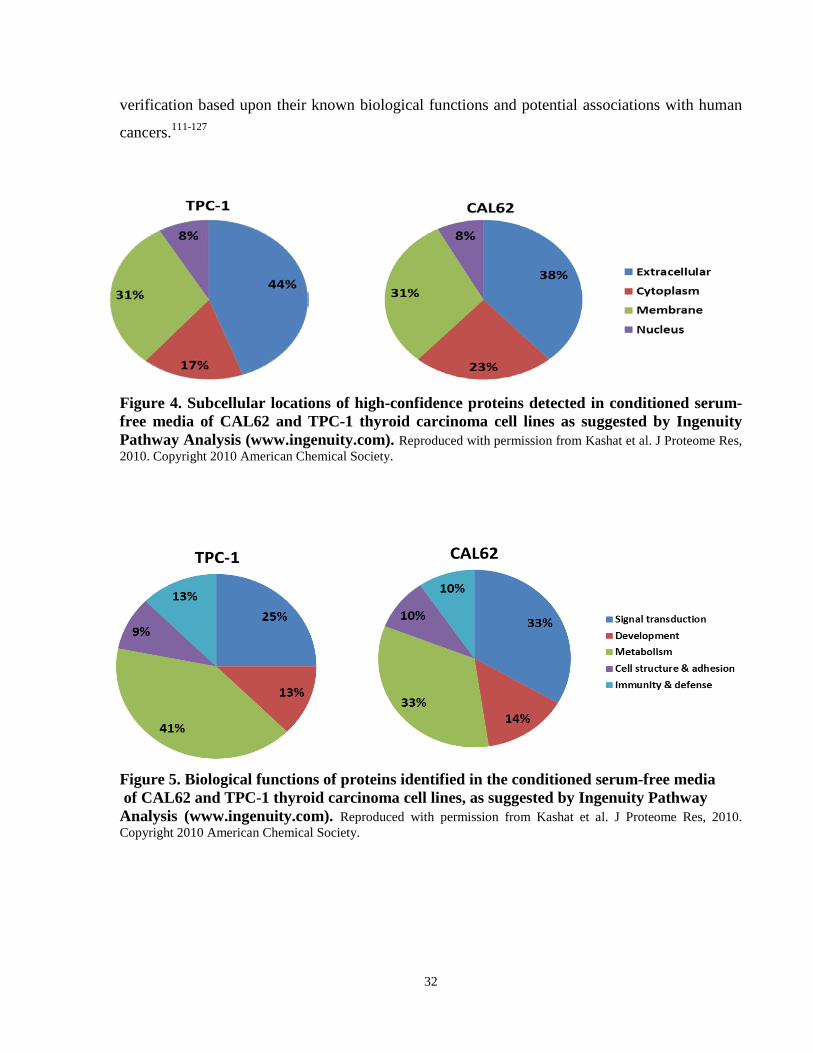
32
verification based upon their known biological functions and potential associations with human
cancers.111-127
Figure 4. Subcellular locations of high-confidence proteins detected in conditioned serum-free media of CAL62 and TPC-1 thyroid carcinoma cell lines as suggested by Ingenuity Pathway Analysis (www.ingenuity.com). Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society. Figure 5. Biological functions of proteins identified in the conditioned serum-free media of CAL62 and TPC-1 thyroid carcinoma cell lines, as suggested by Ingenuity Pathway Analysis (www.ingenuity.com). Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.
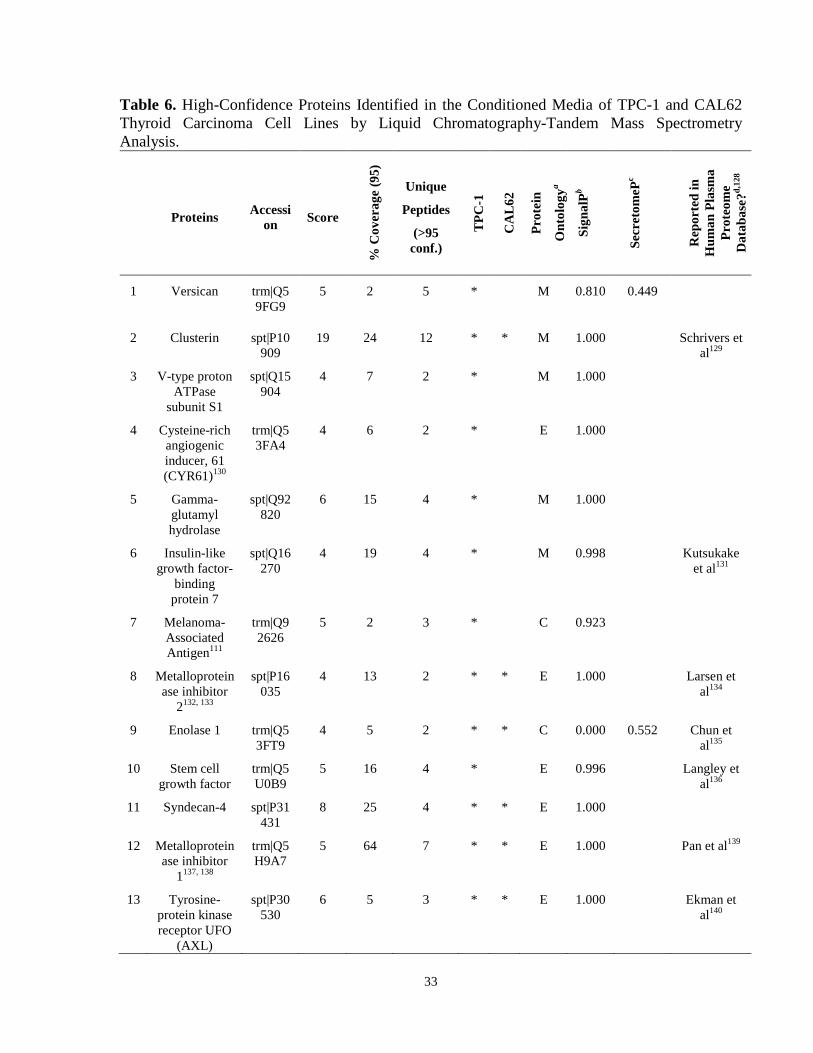
33
Table 6. High-Confidence Proteins Identified in the Conditioned Media of TPC-1 and CAL62 Thyroid Carcinoma Cell Lines by Liquid Chromatography-Tandem Mass Spectrometry Analysis.
Proteins Accession Score
% C
over
age
(95)
Unique
Peptides
(>95 conf.)
TPC
-1
CA
L62
Prot
ein
Ont
olog
ya
Sign
alPb
Secr
etom
ePc
Rep
orte
d in
H
uman
Pla
sma
Prot
eom
e D
atab
ase?
d,12
8
1 Versican trm|Q59FG9
5 2 5 *
M 0.810 0.449
2 Clusterin spt|P10909
19 24 12 * * M 1.000 Schrivers et al129
3 V-type proton ATPase
subunit S1
spt|Q15904
4 7 2 *
M 1.000
4 Cysteine-rich angiogenic inducer, 61 (CYR61)130
trm|Q53FA4
4 6 2 * E 1.000
5 Gamma-glutamyl hydrolase
spt|Q92820
6 15 4 * M 1.000
6 Insulin-like growth factor-
binding protein 7
spt|Q16270
4 19 4 * M 0.998 Kutsukake et al131
7 Melanoma-Associated Antigen111
trm|Q92626
5 2 3 * C 0.923
8 Metalloproteinase inhibitor
2132, 133
spt|P16035
4 13 2 * * E 1.000 Larsen et al134
9 Enolase 1 trm|Q53FT9
4 5 2 * * C 0.000 0.552 Chun et al135
10 Stem cell growth factor
trm|Q5U0B9
5 16 4 * E 0.996 Langley et al136
11 Syndecan-4 spt|P31431
8 25 4 * * E 1.000
12 Metalloproteinase inhibitor
1137, 138
trm|Q5H9A7
5 64 7 * * E 1.000 Pan et al139
13 Tyrosine-protein kinase receptor UFO
(AXL)
spt|P30530
6 5 3 * * E 1.000 Ekman et al140
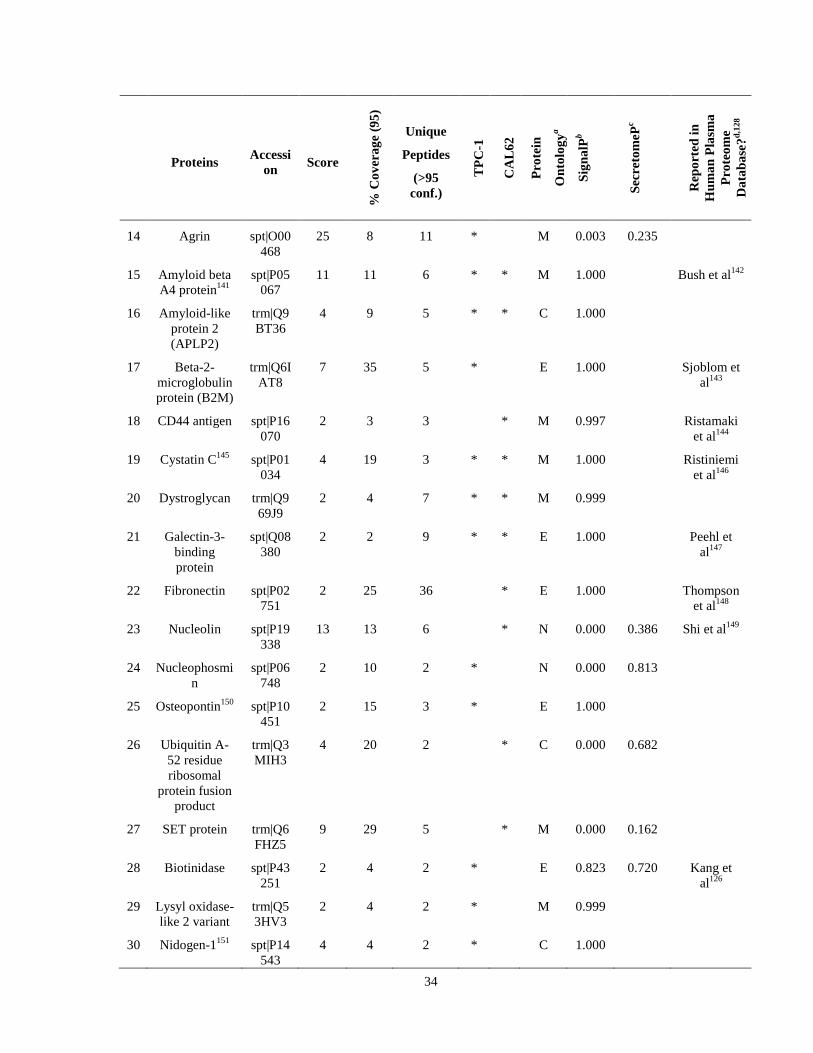
34
Proteins Accession Score
% C
over
age
(95)
Unique
Peptides
(>95 conf.)
TPC
-1
CA
L62
Prot
ein
Ont
olog
ya
Sign
alPb
Secr
etom
ePc
Rep
orte
d in
H
uman
Pla
sma
Prot
eom
e D
atab
ase?
d,12
8
14 Agrin spt|O00468
25 8 11 * M 0.003 0.235
15 Amyloid beta A4 protein141
spt|P05067
11 11 6 * * M 1.000 Bush et al142
16 Amyloid-like protein 2 (APLP2)
trm|Q9BT36
4 9 5 * * C 1.000
17 Beta-2-microglobulin protein (B2M)
trm|Q6IAT8
7 35 5 * E 1.000 Sjoblom et al143
18 CD44 antigen spt|P16070
2 3 3 * M 0.997 Ristamaki et al144
19 Cystatin C145 spt|P01034
4 19 3 * * M 1.000 Ristiniemi et al146
20 Dystroglycan trm|Q969J9
2 4 7 * * M 0.999
21 Galectin-3-binding protein
spt|Q08380
2 2 9 * * E 1.000 Peehl et al147
22 Fibronectin spt|P02751
2 25 36 * E 1.000 Thompson et al148
23 Nucleolin spt|P19338
13 13 6 * N 0.000 0.386 Shi et al149
24 Nucleophosmin
spt|P06748
2 10 2 * N 0.000 0.813
25 Osteopontin150 spt|P10451
2 15 3 * E 1.000
26 Ubiquitin A-52 residue ribosomal
protein fusion product
trm|Q3MIH3
4 20 2 * C 0.000 0.682
27 SET protein trm|Q6FHZ5
9 29 5 * M 0.000 0.162
28 Biotinidase spt|P43251
2 4 2 * E 0.823 0.720 Kang et al126
29 Lysyl oxidase-like 2 variant
trm|Q53HV3
2 4 2 * M 0.999
30 Nidogen-1151 spt|P14543
4 4 2 * C 1.000

35
Proteins Accession Score
% C
over
age
(95)
Unique
Peptides
(>95 conf.)
TPC
-1
CA
L62
Prot
ein
Ont
olog
ya
Sign
alPb
Secr
etom
ePc
Rep
orte
d in
H
uman
Pla
sma
Prot
eom
e D
atab
ase?
d,12
8
31 Nucleobindin 1
trm|Q53GX6
4 5 2 * E 1.000
32 Plasminogen activator,
urokinase18, 19
trm|Q5SWW9
15 23 9 * * E 0.999 Miyake et al152
33 Dickkopf-related protein
3 (DKK-3)
trm|Q6PQ81
4 7 2 * * E 1.000
34 Thrombospondin 1153
trm|Q59E99
34 14 15 * E 0.994 Liu et al154
35 Calsyntenin-1 trm|Q5UE58
11 8 6 * * M 1.000
36 Basement Membrane
Specific Heparan
Sulfate Core Protein
spt|P98160
4 1 2 * E 1.000
37 Prothymosin-alpha
(PTMA)d,112
trm|Q9NYD3
2 10 1 * N 0.000 0.701
38 Cadherin-2 (N-Cadherin)
spt|P19022
3 3 2 * M 0.999
39 Granulins (proepithelin)
spt|P28799
4 5 2 * E 0.999
40 Activated leukocyte cell
adhesion molecule
(ALCAM)
trm|Q1HGM9
2 4 2 * * M 0.985 Vaisocherova et al155
41 Peptidylproyl isomerase A (cyclophilin
A)
trm|Q3KQW3
5 12 2 * C 0.001 0.339 Tegeder et al156
42 Vimentin22, 23 spt|P08670
2 5 2 * * C 0.015 0.512 Sun et al157
43 Cathepsin Z trm|Q5U000
3 7 2 * O 1.000
44 Superoxide dismutase158
trm|Q6NR85
4 17 2 * * C 0.001 0.648 Spranger et al159
45 Putative trm|Q8 7 11 3 * C 0.000 0.494

36
Proteins Accession Score
% C
over
age
(95)
Unique
Peptides
(>95 conf.)
TPC
-1
CA
L62
Prot
ein
Ont
olog
ya
Sign
alPb
Secr
etom
ePc
Rep
orte
d in
H
uman
Pla
sma
Prot
eom
e D
atab
ase?
d,12
8
uncharacterized protein
WVW5
46 Insulin-like growth factor-
binding protein 6
(IGFBP-6)160
spt|P24592
2 27 5 * E 1.000 Baxter et al161
Table reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society. C – cytoplasm, E – extracellular, M – plasma membrane, N – nucleus, O – other, U – unknown. a The ontologies of identified proteins were analyzed using IPA and GoMiner. b The signal peptides were predicted using the hidden Markov model of SignalP 3.0 (protein with SignalP probability ≥0.900 is considered secretory). cThe nonclassical secretion of proteins was evaluated by the neural network output score of SecretomeP 2.0 (protein with SecretomeP probability ≥0.500 is considered secretory). dProteins were searched in the Plasma Blood Proteome database to determine previous reports of detection in human sera (http://www.plasmaproteomedatabase.org/) ePTMA was identified from using 1 unique of 99% confidence (sequence: EVVEEAENGR, detected twice). Its observed precursor mass was 1130 Da, charge +2. If protein has been reported in thyroid carcinoma blood and/or tissue, reference is cited next to protein name.
4.3 Clinical Verification of Identified Proteins As part of our initial analysis of the TPC-1 and CAL62 secretome, we have selected six of the 46
high-confidence proteins for further verification based upon their known biological functions and
potential associations with human cancers.111-127 These proteins are nucleolin, prothymosin-
alpha (PTMA), biotinidase, cysteine-rich angiogenic inducer 61 variant (CYR61), clusterin, and
enolase 1. Although PTMA was identified by only one unique peptide (Figure 2), it was
considered for further verification because it has previously been reported to have an important
role in human cancers, including earlier studies by our group.109, 111-114
Fluorescence microscopy of nucleolin and PTMA in TPC-1 and CAL62 human thyroid
carcinoma cells
The subcellular localization of PTMA and nucleolin was determined in TPC-1 (Figure 5A) and
CAL62 (Figure 5B) thyroid carcinoma cells. PTMA was detected in the cytoplasm and nuclei of
both cell lines, while nucleolin was detected in the nucleoli of TPC-1 and CAL62 cells.

37
Xenografts of TPC-1 thyroid carcinoma cells in NOD/SCID/γ mice exhibit protein
expression pattern similar to cultured TPC-1 thyroid carcinoma cells
Expressions of PTMA and nucleolin were determined in TPC-1 human-mouse xenografts
(Figure 6). PTMA was detected in the nucleus and cytoplasm of tumor cells, while nucleolin
expression was mainly nuclear, confirming similar pattern of expression of these proteins in
cultured thyroid carcinoma cells and tumor xenografts.
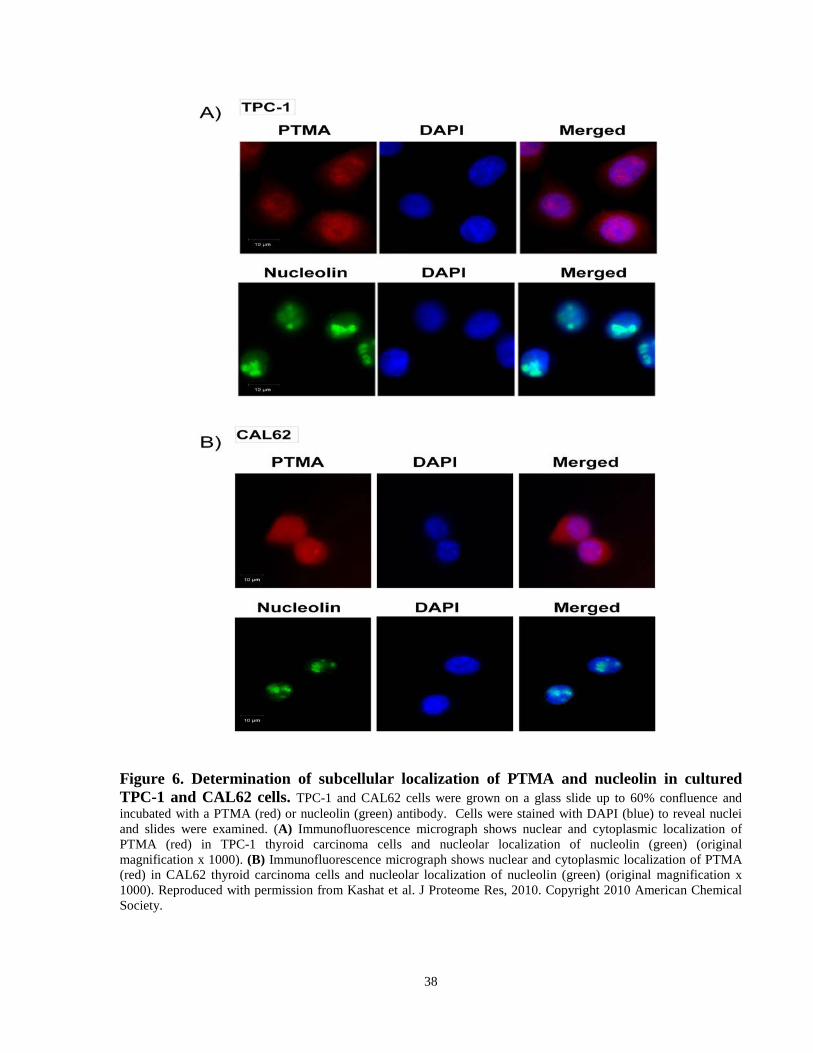
38
Figure 6. Determination of subcellular localization of PTMA and nucleolin in cultured TPC-1 and CAL62 cells. TPC-1 and CAL62 cells were grown on a glass slide up to 60% confluence and incubated with a PTMA (red) or nucleolin (green) antibody. Cells were stained with DAPI (blue) to reveal nuclei and slides were examined. (A) Immunofluorescence micrograph shows nuclear and cytoplasmic localization of PTMA (red) in TPC-1 thyroid carcinoma cells and nucleolar localization of nucleolin (green) (original magnification x 1000). (B) Immunofluorescence micrograph shows nuclear and cytoplasmic localization of PTMA (red) in CAL62 thyroid carcinoma cells and nucleolar localization of nucleolin (green) (original magnification x 1000). Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.
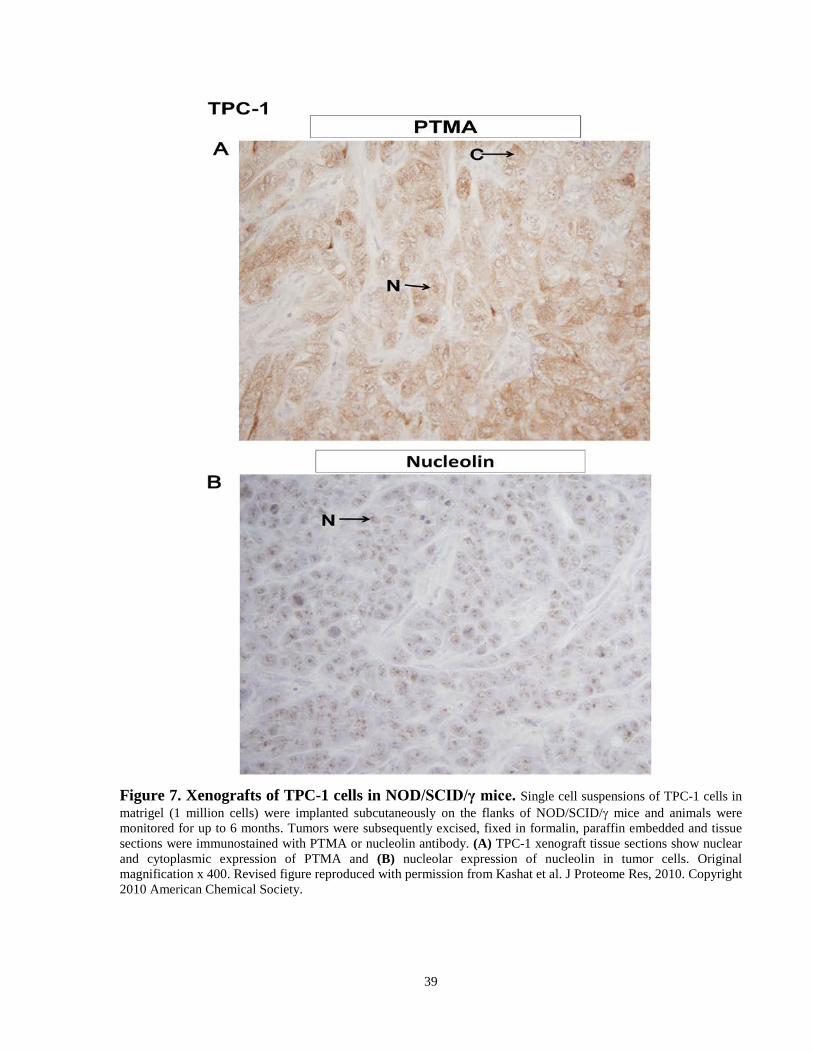
39
Figure 7. Xenografts of TPC-1 cells in NOD/SCID/γ mice. Single cell suspensions of TPC-1 cells in matrigel (1 million cells) were implanted subcutaneously on the flanks of NOD/SCID/γ mice and animals were monitored for up to 6 months. Tumors were subsequently excised, fixed in formalin, paraffin embedded and tissue sections were immunostained with PTMA or nucleolin antibody. (A) TPC-1 xenograft tissue sections show nuclear and cytoplasmic expression of PTMA and (B) nucleolar expression of nucleolin in tumor cells. Original magnification x 400. Revised figure reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.

40
Detection of Secretome Proteins in Human Sera, TPC-1 and CAL62 Thyroid Carcinoma
Cells and their Conditioned Media by Western Blotting
We analyzed 17 sera from thyroid carcinoma patients and 9 cancer-free individuals to determine
if a panel of six proteins, nucleolin, PTMA, clusterin, CYR61, biotinidase and enolase 1, could
be detected in circulation of thyroid carcinoma patients (Figure 7 and Figure 8). In addition, as
proof of principle, we independently verified the detection of these proteins in the whole cell
lysates and conditioned media of TPC-1 and CAL62 thyroid carcinoma cells (Figure 8). These
proteins have all been reported to have possible functions in thyroid and / or other cancers.
Western blotting confirmed all these proteins to be present in TPC-1 and CAL62 cell lysates
(Figure 8). Additionally, all proteins were confirmed in the conditioned media of TPC-1 and/or
CAL62 cells, in accordance with their detection by LC-MS/MS (Figure 8). Notably, all six
proteins were detected in the thyroid carcinoma patient’s sera (Figure 7, Figure 8). In agreement
with our immunohistochemical observations, PTMA appeared to increase in thyroid carcinoma
patients’ sera compared to normal individuals (Figure 7B). Interestingly, clusterin, enolase 1, and
biotinidase appeared to decrease in the sera of many thyroid carcinoma patients compared to the
normal sera (Figure 7). A negative control using goat anti-rabbit secondary antibody failed to
detect any immunoreactive proteins. A 62 kDa protein was detected using a goat anti-mouse
secondary antibody, but at considerably lower levels than the bands detected when mouse
primary antibodies were used.

41
Figure 8. Immunodetection of identified proteins in sera of thyroid carcinoma patients. Protein samples were prepared from the sera (50 μg) of cancer-free individuals and the sera (50 μg) of thyroid carcinoma patients with non-metastatic PTC or metastatic ATC, FTC, or PTC (meta). (A)Western blot analysis confirms the detection of biotinidase, clusterin, and CYR 61 in the sera of thyroid carcinoma patients. A negative control using goat anti-rabbit secondary antibody failed to detect any immunoreactive proteins. (B) Enolase 1, nucleolin, and PTMA were all detected in thyroid carcinoma patient sera. PTMA appears to increase in the sera of thyroid carcinoma patients compared to the sera from cancer-free individuals, consistent with our observations from immunohistochemical analysis of thyroid carcinoma tissues. A faint 62 kDa protein was detected using a goat anti-mouse secondary antibody alone, but at considerably lower levels than the bands detected when mouse primary antibodies were also used. Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.

42
Figure 9. Immunodetection of identified proteins in sera of thyroid carcinoma patients, and in the whole-cell lysates and conditioned-free media of TPC-1 and CAL62 cells. Protein samples were prepared from TPC-1 cells (50-75 μg) and their conditioned serum-free media (CM, 5 μg), and the sera (50 μg) of five thyroid carcinoma patients and one normal sera sample (normal, S-2145, Sigma). As protein loading controls, lysates were examined for actin expression, while conditioned media (CM) and patient sera were stained with Ponceau S (Sigma). Western blot analysis confirms the detection of these six proteins, namely biotinidase, clusterin, CYR 61, enolase 1, nucleolin and prothymosin-a in the thyroid carcinoma patient sera and in the whole-cell lysate and conditioned media of the thyroid carcinoma cell lines. Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.

43
Immunohistochemical analysis of PTMA and nucleolin in human thyroid carcinoma tissues
The expression of nuclear PTMA was observed to increase in ATC compared to normal thyroid
tissues adjacent to benign thyroid disease, papillary, and poorly differentiated (insular)
carcinomas (Figure 9, Figure 12). Notably, ATC displayed markedly increased cytoplasmic
PTMA compared to normal thyroid tissues, papillary, and poorly differentiated (insular)
carcinomas. Increased cytoplasmic expression of PTMA was also observed in poorly
differentiated (insular) carcinomas (Figure 9, Figure 12). Representative photomicrographs of
negative and positive controls are shown in Figure 11A.
Nuclear nucleolin expression was observed in all the thyroid carcinoma subtypes and normal
thyroid tissues adjacent to benign thyroid disease examined (Figure 10, Figure 12). Cytoplasmic
staining of nucleolin was also observed in some of the ATC cases. Interestingly, while most
regions of ATC tissues showed minimal cytoplasmic nucleolin expression, some areas exhibited
strong cytoplasmic staining, occasionally accompanying a loss of nuclear nucleolin expression
(Figure 12); this was only observed in the ATCs. Representative photomicrographs of negative
and positive controls are shown in Figure 11B.
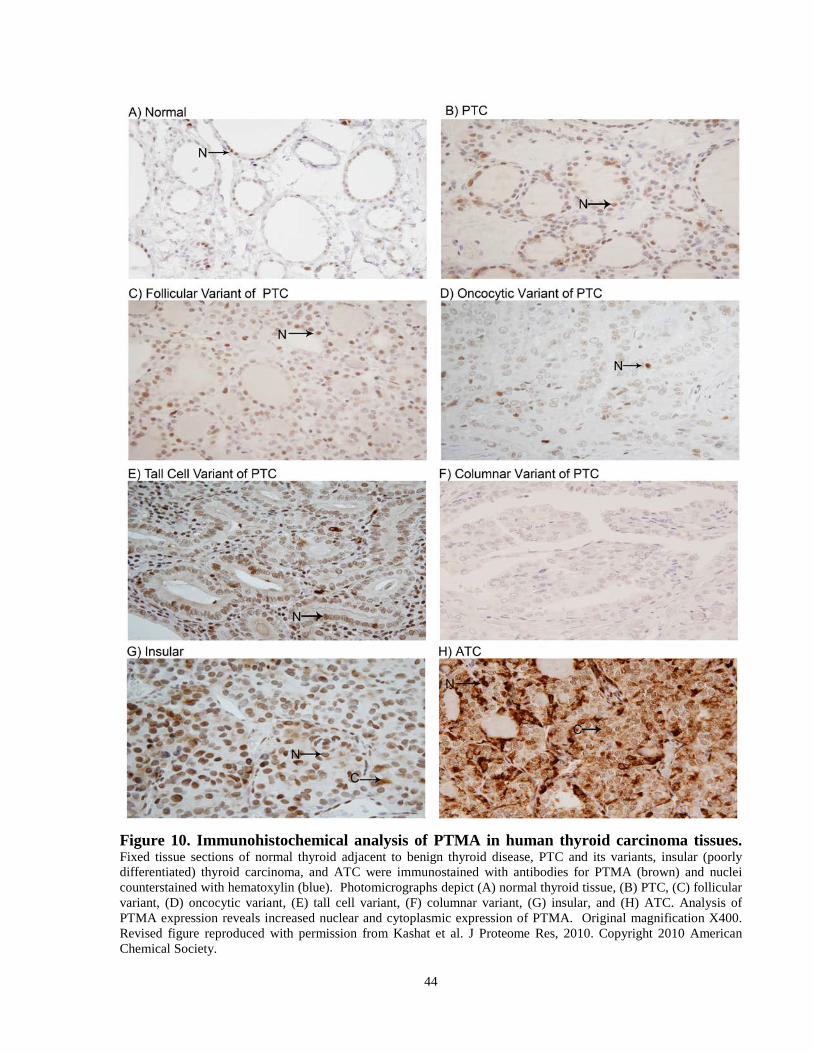
44
Figure 10. Immunohistochemical analysis of PTMA in human thyroid carcinoma tissues. Fixed tissue sections of normal thyroid adjacent to benign thyroid disease, PTC and its variants, insular (poorly differentiated) thyroid carcinoma, and ATC were immunostained with antibodies for PTMA (brown) and nuclei counterstained with hematoxylin (blue). Photomicrographs depict (A) normal thyroid tissue, (B) PTC, (C) follicular variant, (D) oncocytic variant, (E) tall cell variant, (F) columnar variant, (G) insular, and (H) ATC. Analysis of PTMA expression reveals increased nuclear and cytoplasmic expression of PTMA. Original magnification X400. Revised figure reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.

45
Figure 11. Immunohistochemical analysis of nucleolin in human thyroid carcinoma tissues. Fixed tissue sections of normal thyroid adjacent to benign thyroid disease, PTC and its variants, insular (poorly differentiated) thyroid carcinoma, and ATC were immunostained with antibodies for nucleolin (brown) and nuclei counterstained with hematoxylin (blue). Photomicrographs depict (A) normal thyroid tissue, (B) PTC, (C) follicular variant, (D) oncocytic variant, (E) tall cell variant, (F) columnar variant, (G) insular, and (H) ATC. Similar nuclear nuclear staining is seen among the various subtypes of thyroid carcinoma and normal thyroid tissues examined. Some ATC cases revealed cytoplasmic expression of nucleolin. Original magnification X400. Revised figure reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.
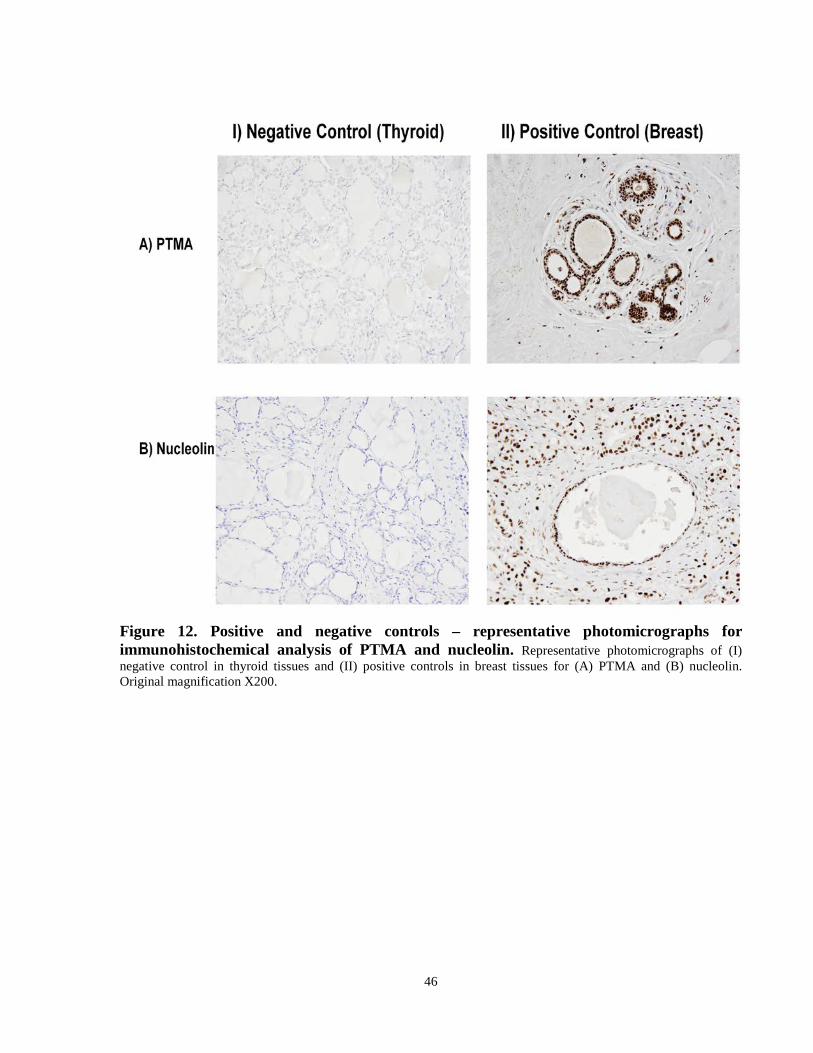
46
Figure 12. Positive and negative controls – representative photomicrographs for immunohistochemical analysis of PTMA and nucleolin. Representative photomicrographs of (I) negative control in thyroid tissues and (II) positive controls in breast tissues for (A) PTMA and (B) nucleolin. Original magnification X200.

