reduced expression oftherenal calcium-sensing...
TRANSCRIPT

J Am Soc Nephrol 9: 2067-2074, 1998
Reduced Expression of the Renal Calcium-Sensing Receptor
in Rats with Experimental Chronic Renal Insufficiency
ROBERT S. MATHIAS,* HIEP T. NGUYEN,t MARTIN Y.H. ZHANG,* and
ANTHONY A. PORTALE*�Division of Pediatric Nephrology, Departments of *pediatrics, tPediatric Urology, and ‘�Medicine, tile
University of California San Francisco, San Francisco, California.
Abstract. Chronic renal insufficiency is associated with ebe-
vated serum parathyroid hormone (PTH) levels (2#{176}HPT), de-
ficiency of 1,25-dihydroxyvitamin D (l,25(OH)2D), and hy-
pocalciuria. In chronic renal insufficiency, the 2#{176}HPT may
result from reduced expression of the parathyroid gland extra-
cellular Ca2tsensing receptor (CaSR). Since the CaSR was
cloned from rat and human kidney, this study examined in rats
whether expression of the renal CaSR is altered in experimen-
tab chronic renal insufficiency. Four weeks after chronic renal
insufficiency was induced by 5/6 nephrectomy (Nx) in Sprague
Dawley rats, the serum creatinine concentration was 0.96 ±
0.06 mg/db compared with 0.35 ± 0.02 mg/dl in sham-operated
animals (P < 0.05). The serum total Ca2� and phosphorus
concentrations were not different. In the Nx group, the serum
concentration of aniino-PTH was higher (65 ± 8 pg/ml), and
the concentration of 1 ,25(OH)2D was significantly lower
(47 ± 5 pg/mb) compared with 45 ± 5 pg/mb and 6 1 ± 4 pg/mb
(P = 0.05) in the sham group, respectively. In a subset of rats
studied, the Nx group was hypocalciuric (1 .4 ± 0.5 mg/kg per
d) compared with the sham group (3.7 ± 0.5 mg/kg per d) (P <
0.05). In the Nx rats, CaSR mRNA expression and CaSR
protein levels were found to be reduced by 35 and 38%,
respectively, than those observed in controls. These results
suggest that reduced renal CaSR expression in chronic renal
insufficiency may play a role in disordered mineral ion ho-
meostasis, including hypocalciuria.
The G protein-coupled extraceblubar calcium (Ca2�)-sensing
receptor (CaSR), recently cloned from bovine parathyroid
gland (1) and rat and human kidney (2,3), provides a mecha-
nism by which changes in the extracellular Ca2� concentration
([Ca210) can regulate, either indirectly or directly, mineral ion
homeostasis. A small decrease in [Ca210 is sensed by the
parathyroid cell, which increases its secretion of parathyroid
hormone (PTH) (4). PTH acts both on the skeleton to increase
bone resorption, and on the kidney to increase the tubular
reabsorption of Ca2� and the synthesis of 1 ,25-dihydroxyvita-
mm D (l,25(OH)2D), thus promoting an increase in [Ca2�j0 to
normal levels. In the kidney, changes in [Ca2�]0 can directly
modulate the synthesis of l,25(OH)2D (5), the expression of
vitamin D-dependent calbindin-D2SK (6), and the tubular reab-
sorption of Ca2� in the thick ascending limb of the loop of
Henle (7,8). The finding that the CaSR has been localized to
segments of the proximal and distal nephron (9) provides a
mechanism by which a direct effect of [Ca2�]0 can modulate
renal tubule function and, thereby, mineral ion homeostasis, at
beast in part, via receptor-mediated events.
Normal expression of the CaSR can be essential in main-
taming mineral ion homeostasis. In patients with familial hy-
Received December 4, 1997. Accepted May 4, 1998.Correspondence to Dr. Robert S. Mathias, Children’s Renal Center, University
of California, San Francisco Medical Center. 533 Parnassus Avenue, RoomU-585, San Francisco, CA 94143-0748.
1046-6673/0901 l-2067$03.00/0
Journal of the American Society of Nephrology
Copyright © 1998 by the American Society of Nephrology
pocabciuric hypercalcemia (FHH) and neonatal hyperparathy-
roidism, inactivating mutations in the CaSR gene have been
identified (10). In such patients, mutations of the CaSR give
rise to disordered mineral-ion homeostasis, specifically, hyper-
parathyroidism, hypocalciuria, and hypercalcemia. In addition,
these symptoms can be induced in mice by knocking out the
CaSR gene (1 1).
With progressive chronic renal insufficiency, disturbances in
mineral ion homeostasis, including secondary hyperparathy-
roidism (2#{176}HPT) (12), play an important role in the pathogen-
esis of metabolic bone disease. Hypocalciuria also is observed
in patients with mild and moderate chronic renal insufficiency
(13,14). In chronic renal insufficiency, there is evidence that
these disturbances reflect, at least in part, an abnormality in the
function of the CaSR. Indeed, expression of the CaSR mRNA
and its protein were reduced in hyperplastic parathyroid glands
from hemodiabysis patients with severe chronic 2#{176}HPT when
compared with that in normal glands from patients with para-
thyroid adenomas (15,16).
In patients with FHH, both the hypocalciuria and the abnor-
mality in [Ca2�i0-regulated PTH secretion, specifically, the
increase in the Ca2� set-point, are thought to reflect abnormal
function of the CaSR (17). Accordingly, as occurs in patients
with FHH and in mice lacking the CaSR gene in which urinary
Ca2� excretion is greatly decreased, a reduction in expression
of the CaSR in the kidney might contribute to the hypocalciuria
observed in patients with chronic renal insufficiency (13,14).
To test this hypothesis, we determined whether renal expres-
sion of the CaSR and urinary calcium excretion were reduced
in the rat with moderate chronic renal insufficiency.

2068 Journal of the American Society of Nephrology J Am Soc Nephrol 9: 2067-2074, 1998
Materials and MethodsAnimals and Diets
Sprague Dawley rats weighing 200 to 250 g were fed normal rat
chow (Purina) throughout the study and were allowed water ad
libitum. The diet contained 0.93% calcium and 0.81% phosphorus.
Rats underwent either sham surgery or one-stage 5/6 nephrectomy
(Nx) ( 18, 19). Briefly, rats were anesthetized with Nembutal, and an
incision was made through the skin and body wall along the midline.
The left kidney was exposed, decapsulated, and in rats subjected to5/6 Nx, the upper and lower branches of renal arteries were ligated.
The right kidney was then exposed, decapsulated, and in rats sub-
jected to 5/6 Nx, the renal pedicle was ligated and the kidney was
removed. The incision was then closed. Four weeks after surgery,
urine was obtained for a 24-h period for the determination of creati-
nine (Cr) and total Ca2� (TCa2� ) concentration. At the time of
sacrifice, blood was obtained for determination of serum concentra-
tions of Cr, TCa2�, phosphorus (P), PTH, and l,25(OH),D, and the
kidneys were snap-frozen in liquid nitrogen. The procedures were
approved by the Committee on Animal Research of the University of
California, San Francisco.
