reconstitution of influenza virus rna … binding mode of influenza virus nucleoprotein to rna tive-...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 265, No. 19. Iasue of July 5, pp. 11151-11155, 1990 0 1990 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A.
Reconstitution of Influenza Virus RNA-Nucleoprotein Complexes Structurally Resembling Native Viral Ribonucleoprotein Cores*
(Received for publication, March 5, 1990)
Kunitoshi Yamanaka@, Akira Ishihama$, and Kyosuke Nagata$ll From the *Department of Molecular Genetics, National Institute of Genetics, Mishima, Shizuoka 411 and the SDepartment of Genetics, Osaka University School of Medicine, Naknnoshima, Kituku, Osaka 530, Japan
Reconstitution of influenza virus nucleoprotein (NP)-RNA complexes was performed with segment 8 RNA, which was synthesized in vitro from cDNA, and NP purified from virions. Under optimum conditions established using a filter binding assay and a gel retar- dation assay, NP was found to bind any RNA longer than 15 nucleotides. NP-RNA complexes formed at 30 “C are more resistant to high concentrations of NaCl than those formed at 0 “C. Treatment of NP with N- ethylmaleimide gave no effect on its RNA binding ac- tivity, whereas treatment with alkaline phosphatase enhanced its RNA binding activity. The newly devel- oped “reverse-printing” method of RNase V 1 -treated complexes revealed that reconstituted NP-RNA com- plexes carry RNase Vl-sensitive sites as do native ribonucleoprotein (RNP) cores (RNA polymerase-NP- RNA complexes), implying that RNA-NP complexes structurally similar to native RNP cores are reconsti- tuted from isolated components.
Influenza virus contains eight segments of single-stranded RNA of negative polarity as the genome (1). In the virion, RNA is associated with RNA polymerase and nucleoprotein (NP)’ forming ribonucleoprotein (RNP) cores (2, 3). RNA polymerase is composed of three species of P protein, PBl, PB2, and PA, and plays central role in transcription and replication of the viral genome (4,5). RNP cores were isolated from detergent-treated influenza virus A/PR/8/34 by glycerol gradient centrifugation (6). RNP cores were dissociated into P-RNA (RNA polymerase-RNA) complexes and NP by CsCl centrifugation (7). The P-RNA complexes were capable of synthesizing only small-sized RNA, which was elongated by the addition of NP, suggesting that NP is required for the elongation of RNA chains. In the presence of excess levels of NP, however, the synthesis of template-sized RNA was not observed. This implied that proper arrangement of NP is needed for the efficient elongation of RNA chains. In addition, it has been suggested that NP is implicated in the switchover from transcriptive to replicative RNA synthesis (8).
In this study, we analyzed the binding mode of NP to RNA. First, we purified NP from virions and optimized the condi- tions for binding of NP to RNA using a filter binding assay
* This work was supported by grants-in-aid from the Ministry of Education, Science, and Culture of Japan. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Y To whom corresuondence should be addressed. Tel.: 0559-75- - 0771 (ext. 636).
’ The abbreviations used are: NP, nucleoprotein; RNP, ribonucle- oprotein; DTT, dithiothreitol; SDS, sodium dodecyl sulfate; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid.
and a gel retardation assay. Reconstituted NP-RNA com- plexes were then analyzed by RNase “reverse-printing” method using RNase Vl. The structure of NP-RNA com- plexes is discussed.
MATERIALS AND METHODS
Virus-Influenza virus A/PR/8/34 (HlNl) was used throughout this study. Virus was grown in allantoic cavity of lo-day-old embryon- ated eggs and purified as described previously (9). The purified virions were suspended in 10 mM Tris-HCI (pH 7.8). 1 mM dithiothreitol (DTT), 20% (w/v) glycerol and stored at -80 “C until use.
