purification, properties, and immunohistochemical localisation of human brain 14-3-3 protein
TRANSCRIPT

Joi imri l of Nerrroc,hemistrv Raven Press, New York @ 1982 International Society for Neurochemistry
0022-3042/82’0501- 1466/$02.75/0
Purification, Properties, and Immunohistochemical Localisation of Human Brain 14-3-3 Protein
Paul F. Boston, Peter Jackson, Pamela A. M. Kynoch, and R. J. Thompson
Deptirttvient of Clinictil Biochemistry, School qf Cliniccrl Medicine, University of Catnbridge, Addenhrooke’.t Hospi f ct 1, Cam bridge, U . K .
Abstract: A protein has been purified from human brain that appears to be the human equivalent of bovine 14-3-3 protein. On polyacrylamide gel electropho- resis the protein migrates as a faster major component, termed 14-3-3-2 protein, and a slower minor component, termed 14-3-3-1 protein, which consists of approximately 12% of the total protein. Both 14-3-3-1 and 14-3-3-2 have a native molecular weight of approximately 67,000. 14-3-3-2 appears to have the subunit composition (YO; 14-3-3-1 has the composition P’P ’ . Peptide mapping with Stuphvlococcus ciureus V8 proteinase shows that a and /3 subunits are unrelated but the p and /3‘ subunits show some common peptides. Im- munoperoxidase labelling shows that 14-3-3 is localised in neurones in the human cerebral cortex. 14-3-3 shows no enolase, creatine kinase, triose phos- phate isomerase, ATPase, cyclic nucleotide-dependent protein kinase, o r purine nucleoside phosphorylase activity. 14-3-3 does not bind calcium and does not appear to be related to calmodulin, calcineurin, tubulin, neurofilament proteins, clathrin-associated proteins, o r tropomyosin. The functional signifi- cance of this neuronal protein remains obscure. Key Words: Human brain- 14-3-3 Protein-Immunoperoxidase-Neurones. Boston P. F. et al. Purifica- tion, properties, and immunohistochemical localisation of human brain 14-3-3 protein. J . Neurochem. 38, 1466- 1474 (1982).
The mammalian nervous system is known to contain a group of soluble acidic proteins that ap- pear to be both nervous system specific and highly conserved in evolutionary terms (Moore and Perez, 1967). On polyacrylamide gel electrophoresis (PAGE) under nondenaturing conditions (Davis, 1964), S-100 protein has the greatest mobility, moving at the dye front (Isobe and Okuyama, 1978) whereas 14-3-2 protein (neurone-specific enolase) has a relative mobility of approximately 0 .8 (Moore and Perez, 1967). Electrophoretograms of human brain soluble proteins show only one protein migrating between S-100 and 14-3-2 protein; this protein appears to be the human equivalent of bovine brain 14-3-3 protein (Moore and Perez,
1967). This protein, previously purified only from bovine sources (Grasso e t al., 19771, appears to consist of two dissimilar polypeptide chains of ap- proximately 25,000 MW and is subject to axonal transport in the rabbit visual system (Erickson and Moore, 1980). The function of bovine brain 14-3-3 protein is unknown.
As part of a wider programme investigating the properties and possible diagnostic uses of human brain-specific proteins (Hullin et al., 1980; Willson et al., 1980; Thompson et al., 1980; Jackson and Thompson, 1981) we purified a protein from human brain that appears analogous to bovine 14-3-3 pro- tein in electrophoretic mobility and subunit compo- sition. The protein appears to be localised in
Received August 13, 1981; revised November 10, 1981; ac- cepted November 20, 1981.
Address correspondence and reprint requests to Dr. R. J . Thompson, Department of Clinical Biochemistry, School of Clinical Medicine, University of Cambridge, Addenbrooke’s Hospital, Hills Road, Cambridge CB2 2QR, U . K .
Ahhreviarions used: PAGE, Polyacrylamide gel electrophore- sis: SDS, Sodium dodecyl sulphate; SSV, Single-strength ver- onal buffer; TBS, Tris-buffered saline.