Biochemical Determinations
Plasma (TCa2�, P, Cr) and urine (Ca2�, Cr) concentrations were
determined by an autoanalyzer (model 747; Boehringer Mannheim,
Indianapolis, IN). Serum immunoreactive N-terminal PTH (iPTH)
was determined in duplicate with a rat PTH (immunoradiometric
assay) kit (Nichols Institute Diagnostics, San Juan Capistrano, CA).
Serum levels of l,25(OH)2D were determined in duplicate by radio-
receptor assay (Nichols Institute Diagnostics).
Isolation of Total RNA
Total tissue RNA was extracted by homogenizing kidney tissue
with a glass tissue grinder in RNA STAT-60 (Tel-Test “B”, Inc.,
Friendswood, TX) as a modification of the guanidinium isothiocya-
nate method (20). The RNA concentration was determined by absor-
bance at 260 nm.
Ribonuclease Protection Analysis
The RNase protection assay was performed as described (Hyb-
Speed RPA assay kit, Ambion, Austin, TX). A l272-bp (PstI-SacI)
fragment of the rat kidney CaSR cDNA (NPS Pharmaceuticals, Salt
Lake City, UT) (2) was subcloned into pBlueScript SK+ immediately
downstream from the bacteriophage T3 promoter. The rat kidney
CaSR fragment consists of the coding region sequence starting at
transmembrane region 4 and extending beyond the C-terminal stop
codon (from exon 7 of the CaR gene). A 250-bp fragment of theglyceraldehyde phosphate dehydrogenase (GAPDH) eDNA was sub-
cloned into pT7Blue(R) immediately downstream from the T7 pro-
moter. The CaSR and OAPDH eDNA were linearized with ApaI
(Promega, Madison, WI) and HphI (New England Biolabs, Beverly,
MA), respectively. The radiolabeled antisense RNA probes weretranscribed separately from the linearized plasmids using T3 polymer-
ase and T7 polymerase, respectively (Maxiscript protocol, Ambion),
in the presence of [a-32P]-labeled UTP (New England Nuclear, Bos-
ton, MA). The predicted sizes of the protected fragments for the CaSR
and GAPDH were 336 bp and I 64 bp, respectively. The labeled
probes (2 p.1; 5 X b0� cpm) were placed together with sample total
RNA and hybridized for 20 mm at 68#{176}C.After hybridization, the
mixture was treated with ribonuclease at 37#{176}Cfor 30 mm to degrade
single-stranded RNA and unhybridized probe. The remaining pro-
tected RNA fragments were precipitated and dried. Each pellet was
dissolved in loading buffer (Ambion), denatured at 90#{176}C,and resolved
on a 5% acrylamide!8 M urea sequencing gel. The gel was placed on
x-ray film (Hyper-fllm MP, Amersham, Arlington Heights, IL) with
an intensifying screen and exposed at -80#{176}C for I to 5 d for
autoradiography. The intensity of the CaSR mRNA was visualized
and normalized to that of GAPDH mRNA using densitometric values
obtained by National Institutes of Health Image I .60 software (Be-
thesda, MD). Results are expressed as the ratio CaSR/GAPDH
mRNA. Total RNA from bovine parathyroid glands was kindly sup-
plied by Dr. Dolores Shoback (Veteran’s Administration Medical
Center, San Francisco, CA).
Protein Preparation
Crude membranes from kidney tissue were prepared by a modifi-
cation of the method by Chattopadhyay et al. (2 1). Briefly, 250 mg of
kidney tissue was homogenized in buffer: ice-cold 50 mM Tris-HC1,
pH 7.4, 0.30 M sucrose, 1 mM ethyleneglycol-bis-(�-aminoethylether)-N,N,N’,N’-tetra-acetic acid, ethylenediamine tetra-acetic acid,
and protease inhibitors (80 p�g/ml aprotinin, 30 p.g/ml leupeptin, I
mg/ml Pefabloc SC, 50 p.g!ml calpain inhibitor, 50 p.g/ml bestatin,
and 5 �tg/ml pepstatin). Nuclei and cell debris were removed from the
resultant homogenate by centrifugation (12,000 X g) for 20 mm at
4#{176}C.The supernatant was centrifuged at 42,000 X g for 20 mm at
4#{176}C,and the remaining pellet, representing a crude membrane prep-
aration, was solubilized with 1% Triton X-lOO in 100 p.1 of lysis
buffer and mixed with 2X Laemmli sample buffer containing 1 mM
dithiothreitol. Cultured human embryonic kidney-293 cells (HEK-
CaSR) stably expressing the human parathyroid Ca2�-sensing recep-
tor (NPS Pharmaceuticals) were grown to confluence and rinsed with
phosphate-buffered saline. Cells were scraped, then pelleted at 3500
rpm for 3 mm at 4#{176}C.Supernatant was removed, and lysis buffer
containing I X Hepes-buffered saline, pH 7.4, 1% Triton X-bOO, and
protease inhibitors (10 p.g/ml each ofaprotinin, leupeptin, and calpain
inhibitor) was added to the pellet. Protein concentrations were deter-
mined by the BCA protein reagent kit (Pierce, Rockford, IL).
Western Blot Analysis
Proteins obtained from HEK-CaSR cells and kidney from sham-
operated and 5!6 Nx rats were separated by 6% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (PAGE) (22) and trans-
ferred to nitrocellubose filters (Amersham) electrophoretically at 0.2
mV for 3 h in transfer buffer contain 3 mM Tris-base, 190 mM
glycine, 0.01% sodium dodecyl sulfate, and 20% methanol. The filters
were blocked with 5% milk containing 20 mM Tris-base, pH 7.6, 137
mM NaC1, and 0. 1 % Tween-20 (TBS-T) overnight, then washed 3
times in 0.1% milk and one time in 1% milk in TBS-T (5 mm each)
at room temperature. The filters were then incubated with protein
A-purified, anti-CaSR antibody 4641 (1:10,000 dilution) in 1% milk
containing TBS-T for 1 h at room temperature. This anti-CaSR
primary antibody was raised to a peptide corresponding to the de-
duced sequence of amino acids 2 15 to 237 (DDDYORPGIEKFREE-
AEERDIC) of the extracellular domain of the bovine parathyroidCaSR (1), which has a sequence identical to that of the rat kidney
CaSR (2). The filters were washed 3 times in 0. 1% milk and one time
in 1% milk in TBS-T (5 mm each) at room temperature. The second-
ary antibody (horseradish peroxidase-coupled anti-rabbit IgO; Amer-
sham) in 1% milk and TBS-T (1:10,000 dilution) was incubated with
the filters for 1 h at room temperature. In a similar sequence, the filters
were washed with milk-containing solutions as before. The specific
bands were visualized using enhanced chemiluminescence (ECL)
detection (Amersham) after exposing the filter to film (Hyperfilm-
J Am Soc Nephrol 9: 2067-2074. 1998 Reduced Renal CaSR in Renal Insufficiency 2069
ECL, Amersham). Results were expressed as arbitrary densitometric
units per milligram of protein added to each lane, using National
Institutes of Health Image 1 .60 software (Bethesda, MD). Crude
membrane protein from HEK-CaSR cells was used as a positive
control.
