recent progress on coconut micropropagation through...
TRANSCRIPT

LI’ c
Recent progress on coconut micropropagation through a joined effort involving different countries
t I I J-L. VERDEIL’, R. HORNUNG2, H-J. JACOBSEN3, E. RILLO’, C, OROPEZA4,
R. BOURDEIX6, Y-P. N’CH06, V/HOCHER’, S. /HALVION1 and A SANGARE6 1
ORSTOM/CIRAD-CP, Laboratoire des Ressources Génétiques et d Xniélioration des Plantes Tropicales. BP 5045, 34032 Montpellier Cedex 01, FRANCE. E-mail: verdeì[email protected] Uiirversity ofLoridon, [Yye College, Wye, Ashford, TN25 5AH, U.K. University of Hannover, Herreiihausser str, 2, Hannover D-3000, G W N Y P U , Albay Research Center, Banao, 4503 Guinobatan, &bay, PHILIPPINES.
i
’ Cenho de Investigacidii Cientifica de Yucatán, A. P. 87, Mérida 9731 O, Yucatán, *CO. 61DEFOWDP0 - Station Marc Delorme, Port Boutit. 07BP13, Abidjan 07, CdTE D TVOIm.
1. Introduction
Coconut is one of the most important perennial crops for the economy of intertropical countries (Persley, 1992). Thanks to breeding, high-performance hybrids can be achieved. However, conventional selection methods have some constraints derived from the features of the reproduction of these perennial plants: great variability among progeny (consequence of the allogamy), very long selection cycle, low multiplication rate (coconut is only multiplied from seeds), and absence of natural vegetative propagation.
Fortunately, coconut asexual propagation can be achieved through in vitro culture techniques, and more particularly through somatic embryogenesis. Plantation productivity could be significantly increased if homogeneous materials obtained by cloning high-yielding individuals were available. Vegetative propagation of tolerant or resistant specimens would also offer an effective control of various diseases that are a real threat to coconut plantations in certain parts of the world (Santos, this volume). Therefore, somatic embryogenesis is the way to obtain homogeneous and high-yielding plant material on a large scale.
More than 25 years ago, several research teams around the world started studies to clone the coconut palm that showed to be extremely recalcitrant to in vitro culture. With a poor basic knowledge on in vitro tissue physiology of this species, only
I l
1 I
I Fonds Dscumenhir~ B R D


few plantlets were regenerated through unreliable protocols (Blake, 1990; Verdeil and Buffard-Morel, 1995). Until 1995 plantlet regeneration was considered as “the great challenge” and this led to competition and lack of collaboration between the different teams, resulting in a very poor exchange of scientific information. During the last four years (1995-1998), a ‘collaborative effort directed towards the improvement of coconut micropropagation through somatic embryogenesis, started within an EC STD3-project by five teams: three European laboratories (Wye College in the UK, the University of Hannover in Germany and ORSTOM- CIR4D in France), and two laboratories from producing countries (Philippines Coconut Authority - PCA in Philippines; and Centro de Investigación Científica de Yucatán - CICY in México). The present paper reports highlights of the results obtained in this project,
I
1
The main particular goals of this project were:
a. To obtain highly embryogenic callus lines using different explant sources (immature leaves, immature inflorescences and plumules from zygotic embryos).
b. The identification of markers for embryogenic competence (histological and protein markers).
c. The quantification of the endogenous growth regulators and understanding of the fate of the exogenous ones, during the initiation of somatic embryogenesis in the presence of activated charcoal.
d. The study of vitroplant physiology using zygotic embryos grown in vitro as a model for the optimisation of plantlet development.
The different phases of the in vitro protocols have been improved by the different participants of the project and plantlets are now regularly obtained in most laboratories of the consortium.
2. Isolation of embryogenic callus lines
2. I . On solid media containing charcoal (Partners involved: PCA, CICY, Wye, ORsToM/CIIRA 0)
It has been demonstrated for a long time that tissues collected fiom adult coconut palms can be maintained on a medium containing charcoal, (the latter controlling browning, which is one of the major constraints of coconut tissue culture), to allow calli production in the presence of auxin growth regulators such as 2,4- dichlorophenoxyacetic acid (2,4-D) (Blake and Eeuwens, 1980). As the culture of
3 92
t

