recruitment of crabs clawto promote nectary development
TRANSCRIPT

5021
IntroductionA primary issue in biology concerns the origin ofmorphological diversity over evolutionary time. Theconvergence of evolutionary biology and developmentalgenetics provides a step toward understanding this complexissue. Approximately 90% of land plants are flowering plantspecies; despite their enormous diversity in morphology,flowers are recognizable owing to the regularity in theirorganization with whorls of floral organs in a stereotypicalarrangement. Recent molecular genetic studies suggestunderlying genetic programs directing flower development areconserved throughout angiosperms, consistent with a singleorigin of flowers (Ambrose et al., 2000; Ferrario et al., 2004;Kyozuka et al., 2000). By contrast, it is often difficult toidentify a common ontology in structures highly modifiedwithin lineages over evolutionary time. Increasing geneticinformation about the development of such structures canprovide clues to their origin.
With the rapid progress in developmental genetics andconfidence in the establishment of the flowering plantphylogeny, a gene(s) involved in particular developmentalprocesses in model species can be evaluated in anevolutionarily context by examining orthologs in divergentangiosperm species. Through comparative expression andfunctional studies, it is possible to gain insight into therelationship between the evolution of genes and morphology.
The nectary provides an interesting example with which toaddress this question owing to its diversity in both structureand ontogeny. Nectaries are highly variable in theirmorphologies, anatomies and locations, and are defined basedon their shared function: the secretion of nectar (Fahn, 1979).Depending on location, nectaries either serve to attractpollinators or protect against herbivores. Although nectariesreportedly occur in ferns (Darwin, 1877) and Gnetales (Porsch,1910), they are most widespread in angiosperms,predominantly developing or occurring within flowers, whencompared with other parts of the angiosperm plant body. Manyangiosperm flowers are animal pollinated (Eriksson andBremer, 1992), with pollinators attracted to flowers to gathernectar or pollen as food sources. Fossil records of angiospermsand insects suggest that the timing of the radiations ofangiosperms and certain insect classes were coincident (Crepetand Friis, 1987; Meeuse, 1978; Pellmyr, 1992). Therefore,along with the other floral organs, the innovation of nectariesmay have played a major role in angiosperm and metazoanevolution.
Although locations of nectaries within flowers are constantat the family level, in broader taxonomic terms, their locationsare highly variable (Brown, 1938). In basal angiosperms,nectaries tend to be associated with the perianth (the non-reproductive floral organs) (Endress, 2001), while in theeudicots, nectaries are usually associated with carpels and
Nectaries are secretory organs that are widely present inflowering plants that function to attract floral pollinators.Owing to diversity in nectary positions and structures, theyare thought to have originated multiple times duringangiosperm evolution, with their potential contribution tothe diversification of flowering plants and pollinatinganimals being considerable. We investigated the geneticbasis of diverse nectary forms in eudicot angiospermspecies using CRABS CLAW (CRC), a gene required fornectaries in Arabidopsis. CRC expression is conserved inmorphologically different nectaries from several coreeudicot species and is required for nectary development inboth rosids and asterids, two major phylogenetic lineagesof eudicots. However, in a basal eudicot species, no evidence
of CRC expression in nectaries was found. Considering thephylogenetic distribution of nectary positions and CRCexpression analyses in eudicots, we propose that diversenectaries in core eudicots share conserved CRC generegulation, and that derived nectary positions in eudicotshave altered regulation of CRC. As the ancestral functionof CRC lies in the regulation of carpel development, it mayhave been co-opted as a regulator of nectary developmentwithin the eudicots, concomitant with the association ofnectaries with reproductive organs in derived lineages.
Key words: Nectary, Carpel, CRABS CLAW, YABBY, Eudicot,Arabidopsis
Summary
Recruitment of CRABS CLAW to promote nectary developmentwithin the eudicot cladeJi-Young Lee1, Stuart F. Baum1, Sang-Hun Oh2, Cai-Zhong Jiang3, Jen-Chih Chen3 and John L. Bowman1,*1Section of Plant Biology, University of California Davis, Davis, CA 95616, USA2Department of Biology, Duke University, Durham, NC 27708, USA3Department of Plant Sciences, University of California Davis, Davis, CA 95616, USA*Author for correspondence (e-mail: [email protected])
Accepted 26 August 2005
Development 132, 5021-5032Published by The Company of Biologists 2005doi:10.1242/dev.02067
Research article
Dev
elop
men
t

5022
stamens. Thus, Fahn (Fahn, 1953) argued that there is a trendin nectary position within flowers, shifting from peripheralperianth positions in basal taxa to central positions associatedwith reproductive organs in more derived taxa. In addition,extrafloral nectaries are currently known in 68 angiospermfamilies (Elias, 1983). Their structures and locations are alsodiverse across the families, although they occur most often ‘onthe upper half of the petiole at or near the base of the leaf bladethan any other site’ (Elias, 1983). The diversity of nectaryforms and distributions within flowering plants suggest thatthey may have multiple independent origins. However, thisdoes not preclude diverse nectaries from sharingdevelopmental genetic machinery.
In Arabidopsis, crabs claw (crc) is the only known singlemutant that lacks floral nectaries (Baum et al., 2001; Bowmanand Smyth, 1999). CRC is expressed in nectaries, withexpression commencing before the emergence of nectaryglands and continuing until after anthesis. Although ectopicexpression of CRC alone does not result in ectopic nectaries,ectopic expression of CRC in conjunction with other genes,such as UFO, or in specific mutant backgrounds, results in thedevelopment of ectopic nectaries at the bases of flower pedicels(Baum et al., 2001). Thus, CRC is required for nectarydevelopment and ectopic CRC expression in some contexts issufficient for nectary formation (Baum et al., 2001). CRCactivation in nectaries is mediated by a combination of florallyexpressed MADS box proteins, although the tissue-specificfactors limiting expression to nectaries and carpels areunknown (Lee et al., 2005).
CRC encodes a putative transcription factor of the YABBYgene family (Bowman and Smyth, 1999). Several members ofthe YABBY gene family are expressed abaxially in developingleaf primordia and floral organs with their ectopic adaxialexpression transforming the adaxial leaf surface into one withabaxial characteristics, implicating members of this genefamily in establishing or interpreting leaf polarity (Sawa et al.,1999; Siegfried et al., 1999). Although crc single mutants donot affect carpel polarity, when combined with other mutations,such as kanadi, adaxial tissues develop in abaxial positions,indicating that CRC is required for proper carpel polarity(Eshed et al., 1999). Consistent with CRC promoting abaxialdifferentiation, CRC is expressed abaxially in the carpels(Bowman and Smyth, 1999). In addition, genetic studies withfloral homeotic ABC mutants in Arabidopsis showed that CRCspecifies carpel identity in parallel with AGAMOUS (Alvarezand Smyth, 1999). Studies of CRC orthologs in Oryza andAmborella, which together with Arabidopsis span thephylogenetic diversity of angiosperms, suggest that a role incarpel development is likely to be the ancestral function ofCRC in angiosperms (Fourquin et al., 2005; Yamaguchi et al.,2004). In this study, we examined whether CRC has anancestral function in the nectaries of angiosperms or whetherCRC was recruited for a role in nectary development within theangiosperm lineage.
Materials and methodsTaxon samplingTaxa were sampled to represent diverse nectary forms inrepresentative eudicot lineages. For the analysis of CRC expressionpatterns from close relatives of Arabidopsis, Lepidium africanum and
Cleome sparsifolia [Cleomaceae based on work by Hall et al. (Hall etal., 2002)] were chosen because the two species show variation in theorganization of floral nectaries. For the analysis in extrafloralnectaries, Capparis flexuosa in Capparaceae [sister toBrassicaceae/Cleomaceae (Hall et al., 2002)] and Gossypiumhirsutum in Malvaceae were used. As representatives of asterids(APG, 1998), CRC homologues from Nicotiana tabacum and Petuniahybrida (both Solanaceae) were isolated, and the expression of CRCin P. hybrida was examined. For the analysis in flowers of basaleudicots (Soltis et al., 2000), Aquilegia formosa (Ranunculaceae) waschosen (Fig. 1).
Cloning of CRABS CLAW orthologsTotal RNA was isolated from nectary or carpel tissue using theRNeasy Plant Mini kit (Qiagen, Hilden, Germany). First strand cDNAwas synthesized using SuperScript II reverse transcriptase (Invitrogen,Carlsbad, USA). Partial fragments of CRC cDNAs were amplified byusing degenerate primers dCRC-ZnF (5�-CDGTRACRGTGAAAT-GYGGYCATTGYRGYA-3�) and dCRC-YB (5�-AIGCGATGGRA-GYCTSTGYTTCTTCTCRGG-3�) (see Fig. S1 in the supplementarymaterial). Polymerase chain reactions (PCR) were conducted withTakara ExTaq polymerase (Madison, USA) on a RoboCycler(Stratagene, La Jolla, USA) with the following protocol: 2 minutes at94°C, 35 cycles of 30 seconds at 94°C, 30 seconds at 42-60°C and 30seconds at 72°C, followed by one cycle of 2 minutes at 72°C. PCRproducts were separated on 0.7% TAE agarose gels, gel isolated(Quiagen, Hilden, Germany) and TOPO TA cloned (Invitrogen,Carlsbad, USA). Purified DNA was sequenced using an ABI PRISM377 DNA sequencer. Following confirmation of CRC orthologsequences, full-length cDNA sequences were obtained by 5� and 3�RACE using a SMART RACE cDNA amplification kit (Clontech,Palo Alto, USA).
Phylogenetic analysesTo determine whether genes from various angiosperm groupsrepresent orthologs, phylogenetic analyses were conducted employingthe Bayesian method. We included CRC genes determined in thisstudy, as well as sequences of YABBY gene family members fromGenBank (the gene list is provided in Table S1 in supplementarymaterial). Deduced amino acid sequences of the YABBY genes werealigned using ClustalX (Thompson et al., 1997) and manuallyadjusted using the program Se-Al (A. Rambaut, seehttp://evolve.zoo.ox.ac.uk/). The alignment confirmed the twodomains recognized by Bowman and Smyth (Bowman and Smyth,1999) (zinc finger and YABBY domains) are highly conserved acrossgenes from angiosperms and gymnosperms (see amino acid alignmentin Fig. S1 in the supplementary material). However, it is difficult toassume positional homology in the remaining regions of the genesowing to a higher level of variability. Thus, we analyzed onlynucleotide sequences of the two conserved domains. The aligned datamatrix was submitted to the TreeBase database(http://www.treebase.org) and is available upon request.
Bayesian phylogenetic analyses were performed with MrBayes3.0 (Huelsenbeck and Ronquist, 2001) using the sequences fromgymnosperms as outgroups. A Metropolis-coupled Markov chainMonte Carlo algorithm was employed for 2 million generations,sampling trees every 100 generations, with four independentchains running simultaneously. The general time-reversal model(Swofford et al., 1996) with six rate parameters and the gammadistribution, as determined by the hierarchical likelihood ratiotest using Modeltest (Posada and Crandall, 1998), was used toestimate the likelihood values. All 20,001 resulting trees wereimported into PAUP* 4.0b10 (Swofford, 2002), and a 50%majority-rule consensus tree was generated after discarding the first1001 trees (100,000 generations). These ‘burn-in’ generations, forwhich the log-likelihood values had not reached a plateau, weredetermined by plotting a graph of the log-likelihoods of each
Development 132 (22) Research article
Dev
elop
men
t

5023CRABS CLAW recruitment
generation versus generation numbers (Huelsenbeck and Ronquist,2001).
MicroscopyFor Scanning Electron Microscopy (SEM), tissue was fixed overnightwith 3% glutaraldehyde, phosphate buffered to pH 7, followed by theovernight fixation in 0.5% osmium tetraoxide. Tissue was dehydratedand critical-point dried. After sputter coating with gold/palladium,tissue was observed on a Hitachi S-3500N scanning electronmicroscope. Anatomical analysis was carried out according to themethod of Baum and Rost (Baum and Rost, 1996). Tissue sectionswere stained using the PAS reaction.
In situ hybridizations and semi-quantitative RT-PCRIn situ RT-PCR was performed following Xoconostle-Cázares et al.(Xoconostle-Cázares et al., 1999) on thin hand-sectioned tissues fromleaves and involucres of Gossypium hirsutum. Gene-specific primersGhCRC-1 (GGTTCCACAATCCGAGCATCTTT) and GhCRC-2(CACAAACGGATGATGCTGCTGGAGAA) were added to a RT-PCR mixture containing Oregon Green-labeled dUTP (MolecularProbes, Eugene, USA). Slides with fresh tissue and PCR mixture weresealed and the PCR was run at 60°C for 20 minutes followed by 10cycles of 30 seconds at 94°C, 30 seconds at 60°C and 1 minute at72°C. As a negative control, RT-PCR was run in parallel without gene-specific primers. After washing out free dNTPs, fluorescent signalswere observed using confocal laser scanning microscopy (CLSM).
For semi-quantitative RT-PCR, the above gene-specific primerswere used along with actin primers as a quantitative control:Ghactin-F (CCTCTTCCAGCCATCTTTCATC) and Ghactin-R(ATTCATCATACTCACCCTTGGA). Total RNA was extracted fromvarious regions of plant tissue using a Qiagen RNeasy Plant Mini kit.Total RNA (2.5 �g) was used for cDNA synthesis and PCR wasperformed using Takara ExTaq polymerase. PCR was run at 94°C for1 minute followed by 30 or 35 cycles of 30 seconds at 94°C, 30seconds at 58°C, and 1 minute at 72°C, and an extension of 5 minutesat 72°C.
Standard RNA in situ hybridization was as described by VielleCalzada et al. (Vielle Calzada et al., 1999), except tissue in paraffinwas sectioned at 10-15 �m and hybridization was carried out at 55°Cfor 36 hours.
Virus induced gene silencing (VIGS)A previously described Tobacco Rattle Virus (TRV)-based VIGSsystem was used to silence CRC in Petunia and Nicotiana (Liu et al.,2002; Chen et al., 2004). A 575 bp fragment of the petunia CRC genecorresponding to entire coding region was cloned into pTRV2 to formpTRV2 CRC. The constructs pTRV1 and pTRV2 or pTRV2 (see Liuet al., 2002) CRC were transformed into Agrobacterium strainGV3101 by electroporation. Virus infection was achieved byAgrobacterium-mediated infection of Petunia hybrida or Nicotianabethamiana as described by Chen et al. (Chen et al., 2004). Flowerswere examined for aberrant phenotypes 2-6 weeks post infection.Selection of Petunia flowers for examination was facilitated by thesimultaneous VIGS mediated silencing of chalcone synthase (Chen etal., 2004).
ResultsPhylogenetic distribution of angiosperm nectariesMapping of nectary positions onto the currently acceptedphylogeny of flowering plants (Qiu et al., 1999; Soltis et al.,1999; Soltis et al., 2000) indicates that structural diversificationof nectaries has occurred in the core eudicots, with nectariesprimarily associated with reproductive organs in these species(Fig. 1). Brown (Brown, 1938) provided a thorough descriptionon the distribution of floral nectary positions in angiosperms
based on observations in more than 3000 species. Focusing ontheir diverse nature, he stated, ‘Nectaries appear to have arisenindependently in different lines of development and then tohave undergone modifications characteristic of variousgroups’. However, mapping nectary positions on the currentlyaccepted angiosperm phylogeny shows a pattern that supportsFahn’s (Fahn, 1953) argument of an evolutionaryacrocentripetal movement of nectary position withinangiosperms (Fig. 1). In general, nectaries are not welldifferentiated in basal angiosperms. Nymphaeaceae andIlliciaceae in the ANITA grade have nectaries on perianth parts,and Amborella generates liquid exudates to the carpel tipwhose identity as nectar has not been characterized (Endress,2001). A similar situation is found in most of the eumagnoliidlineage, except in the monocot clade. Floral nectaries inmonocots are defined as septal glands, the septa being placeswhere adjacent walls of two carpels have not fused (Brown,1938). Almost all monocots possessing nectaries have septalglands. In eudicots, nectaries are more conspicuous, elaborateand usually associated with reproductive floral organs, withperianth nectaries being relatively rare (e.g. Gossypium sp.). Atleast 68 families are reported to have species with extrafloralnectaries (Elias, 1983) and the mapping of those species withinthe current angiosperm phylogeny indicates that extrafloralnectaries are mostly distributed in core eudicot species andmonocots, and have independently evolved in several lineages(Fig. 1).
Single origin of CRC in angiospermsBayesian phylogenetic analysis based on nucleotide sequencesof two domains of several members of the YABBY family froma broad spectrum of angiosperm species and fourgymnosperms resolved five major gene lineages, representedby INO, CRC, YABBY2, FIL/YABBY3 and YABBY5 (Fig. 2).INO, CRC and YABBY5 genes formed strongly supportedclades with the posterior probabilities (i.e. probability of thehypothesis given the data) of 100. YABBY2 and FIL/YABBY3genes were, however, weakly supported as monophyleticlineages. Each clade contains members from basalangiosperms or monocots, suggesting the ancestralangiosperms had five YABBY family genes, consistent withthe results of Yamada et al. (Yamada et al., 2004). Our genetrees weakly suggested that INO and CRC genes, both of whichare involved in developmental processes in flowers, are derivedfrom other YABBY genes that are expressed in all lateralorgans (Siegfried et al., 1999). Further sampling ingymnosperms is required to ascertain the antiquity of thedifferent clades of YABBY gene family members.
Our phylogenetic analysis of YABBY genes indicated thatCRC genes originated once during the evolution of the YABBYgenes. Thus, CRC genes in various diploid angiosperms areorthologs (Hillis, 1994) that have diversified via speciation notby gene duplication. The topology within the CRC gene cladeis consistent with Soltis et al. (Soltis et al., 2000), even thoughour sampling of the genes is limited. The CRC genes frommonocots and Brassicales both formed strongly supportedclades, as did two cotton genes.
CRC expression in floral nectaries is well conservedin Brassicaceae/CleomaceaeIn Brassicaceae/Cleomaceae species (Hall et al., 2002),
Dev
elop
men
t