47
Figure 13. Scatter plot Analysis of immunohistochemical scoring of PTMA and nucleolin. The PTMA and nucleolin immunohistochemically stained tissues were scored based on percentage positivity and immunostaining intensity. Sections were scored as positive if epithelial cells showed immunoreactivity in the plasma membrane, cytoplasm, and/or nucleus when observed by evaluators blinded to the clinical history and outcome. The percentage positive scores were assigned according to the following scale: 0, < 10% cells; 1, 10-30% cells; 2, 30-50% cells; 3, 50-70% cells; and 4, >70%. The intensity of the staining was also scored semi-quantitatively as follows: 0, no staining; 1, mild; 2, moderate; and 3, intense. The total score (0-7) was obtained by adding the percentage positivity scores and intensity scores for each section. (A) ATC displayed elevated nuclear expression of PTMA compared to normal thyroid adjacent to benign thyroid disease, insular, and papillary thyroid carcinomas. ATC cases displayed strikingly elevated cytoplasmic expression of PTMA compared to PTC and insular cells. PTC staining of cytoplasmic PTMA was low. Insular (poorly differentiated) thyroid carcinomas also demonstrated an increased expression of cytoplasmic PTMA compared to PTC and normal thyroid adjacent to benign thyroid disease. (B) Nuclear expression of nucleolin was seen in all thyroid carcinoma subtypes and normal thyroid adjacent to benign thyroid disease examined. Faint cytoplasmic expression was also observed in ATC cases only. Revised figure reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.

48
4.4 Expanded Proteomic Analysis of Thyroid Carcinoma Cell Lines
Based upon our experimental findings from the thyroid carcinoma secretomes of the cell lines
TPC-1 and BCPAP, we expanded our proteomic analysis to improve upon our list of candidate
biological markers for aggressive thyroid carcinomas. In our second proteomic analysis, we
examined two additional anaplastic-derived thyroid carcinoma cell lines (SW1736, C643) and
one papillary-derived (BCPAP). In these cell lines, a total of 83 proteins were identified from at
least 2 unique peptides with confidences ≥95% (Table 7). Of these proteins, 27 were previously
identified in CAL62 and/or TPC-1 (Table 7). Furthermore, an additional 4 proteins were
identified with a single high-confidence peptide each (≥95% confidence) and were previously
identified by at least 2 high-confidence peptides in TPC-1 and/or CAL62 (Table 7). Literature
searches of these 87 high-confidence proteins found that 61 have not been previously reported in
thyroid carcinoma tissues and/or blood samples. Of the 87 high-confidence proteins, 11 were
found in all three cell lines, 12 in SW1736/C643 only, 17 in SW1736/BCPAP only, while 2 were
unique to C643 and 45 were unique to SW1736. The proteins found only in SW1736 and C643
were 14-3-3 protein zeta, collagen alpha-1 (V) chain, collagen alpha 1 (XII) chain, filamin A,
fructose-bisphosphate aldolase, heterogeneous ribonuclear protein K, melanoma associated
antigen, nucleoside diphosphate kinase, phosphoglycerate kinase, PKM2, thrombospondin 1,
translation elongation factor 1 alpha 1-like 14 protein. The complete peptide summary for all
high confidence identifications is available in the appendix (Appendix III). The majority of
these proteins (62/87) were deemed to be secretory according to their SignalP or SecretomeP
scores. Although this is a lower percentage than in our first analysis, a comparative review of
the secretomes of 4 other cancers found nearly all proteins in our study have been reported to be
secreted in at least one of the other studies examined (Appendix IV).
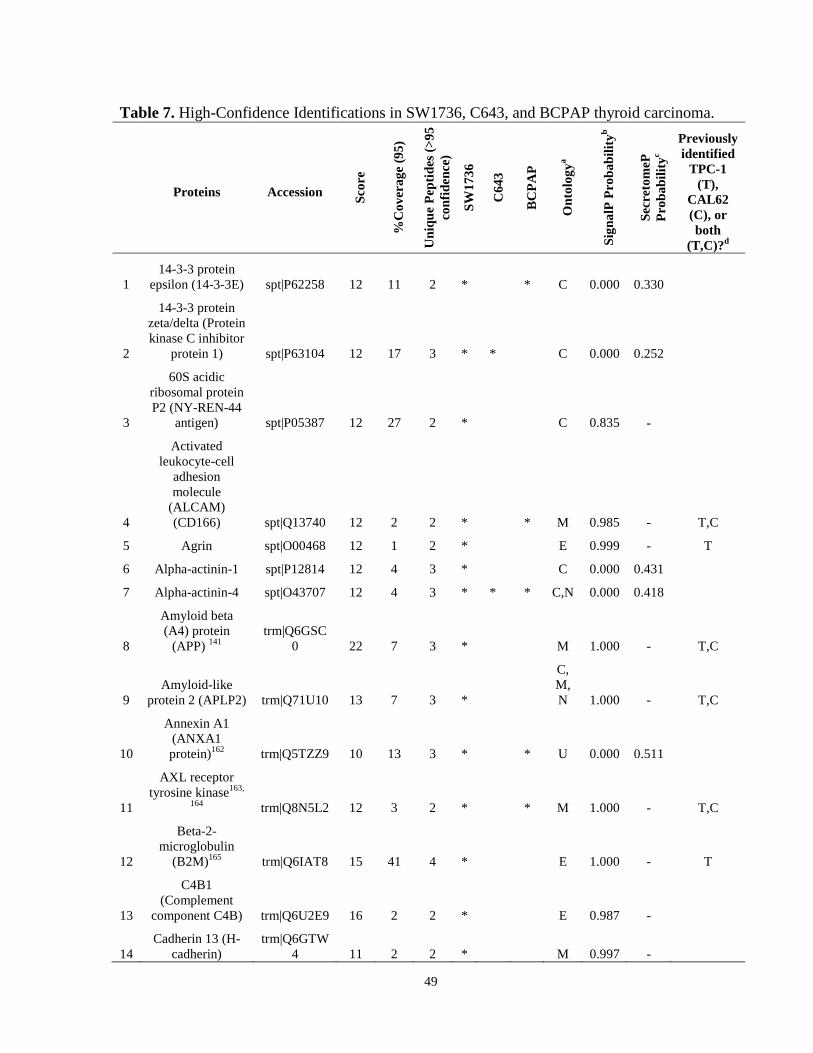
49
Table 7. High-Confidence Identifications in SW1736, C643, and BCPAP thyroid carcinoma.
Proteins Accession Scor
e
%C
over
age
(95)
Uni
que
Pept
ides
(>95
co
nfid
ence
)
SW17
36
C64
3
BC
PAP
Ont
olog
ya
Sign
alP
Prob
abili
tyb
Secr
etom
eP
Prob
abili
tyc
Previously identified
TPC-1 (T),
CAL62 (C), or both
(T,C)?d
1 14-3-3 protein
epsilon (14-3-3E) spt|P62258 12 11 2 * * C 0.000 0.330
2
14-3-3 protein zeta/delta (Protein kinase C inhibitor
protein 1) spt|P63104 12 17 3 * * C 0.000 0.252
3
60S acidic ribosomal protein P2 (NY-REN-44
antigen) spt|P05387 12 27 2 * C 0.835 -
4
Activated leukocyte-cell
adhesion molecule
(ALCAM) (CD166) spt|Q13740 12 2 2 * * M 0.985 - T,C
5 Agrin spt|O00468 12 1 2 * E 0.999 - T
6 Alpha-actinin-1 spt|P12814 12 4 3 * C 0.000 0.431
7 Alpha-actinin-4 spt|O43707 12 4 3 * * * C,N 0.000 0.418
8
Amyloid beta (A4) protein
(APP) 141 trm|Q6GSC
0 22 7 3 * M 1.000 - T,C
9 Amyloid-like
protein 2 (APLP2) trm|Q71U10 13 7 3 *
C, M, N 1.000 - T,C
10
Annexin A1 (ANXA1 protein)162 trm|Q5TZZ9 10 13 3 * * U 0.000 0.511
11
AXL receptor tyrosine kinase163,
164 trm|Q8N5L2 12 3 2 * * M 1.000 - T,C
12
Beta-2-microglobulin
(B2M)165 trm|Q6IAT8 15 41 4 * E 1.000 - T
13
C4B1 (Complement
component C4B) trm|Q6U2E9 16 2 2 * E 0.987 -
14 Cadherin 13 (H-
cadherin) trm|Q6GTW
4 11 2 2 * M 0.997 -
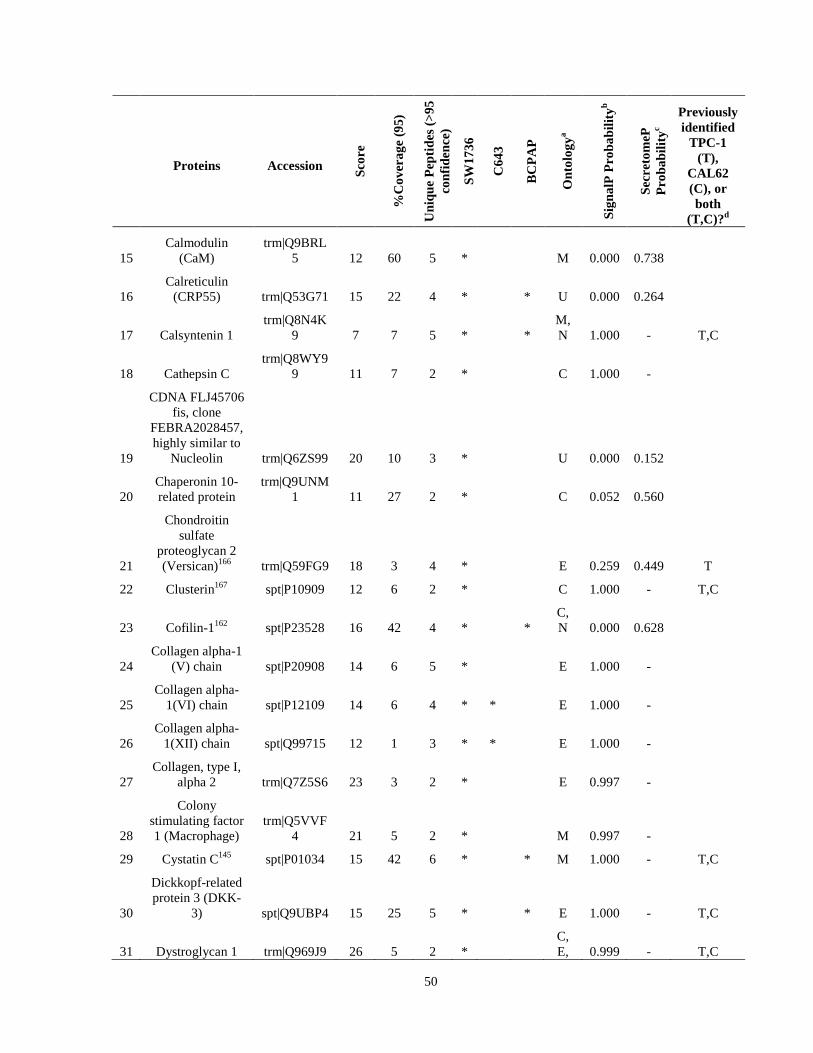
50
Proteins Accession Scor
e
%C
over
age
(95)
Uni
que
Pept
ides
(>95
co
nfid
ence
)
SW17
36
C64
3
BC
PAP
Ont
olog
ya
Sign
alP
Prob
abili
tyb
Secr
etom
eP
Prob
abili
tyc
Previously identified
TPC-1 (T),
CAL62 (C), or both
(T,C)?d
15 Calmodulin
(CaM) trm|Q9BRL
5 12 60 5 * M 0.000 0.738
16 Calreticulin
(CRP55) trm|Q53G71 15 22 4 * * U 0.000 0.264
17 Calsyntenin 1 trm|Q8N4K
9 7 7 5 * * M, N 1.000 - T,C
18 Cathepsin C trm|Q8WY9
9 11 7 2 * C 1.000 -
19
CDNA FLJ45706 fis, clone
FEBRA2028457, highly similar to
Nucleolin trm|Q6ZS99 20 10 3 * U 0.000 0.152
20 Chaperonin 10-related protein
trm|Q9UNM1 11 27 2 * C 0.052 0.560
21
Chondroitin sulfate
proteoglycan 2 (Versican)166 trm|Q59FG9 18 3 4 * E 0.259 0.449 T
22 Clusterin167 spt|P10909 12 6 2 * C 1.000 - T,C
23 Cofilin-1162 spt|P23528 16 42 4 * * C, N 0.000 0.628
24 Collagen alpha-1
(V) chain spt|P20908 14 6 5 * E 1.000 -
25 Collagen alpha-
1(VI) chain spt|P12109 14 6 4 * * E 1.000 -
26 Collagen alpha-
1(XII) chain spt|Q99715 12 1 3 * * E 1.000 -
27 Collagen, type I,
alpha 2 trm|Q7Z5S6 23 3 2 * E 0.997 -
28
Colony stimulating factor 1 (Macrophage)
trm|Q5VVF4 21 5 2 * M 0.997 -
29 Cystatin C145 spt|P01034 15 42 6 * * M 1.000 - T,C
30
Dickkopf-related protein 3 (DKK-
3) spt|Q9UBP4 15 25 5 * * E 1.000 - T,C
31 Dystroglycan 1 trm|Q969J9 26 5 2 * C, E, 0.999 - T,C
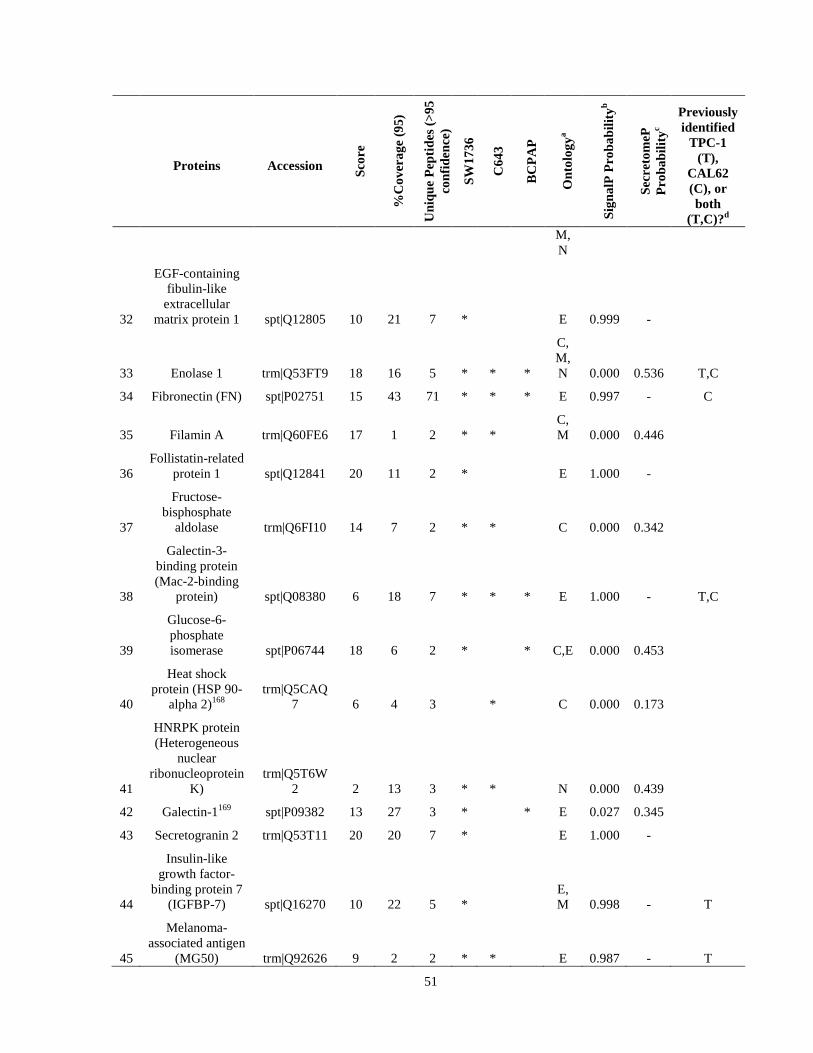
51
Proteins Accession Scor
e
%C
over
age
(95)
Uni
que
Pept
ides
(>95
co
nfid
ence
)
SW17
36
C64
3
BC
PAP
Ont
olog
ya
Sign
alP
Prob
abili
tyb
Secr
etom
eP
Prob
abili
tyc
Previously identified
TPC-1 (T),
CAL62 (C), or both
(T,C)?d M, N
32
EGF-containing fibulin-like
extracellular matrix protein 1 spt|Q12805 10 21 7 * E 0.999 -
33 Enolase 1 trm|Q53FT9 18 16 5 * * *
C, M, N 0.000 0.536 T,C
34 Fibronectin (FN) spt|P02751 15 43 71 * * * E 0.997 - C
35 Filamin A trm|Q60FE6 17 1 2 * * C, M 0.000 0.446
36 Follistatin-related
protein 1 spt|Q12841 20 11 2 * E 1.000 -
37
Fructose-bisphosphate
aldolase trm|Q6FI10 14 7 2 * * C 0.000 0.342
38
Galectin-3-binding protein (Mac-2-binding
protein) spt|Q08380 6 18 7 * * * E 1.000 - T,C
39
Glucose-6-phosphate isomerase spt|P06744 18 6 2 * * C,E 0.000 0.453
40
Heat shock protein (HSP 90-
alpha 2)168 trm|Q5CAQ
7 6 4 3 * C 0.000 0.173
41
HNRPK protein (Heterogeneous
nuclear ribonucleoprotein
K) trm|Q5T6W
2 2 13 3 * * N 0.000 0.439
42 Galectin-1169 spt|P09382 13 27 3 * * E 0.027 0.345
43 Secretogranin 2 trm|Q53T11 20 20 7 * E 1.000 -
44
Insulin-like growth factor-
binding protein 7 (IGFBP-7) spt|Q16270 10 22 5 *
E, M 0.998 - T
45
Melanoma-associated antigen
(MG50) trm|Q92626 9 2 2 * * E 0.987 - T

52
Proteins Accession Scor
e
%C
over
age
(95)
Uni
que
Pept
ides
(>95
co
nfid
ence
)
SW17
36
C64
3
BC
PAP
Ont
olog
ya
Sign
alP
Prob
abili
tyb
Secr
etom
eP
Prob
abili
tyc
Previously identified
TPC-1 (T),
CAL62 (C), or both
(T,C)?d (KIAA0230) 111
46
L-lactate dehydrogenase A
chain spt|P00338 16 18 4 * * C 0.000 0.549
47
Matrix metalloproteinase
1 (MMP-1) trm|Q5TZP0 17 33 8 * * * E 1.000 -
48
Matrix metalloproteinase
1 preprotein variant trm|Q53G75 2 15 2 * * E 0.000 0.442
49
Matrix metalloproteinase-
2 (MMP-2)170 spt|P08253 12 19 6 *
C, M, N 1.000 -
50
Metalloproteinase inhibitor 2
(TIMP-2)132, 133 spt|P16035 19 49 5 * * E 1.000 - T,C
51
Niemann-Pick disease, type C2
variant trm|Q53HV
6 18 26 2 * U 1.000 -
52
Nucleoside diphosphate
kinase (NME1-NME2) trm|Q32Q12 12 10 2 * * C,N 0.649 0.513
53 Nucleobindin 1 trm|Q53GX
6 16 16 5 * * E 1.000 - T
54 OAF homolog trm|Q86UD
1 18 15 3 * U 1.000 -
55 Periostin (PN)171 spt| Q15063 12 4 2 * E 0.999 -
56
Peptidylprolyl isomerase A
(Cyclophilin A) trm|Q3KQ
W3 14 25 3 * * * C 0.001 0.339 C
57 Phosphoglycerate
kinase trm|Q5J7W1 16 7 2 * * C 0.000 0.389
58 PKM2 protein trm|Q8WU
W7 3 7 2 * * C, N 0.000 0.385
59
Plasminogen activator inhibitor
1 (PAI-1)172 spt|P05121 4 4 2 * E 0.999 -
60 Tissue-type trm|Q6PJA5 17 12 4 * E 0.913 -

53
Proteins Accession Scor
e
%C
over
age
(95)
Uni
que
Pept
ides
(>95
co
nfid
ence
)
SW17
36
C64
3
BC
PAP
Ont
olog
ya
Sign
alP
Prob
abili
tyb
Secr
etom
eP
Prob
abili
tyc
Previously identified
TPC-1 (T),
CAL62 (C), or both
(T,C)?d plasminogen
activator (PLAT protein)
61 Protein CutA spt|O60888 11 23 2 * M 0.973 -
62 Protein FAM3C spt|Q92520 19 15 2 * M 0.906 -
63 Protein S100-A9 spt|P06702 9 23 2 *
C, E, M 0.000 0.425
64 Pyruvate kinase
isozymes M1/M2 trm|Q53GK
4 9 5 2 * C, N 0.089 0.420
65 Ribosomal protein
S27a spt|P62979 13 16 2 * C, N 0.000 0.879
66 SET protein trm|Q6FHZ
5 10 16 2 * C, N 0.000 0.162 C
67
SPARC (Secreted protein acidic and rich in cysteine) spt|P09486 13 87 19 * * *
E, M 1.000 -
68 Stem cell growth
factor (SCGF) trm|Q5U0B
9 19 13 3 * U 0.996 - T
69
Sulfhydryl oxidase 1
(Quiescin Q6) (hQSOX) spt|O00391 13 4 2 *
E, M 1.000 -
70
Superoxide dismutase [Cu-
Zn]158 trm|Q6NR8
5 12 48 3 * C 0.001 0.648 T,C
71 Tenascin C173 trm|Q5T7S3 12 16 17 * E 0.998 -
72 Thioredoxin
(TXN protein)174 spt|P10599 14 60 4 *
C, E, N 0.000 0.688
73 Thioredoxin reductase 174
trm|Q9UES8 13 7 2 *
C, N 0.002 0.359
74 Thrombospondin
1175, 176 trm|Q59E99 8 15 13 * * E 0.994 - T
75 Thrombospondin
2 trm|Q5RI52 8 2 2 * E 1.000 -
76
Tissue inhibitor of metalloproteinase 1 (TIMP-1)137 138 spt|P01033 10 82 11 * * E 1.000 - T,C
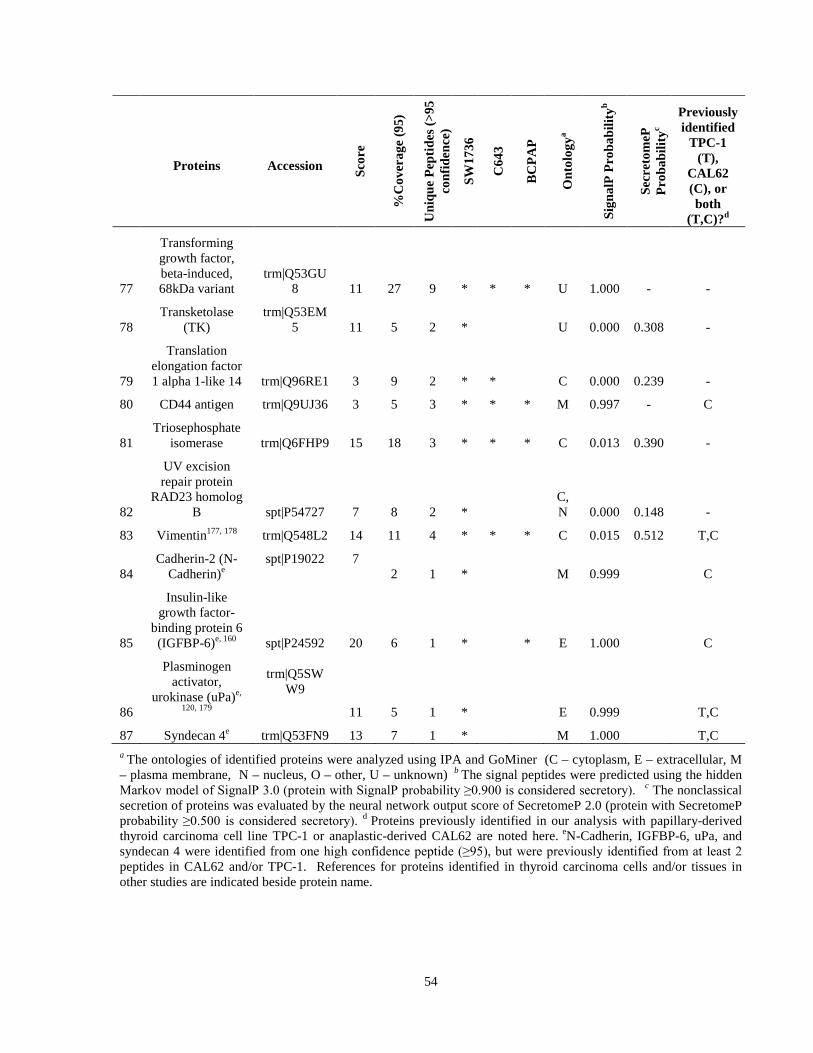
54
Proteins Accession Scor
e
%C
over
age
(95)
Uni
que
Pept
ides
(>95
co
nfid
ence
)
SW17
36
C64
3
BC
PAP
Ont
olog
ya
Sign
alP
Prob
abili
tyb
Secr
etom
eP
Prob
abili
tyc
Previously identified
TPC-1 (T),
CAL62 (C), or both
(T,C)?d
77
Transforming growth factor, beta-induced, 68kDa variant
trm|Q53GU8 11 27 9 * * * U 1.000 - -
78 Transketolase
(TK) trm|Q53EM
5 11 5 2 * U 0.000 0.308 -
79
Translation elongation factor 1 alpha 1-like 14 trm|Q96RE1 3 9 2 * * C 0.000 0.239 -
80 CD44 antigen trm|Q9UJ36 3 5 3 * * * M 0.997 - C
81 Triosephosphate
isomerase trm|Q6FHP9 15 18 3 * * * C 0.013 0.390 -
82
UV excision repair protein
RAD23 homolog B spt|P54727 7 8 2 *
C, N 0.000 0.148 -
83 Vimentin177, 178 trm|Q548L2 14 11 4 * * * C 0.015 0.512 T,C
84 Cadherin-2 (N-
Cadherin)e spt|P19022 7
2 1 * M 0.999 C
85
Insulin-like growth factor-
binding protein 6 (IGFBP-6)e, 160 spt|P24592 20 6 1 * * E 1.000 C
86
Plasminogen activator,
urokinase (uPa)e,
120, 179
trm|Q5SWW9
11 5 1 * E 0.999 T,C
87 Syndecan 4e trm|Q53FN9 13 7 1 * M 1.000 T,C a The ontologies of identified proteins were analyzed using IPA and GoMiner (C – cytoplasm, E – extracellular, M – plasma membrane, N – nucleus, O – other, U – unknown) b The signal peptides were predicted using the hidden Markov model of SignalP 3.0 (protein with SignalP probability ≥0.900 is considered secretory). c The nonclassical secretion of proteins was evaluated by the neural network output score of SecretomeP 2.0 (protein with SecretomeP probability ≥0.500 is considered secretory). d Proteins previously identified in our analysis with papillary-derived thyroid carcinoma cell line TPC-1 or anaplastic-derived CAL62 are noted here. eN-Cadherin, IGFBP-6, uPa, and syndecan 4 were identified from one high confidence peptide (≥95), but were previously identified from at least 2 peptides in CAL62 and/or TPC-1. References for proteins identified in thyroid carcinoma cells and/or tissues in other studies are indicated beside protein name.

55
Chapter 5 Discussion, Conclusion, and Future Directions
5.1 Discussion Herein we demonstrated the potential of secretome analysis of thyroid carcinoma cell lines to
identify secreted proteins that can be independently verified in cell lines, tumor xenografts,
human tumor tissues and blood samples of thyroid carcinoma patients. The majority of the 46
high-confidence proteins identified in our first proteomic analysis were deemed to be secretory
according to their SignalP and SecretomeP scores, lending support to our strategy of finding
secreted proteins using proteomic analysis of conditioned media of cultured thyroid carcinoma
cells. Literature searches conducted on these identified proteins revealed that 31 of them have
not yet been reported in thyroid carcinoma tissues and/or sera, demonstrating the ability of
secretome analysis to reveal novel potential candidates for consideration as biological markers in
the management of thyroid carcinoma. Of the remaining 15 proteins, 12 have been previously
detected in thyroid carcinoma tissues, namely, CYR61, melanoma-associated antigen, tyrosine-
protein kinase receptor UFO (AXL), amyloid-beta A4 (APP), osteopontin, plasminogen activator
urokinase, thrombospondin, PTMA, vimentin, superoxide dismutase, insulin-like growth factor
binding protein 6 (IGFBP6), and nidogen 1.111, 112, 120, 130, 141, 150, 151, 153, 158, 160, 177-180 Our
identification of these proteins in the thyroid carcinoma secretome suggests that they should be
explored as potential blood-based biomarkers due to their secretory potential.
Our study selected a total of six proteins for further verification and clinical validation based
upon their known biological functions and potential associations with cancer.111-127 Two of these
proteins were also examined in two thyroid carcinoma cell lines (papillary-derived TPC-1 and
anplastic-derived CAL62), xenografts of TPC-1 from immunocompromised (NOD/SCID/γ)
mice, and in 48 human thyroid tumors. A previous study examining PTMA in thyroid carcinoma
had suggested its mRNA levels are elevated in differentiated thyroid carcinomas, compared to
adenomas and goitres (P<0.05).112 PTMA is an interesting protein because mixed-expression of
the protein (nuclear and cytoplasmic) has been reported to be involved in the progression of
prostate, bladder, and head and neck carcinomas.108, 111, 113, 114 Nucleolin was selected for further
verification because it has been suggested to play an important role in cancer processes.
Recently, it has known to promote cellular proliferation and growth in vivo and in vitro and to
simultaneously interact with RAS and EGFR (ErbB1) in vivo.181, 182 Our detection of these

56
proteins in the cultured cells and their conditioned media confirmed these secretome proteins
originated from thyroid carcinoma cells. Until our study, the expression of these proteins in
human thyroid carcinoma tissues and detectability in the serum of thyroid carcinoma patients had
not been examined.
The subcellular localization of PTMA and nucleolin was similar in TPC-1 cells and their
xenografts in NOD/SCID/γ mice. The unchanged localization of these proteins in vitro and in
vivo suggests these cell lines retain some of their characteristics in vivo. Furthermore, the mixed
expression of PTMA (nuclear and cytoplasmic) in TPC-1 cells and their xenografts corresponded
with its expression in aggressive thyroid carcinomas and the reported expression found in
aggressive tumors of other cancer types (prostate, bladder, and upper urinary tract-transitional
cell).111, 113, 114 These findings, both in culture and in vivo, provide support to previous findings
suggesting TPC-1 may share properties of aggressive tumors.183 Our immunohistochemical
analysis of PTMA expression in thyroid carcinoma tissues suggests its nuclear and cytoplasmic
expression is elevated in ATC compared to normal thyroid, PTC and poorly differentiated
carcinomas, indicating it may serve as a marker for aggressive carcinomas upon validation in a
larger study and that changes in its expression may possibly be involved in the progression of
poorly and undifferentiated carcinomas. This is further supported by our observation that
cytoplasmic expression of PTMA was also increased in poorly differentiated carcinomas
compared to normal thyroid and PTC, which both showed low cytoplasmic PTMA staining.
These findings in thyroid carcinoma and other cancers, suggest that PTMA may play an
important role in the progression of tumors and its functions should be further examined. The
clinical significance of increased nuclear and cytoplasmic PTMA in thyroid tumor
aggressiveness warrants confirmation in a larger longitudinal study. Furthermore, PTMA was
detected and appeared elevated in sera of thyroid carcinoma patients compared to cancer-free
individuals, making it a potential serological and histological marker for thyroid carcinomas.
Our study was also the first to examine the immunohistochemical expression of nucleolin in
thyroid carcinoma. High expression of the protein was observed in tumor nuclei in all subtypes
of thyroid carcinomas and normal thyroid tissues. Importantly, cytoplasmic localization of
nucleolin was observed in ATCs only, suggesting cytoplasmic expression may be associated
with aggressiveness of these cancers. The significance of these findings remains to be
determined and warrants verification in future larger-scale studies. Nucleolin was found to

57
localize in the nucleoli of TPC-1, CAL62 and the xenografts of TPC-1 cells in NOD/SCID/γ
mice. Similar nucleolar localization of nucleolin has also been reported in breast cancer tissues
and cell lines.117 Herein our observations suggest it may be a marker for proliferation, but might
have limited utility as a biomarker for thyroid carcinoma due to the lack of differential
expression between normal thyroid tissues and the thyroid carcinoma subtypes examined.
In addition to these two proteins, we have also independently verified the expression of four
other secretome proteins in the sera of thyroid carcinoma patients, biotinidase, enolase 1, CYR61
and clusterin also based upon their reported implication in cancer.111-127 Our study’s
identification of these six proteins in the secretome of thyroid carcinoma cells and their
subsequent verification in thyroid carcinoma patients’ sera is an example of how analysis of
secretome proteins can identify candidate biomarkers for use in the creation of minimally
invasive blood-based diagnostic tests in future studies. The remaining proteins we have
examined are discussed here indepth and warrant analysis in large-scale study of thyroid
carcinoma patients’ sera to determine their potential as minimally-invasive thyroid carcinoma
markers. In addition to these six proteins with known roles in cancer, many other proteins have
been identified in our study whose role in cancer remains unclear and may now be investigated
for future mechanistic studies as well as exploration of therapeutic and diagnostic potential.
Enolase 1 appeared to decrease in thyroid carcinoma patients’ sera suggesting it may have
potential to develop into a biological marker upon futher verification in a larger cohort of
patients. Previously, it has been previously shown to be upregulated in male breast cancer tissue
of the infiltrating ductal carcinoma subtype.127 184 Herein we also report a potential decrease in
biotinidase levels in thyroid carcinoma patients’ sera. These findings are supported by a similar
decrease in biotinidase levels recently reported in sera and tumor tissues of breast cancer
patients.126 In this study biotinidase discriminated breast cancer patients from normal subjects
with 47.6% sensitivity and 90.5% specificity. These findings in a different cancer type supports
our observation of these proteins being secreted in the secretomes of other cancers and provides
further evidence that they may have potential applications beyond the field of thyroid oncology.
Of course, due to the limited number of samples examined in our study, the suitability of all
proteins we have examined as blood-based thyroid carcinoma biomarkers remains to be
determined and warrants further investigation.