Statistical AnalysisAll data are expressed as means ± SEM. Statistical analysis was
performed by unpaired t test, using StatView for the Macintosh. A P
value of �0.05 was taken to indicate a statistically significant differ-
ence.
ResultsBiochemical Results
Four weeks after surgery, the body weights of the 5/6 Nx
animals were significantly lower than those of sham-operated
animals (P < 0.05) (Table 1). The serum Cr concentrations in
the Nx animals were higher than those in sham animals (P <
0.05). The serum concentrations of TCa2� and P were not
significantly different between the two groups. The concentra-
tions of N-terminal PTH in the Nx group were higher and of
l,25(OH)2D were significantly lower (P = 0.05) than those in
the sham-operated animals.
Because decreased urinary Ca2� excretion has been noted in
patients with mild-to-moderate chronic renal insufficiency
( 13, 14), we determined whether urinary Ca2� excretion was
decreased in experimental chronic renal insufficiency in the
rat. After 4 wk of surgery, nine animals from each group were
placed in metabolic cages. After a 48-h acclimation period,
urine was collected over a 24-h period for determination of
Ca2� and Cr (Table 2). Urinary excretion of Ca2� and the
creatinine clearance rate (Car) were significantly lower in the
Nx animals than in the sham-operated animals (P < 0.05). In
this subset of animals, the serum bevels of TCa2�, P, and iPTH
Table 1. Body weights and biochemical determinations
obtained at the time of sacrifice in sham-operated
and 5/6 Nx ratsa
Characteristic Sham 5/6 NxValue
n 15 19
Body weight (g)
presurgery 237 ± 3 236 ± 3 NS
at sacrifice 346 ± 4 316 ± 7 <0.05
Serum
Cr (mg/dl) 0.35 ± 0.02 0.96 ± 0.06 <0.05total Ca2� (mg/db) 9.9 ± 0.2 10.0 ± 0.1 NS
P (mg/dl) 9.2 ± 0.2 9.7 ± 0.2 NS
iPTH (pg/mI) 45 ± 5b 65 ± 8C 0.06
l,25(OH)2D (pg/ml) 61 ± 4b 0.05
a Results are expressed as means ± SEM. Cr, creatinine; P,
phosphorus; iPTH, immunoreactive N-terminal parathyroid
hormone; l,25(OH),D, 1,25-dihydroxyvitamin D.C, Represents 14 animals.
C Represents 16 animals.
Table 2. Biochemical determinations obtained at the time of
sacrifice in sham-operated and 5/6 Nx ratsa
..
Charactenstic Sham 5/6 NxP
Value
n 9 9
Serum
total Ca2� (mg/db) 9.9 ± 0.1 10.2 ± 0.1 <0.05
P (mg/dl) 9.3 ± 0.2 9.7 ± 0.2 NS
iPTH (pg/mb) 53 ± 5 72 ± 6 <0.05
l,25(OH)2D (pg/ml) 46 ± 3 33 ± 4 <0.05
Urine’�
C�r (mb/mm per kg) 10.2 ± 0.6 1.9 ± 0.4 <0.05
24-h urine Ca2� (mg/kg) 3.7 ± 0.5 1 .4 ± 0.5 <0.05
a All values are the mean ± SEM. C�r creatinine clearance rate.
Other abbreviations as in Table I.
b Calculated using body weight at time of sacrifice.
(P < 0.05) were higher, and those of 1 ,25(OH)2D (P < 0.05)
were lower in the Nx group than in the sham-operated group.
Ribonuclease Protection Analysis
To determine CaSR mRNA expression in rat kidney and in
bovine parathyroid tissue, RNase protection assay was per-
formed using total RNA prepared from normal kidney and
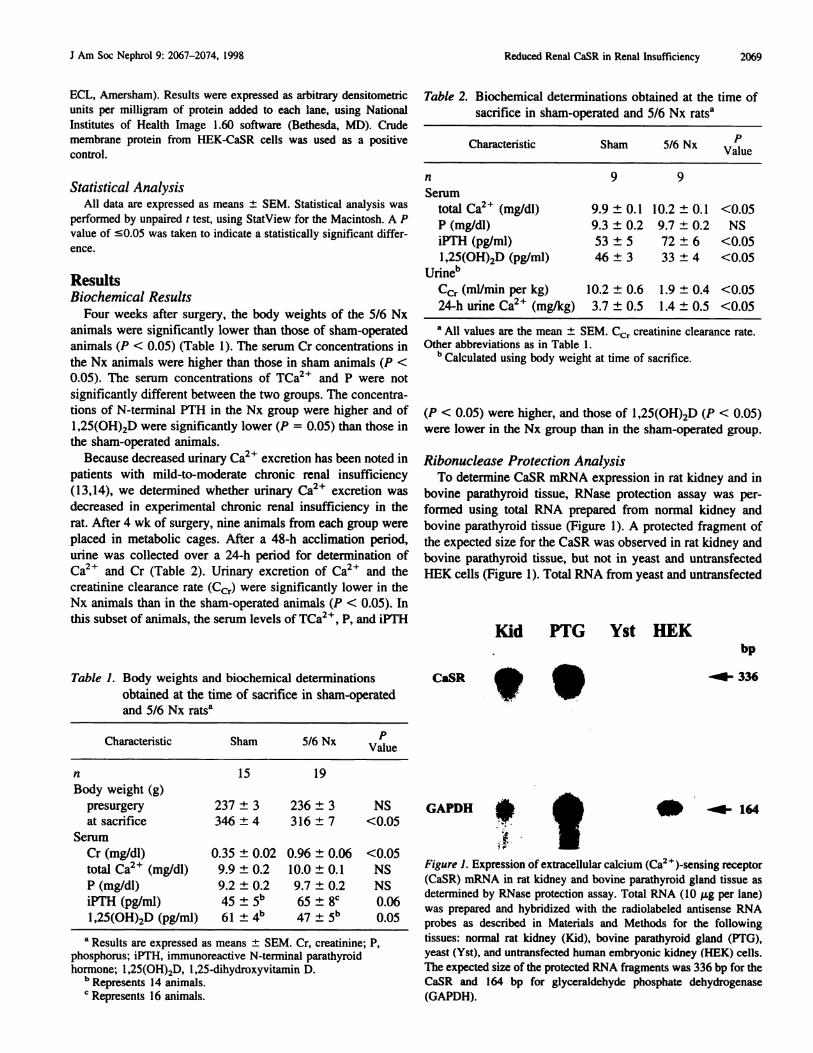
bovine parathyroid tissue (Figure 1). A protected fragment of
the expected size for the CaSR was observed in rat kidney and
bovine parathyroid tissue, but not in yeast and untransfected
HEK cells (Figure 1). Total RNA from yeast and untransfected
Kid PTG Yst HEK
CaSR �
bp
�- 336
GAPDH � � ..l�- 164
Figure 1. Expression of extracellular calcium (Ca2 � )-sensing receptor
(CaSR) mRNA in rat kidney and bovine parathyroid gland tissue asdetermined by RNase protection assay. Total RNA (10 p.g per lane)
was prepared and hybridized with the radiolabeled antisense RNA
probes as described in Materials and Methods for the following
tissues: normal rat kidney (Kid), bovine parathyroid gland (PTO),
yeast (Yst), and untransfected human embryonic kidney (HEK) cells.