5024
nectaries develop at the bases of stamen filaments (Fahn,1979). Norris (Norris, 1941) classified Brassicaceae nectariesinto three types: annular, four-nectary and two-nectary types.Arabidopsis thaliana is a four-nectary type with two medialand two lateral nectaries (Baum et al., 2001); Lepidiumafricanum is a two-nectary type with only two medialnectaries (Fig. 3A); and Cleome sparsifolia has annular typenectaries in which glands develop continuously around thecircumference of the androecium (Fig. 3F). Although thesizes of nectary glands vary among these species, theirexternal organizations are similar. Stomata develop at theabaxial tips of nectary glands (Fig. 3B), and epidermal cellsof nectaries have distinctive reticulate cuticular thickenings(Baum et al., 2001). In these species, CRC expressioncommences at or before initiation of nectary development,increases during growth of the nectary, and decreases afteranthesis (Fig. 3C,D,G). In carpels, CRC expressioncommences early, and is associated with abaxial regions ofthe carpel walls (data not shown), similar to Arabidopsis(Bowman and Smyth, 1999).
Expression of CRC in extrafloral nectaries of rosidsExtrafloral nectaries develop in leaf axils in Capparis flexuosa(Zimmerman, 1932). Nectaries are covered with sheaths andconsist of cytoplasmically dense cells innervated with phloemextending from the stem (Fig. 3H-L). Stomata present in theepidermis of the nectary might be involved in nectar secretion(Fig. 3K). As with floral nectaries, CRC expression correlateswith the proliferation of nectariferous cells underneath thesheaths with CfCRC RNA levels increasing in developingnectaries and decreasing in mature nectaries (Fig. 3M-P).
To determine whether CRC expression correlates withextrafloral nectary development in phylogenetically distantspecies, GhCRC expression in extrafloral nectaries ofGossypium hirsutum (cotton) was analyzed. In G. hirsutum,extrafloral nectaries develop from the midvein on the abaxialsides of leaves and cotyledons, and on involucre bracts, leaf-like organs that subtend flowers. Nectaries consist ofmulticellular secretory structures that proliferate in a concaveindentation on the abaxial midvein, positioned approximatelyone-third of the distance to the distal tip of the leaf (Fig. 4A-
Development 132 (22) Research article
Fig. 1. Phylogenetic distribution of nectaries in angiosperms. Non-eudicot angiosperms have floral nectaries in the perianth, or nectary tissuesare not conspicuously developed. In eudicots, floral nectaries are mostly associated with reproductive organs, although some examples ofperianth nectaries exist. The angiosperm phylogeny is based on that of Soltis et al. (Soltis et al., 2000). Orders with extrafloral nectaries aremarked bold with an asterisk. Extrafloral nectaries are found only within eudicots and monocots. The extrafloral nectary distribution is based onthat of Elias (Elias, 1983) and locations of floral nectaries are primarily based on those of Brown (Brown, 1938) and others (Endress, 1986;Endress, 2001; Proctor et al., 1996; Ronse DeCraene, 2004; von Balthazar and Endress, 2002). Groups reported as not having nectaries areindicated (–). Austrobaileyaceae and Amborellaceae were marked with a question mark, as the possible nectar secretion on the tip of carpelshas not been characterized. Taxa used in this study and the positions of nectaries in these taxa are indicated. ax, leaf axil (Capparis flexuosa); le,abaxial midvein of leaf (Gossypium hirsutum); se, sepal (Gossypium hirsutum); pe, petal (Aquilegia formosa); st, stamen (Arabidopsis thaliana,Lepidium africanum, Cleome sparsifolia); ca, carpel (Petunia hybrida, Nicotiana benthamiana, Grevillea robusta).
Dev
elop
men
t

5025CRABS CLAW recruitment
D). Owing to high concentrations ofsecondary metabolites, RNA in situhybridization was difficult in this speciesand our experimental results wereequivocal. Thus, GhCRC expression wasanalyzed by RT-PCR and in situ RT-PCR,which detected GhCRC expression in thenectariferous cells of the leaf and bract(Fig. 4E,F). To evaluate the specificityand levels of GhCRC expression duringnectary development, semi-quantitativeRT PCR was performed using actin as aquantitative control (Holland et al.,2000). GhCRC is not only expressed indeveloping nectaries but also in otherregions of the leaves, albeit at lowerlevels. GhCRC expression commences atan early stage of leaf development beforeany visible signs of secretory cells and ismore abundant in the region ofdeveloping extrafloral nectaries. Thisexpression pattern is also observed inextrafloral nectaries on involucre bracts.At a stage when nectaries aremorphologically visible, levels ofGhCRC mRNA are clearly higher than inother regions within the bract (Fig. 4G).Although not evident at the cellular level,RNA analysis suggests that GhCRC iscorrelated with extrafloral nectarydevelopment in G. hirsutum.
Gossypium tomentosum, a nativeHawaiian species closely related to G.hirsutum, does not develop extrafloralnectaries, a derived condition within thegenus (Small et al., 1998). Previousgenetic analysis of the nectarilessphenotype suggested that two loci areinvolved, with the G. tomentosum allelesbeing recessive (Meyer and Meyer,1961). As these species are AD genomeallotetraploids, and the two nectarilessloci mapped to syntenous regions of thetwo genomes in G. tomentosum, changesin a single gene may be responsible forthe nectariless phenotype. To determinewhether mutations in CRC could beresponsible for the nectariless phenotype, two experimentswere conducted. First, we analyzed F2 plants derived from anF1 hybrid between G. hirsutum and G. tomentosum, andexamined the segregation of CRC orthologs in nectarilessindividuals by RFLP analyses (data not shown). That thenectariless individuals did not predominantly have the G.tomentosum CRC allele indicates that alleles at this locus arenot involved in the nectariless phenotype. In addition, mappingstudies using CRC RFLP markers positioned the CRCorthologs on chromosomes 6 and 14, which does notcorrespond to the map positions of the nectariless phenotype[chromosomes 5 and 9 according to Meyer and Meyer (Meyerand Meyer, 1961)]. In G. tomentosum, CRC expression isreduced in seedlings when compared with G. hirsutum. In the
apices of seedlings at the stage when cotyledons haveexpanded, CRC is expressed at high level in G. hirsutum,whereas there is no expression detected in G. tomentosum(compare lane 29 with lane 32 in Fig. 4G).
CRC: a general regulator of nectary development inthe core eudicotsThe correlation of CRC expression with developing extrafloralnectaries of G. hirsutum and C. flexuosa raises the question ofthe genetic independence of ontogenetically unrelatednectaries. As CRC expression is observed in structurallydifferent nectaries in closely related species, we examined CRCin nectaries in a broader range of eudicot species. In Petuniahybrida, a core asterid species, nectaries develop as a disc at
Fig. 2. Phylogenetic analysis of YABBY genes. Bayesian phylogenetic tree based on thenucleotide sequences of the zinc finger and YABBY domains of the YABBY family genemembers. Posterior probabilities for clades are shown above branches.
Dev
elop
men
t

5026
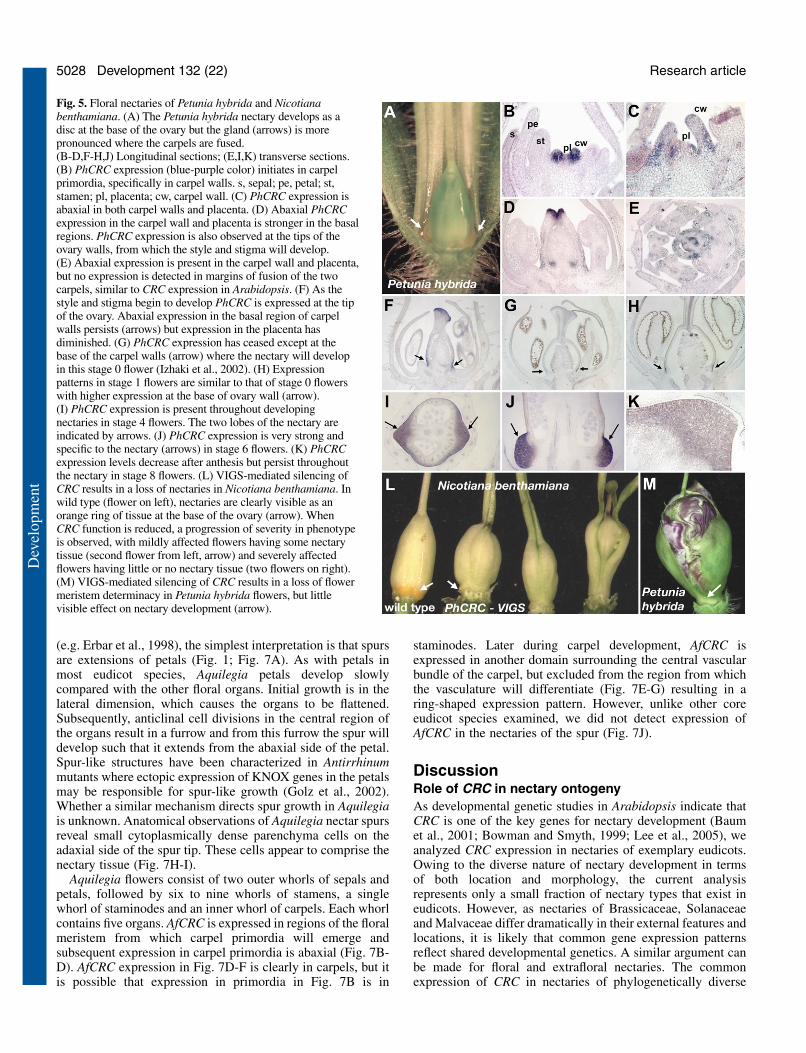
the base of the ovary (Fig. 5A), which is typical for asteridspecies, and PhCRC expression is associated with thedevelopment of carpels and nectaries. PhCRC is expressed incarpel primordia at initiation, abaxially in ovary walls, andsubsequently abaxially in placental tissue developing interiorto the ovary walls (Fig. 5B-E). In addition, CRC is expressedat a high level in the developing style and stigma (Fig. 5D,F).As carpel development progresses, abaxial expression of CRCabates and remains only in the basal regions of the ovary wallsfrom where nectaries arise (Fig. 5F-H). In a stage 2 flower(Izhaki et al., 2002), nectary development is conspicuous asoutgrowths on opposite sides of the septa and PhCRCexpression is specific to the growing nectary tissue (Fig. 5I).Ovary walls adjacent to the nectary do not show detectablePhCRC expression (Fig. 5J). By stage 4, nectary tissuesurrounds the base of the ovary walls (Fig. 5I,J) and PhCRCexpression in nectaries continues through anthesis (Fig. 5K).
A virus-induced gene-silencing (VIGS) system based ontobacco rattle virus (TRV) was employed to determine thefunction of CRC in Petunia hybrida and Nicotianabenthamiana flowers (Chen et al., 2004). Nectaries in most
Solanaceae species are easily distinguished by their orangecolor owing to carotenoid accumulation (Fig. 5A,L). VIGSusing the PhCRC gene resulted in a loss of nectaries, loss ofcarpel fusion and reduced determinacy in N. benthamianaflowers (Fig. 5L), all characteristics of crc mutations inArabidopsis (Alvarez and Smyth, 1999; Bowman and Smyth,1999). In all infected plants, ~80% of N. benthamiana flowersexhibited a severe phenotype, as depicted in Fig. 5. In Petunia,concomitant downregulation of chalcone synthase allows oneto select flowers exhibiting VIGS. The most severe phenotypeobserved was a loss of floral meristem determinacy, with thecells normally giving rise to the placenta instead developinginto another flower (Fig. 5M). However, in Petunia hybrida,downregulation of PhCRC via VIGS was less severe, with aconspicuous phenotype in only two plants out of eightexamined, and less than 50% of white flowers exhibitingindeterminacy. We interpret these results as a partial loss ofCRC function induced by VIGS in Petunia and a more severeloss of CRC function in N. benthamiana, perhaps owing to N.benthamiana being more susceptible to virus infection (Yanget al., 2004).
Development 132 (22) Research article
Fig. 3. Nectaries of the Brassicales.Nectaries and expression of CRC orthologsin Lepidium africanum (A-D), Cleomesparsifolia (E-G) and Capparis flexuosa (H-P). (A) Floral nectaries (n) of Lepidiumafricanum are located between stamens andpetals (p). Nectary glands flank each of thetwo stamens, such that each flower has fourglands. (B) Nectary glands have apicalstomata (arrow). (C,D) LaCRC is expressedin all cells of the nectary glands (arrow in C),from inception beyond anthesis.(E,F) Cleome sparsifolia floral nectaries(arrow) form a ring at the abaxial bases ofthe stamens. (G) CsCRC is expressed at highlevels throughout nectaries. (H,I) Extrafloralnectaries of Capparis flexuosa (arrows) arelocated in leaf axils and are covered withscales, dissected away in I. (J) Longitudinalsection of an extrafloral nectary reveals theinternal anatomy (arrow). An axillary bud isadjacent and below the extrafloral nectary.(K) The covering sheath is composed mostlyof thickened collenchyma cells and fibers,and the nectary has stomatal openings on itssurface (arrows). (L) Nectaries comprisemostly parenchyma cells with phloem tracescontaining sieve tube members andcompanion cells. (M-P) Before nectarytissue develops inside the sheath, CfCRCexpression is not observed (M, the dark spotis not signal). As nectary tissue initiatesdevelopment, CfCRC expression increasesdramatically and levels remain highthroughout the nectary (N,O). As nectarygrowth ceases CfCRC expression decreasesand only a minimal expression level remains(P).
Dev
elop
men
t