!
coconut tissues has in the past been practised in a number of laboratoriks there has been a lot of discussion about choice of media and related matters. One of most important objectives has been to compare and to improve the different protocols for callogenesis and embryogenesis using different explant sources (immature inflorescences, immature leaves and plumular tissues removed from zygotic embryos).
Protocols available for indirect somatic embryogenesis were published at the beginning of the project in a compendium of the protocols describing in detail plant material and media composition. Comparative protocol trials have been conducted by the different partners, and different factors have been studied to
Different types were tested: the auxin 2,4-D on callogenesis; thidiazuron (TDZ), 2-isopentenyl adenine (2iP) and others cytokinins in combination with 2,4-D on somatic embryogenesis; and the cytokinin 6-benzylaminopurine (BAP) and gibberellic acid (GA3) on somatic embryo germination.
- :.
improve their performances. One particularly important is growth regulators, .!
Embryogenic calli and complete embryos able to germinate were obtained in all the laboratories involved in the project, working with media containing 2,4-D and activated charcoal, and starting with different sources of explants. In Wye College from immature inflorescences and plumular tissues; in CICY from plumular tissues (Chan et al., 1998; Sáenz et al., this volume); in PCA from immature leaves, plumular tissues and immature inflorescences; and in ORSTOM-CIRAD from immature leaves, plumular tissues and immature inflorescences.
Also, important achievements have been reported by Wye College in the production of embryogenic callus from immature inflorescence tissues, using a combination of the ORSTOMKIRAD and Wye methods (Hornung and Verdeil, this volume). The formation of embryos able to germinate from plumular tissues has been also reported by the tissue culture staff in the Philippines, using a
own protocol for plantlet regeneration. Other examples of positive interactions reflect the quality and the volume of the information exchanged between the partners, who have succeeded in strengthening their links within the framework of this consortium.
The trials carried out by the different partners allow the identification and confirmation of some key factors for coconut regeneration through somatic embryogenesis:
I
I
i
! I
combination of the Wye College protocol for calloid initiation followed by their i.
393

..
a. The optimum 2,4-D level for the formation of embryogenic calli depends on the genotype.
b. A study conduced by PCA on the response of different cultivars to their callus initiation media showed that there are significant differences among cultivars in terms of percentage of calloid formation and that the optimum 2,4-D concentration required for this varies depending on the genotype used. It is interesting to note that the use of different levels of auxin allowed the formation of calloids on all the genotypes tested in different laboratories (Wye College, PCA and ORSTOM-CIRAD).
c. Regeneration of complete somatic embryos require a decrease in the 2,4-D level and the introduction of a cytokinin in the medium. TDZ or 2iP were found to be two of the most efficient cytokinins
d. Germination of the somatic embryos (obtained fiom all the experimented explant sources) is initiated on a medium containing BAP.
e. GA3 promotes somatic embryo germination in the presence of BAP. This promising result obtained by CICY with plumular tissue, need to be repeated.
$
This study based on the exchange of know-how and complementary experimental results obtained in the different laboratories has allowed the regeneration of vitroplants (fiom plumules, immature inflorescences and leaf fragments) in all the laboratories involved in the project. Reliable protocols for plantlet regeneration have been developed fiom plumules and immature inflorescences and could be duplicated in other laboratories, This represents an important breakthrough for coconut regeneration.
2.2. On solid charcoal-jììee media (Partners involved: Hannover, Wye, PCA, CICK ORSTOM/CIRAD)
The use of activated charcoal in tissue culture media is a controversial subject since it is beneficial, but has some disadvantages at the same time. It is now well established that charcoal not only adsorbs unwanted polyphenols (Weatherhead et al:, 1978), but also important media compounds like plant growth regulators (Ebert and Taylor, 1990; Ebert et al., 1993). This absorption of phytohormones is a major obstacle for mastering in vitro coconut regeneration. Although, trials to remove charcoal carried out in the past have been unsuccessful, it was considered important within the project to perform new experiments in order to develop protocols for coconut tissue culture in charcoal-free media.
In 1995 the Hannover group carried out an experiment using charcoal-free medium and reported calli formation from immature inflorescence explants. They
3 94

used a Murashige and Skoog basal medium with sucrose (50 g I-'), containing TDZ in combination with polyvinylpyrrolidone (PVP, 2.5 g 1-') (a compound that adsorbs polyphenols involved in tissue browning). The medium was solidified with gelrite (3 g 1-') and tissues were subcultured every 2 weeks.
They observed a high rate of callus formation (75% with inflorescence F-7 and TDZ 3 x M) after only three weeks of culture (Köhne and Jacobsen, this volume). Further development of the cultures led to shoot-like structures. These structures were transferred onto media for the induction of shoot elongation and rooting (Köhne and Jacobsen, this volume). Unfortunately, most of the cultures were lost because of contamination by a bacterium (presumed to be an endogenous bacterium) or by fimgus.
.-
In contrast, experiments carried out in Mérida, Albay, Wye and Montpellier with immature inflorescences according to the Hannover charcoal-free protocol led to callus formation but with a very low percentage (less than 5%, 4-6 months after the start of culture). Intense tissue browning and slow calli growth was noted. During the subsequent transfer of the calli to fresh medium, they turned brown and died.
Some of the calli obtained in Hannover were sent to Montpellier for histological analysis. It turned out that most of them were expanded flower buds that did not show features generally observed in embryogenic calli. However, three of the callus samples (out of 19 analysed) showed organogenic potential (see below 4.1.). Unfortunately, in terms of avoiding the use of charcoal, these trials were not successful. It seems that we will have to continue depending on the use of this additive, at least for the culture of inflorescence explants.
In CICY using plumular tissues, embryogenic callus formation and embryo development were obtained in charcoal-free medium (with or without PVPP, an insoluble form of PVP), and containing 2,4-D instead of TDZ (Sáenz el al., this volume). In fact, in contrast with immature inflorescences, plumular tissue, which
any adsorbent. According to CICY, further research is still required to determine whether the performance of plumule explants can be as good in charcoal-free medium as that achieved already on medium containing charcoal (see Sáenz et al., this volume).
is a very immature tissue, shows very little browning and can be cultured without l I ,
395
1 I