The expected size of the protected RNA fragments was 336 bp for theCaSR and I 64 bp for glyceraldehyde phosphate dehydrogenase
(OAPDH).
.� 1
C-.
� 1
�
� 0.5
C-.) 0.25
p<0.05
2070 Journal of the American Society of Nephrology J Am Soc Nephrol 9: 2067-2074, 1998
Nx
,,,,‘ ,
Sham
CaSR
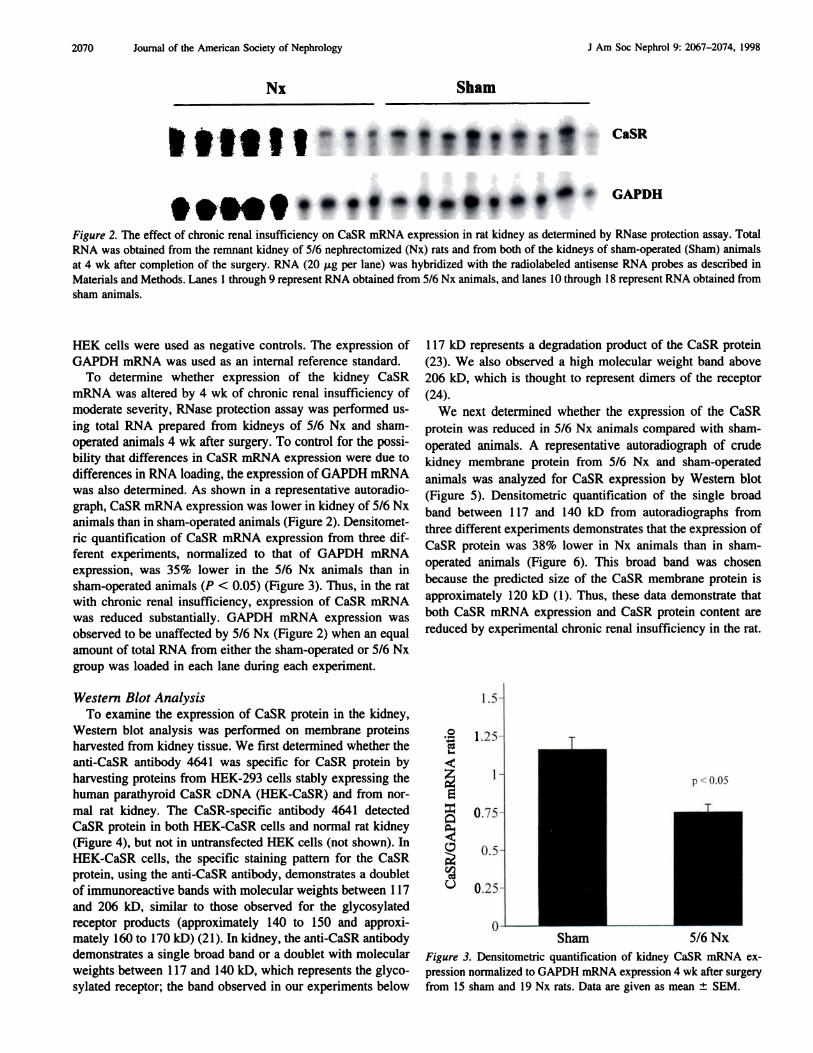
GAPDH
Figure 2. The effect of chronic renal insufficiency on CaSR mRNA expression in rat kidney as determined by RNase protection assay. Total
RNA was obtained from the remnant kidney of 5/6 nephrectomized (Nx) rats and from both of the kidneys of sham-operated (Sham) animals
at 4 wk after completion of the surgery. RNA (20 p.g per lane) was hybridized with the radiolabeled antisense RNA probes as described in
Materials and Methods. Lanes 1 through 9 represent RNA obtained from 5/6 Nx animals, and lanes 10 through I 8 represent RNA obtained from
sham animals.
HEK cells were used as negative controls. The expression of
GAPDH mRNA was used as an internal reference standard.
To determine whether expression of the kidney CaSR
mRNA was altered by 4 wk of chronic renal insufficiency of
moderate severity, RNase protection assay was performed us-
ing total RNA prepared from kidneys of 5/6 Nx and sham-
operated animals 4 wk after surgery. To control for the possi-
bility that differences in CaSR mRNA expression were due to
differences in RNA loading, the expression of GAPDH mRNA
was also determined. As shown in a representative autoradio-
graph, CaSR mRNA expression was lower in kidney of 5/6 Nx
animals than in sham-operated animals (Figure 2). Densitomet-
nc quantification of CaSR mRNA expression from three dif-
ferent experiments, normalized to that of GAPDH mRNA
expression, was 35% lower in the 5/6 Nx animals than in
sham-operated animals (P < 0.05) (Figure 3). Thus, in the rat
with chronic renal insufficiency, expression of CaSR mRNA
was reduced substantially. GAPDH mRNA expression was
observed to be unaffected by 5/6 Nx (Figure 2) when an equal
amount of total RNA from either the sham-operated or 5/6 Nx
group was loaded in each lane during each experiment.