5027CRABS CLAW recruitment
Conservation and changes in regulating CRC:mechanisms to retain CRC as a nectary regulator?CRC expression in extrafloral nectaries of Gossypium hirsutumimplies alterations in spatial and temporal control of CRC generegulation relative to that of Arabidopsis. Thus, we examinedwhether changes have occurred in the GhCRC promoter itselfby examining its activity in Arabidopsis. The GUS reportergene was transcriptionally fused with 8 kb of sequence 5� tothe coding region of GhCRC and transformed into Arabidopsis.Whereas the Arabidopsis CRC promoter drives expression innectaries and carpels, the Gossypium CRC promoter drivesGUS expression in a pattern reminiscent of CRC expression inGossypium (Fig. 6A-E). Expression initiates in seedlings,primarily in the vasculature, with maximum expressionproximally in the midvein where nectaries normally develop incotton (Fig. 6A-C). GUS expression was never found in floral
nectaries of Arabidopsis, though floral expression is detectedin the pedicel and receptacle, as well as the style and stigmaof the gynoecium (Fig. 6D,E). In contrast to the dramaticdifferences in the GhCRC promoter, a GhCRC cDNA is fullyfunctional in Arabidopsis, rescuing the phenotypic defects ofcrc-1 mutants when expressed under control of the ArabidopsisCRC promoter (Fig. 6G). Consistent with the change inexpression domain, pGhCRC 8kb::GhCRC never rescuednectary development in crc-1 mutants, although it frequentlyrescued carpel fusion defects (data not shown), suggesting thatpromoter elements directing carpel expression might bepartially functionally conserved. As a positive control, theheterologous Lepidium promoter (pLaCRC, 4.3 kb)transcriptionally fused with GUS results in an expressionpattern indistinguishable from Arabidopsis CRC and pLaCRCfused to the LaCRC-coding region can fully rescue crc-1mutants (Fig. 6F).
The dramatic alterations in CRC regulation are not due tothe greater evolutionary distance between Arabidopsis andGossypium than between Arabidopsis and Lepidium, as apAtCRC::GUS transgene in Nicotiana tabacum (an asterid,whereas the other three species are rosids) drives GUSexpression in carpels, primarily in style and stigma tissues, andlater at the base of the ovary where the nectary develops (Fig.6H). This pattern is reminiscent of CRC expression pattern inPetunia (Fig. 5), another member of the Solanaceae, suggestingconservation of CRC regulation in distantly related eudicotspecies. Thus, the lineage-specific changes in regulatoryregions of cotton CRC probably contributed to the alterationsin nectary position.
CRC expression in Aquilegia flowers(Ranunculaceae): a basal eudicotNectaries in Aquilegia flowers develop at the tips of petal spurs,the length of which is pollinator specific. Although there hasbeen some debate on the identity of petal spurs of Aquilegia
Fig. 4. The extrafloral nectaries of Gossypium. (A) Extrafloralnectaries are located on midveins on the abaxial sides of the leaves.(B) The secretory structure is composed of trichomes that denselypopulate the concave region of the midvein. (C,D) Secretorytrichomes are multicellular and small cytoplasmically denseparenchyma cells are located below the trichomes. (E) Expression ofGhCRC in an involucral bract nectary localized by in situ PCR with agreen signal representing the fluorescence signal from in situ PCRproducts of GhCRC. (F) GhCRC is highly expressed in the papillarcells and subtending cells in a leaf nectary. (G) Expression ofGhCRC during extrafloral nectary development as monitored bysemi-quantitative RT-PCR. 1-29, G. hirsutum; 30-32, G. tomentosum;left lane, actin; right lane, GhCRC (or GtCRC) amplified from samecDNA for actin under the same PCR condition. The upper bands inlanes 27-29 are from genomic DNA; the lower bands representcDNA. 1, 3 mm whole leaf; 2, 5 mm whole leaf; 3, 1 cm whole leaf;4, 1.4 cm leaf blade; 5, 1.4 cm leaf vein; 6, 2.8 cm leaf blade; 7, 2.8cm leaf vein; 8, 2.8 cm leaf nectary; 9, 7 cm leaf blade; 10, 7 cm leafvein; 11, 7 cm leaf nectary; 12, 12 cm leaf blade; 13, 12 cm leaf vein;14, 12 cm leaf nectary; 15, bract nectary of 1.5 cm flower; 16, rest ofbracts; 17, sepals; 18, petals; 19, stamens; 20, carpels; 21, bractnectary of 6 cm long flower; 22, rest of bracts; 23, sepals; 24, petals;25, stamens; 26, carpels; 27, root of seedlings; 28, hypocotyl ofseedlings; 29, apex of seedlings; 30, root of seedlings; 31, hypocotylof seedlings; 32, apex of seedlings.
Dev
elop
men
t
5028
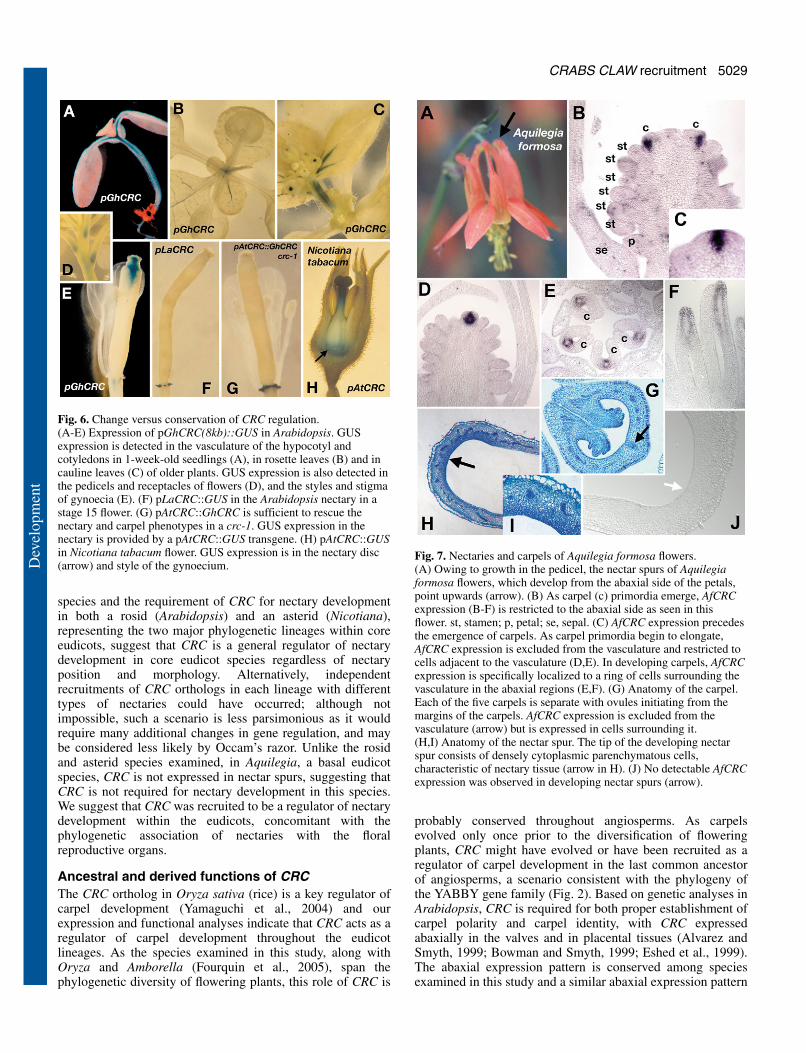
(e.g. Erbar et al., 1998), the simplest interpretation is that spursare extensions of petals (Fig. 1; Fig. 7A). As with petals inmost eudicot species, Aquilegia petals develop slowlycompared with the other floral organs. Initial growth is in thelateral dimension, which causes the organs to be flattened.Subsequently, anticlinal cell divisions in the central region ofthe organs result in a furrow and from this furrow the spur willdevelop such that it extends from the abaxial side of the petal.Spur-like structures have been characterized in Antirrhinummutants where ectopic expression of KNOX genes in the petalsmay be responsible for spur-like growth (Golz et al., 2002).Whether a similar mechanism directs spur growth in Aquilegiais unknown. Anatomical observations of Aquilegia nectar spursreveal small cytoplasmically dense parenchyma cells on theadaxial side of the spur tip. These cells appear to comprise thenectary tissue (Fig. 7H-I).
Aquilegia flowers consist of two outer whorls of sepals andpetals, followed by six to nine whorls of stamens, a singlewhorl of staminodes and an inner whorl of carpels. Each whorlcontains five organs. AfCRC is expressed in regions of the floralmeristem from which carpel primordia will emerge andsubsequent expression in carpel primordia is abaxial (Fig. 7B-D). AfCRC expression in Fig. 7D-F is clearly in carpels, but itis possible that expression in primordia in Fig. 7B is in
staminodes. Later during carpel development, AfCRC isexpressed in another domain surrounding the central vascularbundle of the carpel, but excluded from the region from whichthe vasculature will differentiate (Fig. 7E-G) resulting in aring-shaped expression pattern. However, unlike other coreeudicot species examined, we did not detect expression ofAfCRC in the nectaries of the spur (Fig. 7J).
DiscussionRole of CRC in nectary ontogenyAs developmental genetic studies in Arabidopsis indicate thatCRC is one of the key genes for nectary development (Baumet al., 2001; Bowman and Smyth, 1999; Lee et al., 2005), weanalyzed CRC expression in nectaries of exemplary eudicots.Owing to the diverse nature of nectary development in termsof both location and morphology, the current analysisrepresents only a small fraction of nectary types that exist ineudicots. However, as nectaries of Brassicaceae, Solanaceaeand Malvaceae differ dramatically in their external features andlocations, it is likely that common gene expression patternsreflect shared developmental genetics. A similar argument canbe made for floral and extrafloral nectaries. The commonexpression of CRC in nectaries of phylogenetically diverse
Development 132 (22) Research article
Fig. 5. Floral nectaries of Petunia hybrida and Nicotianabenthamiana. (A) The Petunia hybrida nectary develops as adisc at the base of the ovary but the gland (arrows) is morepronounced where the carpels are fused.(B-D,F-H,J) Longitudinal sections; (E,I,K) transverse sections.(B) PhCRC expression (blue-purple color) initiates in carpelprimordia, specifically in carpel walls. s, sepal; pe, petal; st,stamen; pl, placenta; cw, carpel wall. (C) PhCRC expression isabaxial in both carpel walls and placenta. (D) Abaxial PhCRCexpression in the carpel wall and placenta is stronger in the basalregions. PhCRC expression is also observed at the tips of theovary walls, from which the style and stigma will develop.(E) Abaxial expression is present in the carpel wall and placenta,but no expression is detected in margins of fusion of the twocarpels, similar to CRC expression in Arabidopsis. (F) As thestyle and stigma begin to develop PhCRC is expressed at the tipof the ovary. Abaxial expression in the basal region of carpelwalls persists (arrows) but expression in the placenta hasdiminished. (G) PhCRC expression has ceased except at thebase of the carpel walls (arrow) where the nectary will developin this stage 0 flower (Izhaki et al., 2002). (H) Expressionpatterns in stage 1 flowers are similar to that of stage 0 flowerswith higher expression at the base of ovary wall (arrow).(I) PhCRC expression is present throughout developingnectaries in stage 4 flowers. The two lobes of the nectary areindicated by arrows. (J) PhCRC expression is very strong andspecific to the nectary (arrows) in stage 6 flowers. (K) PhCRCexpression levels decrease after anthesis but persist throughoutthe nectary in stage 8 flowers. (L) VIGS-mediated silencing ofCRC results in a loss of nectaries in Nicotiana benthamiana. Inwild type (flower on left), nectaries are clearly visible as anorange ring of tissue at the base of the ovary (arrow). WhenCRC function is reduced, a progression of severity in phenotypeis observed, with mildly affected flowers having some nectarytissue (second flower from left, arrow) and severely affectedflowers having little or no nectary tissue (two flowers on right).(M) VIGS-mediated silencing of CRC results in a loss of flowermeristem determinacy in Petunia hybrida flowers, but littlevisible effect on nectary development (arrow).
Dev
elop
men
t
5029CRABS CLAW recruitment
species and the requirement of CRC for nectary developmentin both a rosid (Arabidopsis) and an asterid (Nicotiana),representing the two major phylogenetic lineages within coreeudicots, suggest that CRC is a general regulator of nectarydevelopment in core eudicot species regardless of nectaryposition and morphology. Alternatively, independentrecruitments of CRC orthologs in each lineage with differenttypes of nectaries could have occurred; although notimpossible, such a scenario is less parsimonious as it wouldrequire many additional changes in gene regulation, and maybe considered less likely by Occam’s razor. Unlike the rosidand asterid species examined, in Aquilegia, a basal eudicotspecies, CRC is not expressed in nectar spurs, suggesting thatCRC is not required for nectary development in this species.We suggest that CRC was recruited to be a regulator of nectarydevelopment within the eudicots, concomitant with thephylogenetic association of nectaries with the floralreproductive organs.
Ancestral and derived functions of CRCThe CRC ortholog in Oryza sativa (rice) is a key regulator ofcarpel development (Yamaguchi et al., 2004) and ourexpression and functional analyses indicate that CRC acts as aregulator of carpel development throughout the eudicotlineages. As the species examined in this study, along withOryza and Amborella (Fourquin et al., 2005), span thephylogenetic diversity of flowering plants, this role of CRC is
probably conserved throughout angiosperms. As carpelsevolved only once prior to the diversification of floweringplants, CRC might have evolved or have been recruited as aregulator of carpel development in the last common ancestorof angiosperms, a scenario consistent with the phylogeny ofthe YABBY gene family (Fig. 2). Based on genetic analyses inArabidopsis, CRC is required for both proper establishment ofcarpel polarity and carpel identity, with CRC expressedabaxially in the valves and in placental tissues (Alvarez andSmyth, 1999; Bowman and Smyth, 1999; Eshed et al., 1999).The abaxial expression pattern is conserved among speciesexamined in this study and a similar abaxial expression pattern
Fig. 6. Change versus conservation of CRC regulation.(A-E) Expression of pGhCRC(8kb)::GUS in Arabidopsis. GUSexpression is detected in the vasculature of the hypocotyl andcotyledons in 1-week-old seedlings (A), in rosette leaves (B) and incauline leaves (C) of older plants. GUS expression is also detected inthe pedicels and receptacles of flowers (D), and the styles and stigmaof gynoecia (E). (F) pLaCRC::GUS in the Arabidopsis nectary in astage 15 flower. (G) pAtCRC::GhCRC is sufficient to rescue thenectary and carpel phenotypes in a crc-1. GUS expression in thenectary is provided by a pAtCRC::GUS transgene. (H) pAtCRC::GUSin Nicotiana tabacum flower. GUS expression is in the nectary disc(arrow) and style of the gynoecium.
Fig. 7. Nectaries and carpels of Aquilegia formosa flowers.(A) Owing to growth in the pedicel, the nectar spurs of Aquilegiaformosa flowers, which develop from the abaxial side of the petals,point upwards (arrow). (B) As carpel (c) primordia emerge, AfCRCexpression (B-F) is restricted to the abaxial side as seen in thisflower. st, stamen; p, petal; se, sepal. (C) AfCRC expression precedesthe emergence of carpels. As carpel primordia begin to elongate,AfCRC expression is excluded from the vasculature and restricted tocells adjacent to the vasculature (D,E). In developing carpels, AfCRCexpression is specifically localized to a ring of cells surrounding thevasculature in the abaxial regions (E,F). (G) Anatomy of the carpel.Each of the five carpels is separate with ovules initiating from themargins of the carpels. AfCRC expression is excluded from thevasculature (arrow) but is expressed in cells surrounding it.(H,I) Anatomy of the nectar spur. The tip of the developing nectarspur consists of densely cytoplasmic parenchymatous cells,characteristic of nectary tissue (arrow in H). (J) No detectable AfCRCexpression was observed in developing nectar spurs (arrow).
Dev
elop
men
t