Isolation of RNF Cores-Purified virions were treated with a dis- ruption buffer consisting of 100 mM Tris-HCl (pH 8.0), 100 mM NaCl, 5 mM MgC12, 1% lysolecitin, 1% Triton X-100, 1 mM DTT, and 5% glycerol for 10 min at 30 “C, and then directly subjected to centrifu- gation on a 30-60% linear gradient of glycerol on a 70% glycerol cushion in 50 mM Tris-HCl (pH 7.8), 100 mM NaCl, and 1 mM DTT in a Beckman SW 50.1 rotor at 45,000 rpm for 5 hat 4 “C as described previously (6, 7). The distribution of proteins was analyzed by elec- trophoresis on 12.5% polyacrylamide gels in the presence of sodium dodecyl sulfate (SDS) while that of RNAs was analyzed by electro- phoresis on 2.8% gels in the presence of 8 M urea. The gels were stained with a silver reagent (Bio-Rad). Fractions consisting of eight kinds of viral RNA, RNA polymerase, and NP were pooled as RNP cores.
isolation of Nucteoprotein-Isolated RNP cores were directly sub- jected to centrifugation on a discontinuous step gradient of CsCl and glycerol as described previously (7). The gradient was composed of three layers, from top to bottom: 0.5 M CsCl, 20% glycerol; 1.0 M CsCl, 30% glycerol; and 3.0 M CsCI, 50% glycerol. Centrifugation was performed at 45,000 rpm for 25 h at 4 “C in a Beckman SW 50.1 rotor. The distribution of proteins and RNAs was analyzed by gel electrophoresis as described above. The scheme for isolation of NP is summarized in Fig. lA. Isolated NP was dialyzed against 10 mM Hepes-NaOH (pH 7.0), 25 mM NaCl, 5 mM MgQ, 0.5 mM EDTA, 20% (w/v) glycerol, 2.5 mM DTT, and 0.01% Nonidet P-40 at 4 “C for 15 h. Protein concentration was determined by 0.1% SDS, 12.5% polyacrylamide gel electrophoresis and by densitometric scanning of silver-stained gels. Bovine serum albumin was used as a standard.
Preparation of “P-Labeled RNA Template-cDNA for influenza virus genome segment 8 RNA was cloned into pSP65 vector in both orientation relative to the promoter of SP6 RNA polymerase (10). In order to prepare RNA for filter binding assay, the plasmids pSPNSG1 and pSPNSG2 were linearized by digestion with SmaI. To prepare RNA for gel retardation assay, the plasmid pSPNSG2 was linearized by MnlI. RNAs were synthesized by transcribing linearized plasmid DNAs with SP6 RNA polymerase at 37 “C for I h in a final volume of 50 ~1, which contained 40 mM Tris-HCl (pH 7.5), 20 mM NaCl, 10 mM MgCl*, 10 mM DTT, 0.5 mM ATP, 0.5 mM CTP, 0.5 mM GTP, 100 &i of UTP (400 Ci/mmol, Amersham Corp.), 4 mM spermidine, 1000 units/ml RNase inhibitor, and 600 units/ml SP6 RNA polym- erase (11). After electrophoresis on 2.8% polyacrylamide in the pres- ence of 50% urea, synthesized RNAs were eluted from the gel and stored at -80 “C until use. RNA synthesized from SmaI-digested pSPNSG1 was 923 nucleotides tong and of positive polarity, while that from SmaI-digested pSPNSG2 was 923 nucleotides long and of negative polarity. From MnlI-digested pSPNSG2, 446-nucleotide- long RNA of negative polarity was synthesized. Preparation of dou- ble-stranded 32P-labeled RNAs was performed as follows. Both posi-
11151
by guest on March 21, 2019
http://ww
w.jbc.org/
Dow
nloaded from

11152 Binding Mode of Influenza Virus Nucleoprotein to RNA
tive- and negative-strand RNAs were hybridized, treated with Sl nuclease, extracted with phenol/chloroform, and then precipitated
virions of influenza virus A/PR/8/34. Isolated RNP cores
with ethanol (10). were subjected to CsCl centrifugation. NP free of viral RNA
Filter Binding Assay--Synthesized “‘P-labeled RNA was incubated and RNA polymerase was recovered in top fractions of the with purified NP at 30 “C for 10 min in a final volume of 50 ~1, which gradient. Purified NP is shown in Fig. 1B (lanes 1 and 2). contained 10 mM Hepes-NaOH (pH 7.0), 25 mM NaCl, 5 mM MgCl?, The RNA binding activity of NP was examined using 32P- 0.5 mM EDTA, 20% (w/v) glycerol, and 2.5 mM DTT. The mixture labeled segment 8 RNA, which was prepared by transcribing was filtered through a nitrocellulose filter (HAWP, Millipore) and [“YP]RNA retained on the filter was counted with a scintillation
segment 8-cDNA with SP6 RNA polymerase. The purified
counter (12). NP exhibited RNA binding activity as measured by a filter