Western Blot Analysis
To examine the expression of CaSR protein in the kidney,
Western blot analysis was performed on membrane proteins
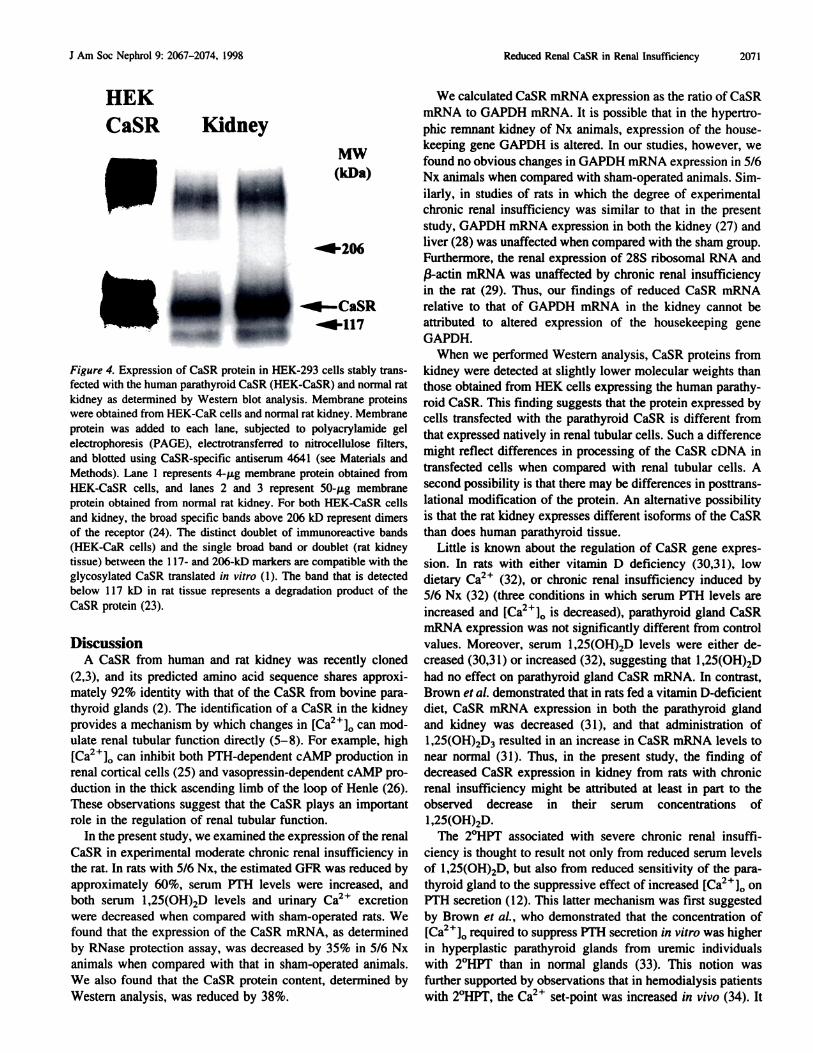
harvested from kidney tissue. We first determined whether the
anti-CaSR antibody 4641 was specific for CaSR protein by
harvesting proteins from HEK-293 cells stably expressing the
human parathyroid CaSR cDNA (HEK-CaSR) and from nor-
mal rat kidney. The CaSR-specific antibody 4641 detected
CaSR protein in both HEK-CaSR cells and normal rat kidney
(Figure 4), but not in untransfected HEK cells (not shown). In
HEK-CaSR cells, the specific staining pattern for the CaSR
protein, using the anti-CaSR antibody, demonstrates a doublet
of immunoreactive bands with molecular weights between 1 17
and 206 kD, similar to those observed for the glycosylated
receptor products (approximately 140 to 150 and approxi-
mately 160 to 170 kD) (2 1 ). In kidney, the anti-CaSR antibody
demonstrates a single broad band or a doublet with molecular
weights between 1 17 and 140 kD, which represents the glyco-
sylated receptor; the band observed in our experiments below
1 17 kD represents a degradation product of the CaSR protein
(23). We also observed a high molecular weight band above
206 kD, which is thought to represent dimers of the receptor
(24).
We next determined whether the expression of the CaSR
protein was reduced in 5/6 Nx animals compared with sham-
operated animals. A representative autoradiograph of crude
kidney membrane protein from 5/6 Nx and sham-operated
animals was analyzed for CaSR expression by Western blot
(Figure 5). Densitometric quantification of the single broad
band between 1 17 and 140 kD from autoradiographs from
three different experiments demonstrates that the expression of
CaSR protein was 38% lower in Nx animals than in sham-
operated animals (Figure 6). This broad band was chosen
because the predicted size of the CaSR membrane protein is
approximately 120 kD (1). Thus, these data demonstrate that
both CaSR mRNA expression and CaSR protein content are
reduced by experimental chronic renal insufficiency in the rat.
Sham 5/6Nx
Figure 3. Densitometric quantification of kidney CaSR mRNA ex-
pression normalized to OAPDH mRNA expression 4 wk after surgery
from 15 sham and 19 Nx rats. Data are given as mean ± SEM.
J Am Soc Nephrol 9: 2067-2074, 1998 Reduced Renal CaSR in Renal Insufficiency 2071
HEK
CaSR Kidney
MW(kDa)
-�-2O6
-Is CaSR�-117
Figure 4. Expression of CaSR protein in HEK-293 cells stably trans-
fected with the human parathyroid CaSR (HEK-CaSR) and normal rat
kidney as determined by Western blot analysis. Membrane proteins
were obtained from HEK-CaR cells and normal rat kidney. Membrane
protein was added to each lane, subjected to polyacrylamide gel
electrophoresis (PAGE), electrotransferred to nitrocellulose filters,
and blotted using CaSR-specific antiserum 4641 (see Materials and
Methods). Lane I represents 4-p.g membrane protein obtained from
HEK-CaSR cells, and lanes 2 and 3 represent 50-p.g membraneprotein obtained from normal rat kidney. For both HEK-CaSR cells
and kidney, the broad specific bands above 206 kD represent dimers
of the receptor (24). The distinct doublet of immunoreactive bands
(HEK-CaR cells) and the single broad band or doublet (rat kidneytissue) between the 1 17- and 206-kD markers are compatible with the
glycosylated CaSR translated in vitro ( 1 ). The band that is detected
below I 17 kD in rat tissue represents a degradation product of the
CaSR protein (23).
DiscussionA CaSR from human and rat kidney was recently cloned
(2,3), and its predicted amino acid sequence shares approxi-
mateby 92% identity with that of the CaSR from bovine para-
thyroid glands (2). The identification of a CaSR in the kidney
provides a mechanism by which changes in [Ca2�]() can mod-
ulate renal tubular function directly (5-8). For example, high
[Ca2110 can inhibit both PTH-dependent cAMP production in
renal cortical cells (25) and vasopressin-dependent cAMP pro-
duction in the thick ascending limb of the loop of Henle (26).
These observations suggest that the CaSR plays an important
role in the regulation of renal tubular function.
In the present study, we examined the expression of the renal
CaSR in experimental moderate chronic renal insufficiency in
the rat. In rats with 5/6 Nx, the estimated GFR was reduced by
approximately 60%, serum PTH levels were increased, and
both serum l,25(OH)2D levels and urinary Ca2� excretion
were decreased when compared with sham-operated rats. We
found that the expression of the CaSR mRNA, as determined
by RNase protection assay, was decreased by 35% in 5/6 Nx
animals when compared with that in sham-operated animals.
We also found that the CaSR protein content, determined by
Western analysis, was reduced by 38%.
We calculated CaSR mRNA expression as the ratio of CaSR
mRNA to GAPDH mRNA. It is possible that in the hypertro-
phic remnant kidney of Nx animals, expression of the house-
keeping gene GAPDH is altered. In our studies, however, we
found no obvious changes in GAPDH mRNA expression in 5/6
Nx animals when compared with sham-operated animals. Sim-
ilarly, in studies of rats in which the degree of experimental
chronic renal insufficiency was similar to that in the present
study, GAPDH mRNA expression in both the kidney (27) and
liver (28) was unaffected when compared with the sham group.