58
CYR61, also known as CCN1, belongs to the CCN family of proteins, initially identified as
secretory proteins whose production is induced by oncogenes,185 and has been shown to promote
celllular proliferation, angiogenesis, and differentiation.186 Paradoxically, while having
demonstrated importance in cancer cell proliferation, it has also been shown to play an important
role in the induction of apotosis.187 It has previously been shown to be reduced to less than 50%
of its normal levels in PTC and has also been proposed to play an important role to the
proliferation of prostate cancer cells.123, 130 CYR61 has previously been found in thyroid
carcinoma tumor tissues, but herein we demonstrate the presence of CYR61 in sera of thyroid
carcinoma patients, suggesting the possibility of development of serum based immunoassays for
investigation of the diagnostic and prognostic potential of this protein in future studies.
Our identification of clusterin in the thyroid carcinoma secretome illustrates the powerful ability
of secretome analysis to guide researchers to proteins with critical importance in the
development and progression of cancer. Clusterin is a heterodimeric protein involved in
numerous cellular functions including lipid transport, complement inhibition, apoptosis, DNA
repair and cellular differentiation.118, 188 Its secreted form has been shown to promote cellular
survival and resistance to chemotherapy and radiation therapy.120, 188, 189 While the pathways
involved in clusterin action are still being elucidated, it has been suggested that clusterin serves
as a ubiquitin binding protein that enhances the activity of the transcription factor NF-kappaB by
increasing the degradation of I-kappaB.122 The overexpression of clusterin in tumors has also
been correlated with unfavourable survival, lymph-node metastasis, tumor invasion and TNM
stage in gastric cancers119 and impaired survival in ovarian cancer.121 Clusterin appeared to be
markedly decreased in the sera of thyroid carcinoma patients compared to the sera of cancer-free
individuals. These observations merit futher verification and should be quantified in a larger
cohort of patients. As clusterin has not yet been reported in thyroid carcinomas, its potential for
improving the diagnosis, management, and treatment of thyroid carcinomas should be further
examined. The finding that many of these proteins have important roles in a wide variety of
cancers is a testament to the notion that secretome analysis has the potential to identify proteins
critical to the progression and aggressive behaviour of certain cancers.
The detection of all six proteins in the sera of thyroid carcinoma patients supports the idea of
using secretome analysis to identify candidate blood-based biomarkers. It is important to be
cautious when interpreting western blot results of patient sera samples. The secondary antibodies

59
antibodies used may bind circulating heavy and light chains of IgG in patient sera (molecular
weights near 50 and 25 kda, respectively). Therefore, it is important to realize that bands
appearing at these sizes are possibly attributable to cross-reactivity with endogenous blood IgG
and may not be solely indicative of the protein of interest.190, 191 We used a secondary antibody
alone control on the patient sera to address these concerns. In our study, a faint band near to the
size of the heavy immunoglobulin chain (50 kDa) was observed when using a biotinylated goat
anti-mouse secondary antibody alone control, suggesting possible cross-reactivity with
endogenous IgG. We have, however, confirmed the blood-based detection of nucleolin, enolase,
biotinidase, and clusterin in the human plasma proteome database
(http://www.plasmaproteomedatabase.org/) providing support to our detection of these proteins
in thyroid carcinoma patient sera.126, 128, 129, 135 Furthermore, CYR61 was detected using a rabbit
polyclonal antibody and our goat anti-rabbit secondary antibody alone control did not show any
immunoreactivity with immunoglobulin light and heavy chains in patient sera. These
observations, in addition to our verification of the detection of all six proteins in the lysates and
conditioned media of TPC-1 and/or CAL62 cells and the independent verification of PTMA and
nucleolin in the cultured thyroid carcinoma cell lines, xenografts, and human thyroid carcinoma
tissues lends strong support for the detection of our panel of proteins in the sera of thyroid
carcinoma patients.
As already discussed, our results suggest although TPC-1 is a papillary-derived cell line, it
retains properties of aggressive thyroid tumors. We expanded proteomic analysis to include
additional papillary and anaplastic cell lines to aid in the identification of markers to distinguish
aggressive and non-aggressive thyroid carcinoma. The use of additional anaplastic (SW1736,
C643) and papillary-derived cell lines (BCPAP) allowed us to greatly improve upon the number
of potential biomarkers identified in our study. As we found in our initial analysis, a large
majority of the proteins identified from this secretome dataset have never been reported in
thyroid carcinoma before (61 of 87 high-confidence identifications). This continues to
demonstrate how secretome-based approaches may unravel novel targets for consideration.
Furthermore, our comparative analysis of both thyroid carcinoma secretome datasets again found
that nearly all of high-confidence proteins have been reported in at least one secretome datasets
of another cancer (breast, pancreatic, lung, and/or nasopharyngeal). This interesting observation
highlights the potential for many of these proteins to serve as biological markers in not only

60
thyroid, but other cancers as well. This work has now paved the way for future studies that may
lead to the characterization of the most promising markers of aggressive and non-aggressive
thyroid carcinomas.
An important limitation of our current methodology stems from the use of one-dimensional
liquid chromatography prior to mass-spectrometry. Our study had a lower amount of total
protein identifications compared to other recent studies in this field. This reduced number of
identifications is likely to improve with the future use of more sensitive proteomic technologies
such as iTRAQ-labelling and multidimensional LC-MS.94, 95, 192, 193 Nevertheless, our study
illustrates the ability of secretome analysis to generate new protein targets for diagnostic and/or
therapeutic consideration. Many of the proteins identified herein have been linked to other cancer
secretomes and the majority has not yet been reported in thyroid carcinoma. These proteins may
have biological implications in thyroid carcinoma, in addition to these other cancer types. Our
work verifying the expression and detection of some of these proteins in thyroid carcinoma
tissues, patient blood samples, thyroid carcinoma cell lines, and xenografts provides researchers
and clinicians with useful information about their potential clinical relevance. It is also important
to understand the limitations of the scoring system we have used for the immunohistochemical
analysis of PTMA and nucleolin in human tissues. The scoring scheme is only semi-quantitative.
The subjectivity of scoring is reduced by comparing the scores of two independent observers and
rescoring all slides with a third objective observer should a discrepancy of >2 occur for the final
score. Furthermore, it is also important to be aware that the intensity of the staining may be
affected by factors including the depth of tissue sections, conditions for antigen retrieval, length
of time sections are incubated with primary and secondary antibodies, and the detection reagent
used. We have standardized our immunohistochemistry protocol to minimize deviations in
intensity due to factors such as these.
Consideration must also be made regarding the use of thyroid carcinoma cell lines for secretome
analysis. Secreted proteins are challenging targets of study because they are frequently found in
lower concentrations due to high dilution in bodily fluids or cell culture media, may be masked
by non-secreted proteins due to cell death and by proteins found in serum in culture media (i.e.
fetal bovine serum).194 For these reasons, careful attention must be paid to limit cell death
induced by cellular stress and to minimize contamination from non-secretome proteins. Due to
their affordability, relative ease of use, and reproducibility of results, cell lines remain a widely

61
used tool in biomarker discovery.195 The conditioned media of cancer cell lines, as discussed
before, contains secreted and shed proteins that have been released via classical and non-classical
secretory pathways. The reduced complexity of this mixture, in comparison to other sources such
as blood and cell lysates, improves the potential to identify lower abundance proteins.196
Furthermore, quantifiable and reproducible results may be produced from a large number of cell
lines characteristic of different stages of a cancer, such as the anaplastic and papillary-derived
thyroid carcinoma cell lines used in our study.196
As with any model system, cancer cell lines have certain limitations that must be carefully
considered by researchers. The cancer cell secretome represents an in vitro system which ignores
potentially important contributions of host-tumor microenvironment.197 Furthermore, it is well
documented that genotypic and phenotypic alterations in the cell lines occur over time and may
produce distinct subpopulations of any given cell line.101 These subpopulations of thyroid
carcinoma cell lines have been thoroughly investigated in a previous study by Schweppe et al
and only confirmed original, unique cell lines were used in this study (confirmed by STR profile
analysis).101 With these limitations in mind, attention must also be drawn to mining strategies for
the selection of the optimal biomarker candidates for further study. Currently, selection of
optimal protein candidates for study past the discovery phase has included the examination of
proteins detected in certain conditions (e.g. aggressive versus non-aggressive carcinoma cell
lines), the use of bioinformatics tools, and even consideration of subcellular localization of
proteins.197 The identification of numerous proteins already in use as cancer biomarkers in the
cancer cell line secretome provides a proof-of-principle for the use of secretome analysis in the
discovery of biomarkers of aggressive carcinomas.194
Our secretome analysis of thyroid carcinoma cell lines has uncovered numerous potential
biological markers for aggressive thyroid carcinomas. We have verified many of these proteins
in patient tissues and sera and determined some may be differentially detected in the blood of
thyroid carcinoma patients. This work has paved the way for the future development of both
histological and blood-based assays that may be used in thyroid carcinoma diagnostics.
Importantly, the potential of secretome approaches extends beyond the ability to quickly identify
candidate biological markers as we have demonstrated. The protein identifications generated
from this analysis may also uncover proteins and important pathways that may be contributors to
tumor progression and aggressiveness. Further exploration of these pathways will allow for a

62
greater understanding of these tumor processes and may ultimately lead to improvements in
patient treatment strategies. It is clear that many of the identified proteins in the secretome
interact with each other and are involved in tumorigenesis and progression. This is illustrated in
our identification of proteins such as vimentin, AXL receptor tyrosine kinase, plasminogen
activator urokinase, and dickkopf-related protein 3 – proteins proposed to interact with each
other and play important roles in epithelial-to-mesenchymal transition.198 Future work that
combines bioinformatics-based analysis with biological exploration of these proteins may help
further our understanding of the most aggressive thyroid carcinomas.
5.2 Conclusion Our detection of proteins in the sera of thyroid carcinoma patients demonstrates the feasibility of
using a proteomics-based secretome anlaysis approach to identify candidate minimally-invasive
biomarkers. We identified many novel proteins for future consideration in the management and
diagnosis of thyroid carcinoma which have been verified in thyroid carcinoma patients’ sera
and/or tissues. Notably, immunohistochemistry revealed increased PTMA expression in both
nucleus and cytoplasm of ATC, compared to normal thyroid adjacent to benign thyroid disease,
poorly differentiated (insular) and papillary carcinomas. Furthermore, cytoplasmic expression of
nucleolin was observed only in ATC tissues, suggesting a possible association with tumor
aggressiveness. Analysis of larger numbers of ATCs and poorly-differentiated thyroid
carcinomas in future studies is likely to establish the clinical relevance of these markers.
Quantification of the levels of these proteins in the sera of thyroid carcinoma patients and
characterization of their expression in thyroid carcinoma tissues may serve as the next step
towards evaluating the suitability of these proteins as potential thyroid carcinoma biomarkers.
5.3 Future Directions This body of work may now form the basis of the future exploration of these identified proteins
as putative diagnostic and/or therapeutic targets.
As detailed in our results and discussion, our work has shown that these proteins are detectable
and may exhibit differential levels in the sera of thyroid carcinoma patients. Quantification of
the levels of these proteins will allow for the creation of minimally-invasive blood-based assays
to aid in prognostic and/or diagnostics in thyroid carcinoma patients. To advance these goals, in

63
the past year we have established an ongoing blood database containing detailed clinical
information for approximately 800 thyroid carcinoma patients, patients with benign tumors, and
normal (cancer-free) individuals who have sought medical care at Mount Sinai Hospital
(Toronto, ON, Canada). The anonymized clinical information available for these patients
include date of birth, sex, date of surgery, the presence of lymph node metastases and location,
multiple foci, extrathyroidal invasion, angioinvasion, TNM staging, histologic features, and
tumor size. Furthermore, our database includes patients with pre- and post-operative blood
samples, along with serial blood samples of patients throughout the course of their treatment.
Follow-up information about disease and treatment course is obtainable and will be added to the
database. Screening of the sera of these patients with many of the proteins from the secretome
may allow for the identification of multiple diagnostic markers, both singly or in a panel, that
may serve as molecular signature to identify patients with more aggressive disease and also
subsequently serve as a tool in clinical decision-making.
Many of the proteins identified here may have important roles in the progression of tumors.
Accordingly, many of these proteins may serve as potential therapeutic targets and require futher
exploration. A crucial tool in any “omics”-based strategy is careful bioinformatics analysis to
identify potentially promising candidates for therapeutic consideration. We are currently using
software tools such as Ingenuity Pathway Analysis to identify key proteins in cellular pathways
that may serve as therapeutic targets. Furthermore, our literature reviews have already revealed
experimentally-verified cross-talk between numerous proteins identified in our study. This is
particularly important given the urgent need for new, innovative approaches in the treatment of
the most aggressive thyroid carcinomas.
From diagnostic and therapeutic perspectives, the resources are in place for rapid analysis of
these proteins in thyroid carcinoma cell lines, tissues, and patient sera. Our extensive amounts of
patient specimens and our establishment of a comprehensive patient database brings this research
much closer to its ultimate goal of translating our findings into important clinical applications
that may be used to improve the care of patients. In the long-term, it is highly likely that some of
these proteins will emerge as important diagnostic markers and/or therapeutic targets.

64
References 1. Reis, E. M.; Ojopi, E. P.; Alberto, F. L.; Rahal, P.; Tsukumo, F.; Mancini, U. M.; Guimaraes, G. S.; Thompson, G. M.; Camacho, C.; Miracca, E.; Carvalho, A. L.; Machado, A. A.; Paquola, A. C.; Cerutti, J. M.; da Silva, A. M.; Pereira, G. G.; Valentini, S. R.; Nagai, M. A.; Kowalski, L. P.; Verjovski-Almeida, S.; Tajara, E. H.; Dias-Neto, E.; Bengtson, M. H.; Canevari, R. A.; Carazzolle, M. F.; Colin, C.; Costa, F. F.; Costa, M. C.; Estecio, M. R.; Esteves, L. I.; Federico, M. H.; Guimaraes, P. E.; Hackel, C.; Kimura, E. T.; Leoni, S. G.; Maciel, R. M.; Maistro, S.; Mangone, F. R.; Massirer, K. B.; Matsuo, S. E.; Nobrega, F. G.; Nobrega, M. P.; Nunes, D. N.; Nunes, F.; Pandolfi, J. R.; Pardini, M. I.; Pasini, F. S.; Peres, T.; Rainho, C. A.; dos Reis, P. P.; Rodrigus-Lisoni, F. C.; Rogatto, S. R.; dos Santos, A.; dos Santos, P. C.; Sogayar, M. C.; Zanelli, C. F., Large-scale transcriptome analyses reveal new genetic marker candidates of head, neck, and thyroid cancer. Cancer Res 2005, 65, (5), 1693-9.
2. Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T.; Thun, M. J., Cancer statistics, 2008. CA Cancer J Clin 2008, 58, (2), 71-96.
3. Sipos, J. A.; Mazzaferri, E. L., Thyroid cancer epidemiology and prognostic variables. Clin Oncol (R Coll Radiol) 2010, 22, (6), 395-404.
4. Hundahl, S. A.; Fleming, I. D.; Fremgen, A. M.; Menck, H. R., A National Cancer Data Base report on 53,856 cases of thyroid carcinoma treated in the U.S., 1985-1995 [see commetns]. Cancer 1998, 83, (12), 2638-48.
5. Smallridge, R. C.; Copland, J. A., Anaplastic thyroid carcinoma: pathogenesis and emerging therapies. Clin Oncol (R Coll Radiol) 2010, 22, (6), 486-97.
6. O'Neill, C. J.; Oucharek, J.; Learoyd, D.; Sidhu, S. B., Standard and Emerging Therapies for Metastatic Differentiated Thyroid Cancer. Oncologist 2010, 15, (2), 146-156.
7. Hundahl, S. A.; Fleming, I. D.; Fremgen, A. M.; Menck, H. R., GLOBOCAN 2000: cancer incidence, mortality and prevalence worldwide.
8. Kilfoy, B. A.; Zheng, T.; Holford, T. R.; Han, X.; Ward, M. H.; Sjodin, A.; Zhang, Y.; Bai, Y.; Zhu, C.; Guo, G. L.; Rothman, N., International patterns and trends in thyroid cancer incidence, 1973-2002. Cancer Causes Control 2009, 20, (5), 525-31.
9. Enewold, L.; Zhu, K.; Ron, E.; Marrogi, A. J.; Stojadinovic, A.; Peoples, G. E.; Devesa, S. S., Rising thyroid cancer incidence in the United States by demographic and tumor characteristics, 1980-2005. Cancer Epidemiol Biomarkers Prev 2009, 18, (3), 784-91.
10. Davies, L.; Welch, H. G., Increasing incidence of thyroid cancer in the United States, 1973-2002. JAMA 2006, 295, (18), 2164-7.
11. Mazzaferri, E. L., Managing small thyroid cancers. JAMA 2006, 295, (18), 2179-82.
12. Chen, A. Y.; Jemal, A.; Ward, E. M., Increasing incidence of differentiated thyroid cancer in the United States, 1988-2005. Cancer 2009, 115, (16), 3801-7.

65
13. Aschebrook-Kilfoy, B.; Ward, M. H.; Sabra, M. M.; Devesa, S. S., Thyroid cancer incidence patterns in the United States by histologic type, 1992-2006. Thyroid 2011, 21, (2), 125-34.
14. Albores-Saavedra, J.; Wu, J., The many faces and mimics of papillary thyroid carcinoma. Endocr Pathol 2006, 17, (1), 1-18.
15. Silver, C. E.; Owen, R. P.; Rodrigo, J. P.; Rinaldo, A.; Devaney, K. O.; Ferlito, A., Aggressive variants of papillary thyroid carcinoma. Head Neck 2010.
16. Mazzaferri, E. L.; Jhiang, S. M., Long-term impact of initial surgical and medical therapy on papillary and follicular thyroid cancer. Am J Med 1994, 97, (5), 418-28.
17. Kouvaraki, M. A.; Shapiro, S. E.; Fornage, B. D.; Edeiken-Monro, B. S.; Sherman, S. I.; Vassilopoulou-Sellin, R.; Lee, J. E.; Evans, D. B., Role of preoperative ultrasonography in the surgical management of patients with thyroid cancer. Surgery 2003, 134, (6), 946-54; discussion 954-5.
18. Podnos, Y. D.; Smith, D.; Wagman, L. D.; Ellenhorn, J. D., The implication of lymph node metastasis on survival in patients with well-differentiated thyroid cancer. Am Surg 2005, 71, (9), 731-4.
19. Bhattacharyya, N., A population-based analysis of survival factors in differentiated and medullary thyroid carcinoma. Otolaryngol Head Neck Surg 2003, 128, (1), 115-23.
20. Zaydfudim, V.; Feurer, I. D.; Griffin, M. R.; Phay, J. E., The impact of lymph node involvement on survival in patients with papillary and follicular thyroid carcinoma. Surgery 2008, 144, (6), 1070-7; discussion 1077-8.
21. Sampson, E.; Brierley, J. D.; Le, L. W.; Rotstein, L.; Tsang, R. W., Clinical management and outcome of papillary and follicular (differentiated) thyroid cancer presenting with distant metastasis at diagnosis. Cancer 2007, 110, (7), 1451-6.
22. Durante, C.; Haddy, N.; Baudin, E.; Leboulleux, S.; Hartl, D.; Travagli, J. P.; Caillou, B.; Ricard, M.; Lumbroso, J. D.; De Vathaire, F.; Schlumberger, M., Long-term outcome of 444 patients with distant metastases from papillary and follicular thyroid carcinoma: benefits and limits of radioiodine therapy. J Clin Endocrinol Metab 2006, 91, (8), 2892-9.
23. Tang, K. T.; Lee, C. H., BRAF mutation in papillary thyroid carcinoma: pathogenic role and clinical implications. J Chin Med Assoc 2010, 73, (3), 113-28.
24. Cohen, Y.; Rosenbaum, E.; Clark, D. P.; Zeiger, M. A.; Umbricht, C. B.; Tufano, R. P.; Sidransky, D.; Westra, W. H., Mutational analysis of BRAF in fine needle aspiration biopsies of the thyroid: a potential application for the preoperative assessment of thyroid nodules. Clin Cancer Res 2004, 10, (8), 2761-5.
25. Salvatore, G.; Giannini, R.; Faviana, P.; Caleo, A.; Migliaccio, I.; Fagin, J. A.; Nikiforov, Y. E.; Troncone, G.; Palombini, L.; Basolo, F.; Santoro, M., Analysis of BRAF point mutation

66
and RET/PTC rearrangement refines the fine-needle aspiration diagnosis of papillary thyroid carcinoma. J Clin Endocrinol Metab 2004, 89, (10), 5175-80.
26. Khanafshar, E.; Lloyd, R. V., The spectrum of papillary thyroid carcinoma variants. Adv Anat Pathol 2011, 18, (1), 90-7.
27. Passler, C.; Prager, G.; Scheuba, C.; Niederle, B. E.; Kaserer, K.; Zettinig, G.; Niederle, B., Follicular variant of papillary thyroid carcinoma: a long-term follow-up. Arch Surg 2003, 138, (12), 1362-6.
28. Salajegheh, A.; Petcu, E. B.; Smith, R. A.; Lam, A. K., Follicular variant of papillary thyroid carcinoma: a diagnostic challenge for clinicians and pathologists. Postgrad Med J 2008, 84, (988), 78-82.
29. Lin, H. W.; Bhattacharyya, N., Clinical behavior of follicular variant of papillary thyroid carcinoma: presentation and survival. Laryngoscope 2010, 120 Suppl 4, S163.
30. Baloch, Z. W.; Shafique, K.; Flannagan, M.; Livolsi, V. A., Encapsulated classic and follicular variants of papillary thyroid carcinoma: comparative clinicopathologic study. Endocr Pract 2010, 16, (6), 952-9.
31. Ghossein, R., Encapsulated malignant follicular cell-derived thyroid tumors. Endocr Pathol 2010, 21, (4), 212-8.
32. Lee, J.; Hasteh, F., Oncocytic variant of papillary thyroid carcinoma associated with Hashimoto's thyroiditis: a case report and review of the literature. Diagn Cytopathol 2009, 37, (8), 600-6.
33. Gross, M.; Eliashar, R.; Ben-Yaakov, A.; Weinberger, J. M.; Maly, B., Clinicopathologic features and outcome of the oncocytic variant of papillary thyroid carcinoma. Ann Otol Rhinol Laryngol 2009, 118, (5), 374-81.
34. Koo, J. S.; Hong, S.; Park, C. S., Diffuse sclerosing variant is a major subtype of papillary thyroid carcinoma in the young. Thyroid 2009, 19, (11), 1225-31.
35. LiVolsi, V. A., Papillary carcinoma tall cell variant (TCV): a review. Endocr Pathol 2010, 21, (1), 12-5.
36. Rufini, V.; Salvatori, M.; Fadda, G.; Pinnarelli, L.; Castaldi, P.; Maussier, M. L.; Galli, G., Thyroid carcinomas with a variable insular component. Cancer 2007, 110, (6), 1209-1217.
37. Volante, M.; Collini, P.; Nikiforov, Y. E.; Sakamoto, A.; Kakudo, K.; Katoh, R.; Lloyd, R. V.; LiVolsi, V. A.; Papotti, M.; Sobrinho-Simoes, M.; Bussolati, G.; Rosai, J., Poorly differentiated thyroid carcinoma: the Turin proposal for the use of uniform diagnostic criteria and an algorithmic diagnostic approach. Am J Surg Pathol 2007, 31, (8), 1256-64.
38. Pilotti, S.; Collini, P.; Mariani, L.; Placucci, M.; Bongarzone, I.; Vigneri, P.; Cipriani, S.; Falcetta, F.; Miceli, R.; Pierotti, M. A.; Rilke, F., Insular Carcinoma: A Distinct De Novo Entity

67
Among Follicular Carcinomas of the Thyroid Gland. The American Journal of Surgical Pathology 1997, 21, (12), 1466-1473.
39. Sasaki, A.; Daa, T.; Kashima, K.; Yokoyama, S.; Nakayama, I.; Noguchi, S., Insular component as a risk factor of thyroid carcinoma. Pathol Int 1996, 46, (12), 939-46.
40. Nishida, T.; Katayama, S.-i.; Tsujimoto, M.; Nakamura, J.-i.; Matsuda, H., Clinicopathological Significance of Poorly Differentiated Thyroid Carcinoma. The American Journal of Surgical Pathology 1999, 23, (2), 205-211.
41. Volante, M.; Landolfi, S.; Chiusa, L.; Palestini, N.; Motta, M.; Codegone, A.; Torchio, B.; Papotti, M. G., Poorly differentiated carcinomas of the thyroid with trabecular, insular, and solid patterns. Cancer 2004, 100, (5), 950-957.
42. O’Neill, J.; Power, D.; Condron, C.; Bouchier-Hayes, D.; Walsh, M., Anaplastic thyroid cancer, tumorigenesis and therapy. Irish Journal of Medical Science 2010, 179, (1), 9-15.
43. Kebebew, E.; Greenspan, F. S.; Clark, O. H.; Woeber, K. A.; McMillan, A., Anaplastic thyroid carcinoma. Cancer 2005, 103, (7), 1330-1335.
44. Are, C.; Shaha, A., Anaplastic Thyroid Carcinoma: Biology, Pathogenesis, Prognostic Factors, and Treatment Approaches. Annals of Surgical Oncology 2006, 13, (4), 453-464.
45. Pitt, S. C.; Moley, J. F., Medullary, Anaplastic, and Metastatic Cancers of the Thyroid. Seminars in Oncology 2010, 37, (6), 567-579.
46. Ain, K. B., Anaplastic thyroid carcinoma: A therapeutic challenge. Seminars in Surgical Oncology 1999, 16, (1), 64-69.
47. Smallridge, R. C.; Marlow, L. A.; Copland, J. A., Anaplastic thyroid cancer: molecular pathogenesis and emerging therapies. Endocr Relat Cancer 2009, 16, (1), 17-44.
48. Levendag, P. C.; De Porre, P. M.; van Putten, W. L., Anaplastic carcinoma of the thyroid gland treated by radiation therapy. Int J Radiat Oncol Biol Phys 1993, 26, (1), 125-8.
49. Tennvall, J.; Lundell, G.; Wahlberg, P.; Bergenfelz, A.; Grimelius, L.; Akerman, M.; Hjelm Skog, A. L.; Wallin, G., Anaplastic thyroid carcinoma: three protocols combining doxorubicin, hyperfractionated radiotherapy and surgery. Br J Cancer 2002, 86, (12), 1848-53.
50. Smit, J. W.; Schroder-van der Elst, J. P.; Karperien, M.; Que, I.; Stokkel, M.; van der Heide, D.; Romijn, J. A., Iodide kinetics and experimental (131)I therapy in a xenotransplanted human sodium-iodide symporter-transfected human follicular thyroid carcinoma cell line. J Clin Endocrinol Metab 2002, 87, (3), 1247-53.
51. Hsieh, Y. J.; Ke, C. C.; Liu, R. S.; Wang, F. H.; Tang, K. T.; Chi, C. W.; Chen, F. D.; Lee, C. H., Radioiodide imaging and treatment of ARO cancer xenograft in a mouse model after expression of human sodium iodide symporter. Anticancer Res 2007, 27, (4B), 2515-22.

68
52. Presta, I.; Arturi, F.; Ferretti, E.; Mattei, T.; Scarpelli, D.; Tosi, E.; Scipioni, A.; Celano, M.; Gulino, A.; Filetti, S.; Russo, D., Recovery of NIS expression in thyroid cancer cells by overexpression of Pax8 gene. BMC Cancer 2005, 5, 80.
53. Spencer, C. A.; LoPresti, J. S., Technology Insight: measuring thyroglobulin and thyroglobulin autoantibody in patients with differentiated thyroid cancer. Nat Clin Pract End Met 2008, 4, (4), 223-233.
54. Schulz, R., Evidence for immunological differences between circulating and tissue-derived thyroglobulin in men. Eur J Clin Invest 1989, 19, 459-463.
55. Spencer, C. A., Clinical impact of thyroglobulin (Tg) and Tg autoantibody method differences on the management of patients with differentiated thyroid carcinomas. J Clin Endocrinol Metab 2005, 90, 5566-5575.
56. Zanotti-Fregonara, P.; Keller, I.; Calzada-Nocaudie, M.; Al-Nahhas, A.; Devaux, J.-Y.; Grassetto, G.; Marzola, M. C.; Rubello, D.; Hindié, E., Increased serum thyroglobulin levels and negative imaging in thyroid cancer patients: are there sources of benign secretion? A speculative short review. Nuclear Medicine Communications 2010, 31, (12), 1054-1058 10.1097/MNM.0b013e328340e717.
57. Weightman, D. R.; Mallick, U. K.; Fenwick, J. D.; Perros, P., Discordant serum thyroglobulin results generated by two classes of assay in patients with thyroid carcinoma. Cancer 2003, 98, (1), 41-47.
58. Schlumberger, M., Comparison of seven serum thyroglobulin assays in the follow-up of papillary and follicular thyroid cancer patients. J Clin Endocrinol Metab 2007, 92, 2487-2495.
59. Preissner, C. M., Phantoms in the assay tube: heterophile antibody interferences in serum thyroglobulin assays. J Clin Endocrinol Metab 2003, 88, 3069-3074.
60. Spencer, C. A., Serum thyroglobulin autoantibodies: prevalence, influence on serum thyroglobulin measurement and prognostic significance in patients with differentiated thyroid carcinoma. J Clin Endocrinol Metab 1998, 83, 1121-1127.
61. Gorges, R., Development and clinical impact of thyroglobulin antibodies in patients with differentiated thyroid carcinoma during the first 3 years after thyroidectomy. Eur J Endocrinol 2005, 153, 49-55.
62. Piechaczyk, M.; Baldet, L.; Pau, B.; Bastide, J. M., Novel immunoradiometric assay of thyroglobulin in serum with use of monoclonal antibodies selected for lack of cross-reactivity with autoantibodies. Clin Chem 1989, 35, (3), 422-4.
63. Chung, J. K., Clinical significance of elevated level of serum antithyroglobulin antibody in patients with differentiated thyroid cancer after thyroid ablation. Clin Endocrinol (Oxf) 2002, 57, 215-221.
64. Kim, W. G.; Yoon, J. H.; Kim, W. B.; Kim, T. Y.; Kim, E. Y.; Kim, J. M.; Ryu, J.-S.; Gong, G.; Hong, S. J.; Shong, Y. K., Change of Serum Antithyroglobulin Antibody Levels Is

69
Useful for Prediction of Clinical Recurrence in Thyroglobulin-Negative Patients with Differentiated Thyroid Carcinoma. J Clin Endocrinol Metab 2008, 93, (12), 4683-4689.
65. Smallridge, R. C.; Meek, S. E.; Morgan, M. A.; Gates, G. S.; Fox, T. P.; Grebe, S.; Fatourechi, V., Monitoring thyroglobulin in a sensitive immunoassay has comparable sensitivity to recombinant human tsh-stimulated thyroglobulin in follow-up of thyroid cancer patients. J Clin Endocrinol Metab 2007, 92, (1), 82-7.
66. Mazzaferri, E. L., A consensus report of the role of serum thyroglobulin as a monitoring method for low-risk patients with papillary thyroid carcinoma. J Clin Endocrinol Metab 2003, 88, 1433-1441.
67. Cole, T. G.; Johnson, D.; Eveland, B. J.; Nahm, M. H., Cost-effective method for detection of "hook effect" in tumor marker immunometric assays. Clin Chem 1993, 39, (4), 695-6.
68. Eustatia-Rutten, C. F., Diagnostic value of serum thyroglobulin measurements in the follow-up of differentiated thyroid carcinoma, a structured meta-analysis. Clin Endocrinol (Oxf) 2004, 61, 61-74.
69. Pacini, F., Recombinant human thyrotropin-stimulated serum thyroglobulin combined with neck ultrasonography has the highest sensitivity in monitoring differentiated thyroid carcinoma. J Clin Endocrinol Metab 2003, 88, 3668-3673.
70. Castagna, M. G.; Brilli, L.; Pilli, T.; Montanaro, A.; Cipri, C.; Fioravanti, C.; Sestini, F.; Capezzone, M.; Pacini, F., Limited value of repeat recombinant human thyrotropin (rhTSH)-stimulated thyroglobulin testing in differentiated thyroid carcinoma patients with previous negative rhTSH-stimulated thyroglobulin and undetectable basal serum thyroglobulin levels. J Clin Endocrinol Metab 2008, 93, (1), 76-81.
71. Ozata, M.; Suzuki, S.; Miyamoto, T.; Liu, R. T.; Fierro-Renoy, F.; DeGroot, L. J., Serum thyroglobulin in the follow-up of patients with treated differentiated thyroid cancer. J Clin Endocrinol Metab 1994, 79, (1), 98-105.
72. Lo Gerfo, P.; Colacchio, T. A.; Colacchio, D. A.; Feind, C. R., Effect of TSH stimulation on serum thyroglobulin in metastatic thyroid cancer. J Surg Oncol 1980, 14, (3), 195-200.
73. Giovanella, L.; Ghelfo, A., Undetectable serum thyroglobulin due to negative interference of heterophile antibodies in relapsing thyroid carcinoma. Clin Chem 2007, 53, 1871-1872.
74. Spencer, C. A.; Wang, C. C., Thyroglobulin measurement. Techniques, clinical benefits, and pitfalls. Endocrinol Metab Clin North Am 1995, 24, (4), 841-63.
75. Baudin, E., Positive predictive value of serum thyroglobulin levels, measured during the first year of follow-up after thyroid hormone withdrawal, in thyroid cancer patients. J Clin Endocrinol Metab 2003, 88, 1107-1111.