5030
of CRC is observed in Amborella (Fourquin et al., 2005),suggesting this domain of function may be ancestral forangiosperms. In Petunia, CRC is also expressed abaxially inthe placenta, a domain that may be related to the placentalexpression domain in Arabidopsis. Other expression domainssuch as the tips of the carpel in Petunia, and regionssurrounding the vascular bundles in Aquilegia may representindependent recruitments of CRC for specific functions, similarto the recruitment of CRC to leaf midvein development in atleast some grasses (Yamaguchi et al., 2004).
By contrast, expression analyses suggest that the role ofCRC as a regulator of nectary development is restricted to aclade within the eudicots. Although nectaries within coreeudicots differ enormously in their structures, a commonalityis their association with the floral reproductive organs. InArabidopsis, CRC is activated in nectaries by a combination ofMADS box proteins encoded by the B and C class floralhomeotic genes and SEPALLATA genes (Baum et al., 2001; Leeet al., 2005). Because the B, C and SEPALLATA genes areintimately involved in the specification of reproductive floralorgan identity, the recruitment of CRC as a nectary regulatormight have played a role in localizing floral nectaries nearstamens and carpels in core eudicots. Based on thephylogenetic distributions of nectary locations (Fig. 1), wespeculate that this could have occurred in the common ancestorof the core eudicots and the Proteales/Sabiaceae. Analyses ofCRC in additional basal eudicots, such as Grevillea robusta(Fig. 1), are required to lend further support to this hypothesis.
That the Arabidopsis CRC promoter drives expression innectaries and carpels in tobacco is consistent with the idea thatCRC is regulated similarly in these two distantly relatedeudicot species. The regulation of other genes specificallyexpressed in nectaries, which are apparently involved inbiochemical pathways in later stages of nectary development,is conserved between Brassica and Petunia, suggesting thatgene regulation at later stages of nectary development mightalso be conserved among eudicots (Ge et al., 2000; Song et al.,2000). To better understand the genetic homology and diversityof nectaries, additional genes acting in conjunction anddownstream of CRC need to be surveyed across angiospermtaxa.
Derived nectary positions within the eudicotsAlthough nectaries in most eudicots are associated withreproductive organs, some taxa, such as Capparis andGossypium, exhibit extrafloral or perianth associated nectaries.Mutant analyses in Arabidopsis have shown that in somegenetic backgrounds, e.g. leafy and apetala1 mutants, nectariesdevelop extraflorally. In these mutants, ectopic nectarystructures expressing CRC develop at the pedicel base or alongthe pedicel (Baum et al., 2001). However, ectopic expressionof CRC is not sufficient to generate ectopic nectaries, implyingthat additional genes are required for nectary development andthat multiple changes in gene regulation, including that ofCRC, might be required to alter nectary positions in eudicotlineages. When introduced into Arabidopsis, the GossypiumCRC promoter directs expression of a reporter gene in a patternreminiscent of CRC expression in Gossypium, rather than theendogenous Arabidopsis CRC pattern, suggesting that thetrans-acting factors directing the respective CRC expressionpatterns in the two species are conserved, but are largely non-
overlapping. This is in contrast to the conservation of generegulation observed with pAtCRC::GUS in Nicotiana whereboth cis- and trans-acting elements may be functionallyconserved. Thus, while activation of CRC in nectariesassociated with reproductive tissues in core eudicots could beconserved at the level of trans-acting factors, activation of CRCin nectaries in derived positions required the recruitment ofdifferent transcriptional networks.
ConclusionsBased on expression and functional analyses in a phylogeneticcontext, we propose the following scenario. Near the base ofthe eudicots, CRC was recruited to play a role in nectarydevelopment. This recruitment might have been concomitantwith nectaries being associated with floral reproductive organsin the earliest branching eudicot lineages, i.e. possibly justsubsequent to the divergence of the Ranunculids from the othereudicots (Fig. 1). As promoting carpel identity or polarity wasprobably the ancestral function of CRC in angiosperms, therecruitment of CRC to nectary development may have entailedCRC expression being brought under control of additionaltrans-acting factors together with an expansion of thedownstream targets of CRC itself. Derived nectary positionswithin the eudicots appear to result from changes in cis-actingsequences controlling CRC expression rather than alterationsin the encoded protein. Thus, the recruitment of CRC to a rolein nectary development probably entailed similar molecularchanges as proposed for other transcription factors wherechanges in gene expression patterns, rather than alterations inprotein activity, were responsible for driving changes inmorphological evolution (Doebley and Lukens, 1998).
Shared CRC gene expression in diverse nectaries of coreeudicots is in some ways comparable with shared generegulation during eye development. Specification of the eyefield requires homologous members of a retinal determinationgene network, including PAX6; the network is conserved fordifferent types of eyes in Bilateria (Silver and Rebay, 2004).This striking conservation of master regulators of eyedevelopment challenged the long-standing view of multipleorigins of eye evolution (Abouheif, 1997). However,developmental and physiological differences between insectand human eyes are enormous, suggesting differentialrecruitment of downstream programs of eye formation in eachlineage under the same master regulators (Pichaud et al., 2001;Nilsson, 2004). Such genetic conservation of master regulationis explained as a result of evolutionary constraints of regulatorycircuits for cuing some developmental processes (Hodin,2000). Identifying where genetic differences occur in each eyedevelopment pathway will shed light on understanding thedevelopmental differences arising during evolution. Likewise,identification of additional nectary regulators is required togain insight into the conservation and divergence of nectarieswithin angiosperms.
We thank Dr Beatriz Xoconostle-Cazares and Dr Roberto Ruiz-Medrano for providing in situ RT-PCR techniques, Ravit Eshed fortransforming pAtCRC::GUS into tobacco, Dr Jonathan Wendel atTexas A&M University for providing F2 seeds of G. hirsutum � G.tomentosum, Dr Andrew Patterson at University of Georgia forGhCRC mapping, and Fairchild Tropical Botanical Garden (FL, USA)for providing Capparis flexuosa. We thank Sandy Floyd, Chris
Development 132 (22) Research article
Dev
elop
men
t

5031CRABS CLAW recruitment
Zalewski and other members of the Bowman laboratory for commentsand discussions. This work was made possible with funding from theDepartment of Energy, Division of Biosciences: DE-FG03-97ER20272 (J.L.B.).
Supplementary materialSupplementary material for this article is available athttp://dev.biologists.org/cgi/content/full/132/22/5021/DC1
ReferencesAbouheif, E. (1997). Developmental genetics and homology: a hierarchical
approach. Trends Ecol. Evol. 12, 405-408.Alvarez, J. and Smyth, D. R. (1999). CRABS CLAW and SPATULA, two
Arabidopsis genes that control carpel development in parallel withAGAMOUS. Development 126, 2377-2386.
Ambrose, B. A., Lerner, D. R., Ciceri, P., Padilla, C. M., Yanofsky, M. andSchmidt, R. J. (2000). Molecular and genetic analysis of the Silky1 genereveal conservation in floral organ specification between eudicots andmonocots. Mol. Cell 5, 569-579.
APG (1998). An ordinal classification for the families of flowering plants.Annu. Missouri Bot. Gard. 85, 531-553.
Baum, S. F. and Rost, T. L. (1996). Root apical organization in Arabidopsisthaliana 1. Root cap and protoderm. Protoplasma 192, 178-188.
Baum, S. F., Eshed, Y. and Bowman, J. L. (2001). The Arabidopsis nectaryis an ABC-independent floral structure. Development 128, 4657-4667.
Bowman, J. L. and Smyth, D. R. (1999). CRABS CLAW, a gene that regulatescarpel and nectary development in Arabidopsis, encodes a novel protein withzinc finger and helix-loop-helix domains. Development 126, 2387-2396.
Brown, W. H. (1938). The bearing of nectaries on the phylogeny of floweringplants. Proc. Am. Phil. Soc. 79, 549-595.
Chen, J.-C., Jiang, C.-Z., Gookin, T., Hunter, D., Clark, D. and Reid, M.(2004). Chalcone synthase as a reporter in virus-induced gene silencingstudies of flower senescence. Plant Mol. Biol. 55, 521-530.
Crepet, W. L. and Friis, E. M. (1987). The evolution of insect pollination inangiosperms. In The Origin of Angiosperms and their BiologicalConsequences (ed. E. M. Friis, W. G. Chaloner and P. R. Crane), pp. 181-202. Cambridge: Cambridge University Press.
Darwin, F. (1877) On the nectar glands of the common brakeferns. Bot. J.Linn. Soc. 15, 407-409.
Doebley, J. and Lukens, L. (1998). Transcriptional regulators and theevolution of plant form. Plant Cell 10, 1075-1082.
Elias, T. S. (1983). Extrafloral nectaries: their structure and distribution. InThe Biology of Nectaries (ed. B. Bentley and T. S. Elias), pp. 174-203. NewYork: Columbia University Press.
Endress, P. K. (1986). Floral structure, systematics, and phylogeny inTrochodendrales. Annu. Missouri Bot. Gard. 73, 297-324.
Endress, P. K. (2001). The flowers in extant basal angiosperms and inferenceson ancestral flowers. Int. J. Plant Sci. 162, 1111-1140.
Erbar, C., Kusma, S. and Leins, P. (1998). Development and interpretationof nectary organs in Ranunculaceae. Flora 194, 317-332.
Eriksson, O. and Bremer, B. (1992). Pollination systems, dispersal modes,life forms, and diversification rates in angiosperm families. Evolution 46,258-266.
Eshed, Y., Baum, S. F. and Bowman, J. L. (1999). Distinct mechanismspromote polarity establishment in carpels of Arabidopsis. Cell 99, 199-209.
Fahn, A. (1953). The topography of the nectary in the flower and itsphylogenetic trend. Phytomorphology 3, 424-426.
Fahn, A. (1979). Secretory tissues in plants. New York: Academic Press.Ferrario, S., Immink, R. G. and Angenent, G. C. (2004). Conservation and
diversity in flower land. Curr. Opin. Plant Biol. 7, 84-91.Fourquin, C., Vinauger-Douard, M., Fogliani, B., Dumas, C. and Scutt, C.
P. (2005). Evidence that CRABS CLAW and TOUSLED have conserved theirroles in carpel development since the ancestor of the extant angiosperms.Proc. Natl. Acad. Sci. USA 102, 4649-4654.
Ge, Y.-X., Angenent, G. C., Wittich, P. E., Peters, J., Franken, J., Busscher,M., Zhang, L.-M., Dahlhaus, E., Kater, M. M., Wullems, G. J. et al.(2000). NEC1, a novel gene, highly expressed in nectary tissue of Petuniahybrida. Plant J. 24, 725-734.
Golz, J. F., Keck, E. J. and Hudson, A. (2002). Spontaneous mutations inKNOX genes give rise to a novel floral structure in Antirrhinum. Curr. Biol.12, 515-522.
Hall, J. C., Systsma, K. J. and Iltis, H. H. (2002). Phylogeny of Capparaceaeand Brassicaceae based on chloroplast sequences data. Am. J. Bot. 89, 1826-1842.
Hillis, D. M. (1994). Homology in molecular biology. In Homology: TheHierarchical Basis of Comparative Biology (ed. B. K. Hall), pp. 339-367.New York: Academic Press.
Hodin, J. (2000). Plasticity and constraints in development and evolution. J.Exp. Zool. 288, 1-20.
Holland, N., Holland, D., Helentjaris, T., Dhugga, K. S., Xoconostle-Cazares, B. and Delmer, D. P. (2000). A comparative analysis of the plantcellulose synthase (CesA) gene family. Plant Physiol. 123, 1313-1323.
Huelsenbeck, J. P. and Ronquist, F. (2001). MrBayes: Bayesian inference ofphylogeny. Bioinformatics 17, 754-755.
Izhaki, A., Borochov, A., Zamski, E. and Weiss, D. (2002). Gibberellinregulates post-microsporogenesis processes in Petunia anthers. Physiol.Plant 115, 442-447.
Kyozuka, J., Kobayashi, T., Morita, M. and Shimamoto, K. (2000).Spatially and temporally regulated expression of rice MADS box genes withsimilarity to Arabidopsis class A, B and C genes. Plant Cell Physiol. 41,710-718.
Lee, J.-Y., Baum, S. F., Alvarez, J., Patel, A., Chitwood, D. H. andBowman, J. L. (2005). Activation of CRABS CLAW in the nectaries andcarpels of Arabidopsis. Plant Cell 17, 25-36.
Liu, Y., Schiff, M. and Dinesh-Kumar, S. P. (2002). Virus-induced genesilencing in tomato. Plant J. 31, 777-786.
Meeuse, A. D. J. (1978). Nectarial secretion, floral evolution, and thepollination syndrome in early angiosperms. Proc. K. Ned. Akad. B Ph. 81,300-326.
Meyer, J. R. and Meyer, V. G. (1961). Origin and inheritance of nectarilesscotton. Crop Sci. 1, 167-169.
Nilsson, D.-E. (2004). Eye evolution: a question of genetic promiscuity. Curr.Opin. Neurobiol. 14, 407-414.
Norris, T. (1941). Torus anatomy and nectary characteristics as phylogeneticcriteria in the Rhoeadales. Am. J. Bot. 28, 101-113.
Pellmyr, O. (1992). Evolution of insect pollination and angiospermdiversification. Trends Ecol. Evo. 7, 46-49.
Pichaud, F., Treisman, J. and Desplan, C. (2001). Reinventing a commonstrategy for patterning the eye. Cell 105, 9-12.
Porsch, O. (1910). Ephedra campylopoda C. A. Mey, eine entomophyleGymnosperme. Ber. Dtsch. Bot. Ges. 31, 580-590.
Posada, D. and Crandall, K. A. (1998). Modeltest: testing the model of DNAsubstitution. Bioinformatics 14, 817-818.
Proctor, M., Yeo, P. and Lack, A. (1996). The natural history of pollination.Portland, Oregon: Timber Press.
Qiu, Y.-L., Lee, J., Bernasconi-Quadroni, F., Soltis, D. E., Soltis, P. S.,Zanis, M., Zimmer, E. A., Chen, Z., Savolainen, V. and Chase, M. W.(1999). The earliest angiosperms: evidence from mitochondrial, plastid andnuclear genes. Nature 402, 404-407.
Ronse DeCraene, L. P. (2004). Floral development of Berberidopsiscorallina: a crucial link in the evolution of flowers in the core eudicots.Annu. Bot. 94, 741-751.
Sawa, S., Watanabe, K., Goto, K., Kanaya, E., Morita, E. M. and Okada,K. (1999). Filamentous Flower, a meristem and organ identity gene ofArabidopsis encodes a protein with a zinc finger and HMG-related domains.Genes Dev. 13, 1079-1088.
Siegfried, K. R., Eshed, Y., Baum, S. F., Otsuga, D., Drews, G. N. andBowman, J. L. (1999). Members of the YABBY gene family specify abaxialcell fate in Arabidopsis. Development 126, 4117-4128.
Silver, S. and Rebay, I. (2004). Signaling circuitries in development: insightsfrom the retinal determination gene network. Development 132, 3-13.
Small, R. L., Ryburn, J. A., Cronn, R. C., Seelanan, T. and Wendel, J. F.(1998). The tortoise and the hare: choosing between noncoding plastomeand nuclear Adh sequences for phylogeny reconstruction in a recentlydiverged plant group. Am. J. Bot. 85, 1301-1315.
Soltis, P. S., Soltis, D. E. and Chase, M. W. (1999). Angiosperm phylogenyinferred from multiple genes as a tool for comparative biology. Nature 402,402-404.
Soltis, D. E., Soltis, P. S., Chase, M. W., Mort, M. E., Albach, D. C., Zanis,M., Savolainen, V., Hahn, W. H., Hoot, S. B., Fay, M. F. et al. (2000).Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences.Bot. J. Linn. Soc. 133, 381-461.
Song, J. T., Seo, H. S., Song, S. I., Lee, J. S. and Choi, Y. D. (2000). NTR1encodes a floral nectary-specific gene in Brassica campestris L. ssp.pekinensis. Plant Mol. Biol. 42, 647-655.
Dev
elop
men
t