Furthermore, the renal expression of 285 ribosomal RNA and
j3-actin mRNA was unaffected by chronic renal insufficiency
in the rat (29). Thus, our findings of reduced CaSR mRNA
relative to that of GAPDH mRNA in the kidney cannot be
attributed to altered expression of the housekeeping gene
GAPDH.
When we performed Western analysis, CaSR proteins from
kidney were detected at slightly lower molecular weights than
those obtained from HEK cells expressing the human parathy-
roid CaSR. This finding suggests that the protein expressed by
cells transfected with the parathyroid CaSR is different from
that expressed natively in renal tubular cells. Such a difference
might reflect differences in processing of the CaSR cDNA in
transfected cells when compared with renal tubular cells. A
second possibility is that there may be differences in posttrans-
lational modification of the protein. An alternative possibility
is that the rat kidney expresses different isoforms of the CaSR
than does human parathyroid tissue.
Little is known about the regulation of CaSR gene expres-
sion. In rats with either vitamin D deficiency (30,3 1 ), low
dietary Ca2� (32), or chronic renal insufficiency induced by
5/6 Nx (32) (three conditions in which serum PTH levels are
increased and [Ca2�]() is decreased), parathyroid gland CaSR
mRNA expression was not significantly different from control
values. Moreover, serum 1 ,25(OH),D levels were either de-
creased (30,3 1 ) or increased (32), suggesting that I ,25(OH),D
had no effect on parathyroid gland CaSR mRNA. In contrast,
Brown et a!. demonstrated that in rats fed a vitamin D-deficient
diet, CaSR mRNA expression in both the parathyroid gland
and kidney was decreased (3 1 ), and that administration of
l,25(OH),D3 resulted in an increase in CaSR mRNA levels to
near normal (3 1 ). Thus, in the present study, the finding of
decreased CaSR expression in kidney from rats with chronic
renal insufficiency might be attributed at least in part to the
observed decrease in their serum concentrations of
1 ,25(OH),D.
The 2#{176}HPTassociated with severe chronic renal insuffi-
ciency is thought to result not only from reduced serum levels
of 1 ,25(OH),D, but also from reduced sensitivity of the para-
thyroid gland to the suppressive effect of increased [Ca2�]() on
PTH secretion (1 2). This latter mechanism was first suggested
by Brown et a!. , who demonstrated that the concentration of
[Ca2�]0 required to suppress PTH secretion in vitro was higherin hyperplastic parathyroid glands from uremic individuals
with 2#{176}HPT than in normal glands (33). This notion was
further supported by observations that in hemodialysis patients
with 2#{176}HPT,the Ca2� set-point was increased in vivo (34). It

10
8
6
4
2
U)
.� !�
UU)C.)
�0
p<0.05
2072 Journal of the American Society of Nephrology J Am Soc Nephrol 9: 2067-2074, 1998
Nx Sham
‘: �-
Figure 5. The effect of chronic renal insufficiency on CaSR protein expression in rat kidney as determined by Western blot analysis. Membrane
proteins were obtained from the kidney remnant of 5/6 Nx rats and from both kidneys of sham-operated animals 4 wk after surgery. A total
of 50 i.�g of membrane protein was added to each lane, subjected to PAGE, electrotransferred to nitrocellulose filters, and Western blots were
performed using CaSR-specific antiserum 4641 (see Materials and Methods). Lanes 1 through 5 represent protein obtained from 5/6 Nx
animals, and lanes 6 through 9 represent protein obtained from sham-operated (Sham) animals.
Sham 5/6 Nx
Figure 6. Densitometric quantification of kidney protein expression 4
wk after surgery in 15 sham and 18 nephrectomized animals. Results
are shown as mean ± SEM.
should be noted, however, that in some dialysis patients with0 � . �±2 HPT, no increase in the Ca set-point has been detected
(35). Nevertheless, it has been suggested that such abnormal
Ca2� sensing by the parathyroid gland in chronic renal insuf-
ficiency reflects an abnormality in function of the parathyroid
cell CaSR. In fact, the expression of the CaSR was reduced in
parathyroid cells from patients undergoing chronic dialysis
when compared with that in parathyroid tissue from healthy
subjects (15,16).
In contrast to the severe chronic 2#{176}HPTfound in patients
with chronic renal failure undergoing long-term hemodialysis,
the biochemical disturbances observed in our study of 5/6 Nx
animals more likely represent mild-to-moderate 2#{176}HPT and
moderate renal insufficiency (Table 2). Such early 2#{176}HPT
could be the result of decreased serum bevels of 1 ,25(OH)2D.
However, we found that CaSR expression is reduced, at beast in
the kidney, and potentially in the parathyroid gland, early in the
development of renal insufficiency, and suggest that such
changes may play an important role in the early development
of disturbed mineral ion homeostasis, i.e. , 2#{176}HPTand possibly
l,25(OH)2D deficiency as well.
In patients with mild-to-moderate chronic renal insuffi-
ciency, urinary Ca2� excretion is significantly lower than that
in subjects with normal renal function (13,14). In our study, we
also found that urinary Ca2� excretion was reduced in animals
with experimental chronic renal insufficiency when compared
with sham-operated controls. In patients with FHH who harborinactivating mutations in the CaSR gene (10) and in mice in
which the CaSR gene has been knocked out (1 1), both exhibit
hypocalciuria. Furthermore, in patients with FHH rendered
hypoparathyroid by total parathyroidectomy, the reduced un-
nary Ca2� excretion persisted, providing evidence that dys-
function of the renal CaSR can mediate hypocalciunia, inde-
pendent of serum PTH bevels (36). Therefore, the observed
hypocalciuria in our study might also reflect an abnormality in
the function of the CaSR in the kidney that results from a
decrease in its renal expression.
An additional potential mechanism for the observed hy-
pocalciuria could reflect the stimubatory effect of increased
serum levels ofPTH on renal Ca2� reabsorption, but could also
reflect either deficiency of 1 ,25(OH)2D or resistance to its
actions on intestine and bone (14). However, the ability of PTH
to reduce urinary Ca2� excretion might be mitigated in patients
with chronic renal insufficiency because of resistance to the
actions of PTH in target tissues, such as bone (37,38), presum-
ably mediated by reduced expression of PTH receptors in such
tissues. Indeed, in 5/6 Nx rats, expression of PTH-PTH-releas-
ing protein (PTHrP) receptor mRNA was reduced in both
hepatocytes (28) and kidney (29), the batter tissue demonstrat-
ing marked reduction in PTH-dependent adenylate cyclase
activity (29). In patients with mild chronic renal insufficiency
and hypocalciuria, serum concentrations of both Ca2� and
1 ,25(OH)2D are reported to be not different from those in
controls (14). The observation that administration of
1 ,25(OH)2D3 to these patients induced an increase in urinary
Ca2� excretion (14) suggests that l,25(OH)2D3 resistance is
present in target tissues (i.e. , intestine and possibly bone) in
patients with mild-to-moderate chronic renal insufficiency.