70
76. Kim, T. Y.; Kim, W. B.; Kim, E. S.; Ryu, J. S.; Yeo, J. S.; Kim, S. C.; Hong, S. J.; Shong, Y. K., Serum thyroglobulin levels at the time of 131I remnant ablation just after thyroidectomy are useful for early prediction of clinical recurrence in low-risk patients with differentiated thyroid carcinoma. J Clin Endocrinol Metab 2005, 90, (3), 1440-5.
77. Vaisman, A.; Orlov, S.; Yip, J.; Hu, C.; Lim, T.; Dowar, M.; Freeman, J. L.; Walfish, P. G., Application of post-surgical stimulated thyroglobulin for radioiodine remnant ablation selection in low-risk papillary thyroid carcinoma. Head & Neck 2010, 32, (6), 689-698.
78. Robinson, M. J.; Cobb, M. H., Mitogen-activated protein kinase pathways. Curr Opin Cell Biol 1997, 9, (2), 180-6.
79. Nikiforov, Y. E.; Biddinger, P. W.; Thompson, L. D. R., Diagnostic Pathology and Molecular Genetics of the Thyroid. Lippincott Williams & Wilkins: Baltimore, 2009.
80. Adeniran, A. J.; Zhu, Z.; Gandhi, M.; Steward, D. L.; Fidler, J. P.; Giordano, T. J.; Biddinger, P. W.; Nikiforov, Y. E., Correlation Between Genetic Alterations and Microscopic Features, Clinical Manifestations, and Prognostic Characteristics of Thyroid Papillary Carcinomas. The American Journal of Surgical Pathology 2006, 30, (2), 216-222.
81. Nikiforova, M. N.; Kimura, E. T.; Gandhi, M.; Biddinger, P. W.; Knauf, J. A.; Basolo, F.; Zhu, Z.; Giannini, R.; Salvatore, G.; Fusco, A.; Santoro, M.; Fagin, J. A.; Nikiforov, Y. E., BRAF Mutations in Thyroid Tumors Are Restricted to Papillary Carcinomas and Anaplastic or Poorly Differentiated Carcinomas Arising from Papillary Carcinomas. J Clin Endocrinol Metab 2003, 88, (11), 5399-5404.
82. Nikiforova, M. N.; Nikiforov, Y. E., Molecular diagnostics and predictors in thyroid cancer. Thyroid 2009, 19, (12), 1351-61.
83. Brandi, M. L.; Gagel, R. F.; Angeli, A.; Bilezikian, J. P.; Beck-Peccoz, P.; Bordi, C.; Conte-Devolx, B.; Falchetti, A.; Gheri, R. G.; Libroia, A.; Lips, C. J. M.; Lombardi, G.; Mannelli, M.; Pacini, F.; Ponder, B. A. J.; Raue, F.; Skogseid, B.; Tamburrano, G.; Thakker, R. V.; Thompson, N. W.; Tomassetti, P.; Tonelli, F.; Wells, S. A., Jr.; Marx, S. J., CONSENSUS: Guidelines for Diagnosis and Therapy of MEN Type 1 and Type 2. J Clin Endocrinol Metab 2001, 86, (12), 5658-5671.
84. de Groot, J. W. B.; Links, T. P.; Plukker, J. T. M.; Lips, C. J. M.; Hofstra, R. M. W., RET as a Diagnostic and Therapeutic Target in Sporadic and Hereditary Endocrine Tumors. Endocr Rev 2006, 27, (5), 535-560.
85. Carpi, A.; Di Coscio, G.; Iervasi, G.; Antonelli, A.; Mechanick, J.; Sciacchitano, S.; Nicolini, A., Thyroid fine needle aspiration: how to improve clinicians' confidence and performance with the technique. Cancer Lett 2008, 264, (2), 163-71.
86. Damante, G. S., A.; Tell, G.; , Thyroid Tumors: novel insights from proteomics studies. Expert Rev Proteomics 2009, 6, (4), 363-376.
87. Chung, L. W.; Baseman, A.; Assikis, V.; Zhau, H. E., Molecular insights into prostate cancer progression: the missing link of tumor microenvironment. J Urol 2005, 173, (1), 10-20.

71
88. Liotta, L. A.; Ferrari, M.; Petricoin, E., Clinical proteomics: written in blood. Nature 2003, 425, (6961), 905.
89. Villanueva, J.; Nazarian, A.; Lawlor, K.; Yi, S. S.; Robbins, R. J.; Tempst, P., A sequence-specific exopeptidase activity test (SSEAT) for "functional" biomarker discovery. Mol Cell Proteomics 2008, 7, (3), 509-18.
90. Rifai, N.; Gillette, M. A.; Carr, S. A., Protein biomarker discovery and validation: the long and uncertain path to clinical utility. Nat Biotechnol 2006, 24, (8), 971-83.
91. Hu, S.; Arellano, M.; Boontheung, P.; Wang, J.; Zhou, H.; Jiang, J.; Elashoff, D.; Wei, R.; Loo, J. A.; Wong, D. T., Salivary proteomics for oral cancer biomarker discovery. Clin Cancer Res 2008, 14, (19), 6246-52.
92. DeSouza, L. V.; Grigull, J.; Ghanny, S.; Dube, V.; Romaschin, A. D.; Colgan, T. J.; Siu, K. W., Endometrial carcinoma biomarker discovery and verification using differentially tagged clinical samples with multidimensional liquid chromatography and tandem mass spectrometry. Mol Cell Proteomics 2007, 6, (7), 1170-82.
93. Volmer, M. W.; Stuhler, K.; Zapatka, M.; Schoneck, A.; Klein-Scory, S.; Schmiegel, W.; Meyer, H. E.; Schwarte-Waldhoff, I., Differential proteome analysis of conditioned media to detect Smad4 regulated secreted biomarkers in colon cancer. Proteomics 2005, 5, (10), 2587-601.
94. Sardana, G.; Jung, K.; Stephan, C.; Diamandis, E. P., Proteomic analysis of conditioned media from the PC3, LNCaP, and 22Rv1 prostate cancer cell lines: discovery and validation of candidate prostate cancer biomarkers. J Proteome Res 2008, 7, (8), 3329-38.
95. Kulasingam, V.; Diamandis, E. P., Proteomics analysis of conditioned media from three breast cancer cell lines: a mine for biomarkers and therapeutic targets. Mol Cell Proteomics 2007, 6, (11), 1997-2011.
96. Lawlor, K.; Nazarian, A.; Lacomis, L.; Tempst, P.; Villanueva, J., Pathway-based biomarker search by high-throughput proteomics profiling of secretomes. J Proteome Res 2009, 8, (3), 1489-503.
97. Gourin, C. G.; Zhi, W.; Adam, B. L., Proteomic identification of serum biomarkers for head and neck cancer surveillance. Laryngoscope 2009, 119, (7), 1291-302.
98. Bijian, K.; Mlynarek, A. M.; Balys, R. L.; Jie, S.; Xu, Y.; Hier, M. P.; Black, M. J.; Di Falco, M. R.; LaBoissiere, S.; Alaoui-Jamali, M. A., Serum proteomic approach for the identification of serum biomarkers contributed by oral squamous cell carcinoma and host tissue microenvironment. J Proteome Res 2009, 8, (5), 2173-85.
99. Gioanni, J.; Zanghellini, E.; Mazeau, C.; Zhang, D.; Courdi, A.; Farges, M.; Lambert, J. C.; Duplay, H.; Schneider, M., [Characterization of a human cell line from an anaplastic carcinoma of the thyroid gland]. Bull Cancer 1991, 78, (11), 1053-62.

72
100. Tanaka, J.; Ogura, T.; Sato, H.; Hatano, M., Establishment and biological characterization of an in vitro human cytomegalovirus latency model. Virology 1987, 161, (1), 62-72.
101. Schweppe, R. E.; Klopper, J. P.; Korch, C.; Pugazhenthi, U.; Benezra, M.; Knauf, J. A.; Fagin, J. A.; Marlow, L. A.; Copland, J. A.; Smallridge, R. C.; Haugen, B. R., Deoxyribonucleic acid profiling analysis of 40 human thyroid cancer cell lines reveals cross-contamination resulting in cell line redundancy and misidentification. J Clin Endocrinol Metab 2008, 93, (11), 4331-41.
102. Meireles, A. M.; Preto, A.; Rocha, A. S.; Rebocho, A. P.; Maximo, V.; Pereira-Castro, I.; Moreira, S.; Feijao, T.; Botelho, T.; Marques, R.; Trovisco, V.; Cirnes, L.; Alves, C.; Velho, S.; Soares, P.; Sobrinho-Simoes, M., Molecular and genotypic characterization of human thyroid follicular cell carcinoma-derived cell lines. Thyroid 2007, 17, (8), 707-15.
103. Al-Nedawi, K.; Szemraj, J.; Cierniewski, C. S., Mast cell-derived exosomes activate endothelial cells to secrete plasminogen activator inhibitor type 1. Arterioscler Thromb Vasc Biol 2005, 25, (8), 1744-9.
104. DeSouza, L.; Diehl, G.; Rodrigues, M. J.; Guo, J.; Romaschin, A. D.; Colgan, T. J.; Siu, K. W., Search for cancer markers from endometrial tissues using differentially labeled tags iTRAQ and cICAT with multidimensional liquid chromatography and tandem mass spectrometry. J Proteome Res 2005, 4, (2), 377-86.
105. Shilov, I. V.; Seymour, S. L.; Patel, A. A.; Loboda, A.; Tang, W. H.; Keating, S. P.; Hunter, C. L.; Nuwaysir, L. M.; Schaeffer, D. A., The Paragon Algorithm, a Next Generation Search Engine That Uses Sequence Temperature Values and Feature Probabilities to Identify Peptides from Tandem Mass Spectra. Molecular & Cellular Proteomics 2007, 6, (9), 1638-1655.
106. Bendtsen, J. D.; Nielsen, H.; von Heijne, G.; Brunak, S., Improved prediction of signal peptides: SignalP 3.0. J Mol Biol 2004, 340, (4), 783-95.
107. Bendtsen, J. D.; Jensen, L. J.; Blom, N.; Von Heijne, G.; Brunak, S., Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng Des Sel 2004, 17, (4), 349-56.
108. Tripathi, S. C.; Matta, A.; Kaur, J.; Grigull, J.; Chauhan, S. S.; Thakar, A.; Shukla, N. K.; Duggal, R.; Choudhary, A. R.; Dattagupta, S.; Sharma, M. C.; Ralhan, R.; Siu, K. W., Overexpression of prothymosin alpha predicts poor disease outcome in head and neck cancer. PLoS One 2011, 6, (5), e19213.
109. Ralhan, R.; Desouza, L. V.; Matta, A.; Chandra Tripathi, S.; Ghanny, S.; Datta Gupta, S.; Bahadur, S.; Siu, K. W., Discovery and verification of head-and-neck cancer biomarkers by differential protein expression analysis using iTRAQ labeling, multidimensional liquid chromatography, and tandem mass spectrometry. Mol Cell Proteomics 2008, 7, (6), 1162-73.
110. Ito, M.; Hiramatsu, H.; Kobayashi, K.; Suzue, K.; Kawahata, M.; Hioki, K.; Ueyama, Y.; Koyanagi, Y.; Sugamura, K.; Tsuji, K.; Heike, T.; Nakahata, T., NOD/SCID/gamma(c)(null)

73
mouse: an excellent recipient mouse model for engraftment of human cells. Blood 2002, 100, (9), 3175-82.
111. Jou, Y.-C.; Tung, C.-L.; Tsai, Y.-S.; Shen, C.-H.; Syue-Yi, C.; Shiau, A.-L.; Tsai, H.-T.; Wu, C.-L.; Tzai, T.-S., Prognostic Relevance of Prothymosin-[alpha] Expression in Human Upper Urinary Tract Transitional Cell Carcinoma. Urology 2009, 74, (4), 951-957.
112. Letsas, K. P.; Vartholomatos, G.; Tsepi, C.; Tsatsoulis, A.; Frangou-Lazaridis, M., Fine-needle aspiration biopsy-RT-PCR expression analysis of prothymosin alpha and parathymosin in thyroid: novel proliferation markers? Neoplasma 2007, 54, (1), 57-62.
113. Suzuki, S.; Takahashi, S.; Takeshita, K.; Hikosaka, A.; Wakita, T.; Nishiyama, N.; Fujita, T.; Okamura, T.; Shirai, T., Expression of prothymosin alpha is correlated with development and progression in human prostate cancers. Prostate 2006, 66, (5), 463-9.
114. Tsai, Y. S.; Jou, Y. C.; Lee, G. F.; Chen, Y. C.; Shiau, A. L.; Tsai, H. T.; Wu, C. L.; Tzai, T. S., Aberrant prothymosin-alpha expression in human bladder cancer. Urology 2009, 73, (1), 188-92.
115. Destouches, D.; El Khoury, D.; Hamma-Kourbali, Y.; Krust, B.; Albanese, P.; Katsoris, P.; Guichard, G.; Briand, J. P.; Courty, J.; Hovanessian, A. G., Suppression of tumor growth and angiogenesis by a specific antagonist of the cell-surface expressed nucleolin. PLoS One 2008, 3, (6), e2518.
116. Soundararajan, S.; Wang, L.; Sridharan, V.; Chen, W.; Courtenay-Luck, N.; Jones, D.; Spicer, E. K.; Fernandes, D. J., Plasma membrane nucleolin is a receptor for the anticancer aptamer AS1411 in MV4-11 leukemia cells. Mol Pharmacol 2009, 76, (5), 984-91.
117. Tulchin N Fau - Chambon, M.; Chambon M Fau - Juan, G.; Juan G Fau - Dikman, S.; Dikman S Fau - Strauchen, J.; Strauchen J Fau - Ornstein, L.; Ornstein L Fau - Billack, B.; Billack B Fau - Woods, N. T.; Woods Nt Fau - Monteiro, A. N. A.; Monteiro, A. N., BRCA1 protein and nucleolin colocalize in breast carcinoma tissue and cancer cell lines. (1525-2191 (Electronic)).
118. Pucci, S.; Mazzarelli, P.; Nucci, C.; Ricci, F.; Spagnoli, L. G., CLU "in and out": looking for a link. Adv Cancer Res 2009, 105, 93-113.
119. Bi, J.; Guo, A. L.; Lai, Y. R.; Li, B.; Zhong, J. M.; Wu, H. Q.; Xie, Z.; He, Y. L.; Lv, Z. L.; Lau, S. H.; Wang, Q.; Huang, X. H.; Zhang, L. J.; Wen, J. M.; Guan, X. Y., Overexpression of clusterin correlates with tumor progression, metastasis in gastric cancer: a study on tissue microarrays. Neoplasma 2010, 57, (3), 191-7.
120. Chung, J.; Kwak, C.; Jin, R. J.; Lee, C. H.; Lee, K. H.; Lee, S. E., Enhanced chemosensitivity of bladder cancer cells to cisplatin by suppression of clusterin in vitro. Cancer Lett 2004, 203, (2), 155-61.
121. Yang, G. F.; Li, X. M.; Xie, D., Overexpression of clusterin in ovarian cancer is correlated with impaired survival. Int J Gynecol Cancer 2009, 19, (8), 1342-6.

74
122. Zoubeidi, A.; Ettinger, S.; Beraldi, E.; Hadaschik, B.; Zardan, A.; Klomp, L. W.; Nelson, C. C.; Rennie, P. S.; Gleave, M. E., Clusterin Facilitates COMMD1 and I-{kappa}B Degradation to Enhance NF-{kappa}B Activity in Prostate Cancer Cells. Mol Cancer Res 2010.
123. Franzen, C. A.; Chen, C. C.; Todorovic, V.; Juric, V.; Monzon, R. I.; Lau, L. F., Matrix protein CCN1 is critical for prostate carcinoma cell proliferation and TRAIL-induced apoptosis. Mol Cancer Res 2009, 7, (7), 1045-55.
124. Jiang, W.; Li, X.; Rao, S.; Wang, L.; Du, L.; Li, C.; Wu, C.; Wang, H.; Wang, Y.; Yang, B., Constructing disease-specific gene networks using pair-wise relevance metric: application to colon cancer identifies interleukin 8, desmin and enolase 1 as the central elements. BMC Syst Biol 2008, 2, 72.
125. Katanasaka, Y.; Asai, T.; Naitou, H.; Ohashi, N.; Oku, N., Proteomic characterization of angiogenic endothelial cells stimulated with cancer cell-conditioned medium. Biol Pharm Bull 2007, 30, (12), 2300-7.
126. Kang, U.-B.; Ahn, Y.; Lee, J.; Kim, Y.-H.; Kim, J.; Yu, M.-H.; Noh, D.-Y.; Lee, C., Differential profiling of breast cancer plasma proteome by isotope-coded affinity tagging method reveals biotinidase as a breast cancer biomarker. BMC Cancer 2010, 10, (1), 114.
127. Chahed, K.; Kabbage, M.; Hamrita, B.; Guillier, C. L.; Trimeche, M.; Remadi, S.; Ehret-Sabatier, L.; Chouchane, L., Detection of protein alterations in male breast cancer using two dimensional gel electrophoresis and mass spectrometry: the involvement of several pathways in tumorigenesis. Clin Chim Acta 2008, 388, (1-2), 106-14.
128. Muthusamy, B.; Hanumanthu, G.; Suresh, S.; Rekha, B.; Srinivas, D.; Karthick, L.; Vrushabendra, B. M.; Sharma, S.; Mishra, G.; Chatterjee, P.; Mangala, K. S.; Shivashankar, H. N.; Chandrika, K. N.; Deshpande, N.; Suresh, M.; Kannabiran, N.; Niranjan, V.; Nalli, A.; Prasad, T. S. K.; Arun, K. S.; Reddy, R.; Chandran, S.; Jadhav, T.; Julie, D.; Mahesh, M.; John, S. L.; Palvankar, K.; Sudhir, D.; Bala, P.; Rashmi, N. S.; Vishnupriya, G.; Dhar, K.; Reshma, S.; Chaerkady, R.; Gandhi, T. K. B.; Harsha, H. C.; Mohan, S. S.; Deshpande, K. S.; Sarker, M.; Pandey, A., Plasma Proteome Database as a resource for proteomics research. Proteomics 2005, 5, (13), 3531-3536.
129. Schrijvers, E. M.; Koudstaal, P. J.; Hofman, A.; Breteler, M. M., Plasma clusterin and the risk of Alzheimer disease. JAMA 2011, 305, (13), 1322-6.
130. Wasenius, V. M.; Hemmer, S.; Kettunen, E.; Knuutila, S.; Franssila, K.; Joensuu, H., Hepatocyte growth factor receptor, matrix metalloproteinase-11, tissue inhibitor of metalloproteinase-1, and fibronectin are up-regulated in papillary thyroid carcinoma: a cDNA and tissue microarray study. Clin Cancer Res 2003, 9, (1), 68-75.
131. Kutsukake, M.; Ishihara, R.; Momose, K.; Isaka, K.; Itokazu, O.; Higuma, C.; Matsutani, T.; Matsuda, A.; Sasajima, K.; Hara, T.; Tamura, K., Circulating IGF-binding protein 7 (IGFBP7) levels are elevated in patients with endometriosis or undergoing diabetic hemodialysis. Reprod Biol Endocrinol 2008, 6, 54.

75
132. Yu, H. Z.; Li, Z. P.; Li, S. G.; Wu, C. M., Correlations of serous levels of matrix metalloproteinases (MMP-2 and MMP-9) to invasion and metastasis of papillary thyroid carcinoma. Ai Zheng. 2005, 24, (6), 740-742.
133. Maeta, H.; Ohgi, S.; Terada, T., Protein expression of matrix metalloproteinases 2 and 9 and tissue inhibitors of metalloproteinase 1 and 2 in papillary thyroid carcinomas. Virchows Arch 2001, 438, (2), 121-8.
134. Larsen, M. B.; Stephens, R. W.; Brunner, N.; Nielsen, H. J.; Engelholm, L. H.; Christensen, I. J.; Stetler-Stevenson, W. G.; Hoyer-Hansen, G., Quantification of tissue inhibitor of metalloproteinases 2 in plasma from healthy donors and cancer patients. Scand J Immunol 2005, 61, (5), 449-60.
135. Chun, J. K.; Lee, T. J.; Choi, K. M.; Lee, K. H.; Kim, D. S., Elevated anti-alpha-enolase antibody levels in Kawasaki disease. Scand J Rheumatol 2008, 37, (1), 48-52.
136. Langley, K. E.; Bennett, L. G.; Wypych, J.; Yancik, S. A.; Liu, X. D.; Westcott, K. R.; Chang, D. G.; Smith, K. A.; Zsebo, K. M., Soluble stem cell factor in human serum. Blood 1993, 81, (3), 656-60.
137. Komorowski, J.; Pasieka, Z.; Jankiewicz-Wika, J.; Stepien, H., Matrix metalloproteinases, tissue inhibitors of matrix metalloproteinases and angiogenic cytokines in peripheral blood of patients with thyroid cancer. Thyroid 2002, 12, (8), 655-62.
138. Hawthorn, L.; Stein, L.; Varma, R.; Wiseman, S.; Loree, T.; Tan, D., TIMP1 and SERPIN-A overexpression and TFF3 and CRABP1 underexpression as biomarkers for papillary thyroid carcinoma. Head Neck 2004, 26, (12), 1069-83.
139. Pan, S.; Chen, R.; Crispin, D. A.; May, D.; Stevens, T.; McIntosh, M. W.; Bronner, M. P.; Ziogas, A.; Anton-Culver, H.; Brentnall, T. A., Protein alterations associated with pancreatic cancer and chronic pancreatitis found in human plasma using global quantitative proteomics profiling. J Proteome Res 2011, 10, (5), 2359-76.
140. Ekman, C.; Linder, A.; Akesson, P.; Dahlback, B., Plasma concentrations of Gas6 (growth arrest specific protein 6) and its soluble tyrosine kinase receptor sAxl in sepsis and systemic inflammatory response syndromes. Crit Care 2010, 14, (4), R158.
141. Krause, K.; Karger, S.; Sheu, S. Y.; Aigner, T.; Kursawe, R.; Gimm, O.; Schmid, K. W.; Dralle, H.; Fuhrer, D., Evidence for a role of the amyloid precursor protein in thyroid carcinogenesis. J Endocrinol 2008, 198, (2), 291-9.
142. Bush, A. I.; Whyte, S.; Thomas, L. D.; Williamson, T. G.; Van Tiggelen, C. J.; Currie, J.; Small, D. H.; Moir, R. D.; Li, Q. X.; Rumble, B.; et al., An abnormality of plasma amyloid protein precursor in Alzheimer's disease. Ann Neurol 1992, 32, (1), 57-65.
143. Sjoblom, K. G.; Saxne, T.; Wollheim, F. A., Plasma levels of beta 2-microglobulin in rheumatoid arthritis. Ann Rheum Dis 1980, 39, (4), 333-9.

76
144. Ristamäki, R.; Joensuu, H.; Lappalainen, K.; Teerenhovi, L.; Jalkanen, S., Elevated Serum CD44 Level Is Associated With Unfavorable Outcome in Non-Hodgkin's Lymphoma. Blood 1997, 90, (10), 4039-4045.
145. Manetti, L.; Pardini, E.; Genovesi, M.; Campomori, A.; Grasso, L.; Morselli, L. L.; Lupi, I.; Pellegrini, G.; Bartalena, L.; Bogazzi, F.; Martino, E., Thyroid function differently affects serum cystatin C and creatinine concentrations. J Endocrinol Invest 2005, 28, (4), 346-9.
146. Ristiniemi, N.; Savage, C.; Bruun, L.; Pettersson, K.; Lilja, H.; Christensson, A., Evaluation of a new immunoassay for cystatin C, based on a double monoclonal principle, in men with normal and impaired renal function. Nephrol Dial Transplant 2011.
147. Peehl, D. M.; Chen, Z.; Nolley, R., Serum Mac-2BP does not distinguish men with high grade, large volume prostate cancer from men with benign prostatic hyperplasia. Prostate 2011, 71, (1), 26-31.
148. Thompson, C.; Blumenstock, F. A.; Saba, T. M.; Feustel, P. J.; Kaplan, J. E.; Fortune, J. B.; Hough, L.; Gray, V., Plasma fibronectin synthesis in normal and injured humans as determined by stable isotope incorporation. The Journal of Clinical Investigation 1989, 84, (4), 1226-1235.
149. Shi, H.; Huang, Y.; Zhou, H.; Song, X.; Yuan, S.; Fu, Y.; Luo, Y., Nucleolin is a receptor that mediates antiangiogenic and antitumor activity of endostatin. Blood 2007, 110, (8), 2899-2906.
150. Fluge, O.; Bruland, O.; Akslen, L. A.; Lillehaug, J. R.; Varhaug, J. E., Gene expression in poorly differentiated papillary thyroid carcinomas. Thyroid 2006, 16, (2), 161-175.
151. Fujarewicz, K.; Jarzab, M.; Eszlinger, M.; Krohn, K.; Paschke, R.; Oczko-Wojciechowska, M.; Wiench, M.; Kukulska, A.; Jarzab, B.; Swierniak, A., A multi-gene approach to differentiate papillary thyroid carcinoma from benign lesions: gene selection using support vector machines with bootstrapping. Endocr Relat Cancer 2007, 14, (3), 809-26.
152. Miyake, H.; Hara, I.; Yamanaka, K.; Gohji, K.; Arakawa, S.; Kamidono, S., Elevation of serum levels of urokinase-type plasminogen activator and its receptor is associated with disease progression and prognosis in patients with prostate cancer. Prostate 1999, 39, (2), 123-9.
153. Ghoneim, C.; Soula-Rothhut, M.; Blanchevoye, C.; Martiny, L.; Antonicelli, F.; Rothhut, B., Activating transcription factor-1-mediated hepatocyte growth factor-induced down-regulation of thrombospondin-1 expression leads to thyroid cancer cell invasion. J Biol Chem 2007, 282, (21), 15490-7.
154. Liu, Z.; Zhao, Q.; Han, Q.; Gao, M.; Zhang, N., Serum thrombospondin-1 is altered in patients with hemorrhagic fever with renal syndrome. J Med Virol 2008, 80, (10), 1799-803.
155. Vaisocherová, H.; Faca, V. M.; Taylor, A. D.; Hanash, S.; Jiang, S., Comparative study of SPR and ELISA methods based on analysis of CD166/ALCAM levels in cancer and control human sera. Biosensors and Bioelectronics 2009, 24, (7), 2143-2148.

77
156. Tegeder, I.; Schumacher, A.; John, S.; Geiger, H.; Geisslinger, G.; Bang, H.; Brune, K., Elevated Serum Cyclophilin Levels in Patients with Severe Sepsis. Journal of Clinical Immunology 1997, 17, (5), 380-386.
157. Sun, S.; Poon, R. T.; Lee, N. P.; Yeung, C.; Chan, K. L.; Ng, I. O.; Day, P. J.; Luk, J. M., Proteomics of hepatocellular carcinoma: serum vimentin as a surrogate marker for small tumors (<or=2 cm). J Proteome Res 2010, 9, (4), 1923-30.
158. Lassoued, S.; Mseddi, M.; Mnif, F.; Abid, M.; Guermazi, F.; Masmoudi, H.; El Feki, A.; Attia, H., A Comparative Study of the Oxidative Profile in Graves' Disease, Hashimoto's Thyroiditis, and Papillary Thyroid Cancer. Biol Trace Elem Res 2010.
159. Spranger, M.; Krempien, S.; Schwab, S.; Donneberg, S.; Hacke, W., Superoxide Dismutase Activity in Serum of Patients With Acute Cerebral Ischemic Injury : Correlation With Clinical Course and Infarct Size. Stroke 1997, 28, (12), 2425-2428.
160. Aldred, M. A.; Huang, Y.; Liyanarachchi, S.; Pellegata, N. S.; Gimm, O.; Jhiang, S.; Davuluri, R. V.; de la Chapelle, A.; Eng, C., Papillary and follicular thyroid carcinomas show distinctly different microarray expression profiles and can be distinguished by a minimum of five genes. J Clin Oncol 2004, 22, (17), 3531-9.
161. Baxter, R. C.; Saunders, H., Radioimmunoassay of insulin-like growth factor-binding protein-6 in human serum and other body fluids. Journal of Endocrinology 1992, 134, (1), 133-139.
162. Giusti, L.; Iacconi, P.; Ciregia, F.; Giannaccini, G.; Donatini, G. L.; Basolo, F.; Miccoli, P.; Pinchera, A.; Lucacchini, A., Fine-Needle Aspiration of Thyroid Nodules: Proteomic Analysis To Identify Cancer Biomarkers. Journal of Proteome Research 2008, 7, (9), 4079-4088.
163. Avilla, E.; Guarino, V.; Visciano, C.; Liotti, F.; Svelto, M.; Krishnamoorthy, G.; Franco, R.; Melillo, R. M., Activation of TYRO3/AXL Tyrosine Kinase Receptors in Thyroid Cancer. Cancer Research 2011, 71, (5), 1792-1804.
164. Ito, T.; Ito, M.; Naito, S.; Ohtsuru, A.; Nagayama, Y.; Kanematsu, T.; Yamashita, S.; Sekine, I., Expression of the Axl receptor tyrosine kinase in human thyroid carcinoma. Thyroid 1999, 9, (6), 563-7.
165. da Silveira Mitteldorf, C. A. T.; de Sousa-Canavez, J. M.; Massumoto, C.; da Camara-Lopes, L. H., Fine-needle aspiration biopsy of thyroid nodules as a possible source for molecular studies: Analysis of RNA obtained from routine cases. Diagnostic Cytopathology 2008, 36, (12), 899-903.
166. Bhardwaj, A.; Frankel, W. L.; Pellegata, N. S.; Wen, P.; Prasad, M. L., Intracellular Versican Expression in Mesenchymal Spindle Cell Tumors Contrasts With Extracellular Expression in Epithelial and Other Tumors-A Tissue Microarray-based Study. Applied Immunohistochemistry & Molecular Morphology 2008, 16, (3), 263-266 10.1097/PAI.0b013e31812cb5bc.

78
167. Wiseman, S.; Masoudi, H.; Niblock, P.; Turbin, D.; Rajput, A.; Hay, J.; Bugis, S.; Filipenko, D.; Huntsman, D.; Gilks, B., Anaplastic Thyroid Carcinoma: Expression Profile of Targets for Therapy Offers New Insights for Disease Treatment. Annals of Surgical Oncology 2007, 14, (2), 719-729.
168. Park, J.-W.; Yeh, M. W.; Wong, M. G.; Lobo, M.; Hyun, W. C.; Duh, Q.-Y.; Clark, O. H., The Heat Shock Protein 90-Binding Geldanamycin Inhibits Cancer Cell Proliferation, Down-Regulates Oncoproteins, and Inhibits Epidermal Growth Factor-Induced Invasion in Thyroid Cancer Cell Lines. J Clin Endocrinol Metab 2003, 88, (7), 3346-3353.
169. Saussez, S.; Glinoer, D.; Chantrain, G.; Pattou, F.; Carnaille, B.; Andre, S.; Gabius, H. J.; Laurent, G., Serum galectin-1 and galectin-3 levels in benign and malignant nodular thyroid disease. Thyroid 2008, 18, (7), 705-12.
170. Liang, H.; Zhong, Y.; Luo, Z.; Huang, Y.; Lin, H.; Luo, M.; Zhan, S.; Xie, K.; Ma, Y.; Li, Q. Q., Assessment of biomarkers for clinical diagnosis of papillary thyroid carcinoma with distant metastasis. Int J Biol Markers 2010, 25, (1), 38-45.
171. Puppin, C.; Fabbro, D.; Dima, M.; Di Loreto, C.; Puxeddu, E.; Filetti, S.; Russo, D.; Damante, G., High periostin expression correlates with aggressiveness in papillary thyroid carcinomas. J Endocrinol 2008, 197, (2), 401-8.
172. Ulisse, S.; Baldini, E.; Sorrenti, S.; Barollo, S.; Gnessi, L.; Catania, A.; Pellizzo, M. R.; Nardi, F.; Mian, C.; De Antoni, E.; D'Armiento, M.; Frati, L., High expression of the urokinase plasminogen activator and its cognate receptor associates with advanced stages and reduced disease-free interval in papillary thyroid carcinoma. J Clin Endocrinol Metab 2011, 96, (2), 504-8.
173. Tseleni-Balafouta, S.; Gakiopoulou, H.; Fanourakis, G.; Voutsinas, G.; Balafoutas, D.; Patsouris, E., Tenascin-c protein expression and mRNA splice variants in thyroid carcinoma. Experimental and Molecular Pathology 2006, 80, (2), 177-182.
174. LINCOLN, D. T.; AL-YATAMA, F.; MOHAMMED, F. M. A.; AL-BANAW, A. G.; AL-BADER, M.; BURGE, M.; SINOWATZ, F.; SINGAL, P. K., Thioredoxin and Thioredoxin Reductase Expression in Thyroid Cancer Depends on Tumour Aggressiveness. Anticancer Research 2010, 30, (3), 767-775.
175. Sid, B.; Langlois, B.; Sartelet, H.; Bellon, G.; Dedieu, S.; Martiny, L., Thrombospondin-1 enhances human thyroid carcinoma cell invasion through urokinase activity. The International Journal of Biochemistry & Cell Biology 2008, 40, (9), 1890-1900.
176. Ghoneim, C.; Soula-Rothhut, M.; Rothhut, B., Thrombospondin-1 in differentiated thyroid cancer: Dr. Jekyll and Mr. Hyde. Connect Tissue Res 2008, 49, (3), 257-60.
177. Sato, K.; Waseda, R.; Tatsuzawa, Y.; Soma, R.; Ueda, Y.; Katsuda, S., Papillary thyroid carcinoma with anaplastic transformation showing a rhabdoid phenotype solely in the cervical lymph node metastasis. Pathol Res Pract 2006, 202, (1), 55-9.