1466
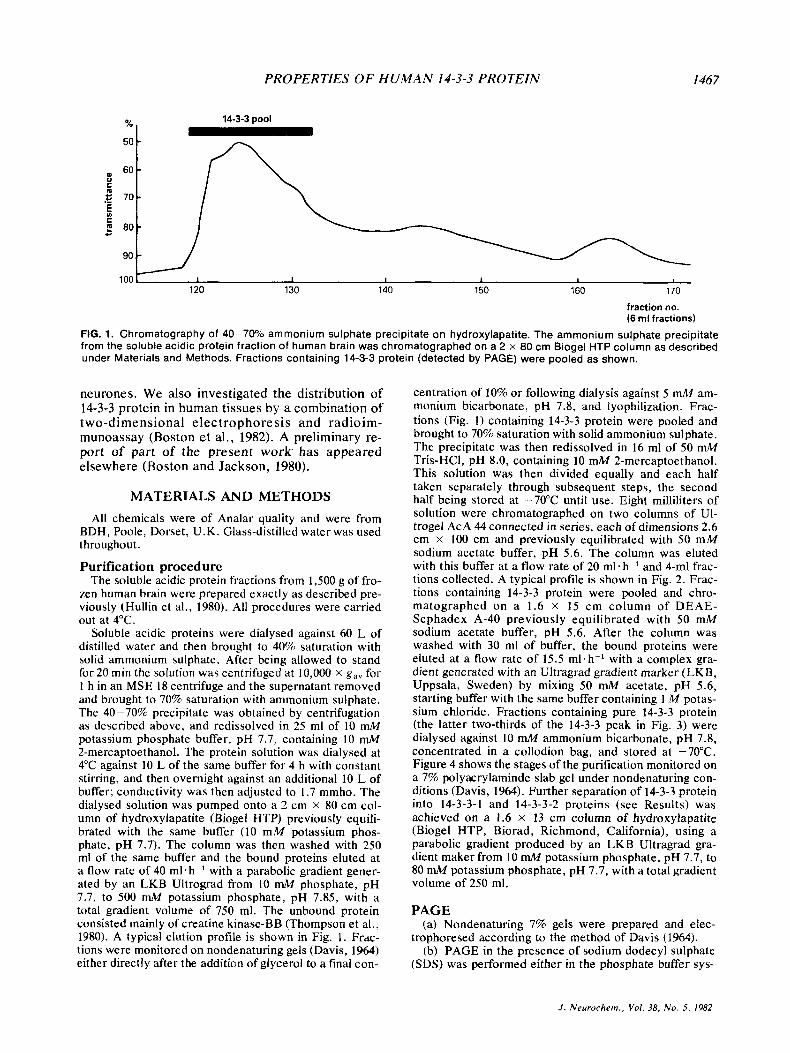
PROPERTIES OF H U M A N 14-3-3 PROTEIN I467
8 .- B i L c
FIG. 1. Chromatography of 40-70% ammonium sulphate precipitate on hydroxylapatite. The ammonium sulphate precipitate from the soluble acidic protein fraction of human brain was chromatographed on a 2 x 80 cm Biogel HTP column as described under Materials and Methods. Fractions containing 14-3-3 protein (detected by PAGE) were pooled as shown.
neurones. We also investigated the distribution of 14-3-3 protein in human tissues by a combination of two-dimensional electrophoresis and radioim- munoassay (Boston e t al., 1982). A preliminary re- port of part of the present work has appeared elsewhere (Boston and Jackson, 1980).
MATERIALS AND METHODS
All chemicals were of Analar quality and were from BDH, Poole, Dorset, U.K. Glass-distilled water was used throughout.
Purification procedure The soluble acidic protein fractions from 1,500 g of fro-
zen human brain were prepared exactly as described pre- viously (Hullin et al., 1980). All procedures were carried out at 4°C.
Soluble acidic proteins were dialysed against 60 L of distilled water and then brought to 40% saturation with solid ammonium sulphate. After being allowed to stand for 20 min the solution was centrifuged at 10,000 x g , , for 1 h in an MSE 18 centrifuge and the supernatant removed and brought to 70% saturation with ammonium sulphate. The 40-70% precipitate was obtained by centrifugation as described above, and redissolved in 25 ml of 10 mM potassium phosphate buffer, pH 7.7, containing 10 mM 2-mercaptoethanol. The protein solution was dialysed at 4°C against 10 L of the same buffer for 4 h with constant stirring, and then overnight against an additional 10 L of buffer; conductivity was then adjusted to 1.7 mmho. The dialysed solution was pumped onto a 2 cm x 80 cm col- umn of hydroxylapatite (Biogel HTP) previously equili- brated with the same buffer (10 mM potassium phos- phate, pH 7.7). The column was then washed with 250 ml of the same buffer and the bound proteins eluted at a flow rate of 40 ml. h-' with a parabolic gradient gener- ated by an LKB Ultrograd from 10 mM phosphate, pH 7.7, to 500 mM potassium phosphate, pH 7.85, with a total gradient volume of 750 ml. The unbound protein consisted mainly of creatine kinase-BB (Thompson et al., 1980). A typical elution profile is shown in Fig. 1. Frac- tions were monitored on nondenaturing gels (Davis, 1964) either directly after the addition of glycerol to a final con-
centration of 10% or following dialysis against 5 mM am- monium bicarbonate, pH 7.8, and lyophilization. Frac- tions (Fig. 1 ) containing 14-3-3 protein were pooled and brought to 70% saturation with solid ammonium sulphate. The precipitate was then redissolved in 16 ml of 50 mM Tris-HC1, pH 8.0, containing 10 mM 2-mercaptoethanol. This solution was then divided equally and each half taken separately through subsequent steps, the second half being stored at -70°C until use. Eight milliliters of solution were chromatographed on two columns of UI- trogel AcA 44 connected in series, each of dimensions 2.6 cm x 100 cm and previously equilibrated with 50 mM sodium acetate buffer, pH 5.6. The column was eluted with this buffer at a flow rate of 20 ml. h-' and 4-ml frac- tions collected. A typical profile is shown in Fig. 2. Frac- tions containing 14-3-3 protein were pooled and chro- matographed on a 1.6 x 15 cm column of DEAE- Sephadex A-40 previously equilibrated with 50 mM sodium acetate buffer, pH 5.6. After the column was washed with 30 ml of buffer, the bound proteins were eluted at a flow rate of 15.5 ml. h-' with a complex gra- dient generated with an Ultragrad gradient marker (LKB, Uppsala, Sweden) by mixing 50 mM acetate, pH 5.6, starting buffer with the same buffer containing 1 M potas- sium chloride. Fractions containing pure 14-3-3 protein (the latter two-thirds of the 14-3-3 peak in Fig. 3) were dialysed against 10 mM ammonium bicarbonate, pH 7.8, concentrated in a collodion bag, and stored at -70°C. Figure 4 shows the stages of the purification monitored on a 7% polyacrylaminde slab gel under nondenaturing con- ditions (Davis, 1964). Further separation of 14-3-3 protein into 14-3-3-1 and 14-3-3-2 proteins (see Results) was achieved on a 1.6 x 13 cm column of hydroxylapatite (Biogel HTP, Biorad, Richmond, California), using a parabolic gradient produced by an LKB Ultragrad gra- dient maker from 10 mM potassium phosphate, pH 7.7, to 80 mM potassium phosphate, pH 7.7, with a total gradient volume of 250 ml.