Table 1. Quantification of free 2,4-D level in the presence of different charcoal brand.
Free 2,4-D level** (mg rl) in the presence of 2 g 1" of activated charcoal frnm.
Medium age RPQM (México) Acidic washed (Sigma) Neutralised (Sigma)
:! 4days 3.5 f 0.42 o. 12 k 0.02 0.09 f 0.02
8 days 1.36 f 0.28 0.08 0.02 0.03 f 0.02
Figures shown are means f SD (n = 5). Initial [2,4-D] = 66 mg I-'. ** Not adsorbed by charcoal.
3. Studies on growth reguiators
3.1. Studies of growth regulators adsorption by activated charcoal (Partners involved: CICZ Wye Collège, PCA and ORSTOMCIRAD)
In parallel with experiments to test compounds that could substitute charcoal (described in 2.2), research has been carried out to explain the behaviour of added auxins and cytokinins in the presence of charcoal. In Wye College and CICY using radiotracer-based techniques (Ebert et al., 1993), and in Montpellier using an HPLC-based quantification method developed by ORSTOM-CIRAD (Verdeil, 1993). The results confirmed the strong adsorption of 2,4-D and cytokinins by charcoal, and showed that:
a.
b.
C.
d.
Growth regulator adsorption by charcoal is a progressive phenomenon that reaches an equilibrium. Freshly prepared media of uncertain composition should not be used. Instead media should be kept 10-15 days until a steady level of plant growth regulators is available. The adsorption capacity is affected by charcoal particle size and can depend on the charcoal brand used. It can also be different from one batch to another for the same brand (Table 1). The free auxin level can be modified by the introduction of a cytokinin (i.e. BAP, iPA) (Table 2). As in tissue culture the choice of a new medium composition is based on the auxin-cytokinin ratio that can be modified by the introduction of charcoal. Quantification of free auxin and cytokinin levels is essential for a better control of in vitro morphogenesis. As 2,4-D can be released from activated charcoal particles, it seems that in some conditions activated charcoal can
3 96
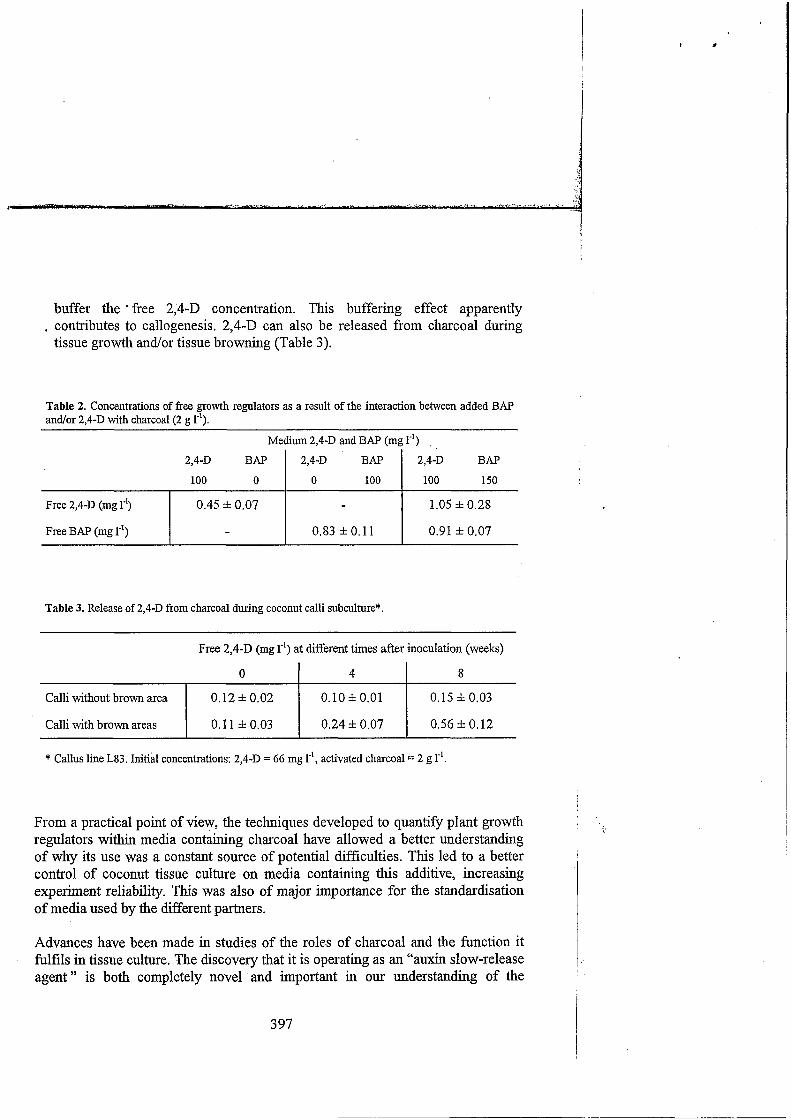
buffer the * free 2,4-D concentration. This buffering effect apparently . contributes to callogenesis. 2,4-D can also be released from charcoal during
tissue growth and/or tissue browning (Table 3).
Free 2,4-D (mg r’) Free BAP (mg 1-’)
Table 2. Concentrations of free growth regulators as a result of the interaction between added BAP and/or 2,4-D with charcoal (2 g 1-I).
0.45 f 0.07 1.05 f 0.28
- 0.83 f O. 11 0.91 f 0.07
Medium 2,4-D and BAP (mg I-’) .
2 ,4D BAP
100 O BAP I 100 150 100
2,4-D 2,4-D
Calli without brown area
Calli with brown areas
o. 12 f 0.02 0.10 f 0.01 0.15 f 0.03
0.1 1 f 0.03 0.24 f 0.07 0.56 f 0.12
Table 3. Release of 2,4-D from charcoal during coconut calli subculture*.
Free 2,4-D (mg le’) at different times after inoculation (weeks)
O I 4 I 8
* Callus line L83. Initial concentrations: 2 , 4 D = 66 mg r’, activated charcoal = 2 g 1-’
From a practical point of view, the techniques developed to quan?ify plant growth regulators within media containing charcoal have allowed a better understanding o f why its use was a constant source of potential difficulties. This led to a better control of coconut tissue culture on media containing this additive, increasing experiment reliability. This was also of major importance for the standardisation of media used by the different partners.
Advances have been made in studies of the roles of charcoal and the function it fulfils in tissue culture. The discovery that it is operating as an “auxin slow-release agent” is both completely novel and important in our understanding of the
3 97