5032
Swofford, D. L. (2002). PAUP*: phylogenetic analysis using parsimony (*andother methods), version 4.0b10. Sunderland, MA: Sinauer Press.
Swofford, D. L., Olsen, G. K., Waddell, P. J. and Hillis, D. M. (1996).Phylogeny reconstruction. In Molecular Systematics, 2nd ed. (ed. D. M.Hillis, C. Moritz and B. K. Mable), pp. 407-514. Sunderland, MA: SinauerPress.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. and Higgins,D. G. (1997). The Clustal-X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. Nucleic AcidsRes. 25, 4876-4882.
Vielle-Calzada, J. P., Thomas, J., Spillane, C., Coluccio, A., Hoeppner, M.A. and Grossniklaus, U. (1999). Maintenance of genomic imprinting at theArabidopsis medea locus requires zygotic DDM1 activity. Genes Dev. 13,2971-2982.
Von Balthazar, M. and Endress, P. K. (2002). Development of inflorescencesand flowers in Buxaceae and the problem of perianth interpretation. Int. J.Plant Sci. 163, 847-876.
Xoconostle-Cázares, B., Xiang, Y., Ruiz-Medrano, R., Wang, H.-L.,Monzer, J., Yoo, B.-C., McFarland, K. C., Franceschi, V. R. and Lucas,W. J. (1999). Plant paralog to viral movement protein that potentiatestransport of mRNA into the phloem. Science 283, 94-98.
Yamada, T., Ito, M. and Kato, M. (2004). YABBY2-homologue expressionin lateral organs of Amborella trichopoda (Amborellaceae). Int. J. Plant Sci.165, 917-924.
Yamaguchi, T., Nagasawa, N., Kawasaki, S., Matsuoka, M., Nagato, Y. andHirano, H. Y. (2004). The YABBY gene DROOPING LEAF regulatescarpel specification and midrib development in Oryza sativa. Plant Cell 16,500-509.
Yang, S. J., Carter, S. A., Cole, A. B., Cheng, N. H. and Nelson, R. S.(2004). A natural variant of a host RNA-dependent RNA polymerase isassociated with increased susceptibility to viruses by Nicotianabenthamiana. Proc. Natl. Acad. Sci. USA 101, 6297-6302.
Zimmerman, J. (1932). Über die extrafloralen nektarien der Angiospermen.Bot. Cent. Beih. 49, 99-196.
Development 132 (22) Research article
Dev
elop
men
t

[ 10 20 30 40 50 60][ . . . . . .]
INO MTTTLNHL---------------------------------------------------- [8]Antirrhinum_majus_B ML-TLDSL---------------------------------------------------- [7]Oryza_sativa_C MSSAARHHC--------------------------------------------------- [9]Nymphaea_alba MSQL-------------------------------------------------------- [4]CRC MNLEEKPTMTASRA---------------------------------------------- [14]Lepidium_africanum_CRC MNLDQEKPTMTSRA---------------------------------------------- [14]Capparis_flexuosa_CRC MNLEEKSVMDSQA----------------------------------------------- [13]Cleome_sparsifolia_CRC MNLEEKPAMASR------------------------------------------------ [12]Gossypium_hirsutum_CRC2 ------------------------------------------------------------ [0]Gossypium_hirsutum_CRC1 ------------------------------------------------------------ [0]Citrus_sinensis_A MNLEDNISTDLQ------------------------------------------------ [12]Antirrhinum_majus_A MDMAN------------------------------------------------------- [5]Lycopersicon_esculentum_B MDY--------------------------------------------------------- [3]Nicotiana_tabacum_CRC1 MDL--------------------------------------------------------- [3]Nicotiana_tabacum_CRC2 MDL--------------------------------------------------------- [3]Petunia_hybrida_CRC MDL--------------------------------------------------------- [3]Hedyotis_centranthoides MDL--------------------------------------------------------- [3]Grevillea_robusta_CRC MEF--------------------------------------------------------- [3]Aquilegia_formosa_CRC MDL--------------------------------------------------------- [3]Triticum_aestivum_A MDL--------------------------------------------------------- [3]Zea_mays_A MDM--------------------------------------------------------- [3]Oryza_sativa_DL MDL--------------------------------------------------------- [3]Amborella_CRC MDFL-------------------------------------------------------- [4]FIL MSMSSMSSPSSA--------VCS-PDH--------------------------------- [18]YABBY3 MSSMSMSSSSAP--------AFP-PDH--------------------------------- [18]Populus_tremula MSSSSS---------------LS-LDH--------------------------------- [11]Malus_x_domestica_A MSSAATSSTVFNSPDQQQHL---------------------------------------- [20]Vitis_shuttleworthii_A MSSSST---------------LS-LDH--------------------------------- [11]Citrus_sinensis_B MSSSTPAA-------------LS-PDH--------------------------------- [13]Antirrhinum_majus_GRAM MSSSSA---------------FA-PDHHH------------------------------- [13]Solanum_tuberosum_A MSSSNS---------------LS-LDHLPPPPPP-------------------------- [18]Lycopersicon_esculentum_A MSSSSAA--------------FA-PDHHLHHHHLS------------------------- [20]Chrysanthemum_x_morifolium MSSSAS---------------FS-PDHHHHHL---------------------------- [16]Aquilegia MSSSSA---------------FS-LDH--------------------------------- [11]ChloYAB324 MSSSSA---------------FS-LDH--------------------------------- [11]ChloYAB329 MSSSS----------------FSQLDH--------------------------------- [11]Oryza_sativa_A MSSSSSSSA-----------VFP-LDHLAA------------------------------ [18]Oryza_sativa_B MSSSSSSSASSAAAAAFRPAVVQ-REQQVVEEKFPAAAAAMREMVLPPVAAAAA------ [53]Triticum_aestivum_B MSSSSSSGASAA---------FT-PVSQQQQQMAPENLSSELQPPAPMQP---------- [40]Zea_mays_B -----------------GTSCFP-LDHLA------------------------------- [11]Nuphar_advena_A_ MSSSSIFGL--------------------------------------------------D [10]Amborella_trichopoda_A ------------------------------------------------------------ [0]YABBY5 MANS-------------------------------------------------------- [4]Populus_euphratica MSSC------------------------------------------------------ID [6]Malus_x_domestica_B MTSC-------------------------------------------------------V [5]Vitis_shuttleworthii_B ------------------------------------------------------------ [0]Gossypium_raimondii MSTCT------------------------------------------------------- [5]Antirrhinum_majus_PROL M----------------------------------------------------------- [1]Lycopersicon_esculentum_C MASCI------------------------------------------------------- [5]Solanum_tuberosum_B MASC-------------------------------------------------------I [5]YABBY2 MS---------------------------------------------------------V [3]Populus_balsamifera MSP--------------------------------------------------------- [3]Vitis_vinifera MS---------------------------------------------------------L [3]Malus_x_domestica_C MS--------------------------------------------------------MD [4]Citrus_sinensis_C MS---------------------------------------------------------L [3]Antirrhinum_majus_C MS---------------------------------------------------------L [3]Lycopersicon_esculentum_E MT--------------------------------------------------------YS [4]Persea_americana MS---------------------------------------------------------V [3]Oryza_sativa_FIL1 MS---------------------------------------------------------V [3]Oryza_sativa_D MS---------------------------------------------------------V [3]Oryza_sativa_FIL2 MSAQ-------------------------------------------------------I [5]Asparagus_officinalis ------------------------------------------------------------ [0]Lycoris_longituba ------------------------------------------------------------ [0]Nuphar_advena_ MS---------------------------------------------------------M [3]Amborella_trichopoda_B ------------------------------------------------------------ [0]Amborella_trichopoda_C MS---------------------------------------------------------L [3]Welwitschia_mirabilis MSACN------------------------------------------------GLDVDP- [11]

Cycas_rumphii_ M------------------------------------------------------EAS-- [4]Pinus_taeda MSTS-------------------------------------------------GFEVG-- [9]Picea_sitchensis MSTS-------------------------------------------------GFEVG-- [9]
[ 70 80 90 100 110 120][ . . . . . .] dCRC-ZnFINO FDLPGQICHVQCGFCTTILLVSVPFTSLSMVVTVRCGHCTSLLS--VNLMKASFIPLHLL [66]Antirrhinum_majus_B FDFQEQICYVQCGFCTTILLVSVPKNCLSMAVTVRCGHCSTILS--VNIPDAYFVPLHFF [65]Oryza_sativa_C SGLRERLGCVQCSFCATVLLVSVPCSSVLRVVAVQCGHCSGILSA-VNLPPSPVSASIEL [68]Nymphaea_alba LDLTEQLCYVQCSFCDTILLVSVPCSSLLKVVPVRCGHCSNLFS--VNMLKASFLPLQLL [62]CRC SPQAEHLYYVRCSICNTILAVGIPLKRMLDTVTVKCGHCGNLSF--LTTTPP-------L [65]Lepidium_africanum_CRC SPQAEHLYYVRCSICNTILAVGIPMKRMLDTVTVKCGHCGNLSF--LTTSLP-------L [65]Capparis_flexuosa_CRC PTQSEHLCYVRCNFCNTVLAVGIPCKRMLDTVTVKCGHCSNLSF--LSVRPP-------L [64]Cleome_sparsifolia_CRC ANQSDHLCYVRCNFCSTILAVGIPLTRMLDTVTVKCGHCGNLSF--LTTTKP-------L [63]Gossypium_hirsutum_CRC2 VPQSEHLCYVRCNFCNTVLAVGIPCKRLLETVTVKCGHCSNLSF--LSTRPP-------L [51]Gossypium_hirsutum_CRC1 VPQSEHLCYVRCNFCNTVLAVGIPCKRLLETVTVKCGHCSNLSF--LSTRPP-------L [51]Citrus_sinensis_A VPQSEHLCYVRCNFCNTVLAVGIPCKRLLDTVTVKCGHCSNLSF--LSTRPP-------Q [63]Antirrhinum_majus_A QSSSEHLCYVRCNFCSTVLAVGIPCKRLMDTVTVKCGHCSNLSF--LSTRPP-------I [56]Lycopersicon_esculentum_B VQSSEHLCYVRCNFCNTVLAVGIPYKRLMDTVTVKCGHCSNLSF--LTTRPP-------I [54]Nicotiana_tabacum_CRC1 VQSSEHLCYVRCSFCNTVLAVGIPYKRLLDTVTVKCGHCSNLSF--LSTRPP-------L [54]Nicotiana_tabacum_CRC2 VQSSEHLCYVRCSFCNTVLAVGIPYKRLLDTVTVKCGHCSNLSF--LSTRPP-------L [54]Petunia_hybrida_CRC AQTSEHLCYVRCSFCNTVLAVGIPFKRLLDTVTVKCGHCSNLSF--LSTRPP-------L [54]Hedyotis_centranthoides VQQSEHLCYVRCNFCNTVLAVGIPCKRLLDTVTVKCGHCSNLSF--LSTRPPPPPPPPTL [61]Grevillea_robusta_CRC SSSSEHLCYVRCNFCNTVLAVGIPCKRLMDTVTVKCGHCSNLSF--LSTRPPP------L [55]Aquilegia_formosa_CRC IPSPEHLCYVRCNFCSTVLAVGIPCKRTLDTVTVKCGHCGNISF--LSTRPP-------I [54]Triticum_aestivum_A VSPSEHLCYVRCTYRNTVLSVGVPCKRLMDTVTVKCGHCNNLSF--LSPRPP-----PMV [56]Zea_mays_A VSQSEHLCYVRCTYCNTVLAVGVPCKRLMDTVTVKCGHCNNLSY--LSPRPP------MV [55]Oryza_sativa_DL VSPSEHLCYVRCTYCNTVLAVGVPCKRLMDTVTVKCGHCNNLSF--LSPRPP------MV [55]Amborella_CRC PGSTDHLCYVRCNFCDTLLAVGVPCRRLMDTVTVKCGHCSHLSF--LSARP-------LL [55]FIL FSPSDHLCYVQCNFCQTILAVNVPYTSLFKTVTVRCGCCTNLLS--VNMRS-------YV [69]YABBY3 FSSTDQLCYVHCSFCDTVLAVSVPPSSLFKTVTVRCGHCSNLLSVTVSMRA-------LL [71]Populus_tremula LPPSEQLCYVHCNICDTVLAVSVPCTSLFKTVTVRCGHCTNLLP--VNMRG-------LF [62]Malus_x_domestica_A LSPTDQLCYVHCNFCDTVLAVSVPCSSLFKTVTVRCGHCSNLLS--VNMRA-------LL [71]Vitis_shuttleworthii_A LPPSEQLCYVHCNICDTVLAVSVPCTSLFKTVTVRCGHCTNLLP--VNLRG-------LL [62]Citrus_sinensis_B LSSSEQLCYVHCTFCDTVLAVSVPCTSLFKTVTVRCGHCTNLLS--VNMRG-------LL [64]Antirrhinum_majus_GRAM LTPSEQLCYVHCNFCDTVLAVSVPCTSLFKTVTVRCGHCTNLLS--VNMRG-------LL [64]Solanum_tuberosum_A PPPSEQLCYVHCNVCDTVLAVSVPCTSLFKTVTVRCGHCTNLLP------G-------WL [65]Lycopersicon_esculentum_A PPSGEQLCYVQCNFCDTVLAVSVPCSSFFKNVTVRCGHCTNLLS--VN----------II [68]Chrysanthemum_x_morifolium SPTSEQLCYVQCNFCDTVLAVSVPCGSLFTTVTVRCGHCTNLLS--VNMRA-------LL [67]Aquilegia LSPSEQLCYVHCNFCDTVLAVSVPCSSLFKTVTVRCGHCTNLLS--VNMRG-------LL [62]ChloYAB324 ISPSDQLCYVHCNFCDTVLAVSV?CSILYKTVTVRCGHCTNLLS--VNMRG-------LL [62]ChloYAB329 IPPSDQLCYVHCNLCDTVLVVSVPYTSFFKTVTVRCGQCTNLLS--VNMRG-------LL [62]Oryza_sativa_A PSPTEQLCYVHCNCCDTILAVGVPCSSLFKTVTVRCGHCANLLS--VNLRG-------LL [69]Oryza_sativa_B DSEQEQLCYVHCHYCDTVLVVSVPSSSLFETVTVRCGHCSSLLT--VNMRG-------LL [104]Triticum_aestivum_B EAPSEQLCYVHCHFCDTVLVVSVPSSSLFKTVTVRCGHCSSLLT--VDMRG-------LL [91]Zea_mays_B PSPTEQLCYVHCNCCDTILAVGVPCSSLFKTVTVRCGHCANLLS--VNLRG-------LL [62]Nuphar_advena_A_ HLQPEQLCYVHCNFCNTVLAVSVPCSSLFKIVTVRCGHCTNLLS--VNMRA-------LL [61]Amborella_trichopoda_A GLASEQLCYVHCNFCDTVLAVSVPCSSLFKMVTVRCGHCTNVLS--VDTRG-------LL [51]YABBY5 VMATEQLCYIPCNFCNIILAVNVPCSSLFDIVTVRCGHCTNLWS--VNMAAA------LQ [56]Populus_euphratica AVPNEQLRYIPCNFCNIVLAVSVPCSSLFDIVTVRCGHCTNIWS--VNMAAT------FQ [58]Malus_x_domestica_B DVATEQLCYIPCNFCSIVLAVSVPCSSLFDIVTVRCGHCTNLWS--VNMAAA------FQ [57]Vitis_shuttleworthii_B DVASEQLCYIPCNFCNIVLAVSVPCSSLFDIVTVRCGHCTNLWS--VNMAAA------FQ [52]Gossypium_raimondii NLAPEQLCYIPCNFCNIVLAVSVPCSSLFDIVTVRCGHCTNLWS--VNMAAA------FQ [57]Antirrhinum_majus_PROL DV-PEQLCYISCNFCSIVLAVSVPCSSLFDVVTVRCGHCTNLWS--VNMAAAAA----FQ [54]Lycopersicon_esculentum_C DVASDQLCYIPCNFCNIVLAVSVPCSSLFDIVTVRCGHCTNLWS--VNMAAA------FQ [57]Solanum_tuberosum_B DVASDQLCYIPCNFCNIVLAVSVPCSSLLDIVTVRCGHCTNLWS--VNMAAA------FQ [57]YABBY2 DFSSERVCYVHCSFCTTILAVSVPYASLFTLVTVRCGHCTNLLS--LNIGVS------LH [55]Populus_balsamifera ELVSEPVCYVHCNLCNTILAVSVPSSSLFNIVTVRCGHCGNLLS--VNMGAS------LQ [55]Vitis_vinifera DITPERVCYVHCNFCNTILAVSVPCTSLFTIVTVRCGHCANLLS--VNMGAL------LQ [55]Malus_x_domestica_C LMASERVCYVHCNFCNTILAVSVPCNSSMNMVTVRCGHCANLLS--VNIGGS------LQ [56]Citrus_sinensis_C DLASERVCYVHCNYCNTILAVSVPCSSLFNIVTVRCGHCANLLN--VNMVSA------LQ [55]Antirrhinum_majus_C DMTSERVCYVHCNFCNTILAVSVPCSSMFTIVTVRCGHCANLLS--VNMGAL------LQ [55]Lycopersicon_esculentum_E SSSSERVCYVHCNFCSTILAVSVPCNSMMTIVTVRCGHCANLLS--VNIAPS------LQ [56]Persea_americana DVSCERMCYVHCNLCNTILAVSVPYSNLFNVVTVRCGHCAYLLP--VNVGSM------IQ [55]Oryza_sativa_FIL1 QFTSEHVCYVNCNYCNTILVVNVPNNCSYNIVTVRCGHCTMVLS--MDLAP-------FH [54]Oryza_sativa_D QFTSEHVCYVNCNYCNTILVVNVPNNCSYNIVTVRCGHCTMVLS--MDLAP-------FH [54]Oryza_sativa_FIL2 VPAPEHVCYVHCNFCNTIFAVSVPSNSMLNIVTVRCGHCTSLLS--VNLRGL------VQ [57]Asparagus_officinalis VIAPEHVCYVHCNFCNTILAVNVPSNNLFSIVTVRCGHCANLLS--VNMGAL------VQ [52]Lycoris_longituba QLATEHVCYVHCNFCNTILAVNVPGNYLLNIVTVRCGHCTNLLS--VNMGSL------LQ [52]Nuphar_advena_ EMGNEHVYYLPCNFCNTILAVSVPCTNLFNIVTIRCGHCSNLQS--VNMGAL------LP [55]Amborella_trichopoda_B ---------------------------------VRCGHCTNLLS--VNMAAV------LH [19]Amborella_trichopoda_C ENPSEHVCYVQCNLCNTILAVSVPGSCLFGIVTVRCGHCTNLLS--MNMGAL------LQ [55]Welwitschia_mirabilis ---REQLCYVPCTNCSTVLAVSVPLKSLFKVVTIRCGHCNNLIS--VDMRG-------LT [59]