PAGE (a) Nondenaturing 7% gels were prepared and elec-
trophoresed according to the method of Davis (1964). (b) PAGE in the presence of sodium dodecyl sulphate
(SDS) was performed either in the phosphate buffer sys-
J . Neurochem., Vol. 38, N o . 5 , 1982

1468
-9
-8
-7
-6
- -5
P . F. BOSTON ET AL.
14-3-3 pool - .
1001 I I I t I
350 400 450 500 550 600 Veml
FIG. 2. Chromatography of hydroxylapatite 14-3-3 fractions on Ultrogel. The hydroxylapatite 14-3-3 pool (Fig. 1) was precipitated with ammonium sulphate and chromatographed on a 2.6 x 200 cm Ultrogel Ac 44 column as described under Materials and Methods. Fractions containing 14-3-3 were pooled as shown
tern of Weber and Osborn (1969), using 7.5% gels, o r in the stacking system of Laemmli (1970), using 10% gels. Peptide mapping on SDS gels was performed exactly as described by Cleveland et al. (1977).
Staining of polyacrylamide gels for carbohydrate with periodic acidischiff reagent was performed by the method of Segrest and Jackson (1972), using 2.5 pI of human serum electrophoresed on a parallel gel as control.
Protein, enzyme assays, and calcium-binding activity
Protein was measured by the method of Lowry et al. (1951). Enolase (EC 4.2.1.11) and creatine kinase (EC 2.7.3.2) activities were determined as described previ- ously (Hullin e t al., 1980; Thompson et al., 1980).
Triosephosphate isomerase (EC 5.3.1.1) activity was
% TIO enobse activity IU mi-'
I !2
50 c CI
enolase activity A I \
I \ I \
\ I
\ I I I
i I t I I
I I i I I I I I I
I I
I
assayed in a medium containing 50 mM Tris-HCI, pH 8.5, 130 pg 'ml- ' NAD, 1.5 mg.ml-' sodium arsenate, 10 p g . ml-I glyceraldehyde-3-phosphate dehydrogenase, and 10 p1.ml-' of 50 mM solution of dihydroxyacetone phos- phate. Purine nucleoside phosphorylase (EC 2.4.2.1) ac- tivity was assayed as described by Lewis (1978). Cyclic AMP- and cyclic GMP-dependent protein kinase ac- tivities were assayed as described by Walter et al. (1980). Cyclic nucleotide phosphodiesterase activity and calcium binding activity were assayed as described by Hutton e t al. (1981). ATPase activity was measured in a medium containing 6 mM histidine chloride, 0.6 mM MgCI,, 26 mM NaCI, 4 mM KCI, 1 mM CaClp, 1.9 mg.ml-' ATP, I mg.rn1-I phosphoenol pyruvate, 0.13 mg'rnl-' NADH, 1 pg.rn1-I pyruvate kinase, and 1 pg.rn1-I lactate dehydro- genase.
conductivity rnmho 40
J 14-3-3 pool
I
1 \ \ -- -3 100 - I - I J - - -
50 60 70 80 90 10 fraction no.
(3.1 ml fractions) FIG. 3. Separation of 14-3-2 and 14-3-3 proteins on DEAE-Sephadex. Fractions containing 14-3-3 protein from the Ultrogel pool (Fig. 2) were chromatographed on DEAE-Sephadex A-50 column as described under Materials and Methods.
J . Nrwoc.hrin.. Y o / . 38. No. 5, I982

PROPERTIES OF HUMAN 14-3-3 PROTEIN 1469
Amino acid and N-terminal analysis Hydrolysates of pure 14-3-3 protein and pure 14-3-3-2
protein were prepared as described by Moore and Stein (1963). Amino acid analyses were performed by Dr. A. Moir of the Department of Biochemistry, University of Birmingham, on an LKB 4400 analyser. N-terminal analyses were performed by the dansyl method of Weiner et al. (1972).