Gel Retardation Assay--:“P-Labeled RNA was incubated with pu- binding assay (Fig. 2) and a gel retardation assay (Fig. 3). The rified NP at 0 “C for 10 min in a final volume of 10 ~1, which contained RNA binding activity at 0 “C was about one-half the levels at the same ingredients as those for the filter binding assay. The mixture 25, 30, 37, or 42 “C (Fig. 20). As to NaCl concentration, NP- was then subjected to electrophoresis at 4 “C in 4% polyacrylamide gel containing 2.8% (w/v) glycerol, 25 mM Tris-HCl (pH 8.3), 1 mM
RNA complexes were formed at 0.5 M NaCl at 30 “C, while at
EDTA, and 190 mM glycine (13, 14). 0 “C no RNA binding was observed at 0.5 M (Fig. 2E). This
RNuse A Protection Assay--‘“P-Labeled RNA was incubated with suggests that the binding mode is different between at 0 and
purified NP at 30 “C for 10 min as described above. After addition of 30 “C. The maximum binding was achieved at 125 mM NaCl RNase A, the reaction mixture was further incubated at 30 “C for 5 at both temperatures. The optimum condition of pH was min. RNA was extracted with phenol/chloroform, precipitated with broad between 5.0 and 8.0 (Fig. 2F). ethanol, and analyzed by electrophoresis on 6% polyacrylamide gel Pretreatment of NP at 90 “C for 3 min resulted in loss of containing 50% urea.
RNase Reverse-printing Assay-Oligonucleotide, 5’-GGGTGACA- AAGACATAATG-3’, complementary to the region near the 3’ ter- minus of influenza virus genome segment 8 RNA (15) was synthesized with an Applied Biosystems model 380B DNA synthesizer, and la- beled at the 5’ end with T4 polynucleotide kinase and [y-‘LP]ATP (>7000 Ci/mmol, ICN Radiochemicals). Influenza virus genome RNA was extracted from purified virions and incubated with purified NP as described above. These reconstituted NP-RNA complexes and native RNP cores were treated with 5 units/ml RNase Vl at 30 “C for 5 min in a final volume of 20 gl, which contained 20 mM Tris- HCl (pH 7.5) and 125 mM NaCl. RNA was extracted with phenol/ chloroform, and precipitated with ethanol. RNA was hybridized with the lLP-labeled oligonucleotide and incubated with 500 units/ml re- verse transcriptase at 42 “C for 1 h in a final volume of 20 ~1, which contained 50 mM Tris-HCI (pH 8.3), 10 mM MgCl*, 10 mM DTT, 60 mM NaCl, 1 mM each of dATP, dGTP, dCTP, and TTP. Elongated DNA were analyzed by electrophoresis on 6% polyacrylamide gel containing 50% urea. The scheme of the RNase reverse-printing assay is summarized in Fig. 7A. Reverse transcriptase was purified from avian myeloblastosis virus as described previously (16).
Chemicals and Enzymes-All restriction enzymes used were pur- chased from Takara Shuzo, Japan. SP6 RNA-polymerase was- ob- tained from Boehrineer Mannheim. RNase Vl and unlabeled nucle- otides from PharmaGa LKB Biotechnology Inc., [a-“2P]UTP (>400 Ci/mmol) from Amersham Corp., and [r-“P]ATP (>7000 Ci/mmol) from ICN Radiochemicals. All reagents used in this study were analytical grade.
RESULTS
Biochemical Parameters for RNA Binding Activity of NP Purified from Virions-The scheme for isolation of NP is summarized in Fig. 1A. Briefly, RNP cores consisting of viral RNA, RNA polymerase, and NP were first isolated from
.--- - -
NP P-RNA complexes
FIG. 1. Purification of NP protein from influenza virus. A, purification procedures are summarized. Details were described under “Materials and Methods.” B, purified NP was analyzed by electro- phoresis on 12.5% polyacrylamide in the presence of 0.1% SDS (lanes I and 2). As a standard, bovine serum albumin was also shown (lanes 3-7, 1, 0.75, 0.5, 0.25, and 0.1 rg, respectively).
its RNA binding activity, while pretreatment at 37 “C gave no effect (Table I). Pretreatment of NP with N-ethylmaleim- ide, a SH-blocking agent, gave no effect on its RNA binding activity (Table I), suggesting that an S-S bond is not involved in the RNA binding. When NP was treated with bacterial alkaline phosphatase, the RNA binding activity was slightly increased (Table I). It is possible that virus-associated NP is phosphorylated, and that phosphorylation of NP regulates its RNA binding activity and thereby controls the transcription and/or replication (17).