79
178. Watanabe, R.; Hayashi, Y.; Sassa, M.; Kikumori, T.; Imai, T.; Kiuchi, T.; Murata, Y., Possible involvement of BRAFV600E in altered gene expression in papillary thyroid cancer. Endocr J 2009, 56, (3), 407-14.
179. Horvatic Herceg, G.; Herceg, D.; Kralik, M.; Bence-Zigman, Z.; Tomic-Brzac, H.; Kulic, A., Urokinase-type plasminogen activator and its inhibitor in thyroid neoplasms: a cytosol study. Wien Klin Wochenschr 2006, 118, (19-20), 601-9.
180. Sainaghi, P. P.; Castello, L.; Bergamasco, L.; Galletti, M.; Bellosta, P.; Avanzi, G. C., Gas6 induces proliferation in prostate carcinoma cell lines expressing the Axl receptor. J Cell Physiol 2005, 204, (1), 36-44.
181. Gorsler, T.; Murzik, U.; Ulbricht, T.; Hentschel, J.; Hemmerich, P.; Melle, C., DNA damage-induced translocation of S100A11 into the nucleus regulates cell proliferation. BMC Cell Biology 2010, 11, (1), 100.
182. Farin, K.; Schokoroy, S.; Haklai, R.; Cohen-Or, I.; Elad-Sfadia, G.; Reyes-Reyes, M. E.; Bates, P. J.; Cox, A. D.; Kloog, Y.; Pinkas-Kramarski, R., Oncogenic Synergism between ErbB1, Nucleolin, and Mutant Ras. Cancer Research 2011, 71, (6), 2140-2151.
183. van Staveren, W. C. G.; Solís, D. W.; Delys, L.; Duprez, L.; Andry, G.; Franc, B.; Thomas, G.; Libert, F.; Dumont, J. E.; Detours, V.; Maenhaut, C., Human Thyroid Tumor Cell Lines Derived from Different Tumor Types Present a Common Dedifferentiated Phenotype. Cancer Research 2007, 67, (17), 8113-8120.
184. León-Del-Río, A., Biotin-dependent regulation of gene expression in human cells. The Journal of Nutritional Biochemistry 2005, 16, (7), 432-434.
185. Chen, C.-C.; Lau, L. F., Functions and mechanisms of action of CCN matricellular proteins. The International Journal of Biochemistry & Cell Biology 2009, 41, (4), 771-783.
186. Chen, Y.; Du, X. Y., Functional properties and intracellular signaling of CCN1/Cyr61. J Cell Biochem 2007, 100, (6), 1337-45.
187. Leask, A., A sticky situation: CCN1 promotes both proliferation and apoptosis of cancer cells. Journal of Cell Communication and Signaling 2009.
188. Watari, H.; Kanuma, T.; Ohta, Y.; Hassan, M. K.; Mitamura, T.; Hosaka, M.; Minegishi, T.; Sakuragi, N., Clusterin Expression Inversely Correlates with Chemosensitivity and Predicts Poor Survival in Patients with Locally Advanced Cervical Cancer Treated with Cisplatin-Based Neoadjuvant Chemotherapy and Radical Hysterectomy. Pathol Oncol Res 2010.
189. Zellweger, T.; Chi, K.; Miyake, H.; Adomat, H.; Kiyama, S.; Skov, K.; Gleave, M. E., Enhanced radiation sensitivity in prostate cancer by inhibition of the cell survival protein clusterin. Clin Cancer Res 2002, 8, (10), 3276-84.
190. Magnarelli, L. A.; Miller, J. N.; Anderson, J. F.; Riviere, G. R., Cross-reactivity of nonspecific treponemal antibody in serologic tests for Lyme disease. J. Clin. Microbiol. 1990, 28, (6), 1276-1279.

80
191. Marinova, I.; Nikolov, G.; Michova, A.; Kurdova, R.; Petrunov, B., Quantitative assessment of serum-specific IgE in the diagnosis of human cystic echinococcosis. Parasite Immunology 2011, 33, (7), 371-376.
192. Adav, S. S.; Li, A. A.; Manavalan, A.; Punt, P.; Sze, S. K., Quantitative iTRAQ secretome analysis of Aspergillus niger reveals novel hydrolytic enzymes. J Proteome Res 2010, 9, (8), 3932-40.
193. Planque, C.; Kulasingam, V.; Smith, C. R.; Reckamp, K.; Goodglick, L.; Diamandis, E. P., Identification of five candidate lung cancer biomarkers by proteomics analysis of conditioned media of four lung cancer cell lines. Mol Cell Proteomics 2009, 8, (12), 2746-58.
194. Makridakis, M.; Vlahou, A., Secretome proteomics for discovery of cancer biomarkers. Journal of Proteomics 2010, 73, (12), 2291-2305.
195. Ringel, M. D., "Thyroid cancer" cell line misidentification: a time for proactive change. J Clin Endocrinol Metab 2008, 93, (11), 4226-7.
196. Karagiannis, G. S.; Pavlou, M. P.; Diamandis, E. P., Cancer secretomics reveal pathophysiological pathways in cancer molecular oncology. Molecular Oncology 2010, 4, (6), 496-510.
197. Pavlou, M. P.; Diamandis, E. P., The cancer cell secretome: A good source for discovering biomarkers? Journal of Proteomics 2010, 73, (10), 1896-1906.
198. Vuoriluoto, K.; Haugen, H.; Kiviluoto, S.; Mpindi, J. P.; Nevo, J.; Gjerdrum, C.; Tiron, C.; Lorens, J. B.; Ivaska, J., Vimentin regulates EMT induction by Slug and oncogenic H-Ras and migration by governing Axl expression in breast cancer. Oncogene 2010.
199. Chang, K.-P.; Wu, C.-C.; Chen, H.-C.; Chen, S.-J.; Peng, P.-H.; Tsang, N.-M.; Lee, L.-Y.; Liu, S.-C.; Liang, Y.; Lee, Y.-S.; Hao, S.-P.; Chang, Y.-S.; Yu, J.-S., Identification of candidate nasopharyngeal carcinoma serum biomarkers by cancer cell secretome and tissue transcriptome analysis: Potential usage of cystatin A for predicting nodal stage and poor prognosis. Proteomics 2010, 10, (14), 2644-2660.
200. Schiarea, S.; Solinas, G.; Allavena, P.; Scigliuolo, G. M.; Bagnati, R.; Fanelli, R.; Chiabrando, C., Secretome Analysis of Multiple Pancreatic Cancer Cell Lines Reveals Perturbations of Key Functional Networks. Journal of Proteome Research 2010, 9, (9), 4376-4392.
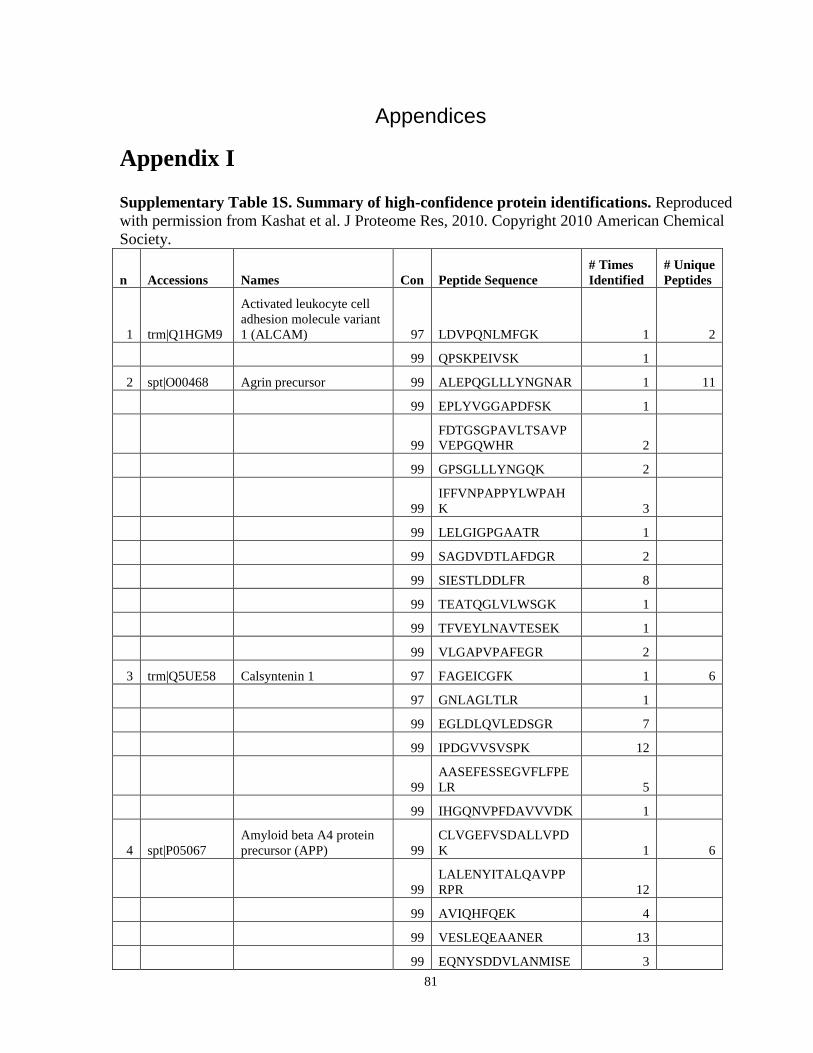
81
Appendices
Appendix I
Supplementary Table 1S. Summary of high-confidence protein identifications. Reproduced with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
1 trm|Q1HGM9
Activated leukocyte cell adhesion molecule variant 1 (ALCAM) 97 LDVPQNLMFGK 1 2
99 QPSKPEIVSK 1
2 spt|O00468 Agrin precursor 99 ALEPQGLLLYNGNAR 1 11
99 EPLYVGGAPDFSK 1
99 FDTGSGPAVLTSAVPVEPGQWHR 2
99 GPSGLLLYNGQK 2
99 IFFVNPAPPYLWPAHK 3
99 LELGIGPGAATR 1
99 SAGDVDTLAFDGR 2
99 SIESTLDDLFR 8
99 TEATQGLVLWSGK 1
99 TFVEYLNAVTESEK 1
99 VLGAPVPAFEGR 2
3 trm|Q5UE58 Calsyntenin 1 97 FAGEICGFK 1 6
97 GNLAGLTLR 1
99 EGLDLQVLEDSGR 7
99 IPDGVVSVSPK 12
99 AASEFESSEGVFLFPELR 5
99 IHGQNVPFDAVVVDK 1
4 spt|P05067 Amyloid beta A4 protein precursor (APP) 99
CLVGEFVSDALLVPDK 1 6
99 LALENYITALQAVPPRPR 12
99 AVIQHFQEK 4
99 VESLEQEAANER 13
99 EQNYSDDVLANMISE 3
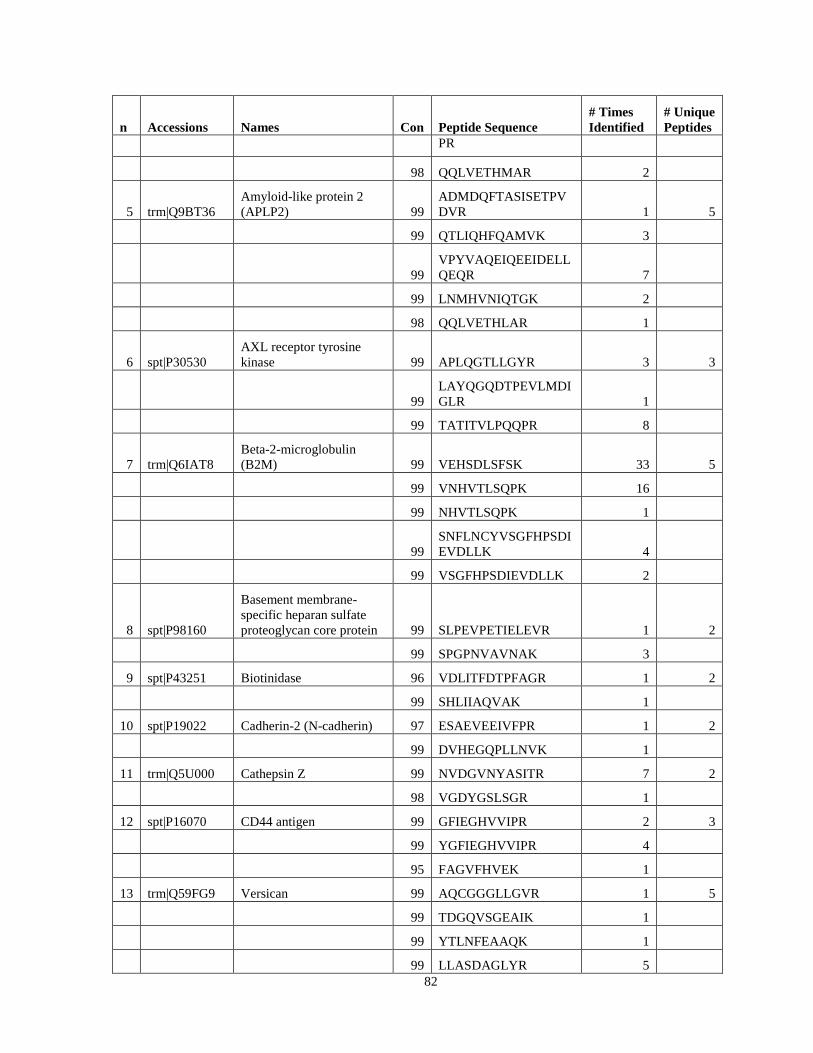
82
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
PR
98 QQLVETHMAR 2
5 trm|Q9BT36 Amyloid-like protein 2 (APLP2) 99
ADMDQFTASISETPVDVR 1 5
99 QTLIQHFQAMVK 3
99 VPYVAQEIQEEIDELLQEQR 7
99 LNMHVNIQTGK 2
98 QQLVETHLAR 1
6 spt|P30530 AXL receptor tyrosine kinase 99 APLQGTLLGYR 3 3
99 LAYQGQDTPEVLMDIGLR 1
99 TATITVLPQQPR 8
7 trm|Q6IAT8 Beta-2-microglobulin (B2M) 99 VEHSDLSFSK 33 5
99 VNHVTLSQPK 16
99 NHVTLSQPK 1
99 SNFLNCYVSGFHPSDIEVDLLK 4
99 VSGFHPSDIEVDLLK 2
8 spt|P98160
Basement membrane-specific heparan sulfate proteoglycan core protein 99 SLPEVPETIELEVR 1 2
99 SPGPNVAVNAK 3
9 spt|P43251 Biotinidase 96 VDLITFDTPFAGR 1 2
99 SHLIIAQVAK 1
10 spt|P19022 Cadherin-2 (N-cadherin) 97 ESAEVEEIVFPR 1 2
99 DVHEGQPLLNVK 1
11 trm|Q5U000 Cathepsin Z 99 NVDGVNYASITR 7 2
98 VGDYGSLSGR 1
12 spt|P16070 CD44 antigen 99 GFIEGHVVIPR 2 3
99 YGFIEGHVVIPR 4
95 FAGVFHVEK 1
13 trm|Q59FG9 Versican 99 AQCGGGLLGVR 1 5
99 TDGQVSGEAIK 1
99 YTLNFEAAQK 1
99 LLASDAGLYR 5
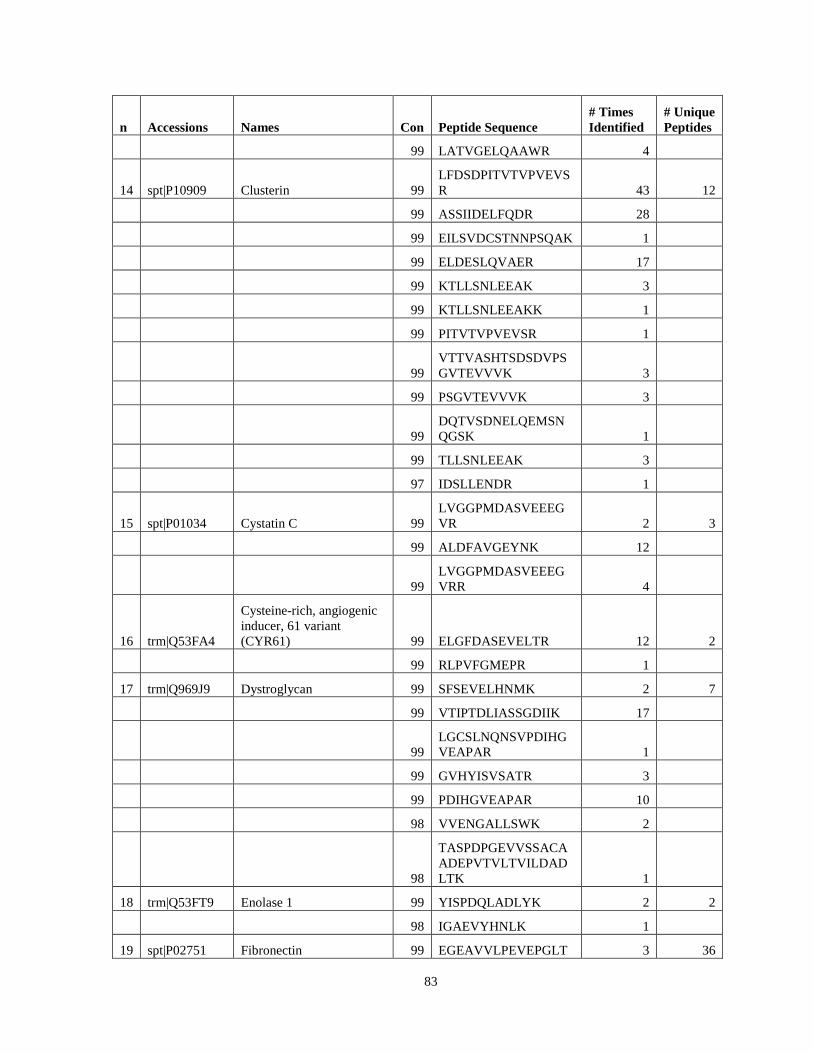
83
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
99 LATVGELQAAWR 4
14 spt|P10909 Clusterin 99 LFDSDPITVTVPVEVSR 43 12
99 ASSIIDELFQDR 28
99 EILSVDCSTNNPSQAK 1
99 ELDESLQVAER 17
99 KTLLSNLEEAK 3
99 KTLLSNLEEAKK 1
99 PITVTVPVEVSR 1
99 VTTVASHTSDSDVPSGVTEVVVK 3
99 PSGVTEVVVK 3
99 DQTVSDNELQEMSNQGSK 1
99 TLLSNLEEAK 3
97 IDSLLENDR 1
15 spt|P01034 Cystatin C 99 LVGGPMDASVEEEGVR 2 3
99 ALDFAVGEYNK 12
99 LVGGPMDASVEEEGVRR 4
16 trm|Q53FA4
Cysteine-rich, angiogenic inducer, 61 variant (CYR61) 99 ELGFDASEVELTR 12 2
99 RLPVFGMEPR 1
17 trm|Q969J9 Dystroglycan 99 SFSEVELHNMK 2 7
99 VTIPTDLIASSGDIIK 17
99 LGCSLNQNSVPDIHGVEAPAR 1
99 GVHYISVSATR 3
99 PDIHGVEAPAR 10
98 VVENGALLSWK 2
98
TASPDPGEVVSSACAADEPVTVLTVILDADLTK 1
18 trm|Q53FT9 Enolase 1 99 YISPDQLADLYK 2 2
98 IGAEVYHNLK 1
19 spt|P02751 Fibronectin 99 EGEAVVLPEVEPGLT 3 36
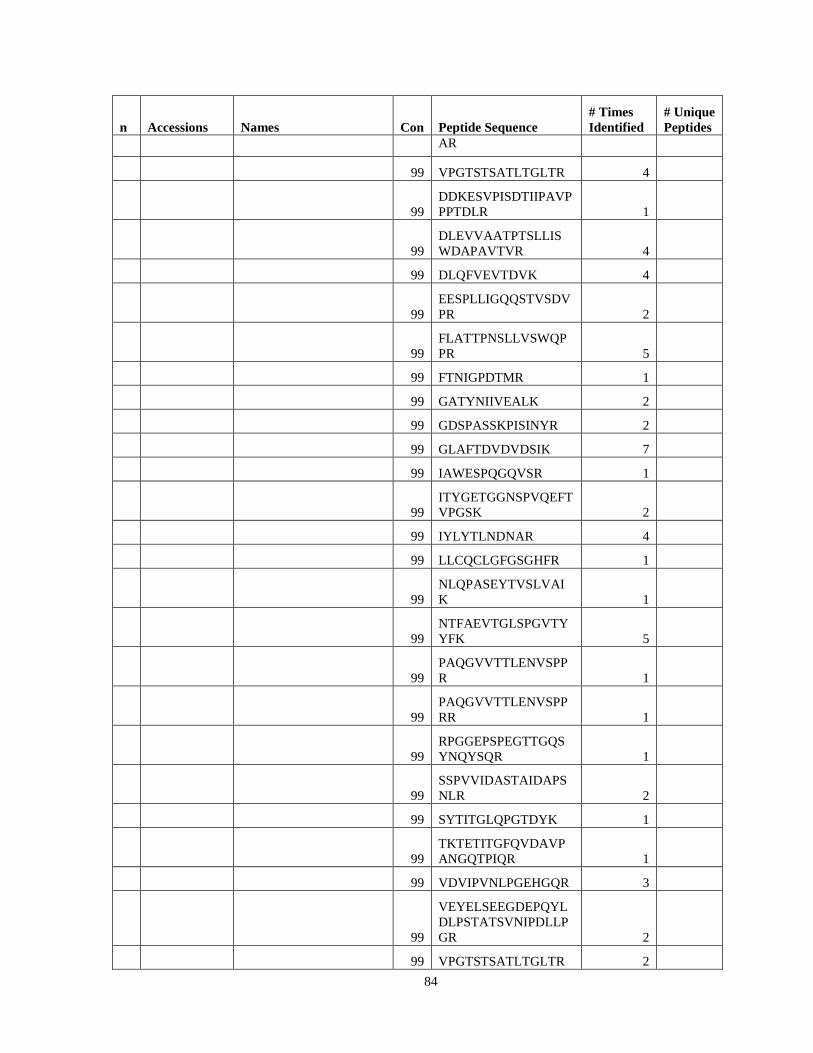
84
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
AR
99 VPGTSTSATLTGLTR 4
99 DDKESVPISDTIIPAVPPPTDLR 1
99 DLEVVAATPTSLLISWDAPAVTVR 4
99 DLQFVEVTDVK 4
99 EESPLLIGQQSTVSDVPR 2
99 FLATTPNSLLVSWQPPR 5
99 FTNIGPDTMR 1
99 GATYNIIVEALK 2
99 GDSPASSKPISINYR 2
99 GLAFTDVDVDSIK 7
99 IAWESPQGQVSR 1
99 ITYGETGGNSPVQEFTVPGSK 2
99 IYLYTLNDNAR 4
99 LLCQCLGFGSGHFR 1
99 NLQPASEYTVSLVAIK 1
99 NTFAEVTGLSPGVTYYFK 5
99 PAQGVVTTLENVSPPR 1
99 PAQGVVTTLENVSPPRR 1
99 RPGGEPSPEGTTGQSYNQYSQR 1
99 SSPVVIDASTAIDAPSNLR 2
99 SYTITGLQPGTDYK 1
99 TKTETITGFQVDAVPANGQTPIQR 1
99 VDVIPVNLPGEHGQR 3
99
VEYELSEEGDEPQYLDLPSTATSVNIPDLLPGR 2
99 VPGTSTSATLTGLTR 2

85
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
99 VTIMWTPPESAVTGYR 4
99 VTWAPPPSIDLTNFLVR 22
99 VVTPLSPPTNLHLEANPDTGVLTVSWER 9
99 WLPSSSPVTGYR 3
97 GLAFTDVDVDSIK 3
97 HYQINQQWER 1
97 STATISGLKPGVDYTITVYAVTGR 1
96 VTDATETTITISWR 56
96 GEWTCIAYSQLR 1
96 SLLVSWQPPR 1
20 spt|Q08380 Galectin-3 binding protein 99 ELSEALGQIFDSQR 2 9
99 ASHEEVEGLVEK 2
99 AVDTWSWGER 2
99 ELSEALGQIFDSQR 3
99 IYTSPTWSAFVTDSSWSAR 3
99 SDLAVPSELALLK 4
99 TLQALEFHTVPFQLLAR 19
99 YYPYQSFQTPQHPSFLFQDK 1
96 GQWGTVCDNLWDLTDASVVCR 1
21 spt|Q92820 Gamma-glutamyl hydrolase 99 FFNVLTTNTDGK 2 4
99 NLDGISHAPNAVK 1
99 YPVYGVQWHPEK 1
99 KPIIGILMQK 1
22 spt|P28799 Granulins (proepithelin) 99 APAHLSLPDPQALK 1 2
97 EVVSAQPATFLAR 1
23 trm|Q8WVW5 Putative uncharacterized protein 99 HQGVMVGMGQK 1 3
99 SYELPDGQVITIGNER 5
96 GIHETTFNSIMK 1

86
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
24 spt|P24592
Insulin-like growth factor-binding protein 6 (IGFBP-6) 98 APAVAEENPK 1 5
99 ITVVDALHEIPVK 1
99 TELLPGDRDNLAIQTR 2
99 HEVTGWVLVSPLSK 2
99 VVDALHEIPVK 1
25 trm|Q92626 Melanoma-associated antigen 99 AEGNPKPEIIWLR 1 3
99 SPNDLLALFR 3
96 IVNEGGIDPLLR 1
26 trm|Q53HV3 Lysyl oxidase-like 2 variant 99
LGQGIGPIHLNEIQCTGNEK 1 2
99 TPVMEGYVEVK 3
27 spt|P16035
Metalloproteinase inhibitor 2 precursor (TIMP-2) 99 EVDSGNDIYGNPIK 2 2
99 GAAPPKQEFLDIEDP 1
28 spt|P14543 Nidogen-1 precursor (Entactin) 99
QELFPFGPGQGDLELEDGDDFVSPALELSGALR 1 2
95 QDLGSPEGIAVDHLGR 1
29 trm|Q53GX6 Nucleobindin 1 99 LPEVEVPQHL 1 2
99 LVTLEEFLASTQR 1
30 spt|P19338 Nucleolin (Protein C23) 99 GFGFVDFNSEEDAK 2 6
99 FGYVDFESAEDLEK 2.0
97 NDLAVVDVR 1.0
99 SISLYYTGEK 1.0
99 TLVLSNLSYSATEETLQEVFEK 6.0
99 VEGTEPTTAFNLFVGNLNFNK 4.0
31 spt|P06748 Nucleophosmin (NPM) 99 MSVQPTVSLGGFEITPPVVLR 1 2
99 GPSSVEDIK 1
32 spt|P10451 Osteopontin 99 AIPVAQDLNAPSDWDSR 9 3
99 YPDAVATWLNPDPSQ 11

87
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
K
99 ISHELDSASSEVN 1
33 trm|Q3KQW3 Peptidylprolyl isomerase A (Cyclophilin A) 99 VSFELFADK 1 2
98 EGMNIVEAMER 1
34 trm|Q5SWW9 Plasminogen activator, urokinase activator 96 SHTKEENGLAL 1 9
98 SDALQLGLGK 9
99 FEVENLILHK 6
99 IIGGEFTTIENQPWFAAIYR 12
99 KEDYIVYLGR 2
99 KPSSPPEELK 11
99 MTLTGIVSWGR 16
99 DYSADTLAHHNDIALLK 1
99 SADTLAHHNDIALLK 1
35 trm|Q9NYD3 Prothymosin-alpha (PTMA) 99 EVVEEAENGR 1 1
36 trm|Q6PQ81 Dickkopf-related protein 3 (DKK-3) 98 DQDGEILLPR 6 2
98 LLDLITWELEPDGALDR 1
37 trm|Q6FHZ5 SET protein 95 EQQEAIEHIDEVQNEIDR 1.0 5
99 IDFYFDENPYFENK 3.0
96
IPNFWVTTFVNHPQVSALLGEEDEEALHYLTR 1.0
99 LNEQASEEILK 1.0
99 VEVTEFEDIK 2.0
38 trm|Q5U0B9
Stem cell growth factor; lymphocyte secreted C-type lectin 99
AALAPYNWPVWLGVHDR 10 4
99 DAVQALQEAQGR 2
99 DFEAQAAAQAR 1
99 HLQEALGLPAGR 3
39 trm|Q6NR85 Superoxide dismutase [Cu-Zn] 99 GDGPVQGIINFEQK 4 2

88
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
99 HVGDLGNVTADK 2
40 spt|P31431 Syndecan-4 99 AGSGSQVPTEPK 34 4
99 ETEVIDPQDLLEGR 23
99 ISPVEESEDVSNK 26
99 KLEENEVIPK 8
41 trm|Q59E99 Thrombospondin 1 variant (Fragment) 99 AGTLDLSLTVQGK 22 15
99 DHSGQVFSVVSNGK 1
99 FVFGTTPEDILR 7
99 GGVNDNFQGVLQNVR 2
99 GPDPSSPAFR 1
99 IEDANLIPPVPDDK 1
99 IEDANLIPPVPDDKFQDLVDAVR 7
99 IPESGGDNSVFDIFELTGAAR 13
99 MENAELDVPIQSVFTR 2
99 NRIPESGGDNSVFDIFELTGAAR 79
99 PPVPDDKFQDLVDAVR 1
99 QHVVSVEEALLATGQWK 20
99 QVTQSYWDTNPTR 1
99 TIVTTLQDSIR 6
97 FQDLVDAVR 1
42 trm|Q5H9A7 Metalloproteinase inhibitor 1(TIMP-1) 99 GFQALGDAADIR 24 7
99 SFVAPWNSLSLAQR 4
98 SEEFLIAGK 1
99 LQDGLLHITTCSFVAPWNSLSLAQR 5
99 EPGLCTWQSLR 1
99 FVYTPAMESVCGYFHR 2
99 LQSGTHCLWTDQLLQGSEK 3
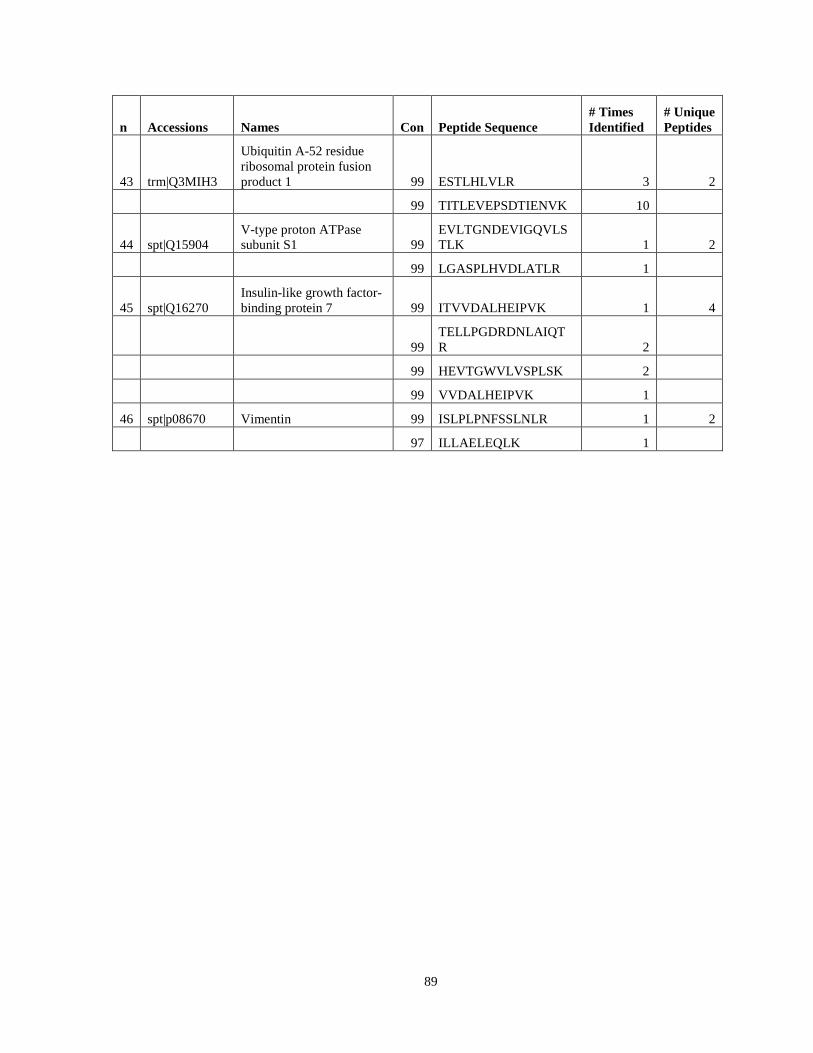
89
n Accessions Names Con Peptide Sequence # Times Identified
# Unique Peptides
43 trm|Q3MIH3
Ubiquitin A-52 residue ribosomal protein fusion product 1 99 ESTLHLVLR 3 2
99 TITLEVEPSDTIENVK 10
44 spt|Q15904 V-type proton ATPase subunit S1 99
EVLTGNDEVIGQVLSTLK 1 2
99 LGASPLHVDLATLR 1
45 spt|Q16270 Insulin-like growth factor-binding protein 7 99 ITVVDALHEIPVK 1 4
99 TELLPGDRDNLAIQTR 2
99 HEVTGWVLVSPLSK 2
99 VVDALHEIPVK 1
46 spt|p08670 Vimentin 99 ISLPLPNFSSLNLR 1 2
97 ILLAELEQLK 1
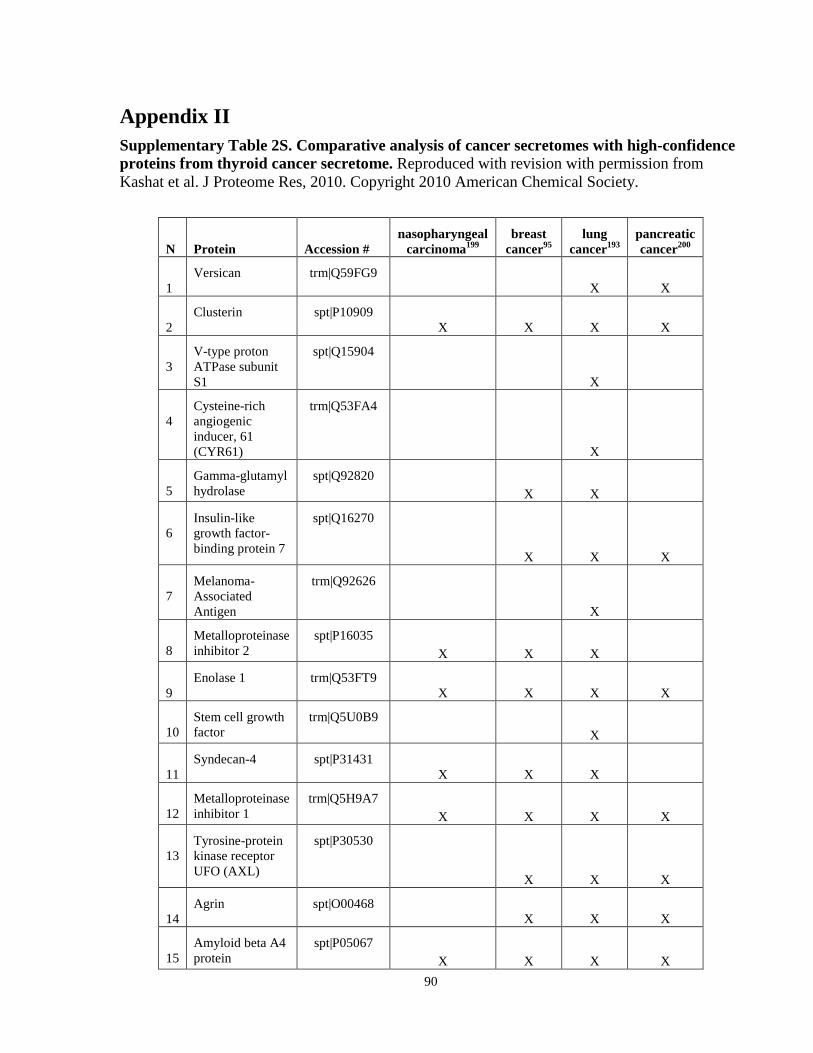
90
Appendix II Supplementary Table 2S. Comparative analysis of cancer secretomes with high-confidence proteins from thyroid cancer secretome. Reproduced with revision with permission from Kashat et al. J Proteome Res, 2010. Copyright 2010 American Chemical Society.
N Protein Accession # nasopharyngeal
carcinoma199 breast
cancer95 lung
cancer193 pancreatic cancer200
1 Versican trm|Q59FG9
X X
2 Clusterin spt|P10909
X X X X
3 V-type proton ATPase subunit S1
spt|Q15904
X
4 Cysteine-rich angiogenic inducer, 61 (CYR61)
trm|Q53FA4
X
5 Gamma-glutamyl hydrolase
spt|Q92820 X X
6 Insulin-like growth factor-binding protein 7
spt|Q16270
X X X
7 Melanoma-Associated Antigen
trm|Q92626
X
8 Metalloproteinase inhibitor 2
spt|P16035 X X X
9 Enolase 1 trm|Q53FT9
X X X X
10 Stem cell growth factor
trm|Q5U0B9 X
11 Syndecan-4 spt|P31431
X X X
12 Metalloproteinase inhibitor 1
trm|Q5H9A7 X X X X
13 Tyrosine-protein kinase receptor UFO (AXL)
spt|P30530
X X X
14 Agrin spt|O00468
X X X
15 Amyloid beta A4 protein
spt|P05067 X X X X
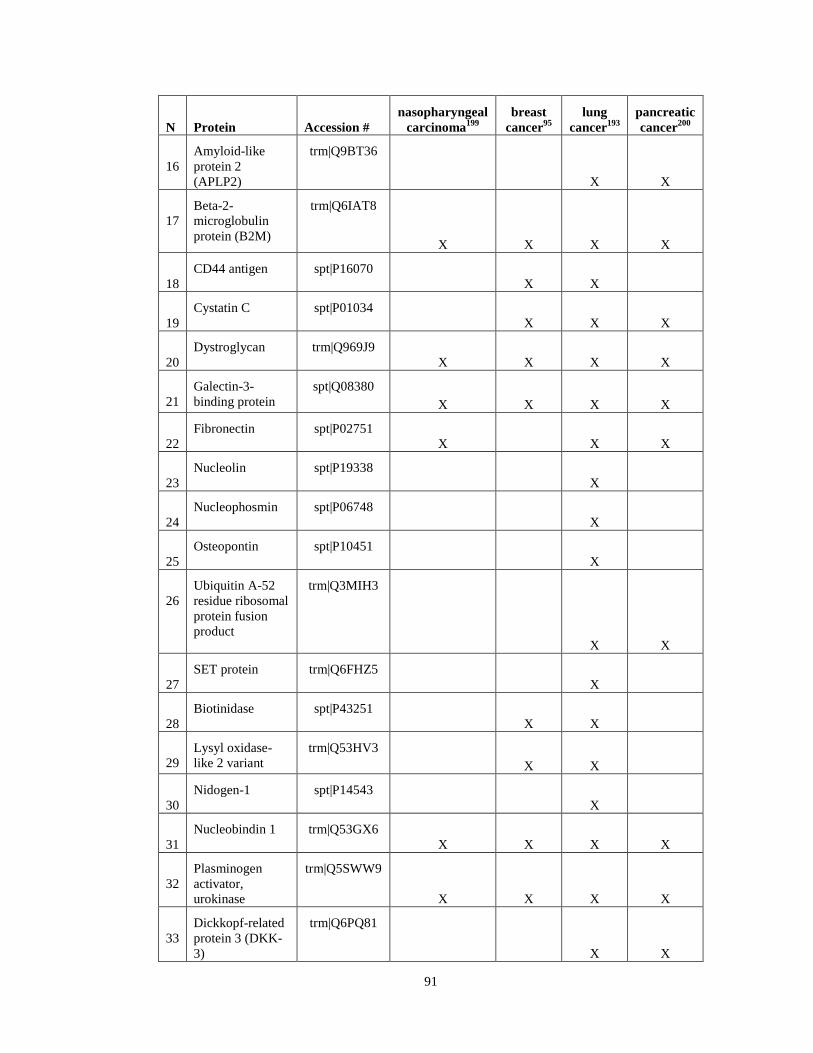
91
N Protein Accession # nasopharyngeal
carcinoma199 breast
cancer95 lung
cancer193 pancreatic cancer200
16 Amyloid-like protein 2 (APLP2)
trm|Q9BT36
X X
17 Beta-2-microglobulin protein (B2M)
trm|Q6IAT8
X X X X
18 CD44 antigen spt|P16070
X X
19 Cystatin C spt|P01034
X X X
20 Dystroglycan trm|Q969J9
X X X X
21 Galectin-3-binding protein
spt|Q08380 X X X X
22 Fibronectin spt|P02751
X X X
23 Nucleolin spt|P19338
X
24 Nucleophosmin spt|P06748
X
25 Osteopontin spt|P10451
X
26 Ubiquitin A-52 residue ribosomal protein fusion product
trm|Q3MIH3
X X
27 SET protein trm|Q6FHZ5
X
28 Biotinidase spt|P43251
X X
29 Lysyl oxidase-like 2 variant
trm|Q53HV3 X X
30 Nidogen-1 spt|P14543
X
31 Nucleobindin 1 trm|Q53GX6
X X X X
32 Plasminogen activator, urokinase
trm|Q5SWW9
X X X X
33 Dickkopf-related protein 3 (DKK-3)
trm|Q6PQ81
X X