Immunoperoxidase localisation in human brain sections
Sections of human cerebrum were processed exactly as described elsewhere (Thompson et al., 1980). Pure 14-3-3 protein (6 mg) was coupled to 10 mg of diazocellulose (Miles and Hales, 1968). Approximately 6 mg of the 14-3-3 immunoadsorbent was incubated with 1.5 ml of rabbit anti-human 14-3-3 antiserum [prepared as described in the accompanying paper (Boston et al., 1982)] for 72 h at 4°C with shaking. After washing the immunoadsorbent twice in 0.2 M borate buffer, pH 8.2, containing 14.9 g-L-' KCI and three times in single-strength veronal (SSV) buffer, pH 8.1, containing 10.3 g- L-' sodium barbitone, 5 g.L-' NaCI, and 0.1 g.L-' sodium azide the antibody- immunoadsorbent complex was stored in aliquots at -20°C. Prior to use an aliquot equivalent to approxi- mately 3 mg of cellulose was thawed and washed by cen- trifugation in 5 ml of SSV buffer and bound antibodies were eluted as described elsewhere (Willson et al., 1981). The final antibody elution with 5 ml of HCI (pH 2) was mixed with 5 ml of double-strength SSV buffer (at twice the above concentration), which also contained 10 g. L-' bovine serum albumin.
Sections of human cerebrum were pretreated with methanol and hydrogen peroxide as described elsewhere (Thompson et al., 1981) and then incubated with nonim- mune sheep serum at room temperature for 10 min. The sections were washed three times with Tris-buffered saline (TBS: 10 mM Tris-HCI, pH 7.4, in saline), then incubated overnight at 4°C with rabbit anti-human 14-3-3 antibodies eluted as above. Sections were washed three times with TBS and then incubated for 30 min at room temperature with sheep anti-rabbit IgG coupled to horse- radish peroxidase.
This reagent was prepared exactly as described by Nakane and Kawaoi (1974) from sheep anti-rabbit IgG (DAKO immunoglobulins, Copenhagen, Denmark) and horseradish peroxidase (Sigma, London). Further stages for colour development and counterstaining were carried out as described elsewhere (Thompson et al., 1981). As controls, sections were treated in parallel with (a) nonimmune rabbit serum, (b) nonimmune rabbit serum taken through the same immunoadsorbent procedure as above, and (c) solutions of anti-14-3-3 antibodies eluted as described above that had been absorbed overnight at 4°C with pure 14-3-3 protein and centrifuged before being applied to the sections.
Molecular weight estimations Molecular weight estimations were performed on the
same Ultrogel AcA 44 columns described under the purification procedure by the method of Andrews (1964), using bovine serum albumin, carbonic anhydrase, and myoglobin as standards.
Miscellaneous procedures Bovine and human brain tropomyosin were prepared in
parallel using the method of Cohen and Cohen (1972).
Bovine brain tubulin, chick brain tubulin, 1251-~hi~k brain tubulin, and anti-bovine brain tubulin were a gift of Dr. J. Kilmartin (MRC Laboratory of Molecular Biology, Cam- bridge). Coated vesicles from pig brain were a gift of Dr. F. Winkler (EMBO Laboratory, Heidelberg).
Densitometry of stained polyacrylamide gels was per- formed on a Gilford 250 spectrophotometer.
RESULTS
The purity of 14-3-3 protein at various stages of the purification procedure is shown in Fig. 4. The only protein appearing between neurone-specific enolase and S-100 at the dye front [i.e., in the po- sition described by Moore and Perez (1967) for bovine 14-3-3 protein] on electrophoresis is a dou- blet consisting of a minor more slowly migrating component and a major more rapidly migrating component. Since peptide mapping with S. aweus
a b C d e f
14-3-2 L
14-3-3- 1 + b
14-3-3-2
FIG. 4. Stages in the purification of 14-3-3 protein. Frac- tions from progressive stages of the purification were elec- trophoresed on a polyacrylamide slab gel as described in Materials and Methods. (a) Total soluble proteins from hu- man brain; (b) soluble acidic protein fraction from the initial DEAE-cellulose column; (c) 40-70% ammonium sulphate precipitate from the soluble acidic protein fraction; (d) the pooled fractions from the hydroxylapatite column (Fig. 1). The Ultrogel pool (not shown) consisted of 14-3-2 and 14-3-3 proteins. (e) The initial portion of the 14-3-3 peak from the DEAE-Sephadex column (Fig. 3); (1) the latter portion of the 14-3-3 peak from the DEAE-Sephadex column (Fig. 3).
J . Neurochem.. Vol. 38, N o . 5 , 1982

1470 P . F . BOSTON ET AL.
TABLE 1. Recovery of protein utid 14-3-3-2 immunoreuctivity during purifirution
Total 14-3-3-2 protein protein
(mg) (mg)
Total soluble proteins 9543 126.1 Soluble acidic protein fraction 1896 89.8 40-70% ammonium sulphate 900 76.5
Hydroxyhpdtite 14-3-3 pool 259 41.0 Ultrogel Ac 44 pool 58 23.1 DEAE-Sephadex 14-3-3 pool 19.6 11.6
precipitate
Total protein was measured according to the method of Lowry et al. (1951). 14-3-3-2 was measured by radioimmunoassay (Bos- ton et al., 1982).
V8 proteinase showed the presence of common peptides (see Fig. 6), the more slowly migrating component (which forms 12% of the total protein present as estimated by densitometry) has been termed 14-3-3-1 and the major more rapidly migrating component has been termed 14-3-3-2. In some elec- trophoreses on slab gels (but not tube gels) the 14- 3-3-2 major component itself showed a fine splitting into two bands (e.g., see Fig. 4). The recovery of protein and 14-3-3 immunoreactivity as determined by radioimmunoassay (Boston et al., 1982) obtained during the purification procedure is shown in Table 1. There was some separation of 14-3-3-1 and 14- 3-3-2 on DEAE-Sephadex (Fig. 4) but complete separation was achieved on hydroxylapatite as de- scribed under Materials and Methods. This proce-
a C b d e
dure led to considerable losses, with 24 mg of 14-3-3 protein yielding 0.6 mg of pure 14-3-3-1 and 3.1 mg of pure 14-3-3-2 protein.