RNA Specificity of NP Binding-Using the optimized con- ditions, we tested the effect of RNA polarity on the binding of NP. As shown in Fig. 2, A-C, NP binds equally well to both the negative and positive strand of viral RNA. The binding activity was less to double-stranded RNA than single- stranded RNA. For all the segment 8 RNAs tested, a sigmoidal binding character was observed, suggesting that NP binds cooperatively to RNA. This is also supported by the gel retardation assay (Fig. 3). At low concentrations of NP (lanes 2-4), only one large NP-RNA complex band was formed but no intermediate complexes appeared even though unbound RNA remained. Addition of ATP leads to the formation of intermediate complexes (lanes 8 and 9). It is, however, unclear how ATP alters the binding mode of NP to RNA.
To elucidate the sequence specificity of NP binding, we carried out a competition filter binding assay with poly- and oligonucleotides. Fig. 4 shows the RNA binding activity of NP as a function of increasing amount of poly(A) as a com- petitor. The 50% competition was observed in the presence of approximately IOOO-fold excess poly(A) in weight. The competition filter binding assay was carried out using other competitors at the competitor:RNA ratio of 1000 (Table II). Poly(C), poly(U), and heat-denatured calf thymus DNA com- peted more than poly(A). Other RNA and DNA exhibited the competition activity as high as that of poly(A).
The effect of RNA chain length on the RNA binding activity of NP was examined (Fig. 5A). Labeled RNA was partially digested with RNase A and then incubated with NP. Filter-trapped RNA was then analyzed by polyacrylamide gel electrophoresis in the presence of urea. The ratio of trapped RNA (lane 3) to input RNA (lane 1) were shown in Fig. 5B after densitometric scanning of the autoradiograms. It is clear that NP binds to RNA longer than 15 nucleotides. The weak competition with oligo(dT)12-18 might therefore be due to its short chain length.
Binding Mode of NP to RNA-Vesicular stomatitis virus RNA-nucleoprotein complexes are totally resistant to RNase digestion (18), while influenza virus RNP complexes are par-
by guest on March 21, 2019
http://ww
w.jbc.org/
Dow
nloaded from
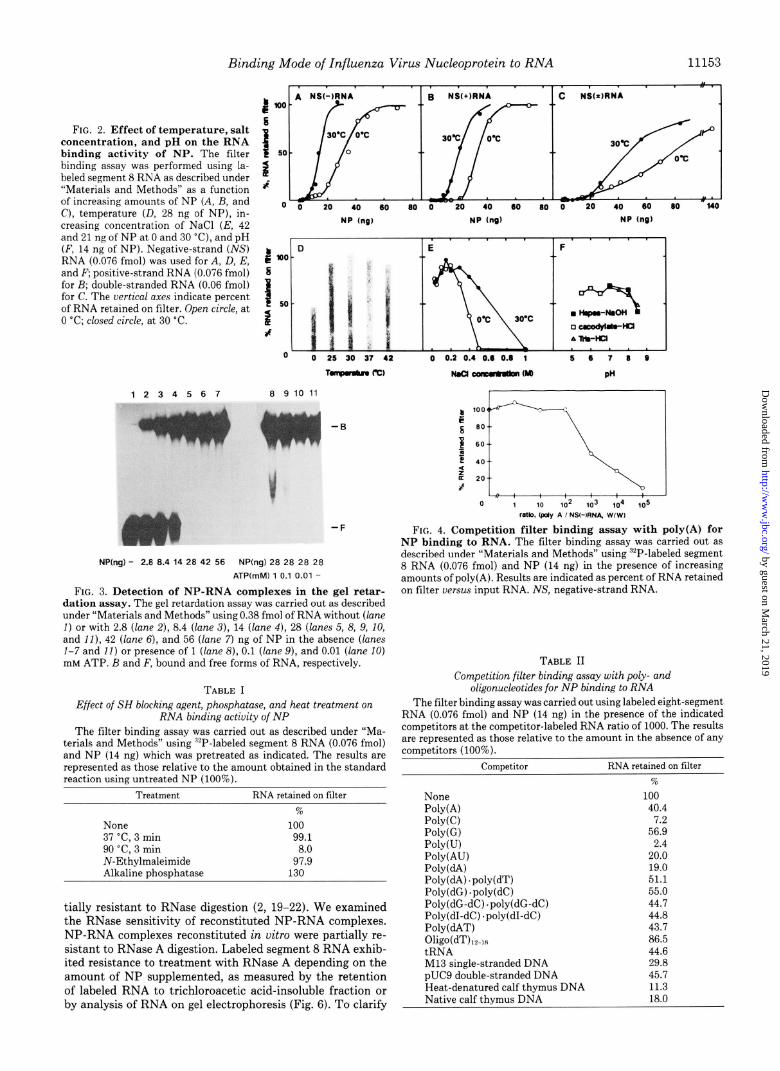
Binding Mode of Influenza Virus Nucleoprotein to RNA
FIG. 2. Effect of temperature, salt concentration, and pH on the RNA binding activity of NP. The filter binding assay was performed using la- beled segment 8 RNA as described under “Materials and Methods” as a function of increasing amounts of NP (A, B, and C), temperature (D, 28 ng of NP), in- creasing concentration of NaCl (E, 42 and 21 nr of NP at 0 and 30 “C), and DH
$ loo s
1 50
2 af