: ” n
beneficial gffects of activated charcoal. This is of a wider interest for tissue culturists in general (Homung and Verdeil, this volume).
3.2. Studies of iiietabolisnz and the interaction of growth regulators during somatic eiiibïyogenesis
a. Fate of 2,4-0 in coconut tissues cultured in vitro (Partners involved: CICY and Wye College)
‘f
The synthetic auxin 2,4-D is required in coconut in vitro culture to promote morphogenic responses such as callus formation and somatic embryo development but very little is known of what is the fate of this auxin in coconut tissues that might determine success or failure in tissue evolution. Investigations were designed to try to clarify this gap in our knowledge. Studies on 2,4-D uptake and metabolism have been done at Wye College and CICY with inflorescence, plumule and immature leaf explants, using radiolabelled synthetic auxin.
Results obtained in charcoal-free medium, showed that coconut tissues take up 14C-2,4-D very rapidly. Data support the hypothesis that explant 2,4-D uptake depends on acidic pH, involving a requirement of energy for pH acidification and uptake. It was found that medium sucrose is a major factor in promoting 2,4-D metabolism and tissue uptake. Up to one-third of the auxin was metabolised, presumably to sugar conjugates that accumulate in the tissues. Accumulated polar metabolites, such as the ß-D-glucose ester, have been reported in other species and are believed to be important as reservoirs of auxins. Regarding 2,4-D uptake, sucrose is required as an energy source. Probably this explains why high sucrose levels reduce the concentration of 2,4-D required for promoting morphogenesis (Magnaval, 1995). This information is very useful since any way of reducing the levels of 2,4-D in the media formulations would be extremely important to avoid or limit the detrimental auxin effects (this synthetic auxin is also known to promote somaclonal variation).
b. Endogenous plant growth regulators (Partner involved: ORSTOM- CIRA D)
An important point for the mastering of coconut regeneration is to understand the physiology of the coconut tissues cultured in vitro. Plant growth regulators are “the” controlling factors of coconut somatic embryogenesis and in this context it is essential to characterise the endogenous equilibrium of hormones in the somatic structures. A better knowledge of the hormonal status of the embryos would be
398