Cycas_rumphii_ ---SEHLCYVSCSFCSTILAVSVPCSSLFKIVTVRCGHCTNLLS--VNMGGL------LQ [53]Pinus_taeda ---SDHLCYVHCDYCNTILAVNVPCNSLYRTVTVRCGHCNNLLS--VNMAPL------LQ [58]Picea_sitchensis ---QDHLCYVHCDFCSTILAVNVPCNSLYRIVTVRCGHCTNLLS--VNMAPL------LQ [58]
[ 130 140 150 160 170 180][ . . . . . .]
INO AS---------------------------------LSHLDETGKEEVAATDGVEEEAWKV [93]Antirrhinum_majus_B SS------------------------------INQQEQMSIQPKQEACSVEMAGDHKKAG [95]Oryza_sativa_C TP---------------------------------------QMKQELDAGPPP------- [82]Nymphaea_alba AS------------------------------------INNEAKQDSFENAPVKI----- [81]CRC QG------------------------------------------HVSLTLQ--MQSFGGS [81]Lepidium_africanum_CRC HG------------------------------------------HVSLTLQ--MQSFGGS [81]Capparis_flexuosa_CRC HG--QC-LDH----------------------------------QVNLTLQ--TQSFCGN [85]Cleome_sparsifolia_CRC QG--QC-LDR----------------------------------HVSLTLQ--MQSFGGS [84]Gossypium_hirsutum_CRC2 QG--QC-LDP----------------------------------QTSLTLQ----SFCG- [69]Gossypium_hirsutum_CRC1 QG--QC-LDP----------------------------------QTSLTLQ----SFCG- [69]Citrus_sinensis_A QGPSQM-SLR---------------------------------------FQ-EKQSFCN- [81]Antirrhinum_majus_A QG--QY-YDH----------------------------------QT--SLH--HQSLCS- [74]Lycopersicon_esculentum_B QG--QC-FDH----------------------------------QP--NIQ----GYCS- [70]Nicotiana_tabacum_CRC1 QG--QC-FDH----------------------------------QS--ALQ--HQTFFS- [72]Nicotiana_tabacum_CRC2 QG--QC-FDH----------------------------------QT--ALQ--HQAFFS- [72]Petunia_hybrida_CRC QG--QC-FDH----------------------------------QT--ALQ--HQAFFS- [72]Hedyotis_centranthoides QP--QS-FDH--------------------------------------PPS--IQSFFS- [77]Grevillea_robusta_CRC QG--QF-LDR--------------------------------HYPVPLPCQ--MQGFSA- [77]Aquilegia_formosa_CRC QG--QC-LDH----------------------------------QV-DAFQSFRN----- [71]Triticum_aestivum_A QP--LSPNDH---------------------------------HHPMGPFQGWT------ [75]Zea_mays_A QP--LSPTDH-----------------------------------PLGPFQCQGPCN--- [75]Oryza_sativa_DL QP--LSPTDH-----------------------------------PLGPFQGPCT----- [73]Amborella_CRC QN--QSLELLST--------------------------------QN---FCG-------- [70]FIL LPA--------------------S---------------------NQLQLQLGPHSYF-- [86]YABBY3 LPS--------------------VS-------------------------NLG-HSFL-- [83]Populus_tremula LPS--------------------AS-----------------------QFPLG-HNFY-- [76]Malus_x_domestica_A LPP--------------------P--------------------NNNNQLHLP-HHFF-- [88]Vitis_shuttleworthii_A LPS--------------------A-----------------------NQLHLG-HAFF-- [76]Citrus_sinensis_B LPA--------------------A-----------------------NQLHLG-HPFF-- [78]Antirrhinum_majus_GRAM LPA--------------------A-----------------------NQLHLG-HSFF-- [78]Solanum_tuberosum_A LPS--------------------T----------------------NHHHHFG-HTYF-- [80]Lycopersicon_esculentum_A LPT--------------------AA----------------------NQLHLP-HSFF-- [83]Chrysanthemum_x_morifolium FPA--------------------SVTTTAAA----------------NQFHLG-HNFF-- [88]Aquilegia LPT--------------------GA----------------------NQLHFG-HSFF-- [77]ChloYAB324 LPS--------------------A-----------------------NQLHLG-HTFF-- [76]ChloYAB329 LAS--------------------A-----------------------HQLNLG-HSLF-- [76]Oryza_sativa_A LPA--------------------PAPAPA------------------NQLHFG-PSLL-- [88]Oryza_sativa_B LPT--------------------TAAAAPPPPPPPPPPPPPPAA------HFP-HSLNLA [137]Triticum_aestivum_B FPT----------------TTTTVAAESAASAVTTTTSPPPAAAAHHGQFHYP-SSLNLA [134]Zea_mays_B LPP--------------------AAPAPP------------------NHLNFA-HSLLSP [83]Nuphar_advena_A_ QPS-----------------PPPEPA--------------------LTPPSLPNHQFF-- [82]Amborella_trichopoda_A HPT-----------------AAT-------------------------QLHLG-HAFF-- [66]YABBY5 S--LSRPN----FQ---------------------------------------------- [64]Populus_euphratica S--LP-------WQDQV--Q---------------------------------------- [67]Malus_x_domestica_B S--PS-------WQD-V--Q---------------------------------------- [65]Vitis_shuttleworthii_B S--LS-------WQD-V--Q---------------------------------------- [60]Gossypium_raimondii S--LS-------WQD-V--QV--------------------------------------- [66]Antirrhinum_majus_PROL S--LQ-PH----WQDAVVHQ---------------------------------------- [67]Lycopersicon_esculentum_C S--SSSAS----WQN---HQV--------------------------------------- [69]Solanum_tuberosum_B S--SSS-------QNHH-IQV--------------------------------------- [68]YABBY2 QTSAPPI-----HQDLQ---PHRQHTTSLVTR---------------------------- [79]Populus_balsamifera T--LP-------LQDPQ------KLLLINS-------EDLN------------------- [74]Vitis_vinifera T--VPT-------QDLQ----SQKQQLSC--------GDPS------------------- [75]Malus_x_domestica_C STTIPP------HQDPQ------KQHLSS--------EDSN------------------- [77]Citrus_sinensis_C A--VP-------LQDPQ----LQKQHINL--------EDSIT------------------ [76]Antirrhinum_majus_C S--VH-------LQDFQ------KQQH----------GEAAA------------------ [72]Lycopersicon_esculentum_E S--LP-------IQDLQ------RQNESSI-------EDGMS------------------ [76]Persea_americana T--FP-------FQDFQ------HQNFSCQ------DSD--------------------- [73]Oryza_sativa_FIL1 Q--ART------VQD--------HQVQNRGFQGN-------------------------- [72]Oryza_sativa_D Q--ART------VQD--------HQVQNRGFQGN-------------------------- [72]Oryza_sativa_FIL2 A--LPAEDH---LQDNL------KMHNMSFR-------ENYS------------------ [81]Asparagus_officinalis A--L----H---LQDFQ------KLQNHQVAS-----QDSHG------------------ [74]Lycoris_longituba A--LP-------LQDLQ------KFQNHQKAS-----QGSRG------------------ [74]Nuphar_advena_ AA-IP-------IQGVQLQN---HAYNS---------QQHGM------------------ [77]Amborella_trichopoda_B APSLTPAPAN--LQHDQLL----RNQSLTGSHGYNRMMMMMTTM---------------- [57]Amborella_trichopoda_C T--IPFHD----LQN--------QSVAPQER-QRM------------------------- [75]Welwitschia_mirabilis ---VPQFFQSQP-VPALSVG---------------------------------------- [75]

Cycas_rumphii_ P--LP-YQAPTGQQNQSFNS----YDHHRK------------------------------ [76]Pinus_taeda S--LA-------QDPQQQGQNTGTYDYNR------------------------------- [78]Picea_sitchensis S--LP-------QQAQQQGPNTGPHDYTR------------------------------- [78]
[ 190 200 210 220 230 240][ . . . . . .]
INO NQEKENSPTTL--VSSSDNED--------------------------------------- [112]Antirrhinum_majus_B MTLCFSS----------DEEEY-------------------------------------- [107]Oryza_sativa_C --------GEYSDESSGDDREGRDAE---------------------------------- [100]Nymphaea_alba --GD----TTF------MESLY-------------------------------------- [91]CRC ----------------DYK--KGSSSSSSS-------STSS------------------- [97]Lepidium_africanum_CRC ----------------EYK--KGSSSSSSS-------STSS------------------- [97]Capparis_flexuosa_CRC ----------------ELK--KGSSSSSSSS------STSS------------------- [102]Cleome_sparsifolia_CRC N---------------ELK--KGGSSSSSSS------STSS------------------- [102]Gossypium_hirsutum_CRC2 ----------------DFR--KGTQFPSPSS------STSS------------------- [86]Gossypium_hirsutum_CRC1 ----------------DFR--KGTQFPSPSS------STSS------------------- [86]Citrus_sinensis_A ----------------DFK--LGNASSSSS-------STSS------------------- [97]Antirrhinum_majus_A ----------------EFK--KGGSSSFSS-------STSS------------------- [90]Lycopersicon_esculentum_B ----------------ELK--NKGQASSSTS------STSS------------------- [87]Nicotiana_tabacum_CRC1 ----------------DFK--KG-QSSS---------SSSS------------------- [85]Nicotiana_tabacum_CRC2 ----------------DFK--KG-QSSS---------SSSS------------------- [85]Petunia_hybrida_CRC ----------------DYK--KG-QSSSSFS------SSSS------------------- [88]Hedyotis_centranthoides ----------------KFK--KG-QTSSSASS-----STSS------------------- [94]Grevillea_robusta_CRC ----------------DYR--KG-QSSQSLTSS----STSS------------------- [95]Aquilegia_formosa_CRC ----------------EYR--KG-QSSSSSSSTSCGQPTSPN------------------ [94]Triticum_aestivum_A ----------------DCR--RN-QPLPPLAS-----PTSS------------------- [92]Zea_mays_A ----------------DCR--RN-QPLPLAS------PSST------------------- [91]Oryza_sativa_DL ----------------DCR--RN-QPLPLVS------PTS-N------------------ [89]Amborella_CRC ----------------DNK--KSQQSSSSS-------PLTP------------------- [86]FIL --------NPQD--ILEELRDAPSNMNMMMMNQHPTM----NDIPSFM------------ [120]YABBY3 --PPPPPPPPPN--LLEEMRSGGQNINMNMMMSHHASAHHPNEHLVMAT-RNGRSV---- [134]Populus_tremula -------SPSRN--LLDDQIPNPTPN-FLI-NQT-HV----NDFSV-TV-RGMS------ [112]Malus_x_domestica_A -------STPHN--LLEEIRNTPSNT-ITN-HQPNYT----NES?M-PI-RGGGF----- [126]Vitis_shuttleworthii_A -------SPSHN--ILEEMPNPSPN--FLI-NQTTA-----NDFSV-SA-RGGA------ [111]Citrus_sinensis_B --------TPQN--LLEEIRNVPAN--MLM-INPPNP----ADQSVMPV-RVV------- [113]Antirrhinum_majus_GRAM --------SPQN--LLEEIRNSPSN--LLM-NQP-NP----ND-SMMPV-RGL------- [111]Solanum_tuberosum_A -------SPSHS--LLEEISNATPN--FLM-NQS-NSA---HEFVQLPA-RPGF------ [117]Lycopersicon_esculentum_A --------SPHN--LLDEIRNNPPS--LLM-NQQINP----NE-SLMPV-RGGV------ [118]Chrysanthemum_x_morifolium --------SAQS--LMEEMRNTPAN--LFL-NQP-NP----ND-HFGPV-RV-------- [120]Aquilegia -------SPTHN--LLEEIPSPPPN--LLI-DQS-NP----NDHLMMPVIRGGT------ [114]ChloYAB324 -------SPSHN--LLEEIPSPHPS--LML-DQT-NQ----NE--AMALAKGA------- [110]ChloYAB329 -------TPSHN--LLEEIPSLPPS--LLV-DQP-NS----NH--SGISVRGT------- [110]Oryza_sativa_A -----SPTSPHG--LLDEVAFQTPS--LLM-EQAASASLS------SITGRSS-SSCASN [131]Oryza_sativa_B PANPP---HHHS--LLDEISTANSPTQLLL-EQHGLGGLM----ASAASCRNNNSPAAAA [187]Triticum_aestivum_B PGNP----PRHS--LLDEISSANPSLQ-LL-EQHGLGGLI-----AAAGGRNAAAPAP-- [179]Zea_mays_B T-------SPHG--LLDELALQQAPS-FLM-EQASAN-LS-----STMTGRSSNSSCASN [126]Nuphar_advena_A_ PPSLNLPVVRHD--PQEEITAAAS---MLL-EH----SIM-KDSAASSVSRVA------- [124]Amborella_trichopoda_A ------SPTPHN--LLDECSPPSS---LLL-DH----PLMTPSNTGSASTR--------- [101]YABBY5 AT--NYAVP-----------EYGSSSRSH-T-----KIPSR------------------- [86]Populus_euphratica AS--NHNSHDYRI-------DLGSSSKFN-N-----KISVR------------------- [93]Malus_x_domestica_B AQNYNYNSQNYRI-------DSGSSSRCT-K-----KNATR------------------- [93]Vitis_shuttleworthii_B AP--NYTSPDYRI-------DLGSSSKCN-T-----KMAMR------------------- [86]Gossypium_raimondii GA--SYATQDYRT-------DLGSSSKCN-T-----KFSMR------------------- [92]Antirrhinum_majus_PROL AP--NHASTEYKV-------DLGSSSRWN-N-----KMAVQ------------------- [93]Lycopersicon_esculentum_C -P--NYTAPEYRM-------DFGSSTKCNMN-----RMSMR------------------- [95]Solanum_tuberosum_B -P--NYTAPEYRM-------DFGSSTKCNMN-----RMSMR------------------- [94]YABBY2 -------------------KDCASSS----------RSTNNL----SEN----------- [95]Populus_balsamifera -------------------KNFGSSS----------KC-NKVT-A-SES----------- [91]Vitis_vinifera -------------------EDCGSSS----------KC-NKFS-A-FES----------- [92]Malus_x_domestica_C -------------------KNCGSSSSSSSSSS---KS-NKFGHA-FES----------- [102]Citrus_sinensis_C -------------------KDRGSSS----------KC-NKFSSA-FEI----------- [94]Antirrhinum_majus_C -------------------KDNGSSS----------KS-NRYAP--LQ------------ [88]Lycopersicon_esculentum_E -------------------RGYGSSSSST-------NSFHRFSP--IP------------ [96]Persea_americana -------------------KECGSSS----------KT-NKMS---DST----------- [89]Oryza_sativa_FIL1 --------------------NFGSYDIAS-------RN-QRTSTAMY------------- [91]Oryza_sativa_D --------------------NFGSYDIAS-------RN-QRTSTAMY------------- [91]Oryza_sativa_FIL2 --------------------EYGSSS----------RY-DRVPMM-FS------------ [97]Asparagus_officinalis --------------------DNGSSS----------RC-NRPSIM-LT------------ [90]Lycoris_longituba --------------------DCSSTS----------KY-NSTSVMTFS------------ [91]Nuphar_advena_ --------------------ECGSSSS---------RF-SMIPMM--G------------ [93]Amborella_trichopoda_B --------------------DSSSTSNNFNHDDINNKV--LIPNESI------------- [82]Amborella_trichopoda_C -------------------EDGSSSKSI--KDSE-------------------TIPS--- [92]Welwitschia_mirabilis --------------------QASTSNPPFKNTLNHNFNYHEN-----------AYNPNLY [104]