Molecular weight estimations of 14-3-3 protein under nondenaturing conditions gave a value of 67-70,000. Figure 5 shows the appearance of poly- acrylamide gel electrophoresis in the presence of SDS of 14-3-3 protein, purified 14-3-3-1, and purified 14-3-3-2. Two bands with molecular weights of 29,000 and 32,000 were obtained from 14-3-3 protein; pure 14-3-3-1 showed only the smaller band, whereas pure 14-3-3-2 showed both bands. The smaller band of 14-3-3-1 protein comi- grated with the more rapidly moving band of 14-3-3-2 (Fig. 5) .
The relationships between the polypeptides com- prising 14-3-3 protein and purified 14-3-3-1 and 14- 3-3-2 proteins were further investigated using pep- tide mapping with S. a w e u s V8 proteinase as de- scribed under Materials and Methods. Figure 6 shows the result of this procedure at a proteinase concentration of 4 pg-ml-'. The two protein stain- ing bands of 14-3-3-2 appear to be structurally unre- lated, whereas the protein band of 14-3-3-1 appears to have some peptides common to the smaller band of 14-3-3-2, but with at least one major peptide ab- sent (Fig. 6). Essentially similar results were ob- tained at V8 proteinase concentrations of 2 pLg.rn1-l and 40 pg.rn1-l. The two finely split forms of 14- 3-3-2 on slab gels (Fig. 4) showed exactly the same subunit composition and peptide maps when cut from gels and examined separately.
f
FIG. 5. Subunit composition of 14-3-3-1 and 14-3-3-2 proteins. Samples were elec- trophoresed on a polyacrylamide slab gel as described under Materials and Methods. The lanes show (a) molecular weight markers (bovine serum albumin, ovalbumin, carbonic anhydrase); (b) the 14-3-3 protein pool from the DEAE-Sephadex A-50 column (Fig. 3); (c) pure 14-3-3-1 protein; (d) pure 14-3-3-2 pro- tein; (e) 3 p g pure 14-3-3-2 protein; (f) 3 pg pure 14-3-3-2 protein and 5 pg pure 14-3-3-1 protein.
J . Neurorhcm., Vo l . 38, N o . 5 , 1982

PROPERTIES OF HUMAN 14-3-3 PROTEIN 1471
a b C d e
FIG. 6. Peptide mapping of 14-3-3 subunits with S. aureus V8 proteinase. The stained bands shown in Fig. 5 were cut out and subjected to V 8 proteinase digestions as described by Cleveland et al. (1977). The lanes show (a) the single ( p ' ) band from purified 14-3-3-1 protein; (b) the smaller molecu- lar weight ( p ) band from purified 14-3-3-2 protein; (c) the smaller molecular weight band from 14-3-3 protein (lane b in Fig. 5); (d) the larger molecular weight (a) band from purified 14-3-3-2; (e) the larger molecular weight band from 14-3-3 (lane b in Fig. 5) at a lower loading than lane d.
N-terminal amino acid analysis of 14-3-3 protein showed dansyl-E-lysine but no evidence of a free N-terminal residue. The results of amino acid analyses of pure 14-3-3 and 14-3-3-2 are shown in Table 2. Insufficient material was available for a similar analysis of 14-3-3-1. Pure 14-3-3 showed an absorption maximum at 276 nm and an Eh% of 11.8. Native 14-3-3 protein showed no calcium-binding activity and did not stain for carbohydrate when tested as described under Materials and Methods.
Purified 14-3-3 protein did not show any creatine kinase, tr iosephosphate isomerase, purine nu- cleoside phosphorylase, cyclic nucleotide-dependent protein kinase, o r ATPase activity, nor did the purified protein show any effect on cyclic nucleo- tide phosphodiesterase activity. 14-3-3 was devoid of enolase activity; further, the purified protein cross-reacted to only 0.03% on a weight basis with pure 14-3-2 (neurone-specific enolase) in a 14-3-2 radioimmunoassay (Hullin et al., 1980).
Comparison of bovine brain tubulin with human 14-3-3-2 a and /3 subunits by V8 proteinase mapping showed no common peptides. A radioimmunoassay using rabbit anti-bovine brain tubulin, 1 2 5 1 - ~ h i ~ k brain tubulin, and unlabelled bovine brain tubulin showed no immunological cross-reactivity of 14-3-3 protein with either tubulins.