very useful to manipulate their culture. Therefore, it was decided to use zygotic embryo as a model for the characterisation of hormonal stat& during embryo development.
The technique used is based on the separation of different hormonal forms by HPLC and their quantification by an immunoenzymatic ELISA method. Analyses were carried out on nuts at different developmental stages and also on embryogenic and non-embryogenic calli. Data revealed the presence of different forms of cytokinins in the liquid and solid endosperm as well as in the zygotic embryo. The high levels of endogenous cytokinins found suggest their possible involvement in the accumulation of reserves and their mobilisation during germination. Moreover, given the high concentrations detected in somatic structures with high embryogenic potential it can be postulated that cytokinins may play an important role during the initiation of somatic embryogenesis.
-
These studies support the idea that cytokinins play an important role in coconut embryogenesis of both zygotic and somatic types. This information will eventually enable us to fme tune the protocols to optimise coconut regeneration.
4. Histological and protein markers of embryogenic competence
Because morphogenetic events occur slowly during coconut tissue in vitro culture (it takes around 2 years from the frrst culturing operation to the appearance of plantlets), the search for early markers of embryogenesis is particularly important in order to obtain successful regeneration and thus to avoid keeping non- responsive tissues in culture.
4. I . Histological markers of enzbryogenic competence (Partners involved: CICY, PCA, Hannover Universi& Wye College , ORSTOM/CIRAD)
Routine histological evaluation is important to detect early signs of somatic embryogenesis in calli and to rapidly observe the effect of a specific treatment. Samples were treated according to previously described histological techniques (Buffard-Morel et al., 1992). The sections were double stained with the periodic acid-Schiff reaction, associated with protein-specific naphthol blue-black (Fisher, 1968). Studies were conducted on in vitro cultured tissues from CICY, PCA, University of Hannover and ORSTOM-CIRAD. Studies performed at PCA and CICY in collaboration with ORSTOM on plumular tissues allowed us:
399

n
a. to localise callus origin (first divisions occurred in pmvascular strands of the leaf primordia);
b. to confirm the high embryogenic potential of calli obtained according to the Wye College protocol. (Hornung, 1995); and
c. to show that embryogenic structures have a multicellular origin (they are formed by the fragmentation of the outer cambium-like zone, normally ensuring calli growth (Chan et al., 1998). !,
Single embryogenic cell individualisation could also be identified but this route led to abortion after the first few embryogenic cell divisions. This type of embryogenic pathway could be interesting for obtaining embryogenic suspensions.
Concerning immature inflorescences, it could be demonstrated that TDZ and 2iP promote shoot meristem differentiation in somatic embryos. Calli and cultures obtained in Hannover on a charcoal-free medium were analysed in Montpellier. The histocytological features generally present in embryogenic calli obtained with charcoal were not observed in &e nodules studied. It seems that some of the meristematic tissues could have potential for superficial shoot meristem organogenesis. However the regeneration potential of tissues formed with the TDZ/PVP is lower compared with tissues cultured on the 2,4-D/charcoal combination.
4.2. Protein markers (Partners involved: Hannover Universi& CICY)
The objective of this study was to test for the presence of specific markers of early somatic embryogenesis in coconut tissues. These markers were available in Hannover and were identified as being species independent. The study started in Hannover in 1997 with the tissues obtained in Hannover but also with material sent by PCA, CICY and ORSTOM-CIRAD, covering a wide range of development stages (from the starting explant to germinated embryos. Three different antibodies were used: 7C5 directed against a protein marker for early embryogenesis (Altherr et al., 1993), TIM 11 B2 recognising a 40 D a glycoprotein (marker of cell elongation) (Reinard et al., 1994) and -Glo-I recognising Glyoxalase I (an enzyme involved in the control of cell-proliferation and cell-differentiation processes) (Thornally, 1990). Results showed that following screening with the 7C5 antibody (marker of early embryogenesis) a strong signal was present in some of the embryogenic structures sent to Hannover. (for more details see Köhne and Jacobsen, this volume).
400