92
N Protein Accession # nasopharyngeal
carcinoma199 breast
cancer95 lung
cancer193 pancreatic cancer200
34 Thrombospondin 1
trm|Q59E99 X X X X
35 Calsyntenin-1 trm|Q5UE58
X X X
36 Basement Membrane Specific Heparan Sulfate Core Protein
spt|P98160
X X
37 Prothymosin-α (PTMA)d
trm|Q9NYD3
X
38 Cadherin-2 (N-Cadherin)
spt|P19022 X X
39 Granulins (proepithelin)
spt|P28799 X X
40 Activated leukocyte cell adhesion molecule (ALCAM)
trm|Q1HGM9
X X X
41 Peptidylproyl isomerase A (cyclophilin A)
trm|Q3KQW3
X
42 Vimentin spt|P08670
X X X
43 Cathepsin Z trm|Q5U000
X X
44 Superoxide dismutase
trm|Q6NR85 X X X
45 Putative uncharacterized protein
trm|Q8WVW5
46 Insulin-like growth factor-binding protein 6 (IGFBP-6)
spt|P24592
X X
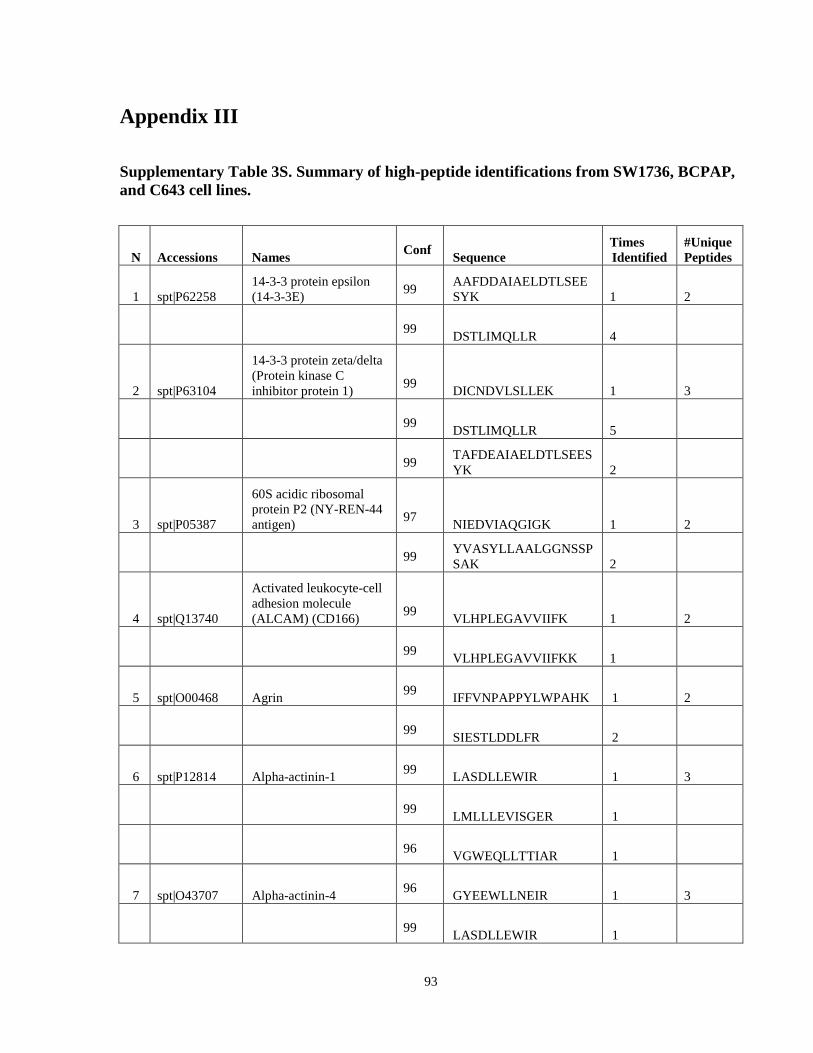
93
Appendix III
Supplementary Table 3S. Summary of high-peptide identifications from SW1736, BCPAP, and C643 cell lines.
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
1 spt|P62258 14-3-3 protein epsilon (14-3-3E) 99 AAFDDAIAELDTLSEE
SYK 1 2
99 DSTLIMQLLR 4
2 spt|P63104
14-3-3 protein zeta/delta (Protein kinase C inhibitor protein 1) 99 DICNDVLSLLEK 1 3
99 DSTLIMQLLR 5
99 TAFDEAIAELDTLSEESYK 2
3 spt|P05387
60S acidic ribosomal protein P2 (NY-REN-44 antigen) 97 NIEDVIAQGIGK 1 2
99 YVASYLLAALGGNSSPSAK 2
4 spt|Q13740
Activated leukocyte-cell adhesion molecule (ALCAM) (CD166) 99 VLHPLEGAVVIIFK 1 2
99 VLHPLEGAVVIIFKK 1
5 spt|O00468 Agrin 99 IFFVNPAPPYLWPAHK 1 2
99 SIESTLDDLFR 2
6 spt|P12814 Alpha-actinin-1 99 LASDLLEWIR 1 3
99 LMLLLEVISGER 1
96 VGWEQLLTTIAR 1
7 spt|O43707 Alpha-actinin-4 96 GYEEWLLNEIR 1 3
99 LASDLLEWIR 1
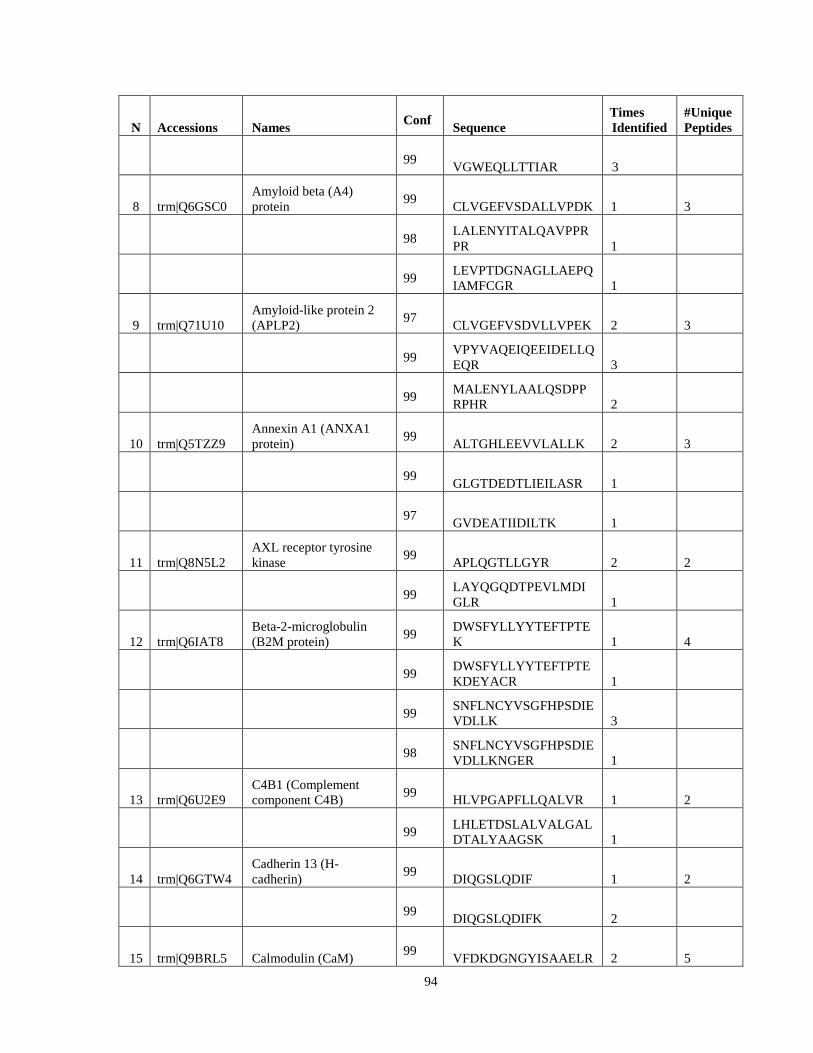
94
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 VGWEQLLTTIAR 3
8 trm|Q6GSC0
Amyloid beta (A4) protein 99 CLVGEFVSDALLVPDK 1 3
98 LALENYITALQAVPPRPR 1
99 LEVPTDGNAGLLAEPQIAMFCGR 1
9 trm|Q71U10
Amyloid-like protein 2 (APLP2) 97 CLVGEFVSDVLLVPEK 2 3
99 VPYVAQEIQEEIDELLQEQR 3
99 MALENYLAALQSDPPRPHR 2
10 trm|Q5TZZ9
Annexin A1 (ANXA1 protein) 99 ALTGHLEEVVLALLK 2 3
99 GLGTDEDTLIEILASR 1
97 GVDEATIIDILTK 1
11 trm|Q8N5L2
AXL receptor tyrosine kinase 99 APLQGTLLGYR 2 2
99 LAYQGQDTPEVLMDIGLR 1
12 trm|Q6IAT8
Beta-2-microglobulin (B2M protein) 99 DWSFYLLYYTEFTPTE
K 1 4
99 DWSFYLLYYTEFTPTEKDEYACR 1
99 SNFLNCYVSGFHPSDIEVDLLK 3
98 SNFLNCYVSGFHPSDIEVDLLKNGER 1
13 trm|Q6U2E9
C4B1 (Complement component C4B) 99 HLVPGAPFLLQALVR 1 2
99 LHLETDSLALVALGALDTALYAAGSK 1
14 trm|Q6GTW4
Cadherin 13 (H-cadherin) 99 DIQGSLQDIF 1 2
99 DIQGSLQDIFK 2
15 trm|Q9BRL5 Calmodulin (CaM) 99 VFDKDGNGYISAAELR 2 5

95
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 EAFSLFDKDGDGTITTK 2
99 EADIDGDGQVNYEEFVQMMTAK 2
98 HVMTNLGEKLTDEEVDEMIR 1
99 TIDFPEFLTMMAR 1
16 trm|Q53G71 Calreticulin (CRP55) 99 FYALSASFEPFSNK 2 4
99 GQTLVVQFTVK 1
98
GTWIHPEIDNPEYSPDPSIYAYDNFGVLGLDLWQVK 1
99 SGTIFDNFLITNDEAYAEEFGNETWGVTK 3
17 trm|Q8N4K9 Calsyntenin 1 97 GNLAGLTLR 2 5
99 AASEFESSEGVFLFPELR 4
98 GVQIQAHPSQLVLTLEGEDLGELDK 1
99 FAESFEVTVTK 1
99 LIFLFR 1
18 trm|Q8WY99 Cathepsin C 97 ILHLPTSWDWR 1 2
99 LELVHHGPMAVAFEVYDDFLHYK 1
19 trm|Q6ZS99
CDNA FLJ45706 fis, clone FEBRA2028457, highly similar to Nucleolin 99 TLVLSNLSYSATEETL
QEVFEK 2 3
99 VEGTEPTTAFNLFVGNLNFNK 1
99 VTQDELKEVFEDAAEIR 1
20 trm|Q9UNM1
Chaperonin 10-related protein 99 VLQATVVAVGSGSK 2 2
99 VVLDDKDYFLFR 1

96
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
21 trm|Q59FG9
Chondroitin sulfate proteoglycan 2 (Versican) 99
ACLDVGAVIATPEQLFAAYEDGFEQCDAGWLADQTVR 3 4
99 LATVGELQAAWR 2
98 SPQETYDVYCYVDHLDGDVFHLTVPSK 1
99 VSVPTHPEAVGDASLTVVK 1
22 spt|P10909 Clusterin 99 ASSIIDELFQDR 2 2
99 LFDSDPITVTVPVEVSR 2
23 spt|P23528 Cofilin-1 99 KEDLVFIFWAPESAPL
K 3 4
99 LGGSAVISLEGKPL 3
99 NIILEEGKEILVGDVGQTVDDPYATFVK 6
99 YALYDATYETK 1
24 spt|P20908
Collagen alpha-1(V) chain 96 GPQGPAGRDGLQGPV
GL 1 6
99 ILDEEVFEGDIQQLLFVSDHR 2
99 LLSYVDAEGNPVGVVQMTFLR 1
99 QLYPASAFPEDFSILTTVK 1
99 SPVFLYEDHTGKPGPEDYPLFR 2
99 VLDFHNLPDGITK 2
25 spt|P12109
Collagen alpha-1(VI) chain 99 AVAFQDCPVDLFFVLD
TSESVALR 1 4
99 DAEEAISQTIDTIVDMIK 1
99 IALVITDGR 1
99 VPSYQALLR 1
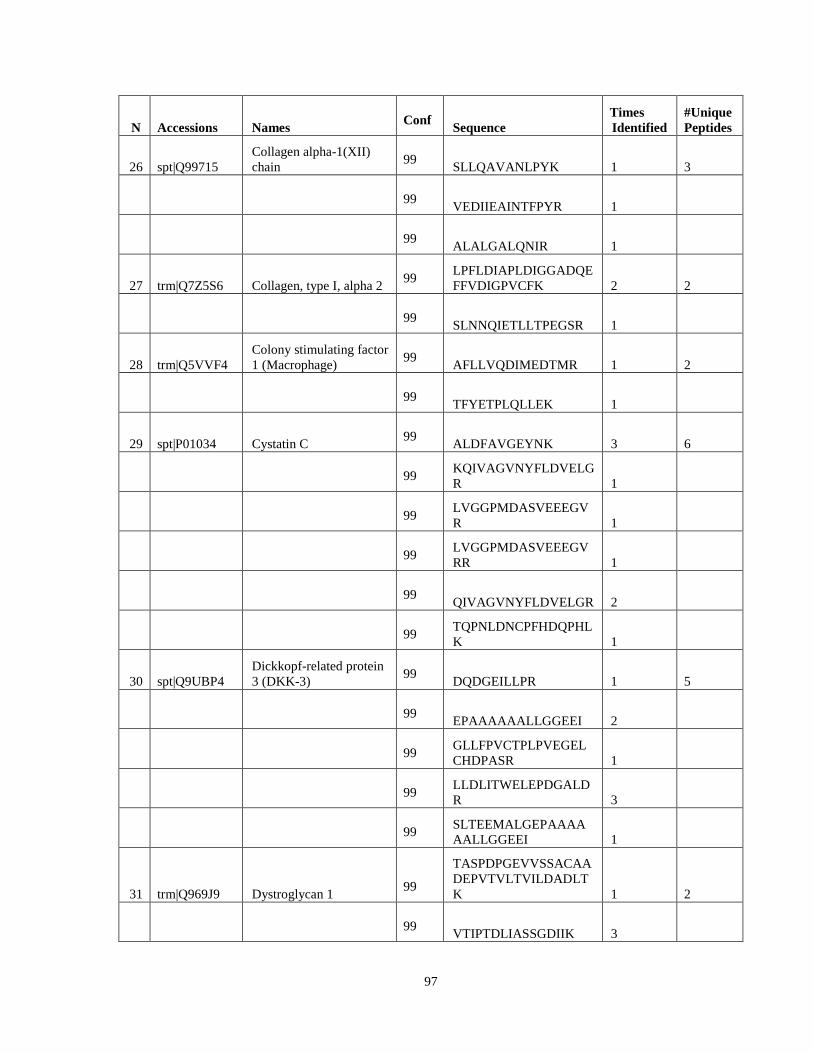
97
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
26 spt|Q99715 Collagen alpha-1(XII) chain 99 SLLQAVANLPYK 1 3
99 VEDIIEAINTFPYR 1
99 ALALGALQNIR 1
27 trm|Q7Z5S6 Collagen, type I, alpha 2 99 LPFLDIAPLDIGGADQE
FFVDIGPVCFK 2 2
99 SLNNQIETLLTPEGSR 1
28 trm|Q5VVF4
Colony stimulating factor 1 (Macrophage) 99 AFLLVQDIMEDTMR 1 2
99 TFYETPLQLLEK 1
29 spt|P01034 Cystatin C 99 ALDFAVGEYNK 3 6
99 KQIVAGVNYFLDVELGR 1
99 LVGGPMDASVEEEGVR 1
99 LVGGPMDASVEEEGVRR 1
99 QIVAGVNYFLDVELGR 2
99 TQPNLDNCPFHDQPHLK 1
30 spt|Q9UBP4
Dickkopf-related protein 3 (DKK-3) 99 DQDGEILLPR 1 5
99 EPAAAAAALLGGEEI 2
99 GLLFPVCTPLPVEGELCHDPASR 1
99 LLDLITWELEPDGALDR 3
99 SLTEEMALGEPAAAAAALLGGEEI 1
31 trm|Q969J9 Dystroglycan 1 99
TASPDPGEVVSSACAADEPVTVLTVILDADLTK 1 2
99 VTIPTDLIASSGDIIK 3
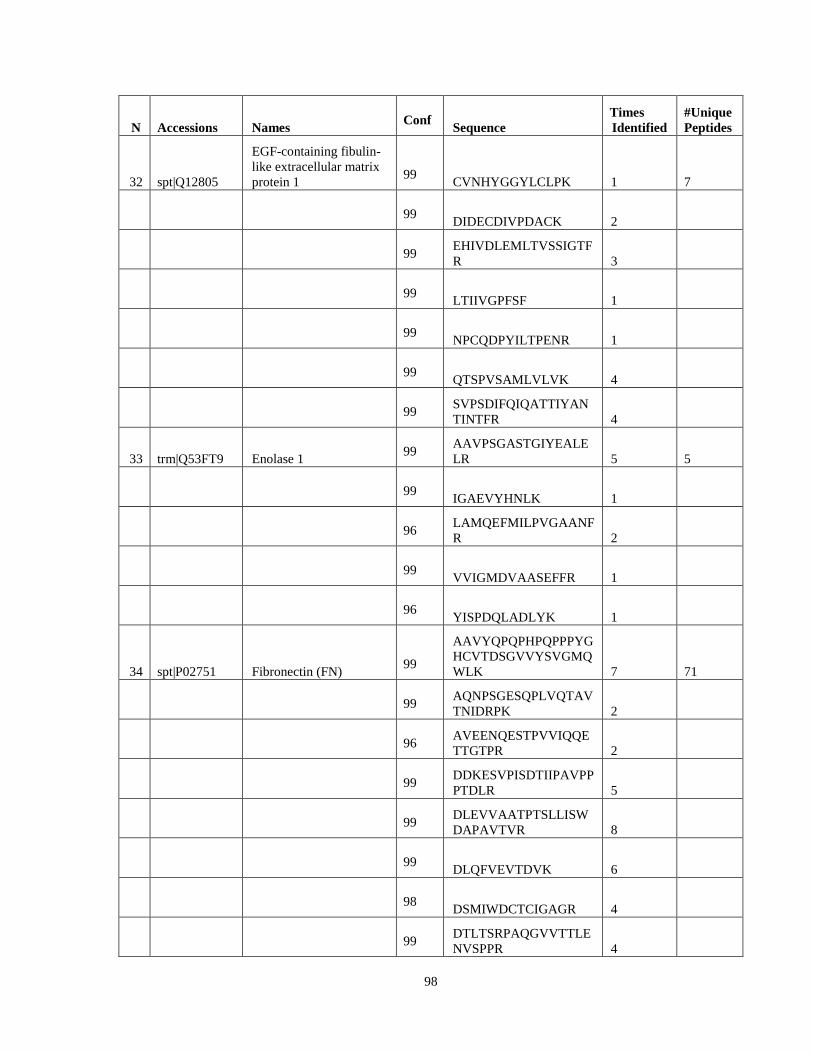
98
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
32 spt|Q12805
EGF-containing fibulin-like extracellular matrix protein 1 99 CVNHYGGYLCLPK 1 7
99 DIDECDIVPDACK 2
99 EHIVDLEMLTVSSIGTFR 3
99 LTIIVGPFSF 1
99 NPCQDPYILTPENR 1
99 QTSPVSAMLVLVK 4
99 SVPSDIFQIQATTIYANTINTFR 4
33 trm|Q53FT9 Enolase 1 99 AAVPSGASTGIYEALE
LR 5 5
99 IGAEVYHNLK 1
96 LAMQEFMILPVGAANFR 2
99 VVIGMDVAASEFFR 1
96 YISPDQLADLYK 1
34 spt|P02751 Fibronectin (FN) 99
AAVYQPQPHPQPPPYGHCVTDSGVVYSVGMQWLK 7 71
99 AQNPSGESQPLVQTAVTNIDRPK 2
96 AVEENQESTPVVIQQETTGTPR 2
99 DDKESVPISDTIIPAVPPPTDLR 5
99 DLEVVAATPTSLLISWDAPAVTVR 8
99 DLQFVEVTDVK 6
98 DSMIWDCTCIGAGR 4
99 DTLTSRPAQGVVTTLENVSPPR 4
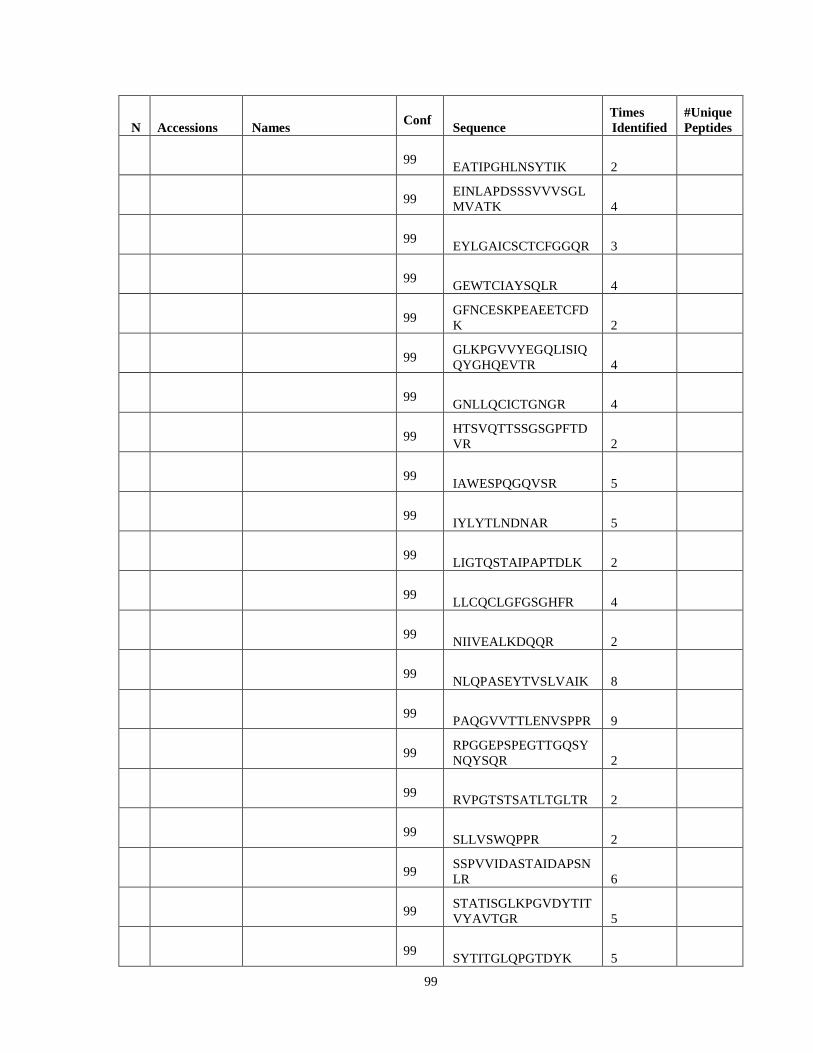
99
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 EATIPGHLNSYTIK 2
99 EINLAPDSSSVVVSGLMVATK 4
99 EYLGAICSCTCFGGQR 3
99 GEWTCIAYSQLR 4
99 GFNCESKPEAEETCFDK 2
99 GLKPGVVYEGQLISIQQYGHQEVTR 4
99 GNLLQCICTGNGR 4
99 HTSVQTTSSGSGPFTDVR 2
99 IAWESPQGQVSR 5
99 IYLYTLNDNAR 5
99 LIGTQSTAIPAPTDLK 2
99 LLCQCLGFGSGHFR 4
99 NIIVEALKDQQR 2
99 NLQPASEYTVSLVAIK 8
99 PAQGVVTTLENVSPPR 9
99 RPGGEPSPEGTTGQSYNQYSQR 2
99 RVPGTSTSATLTGLTR 2
99 SLLVSWQPPR 2
99 SSPVVIDASTAIDAPSNLR 6
99 STATISGLKPGVDYTITVYAVTGR 5
99 SYTITGLQPGTDYK 5

100
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 TGLDSPTGIDFSDITANSFTVHWIAPR 6
99 TYLGNALVCTCYGGSR 8
99 VDVIPVNLPGEHGQR 5
99 VEYELSEEGDEPQYLDLPSTATSVNIPDLLPGR 3
99
VEYELSEEGDEPQYLDLPSTATSVNIPDLLPGRK 3
99 VPGTSTSATLTGLTR 9
99 VTIMWTPPESAVTGYR 6
99 AAHEEICTTNEGVMYR 1
99 EESPLLIGQQSTVSDVPR 4
99 ESVPISDTIIPAVPPPTDLR 1
99 EVTSDSGSIVVSGLTPGVEYVYTIQVLR 3
99 FGFCPMAAHEEICTTNEGVMYR 1
99 FLATTPNSLLVSWQPPR 9
99 FTNIGPDTMR 2
99 FTQVTPTSLSAQWTPPNVQLTGYR 6
99 GATYNIIVEALK 11
99 GATYNIIVEALKDQQR 10
99 GLAFTDVDVDSIK 5
99 ITYGETGGNSPVQEFTVPGSK 4
99 KTDELPQLVTLPHPNLHGPEILDVPSTVQK 5
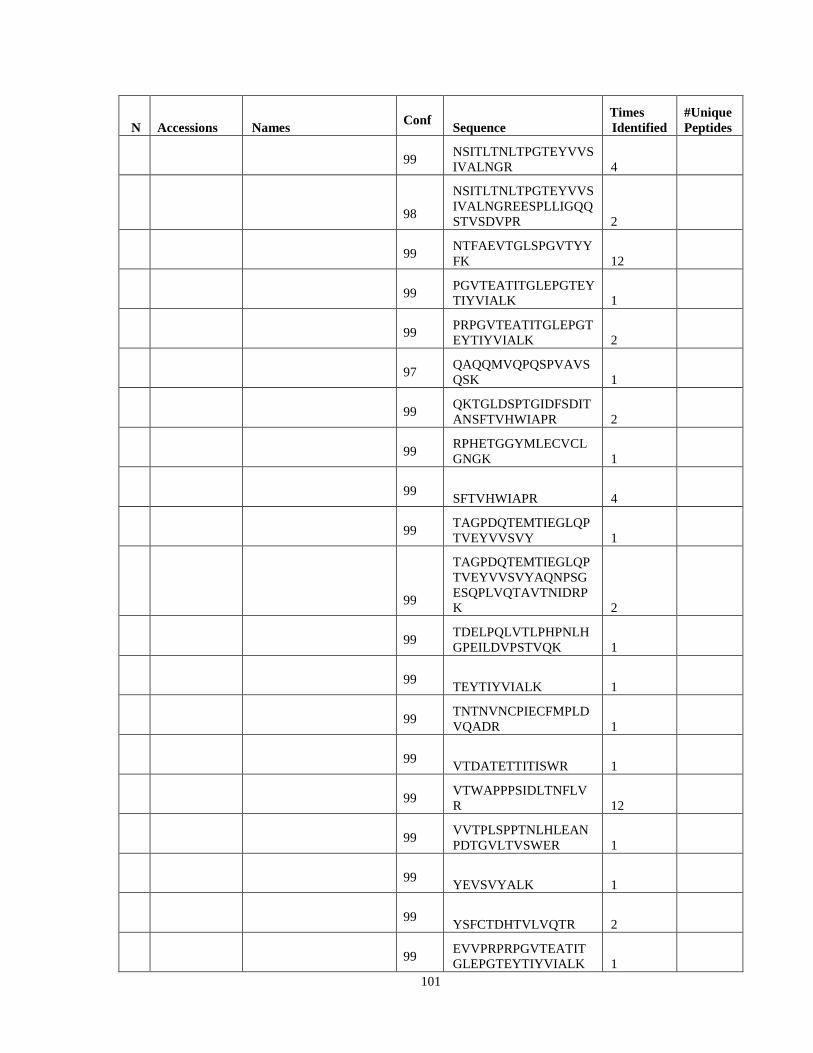
101
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 NSITLTNLTPGTEYVVSIVALNGR 4
98
NSITLTNLTPGTEYVVSIVALNGREESPLLIGQQSTVSDVPR 2
99 NTFAEVTGLSPGVTYYFK 12
99 PGVTEATITGLEPGTEYTIYVIALK 1
99 PRPGVTEATITGLEPGTEYTIYVIALK 2
97 QAQQMVQPQSPVAVSQSK 1
99 QKTGLDSPTGIDFSDITANSFTVHWIAPR 2
99 RPHETGGYMLECVCLGNGK 1
99 SFTVHWIAPR 4
99 TAGPDQTEMTIEGLQPTVEYVVSVY 1
99
TAGPDQTEMTIEGLQPTVEYVVSVYAQNPSGESQPLVQTAVTNIDRPK 2
99 TDELPQLVTLPHPNLHGPEILDVPSTVQK 1
99 TEYTIYVIALK 1
99 TNTNVNCPIECFMPLDVQADR 1
99 VTDATETTITISWR 1
99 VTWAPPPSIDLTNFLVR 12
99 VVTPLSPPTNLHLEANPDTGVLTVSWER 1
99 YEVSVYALK 1
99 YSFCTDHTVLVQTR 2
99 EVVPRPRPGVTEATITGLEPGTEYTIYVIALK 1