Electrophoresis of pure human 14-3-3 protein in parallel with a preparation of coated vesicles from pig brain showed that both polypeptides of 14-3-3 protein were smaller than low-molecular-weight clathrin-associated proteins. Parallel electrophore- sis of 14-3-3 protein, bovine brain tropomyosin, and human brain tropomyosin showed that both
TABLE 2. Amino acid analysis of human 14-3-3 protein
Human Mammalian Amino platelet brain Bovine
acid 14-3-3 14-3-3-2 tropomyosin tubulin calmodulin
LY s 9.3 8. I 8.8 3.9 4.7 His 0.8 1.3 0.45 2.4 0.7 '4% 4.8 4.3 6.8 4.4 4.1 ASP 11.9 11.8 9.8 10.3 15.5 Thr 4.4 4.1 2.9 6.3 8.1 Ser 6.4 6.1 2.8 5.9 2.7 Glu 18.3 18.4 28.6 14.2 18.2 pro 1.9 3.0 0 5.2 1.4 G ~ Y 4.2 4.4 3.7 7.9 7.4 Ala 9.8 9.3 11.7 7.5 7.4 Val 3.8 4.3 4.0 6.3 4.7 Met 2.9 2.9 2.2 2.3 6.1 Ile 4.1 4.1 4.1 4.5 5.4 Leu 10.3 10.2 11.8 7.7 6.1 TY r 4.6 4.4 0.89 3.9 1.4 Phe 2.4 2.5 0.78 4.8 5.4
~
Amino acid analyses were performed as described in Materials and Methods. For comparison the amino acid analyses of human platelet tropomyosin (Cohen and Cohen, 1972), mammalian brain tubulin (Weisenberg et al., 1968), and bovine cal- modulin (Means and Dedman, 1980) are shown. All values are expressed as mole %; tryptophan and cysteine were not determined.
J . Neurochem., Vol. 38, N o . 5 , 1982

1472 P. F . BOSTON ET A L .
tropomyosin preparations contained two polypep- tide chains (Bretscher and Weber, 19781, the smaller of which was slightly larger than the largest polypeptide chain of 14-3-3 protein.
The results of immunohistochemical staining of sections of human cerebrum are shown in Fig. 7. The protein appeared localised to neurones with no evidence of staining of glial elements. Axonal staining was particularly evident. The staining shown in Fig. 7 was abolished by absorption of anti-human 14-3-3 antibodies with 14-3-3 protein (0.5-3 mg per 100 pl) but not with pure 14-3-2 pro- tein (0.1-3 mg per 100 pl), prepared according to the method of Hullin et al. (1980). Substitution of anti-human 14-3-3 with nonimmune rabbit serum also abolished the staining.
DISCUSSION
The present work describes for the first time the purification and properties of a protein from human brain which, on the basis of its electrophoretic mo- bility (Moore and Perez, 1967) and apparent subunit structure (Grass0 et al., 1977), appears to be the human equivalent of bovine 14-3-3 protein. The protein occurs in human brain in similar quantities to other brain-specific proteins, e.g., creatine kinase-BB (Thompson et al., 1980), neurone-specific enolase (Hullin et al., 1980), or aldolase C (Willson
and Thompson, 1980) and the purification proce- dure described produces the protein in reasonable yield (approximately 15%).
14-3-3 protein migrates on PAGE as a major com- ponent (14-3-3-2) and a minor component (14-3-3-l), the latter representing about 12% of the total pro- tein. Both components appear to have native mo- lecular weights of approximately 67,000 and on poly- acrylamide gel electrophoresis in the presence of SDS both appear to have a dimeric structure (Fig. 5); 14-3-3-1 protein appears to be a homodimer of 29,000 molecular weight subunit and 14-3-3-2 ap- pears to be a heterodimer of one subunit of 29,000 and one of 32,000 MW. Peptide mapping with V8 proteinase suggests that the two subunits of 14-3-3-2 protein are unrelated, but that the smaller subunit of 14-3-3-2 and the subunit of 14-3-3-1 are related, but not identical (Fig. 6). It is therefore proposed that 14-3-3 protein consists of a major component, 14-3- 3-2, with the subunit composition of aj3 (a being the larger subunit) and a minor component, 14-3-3-1, with the subunit composition j3’p’. On the basis of the relative intensity of staining of the 14-3-3-2 bands in Fig. 5, it is possible that 14-3-3-2 is a mix- ture of ap and pj3 dimers. The immunological cross-reactivity of 14-3-3 components and the exis- tence of the protein in tissues other than the brain are discussed in the accompanying paper (Boston e t al., 1982).
FIG. 7. lmmunohistochemical localization of 14-3-3 protein to neurones in the human cerebral cortex. lmmunoperoxidase labelling of sections of human cerebral cortex was performed with affinity purified rabbit anti-human 14-3-3 protein as described under Materials and Methods. Note prominent axonal staining (final magnification, x 140).
.I. Nmrochem. , Vol. 38, No. 5 , 1982

PROPERTIES OF H U M A N 14-3-3 PROTEIN 1473
Immunoperoxidase labelling shows that within the brain, 14-3-3 is localised in neurones (Fig. 7); however, the function of the protein in these cells is unknown. An obvious possibility is that the protein represents a new brain-specific isoenzyme; for example, 14-3-2 was studied for some years as a neurone-specific protein before being recognised as neuronal y y enolase (Bock and Dissing, 1975). Clearly it is impracticable to screen 14-3-3 for all known enzyme activities, but in view of the acidic nature, dimeric structure, and precipitability be- tween 35 and 70% ammonium sulphate shown by brain purine nucleoside phosphorylase (Lewis, 1978), 14-3-3 was tested for this activity and also for triosephosphate isomerase activity (suitable dimeric structure and possible isoenzyme forms in other tis- sues; see Norton et al., 1970), both with negative results. Similarly 14-3-3 protein is devoid of ATPase, cyclic AMP- or cyclic GMP-dependent pro- tein kinase, enolase, or creatine kinase activity.