I
I In all the samples screened with the -Glo-I antibody (marker for cell proliferation) a signal was always detected independent of the source and the treatment; however this signal was always weak. This could be related to the very low mitotic index that is generally observed in coconut, and characteristic of the very slow growing in vitro cultures of this species.
Among the markers available in Hannover no candidates could be selected for the early identification of coconut tissues showing competence for somatic embryogenesis.
Complementary to the research done on protein markers an interesting study was initiated in CICY to examine whether protein phosphorylation occurs on tyrosine residues during coconut zygotic embryo development. Protein phosphorylation- dephosphorylation is indeed thought to be one of the major mechanisms regulating cell division and differentiation in microorganisms, animal cells and plant cells. Results obtained in CICY showed the occurrence of kinase activities for the phosphorylation of threonine, serine and tyrosine residues in coconut tissues of developing zygotic embryos (Islas-Flores et al., 1998). A study performed with in vitro cultured plumular tissues showed that some of these proteins might have a potential use as markers of embryogenic capacity.
Further studies are required to find useful markers that will help us to select embryogenic tissues and monitor their response to growth regulator treatments. results obtained by CICY on phosphorylated proteins have already opened up promising lines of research.
5. Studies of vitroplant physiology using zygotic in vitro germinating embryos as a model (Partners involved : PCA, CICY, ORSTOM/CIRAD)
In coconut, in vitro development and acclimatisation of plantlets constitute a major bottleneck for the implementation of tissue culture protocols. Foliar development appears to be very slow and is sometimes accompanied by leaf yellowing. Therefore, experiments were conducted to assess the physiological competence of the plantlet obtained and particularly its ability to control excessive water loss (when transferred to ex vitro conditions), its photosynthetic capabilities and thus its capacity for acclimatisation.
i '
40 1

402
Work to investigate the physiological status of shoots obtained in vitro was conducted using in vitro germinated zygotic embryos as a model system. Net photosynthesïs rate was estimated through COZ exchange measurements on intact leaves (Triques et a1.,1997a) Chlorophyll fluorimeby, a non destructive and highly sensitive method, was used to analyse the potential activity (OpMAx) and the actual efficiency (0p) of photosystem II. To characterise carbon metabolism, carboxylase (Phosphoenolpyruvate carboxylase, PEPC; and Ribulose 1,5- biphosphate carboxylase/oxygenase, RubisCO) activities were measured in crude extracts and the PEPC:RubisCO ratio was used as an indicator of autotrophic CO2 fixation. Quantification of the RubisCO protein was performed using immunoelectrophoresis. The development of the photosynthetic apparatus was assessed by TEM observation of leaf chloroplast ultrastructure, (Triques et al.,1997b)
!
In vitro cultured coconut displayed an early initiation of a photosynthetic metabolism. Concomitant changes in several parameters (OpMm, Op, PEPC:RubisCO ratio and transpiration rates) were measured. However a lower rate of net photosynthesis was recorded in in vitro grown plantlets as compared with the acclimatised palm. This could be explained by a lower RubisCO content and activity together with a lower chlorophyll content compared to the acclimatised palm (Triques et al., 1998).
The effect of sucrose in the medium on the photosynthetic capacity of coconut vitroplants derived from zygotic embryos has been studied by J, Santamaría (CICY) in Montpellier (see Santamaría et al, this volume). It was shown that sucrose decreased the activity of RubisCO in tissues, while slightly increased the activity of PEPC. Since PEPC/RubisCO is taken as a measure of plant photoautotrophy (see Desjardins, 1995), these results suggest that sucrose inhibits the development of vitroplant photoautotrophy. They also suggest that, while sucrose might be important in early stages of coconut embryo cultures, the continuous growth in sucrose-rich medium in later stages during plantlet growth, will affect the development of photoautotrophy in coconut vitroplants, which in turn can affect plant performance when transferred ex vitro.
In studies carried out at CICY, coconut vitroplants (derived from zygotic embryos) showed a reduced capacity to control water loss relative to that shown by field- grown plants. This is due to altered stomatal functioning. Ventilation of the culture containers resulted in an increased capacity of vitroplants to control water loss, similar to that shown by field-grown plants.

These results are promising as a means of in vitro hardening and acclimatisation ’ that is today the major bottleneck for the establishment of clone field trials or
demonstrative plots. Knowledge of vitrbplant physiology has to be increased to help us to improve in vitro shoot growing conditions and will facilitate plantlet transfer to ex vitro culture conditions.
6. Conclusion
Solid progress for the mastery of coconut regeneration has been made in the last three years under this project. The partners have exchanged information (through a publication: Coconut Newsletter) and techniques (through trainings and publication of a compendium of the protocols) thus enabling us to improve our respective protocols. Since coconut regeneration is a slow phenomenon (the actual process from inflorescence explants takes at least three years), it would be too early at this stage to accurately quanti@ the impact of this project on the improvement of each regeneration phase. Nevertheless, in a tentative assessment, taking into account the data available so far using the ORSTOMKIRAD protocol, the efficiencies of embryogenesis induction and embryo maturation have been at least doubled. Although, there is still room to improve, prototype regeneration systems from plumular tissues and fiom immature inflorescence are now available, and clonal plantlets are produced now in the project participating laboratories.
Research is currently focused on somatic embryo germination (current germination rate is about 5%), improvement of plantlet growth and the establishment of plantlets in the greenhouse (still a limiting step for further ex vitro development).
Coconut micropropagation requires long-term research programmes, with guaranteed financial support that several countries cannot reliably provide to their research institutions, and therefore, will depend on international assistance (see Batugal, and Concluding Remarks, this volume). The project reported here demosntrated that intemational cooperation contributes to increase research efficiency, avoiding duplication of work and promoting complementation and synergy of activities. However, in order to master the late phases of coconut micropropagation (plantlet growth and acclimatisation) in the shortest time possible, a continued collaborative effort from the different groups will be required, as well as further international assistance.
403