102
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 LVQTAVTNIDRPK 1
99 WLPSSSPVTGYR 1
35 trm|Q60FE6 Filamin A 99 SPFPLEAVAPTKPSK 1 2
99 LIALLEVLSQK 1
36 spt|Q12841
Follistatin-related protein 1 99 IIQWLEAEIIPDGWFSK 2 2
99 RIIQWLEAEIIPDGWFSK 1
37 trm|Q6FI10
Fructose-bisphosphate aldolase 99 ALANSLACQGK 1 2
96 GILAADESTGSIAK 1
38 spt|Q08380
Galectin-3-binding protein (Mac-2-binding protein) 99 AVDTWSWGER 2 7
99 ELSEALGQIFDSQR 4
99 GQWGTVCDNLWDLTDASVVCR 2
99 IYTSPTWSAFVTDSSWSAR 1
99 SDLAVPSELALLK 4
99 TLQALEFHTVPFQLLAR 4
99 YSSDYFQAPSDYR 1
39 spt|P06744
Glucose-6-phosphate isomerase 99 ILLANFLAQTEALMR 3 2
99 TLAQLNPESSLFIIASK 1
40 trm|Q5CAQ7
Heat shock protein HSP 90-alpha 2 99 ADLINNLGTIAK 1 3
99 APFDLFENR 1
99 DQVANSAFVER 1

103
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
41 trm|Q5T6W2
Heterogeneous nuclear ribonucleoprotein K (HNRNP K) 99 IILDLISESPIK 1 3
99 ILSISADIETIGEILK 1
99 IITITGTQDQIQNAQYLLQNSVK 1
42 trm|Q0JVC4 Galectin-1 98 DSNNLCLHFNPR 2 3
99 LNLEAINYMAADGDFK 4
99 SFVLNLGK 1
43 trm|Q53T11 Secretogranin-2 99 AGTEALPDGLSVEDIL
NLLGMESAANQK 3 7
99 ALEYIENLR 1
99 LFEKPLDSQSIYQLIEISR 3
99 NLQIPPEDLIEMLK 1
99 QMAYENLNDKDQELGEYLAR 1
98 QYWDEDLLMK 1
99 TNEIVEEQYTPQSLATLESVFQELGK 3
44 spt|Q16270
Insulin-like growth factor-binding protein 7 (IGFBP-7) 95 GGPEKHEVTGWVLVS
PLSK 1 5
99 HEVTGWVLVSPLSK 2
99 ITVVDALHEIPVK 2
97 ITVVDALHEIPVKK 1
99 TELLPGDRDNLAIQTR 1
45
trm|Q92626|Q92626_HUMAN
Melanoma-associated antigen MG50 (KIAA0230 protein) 99 LGPTLMCLLSTQFK 1 2
99 SPNDLLALFR 2
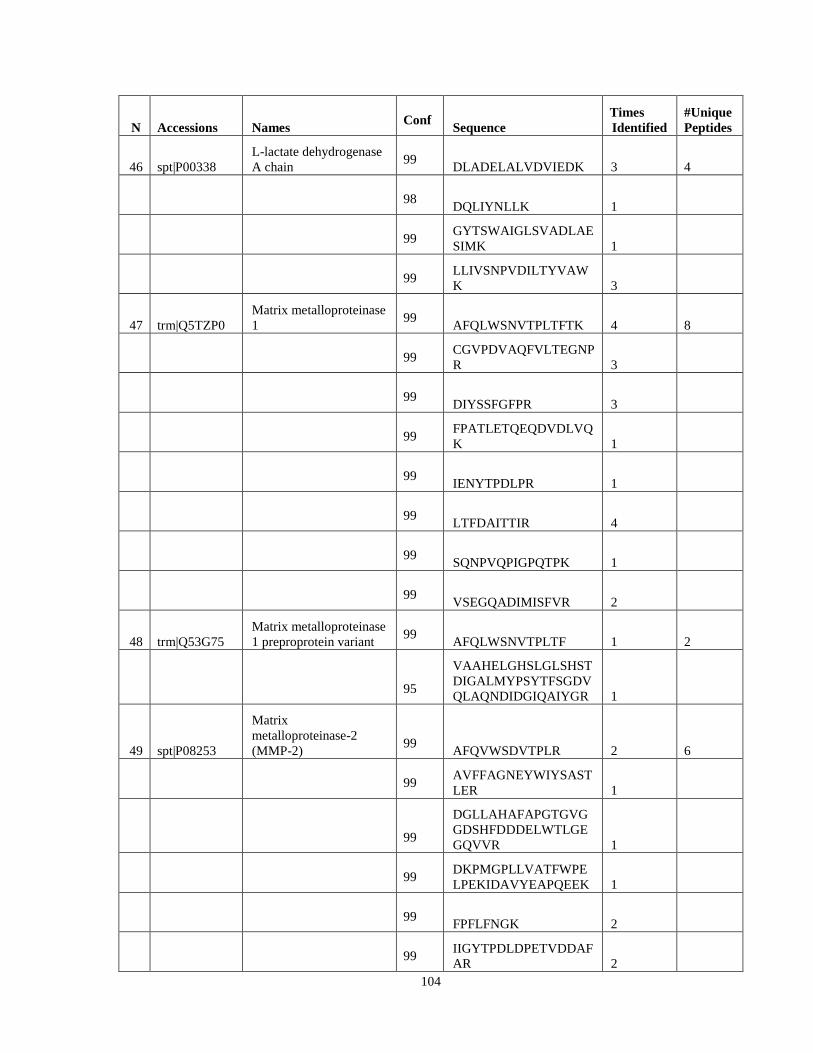
104
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
46 spt|P00338 L-lactate dehydrogenase A chain 99 DLADELALVDVIEDK 3 4
98 DQLIYNLLK 1
99 GYTSWAIGLSVADLAESIMK 1
99 LLIVSNPVDILTYVAWK 3
47 trm|Q5TZP0
Matrix metalloproteinase 1 99 AFQLWSNVTPLTFTK 4 8
99 CGVPDVAQFVLTEGNPR 3
99 DIYSSFGFPR 3
99 FPATLETQEQDVDLVQK 1
99 IENYTPDLPR 1
99 LTFDAITTIR 4
99 SQNPVQPIGPQTPK 1
99 VSEGQADIMISFVR 2
48 trm|Q53G75
Matrix metalloproteinase 1 preproprotein variant 99 AFQLWSNVTPLTF 1 2
95
VAAHELGHSLGLSHSTDIGALMYPSYTFSGDVQLAQNDIDGIQAIYGR 1
49 spt|P08253
Matrix metalloproteinase-2 (MMP-2) 99 AFQVWSDVTPLR 2 6
99 AVFFAGNEYWIYSASTLER 1
99
DGLLAHAFAPGTGVGGDSHFDDDELWTLGEGQVVR 1
99 DKPMGPLLVATFWPELPEKIDAVYEAPQEEK 1
99 FPFLFNGK 2
99 IIGYTPDLDPETVDDAFAR 2
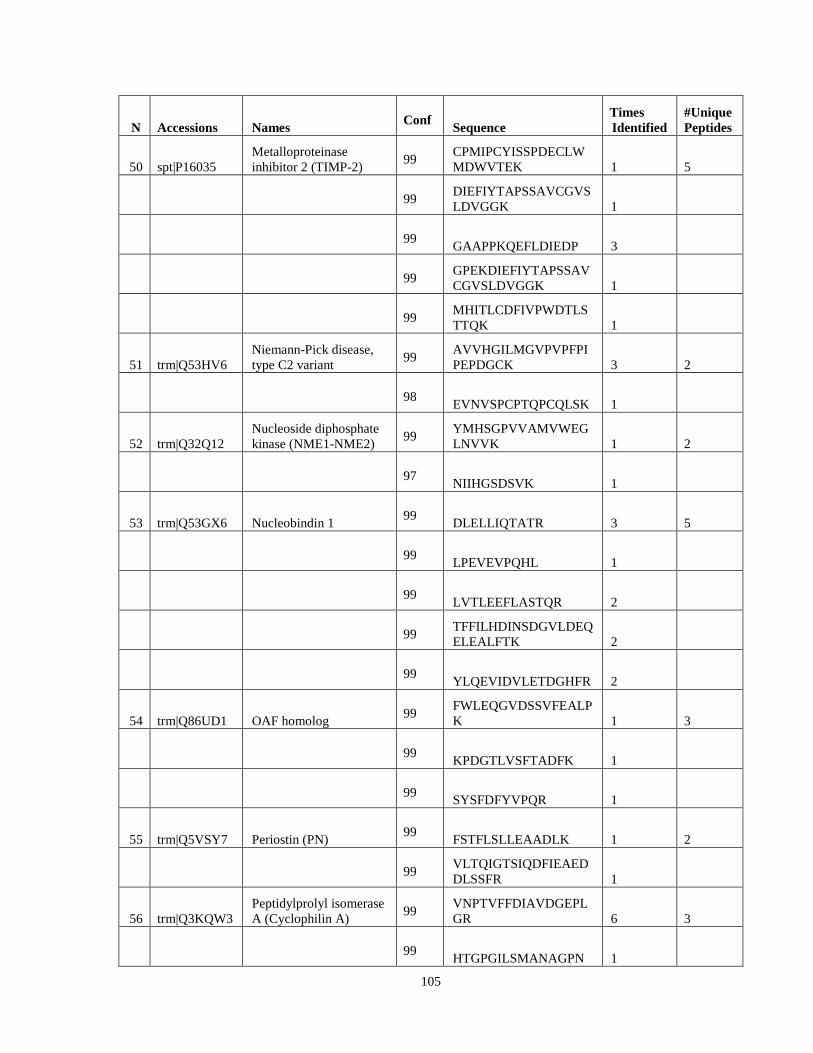
105
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
50 spt|P16035 Metalloproteinase inhibitor 2 (TIMP-2) 99 CPMIPCYISSPDECLW
MDWVTEK 1 5
99 DIEFIYTAPSSAVCGVSLDVGGK 1
99 GAAPPKQEFLDIEDP 3
99 GPEKDIEFIYTAPSSAVCGVSLDVGGK 1
99 MHITLCDFIVPWDTLSTTQK 1
51 trm|Q53HV6
Niemann-Pick disease, type C2 variant 99 AVVHGILMGVPVPFPI
PEPDGCK 3 2
98 EVNVSPCPTQPCQLSK 1
52 trm|Q32Q12
Nucleoside diphosphate kinase (NME1-NME2) 99 YMHSGPVVAMVWEG
LNVVK 1 2
97 NIIHGSDSVK 1
53 trm|Q53GX6 Nucleobindin 1 99 DLELLIQTATR 3 5
99 LPEVEVPQHL 1
99 LVTLEEFLASTQR 2
99 TFFILHDINSDGVLDEQELEALFTK 2
99 YLQEVIDVLETDGHFR 2
54 trm|Q86UD1 OAF homolog 99 FWLEQGVDSSVFEALP
K 1 3
99 KPDGTLVSFTADFK 1
99 SYSFDFYVPQR 1
55 trm|Q5VSY7 Periostin (PN) 99 FSTFLSLLEAADLK 1 2
99 VLTQIGTSIQDFIEAEDDLSSFR 1
56 trm|Q3KQW3
Peptidylprolyl isomerase A (Cyclophilin A) 99 VNPTVFFDIAVDGEPL
GR 6 3
99 HTGPGILSMANAGPN 1
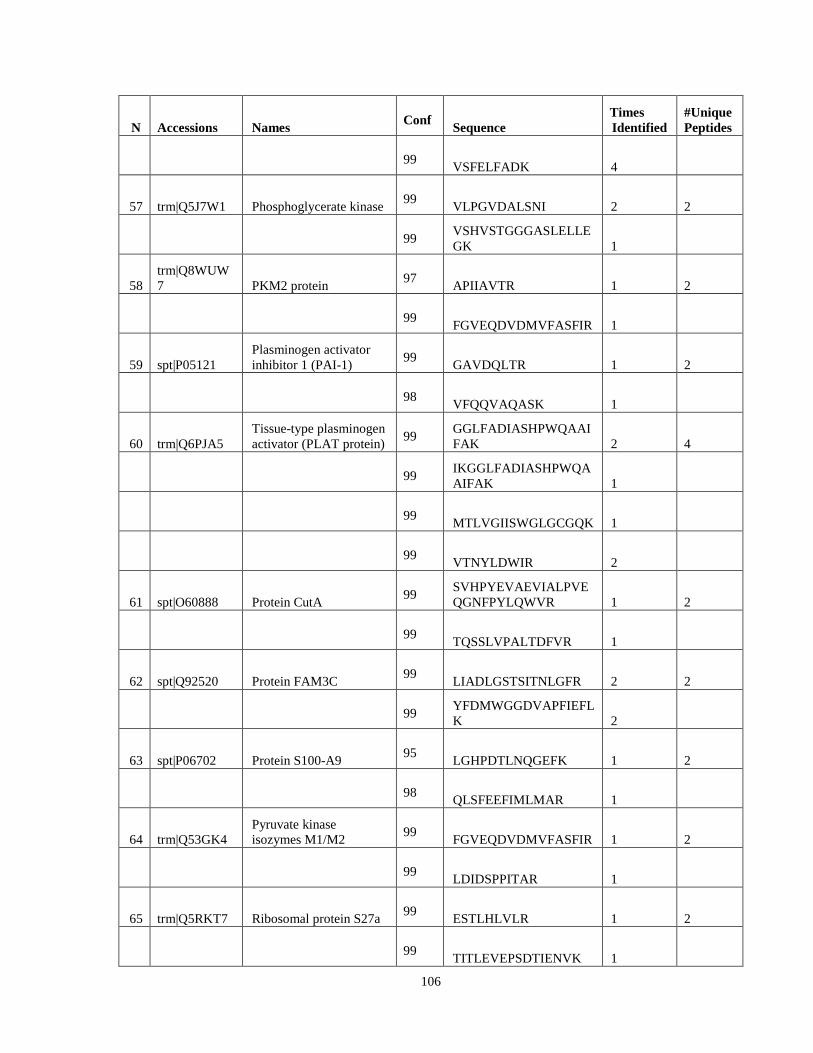
106
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 VSFELFADK 4
57 trm|Q5J7W1 Phosphoglycerate kinase 99 VLPGVDALSNI 2 2
99 VSHVSTGGGASLELLEGK 1
58
trm|Q8WUW7 PKM2 protein 97 APIIAVTR 1 2
99 FGVEQDVDMVFASFIR 1
59 spt|P05121
Plasminogen activator inhibitor 1 (PAI-1) 99 GAVDQLTR 1 2
98 VFQQVAQASK 1
60 trm|Q6PJA5
Tissue-type plasminogen activator (PLAT protein) 99 GGLFADIASHPWQAAI
FAK 2 4
99 IKGGLFADIASHPWQAAIFAK 1
99 MTLVGIISWGLGCGQK 1
99 VTNYLDWIR 2
61 spt|O60888 Protein CutA 99 SVHPYEVAEVIALPVE
QGNFPYLQWVR 1 2
99 TQSSLVPALTDFVR 1
62 spt|Q92520 Protein FAM3C 99 LIADLGSTSITNLGFR 2 2
99 YFDMWGGDVAPFIEFLK 2
63 spt|P06702 Protein S100-A9 95 LGHPDTLNQGEFK 1 2
98 QLSFEEFIMLMAR 1
64 trm|Q53GK4
Pyruvate kinase isozymes M1/M2 99 FGVEQDVDMVFASFIR 1 2
99 LDIDSPPITAR 1
65 trm|Q5RKT7 Ribosomal protein S27a 99 ESTLHLVLR 1 2
99 TITLEVEPSDTIENVK 1
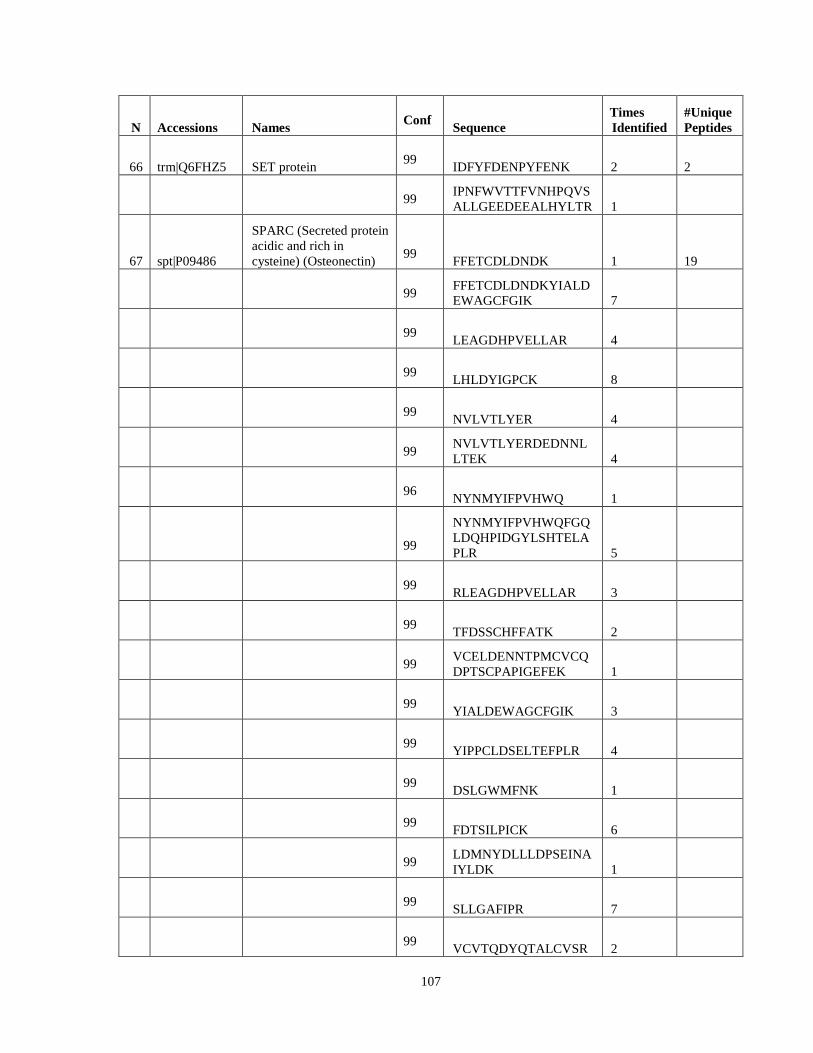
107
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
66 trm|Q6FHZ5 SET protein 99 IDFYFDENPYFENK 2 2
99 IPNFWVTTFVNHPQVSALLGEEDEEALHYLTR 1
67 spt|P09486
SPARC (Secreted protein acidic and rich in cysteine) (Osteonectin) 99 FFETCDLDNDK 1 19
99 FFETCDLDNDKYIALDEWAGCFGIK 7
99 LEAGDHPVELLAR 4
99 LHLDYIGPCK 8
99 NVLVTLYER 4
99 NVLVTLYERDEDNNLLTEK 4
96 NYNMYIFPVHWQ 1
99
NYNMYIFPVHWQFGQLDQHPIDGYLSHTELAPLR 5
99 RLEAGDHPVELLAR 3
99 TFDSSCHFFATK 2
99 VCELDENNTPMCVCQDPTSCPAPIGEFEK 1
99 YIALDEWAGCFGIK 3
99 YIPPCLDSELTEFPLR 4
99 DSLGWMFNK 1
99 FDTSILPICK 6
99 LDMNYDLLLDPSEINAIYLDK 1
99 SLLGAFIPR 7
99 VCVTQDYQTALCVSR 2