The acidic nature of 14-3-3 and its presence in other tissues apart from the brain (Boston et al., 1982) suggests the possibility that 14-3-3 may be re- lated to proteins such as calmodulin (Means and Dedman, 1980) or calcineurin (Klee et al., 1979). However, 14-3-3 does not show calcium-binding activity, has no effect on cyclic nucleotide phos- phodiesterase, and further, is clearly distinguish- able from calmodulin in molecular weight, amino acid composition, (Table 2), and on two-dimension- al electrophoresis (Jackson and Thompson, 1981).
A further possibility is that 14-3-3 protein repre- sents a cytoskeletal element especially in view of the prominent axonal staining produced by the im- munoperoxidase technique (Fig. 7). However, 14-3-3 is too small to represent tubulin o r any known microtubular component; also, bovine brain tubulin does not cross-react in a 14-3-3 radioim- munoassay (Boston et al., 1982). In addition, pep- tide mapping with V8 proteinase does not show any similarity between bovine brain tubulin and human 14-3-3 protein. Glial fibrillary acidic protein is firstly localized in astrocytes rather than neurones and secondly too large to represent 14-3-3 (Dahl and Bignami, 1975); similarly 14-3-3 is of too low a molecular weight to represent any known neurofil- ament protein (Lazarides, 1980). Although (espe- cially working with human postmortem material) it cannot be excluded that 14-3-3 is derived from a higher molecular weight precursor, there is no evi- dence of such a precursor when human brain ex- tract is gel-filtered and the resulting column frac- tions subjected to radioimmunoassay for 14-3-3 (Boston et al., 1982). Further, two-dimensional electrophoresis of fresh rat brain shows a 14-3-3 complex very similar to that seen in human brain (P. Jackson and R. J. Thompson, unpublished work). Coated vesicle-associated proteins (Pearse, 1978) appear to be too large to be related to 14-3-3 and
also 14-3-3 immunoreactivity in brain extracts is not sedimented by centrifugation at 100,000 x gav for 1 h (P. F. Boston, unpublished work). A final possi- bility considered in the present study was that 14-3-3 might represent human brain tropomyosin. However, the peptides from this, which appear identical in mobility to the two polypeptides from bovine brain tropomyosin (Bretscher and Weber, 1978), do not correspond to the polypeptides seen in 14-3-3 protein. There are also some differences in the amino acid composition of 14-3-3 compared with human platelet tropomyosin (Table 2).
Although the function of 14-3-3 protein remains unknown, the neuronal localisation and possible brain specificity of this protein make it a potential candidate as a marker for damage within the CNS in neurological disorders. We have accordingly inves- tigated this possibility in the accompanying paper (Boston et al., 1982).
Acknowledgments: This work was supported by a grant from the Wellcome Trust to Professor C. N. Hales. P.F.B. is a Wellcome Trust Research Scholar.
REFERENCES Andrews P. (1964) Estimation of molecular weight of proteins by
Sephadex gel filtration. Biochrrn. J. 91, 222-233. Bock E. and Dissing J. (1975) Demonstration of enolase activity
connected to the brain-specific protein 14-3-2. Scand. J. Immunol. (Suppl. 2) 4, 31-36.
Boston P. F. and Jackson P. (1980) Purification and properties of a brain-specific protein, human 14-3-3. Biuchem. Soc. Trans.
Boston P. F., Jackson P., and Thompson R. J. (1982) Human 14-3-3 protein: Radioimmunoassay, tissue distribution, and cerebrospinal fluid levels in patients with neurological dis- orders. J. Neurochern. 38, 1475- 1482.
Bretscher A. and Weber K. (1978) Tropomyosin from bovine brain contains two polypeptide chains of slightly different molecular weights. FEBS Lef t . 85, 145- 148.
Cleveland D. W., Fischer S. F., Kirschner M. W., and Laemmli U. K. (1977) Peptide mapping by limited proteolysis in sodium dodecyl sulphate and analysis by gel electrophore- sis. J . B i d . Chem. 252, 1102-1106.
Cohen I . and Cohen C. (1972) A tropomyosin-like protein from human platelets. J . Mu/. Biol. 68, 383-387.
Dahl D. and Bignami A. (1975) Glial fibrillary acidic protein from normal and gliosed human brain-demonstration of multiple related polypeptides. Biochirn. Biophys. Acta 3845, 41 -51.
Davis B. J. (1964) Disc electrophoresis, part 2 (Method and ap- plication to human serum proteins). Ann. NY Acud. Sci.
Erickson P. F. and Moore B. W. (1980) Investigation of the axonal transport of three acidic, soluble proteins (14-3-2, 14-3-3, and S-100) in the rabbit visual system. J . Neurochern. 35, 232-241.
Grasso A., Roda G., Hogue-Angeletti R. A., Moore B. W., and Perez V. J. (1977) Preparation and properties of the brain- specific protein 163-2. Brain Res. 124, 497-507.
8, 617-618.
121, 404-427.
Hullin D.A. (1979) Ph.D. Thesis, University of Wales. Hullin D. A., Brown K. B., Kynoch P. A. M., Smith C., and
Thompson R. J. (1980) Human 14-3-2 protein: Purification radioimmunoassay and distribution in human tissues. Biochim. Biuphys. Acta 628, 98-108.