Cycas_rumphii_ --------------------DSASSSTCS---------------EAAMI----SYA---- [93]Pinus_taeda --------------------DPASSST--------------------VIRD-ASYL---- [93]Picea_sitchensis --------------------DPASSST--------------------AIND--AYP---- [92]
[ 250 260 270 280 290 300][ . . . . . .] dCRC-YBINO -----------------E--DVSRVYQVVNK-PPEKRQRAPSAYNCFIKEEIRRLKAQNP [152]Antirrhinum_majus_B -----------------E--DSLHLNQLVHK-PPEKKQRAPSAYNHFIKKEIKRLKIEYP [147]Oryza_sativa_C -----------------DDAPAPAAAAVANK-PPERKQRTPSAYNCFVKEEIKRIESMEP [142]Nymphaea_alba -----------------E--EERRPAFTVNK-PPEKRHRAPSAYNRFIKEEIQRLKTSEP [131]CRC -----------------DQ-PPSPSPPFVVK-PPEKKQRLPSAYNRFMRDEIQRIKSANP [138]Lepidium_africanum_CRC -----------------DQ-PPSPTPPFVVK-PPEKKQRLPSAYNRFMRDEIQRIKTANP [138]Capparis_flexuosa_CRC -----------------DQ-PSSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [143]Cleome_sparsifolia_CRC -----------------DQ-PPFPTAAFVVK-PPEKKQRLPSAYNRFMREEIQRIKAANP [143]Gossypium_hirsutum_CRC2 -----------------E--PSSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [126]Gossypium_hirsutum_CRC1 -----------------E--PSSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [126]Citrus_sinensis_A -----------------E--PLSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [137]Antirrhinum_majus_A -----------------E--PLSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [130]Lycopersicon_esculentum_B -----------------E--PLSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKSENP [127]Nicotiana_tabacum_CRC1 -----------------E--PSSPKAPFVVK-PPEKKHRLPSAYNRFMKDEIQRIKAANP [125]Nicotiana_tabacum_CRC2 -----------------E--PSSPKAPFVVK-PPEKKHRLPSAYNRFMKDEIQRIKAANP [125]Petunia_hybrida_CRC -----------------E--PSSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [128]Hedyotis_centranthoides -----------------E--PLSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKAANP [134]Grevillea_robusta_CRC -----------------E--QLSPKAPFVVK-PPEKKHRLPSAYNRFMKEEIQRIKSAKP [135]Aquilegia_formosa_CRC -----------------E-----PN--YVVK-PPERKHRLPSAYNRYMKEEIQRIKSANP [129]Triticum_aestivum_A -----------------D---ASPRAPFVVK-PPEKKHRLPSAYNRFMREEIQRIKAAKP [131]Zea_mays_A -----------------E---LSPRMPFVVK-PPEKKHRLPSAYNRFMREEIQRIKAAKP [130]Oryza_sativa_DL -----------------E---GSPRAPFVVK-PPEKKHRLPSAYNRFMREEIQRIKAAKP [128]Amborella_CRC -----------------NQQ-VVPKVPNVVK-PPEKKHRLPSAYNRFMKEEIKRIKAGNP [127]FIL ----DLHQQH-------E--IPK--APPVNR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [164]YABBY3 -----DHLQ--------E--MPR--PPPANR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [176]Populus_tremula -----DH----------E--LPR--PPVISR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [152]Malus_x_domestica_A -----DHHQ--------E--IPK-PPAVVNR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [169]Vitis_shuttleworthii_A -----D-----------E--LPR--PPVINR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [150]Citrus_sinensis_B -----DH-Q--------E--IPK--SPVVNR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [154]Antirrhinum_majus_GRAM -----D-----------E--LPK--PPVANR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [150]Solanum_tuberosum_A -----D-----------D--LPR-PPPVINR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [157]Lycopersicon_esculentum_A -----D-----------E--IPK--PPVANR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [157]Chrysanthemum_x_morifolium -----D-----------E--LPK--PPVANR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [159]Aquilegia -----D-----------E--LPR--QPAANR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [153]ChloYAB324 -----D-----------E--VPR--NPVVNR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [149]ChloYAB329 -----E-----------E--NPK--TPVINR-RQEKRQRVPSAYNQFIKDEIQRIKAVNP [149]Oryza_sativa_A APAMQ-MPPAKPVQQEPE--LPKNAPASANR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [187]Oryza_sativa_B APPPPTSQGKAAAK---E-PSPRTNTAVINR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [242]Triticum_aestivum_B LPPPP-VAGGKGGK---E-PSPRTNP-VVNR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [232]Zea_mays_B LPPPAPMPAAQPVQQEAE--LPKTAPS-VNR-PPEKRQRVPSAYNRFIKDEIQRIKAGNP [182]Nuphar_advena_A_ -----------------DHTTTR-TTT-VNR-PPEKRQRVPSAYYRFIKDEIQRIKAGNP [164]Amborella_trichopoda_A ---LQEN----------E--ALH-SP--VSR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [142]YABBY5 -IST--RTI--------------TEQRIVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [128]Populus_euphratica -TPAANI-V--------------TQERVVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [136]Malus_x_domestica_B -DPTSDH-L--------------TEERGVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [136]Vitis_shuttleworthii_B -APASNI----------------SEERIVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [128]Gossypium_raimondii -APS--KNA--------------TEERVVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [134]Antirrhinum_majus_PROL --PSITK----------------PEQRIVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [134]Lycopersicon_esculentum_C -TP-ITNNV--------------HEERIVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [138]Solanum_tuberosum_B -TP-ITNNV--------------HEERIVNR-PPEKRQRVPSAYNQFIKEEIQRIKANNP [137]YABBY2 ----------------ID--R-EAPRMPPIR-PPEKRQRVPSAYNRFIKEEIQRIKACNP [135]Populus_balsamifera ----------------TE--H-EPPRMPAIR-PPEKRQRVPSAYNRFIKEEIQRIKASNP [131]Vitis_vinifera ----------------AE--H-EQPRMPPIR-PPEKRQRVPSAYNRFIKEEIQRIKASNP [132]Malus_x_domestica_C ----------------LEL-H-DQPRISPIR-PPEKRQRVPSAYNRFIKEEIQRIKASNP [143]Citrus_sinensis_C ----------------AE--H-ETPRMPPIR-PPEKRQRVPSAYNRFIKEEIQRIKASNP [134]Antirrhinum_majus_C ----------------AE--H-EQPKMPPIR-PPEKRQRVPSAYNRFIKEEIQRIKAGNP [128]Lycopersicon_esculentum_E ----------------TD--H-DQPRSPPIR-VPEKRQRVPSAYNRFIKEEIQRIKASNP [136]Persea_americana ----------------ED----DQPRILPIR-PPEKRQRVPSAYNKFIKEEIQRIKASNP [128]Oryza_sativa_FIL1 --PM----------------PTSQQQVSPIR-PPEKRQRVPSAYNRFIKEEIQRIKTSNP [132]Oryza_sativa_D --PM----------------PTSQQQVSPIR-PPEKRQRVPSAYNRFIKEEIQRIKTSNP [132]Oryza_sativa_FIL2 --------------------KNDTEHMLHVR-PPEKRQRVPSAYNRFIKEEIRRIKANNP [136]Asparagus_officinalis ----------------QE--H-DQQQLLPIR-PPEKRQRVPSAYNRCIKEEIQRIKANNP [130]Lycoris_longituba ----------------QQ--H-DQQKLLPIH-TPEKRQRVPSAYNRFIKEEIQRIKANNP [131]Nuphar_advena_ ----------------PE--KNE-QRMPPNR-PPEKRQRVPSAYNRFIKEEIQRIKACDP [133]Amborella_trichopoda_B ----------------AE--KEE--RTVVNR-PPEKRQRVPSAYNRFIKEEIQRIKASNP [121]Amborella_trichopoda_C ----ENEEP----------------RTIPNR-PPEKRQRVPSAYNRFIKEEIQRIKARNP [131]Welwitschia_mirabilis NG--KQETP----------------RTDVPR-APEKKQRISSAYNRFIKEEIQRIKAGNP [145]