Furthermore, the administration of 1 ,25(OH)2D3 to vitamin
D-deficient rats increased renal CaSR expression to near nor-
mal levels (3 1). Thus, another potential explanation for the
observed increased urinary Ca2� excretion in patients with

J Am Soc Nephrol 9: 2067-2074, 1998 Reduced Renal CaSR in Renal Insufficiency 2073
mild-to-moderate chronic renal insufficiency treated with
1 ,25(OH)2D3 is that this hormone resulted in increased renal
CaSR expression, and thereby improved renal tubular handlingof Ca2� . It remains to be determined whether administration of
1 ,25(OH)2D3 to animals with experimental chronic renal in-
sufficiency can alter the expression of the CaSR. Finally,
because we did not measure the urinary Ca2� ultrafiltrate, we
cannot determine whether the hypocalciuria observed in the 5/6
Nx animals was in part due to a decrease in the filtered load of
Ca2� with no change in its fractional reabsorption.
Decreased expression of calciotrophic hormone receptors in
the kidney and in other tissues could contribute to disorders of
mineral homeostasis in chronic renal insufficiency. For exam-
ple, reduced expression of PTH-PTHrP receptors in the kidney
could result in decreased phosphaturic effect of PTH on the
renal tubule and thereby contribute to the development of
hyperphosphatemia in chronic renal insufficiency. Similarly, in
experimental chronic renal insufficiency in the rat, the ob-
served resistance to the actions of growth hormone is due, at
least in part, to reduced expression of hepatic receptors for
insulin-like growth factor-b (39) and growth hormone (40).
These observations raise the fundamental question of the mo-
lecular basis for decreased receptor expression that is believed
to play an important role in the resistance to hormonal actions
in chronic renal insufficiency. Massry et al. observed that basal
levels of intracellular Ca2� were increased in hepatocytes and
cardiomyocytes from rats with experimental chronic renal in-
sufficiency, and hypothesized that such an increase of intra-
cellular Ca2� may induce a decrease in mRNA levels for
PTHrP receptors, and therefore be responsible for decreased
receptor expression in chronic renal insufficiency (28). More
recently, Baudouin-Legros et al. demonstrated that urea-in-
duced inhibition of cAMP production was reversed by protein
kinase inhibitors, thus suggesting that urea may directly inter-
fere with specific signaling pathways by altering cellular pro-
tein phosphorylation (41).
In summary, we demonstrate reduced expression of the
CaSR at the level of mRNA expression and protein content in
the kidney of rats with experimental chronic renal insuffi-
ciency. We hypothesize that reduced expression of the CaSR
gives rise to abnormal Ca2� sensing by the renal tubule, which
results in disordered tubular handling of Ca2�, and thereby
contributes to the hypocalciuria observed in patients with
chronic renal insufficiency. It would be of interest to perform
immunohistochemical localization of the CaSR in kidney from
animals with chronic renal insufficiency to determine whether
altered CaSR expression is specific in tubular segments along
the nephron, which may play a role in the development of
hypocalciuria found in chronic renal insufficiency.
AcknowledgmentsThis work was supported by National Institutes of Health Grant
MO1RRO127I, Pediatric Clinical Research Center, and by a grant
from the Oenentech Foundation. We are grateful to Drs. Forrest
Fuller, Jim Garrett, and Edward Nemeth (NPS Pharmaceuticals, SaltLake City, UT) for the kind gift of the CaSR riboprobe and anti-CaSR
antibody 4641. We also thank Dr. Edward Brown for his continued
support, suggestions, and discussions.
References1. Brown EM, Gamba 0, Riccardi D, Lombardi M, Butters R, Kifor
0, Sun A, Hediger MA, Lytton J, Hebert SC: Cloning and
characterization of an extracellular Ca(2+)-sensing receptorfrom bovine parathyroid. Nature 366: 575-580, 1993
2. Riccardi D, Park J, Lee WS, Oamba 0, Brown EM, Hebert SC:
Cloning and functional expression of a rat kidney extracellularcalcium/polyvalent cation-sensing receptor. Proc Nail Acad Sci
USA 92: 131-135, 1995
3. Aida K, Koishi 5, Tawata M, Onaya T: Molecular cloning of a
putative Ca(2+)-sensing receptor eDNA from human kidney.
Biochem Biophys Res Commun 214: 524-529, 1995
4. Brown EM, Gardner DO, Brennan MF, Marx Si, Spiegel AM,
Attie MF, Downs RW Jr. Doppman IL, Aurbach CD: Calcium-
regulated parathyroid hormone release in primary hyperparathy-roidism: Studies in vitro with dispersed parathyroid cells. Am J
Med 66: 923-931, 1979
5. Weisenger J, Farus M, Langman C, Bushinsky D: Regulation of
l,25-dihydroxyvitamin D3 by calcium in the parathyroidecto-
mized, parathyroid hormone-replete rat. J Bone Miner Res 4:
929-935, 1989
6. Clemens T, McGlade 5, Garrett KP, Craviso G, Hendy 0:Extracellular calcium modulates vitamin D-dependent calbindin-D28k gene expression in chick kidney cells. Endocrinology 124:
1582-1584, 1989
7. Quamme 0, Dirks J: Intraluminal and contraluminal magnesium
on magnesium and calcium transfer in the rat nephron. Am J
Physiol 238: Fl87-Fl98, 1980
8. Quamme 0, Dc Rouffignac C: Renal magnesium handling and
its hormonal control. Physiol Rev 74: 305-322, 1994
9. Riccardi D, Hall A, Xu J, Chattopadhyay N, Brown E, Hebert 5:Localization of calcium (polyvalent cation)-sensing receptor
(CaR) protein in rat kidney. J Am Soc Nephrol 8: 566A, 1997
10. Pollak M, Brown E, Chou Y-W, Hebert 5, Marx 5, Steinmann B,
Levi T, Seidman C, Seidman i: Mutations in the Ca2�-sensing
receptor gene cause familial hypocalciuric hypercalcemia and
neonatal severe hyperparathyroidism. Cell 75: 1 297-1 303, 1993
1 1. Ho C, Conner DA, Pollak MR. Ladd Di, Kifor 0, Warren HB,
Brown EM, Seidman JO, Seidman CE: A mouse model of humanfamilial hypocalciuric hypercalcemia and neonatal severe hyper-
parathyroidism [see Comments]. Nat Genet I 1: 389-394, 1995
12. Llach F: Secondary hyperparathyroidism in renal failure: The
trade-off hypothesis revisited. Am J Kidney Dis 25: 663-679,
1995
13. Heaby M, Malluche H, Goldstein D, Singer F, Massry S: Effects
of long-term therapy with calcitriol in patients with moderaterenal failure.Arch Intern Med 140: 1030-1033, 1980
14. Cremer B, Lubbers E, Klooker P. Schmidt-Gayk H, Ritz E:
Calciuric response to b,25-(OH),D3 in early renal failure. MinerElectrolyteMetab 11: 182-185, 1985
15. Kifor 0, Moore FD Jr. Wang P, Goldstein M, Vassilev P. Kifor
I, Hebert SC, Brown EM: Reduced immunostaining for the
extracellular Ca24-sensing receptor in primary and uremic sec-
ondary hyperparathyroidism. J C/in Endocrinol Metab 81:
1598-1606, 1996
16. Oogusev I, Duchambon P. Hory B, Giovannini M, Ooureau Y,Sarfati E, Drueke TB: Depressed expression of calcium receptorin parathyroid gland tissue of patients with hyperparathyroidism.