Hutton J. C., Penn E. J. , Jackson P., and Hales C. N. (1981)
J . Neurochem., Vol. 38, N o . 5 , 1982

1474 P . F . BOSTON ET AL.
Isolation and characterization of calmodulin from an insulin-secreting tumour. Biochem. J . 193, 875-885.
Isobe T. and Okuyama T. (1978) The amino acid sequence of S-100 protein (PAP I-b protein) and its relation to the calcium-binding proteins. Eur . J . Biochem. 89, 379-388.
Jackson P. and Thompson R. J. (1981) The demonstration ofnew human brain-specific proteins by high-resolution two- dimensional polyacrylamide gel electrophoresis. J . Nrirrol.
Klee C. B., Crouch T. H., and Knnks M. H. (1979) Calcineurin: A calcium and calmodulin-binding protein of the nervous system. Proc. Nat l . Acutl. Sci. USA 76, 6270-6273.
Laemmli U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T,. Nuture 283,
Lazarides E. (1980) Intermediate filaments as mechanical inte- grators of cellular space. Nature 283, 249-256.
Lewis A. S. (1978) Rabbit brain purine nucleoside phosphorylase purification and properties. Arch. Biochem. Biophys . 190,
Lowry 0. H., Rosebrough N. J. , Farr A. L., and Randall R. J. (1951) Protein measurement with the Folin phenol reagent. J . Biol. Chem. 193, 265-275.
Means A. R. and Dedman J. R. (1980) Calmodulin-An in- tracellular calcium receptor. Nature 285, 73-75.
Miles L. E. M. and Hales C. N. (1968) The preparation and properties of purified '"I-labeled antibodies to insulin. Biochem. J . 108, 61 1-618.
Moore B. W. and Perez V. J. (1967) Specific acidic proteins of the nervous system, in Physiologicril and Biochemicul As- pects of' Nervous Integrution (Carlson F. D, ed), pp. 343-359. Prentice-Hall, Englewood Cliffs, New Jersey.
Moore S. and Stein W. H. (1963) Chromatographic determina- tion of amino acids by the use of automatic reading equip- ment, in Methods in Enzymology, Vol. 6, (Colowick S. P. and Kaplan N. O., eds). pp. 819-831. Academic Press, New York and London.
Nakane P. K. and Kawaoi A. (1974) Peroxidase-labelled antibody-a new method of conjugation. J . Histoehem. Cytochem. 22, 1084- 1091.
Norton I. L., Pfuderer P., Stringer C. D., and Hartmann F. (1970) Isolation and characterization of rabbit muscle triose phosphate isomerase. Biochemistry 25, 4952-4958.
Sci. 49, 429-438.
249- 256.
662- 270.
Pearse B. M. F. (1978) On the structural and functional compo- nents of coated vesicles. J . M o l . Bid. 126, 803-812.
Putnam F. W. (1975) Transferrin, in The Plusnza Proteins, Vol. I (Putnam F. W., ed), pp. 265-316. Academic Press, New York, San Francisco, London.
Segrest J . P. and Jackson R. L. (1972) Molecular weight deter- mination of glycoproteins by polyacrylamide gel electropho- resis in sodium dodecyl sulphate, in Methods in Enzynzol- ogy, Vol. 28, (Ginsburg V. , ed), pp. 54-63. Academic Press, New York and London.
Thompson R. J. , Graham J. G . , McQueen I . N. F., Kynoch P. A. M., and Brown K. W. (1980) Radioimmunoassay of brain-type creatine kinase BB isoenzyme in human tissues and in serum of patients with neurological disorders. J . Neurol. Sci. 47, 241 -254.
Thompson R. J., Kynoch P. A. M., and Sajant J. (1981) Im- munohistochemical localization of creatine kinase BB isoenzyme to astrocytes in human brain. Bruin Re.s. 201,
Walter U., Miller P., Willson F., Markes D., and Greengard P. (1980) Immunological distinction between guanosine 3'-5'- monophosphate and adenosine 3'-5'-monophosphate de- pendent protein kinases. J . B i d . Chem. 255, 3757-3762.
Weber K. and Osborn M. (1969) The reliability of molecular weight determinations by dodecyl-sulphate-polyacrylamide gel electrophoresis. J . Biol. Chem. 244, 446-4412.
Weiner A. M., Platt T., and Weber K. (1972) Amino-terminal sequence analysis of proteins purified on a nanomole scale by gel electrophoresis. J . Bid. Chem. 247, 3242-3251.
Weisenberg R. C., Baisy E. G., and Taylor E. W. (1968) The colchicine binding protein of mammalian brain and its rela- tion to microtubules. Biochemistry 7, 4466-4478.
Willson V. J. C. and Thompson R. J. (1980) Human brain al- dolase C, isoenzyme-Purification, radioimmunoassay and distribution in human tissues. Ann. Clin. Biocheni. 17,
Willson V. J . C. , Graham J . E., McQueen I . N . F . , and Thompson R. J. (1980) Immunoreactive aldolase C in cere- brospinal fluid of patients with neurological disorders. Ann. Clin. Biochem. 17, 110-113.
Willson V. J . C . , Jones H. M., and Thompson R. J. (1981) A two-site immunoradiometric assay for the MB isoenzyme of creatine kinase. Clin. Chim. Actu 113, 153- 163.
423-426.
114-121.
J . Neurochem.. Vol. 38, N o . 5 . 1982