1.
2.
?3,
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
References
Altherr S, Stirn S and Jacobsen HJ (1993). Immunobiochemical analysis of a nuclear protein marker for regeneration potential in higher plants. J Plant Physiol 141:415-422. Blake J and Eeuwens CJ (1980). Inflorescence tissue as source material for vegetative propagation of coconut palm. In: Proc. 1ntem.Conf. Cocoa/Coconuts, Kuala Lumpur, pp 549-556. Incorporated Society of Planters, Kuala Lumpur. Blake J (1990). Coconut (Cocos nucifra L.) Micropropagation. In: YPS Bajaj (Ed.). Biotechnology in Agriculture and Forestry, vol. 10, Legumes and Oil Seed Crops, pp 538-554. Springer-Verlag,
BuEard-More! J, Verdeil JL and Pannetier C (1992). Embryogenkse somatique du cocotier (Cocos nucifra L.) A partir de tissus foliaires: étude histologique. Can J Bot 70:735-741. Chan JL, Saénz L, Talavera C, Homung R, Robert M and Oropeza C (1998). Regeneration of coconut (Cocos nucifra L.) from plumule explants through somatic embryogenesis. Plant Cell
Desjardins Y (1995). Factors affecting CO2 fixation in striving to optimize photoautotrophy in micropropagated plantlets. Plant Tiss Cult Biotech 1: 13-25.
concentrations in plant tissue culture media in the presence of activated charcoal. Plant Cell Tiss Org
Ebert A, Taylor FH and Blake J (1993). Changes of 6-benzylaminopurine and 2,4- dichlorophenoxyacetic acid concentrations in plant tissue culture media in the presence of activated charcoal. Plant Cell Tiss Org Cult 33:157-162. Fisher DB (1968). Protein staining of ribboned epon sections for light microscopy. Histochemie
Hornung R (1995). Initiation in callogenesis in coconut palm (Cocos nucfera L). In: C Oropeza, FW Howard, GR Ashburner (Eds.) Lethal yellowing research and practical aspects, pp 203-215. Kluwer Academic Publishers, Dordrecht. Islas-Flores I, Oropeza C and Hernández-Sotomayor SMT (1998). Protein phosphorylation during coconut (Cocos nucifera L.) zygotic embryo development. Plant Physiol 118 :257-263. Magnaval C (1995). Etude des facteurs nutritifs intervenant lors de l'initiation de 'embryogenèse somatique du cocotier (Cocos nucifra L.) en milieu gélosé et mise au point des suspensions embryogènes. PhD thesis, Université Paris Vi. Paris. Persley G (1992). Replanting the Tree of Life. Towards an international agenda for coconut palm research. CAB Intemational, Wallingford, UK. Reinard T, Sprunck S, Altherr S and Jacobsen HJ (1994). Biochemical properties of a novel cell wall
I
Heidelberg. 1
I
Reports 17:5 15-52 1. I !
Ebert A and Taylor HF (1990). Assessment of the changes of 2,4-dichlorophenoxyacetic acid I Cult 20:165-172.
16:92-96.
protein associated with elongation growth in higher plants. J Exp Bot 45:1593-1601. 15. Ri110 EP, Cueto CA, Orense OD, Area MBB. and Ebert AW (1992). Recent development on embryo
culture technology of coconuts. Phil J Crop Science 17 (Suppl. N01):S30. 16. Thornally PJ (1990). The glyoxalase system: new developements towards functional characterisation
of metabolic pathway fundamental to biological life. Biochem J 269:l-11. 17. Triques K (1996). Comportement des embryons zygotiques de cocotier (Cocos nucifra L.) cultivés in
vifro: Nutrition et acquisition de l'autotrophie. DEA, Bases de la production végétale, Université Montpellier II, 13p.
18. Triques K, Rival A, Bedé T, Dussert S, Hocher V, Verdeil JL and Hamon S (1997a) Developmental changes in carboxylase activities in in vitro cultured coconut zygotic embryos: comparison with corresponding activities in seedlings. Plant Cell Tiss Org Cult 49:227-231.
19. Triques K, Rival A, Beult T, Puard M, Roy J, Nato A, Lavergne D, Havaux M, Verdeil JL, Sangare A and Hamon S (1997b). Photosynthetic ability of in vifro grown coconut (Cocus nucifra L.) plantlets derived from zygotic embryos. Plant Sci 127:39-51.
404 I