Cycas_rumphii_ ----ENEEA----------------RTLSTRHVPEKRQRVPSAYNRFIKEEIQRIKACHP [133]Pinus_taeda ----ENEEA----------------RMSSYR-QPEKKQRVPSAYNRFIRDEIQRIKAHNP [132]Picea_sitchensis ----DNEET----------------RIPSYR-QPEKKQRVPSAYNRFIRDEIQRIKANNP [131]
[ 310 320 330 340 350 360][ . . . . . .]
INO SMAHKEAFSLAAKNWAHFPPAHNKRAASDQCFCEEDN------NAILPCNVFEDHEESNN [206]Antirrhinum_majus_B NMTHKQAFSAAAKNWAHNPQSQYKRG--EVQG-RGEV---GRTMHDVDEEVPCS---DSS [198]Oryza_sativa_C NITHKQAFSTAAKNWAHLPRIQQKRGR-DSC---*TTPAKQGKRN*WAR-VPGSSELLGR [197]Nymphaea_alba NISHREAFSTAAKNWAHMPRIQ-HKPDAESGSQRQSN--------KGKDKHVDREDKEGN [182]CRC EIPHREAFSAAAKNWAKYIPNSPT--SITSGGHNM-----IHGLGF--GEKK-------- [181]Lepidium_africanum_CRC EIPHREAFSAAAKNWAKYIPNSPT--SLTSGGNHM----MNVSYTNNPSEKARLYF*--- [189]Capparis_flexuosa_CRC EIPHREAFSAAAKNWARYIPNSPPG-SISAGSSSI------NGFDF--GEMK*------- [187]Cleome_sparsifolia_CRC EIPHREAFSAAAKNWAKYIPNSPT--SISTGGNAI------TGLGL--GAMK*------- [186]Gossypium_hirsutum_CRC2 EIPHREAFSAAAKNWARYIPNSPAASSVCGSSSNG------FYENF--AGTK*------- [171]Gossypium_hirsutum_CRC1 EIPHREAFSAAAKNWARYIPNSPAASSVCGSSSNE------QNDNV*------------- [167]Citrus_sinensis_A EIPHREAFSTAAKNWARYIPNSLAGSTSGSSSHE*------------------------- [172]Antirrhinum_majus_A EIPHREAFSAAAKNWARYIPNTPPPVPVTTSNHNI*------------------------ [166]Lycopersicon_esculentum_B EIPHREAFSAAAKNWARYLPNPPNS---GNT-NNV*------------------------ [159]Nicotiana_tabacum_CRC1 EIPHREAFSAAAKNWARYIPNTPNG-TLAESSNNA*------------------------ [160]Nicotiana_tabacum_CRC2 EIPHREAFSAAAKNWARYIPNTPNG-TLAESSNNA*------------------------ [160]Petunia_hybrida_CRC EIPHREAFSAAAKNWARYIPNTPNG-PLSESRNNA*------------------------ [163]Hedyotis_centranthoides EIPHREAFSAAAKNWARYIPH--NGPAGSITESSNTNNM*-------------------- [172]Grevillea_robusta_CRC EIPHREAFSTAAKNWARYVPSSGSVPENNKNERRTAAQES-------------------- [175]Aquilegia_formosa_CRC EIPHREAFSSAAKNWAKYVPHSQ-AGTVSGGKK--N----E-R-V--PAKESLDGA**-- [176]Triticum_aestivum_A DIPHREAFSMAAKNWAKCDPRCS--STVSAS----NSAP-EPR-IIVPGPQL--QERATE [181]Zea_mays_A DIPHREAFSMAAKNWAKCDPRCSTAASTETS----NSAPAEPR-VV-PTPQLTEPRFDLE [184]Oryza_sativa_DL DIPHREAFSMAAKNWAKCDPRCS--STVSTS----NSNP-EPR-VV-AAP-IPHQERANE [178]Amborella_CRC EIPHREAFSMAAKNWARFDPQLLHGSTTSTQIEKQVKPNQEIHEMVTAGGRVK-QE---- [182]FIL DISHREAFSAAAKNWAHFPHIHFGLVP-DNQPV--KKT-NMPQQEG---EDN-MVMKEGF [216]YABBY3 DISHREAFSAAAKNWAHFPHIHFGLMA-DHPPT--KKA-NVRQQEG---EDG-MMGREGF [228]Populus_tremula DISHREAFSAAAKNWAHFPHIHFGLMP-D-QTM--KKN-NVRHQEG---EDV-LIN-DAF [202]Malus_x_domestica_A DISHREAFSAAAKNWAHFPHIHFGLLP-D-QPV--KKT-NMRQQEG---DDL-LVK-DGF [219]Vitis_shuttleworthii_A DITHREAFSAAAKNWAHFPHIHFGLMP-D-QTM--KKT-NVRQQE--------------- [190]Citrus_sinensis_B DISHREAFSAAAKNWAHFPHIHFGLMPSD-QPV--KKT-SVRQQQEG--ED--AMIKDGF [206]Antirrhinum_majus_GRAM DISHREAFSAAAKNWAHFPHIHFGLMP-D-QPV--KKP-NVCQ-QDG--DD--VLMKDGF [200]Solanum_tuberosum_A DISHREAFSAAAKNWAHFPHIQFGLMP-D-QTV--KRT-NVRQ-QDG--ED--VLMKDGL [207]Lycopersicon_esculentum_A DISHREAFSAAAKNWAHFPHIHFGLMP-D-QPV--KKP-NACQ-QER--ED--VLMKEGF [207]Chrysanthemum_x_morifolium DISHREAFSAAAKNWAHFPHIHFGLMP-D-QPV--KKP-NVCQ-QDG--ED--LLMKDGF [209]Aquilegia DITHREAFSAAAKNWAHFPHIHFGLMP-D-QITAAKKA-NVRQ-QEG-DHD--VLMKDGI [206]ChloYAB324 DITHREAFSAAAKNWAHFPHIHFGLMP-E-QNV--KKT-SVRQ-QEG--EE--ILIKDGF [199]ChloYAB329 DISHRDAFSAAAKNWAHLPHIHFGLMR-E-QSM--NRN-NVRQ-QVM--EGEDVLTKNGL [201]Oryza_sativa_A DISHREAFSAAAKNWAHFPHIHFGLMP-D-QGF--KKT-FK-P-QDGS-ED--ILLKDSL [237]Oryza_sativa_B DISHREAFSAAAKNWAHFPHIHFGLMP-DHQGL--KKT-SLLP-QDHQRKD--GLLKEGL [295]Triticum_aestivum_B DISHREAFSAAAKNWAHFPHIHFGLMP-DHQGL--RKT-SLLP-QDHQRKDGHGLLKEGL [287]Zea_mays_B DITHREAFSAAAKNWAHFPHIHFGLMP-D-QGL--KKT-FKTH-QDGA-ED--MLLKDDL [233]Nuphar_advena_A_ DITHREAFSTAAKNWAHFPHIHFGLMA-D-QNM--KKS-NLR--QQEG-DD--LIISDGF [214]Amborella_trichopoda_A DITHREAFSTAAKNWAHFPHIHFGLMA-D-QSI--KKT-NM---QDG--DD--VLITESF [190]YABBY5 DISHREAFSTAAKNWAHFPHIHFGLML-ESNKQA-K---IA------------------- [164]Populus_euphratica DISHREAFSTAAKNWAHFPHIHFGLML-GTNSQT-K---VDDGSEKHLMSRSALP----- [186]Malus_x_domestica_B DISHREAFSTAAKNWAHFPHIHFGLML-EANNQA-K---LDNDPEKHLMSRAALLNN*-- [189]Vitis_shuttleworthii_B DISHREAFSTAAKNWAHFPHIHFGLML-ETNNQP-K---LDEGSEKHLMSKAALRNK*-- [181]Gossypium_raimondii DISHREAFSTAAKNWAHFPHIHFGLML-E-N-Q--K-------SN*-------------- [168]Antirrhinum_majus_PROL EISHREAFSTAAKNWAHFPHIHFGLML-ETNTQA-K--VLNEGSEKHRSHAKGYFI*--- [187]Lycopersicon_esculentum_C DISHREAFSTAAKNWAHFPHIHFGLML-ESNNQA-K---LACGS-RKL*----------- [181]Solanum_tuberosum_B DISHREAFSTAAKNWAHFPHIHFGLML-ESNNQA-K---LACGS-RKL*----------- [180]YABBY2 EISHREAFSTAAKNWAHFPHIHFGLKL-DGNKKG-K--QL-DQ--SVAG----------- [177]Populus_balsamifera DISHREAFSTAAKNWAHFPHIHFGLKL-DSNKHA-K---L-DHQ-SFAG--------EGT [176]Vitis_vinifera DITHREAFSTAAKNWAHFPHIHFGLKL-DGNKQG-K---L-DQ--AFAG--------EGP [176]Malus_x_domestica_C DISHREAFSTAAKNWAHFPHIHFGLKL-DANREP-K---L-DQ--AFC------------ [183]Citrus_sinensis_C DISHREAFSTAAKNWAHFPHIHFGLKL-DGNKQA-K---L-DKAAAYA---------EGT [179]Antirrhinum_majus_C EISHREAFSTAAKNWAHFPHIHFGLKL-DSNKQA-K---LQDH--SVAGG-------SGT [174]Lycopersicon_esculentum_E DISHREAFSTAAKNWAHFPHIHFGLKL-DGNKQTNK---L-DH--AVAG--------EGP [181]Persea_americana DISHREAFSTAAKNWAHFPHIHFGLTV-DGNKQV-K---V-DE--PFAG--------EGA [172]Oryza_sativa_FIL1 EISHREAFSAAAKNWAHLPRLHFGLNVADGGGGG--------GSN*-------------- [170]Oryza_sativa_D EISHREAFSAAAKNWAHLPRLHFGLSVADGGGGG--------GSN*-------------- [170]Oryza_sativa_FIL2 DISHREAFSTAAKNWAHFPNIHFGLGSHESSK---K---L-DE--AIGAPSP-------- [179]Asparagus_officinalis DISHREAFRAAAKNWAHFPHIHFGLSL-DENKQS-K---L-DD--AIATAHGVG------ [176]Lycoris_longituba DISHREAFSAAAKNWAHFPH---------------------------------------- [151]Nuphar_advena_ DISHKEAFSTAAKNWAHFPHIQFGLAL-EGNKQQNL---M-DL---F*------------ [173]Amborella_trichopoda_B DISHREAFSTAAKNWAHFPHIHFGLML-DTTKKQPK---L-DQ-----GS-GKHLM---- [166]Amborella_trichopoda_C EITHREAFSTAAKNWAHFPHLHYGLSL-E-RNNQVT---L-D------EVLV-------- [171]Welwitschia_mirabilis NMSHREAFSTAAKNWALFPYMQLGLVF-D-EKKQES*----------------------- [180]

Cycas_rumphii_ EITHREAFSTAAKNWAHFPHIHFGLML-E-NHKQAK---I-ADEDGDHDVLNG-AEA--- [183]Pinus_taeda KITHKEAFSAAAKNWAHYPHIHFGMMM-D-SRRQAN-----SEDGGD--VLN--AE---- [177]Picea_sitchensis KITHKEAFSAAAKNWAHYPHIHFGLML-D-NRRQSN-----SEDGGD--VLN--AE---- [176]
[ 370 380 390 400 ][ . . . . ]
INO GFRERKAQRHSIWGKSPFE------------------------- [225]Antirrhinum_majus_B FPKQ--RHSINLFAPVKLYAESGYFSFYGLLKLLYGLNT*---- [236]Oryza_sativa_C FRKNPSCSCPDNLVAHLKHVLPEFFSFM**IQQVFFFDYSVLNQ [241]Nymphaea_alba QIFQQRKVSRQCFLTKVPQQ*----------------------- [203]CRC -------------------------------------------- [181]Lepidium_africanum_CRC -------------------------------------------- [189]Capparis_flexuosa_CRC -------------------------------------------- [187]Cleome_sparsifolia_CRC -------------------------------------------- [186]Gossypium_hirsutum_CRC2 -------------------------------------------- [171]Gossypium_hirsutum_CRC1 -------------------------------------------- [167]Citrus_sinensis_A -------------------------------------------- [172]Antirrhinum_majus_A -------------------------------------------- [166]Lycopersicon_esculentum_B -------------------------------------------- [159]Nicotiana_tabacum_CRC1 -------------------------------------------- [160]Nicotiana_tabacum_CRC2 -------------------------------------------- [160]Petunia_hybrida_CRC -------------------------------------------- [163]Hedyotis_centranthoides -------------------------------------------- [172]Grevillea_robusta_CRC -------------------------------------------- [175]Aquilegia_formosa_CRC -------------------------------------------- [176]Triticum_aestivum_A -----QVVESFDIFKQMERSA*---------------------- [198]Zea_mays_A DRAKGQVIESFDIFKHIERSI*---------------------- [206]Oryza_sativa_DL -----QVVESFDIFKQMERSG*---------------------- [195]Amborella_CRC ------DMRQLQAAARSQIT*----------------------- [197]FIL YAPAAA------NVGV-TPY------------------------ [229]YABBY3 YGSAA-------NVGVAHN------------------------- [240]Populus_tremula FSSS--------NVGVTTPY*----------------------- [215]Malus_x_domestica_A FSTPA-------NVGDS-PY*----------------------- [232]Vitis_shuttleworthii_A -------------------------------------------- [190]Citrus_sinensis_B FAPAA-------NVGVT-PY*----------------------- [219]Antirrhinum_majus_GRAM LAPA--------NVGVS-PY*----------------------- [212]Solanum_tuberosum_A FNTSA-------NVSV*--------------------------- [217]Lycopersicon_esculentum_A LAPA--------NIGVS-PY*----------------------- [219]Chrysanthemum_x_morifolium LATA--------NVGVS-PY*----------------------- [221]Aquilegia FAAA--------NLGVS-PY*----------------------- [218]ChloYAB324 FTTA--------NIGVT-PY*----------------------- [211]ChloYAB329 LSTK---------------------------------------- [205]Oryza_sativa_A YAAAAAAAAAAANMGVT-PF*----------------------- [257]Oryza_sativa_B YAAAAAAAAAA-NMGVA-PY*----------------------- [314]Triticum_aestivum_B YAA---------NMGIA-PY*----------------------- [298]Zea_mays_B YAAAAAAAAA--NMGIT-PF*----------------------- [251]Nuphar_advena_A_ FTTA--------NMGVT-PY*----------------------- [226]Amborella_trichopoda_A FTAA--------NMGVT-PY*----------------------- [202]YABBY5 -------------------------------------------- [164]Populus_euphratica -------------------------------------------- [186]Malus_x_domestica_B -------------------------------------------- [189]Vitis_shuttleworthii_B -------------------------------------------- [181]Gossypium_raimondii -------------------------------------------- [168]Antirrhinum_majus_PROL -------------------------------------------- [187]Lycopersicon_esculentum_C -------------------------------------------- [181]Solanum_tuberosum_B -------------------------------------------- [180]YABBY2 ------------QKSNGYY------------------------- [184]Populus_balsamifera ------------QKTSGLYEINGSL*------------------ [190]Vitis_vinifera ------------HKAHGFY*------------------------ [184]Malus_x_domestica_C ------------RRGNSKV*------------------------ [191]Citrus_sinensis_C ------------QKSNGFY*------------------------ [187]Antirrhinum_majus_C TMAHA-------QKSLGFY------------------------- [186]Lycopersicon_esculentum_E ------------QKTIGLY*------------------------ [189]Persea_americana ------------QKTQGFY*------------------------ [180]Oryza_sativa_FIL1 -------------------------------------------- [170]Oryza_sativa_D -------------------------------------------- [170]Oryza_sativa_FIL2 ------------QKVQRLY*------------------------ [187]Asparagus_officinalis ------------QKSQVLY*------------------------ [184]Lycoris_longituba -------------------------------------------- [151]Nuphar_advena_ -------------------------------------------- [173]Amborella_trichopoda_B ------------NQAAVCIE*----------------------- [175]Amborella_trichopoda_C ----NEGSQSDPIIHQAV*------------------------- [186]Welwitschia_mirabilis -------------------------------------------- [180]

Cycas_rumphii_ VCNK*--------------------------------------- [188]Pinus_taeda VCNN*--------------------------------------- [182]Picea_sitchensis VCNN*--------------------------------------- [181]

Table S1. GenBank accession information on the YABBY genes that were used in ourphylogentic analysis
Species and/or gene Gene lineage GenBank accession number
Amborella trichopoda A FIL/YAB3 CV002899Amborella trichopoda B – CV005012Amborella trichopoda C YAB2 AB126654Amborella trichopoda CRC CRC AJ877257Antirrhinum majus A CRC AJ559642Antirrhinum majus B INO AY451400Antirrhinum majus C YAB2 AJ808095Antirrhinum majus GRAM FIL/YAB3 AJ804783Antirrhinum majus PROL YAB5 AJ559762Aquilegia formosa CRC CRC AY854797 (this study)Aquilegia sp. FIL/YAB3 –Asparagus officinalis YAB2 CV459037Capparis flexuosa CRC CRC AY854798 (this study)ChloYAB324 FIL/YAB3 –ChloYAB329 FIL/YAB3 –Chrysanthemum � morifolium FIL/YAB3 AY173066Citrus sinensis A CRC CN185168Citrus sinensis B FIL/YAB3 CK935155Citrus sinensis C YAB2 CK933417Cleome sparsifolia CRC CRC AY854803 (this study)CRC (Arabidopsis thaliana) CRC AF132606Cycas rumphii – CB093758FIL (Arabidopsis thaliana) FIL/YAB3 AF136538Gossypium hirsutum CRC1 CRC AY854804 (this study)Gossypium hirsutum CRC2 CRC AY854805 (this study)Gossypium raimondii YAB5 CO100015Grevillea robusta CRC CRC (this study)Hedyotis centranthoides CRC CB088054INO (Arabidopsis thaliana) INO AF195047Lepidium africanum CRC CRC AY854802 (this study)Lycopersicon esculentum A FIL/YAB3 AW623191 + BE354691Lycopersicon esculentum B CRC AI771643 + AI483816Lycopersicon esculentum C YAB5 AI484101 + BW684955Lycopersicon esculentum D YAB2 BW69229Lycoris longitu YAB2 CN453541Malus � domestica A FIL/YAB3 CO576666 + CO540988 + CN996971Malus � domestica B YAB5 CN580139 + CO051593Malus � domestica C YAB2 CV631928Nicotiana tabacum CRC1 CRC AY854799 (this study)Nicotiana tabacum CRC2 CRC AY854800 (this study)Nuphar advena A FIL/YAB3 CD474192 + CV001750 + CD474498Nuphar advena B YAB2 CD472928Nymphaea alba INO AB092980Oryza sativa A FIL/YAB3 XM467005Oryza sativa B FIL/YAB3 NM197274Oryza sativa C INO (from genomic sequence)Oryza sativa D YAB2 TC264221Oryza sativa DL CRC AB106553Oryza sativa FIL1 YAB2 AF098752Oryza sativa FIL2 YAB2 AU094964Persea americana YAB2 CV004856 + CV000415Petunia hybrida CRC CRC AY854801 (this study)Pinus sitchensis – DR517093Pinus taeda – DR100835Populus balsamifera YAB2 BU871780Populus euphratica YAB5 AJ773836Populus tremula FIL/YAB3 BU830431Solanum tuberosum A FIL/YAB3 CK266773Solanum tuberosum B YAB5 CV506612Triticum aestivum A CRC AF545436Triticum aestivum B FIL/YAB3 AY330228Vitis shuttleworthii A FIL/YAB3 CV096830Vitis shuttleworthii B YAB5 CV098755Vitis vinifera YAB2 CF373869 + CF609595Welwitschia mirabilis – CV004246 + CV005289YABBY2 (Arabidopsis thaliana) YAB2 AF136539YABBY3 (Arabidopsis thaliana) FIL/YAB3 AF136540YABBY5 (Arabidopsis thaliana) YAB5 AK119091Zea mays A CRC AY104291Zea mays B FIL/YAB3 AY313902
Unannotated genes from a single species are distinguished by a letter, immediately following the species name.