108
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
95 LKDWFGALHEDANR 1
68 trm|Q5U0B9
Stem cell growth factor (SCGF) 99 AALAPYNWPVWLGV
HDR 1 3
99 DAVQALQEAQGR 1
96 LAGLDAGLHQLHVR 1
69 spt|O00391
Sulfhydryl oxidase 1 (Quiescin Q6) (hQSOX) 99 IYMADLESALHYILR 1 2
99 LDVPVWDVEATLNFLK 1
70 trm|Q6NR85
Superoxide dismutase [Cu-Zn] 99 DGVADVSIEDSVISLSG
DHCIIGR 1 3
99 GDGPVQGIINFEQK 2
99
HVGDLGNVTADKDGVADVSIEDSVISLSGDHCIIGR 1
71 trm|Q5T7S3 Tenascin C 99 AATPYTVSIYGVIQGY
R 2 17
99 AVDIPGLEAATPYR 1
99 DLTATEVQSETALLTWRPPR 2
99 DVTDTTALITWFKPLAEIDGIELTYGIK 1
99
GHSTRPLAVEVVTEDLPQLGDLAVSEVGWDGLR 2
99 GLEPGQEYNVLLTAEK 1
99 LEELENLVSSLR 2
99 LIPGVEYLVSIIAMK 3
99 LSWTADEGVFDNFVLK 2
99 PLAVEVVTEDLPQLGDLAVSEVGWDGLR 1
99 QSEPLEITLLAPER 1
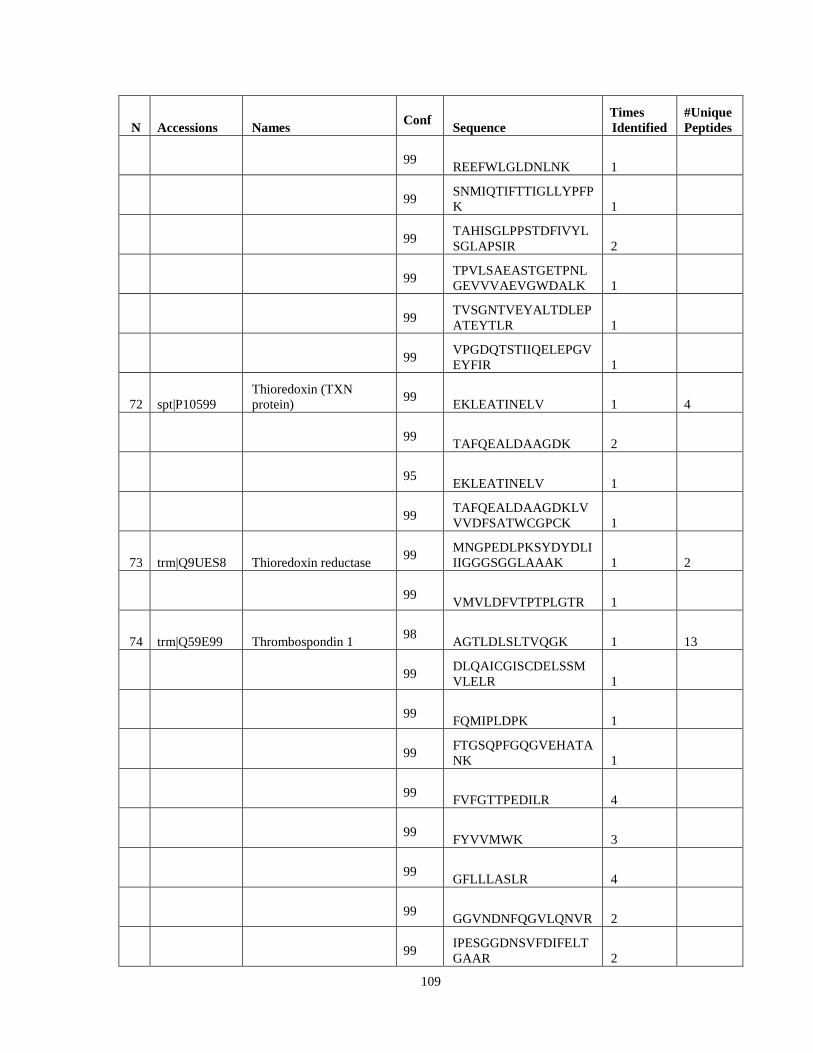
109
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 REEFWLGLDNLNK 1
99 SNMIQTIFTTIGLLYPFPK 1
99 TAHISGLPPSTDFIVYLSGLAPSIR 2
99 TPVLSAEASTGETPNLGEVVVAEVGWDALK 1
99 TVSGNTVEYALTDLEPATEYTLR 1
99 VPGDQTSTIIQELEPGVEYFIR 1
72 spt|P10599
Thioredoxin (TXN protein) 99 EKLEATINELV 1 4
99 TAFQEALDAAGDK 2
95 EKLEATINELV 1
99 TAFQEALDAAGDKLVVVDFSATWCGPCK 1
73 trm|Q9UES8 Thioredoxin reductase 99 MNGPEDLPKSYDYDLI
IIGGGSGGLAAAK 1 2
99 VMVLDFVTPTPLGTR 1
74 trm|Q59E99 Thrombospondin 1 98 AGTLDLSLTVQGK 1 13
99 DLQAICGISCDELSSMVLELR 1
99 FQMIPLDPK 1
99 FTGSQPFGQGVEHATANK 1
99 FVFGTTPEDILR 4
99 FYVVMWK 3
99 GFLLLASLR 4
99 GGVNDNFQGVLQNVR 2
99 IPESGGDNSVFDIFELTGAAR 2
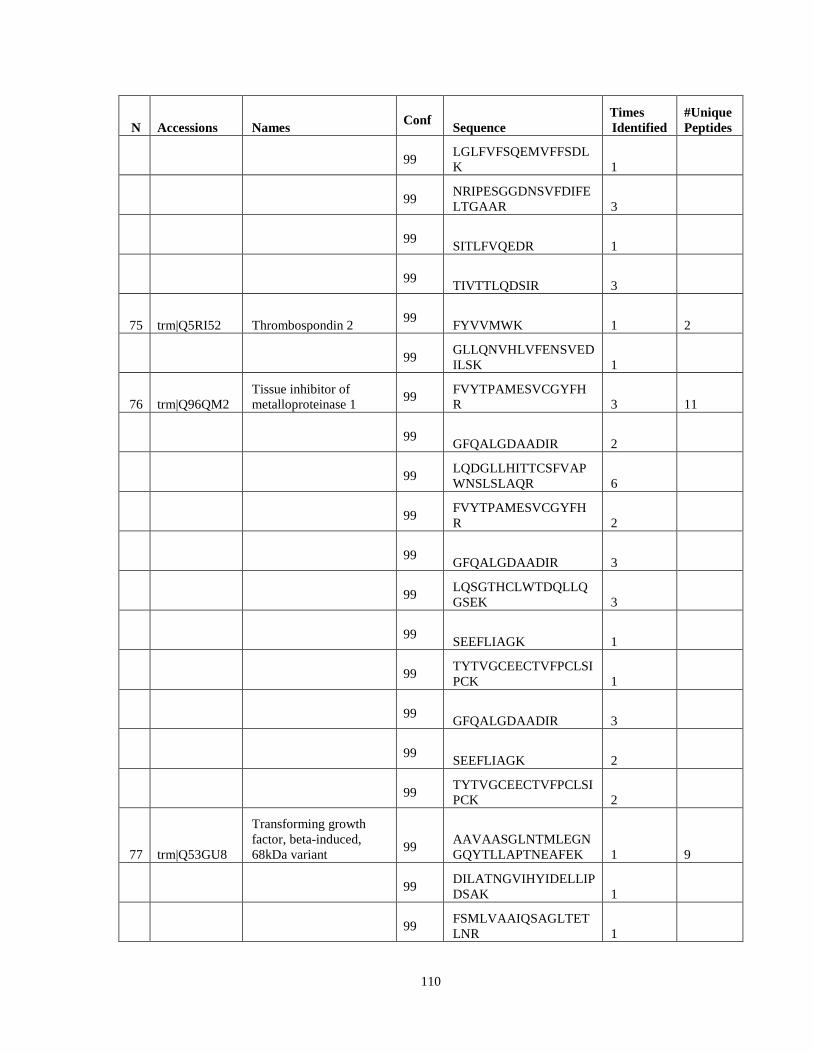
110
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 LGLFVFSQEMVFFSDLK 1
99 NRIPESGGDNSVFDIFELTGAAR 3
99 SITLFVQEDR 1
99 TIVTTLQDSIR 3
75 trm|Q5RI52 Thrombospondin 2 99 FYVVMWK 1 2
99 GLLQNVHLVFENSVEDILSK 1
76 trm|Q96QM2
Tissue inhibitor of metalloproteinase 1 99 FVYTPAMESVCGYFH
R 3 11
99 GFQALGDAADIR 2
99 LQDGLLHITTCSFVAPWNSLSLAQR 6
99 FVYTPAMESVCGYFHR 2
99 GFQALGDAADIR 3
99 LQSGTHCLWTDQLLQGSEK 3
99 SEEFLIAGK 1
99 TYTVGCEECTVFPCLSIPCK 1
99 GFQALGDAADIR 3
99 SEEFLIAGK 2
99 TYTVGCEECTVFPCLSIPCK 2
77 trm|Q53GU8
Transforming growth factor, beta-induced, 68kDa variant 99 AAVAASGLNTMLEGN
GQYTLLAPTNEAFEK 1 9
99 DILATNGVIHYIDELLIPDSAK 1
99 FSMLVAAIQSAGLTETLNR 1
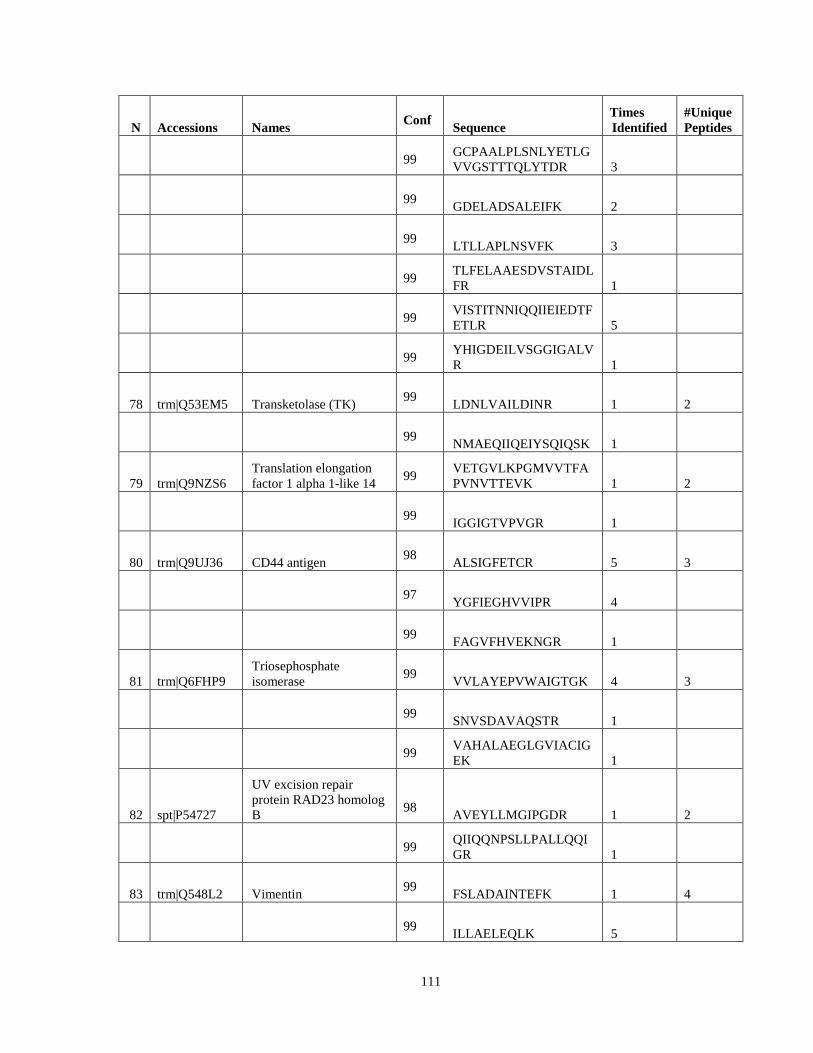
111
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
99 GCPAALPLSNLYETLGVVGSTTTQLYTDR 3
99 GDELADSALEIFK 2
99 LTLLAPLNSVFK 3
99 TLFELAAESDVSTAIDLFR 1
99 VISTITNNIQQIIEIEDTFETLR 5
99 YHIGDEILVSGGIGALVR 1
78 trm|Q53EM5 Transketolase (TK) 99 LDNLVAILDINR 1 2
99 NMAEQIIQEIYSQIQSK 1
79 trm|Q9NZS6
Translation elongation factor 1 alpha 1-like 14 99 VETGVLKPGMVVTFA
PVNVTTEVK 1 2
99 IGGIGTVPVGR 1
80 trm|Q9UJ36 CD44 antigen 98 ALSIGFETCR 5 3
97 YGFIEGHVVIPR 4
99 FAGVFHVEKNGR 1
81 trm|Q6FHP9
Triosephosphate isomerase 99 VVLAYEPVWAIGTGK 4 3
99 SNVSDAVAQSTR 1
99 VAHALAEGLGVIACIGEK 1
82 spt|P54727
UV excision repair protein RAD23 homolog B 98 AVEYLLMGIPGDR 1 2
99 QIIQQNPSLLPALLQQIGR 1
83 trm|Q548L2 Vimentin 99 FSLADAINTEFK 1 4
99 ILLAELEQLK 5
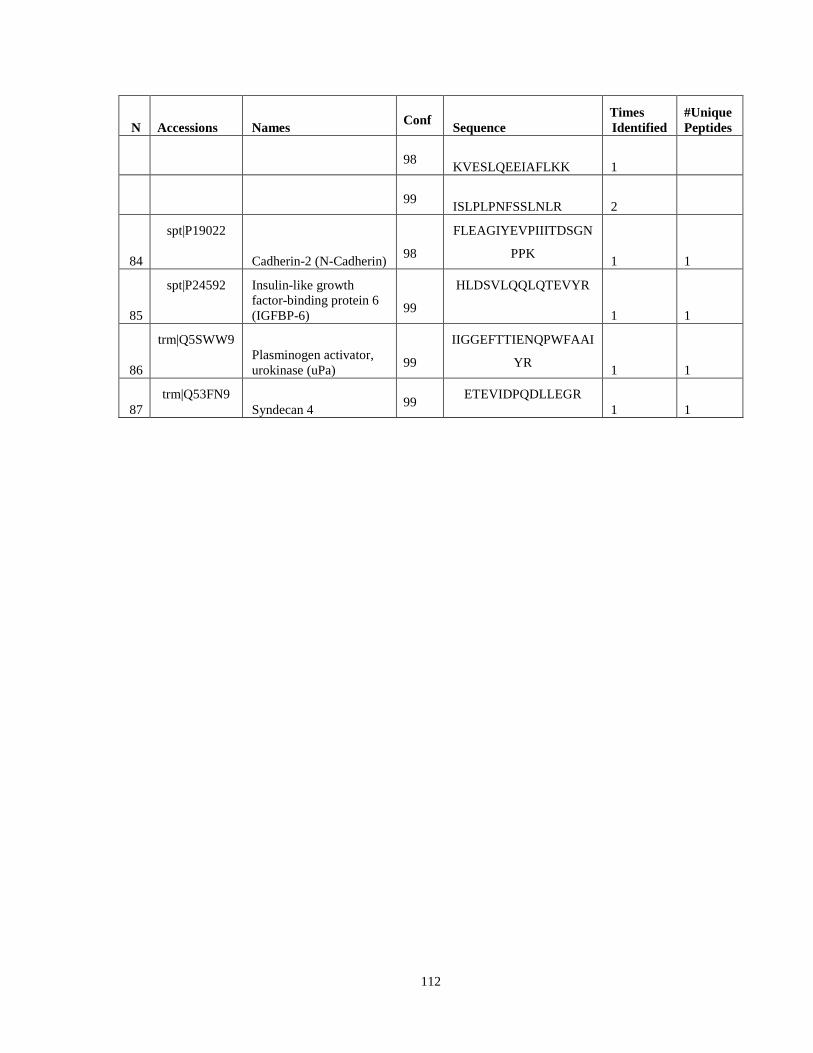
112
N Accessions Names Conf Sequence Times Identified
#Unique Peptides
98 KVESLQEEIAFLKK 1
99 ISLPLPNFSSLNLR 2
84
spt|P19022
Cadherin-2 (N-Cadherin) 98
FLEAGIYEVPIIITDSGN
PPK 1 1
85
spt|P24592 Insulin-like growth factor-binding protein 6 (IGFBP-6) 99
HLDSVLQQLQTEVYR
1 1
86
trm|Q5SWW9 Plasminogen activator, urokinase (uPa) 99
IIGGEFTTIENQPWFAAI
YR 1 1
87 trm|Q53FN9
Syndecan 4 99 ETEVIDPQDLLEGR 1 1
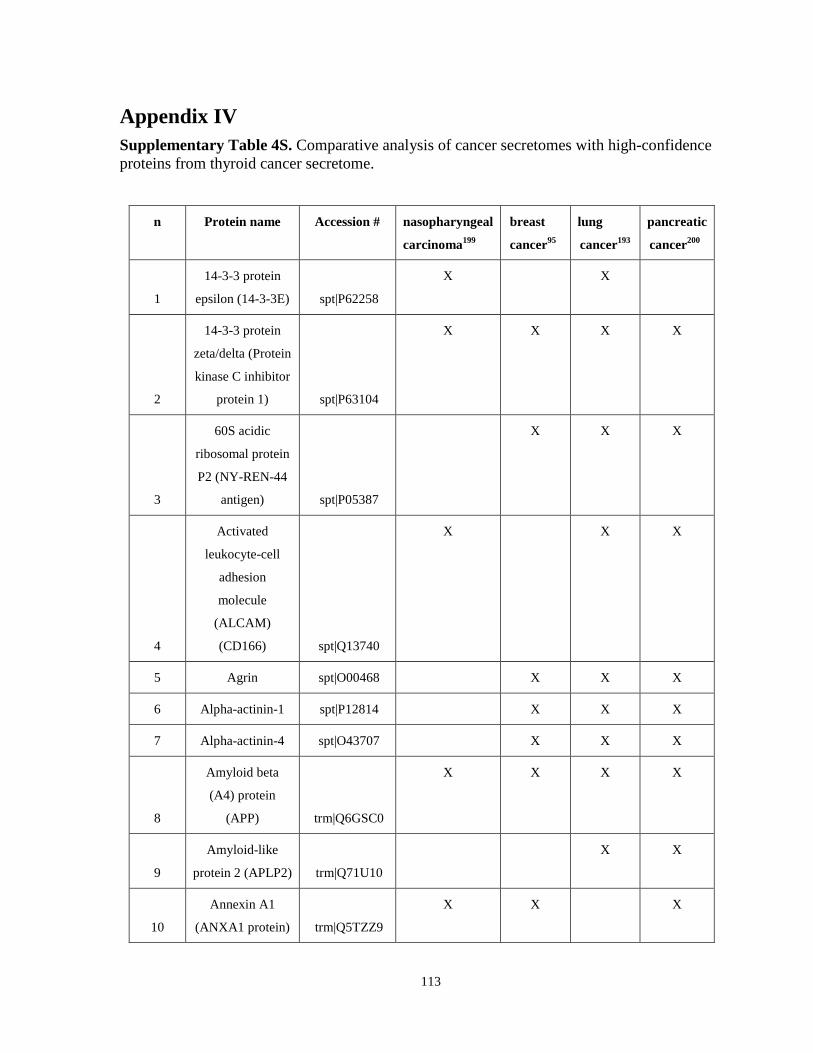
113
Appendix IV Supplementary Table 4S. Comparative analysis of cancer secretomes with high-confidence proteins from thyroid cancer secretome.
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
1
14-3-3 protein
epsilon (14-3-3E) spt|P62258
X X
2
14-3-3 protein
zeta/delta (Protein
kinase C inhibitor
protein 1) spt|P63104
X X X X
3
60S acidic
ribosomal protein
P2 (NY-REN-44
antigen) spt|P05387
X X
X
4
Activated
leukocyte-cell
adhesion
molecule
(ALCAM)
(CD166) spt|Q13740
X X X
5 Agrin spt|O00468 X X X
6 Alpha-actinin-1 spt|P12814 X X X
7 Alpha-actinin-4 spt|O43707 X X X
8
Amyloid beta
(A4) protein
(APP) trm|Q6GSC0
X
X X X
9
Amyloid-like
protein 2 (APLP2) trm|Q71U10
X X
10
Annexin A1
(ANXA1 protein) trm|Q5TZZ9
X X X
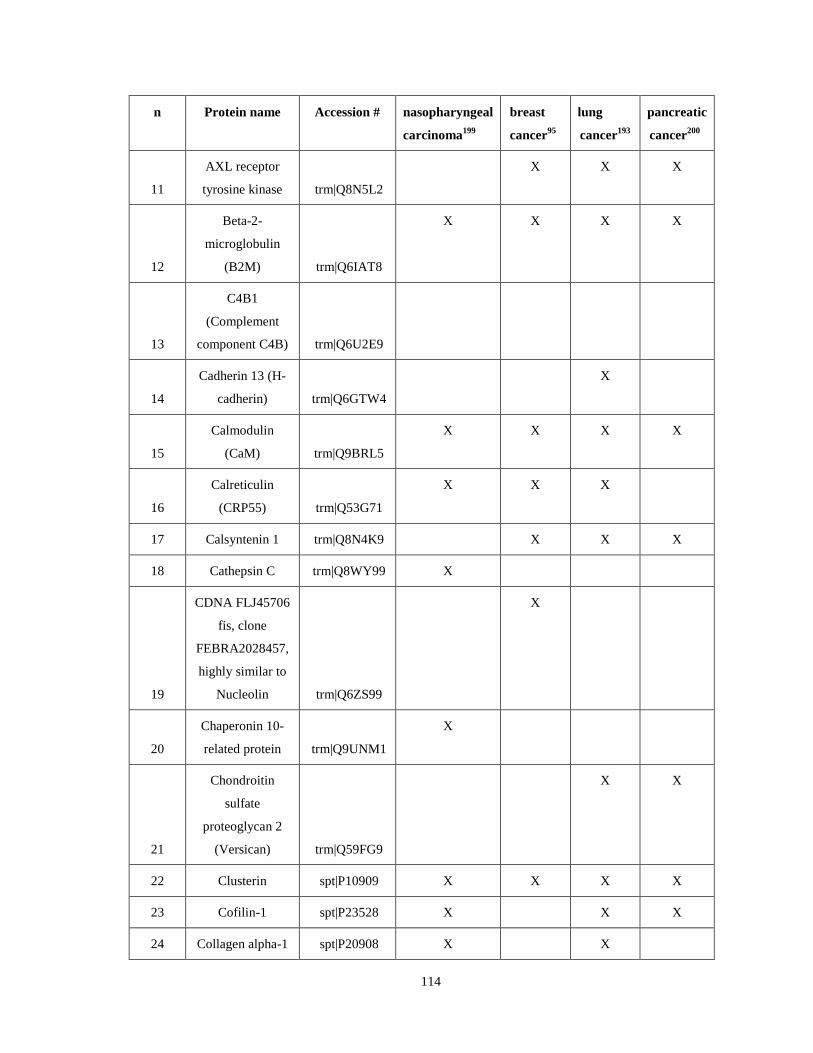
114
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
11
AXL receptor
tyrosine kinase trm|Q8N5L2
X X X
12
Beta-2-
microglobulin
(B2M) trm|Q6IAT8
X X X X
13
C4B1
(Complement
component C4B) trm|Q6U2E9
14
Cadherin 13 (H-
cadherin) trm|Q6GTW4
X
15
Calmodulin
(CaM) trm|Q9BRL5
X X X X
16
Calreticulin
(CRP55) trm|Q53G71
X X X
17 Calsyntenin 1 trm|Q8N4K9 X X X
18 Cathepsin C trm|Q8WY99 X
19
CDNA FLJ45706
fis, clone
FEBRA2028457,
highly similar to
Nucleolin trm|Q6ZS99
X
20
Chaperonin 10-
related protein trm|Q9UNM1
X
21
Chondroitin
sulfate
proteoglycan 2
(Versican) trm|Q59FG9
X X
22 Clusterin spt|P10909 X X X X
23 Cofilin-1 spt|P23528 X X X
24 Collagen alpha-1 spt|P20908 X X
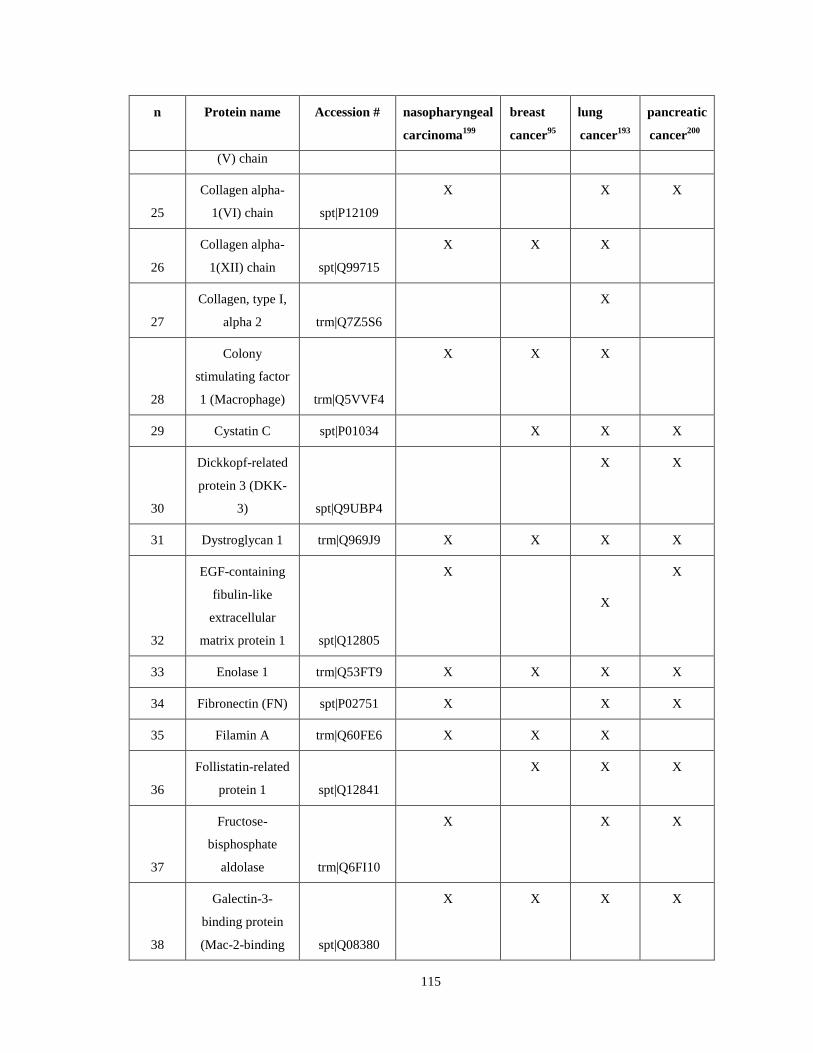
115
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
(V) chain
25
Collagen alpha-
1(VI) chain spt|P12109
X X X
26
Collagen alpha-
1(XII) chain spt|Q99715
X X X
27
Collagen, type I,
alpha 2 trm|Q7Z5S6
X
28
Colony
stimulating factor
1 (Macrophage) trm|Q5VVF4
X
X X
29 Cystatin C spt|P01034 X X X
30
Dickkopf-related
protein 3 (DKK-
3) spt|Q9UBP4
X X
31 Dystroglycan 1 trm|Q969J9 X X X X
32
EGF-containing
fibulin-like
extracellular
matrix protein 1 spt|Q12805
X
X
X
33 Enolase 1 trm|Q53FT9 X X X X
34 Fibronectin (FN) spt|P02751 X X X
35 Filamin A trm|Q60FE6 X X X
36
Follistatin-related
protein 1 spt|Q12841
X X X
37
Fructose-
bisphosphate
aldolase trm|Q6FI10
X
X X
38
Galectin-3-
binding protein
(Mac-2-binding spt|Q08380
X X X X
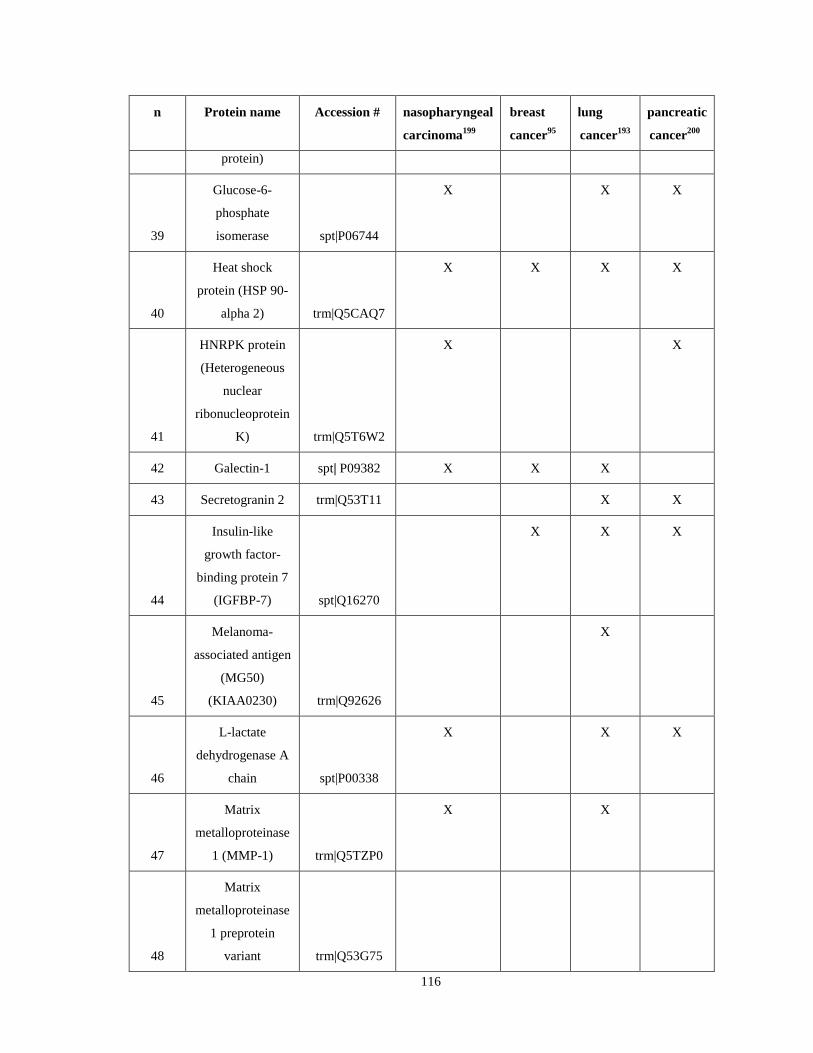
116
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
protein)
39
Glucose-6-
phosphate
isomerase spt|P06744
X X X
40
Heat shock
protein (HSP 90-
alpha 2) trm|Q5CAQ7
X
X X X
41
HNRPK protein
(Heterogeneous
nuclear
ribonucleoprotein
K) trm|Q5T6W2
X
X
42 Galectin-1 spt| P09382 X X X
43 Secretogranin 2 trm|Q53T11 X X
44
Insulin-like
growth factor-
binding protein 7
(IGFBP-7) spt|Q16270
X X X
45
Melanoma-
associated antigen
(MG50)
(KIAA0230) trm|Q92626
X
46
L-lactate
dehydrogenase A
chain spt|P00338
X X X
47
Matrix
metalloproteinase
1 (MMP-1) trm|Q5TZP0
X X
48
Matrix
metalloproteinase
1 preprotein
variant trm|Q53G75
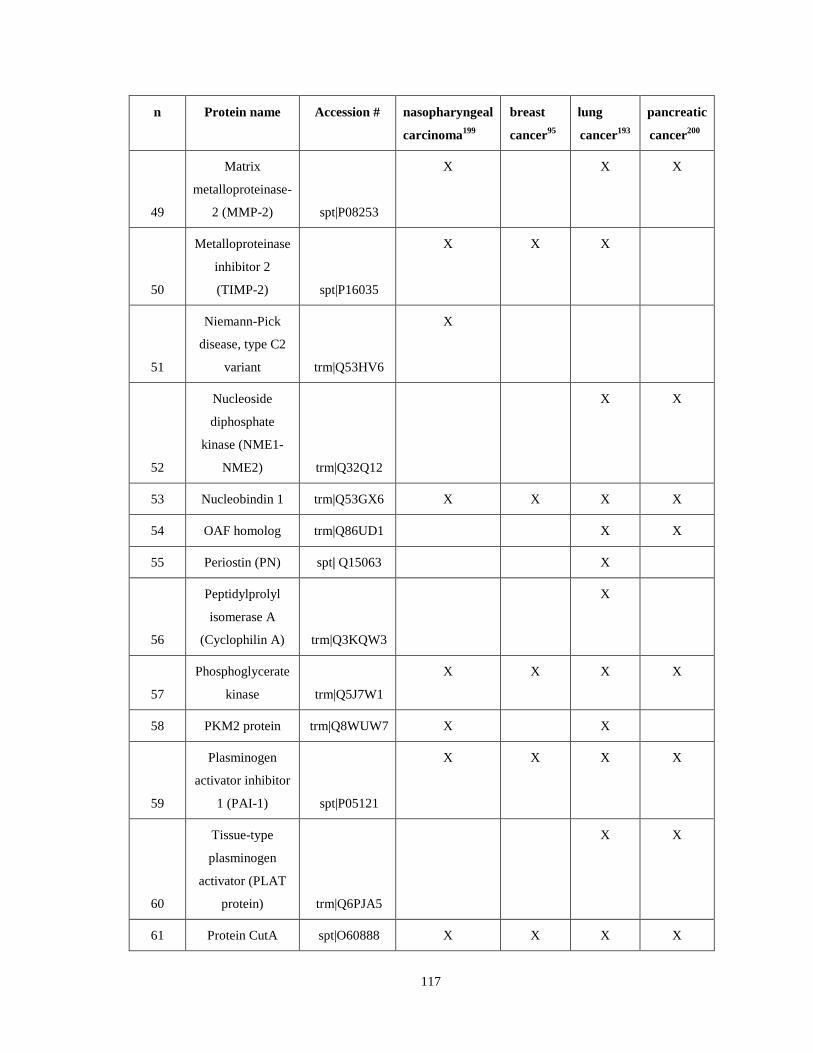
117
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
49
Matrix
metalloproteinase-
2 (MMP-2) spt|P08253
X X X
50
Metalloproteinase
inhibitor 2
(TIMP-2) spt|P16035
X
X X
51
Niemann-Pick
disease, type C2
variant trm|Q53HV6
X
52
Nucleoside
diphosphate
kinase (NME1-
NME2) trm|Q32Q12
X
X
53 Nucleobindin 1 trm|Q53GX6 X X X X
54 OAF homolog trm|Q86UD1 X X
55 Periostin (PN) spt| Q15063 X
56
Peptidylprolyl
isomerase A
(Cyclophilin A) trm|Q3KQW3
X
57
Phosphoglycerate
kinase trm|Q5J7W1
X X X X
58 PKM2 protein trm|Q8WUW7 X X
59
Plasminogen
activator inhibitor
1 (PAI-1) spt|P05121
X X X X
60
Tissue-type
plasminogen
activator (PLAT
protein) trm|Q6PJA5
X
X
61 Protein CutA spt|O60888 X X X X
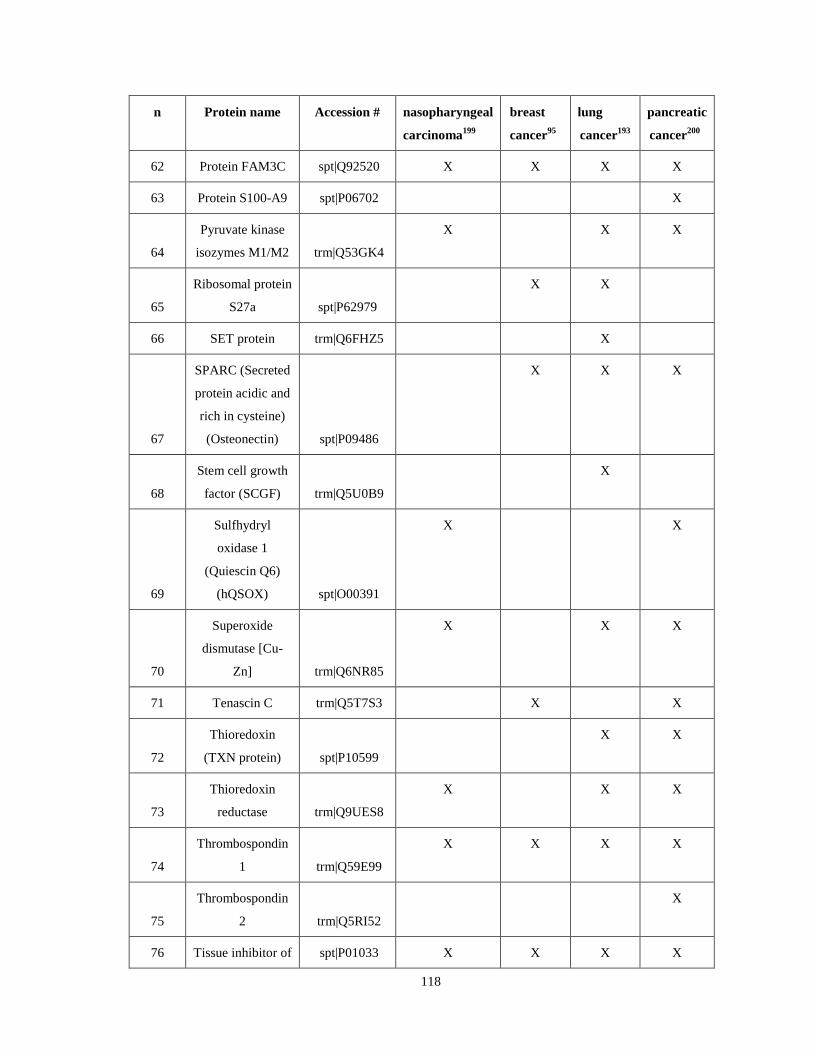
118
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
62 Protein FAM3C spt|Q92520 X X X X
63 Protein S100-A9 spt|P06702 X
64
Pyruvate kinase
isozymes M1/M2 trm|Q53GK4
X X X
65
Ribosomal protein
S27a spt|P62979
X X
66 SET protein trm|Q6FHZ5 X
67
SPARC (Secreted
protein acidic and
rich in cysteine)
(Osteonectin) spt|P09486
X X X
68
Stem cell growth
factor (SCGF) trm|Q5U0B9
X
69
Sulfhydryl
oxidase 1
(Quiescin Q6)
(hQSOX) spt|O00391
X X
70
Superoxide
dismutase [Cu-
Zn] trm|Q6NR85
X X X
71 Tenascin C trm|Q5T7S3 X X
72
Thioredoxin
(TXN protein) spt|P10599
X X
73
Thioredoxin
reductase trm|Q9UES8
X X X
74
Thrombospondin
1 trm|Q59E99
X X X X
75
Thrombospondin
2 trm|Q5RI52
X
76 Tissue inhibitor of spt|P01033 X X X X
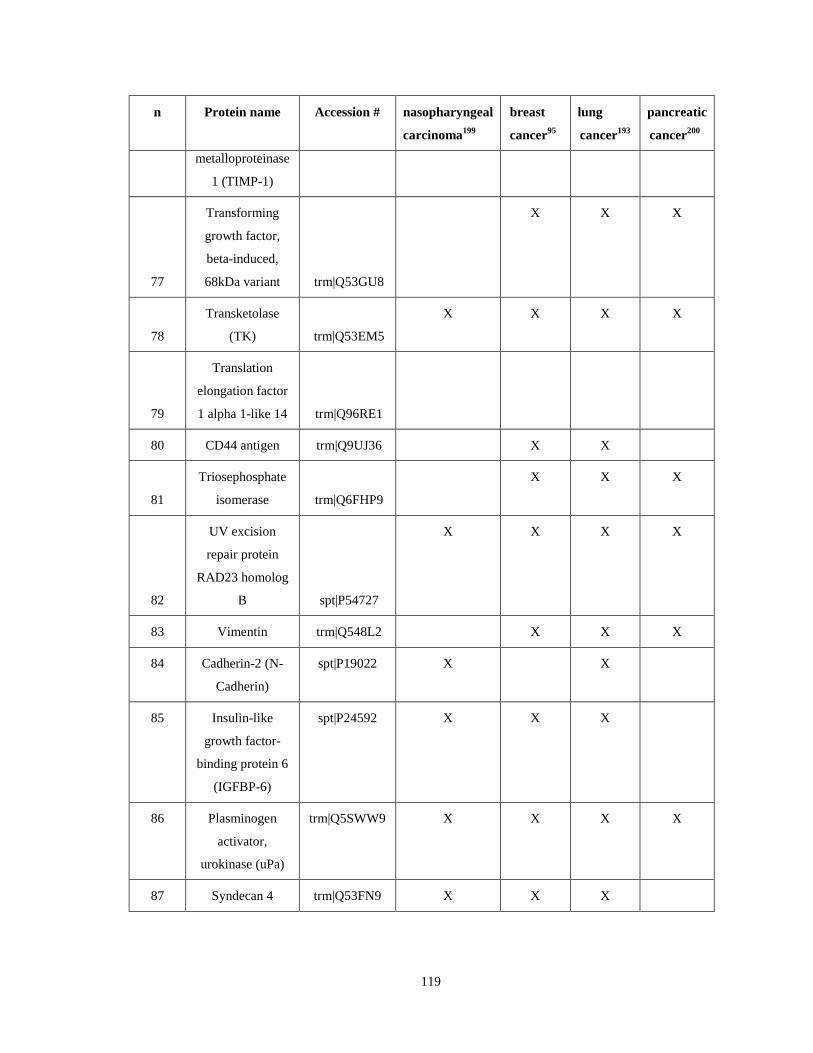
119
n Protein name Accession # nasopharyngeal
carcinoma199
breast
cancer95
lung
cancer193
pancreatic
cancer200
metalloproteinase
1 (TIMP-1)
77
Transforming
growth factor,
beta-induced,
68kDa variant trm|Q53GU8
X X
X
78
Transketolase
(TK) trm|Q53EM5
X X X X
79
Translation
elongation factor
1 alpha 1-like 14 trm|Q96RE1
80 CD44 antigen trm|Q9UJ36 X X
81
Triosephosphate
isomerase trm|Q6FHP9
X X X
82
UV excision
repair protein
RAD23 homolog
B spt|P54727
X X X X
83 Vimentin trm|Q548L2 X X X
84 Cadherin-2 (N-
Cadherin)
spt|P19022 X X
85 Insulin-like
growth factor-
binding protein 6
(IGFBP-6)
spt|P24592 X
X X
86 Plasminogen
activator,
urokinase (uPa)
trm|Q5SWW9
X X X X
87 Syndecan 4 trm|Q53FN9 X X X
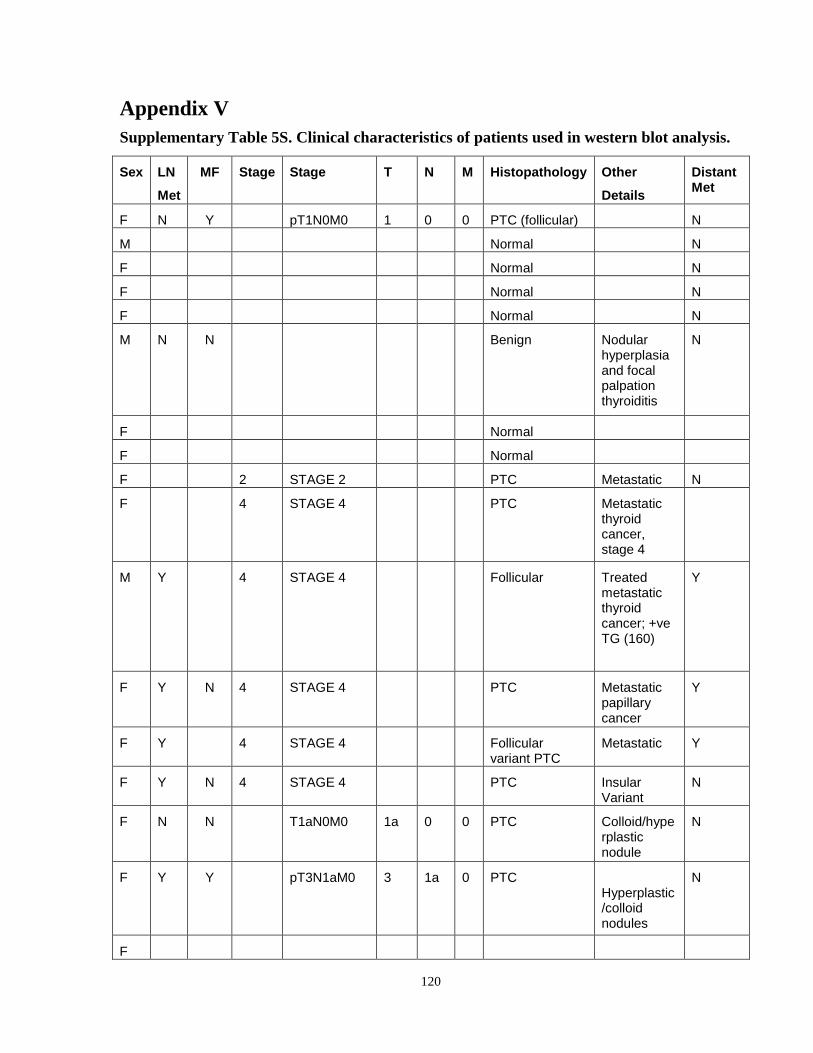
120
Appendix V Supplementary Table 5S. Clinical characteristics of patients used in western blot analysis.
Sex LN
Met
MF Stage Stage T N M Histopathology Other
Details
Distant Met
F N Y pT1N0M0 1 0 0 PTC (follicular) N
M Normal N
F Normal N
F Normal N
F Normal N
M N N Benign Nodular hyperplasia and focal palpation thyroiditis
N
F Normal
F Normal
F 2 STAGE 2 PTC Metastatic N
F 4 STAGE 4 PTC Metastatic thyroid cancer, stage 4
M Y 4 STAGE 4 Follicular Treated metastatic thyroid cancer; +ve TG (160)
Y
F Y N 4 STAGE 4 PTC Metastatic papillary cancer
Y
F Y 4 STAGE 4 Follicular variant PTC
Metastatic Y
F Y N 4 STAGE 4 PTC Insular Variant
N
F N N T1aN0M0 1a 0 0 PTC Colloid/hyperplastic nodule
N
F Y Y pT3N1aM0 3 1a 0 PTC Hyperplastic/colloid nodules
N
F
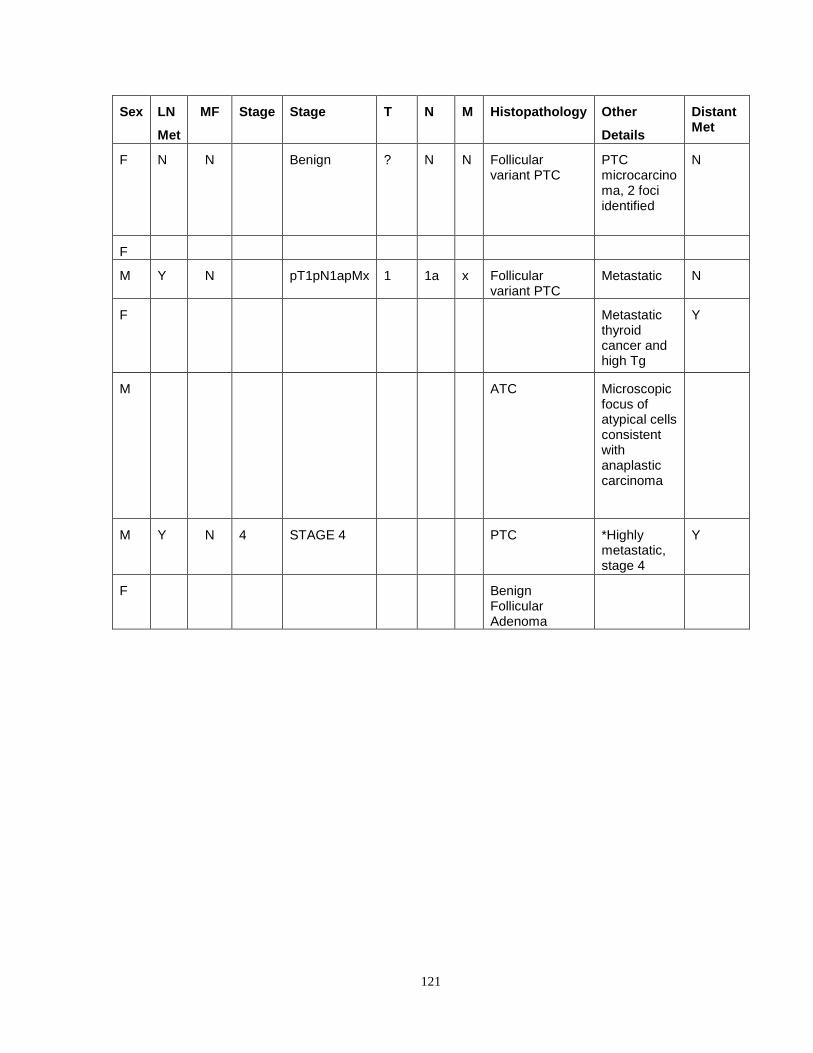
121
Sex LN
Met
MF Stage Stage T N M Histopathology Other
Details
Distant Met
F N N Benign ? N N Follicular variant PTC
PTC microcarcinoma, 2 foci identified
N
F
M Y N pT1pN1apMx 1 1a x Follicular variant PTC
Metastatic N
F Metastatic thyroid cancer and high Tg
Y
M ATC Microscopic focus of atypical cells consistent with anaplastic carcinoma
M Y N 4 STAGE 4 PTC *Highly metastatic, stage 4
Y
F Benign Follicular Adenoma