Kidney hit 51: 328-336, 1997

2074 Journal of the American Society of Nephrology J Am Soc Nephrol 9: 2067-2074, 1998
17. Khosla 5, Ebeling P, Firek A, Burritt M, Kao P, Heath H III:
Calcium infusion suggests a “set-point” abnormality of parathy-
roid gland function in familial benign hypercalcemia and more
complex disturbances in primary hyperparathyroidism. J C/in
Endocrinol Metab 76: 7 15-720, 1993
18. Reyes A, Purkerson M, Karl I, Klahr 5: Dietary supplementation
with L-arginine ameliorates the progression of renal disease in
rats with subtotal nephrectomy. Am J Kidney Dis 20: 168-176,
1992
19. Ingram A, Parbtani A, Clark W, Spanner E, Huff M, Philbrick D,
Hobub B: Effects of flaxseed and flax oil diets in a rat 5/6 renal
ablation model. Am J Kidney Dis 25: 320-329, 1995
20. Chomczynski P, Sacchi N: Single-step method of RNA isolation
by acid guanidinium isothiocyanate-phenol-chloroform extrac-
tion. Anal Biochem 162: 156-159, 1987
21 . Chattopadhyay N, Mithal A, Brown EM: The calcium-sensing
receptor: A window into the physiology and pathophysiology of
mineral ion metabolism. Endocr Rev 17: 289-307, 1996
22. Laemmli U: Cleavage of structural proteins during the assembly
of the head of bacteriophage T4. Nature 227: 680-685, 1970
23. Garrett iE, Tamir H, Kifor 0, Simm RT, Rogers KV, Mithal A,
Gagel RF, Brown EM: Calcitonin-secreting cells of the thyroid
express an extracellular calcium receptor gene. Endocrinology
136: 5202-5211, 1995
24. Chattopadhyay N, Baum M, Bai M, Riccardi D, Hebert SC,
Harris HW, Brown EM: Ontogeny of the extracellular calcium-
sensing receptor in rat kidney. Am J Physiol 271: F736-F743,
I 996
25. Mathias R, Brown E: Divalent cations modulate PTH-dependent
3’,5’-cyclic adenosine monophosphate production in renal prox-
imal tubular cells. Endocrinology 128: 3005-3012, 1991
26. Takaichi K, Uchida 5, Kurokawa K: High Ca2� inhibits AVP-
dependent cAMP production in thick ascending limbs of Henle.
Am J Physiol 250: F770-F776, 1986
27. Wang D, Yao A, Zhao H, DiPette D: Regulation of ANO II
receptor in hypertension: Role of ANG II. Am J Physiol 271:
H120-Hl25, 1996
28. Massry 5, Klin M, Ni Z, Tian J, Kedes L, Smogorzewski M:
Impaired agonist-induced calcium signaling in hepatocytes from
chronic renal failure rats. Kidney mt 48: 1324-1331, 1995
29. Urena P, Kubrusly M, Mannstadt M, Hruby M, Tan M-M, Silve
C, Lacour B, Abou-Samra A-B, Segre G, Drueke T: The renalPTH/PTHRP receptor is down-regulated in rats with chronic
renal failure. Kidney mt 45: 605-61 1, 1994
30. Rogers K, Dunn C, Conklin R, Hadfield 5, Petty V. Brown E,
Hebert 5, Nemeth E, Fox i: Calcium receptor messenger ribo-
nucleic acid levels in the parathyroid glands and kidney of
vitamin D-deficient rats are not regulated by plasma calcium or
1,25-(OH)2D3. Endocrinology 136: 499-504, 1995
3 1. Brown AJ, Zhong M, Finch J, Ritter C, McCracken R, Morrissey
I, Slatopolsky E: Rat calcium-sensing receptor is regulated by
vitamin D but not by calcium. Am J Physiol 270: F454-F460,
1996
32. Fox i, Brown EM, Hebert SC, Rogers KV: Parathyroid gland
calcium receptor gene expression is unaffected by chronic renal
failure or low dietary calcium in rats. JAm Soc Nephrol 5: 879A,
1994
33. Brown E, Wilson R, Eastman R, Pallotta I, Marynick 5: Abnor-
mal regulation of parathyroid hormone release by calcium in
secondary hyperparathyroidism due to chronic renal failure.
J C/in Endocrinol Metab 54: 172-179, 1982
34. Rodriguez M, Caravaca F, Fernandez E, Borrego Mi, Lorenzo V,
Cubero J, Martin-Malo A, Betriu A, Rodriguez AP, Felsenfeld
Ai: Evidence for both abnormal set point of PTH stimulation by
calcium and adaptation to serum calcium in hemodialysis pa-
tients with hyperparathyroidism. J Bone Miner Res 12: 347-355,
1997
35. Sanchez C, Goodman W, Ramirez J, Gales B, Belin T, Segre 0,
Salusky I: Calcium-regulated parathyroid hormone secretion in
adynamic renal insufficiency. Kidney mt 48: 838-843, 1995
36. Attie MF, Gill JR Jr, Stock JL, Spiegel AM, Downs RW Jr,
Levine MA, Marx Si: Urinary calcium excretion in familial
hypocalciuric hypercalcemia: Persistence of relative hypocalci-
uria after induction of hypoparathyroidism. J C/in invest 72:
667-676, 1983
37. Massry 5, Coburn I, Lee D, iowsey I, Kleeman C: Skeletal
resistance to parathyroid hormone in renal failure: Study in 105
human subjects. Ann intern Med 78: 357-364, 1973
38. Drueke T: Abnormal skeletal response to parathyroid hormone
and the expression of its receptor in chronic uremia. Pediatr
Nephrol 10: 348-350, 1996
39. Chan W, Valerie K, Chan J: Expression of insulin-bike growth
factor-b in uremic rats. Kidney mt 43: 790-795, 1993
40. Tonshoff B, Eden 5, Weiser E, Carlsson B, Robinson I, Blum W,
Mehls 0: Reduced hepatic growth hormone (OH) receptor gene
expression and increased plasma OH binding protein in experi-
mental uremia. Kidney In,’ 45: 1085-1092, 1994
41 . Baudouin-Legros M, Asdram L, Tondelier D, Paulais M, Anag-
nostopoulos T: Extracellular urea concentration modulates
cAMP production in the mouse MTAL. Kidney mt 50: 26-33,
I996