o 0 20 40 60 60 0 20 40 60 60 0 20 40 60 60 l40
NP (n61 NP (n6) HP (ng)
(F, 14 niof NP). Negative-strand (ihjS) b RNA (0.076 fmol) was used for A, D, E, and F; positive-strand RNA (0.076 fmol)
: 10’
for B; double-stranded RNA (0.06 fmol) for C. The vertical axes indicate percent of RNA retained on filter. Open circle, at 1
5.
0 “C; closed circle, at 30 “C. 2 J
0 0 25 30 37 42 0 0.2 0.4 0.6 0.6 1 5 6 7 6 0
1234567 6 9 10 11
.F
NP(ng) - 2.6 6.4 14 28 42 56 NP(ng) 28 28 28 28
ATPtmM) 1 0.1 0.01 -
FIG. 3. Detection of NP-RNA complexes in the gel retar- dation assay. The gel retardation assay was carried out as described under “Materials and Methods” using 0.38 fmol of RNA without (lane I) or with 2.8 (lane 2), 8.4 (lane 3), 14 (lane 4), 28 (lanes 5, 8, 9, IO, and II), 42 (lane 6), and 56 (lane 7) ng of NP in the absence (lanes l-7 and II) or presence of 1 (lane 8), 0.1 (lane 9), and 0.01 (lane 10) IIIM ATP. B and F, bound and free forms of RNA, respectively.
TABLE I
Effect of SH blocking agent, phosphamse, and heat treatment on RNA binding actiuity of NP
The filter binding assay was carried out as described under “Ma- terials and Methods” using “P-labeled segment 8 RNA (0.076 fmol) and NP (14 ng) which was pretreated as indicated. The results are represented as those relative to the amount obtained in the standard reaction using untreated NP (100%).
Treatment RNA retained on filter
None 37 “C. 3 min
% 100
99.1 90 ‘Cl 3 min N-Ethylmaleimide Alkaline Dhosnhatase
8.0 97.9
130
tially resistant to RNase digestion (2, 19-22). We examined the RNase sensitivity of reconstituted NP-RNA complexes. NP-RNA complexes reconstituted in vitro were partially re- sistant to RNase A digestion. Labeled segment 8 RNA exhib- ited resistance to treatment with RNase A depending on the amount of NP supplemented, as measured by the retention of labeled RNA to trichloroacetic acid-insoluble fraction or by analysis of RNA on gel electrophoresis (Fig. 6). To clarify
FIG. 4. Competition filter binding assay with poly(A) for NP binding to RNA. The filter binding assay was carried out as described under “Materials and Methods” using “P-labeled segment 8 RNA (0.076 fmol) and NP (14 ng) in the presence of increasing amounts of poly(A). Results are indicated as percent of RNA retained on filter uersu.s input RNA. NS, negative-strand RNA.
TABLE II
Competition filter binding assay with poly- and oligonucleotides for NP binding to RNA
The filter binding assay was carried out using labeled eight-segment RNA (0.076 fmol) and NP (14 ng) in the presence of the indicated competitors at the competitor-labeled RNA ratio of 1000. The results are represented as those relative to the amount in the absence of any comuetitors (100%).