20. Triques K, Rival A, Bedé T, Morcillo F, Hocher V, Verdeil JL and Hamon S (1998). Changes in photosynthetic parameters during in vifro growth and subsequent acclimatization of coconut (Cocos nuciferu L.) zygotic embryos. Acta Hort 461: 275-282.
21. Verdeil JL and Buffard-Morel J (1995). Somatic embryogenesis in coconut (Cocos nuciferu L.). In! YPS Bajaj (Ed.). Biotechnology in Agriculture and Forestry, vol. 30. Somatic embryogenesis and Synthetic Seed, I, pp 299-3 17. Springer-Verlag, Heidelberg.
22. Verdeil JL, Buffard-Morel J, Dussert S, Rival A, Grosdemange F, Huet C and Pannetier C (1993). Coconut clones through somatic embryogenesis. In: MK Nair, HH Khan, P Gopalasundaram and E W Bhaskara Ra0 (Eds.). Advances in Coconut Research and Development, pp 173-179. Oxford & IBH, New Delhi.
23. Verdeil JL, Huet C, Grosdemange F and BuEard-Morel J (1994). Plant regeneration from cultured immature inflorescences of coconut (Cocos nucifera L.): evidence for somatic embryogenesis. Plant Cell Rep 13:218-221.
24. Verdeil JL (1993). Etude de la régénération du cocotier (Cocos nucifra L.) par embryogenèse somatique à partir d'explants intlorescentiels. PhD thesis, l'université P. et M. Curie (Paris VI), 156p.
405

Current Advancesin Coconut Biotechnology
Edited by
C. OROPEZA Centro de Investigación Cientpca de Kicatríiz (Méricìci)
J.L. VERDEIL ORSTOM- CIRA D (Mon tpellicr)
G.R. ASHBURNER I I 1st it 1 it e of Si isto i11 able Irrigated Ag rici1 It i 1 re (Tat 11 ieri)
and
J.M. SANTAMARÍA C n I /)'o de I ì I I v s t ig cic i61 i Ciei I t $ ca de K r cn tr i i I (Mérida)
KLUWER ACADEMIC PUBLISHERS DORDRECHT / BOSTON / LONDON
!I-
if: ;
. . . . , . . . . : . . . . . . .. . ' . . ;:. .:
. . . . . ' . . . . . .. . .
. . . . ._ . .
. . . . , . . . . .

ISBN 0-7923-5823-6
Published by Kluwer Academic Publishers, P.O. Box 17,3300 AA Dordrecht, The Netherlands.
Sold and distributed in North, Central and South America by Kluwer Academic Publishers, 101 Philip Drive, Norwell, MA 02061, U.S.A.
In a11 other countries, sold and distributed by Kluwer Academic Publishers, P.O. Box 322,3300 AH Dordrecht, The Netherlands.
All Rights Reserved O 1999 Kluwer Academic Publishers No part of the material rrotected by this copyright notice may be reproduced or utilized in any form or h;: any means, electronic or mechanical, j including photocopying. recording or by any information storage and retrieval system. withoui written pcrmission from the copyright owner.
Printed in the Netherlan:.s.
I

B l-4' b
Current Plant Science and Biotechnology in Agriculture VOLUME 35
Scientijc Edifor R.J. Summerfield, The University of Renditig, Department ofAgricirlture, P.O. Box 236, Rending RG6 2A1; Berkshire, UK
Scientific Advisoty Board D.F. Bezdicek, \Vashitiglon State Uniiwsiy , Piillmati, USA J. Denecke, University of York, York, UK J, Hamblin, The University of kvesterti Aiutmlin, Nedlands, AiWrdia H.-J. Jacobsen, Utiiversitiit Hannover. Hatinorper; Gertnnny
Aims and Scope The book series is intended for readers ranging from advanced students to senior research scientists and corporate directors interested in acquiring in-depth, state-of-the-art knowledge about research fin- dings and techniques related to all aspects of agricultural biotechnology. Although the previous volu- mes in the series dealt with plant science and biotechnology, the aim is now to also include volumes dealing with animal science, food science and microbiology. While the subject matter will relate more particularly to agricultural applications, timely topics i n basic science and biotechnology will also be explored. Some volumes will report progress in rapidly advancing disciplines through proceedings of symposia and workshops while others w i l l detail fundamental inFormation of an enduring nature that will be referenced repeatedly.
The titles priblidied in this series n-re listed ot the end of this voliittie.
... . .
.. . ....' ':'!;
. .. ' .. .. ... . . . ... , . , ..' . ' ._ . . ., __...