ComDetitor RNA retained on filter
None PolyW POlY(C) POlY(G) POlY(U) Poly(AU) POlY(dA) Poly(dA) .poly(dT) Poly(dG) .poly(dC) Poly(dG-dC) .poly(dG-dC) Poly(dI-dC) .poly(dI-dC) Poly(dAT) Oligo(dT)l?~lH tRNA Ml3 single-stranded DNA pUC9 double-stranded DNA Heat-denatured calf thymus DNA
% 100
40.4 1.2
56.9 2.4
20.0 19.0 51.1 55.0 44.7 44.8 43.7 86.5 44.6 29.8 45.7 11.3
Native calf thymus DNA 18.0
by guest on March 21, 2019
http://ww
w.jbc.org/
Dow
nloaded from

1123
Binding Mode of Influenza Virus Nucleoprotein to RNA
12 14 16 16 20
RNA length (nucleotlder)
c
FIG. 5. Effect of RNA chain length on RNA binding activity of NP. A, ‘LP-labeled segment 8 RNA (7.6 fmol) was partially digested with RNase A (lane 1). These partial digests were incubated without (lane 2) or with 42 ng of NP (lane 3) and subjected to the filter binding assay. Filter-trapped RNA was eluted from filter and then analyzed by electrophoresis on 6% polyacrylamide gel containing 50% urea. “P-Labeled oligonucleotides (MI), “‘P-labeled Hoe111 fragments of pBR322 (M2), and “‘P-labeled RNA treated with NaOH (M3) were used as size markers. B, after densitometric scanning of the autora- diograms, the ratio of trapped RNA to input RNA was calculated and plotted for each size of RNA.
whether the binding mode of NP to RNA is the same as that of native RNP cores, we developed a reverse-printing method (Fig. 7A). Viral RNA extracted from virions was incubated in the presence or absence of NP, and then subjected to limited digestion with RNase Vl, which recognizes double-stranded RNA or helical RNA (23). Digested RNA was extracted and the primer extension analysis was carried out using the spe- cific primer for the 3’ terminus of segment 8 viral RNA. Fig. 7B shows the result of the reverse-printing analysis for RNase W-treated native RNP cores. Major RNase Vl-sensitive sites were visualized approximately every 15 nucleotides (lanes 1 and 2). Such a periodical digestion pattern was not observed using either RNA or NP alone (Fig. 7C, lanes 3-6). In contrast, RNase W-sensitive sites were detected in the NP-RNA com- plexes reconstituted in vitro (Fig. 7C, lanes 1 and 2). Together, these results suggest that NP-RNA complexes reconstituted in vitro as well as the native RNP cores contain the higher ordered structure, which is recognized by RNase Vl.
DISCUSSION
RNP cores of influenza virus support the synthesis of full- sized RNA in vitro (6). P-RNA complexes devoid of NP, however, are capable of synthesizing only small-sized RNA, which can be elongated by the addition of NP (7). These results suggest that NP is involved in the elongation of RNA chains. To reveal the function of NP in these processes in detail, it is important to characterize the mode of its RNA binding. In this paper, we optimized the conditions for binding of NP to RNA by using both a filter binding assay and a gel retardation assay (Figs. 2 and 3). Competitive RNA binding assays using oligo- and polynucleotides indicated that NP is a polynucleotide binding protein with higher affinity to single- stranded than to double-stranded polynucleotides. The bind-
1234567M
1 I i 1 Jk
NPlngl - - 7 21 70 70 70 RNeseAcpg ml) - 1 1 1 1 10 100
l-CA irscdubleta) lO0 0 16 43.6 613 St6 15.6
FIG. 6. Protection of NP-RNA complexes against RNase A digestion. Labeled segment 8 RNA (0.076 fmol) was incubated in the absence (lanes 1 and 2) or presence of 7 (lane 3), 21 (lane 4), and 70 (lanes 5, 6, and 7) ng of NP. After addition of RNase A (lane 1, no adittion; lanes Z-5, 1 &ml; lane 6, 10 Mg/ml; lane 7, 100 @g/ml), the reaction mixture was incubated at 30 “C for 5 min. RNA was extracted and analyzed as described under “Materials and Methods.” The fraction of labeled RNA retained to trichoroacetic acid (Z’CA)-insol- uble fraction is also shown at the bottom. “‘P-Labeled Hue111 frag- ments of pBR322 were used as size markers (lane M).
ing activity seems less to purine than to pyrimidine polynu- cleotides (Fig. 4 and Table II). RNA longer than 15 nucleo- tides is bound by NP (Fig. 5). RNA complexed with NP under the optimum conditions was partially resistant to RNase A (Fig. 6). The reverse-printing method using RNase Vl re- vealed that reconstituted NP-RNA complexes as well as na- tive RNP cores carry the RNase Vl-sensitive sites (Fig. 7, B and C). This indicates that RNA covered with NP contains helical or double-stranded structure. This observation also suggests that the NP-RNA complexes thus formed are struc- turally similar to native RNP cores.
NP is rich in arginine and has a net positive charge of +14 at pH 6.5 (24). There are no clusters of basic residues, and therefore several regions on NP molecule might be involved in the interaction with RNA. Based on the total length of the influenza virus genome and the number of NP molecules associated with one virus particle, it is estimated that approx- imately 20 nucleotides interact with a single NP protein (25). NP can be displaced from RNP cores by polyvinyl sulfate (2, 26) and RNP cores are partially susceptible to RNase diges- tion (2,20-22). All these observations support the notion that RNA is exposed on the surface of the RNP structure (27). Putting these results together, we propose the following model as a possible structure of RNP complexes: RNA polymerase is associated near the 3’ terminus of viral RNA (6) and NP molecules cooperatively bind to viral RNA and constitute the spiral sheets, along which RNA adhere in a wavy form with a pitch of 15 nucleotides. It is not clear whether helical and/or double-stranded RNA structure induced by NP is formed intramolecularly or intermolecularly.
by guest on March 21, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Binding Mode of Influenza Virus Nucleoprotein to RNA 11155
12s . i
106 . 4
m- * _ 80- :
.
M- 9 II- .
r - Sl- ;
123-
104- . -
89- w-
M- . . 57- .
Sl- *-
FIG. 7. Reverse-printing analysis of NP-influenza viral RNA complexes. A, scheme of the reverse-printing analysis is summarized. Details are described under “Materials and Methods.” B, using native RNP cores, the reverse-printing analysis was carried out in the absence (lane I) or presence (lane 2) of RNase Vl. 32P- Labeled Hue111 fragments of pBR322 (&f) and “‘P-labeled primer (P) are used as size markers. C, using NP-viral RNA complexes (lanes 1 and 2), viral RNA (URNA) (lanes 3 and 4), and NP (lanes 5 and 6), reverse-printing analysis was carried out in the absence (lanes 1, 3, and 5) or presence (lanes 2, 4, and 6) of RNase Vl. M and P are the same as in B. Viral RNA used for this analysis is originally degraded to some extent.
A fraction of NP is phosphorylated at 1 serine residue per molecule (2%30), but it is not known yet what percentage of NP molecules is phosphorylated or what role the phosphate group plays. During the preparation of this paper, Kistner et al. (17) reported that during the infectious cycle the phospho- peptide fingerprint of NP underwent significant changes and proposed that a subtle phosphorylation/dephosphorylation control exists during viral multiplication. On this line, it is of interest that the pretreatment of NP with alkaline phospha- tase enhanced its RNA binding activity (Table I). It is possible that the RNA binding activity is subject to change by phos- phorylation and/or dephosphorylation and this control plays a role in the regulation of transcription and replication.
Several lines of genetical evidence indicate that not only RNA polymerase proteins and NP but also one or more proteins encoded by virus are required for replication (31-33). Biochemical studies have revealed that the membrane protein (M) can interact with RNP and controls the transcription from RNP (34-37). By isolating these proteins and reconsti-
tuting ribonucleoprotein complexes with them, we hope to define the regulation of influenza virus transcription and replication mechanisms.
Acknowledgments-We thank Dr. S. Ueda (Nippon Institute for Biological Science) for preparation of influenza virus, Drs. N. Oga- sawara and H. Yoshikawa (Osaka University) for valuable comments, and M. Ogino for typing of this manuscript.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20. 21.
22.
23. 24. 25.
26. 27.
28. 29.
30. 31.
32. 33. 34.
35.
36.
37.
REFERENCES Palese, P. (1979) Cell 10, l-10 Pons, M. W., Schulze, I. T., and Hirst, G. K. (1969) Virology 39,
250-259 Compans, R. W., Content, J., and Deusberg, P. H. (1972) J. Virol.
10,795-800 Krug, R. M. (1983) in Genetics of Influenza Viruses (Palese, P.,
and Kingsbury, D. W., eds) pp. 70-98, Springer-Verlag, Berlin, West Germany
Ishihama, A., and Nagata, K. (1988) CRC Crit. Reu. Biochem. 23, 27-76
Honda, A., Ueda, K., Nagata, K., and Ishihama, A. (1988) J. Biochem. (Tokyo) 102, 1241-1249
Honda, A., Ueda, K., Nagata, K., and Ishihama, A. (1987) J. Biochem. (Tokyo) 104, 1021-1026
Beaton, A. R., and Krug, R. M. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,6282-6286
Kawakami. K.. Ishihama, A., and Hamaguchi, M. (1981) J. Biochem: (Tokyo) 89, 1?51-i757 -
Takeuchi, K., Nagata, K., and Ishihama, A. (1987) J. Biochem. (Tokyo) 101,837-845
Melton. D. A.. Kriee. P. A.. Rebaeliati. M. R.. Maniatis. T.. Zinn. K., and Green, MR. (1984) N&e& Acids bes. 12, ?035-7056
Nagata, K., Guggenheimer, R. A., and Hurwitz, J. (1983) Proc. Natl. Acad. Sci. U. S. A. 80, 6177-6181
Carthew, R. W., Chodosh, L. A., and Sharp, P. A. (1985) Cell 43, 439-448
Matsumoto, K., Nagata, K., Hanaoka, F., and Ui, M. (1989) J. Biochem. (Tokyo) 105,927-932
Winter, G., Fields, S., Gait, M. J., and Brownlee, G. G. (1981) Nucleic Acids Res. 9, 237-245
Kato, A., Ishihama, A., Noda, A., and Ueda, S. (1984) J. Virol. Methods 9,325-339
Kistner, O., Muller, K., and Scholtissed, C. (1989) J. Gen. Virol. 70,2421-2431
Blumberg, B. M., Giorgi, C., and Kolakofsky, D. (1983) Cell 32, 559-567
Kingsbury, D. W., James, I. W., and Murti, K. G. (1987) Virology 156,396-403
Duesberg, P. (1969) J. Mol. Biol. 42,485-499 Kingsbury, D. W., and Webster, R. G. (1969) J. Virol. 4, 219-
225 Murti, K. G., Bean, W. J., Jr., and Webster, R. G. (1980) Virology
104,224-229 Grant. R. A.. and Olesen, S. 0. (1982) Biochemistry 21, 4823- Winter, G., and Fields, S: (1981) Virology 114, 423-428 Comnans. R. W.. Content. J., and Deusberg, P. H. (1972) J. Virol.
l&795-800 Goldstein, E. A., and Pons, M. W. (1970) Virology 41, 382-384 Lamb, R. A. (1983) in Genetics of Influenza Viruses (Palese, P.,
and Kingsbury, D. W., eds) pp. 21-69, Springer-Verlag, Berlin, West Germany
Kamata, T., and Watanabe, Y. (1977) Nature 267,460-462 Privalsky, M. L., and Penhoet, E. E. (1981) J. Biol. Chem. 256,
5368-5376 Petri, T., and Dimmock, N. J. (1981) J. Gen. Virol. 57, 185-190 Sugiura, A., Ueda, M., Tobita, K., and Enomoto, C. (1975) Virol-
ogy 65,363-373 Scholtissek, C., and Bowles, A. L. (1975) Virology 67,576-587 Thierry, F., and Spring, S. B. (1981) Virology 115, 137-148 Zvonarjev, A. Y., and Ghendon, Y. Z. (1980) J. Virol. 33, 583-
586 Rees, P. J., and Dimmock, N. J. (1981) J. Gen. Virol. 53, 125-
132 Te, Z., Pal, R., Fox, J. W., and Wagner, R. R. (1987) J. Virol.
61,239-246 Hankins, R. W., Nagata, K., Bucher, D. J., Popple, S., and
Ishihama, A. (1989) Virus Genes 3, 111-126
by guest on March 21, 2019
http://ww
w.jbc.org/
Dow
nloaded from

K Yamanaka, A Ishihama and K Nagataresembling native viral ribonucleoprotein cores.
Reconstitution of influenza virus RNA-nucleoprotein complexes structurally
1990, 265:11151-11155.J. Biol. Chem.
http://www.jbc.org/content/265/19/11151Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/265/19/11151.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 21, 2019
http://ww
w.jbc.org/
Dow
nloaded from